Клиническая фармакокинетика: теоретические, прикладные и аналитические аспекты: руководство / Под ред. В.Г. Кукеса. - 2009. - 432 с
|
|
|
|
ГЛАВА 5 БИОТРАНСФОРМАЦИЯ ЛЕКАРСТВЕННЫХ СРЕДСТВ
В.Г. Кукес, Д.А. Сычёв, Г.В. Раменская, И.В. Игнатьев
Человек ежедневно подвергается воздействию множества инородных химических веществ, называемых «ксенобиотики». Ксенобиотики попадают в организм человека через лёгкие, кожу и из пищеварительного тракта в составе примесей воздуха, пищи, напитков, ЛС. Некоторые ксенобиотики не оказывают никакого воздействия на организм человека. Однако большинство ксенобиотиков могут вызывать биологические ответные реакции. Организм реагирует на ЛС так же, как и на любой другой ксенобиотик. При этом ЛС становятся объектами различных механизмов воздействия со стороны организма. Это, как правило, приводит к нейтрализации и элиминации (выведению) ЛС. Некоторые, легко растворимые в воде, ЛС элиминируются почками в неизменённом виде, другие вещества предварительно подвергаются воздействию ферментов, изменяющих их химическое строение. Таким образом, биотрансформация - общее понятие, включающее все химические изменения, происходящие с ЛС в организме. Результат биологической трансформации ЛС: с одной стороны - снижается растворимость веществ в жирах (липофильность) и повышается их растворимость в воде (гидрофильность), а с другой стороны - изменяется фармакологическая активность препарата.
Снижение липофильности и повышение гидрофильности лекарственных средств
Небольшое число ЛС способно выводиться почками в неизменён- ном виде. Чаще всего эти препараты представляют «малые молекулы» или они способны находиться в ионизированном состоянии при физиологических значениях рН. Большинство ЛС не обладают такими физико-химическими свойствами. Фармакологически активные органические молекулы чаще липофильны и остаются неионизированными при физиологических значениях рН. Эти ЛС обычно связаны с белками плазмы, плохо фильтруются в почечных клубочках и одновременно легко реабсорбируются в почечных канальцах. Биотрансформация (или система биотрансформации) направлена на повышение растворимости молекулы ЛС (повышение гидрофильности), что способствует выведению его из организма с мочой. Иными словами, липофильные ЛС превращаются в гидрофильные и, следовательно, в более легковыводимые соединения.
Изменение фармакологической активности лекарственных средств
Направления изменения фармакологической активности ЛС в результате биотрансформации.
• Фармакологически активное вещество превращается в фармакологически неактивное (это характерно для большинства ЛС).
• Фармакологически активное вещество на первом этапе превращается в другое фармакологически активное вещество (табл. 5-1).
• Неактивное фармакологическое ЛС превращается в организме в фармакологически активное вещество; такие препараты называют «пролекарства» (табл. 5-2).
Таблица 5-1. Лекарственные средства, метаболиты которых сохраняют фармакологическую активность
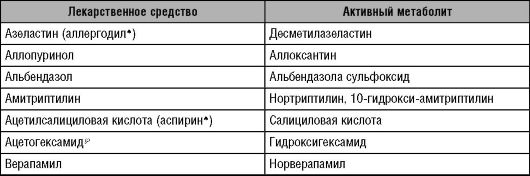 Окончание таблицы 5-1
Окончание таблицы 5-1
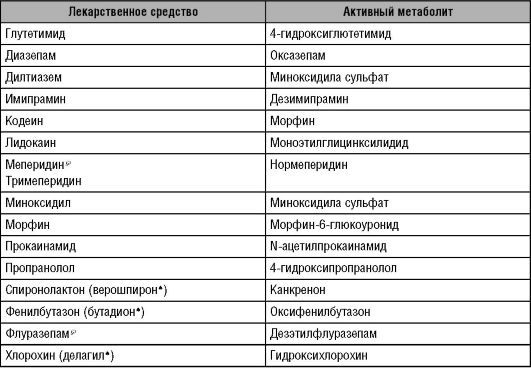 Таблица 5-2. Пролекарства
Таблица 5-2. Пролекарства
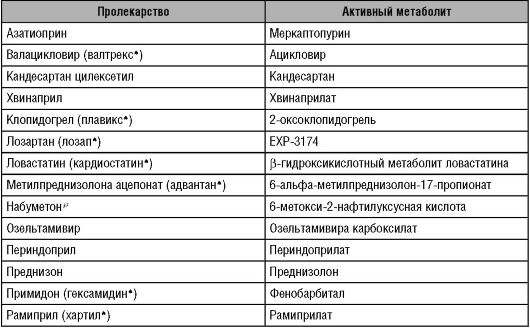 Окончание таблицы 5-2
Окончание таблицы 5-2
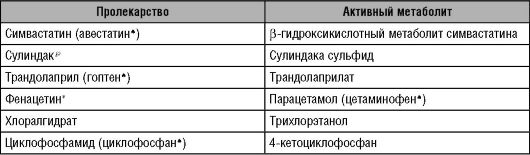 * Фенацетин снят с производства из-за выраженных побочных эффектов, в частности, нефротоксичности («фенацетиновый нефрит»).
* Фенацетин снят с производства из-за выраженных побочных эффектов, в частности, нефротоксичности («фенацетиновый нефрит»).
Следует отметить, что эффективность и безопасность применения ЛС (перечислены в табл. 5-1), имеющих активные метаболиты, зависят не только от фармакокинетики собственно ЛС, но и от фармакокинетики их активных метаболитов.
5.1. ПРОЛЕКАРСТВА
Одна из целей создания пролекарств - улучшение фармакокинетических свойств; это ускоряет и увеличивает всасывание веществ. Так, были разработаны сложные эфиры ампициллина (пивампицинp, талампицинp и бикампицинp), в отличие от ампициллина практически полностью всасывающиеся при приёме внутрь (98-99%). В печени эти препараты под действием карбоксиэстераз гидролизуются до ампициллина, обладающего антибактериальной активностью.
Биологическая доступность противовирусного ЛС валацикловира составляет 54%, в печени он превращается в ацикловир. Следует отметить, что биодоступность собственно ацикловира не превышает 20%. Высокая биодоступность валацикловира обусловлена наличием в его молекуле остатка аминокислоты валина. Именно поэтому валацикловир всасывается в кишечнике путём активного транспорта с помощью транспортёра олигопептидов PEPT 1.
Ещё один пример: ингибиторы аденозинпревращающего фермента, содержащие карбоксильную группу (эналаприл, периндоприл, трандолаприл, хвинаприл, спираприл, рамиприл и др.). Так, эналаприл всасывается при приёме внутрь на 60%, гидролизуется в печени под влиянием карбоксиэстераз до активного эналаприлата. Необходимо отметить: эналаприлат при введении внутрь всасывается лишь на 10%.
Другая цель разработки пролекарств - повышение безопасности лекарственных веществ. Например, учёные создали сулиндакp - НПВС. Данный препарат изначально не блокирует синтез простагландинов. Лишь в печени сулиндакp гидролизуется с образованием активного сульфида сулиндакаp (именно это вещество обладает противовоспалительной активностью). Предполагали, что сулиндакp не будет обладать ульцерогенным действием. Однако ульцерогенность НПВС обусловлена не местным, а «системным» действием, поэтому, как показали исследования, частота возникновения эрозивно-язвенных поражений органов пищеварения при приёме сулиндакаp и других НПВС примерно одинакова.
Ещё одна цель создания пролекарств - повышение избирательности действия ЛС; это увеличивает эффективность и безопасность препаратов. Дофамин используют для усиления почечного кровотока при острой почечной недостаточности, однако препарат влияет на миокард и сосуды. Отмечают повышение АД, развитие тахикардии и аритмий. Присоединение к дофамину остатка глутаминовой кислоты позволило создать новый препарат - глутамил-дофаp. Глутамил-дофаp гидролизуется до дофамина только в почках под влиянием глутамилтранспептидазы и декарбоксилазы L-ароматических аминокислот и таким образом практически не оказывает нежелательного действия на центральную гемодинамику.
 Рис. 5-1. Фазы биотрансформации лекарственных средств (Katzung В., 1998)
Рис. 5-1. Фазы биотрансформации лекарственных средств (Katzung В., 1998)
5.2. ФАЗЫ БИОТРАНСФОРМАЦИИ ЛЕКАРСТВЕННЫХ СРЕДСТВ
Процессы биотрансформации большинства ЛС происходят в печени. Однако биотрансформация ЛС может протекать и в других органах, например, в пищеварительном тракте, лёгких, почках.
В целом, все реакции биотрансформации ЛС можно отнести к одной из двух категорий, обозначаемых как фаза биотрансформации I и фаза биотрансформации II.
Реакции I фазы (несинтетические реакции)
В процессе несинтетических реакций ЛС переходят в более полярные и лучше растворимые в воде (гидрофильные) соединения, чем исходное вещество. Изменения исходных физико-химических свойств ЛС обусловлены присоединением или освобождением активных функциональных групп: например, гидроксильных (-ОН), сульфгидрильных (-SH), аминогрупп (-NH2). Основные реакции I фазы - реакции окисления. Гидроксилирование - наиболее распространённая реакция окисления - присоединение гидроксильного радикала (-ОН). Таким образом, можно считать, что в I фазу биотрансформации происходит «взлом» молекулы ЛС (табл. 5-3). Катализаторы указанных реакций - ферменты, называемые «оксидазы со смешанной функцией». В целом, субстратная специфичность этих ферментов очень низка, поэтому они окисляют различные лекарственные вещества. К другим, менее частым реакциям I фазы, относят процессы восстановления и гидролиза.
Реакции II фазы (синтетические реакции)
Реакции II фазы биотрансформации, или синтетические реакции, представляют соединение (конъюгацию) ЛС и/или его метаболитов с эндогенными веществами, в результате образуются полярные, хорошо растворимые в воде конъюгаты, легко выводимые почками или с желчью. Для вступления в реакцию II фазы молекула должна обладать химически активным радикалом (группировкой), к которому может присоединиться конъюгирующая молекула. Если активные радикалы присутствуют в молекуле ЛС изначально, тогда реакция конъюгации протекает, минуя реакции I фазы. Иногда молекула лекарственного вещества приобретает активные радикалы в ходе реакций I фазы (табл. 5-4).
Таблица 5-3. Реакции I фазы (Katzung 1998; с дополнениями)
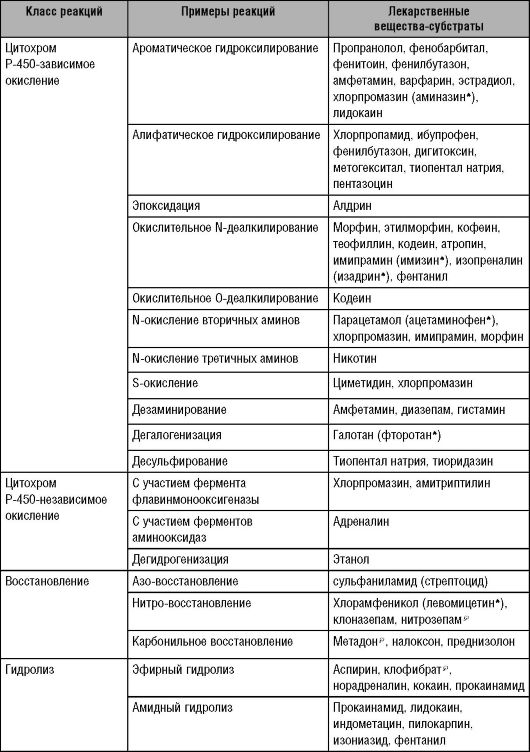 Таблица 5-4. Реакции II фазы (Katzung 1998; с дополнениями)
Таблица 5-4. Реакции II фазы (Katzung 1998; с дополнениями)
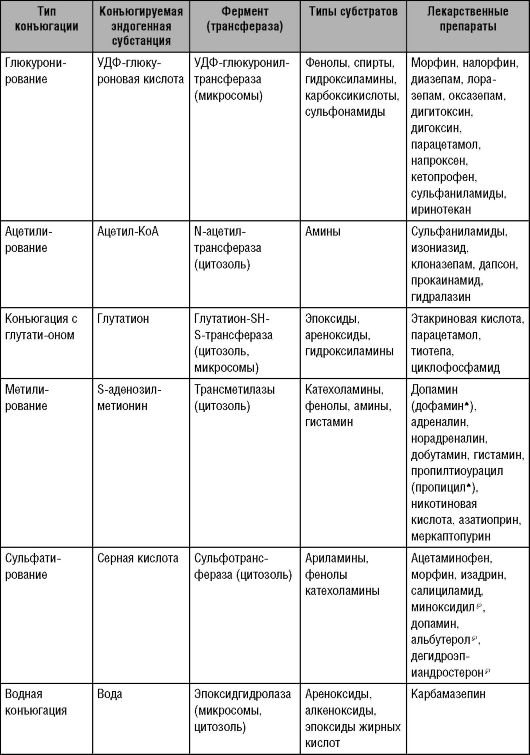 Следует
отметить, что препарат в процессе биотрансформации может превращаться
только за счёт реакций I фазы, либо - исключительно за счёт реакций II
фазы. Иногда часть ЛС метаболизируется путём реакций I фазы, а часть -
путём реакций II фазы. Кроме того, существует возможность
последовательного прохождения реакций I и II фазы (рис. 5-2).
Следует
отметить, что препарат в процессе биотрансформации может превращаться
только за счёт реакций I фазы, либо - исключительно за счёт реакций II
фазы. Иногда часть ЛС метаболизируется путём реакций I фазы, а часть -
путём реакций II фазы. Кроме того, существует возможность
последовательного прохождения реакций I и II фазы (рис. 5-2).
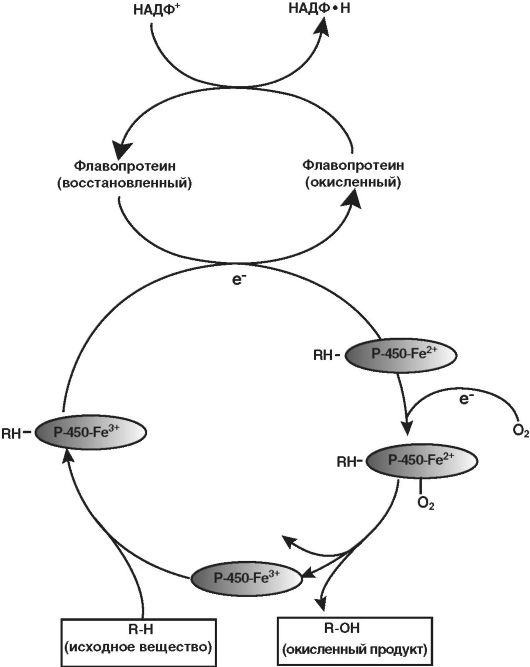 Рис. 5-2. Функционирование системы оксидаз со смешанной функцией
Рис. 5-2. Функционирование системы оксидаз со смешанной функцией
Эффект первого прохождения через печень
Биотрансформация большинства ЛС осуществляется в печени. ЛС, метаболизм которых протекает в печени, подразделяют на две подгруппы: вещества с высоким печёночным клиренсом и вещества с низким печёночным клиренсом.
• Для ЛС с высоким печёночным клиренсом характерна высокая степень извлечения (экстракции) из крови, что обусловлено значительной активностью (ёмкостью) метаболизирующих их ферментных систем (табл. 5-5). Поскольку такие ЛС быстро и легко метаболизируются в печени, клиренс их зависит от величины и скорости печёночного кровотока.
• ЛС с низким печёночным клиренсом. Печёночный клиренс зависит не от скорости печёночного кровотока, а от активности ферментов и степени связывания ЛС с белками крови.
Таблица 5-5. Лекарственные средства с высоким печёночным клиренсом
 При
одинаковой ёмкости ферментных систем лекарственные вещества, в
значительной степени связанные с белками (дифенин, хинидин, толбутамид),
будут иметь низкий клиренс, по сравнению со слабосвязанными с белками
ЛС (теофиллин, парацетамол). Ёмкость ферментных систем - не постоянная
величина. Например, уменьшение ёмкости ферментных систем регистрируют
при увеличении дозы ЛС (вследствие насыщения ферментов); это может
привести к увеличению биологической доступности ЛС.
При
одинаковой ёмкости ферментных систем лекарственные вещества, в
значительной степени связанные с белками (дифенин, хинидин, толбутамид),
будут иметь низкий клиренс, по сравнению со слабосвязанными с белками
ЛС (теофиллин, парацетамол). Ёмкость ферментных систем - не постоянная
величина. Например, уменьшение ёмкости ферментных систем регистрируют
при увеличении дозы ЛС (вследствие насыщения ферментов); это может
привести к увеличению биологической доступности ЛС.
При приёме внутрь ЛС с высоким печёночным клиренсом, они всасываются в тонкой кишке и через систему воротной вены поступают в печень, где подвергаются активному метаболизму (на 50-80%) ещё до поступления в системное кровообращение. Этот процесс известен как пресистемная элиминация, или эффект «первого прохождения» («first-pass effect»). В результате такие ЛС имеют низкую биологическую доступность при приёме внутрь, при этом абсорбция их может составлять почти 100%. Эффект первого прохождения характерен для таких препаратов, как аминазин, ацетилсалициловая кислота, вера-
памил, гидралазин, изопреналин, имипрамин, кортизон, лабетолол, лидокаин, морфин. Метопролол, метилтестостерон, метоклопрамид, нортриптилинp, окспренололp, органические нитраты, пропранолол, резерпин, салициламид, морацизин (этмозин) и некоторые другие препараты также подвергаются пресистемной элиминации. Следует отметить, что незначительная биотрансформация ЛС может проходить и в других органах (просвете и стенке кишечника, лёгких, плазме крови, почках и других органах).
Как показали исследования последних лет, эффект первого прохождения через печень зависит не только от процессов биотрансформации ЛС, но и от функционирования транспортёров ЛС, и, прежде всего, гликопротеина-Р и транспортёров органических анионов и катионов (см. «Роль транспортёров лекарственных средств в фармакокинетических процессах»).
5.3. ФЕРМЕНТЫ I ФАЗЫ БИОТРАНСФОРМАЦИИ ЛЕКАРСТВЕННЫХ СРЕДСТВ
Микросомальная система
Многие ферменты, метаболизирующие ЛС, располагаются на мембранах эндоплазматического ретикулума (ЭПР) печени и других тканей. При изоляции мембран ЭПР путём гомогенизации и фракционирования клетки, мембраны преобразуются в везикулы, называемые «микросомы». Микросомы сохраняют большинство морфологических и функциональных характеристик интактных мембран ЭПР, включая свойство шероховатости или гладкости поверхности, соответственно шероховатого (рибосомального) и гладкого (нерибосомального) ЭПР. В то время как шероховатые микросомы в основном связаны с синтезом белка, гладкие - относительно богаты ферментами, ответственными за окислительный метаболизм лекарственных веществ. В частности, гладкие микросомы содержат ферменты, известные как оксидазы со смешанной функцией, или монооксигеназы. Активность этих ферментов требует присутствия как восстанавливающего агента - никотинамидадениндинуклеотидфосфата (НАДФ-Н), так и молекулярного кислорода. При типичной реакции расходуется (восстанавливается) одна молекула кислорода на одну молекулу субстрата, при этом один кислородный атом включается в продукт реакции, а другой образует молекулу воды.
В этом окислительно-восстановительном процессе ключевую роль играют два микросомальных фермента.
• Флавопротеин НАДФ-Н-цитохром Р-450-редуктаза. Один моль этого фермента содержит по одному молю флавинмононуклеотида и флавинадениндинуклеотида. Поскольку цитохром С может служить акцептором электрона, то указанный фермент часто называют НАДФ-цитохром С-редуктазой.
• Гемопротеин, или цитохром Р-450 выполняет функцию конечной оксидазы. В действительности микросомальная мембрана содержит множество форм данного гемопротеина, и эта множественность возрастает при повторном введении ксенобиотиков. Относительное изобилие цитохрома Р-450, по сравнению с редуктазой печени, делает процесс восстановления гема цитохрома Р-450 лимитирующей стадией в процессе окисления лекарственных веществ в печени.
Процесс микросомального окисления ЛС требует участия цитохрома Р-450, цитохрома Р-450-редуктазы, НАДФ-Н и молекулярного кислорода. Упрощённая схема окислительного цикла представлена на рисунке (рис. 5-3). Окисленный (Fe3+) цитохром Р-450 соединяется с лекарственным субстратом с образованием бинарного комплекса. НАДФ-Н - донор электрона для флавопротеинредуктазы, которая, в свою очередь, восстанавливает окисленный комплекс цитохром Р-450-лекарство. Второй электрон переходит от НАДФ-Н через ту же флавопротеинредуктазу, восстанавливающую молекулярный кислород и формирующую комплекс «активированный кислород»-цитохром Р-450-субстрат. Этот комплекс переносит «активированный кислород» на лекарственный субстрат с образованием окисленного продукта.
Цитохром Р-450
Цитохром Р-450, в литературе часто обозначаемый CYP, представляет группу ферментов, осуществляющих не только метаболизм ЛС и других ксенобиотиков, но и участвующих в синтезе глюкокортикоидных гормонов, желчных кислот, простаноидов (тромбоксана А2, простациклина I2), холестерина. Впервые цитохром Р-450 идентифицировали Klingenberg и Garfincell в микросомах печени крысы в 1958 году. Филогенетические исследования показали, что цитохромы Р-450 появились в живых организмах около 3,5 млрд лет назад. Цитохром Р-450 - гемопротеин: он содержит гем. Название цитохрома Р-450 связано с особыми свойствами этого гемопротеина. В восстановлен-
ной форме цитохром Р-450 связывает монооксид углерода с образованием комплекса с максимальным поглощением света при длине волны 450 нм. Это свойство объясняют тем, что в геме цитохрома Р-450 железо связано не только с атомами азота четырёх лигандов (при этом образуя порфириновое кольцо). Существуют также пятый и шестой лиганды (сверху и снизу кольца гема) - атом азота гистидина и атом серы цистеина, входящие в состав полипептидной цепи белковой части цитохрома Р-450. Наибольшее количество цитохрома Р-450 располагается в гепатоцитах. Однако цитохром Р-450 обнаруживают и в других органах: в кишечнике, почках, лёгких, надпочечниках, головном мозге, коже, плаценте и миокарде. Важнейшее свойство цитохрома Р-450 - способность метаболизировать практически все известные химические соединения. Наиболее важная реакция - гидроксилирование. Как уже указывалось, цитохромы Р-450 ещё называют монооксигеназами, так как они включают один атом кислорода в субстрат, окисляя его, а один - в воду, в отличие от диоксигеназ, которые включают оба атома кислорода в субстрат.
Цитохром Р-450 имеет множество изоформ - изоферментов. В настоящее время выделено более 1000 изоферментов цитохрома Р-450. Изоферменты цитохрома Р-450, по классификации Nebert (1987), принято разделять по близости (гомологии) нуклеотид/амино- кислотной последовательности на семейства. В свою очередь, семейства подразделяют на подсемейства. Изоферменты цитохрома Р-450 с идентичностью аминокислотного состава более 40% объединены в семейства (выделено 36 семейств, 12 из них обнаружены у млекопитающих). Изоферменты цитохрома Р-450 с идентичностью аминокислотного состава более 55% объединены в подсемейства (выделено 39 подсемейств). Семейства цитохромов Р-450 принято обозначать римскими цифрами, подсемейства - римскими цифрами и латинской буквой.
Схема обозначения отдельных изоферментов.
• Первый символ (вначале) - арабская цифра, обозначающая семейство.
• Второй символ - латинская буква, обозначающая подсемейство.
• В конце (третий символ) указывают арабскую цифру, соответствующую изоферменту.
Например, изофермент цитохрома Р-450, обозначенный как CYP3A4, принадлежит к семейству 3, подсемейству IIIA. Изоферменты цитохрома Р-450 - представители различных семействи подсемейств-
различаются регуляторами активности (ингибиторы и индукторы) и субстратной специфичностью1. Например, CYP2C9 метаболизирует исключительно S-варфарин, в то время как R-варфарин метаболизируют изоферменты CYP1A2 и CYP3A4.
Однако члены отдельных семейств, подсемейств и отдельные изоферменты цитохрома Р-450 могут обладать перекрёстной субстратной специфичностью, а также иметь перекрёстные ингибиторы и индукторы. Например, ритонавир (противовирусный препарат) метаболизируют принадлежащие к различным семействам и подсемействам 7 изоферментов (CYP1A2, CYP2A6, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4). Циметидин одновременно ингибирует 4 изофермента: CYP1A2, СYP2C9, CYP2D6 и CYP3A4. В метаболизме ЛС принимают участие изоферменты цитохрома Р-450 I, II и III семейств. CYP1A1, CYP1A2, CYP2A6, CYP2B6, CYP2D6, CYP2C9, CYP209, CYP2E1, CYP3A4 - наиболее важные для метаболизма лекарственных веществ и хорошо изученные изоферменты цитохрома Р-450. Содержание различных изоферментов цитохрома Р-450 в печени человека, а также их вклад в окисление ЛС различны (табл. 5-6). Лекарственные вещества - субстраты, ингибиторы и индукторы изоферментов цитохрома Р-450 представлены в приложении 1.
Таблица 5-6. Содержание изоферментов цитохрома Р-450 в печени человека и их вклад в окисление лекарственных средств (Lewis и соавт., 1999)
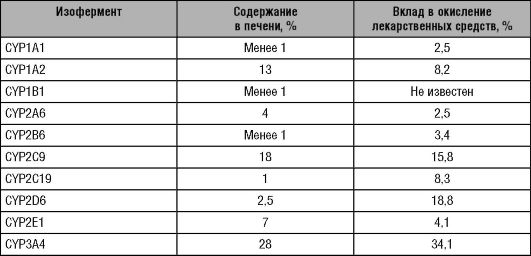 1 Некоторые изоферменты цитохрома Р-450 обладают не только субстратной специфичностью, но и стереоспецифичностью.
1 Некоторые изоферменты цитохрома Р-450 обладают не только субстратной специфичностью, но и стереоспецифичностью.
До сих пор не известны эндогенные субстраты для изоферментов семейства CYPI. Эти изоферменты метаболизируют ксенобиотики: некоторые ЛС и ПАУ - основные компоненты табачного дыма и продукты сжигания органического топлива. Отличительная особенность изоферментов семейства СYPI - их способность к индукции под действием ПАУ, в том числе, диоксина и 2,3,7,8-тетрахлорди- бензо-р-диоксина (TCDD). Поэтому семейство СYPI в литературе называют «цитохром, индуцибельный ПАУ»; «диоксин-индуцибельный цитохром» или «ТСDD-индуцибельный цитохром». В организме человека семейство СYPI представлено двумя подсемействами: IА и IB. В состав подсемейства IA входят изоферменты 1А1 и 1А2. В состав подсемейства IB входит изофермент 1В1.
Подсемейство цитохрома Р-450 CYPIIA
Изофермент 1А1 цитохрома Р-450 (CYP1A1) обнаруживают в основном в лёгких, в меньшей степени - в лимфоцитах и плаценте. CYP1A1 не участвует в метаболизме ЛС, однако в лёгких этот изофермент активно метаболизирует ПАУ. При этом некоторые ПАУ, например, бензопирен и нитрозамины превращаются в канцерогенные соединения, способные спровоцировать развитие злокачественных новообразований, в первую очередь - рака лёгких. Этот процесс получил название «биологическая активация канцерогенов». Как и другие цитохромы семейства CYPI, CYP1A1 индуцируется ПАУ. При этом изучен механизм индукции CYP1A1 под влиянием ПАУ. Проникнув в клетку, ПАУ соединяются с Аh-рецептором (белок из класса регуляторов транскрипции); образовавшийся комплекс ПАУ-Ап-рецептор проникает в ядро при помощи другого белка - ARNT, а затем стимулирует экспрессию гена CYP1A1, связываясь со специфическим диоксин-чувствительным участком (сайтом) гена. Таким образом, у курящих людей процессы индукции CYP1A1 протекают наиболее интенсивно; это приводит к биологической активации канцерогенов. Именно этим объясняют высокий риск возникновения рака легких у курильщиков.
Изофермент 1А2 цитохрома Р-450 (CYP1A2) обнаруживают в основном в печени. В отличие от цитохрома CYP1A1, CYP1A2 метаболизирует не только ПАУ, но и ряд ЛС (теофиллин, кофеин и другие препараты). В качестве маркёрных субстратов для фенотипирования CYP1A2 используют фенацетин, кофеин и антипирин. При этом фенацетин подвергают О-деметилированию, кофеин - 3-деметилированию, а антипирин - 4-гидроксилированию. Оценка
клиренса кофеина - важный диагностический тест, позволяющий определить функциональное состояние печени. В связи с тем, что CYP1A2 - главный метаболизирующий фермент кофеина, по сути, в данном тесте определяют активность указанного изофермента. Пациенту предлагают принять внутрь кофеин, меченный радиоактивным изотопом углерода С13 (С13-кофеин), затем выдыхаемый пациентом воздух в течение часа собирают в специальный резервуар и анализируют. При этом в выдыхаемом пациентом воздухе содержится радиоактивный углекислый газ (С13О2 - образован радиоактивным углеродом) и обычный углекислый газ (С12О2). По соотношению в выдыхаемом воздухе С13О2 к С12О2 (измеряют с помощью масс-спектроскопии) определяют клиренс кофеина. Существует модификация этого теста: методом высокоэффективной жидкостной хроматографии определяют концентрацию кофеина и его метаболитов в плазме крови, моче и слюне, взятых натощак. В этом случае определённый вклад в метаболизм кофеина вносят цитохромы CYP3A4 и CYP2D6. Оценка клиренса кофеина - надёж- ный тест, позволяющий оценить функциональное состояние печени при её выраженном поражении (например, при циррозе печени) и определить степень нарушений. К недостаткам теста относят его недостаточную чувствительность при умеренном поражении печени. На результат теста влияют курение (индукция CYP1A2), возраст, совместное применение ЛС, изменяющих активность изоферментов цитохрома Р-450 (ингибиторов или индукторов).
Подсемейство цитохрома Р-450 CYPIIA
Из изоферментов подсемейства CYPIIA наиболее важную роль в метаболизме ЛС играет изофермент цитохрома Р-450 2А6 (CYP2A6). Общее свойство изоферментов подсемейства CYPIIA - способность к индукции под действием фенобарбитала, поэтому подсемейство CYPIIA называют фенобарбитал-индуцибельными цитохромами.
Изофермент цитохрома Р-450 2А6 (CYP2A6) обнаруживают, в основном, в печени. CYP2A6 метаболизирует небольшое число ЛС. С помощью данного изофермента происходит превращение никотина в котинин, а также котинина в 3-гидроксикотинин; 7-гидроксили- рование кумарина; 7-гидроксилирование циклофосфана. CYP2A6 вносит определённый вклад в метаболизм ритонавира, парацетамола и вальпроевой кислоты. CYP2A6 принимает участие в биологической активации компонентов табачного дыма нитрозоаминов - канцерогенов, вызывающих рак лёгких. CYP2A6 способствует биоактивации
мощных мутагенов: 6-амино-(х)-ризена и 2-амино-3-метилмидазо- (4,5-f)-кванолина.
Подсемейство цитохрома Р450 CYPIIB
Из изоферментов подсемейства CYPIIB наиболее важную роль в метаболизме ЛС играет изофермент CYP2В6. Общее свойство изоферментов подсемейства CYPIIB - способность к индукции под действием фенобарбитала.
Изофермент цитохрома Р-450 2В6 (CYP2В6) участвует в метаболизме небольшого числа ЛС (циклофосфамид, тамоксифен, S-метадонp, бупропионр, эфавиренз). В основном CYP2В6 метаболизирует ксенобиотики. Маркёрный субстрат для CYP2В6 - антиконвульсант.
S-мефенитоинp при этом CYP2В6 подвергает S-мефенитоинp N-деметилированию (определяемый метаболит - N-деметилмефени- тоин). CYP2В6 принимает участие в метаболизме эндогенных стероидов: катализирует 16α-16β-гидроксилирование тестостерона.
Подсемейство цитохрома Р-450 CYPIIU
Из всех изоферментов подсемейства цитохрома CYPIIC наиболее важную роль в метаболизме ЛС играют изоферменты цитохрома Р-450 2С8, 2С9, 2С19. Общее свойство цитохромов подсемейства CYPIIC - 4-гидроксилазная активность по отношению к мефенитоинур (противосудорожное ЛС). Мефенитоинр - маркёрный субстрат изоферментов подсемейства CYPIIC. Именно поэтому изоферменты подсемейства CYPIIC называют ещё мефенитоин-4-гидроксилазами.
Изофермент цитохрома Р-450 2С8 (CYP2C8) принимает участие в метаболизме целого ряда лекарственных веществ (НПВС, статины и другие средства). Для многих ЛС CYP2C8 - «альтернативный» путь биотрансформации. Однако для таких препаратов, как репаглинид (гипогликемическое ЛС, принимаемое внутрь) и таксол (цитостатик), CYP2С8 - основной фермент метаболизма. CYP2С8 катализирует реакцию 6а-гидроксилирования таксола. Маркёрный субстрат CYP2С8 - паклитаксел (цитостатический препарат). В ходе взаимодействия паклитаксела с CYP2С8 происходит 6-гидроксилирование цитостатика.
Изофермент цитохрома Р-450 2С9 (CYP2C9) содержится, в основном, в печени. CYP2С9 отсутствует в фетальной печени, его обнаруживают только через месяц после рождения. Активность CYP2С9 не меняется в течение всей жизни. CYP2С9 метаболизирует различные лекарственные вещества. CYP2С9 - главный фермент метаболизма
многих НПВС, в том числе селективных ингибиторов циклоокси- геназы-2, ингибиторов ангиотензиновых рецепторов (лозартана и ирбесартана), гипогликемических препаратов (производных сульфонилмочевины), фенитоина (дифенина♠), непрямых антикоагулянтов (варфарина1, аценокумарола2), флувастатина3.
Следует отметить, что CYP2С9 имеет «стереоселективность» и метаболизирует в основном S-варфарин и S-аценокумарол, в то время как биотрансформация R-варфарина и R-аценокумарола происходит при помощи других изоферментов цитохрома Р-450: CYP1A2, CYP3A4. Индукторы CYP2С9 - рифампицин и барбитураты. Следует отметить, что практически все сульфаниламидные антибактериальные препараты ингибируют CYP2С9. Однако обнаружен специфический ингибитор CYP2С9 - сульфафеназолр. Существуют данные, что экстракт эхинацеи, пурпурной ингибирует CYP2С9 в исследованиях in vitro и in vivo, а гидролизованный экстракт сои (за счёт содержащихся в нём изофлавонов) ингибирует данный изофермент in vitro. Совместное применение ЛС-субстратов CYP2С9 с его ингибиторами приводит к угнетению метаболизма субстратов. В результате могут возникнуть нежелательные лекарственные реакции субстратов CYP2С9 (вплоть до интоксикации). Например, совместное применение варфарина (субстрат CYP2С9) с сульфаниламидными препаратами (ингибиторы CYP2С9) приводит к усилению антикоагулянтного эффекта варфарина. Именно поэтому при сочетании варфарина с сульфаниламидами рекомендуют выполнять строгий (по крайней мере, 1-2 раза в неделю) контроль международного нормализованного отношения. CYP2С9 обладает генетическим полиморфизмом. «Медленные» аллельные варианты CYP2C9*2 и CYP2C9*3- однонуклеотидные полиморфизмы гена CYP2C9, изученные в настоящее время наиболее полно. У носителей аллельных вариантов CYP2C9*2 и CYP2C9*3 отмечают снижение активности CYP2C9; это приводит к снижению скорости биотрансформации ЛС, метаболизирующихся данным изоферментом и к повышению их концентрации в плазме
1 Варфарин - рацематическая смесь изомеров: S-варфарина и R-вафрарина. Следует отметить, что большей антикоагулянтной активностью обладает S-варфарин.
2 Аценокумарол - рацематическая смесь изомеров: S-аценокумарола и R-ацено- кумарола. Однако, в отличие от варфарина, эти два изомера обладают одинаковой антикоагулянтной активностью.
3 Флувастатин - единственное ЛС из группы гиполипидемических препаратов ингибиторов ГМГ-КоА-редуктазы, метаболизм которого происходит при участии CYP2C9, а не CYP3A4. При этом CYP2C9 метаболизирует оба изомера флувастатина: активный (+)-3R,5S-энантиомер и неактивный (-)-3S,5R-энантиомер.
крови. Поэтому гетерозиготы (CYP2C9*1/*2, CYP2C9*1/*3) и гомозиготы (CYP2C9*2/*2, CYP2C9*3/*3, CYP2C9*2/*3) - «медленные» метаболизаторы по CYP2C9. Так, именно у этой категории пациентов (носителей перечисленных аллельных вариантов гена CYP2C9) наиболее часто отмечают нежелательные лекарственные реакции при применении ЛС, метаболизм которых происходит под влиянием CYP2C9 (непрямые антикоагулянты, НПВС, применяемые внутрь гипогликемические ЛС - производные сульфонилмочевины).
Изофермент цитохрома Р-450 2С18 (CYP2C18) содержится в основном в печени. CYP2Cl8 отсутствует в фетальной печени, его обнаруживают только через месяц после рождения. Активность CYP2Cl8 не изменяется в течение всей жизни. CYP2Cl8 вносит определённый вклад в метаболизм таких ЛС, как напроксен, омепразол, пироксикам, пропранолол, изотретиноин (ретиноевая кислота) и варфарин.
Изофермент цитохрома Р-450 2С19 (CYP2C19) - основной фермент метаболизма ингибиторов протонного насоса. При этом метаболизм отдельных ЛС из группы ингибиторов протонного насоса имеет свои особенности. Так, у омепразола обнаружили два пути метаболизма.
• Под действием CYP2C19 омепразол превращается в гидроксиомепразол. Под действием CYP3A4 гидроксиомепразол переходит в омепразол гидроксисульфон.
• Под действием CYP3A4 омепразол превращается в сульфид омепразола и сульфон омепразола. Под действием CYP2C19 сульфид омепразола и сульфон омепразола переходят в омепразол гидроксисульфон.
Таким образом, вне зависимости от пути биологической трансформации, конечный метаболит омепразола - омепразол гидроксисульфон. Однако следует отметить, что указанные пути метаболизма характерны, прежде всего, для R-изомера омепразола (S-изомер в значительно меньшей степени подвергается биотрансформации). Понимание этого феномена позволило создать эзопразолр - препарат, представляющий S-изомер омепразола (ингибиторы и индукторы CYP2C19, а также генетический полиморфизм этого изофермента в меньшей степени влияют на фармакокинетику эзопразолар).
Метаболизм лансопразола идентичен метаболизму омепразола. Рабепразол метаболизируется при участии CYP2C19 и CYP3A4 до диметилрабепразола и рабепразол сульфона соответственно.
CYP2C19 участвует в метаболизме тамоксифена, фенитоина, тиклопидина, таких психотропных ЛС, как трициклические антидепрессанты, диазепам, некоторые барбитураты.
Для CYP2C19 характерен генетический полиморфизм. Медленные метаболизаторы по CYP2Cl9 - носители «медленных» аллельных вариантов. Применение у медленных метаболизаторов по CYP2CL9 препаратов - субстратов этого изофермента приводит к более частому возникновению нежелательных лекарственных реакций, особенно при использовании препаратов с узкой терапевтической широтой: трициклических антидепрессантов, диазепама, некоторых барбитуратов (мефобарбитала, гексобарбитала). Однако наибольшее количество исследований посвящено влиянию полиморфизма гена CYP2C19 на фармакокинетику и фармакодинамику блокаторов ингибиторов протонного насоса. Как показали фармакокинетические исследования, проведённые при участии здоровых добровольцев, площадь под фармакокинетической кривой, значения максимальной концентрации омепразола, лансопразола и рабепразола достоверно выше у гетерозигот и, особенно, у гомозигот по «медленным» аллельным вариантам гена CYP2C19. Кроме того, более выраженное подавление желудочной секреции при применении омепразола, лансорпразола, рабепразола наблюдали у пациентов (гетерозиготы и гомозиготы по «медленным» аллельным вариантам CYP2C19), страдающих язвенной болезнью и рефлюкс-эзофагитом. Однако частота нежелательных лекарственных реакций ингибиторов протонного насоса не зависит от генотипа по CYP2C19. Существующие данные позволяют предположить, что для достижения «целевого» подавления желудочной секреции у гетерозигот и гомозигот по «медленным» аллельным вариантам гена CYP2C19 необходимы меньшие дозы ингибиторов протонного насоса.
Подсемейство цитохрома Р-450 CYPIID
В состав подсемейства цитохрома Р-450 CYPIID входит единственный изофермент - 2D6 (CYP2D6).
Изофермент цитохрома Р-450 2D6 (CYP2D6) обнаруживают в основном в печени. CYP2D6 метаболизирует около 20% всех известных ЛС, в том числе нейролептики, антидепрессанты, транквилизаторы, β-адреноблокаторы. Доказано: CYP2D6 - главный фермент биотрансформации и трициклического антидепрессанта амитриптилина. Однако, как показали исследования, незначительная часть амитриптилина метаболизируется и другими изоферментами цитохрома Р-450 (CYP2C19, CYP2C9, CYP3A4) до неактивных метаболитов. Дебризохинр, декстрометорфан и спартеин - маркёрные субстраты, используемые для фенотипирования изофермента 2D6. CYP2D6, в отличие от других изоферментов цитохрома Р-450, не имеет индукторов.
Ген CYP2D6 обладает полиморфизмом. Еще в 1977 году Iddle и Mahgoub обратили внимание на различие гипотензивного эффекта у больных артериальной гипертензией, применявших дебризохинр (препарат из группы α-адреноблокаторов). Тогда же сформулировали предположение о различии скорости метаболизма (гидроксилирования) дебризохинар у разных индивидуумов. У «медленных» метаболизаторов дебризохинар зарегистрировали наибольшую выраженность гипотензивного эффекта данного препарата. Позднее доказали, что у «медленных» метаболизаторов дебризохинар замедлен метаболизм и некоторых других ЛС, в том числе фенацетина, нортриптилинар, фенформинар, спартеина, энкаинидар, пропранолола, гуаноксанар и амитриптилина. Как показали дальнейшие исследования, «медленные» метаболизаторы по CYP2D6 - носители (как гомозиготы, так и гетерозиготы) функционально дефектных аллельных вариантов гена CYP2D6. Результат этих вариантов - отсутствие синтеза CYP2D6 (аллельный вариант CYP2D6x5), синтез неактивного белка (аллельные варианты CYP2D6x3, CYP2D6x4, CYP2D6x6, CYP2D6x7, CYP2D6x8, CYP2D6x11, CYP2D6x12, CYP2D6x14, CYP2D6x15, CYP2D6x19, CYP2D6x20), синтез дефектного белка со сниженной активностью (варианты CYP2D6x9, CYP2D6x10, CYP2D6x17,
CYP2D6x18, CYP2D6x36). С каждым годом растёт количество найденных аллельных вариантов гена CYP2D6 (их носительство приводит к изменению активности CYP2D6). Однако ещё Saxena (1994) указывал, что 95% всех «медленных» метаболизаторов по CYP2D6 - носители вариантов CYP2D6x3, CYP2D6x4, CYP2D6x5, остальные варианты обнаруживают гораздо реже. По данным Rau и соавт. (2004), частота аллельного варианта CYP2D6x4 среди пациентов, у которых наблюдали нежелательные лекарственные реакции на фоне приёма трициклических антидепрессантов (артериальная гипотензия, седативный эффект, тремор, кардиотоксичность), почти в 3 раза (20%) превышает таковую у пациентов, при лечении которых указанными препаратами осложнений не регистрировали (7%). Аналогичное влияние генетического полиморфизма CYP2D6 обнаружили и на фармакокинетику и фармакодинамику нейролептиков, в результате продемонстрировали наличие ассоциаций между носительством некоторых аллельных вариантов гена CYP2D6 и развитием индуцированных нейролептиками экстрапирамидных нарушений.
Однако носительство «медленных» аллельных вариантов гена CYP2D6 может сопровождаться не только увеличением риска развития нежелательных лекарственных реакций при применении препа-
ратов, метаболизирующихся данным изоферментом. Если ЛС - пролекарство, а активный метаболит образуется именно под действием CYP2D6, то у носителей «медленных» аллельных вариантов отмечают низкую эффективность препарата. Так, у носителей «медленных» аллельных вариантов гена CYP2D6 регистрируют менее выраженный анальгезирующий эффект кодеина. Этот феномен объясняют снижением О-деметилирования кодеина (в ходе указанного процесса образуется морфин). Обезболивающее действие трамадола также обусловлено активным метаболитом О-деметилтрамадолом (образуется под действием CYP2D6). У носителей «медленных» аллелельных вариантов гена CYP2D6 отмечают значительное уменьшение синтеза О-деметилтрамадола; это может привести к недостаточному анальгезирующему эффекту (аналогично процессам, возникающим при использовании кодеина). Так, Stamer и соавт. (2003), изучив анальгезирующий эффект трамадола у 300 пациентов, перенёсших операции на брюшной полости, обнаружили, что гомозиготы по «медленным» аллельным вариантам гена CYP2D6 не «отвечали» на терапию трамадолом в 2 раза чаще, чем пациенты, не несущие данных аллелей (46,7% против 21,6% соответственно, р=0,005).
В настоящее время выполнили множество исследований, посвя- щённых влиянию генетического полиморфизма CYP2D6 на фармакокинетику и фармакодинамику β-адреноблокаторов. Результаты этих исследований имеют клиническое значение для индивидуализации фармакотерапии данной группой ЛС.
Подсемейство цитохрома Р-450 CYPIIB
Из изоферментов подсемейства цитохрома IIE наиболее важную роль в метаболизме ЛС играет изофермент цитохрома Р-450 2Е1. Общее свойство изоферментов подсемейства CYPIIE - способность к индукции под влиянием этанола. Именно поэтому второе название подсемейства CYPIIE - этанол-индуцибельные цитохромы.
Изофермент цитохрома Р-450 2Е1 (CYP2E1) содержится в печени взрослых. CYP2E1 составляет около 7% всех изоферментов цитохрома Р-450. Субстраты CYP2E1 - малое количество ЛС, а также некоторые другие ксенобиотики: этанол, нитрозоамины, «небольшие» ароматические углеводороды типа бензола и анилина, алифатические хлоруглеводороды. CYP2E1 катализирует превращение дапсона в гидроксиламиндапсон, n1-деметилирование и N7-деметилирование кофеина, дегалогенизацию хлорфторуглеводородов и средств для ингаляционного наркоза (галотан) и некоторые другие реакции.
CYP2E1 вместе с CYP1A2 катализируют важную реакцию превращения парацетамола (ацетаминофена) в N-ацетилбензохинонимин, обладающий мощным гепатотоксическим действием. Существуют данные об участии цитохрома CYP2E1 ватерогенезе. Например, известно, что CYP2E1 - наиболее важный изофермент цитохрома Р-450, окисляющий холестерин липопротеидов низкой плотности (ЛНП). В процессе окисления ЛНП также принимают участие цитохромы и другие изоферменты цитохрома Р-450, а также 15-липооксигеназа и НАДФ-Н-оксидазы. Продукты окисления: 7а-гидроксихолестерол, 7β -гидроксихолестерол, 5β -6β -эпоксихолестерол, 5 α-6β -эпоксихолестерол, 7-кетохолестерол, 26-гидроксихолестерол. Процесс окисления ЛНП происходит в эндотелиоцитах, гладкой мускулатуре кровеносных сосудов, макрофагах. Окисленные ЛНП стимулируют формирование пенистых клеток и таким образом способствуют образованию атеросклеротических бляшек.
Подсемейство цитохрома Р-450 CYPIIIA
Подсемейство цитохрома Р-450 CYPIIIA включает четыре изофермента: 3А3, 3А4, 3А5 и 3А7. Цитохромы подсемейства IIIA составляют 30% всех изоферментов цитохрома Р-450 в печени и 70% всех изоферментов стенки пищеварительного тракта. При этом в печени преимущественно локализован изофермент 3А4 (CYP3A4), в стенках желудка и кишечника - изоферменты 3А3 (CYP3A3) и 3А5 (CYP3A5). Изофермент 3А7 (CYP3A7) обнаруживают только в печени плода. Из изоферментов подсемейства IIIA наиболее важную роль в метаболизме ЛС играет CYP3A4.
Изофермент цитохрома Р-450 3А4 (CYP3A4) метаболизирует около 60% всех известных ЛС, в том числе блокаторы медленных кальциевых каналов, макролидные антибиотики, некоторые антиаритмики, статины (ловастатин, симвастатин, аторвастатин), клопидогрел1 и другие препараты.
CYP3A4 катализирует реакцию 6β-гидроксилирования эндогенных стероидов, в том числе тестостерона, прогестерона, кортизолаp. Маркёрные субстраты для определения активности CYP3A4 - дапсон, эритромицин, нифедипин, лидокаин, тестостерон и кортизолp.
Метаболизм лидокаина протекает в гепатоцитах, где через оксидативное N-деэтилирование CYP3A4 образуется моноэтилглицинксилидид (MEGX).
1 Клопидогрел - пролекарство, под действием CYP3A4 он превращается в активный метаболит, обладающий антиагрегантным действием.
Определение активности CYP3A4 по MEGX (метаболит лидокаина) - наиболее чувствительный и специфичный тест, позволяющий оценить функциональное состояние печени при острых и хронических её заболеваниях, а также при синдроме системного воспалительного ответа (сепсисе). При циррозе печени концентрация MEGX коррелирует с прогнозом заболевания.
В литературе существуют данные о внутривидовой вариабельности метаболизма ЛС под действием CYP3A4. Однако молекулярные подтверждения генетического полиморфизма CYP3A4 появились только в последнее время. Так, A. Lemoin и соавт. (1996) описали случай интоксикации такролимусом (субстрат CYP3A4) у больного после пересадки печени (в клетках печени активность CYP3A4 обнаружить не удалось). Только после обработки клеток пересаженной печени глюкокортикоидами (индукторы CYP3A4) можно определить активность CYP3A4. Существует предположение, что нарушение экспрессии факторов транскрипции гена, кодирующего CYP3A4, - причина вариабельности метаболизма данного цитохрома.
Изофермент цитохрома Р-450 3А5 (CYP3A5), по последним данным, может играть существенную роль в метаболизме некоторых лекарственных веществ. Следует отметить, что CYP3A5 экспрессируется в печени 10-30% взрослых людей. У этих индивидуумов вклад CYP3A5 в активность всех изоферментов подсемейства IIIA составляет от 33 (у европейцев) до 60% (у афроамериканцев). Как показали исследования, под влиянием CYP3A5 происходят процессы биотрансформации тех ЛС, которые традиционно рассматривают как субстраты CYP3A4. Следует отметить, что индукторы и ингибиторы CYP3A4 обладают аналогичным действием и в отношении CYP3A5. Активность CYP3A5 у различных индивидуумов варьирует более чем в 30 раз. Впервые различия активности CYP3A5 описали Paulussen и соавт. (2000): они наблюдали in vitro существенные различия скорости метаболизма мидазолама под влиянием CYP3A5.
Дигидропиримидин дегидрогеназа
Физиологическая функция дигидропиримидин дегидрогеназы (ДПДГ) - восстановление урацила и тимидина - первая реакция трёхэтапного метаболизма этих соединений до β-аланина. Кроме того, ДПДГ - основной фермент, метаболизирующий 5-фторура- цил. Указанный препарат применяют в составе комбинированной химиотерапии рака молочной железы, яичников, пищевода, желудка, толстой и прямой кишки, печени, шейки матки, вульвы. Также
5-фторурацил используют при лечении рака мочевого пузыря, простаты, опухолей головы, шеи, слюнных желёз, надпочечников, поджелудочной железы. В настоящее время известна аминокислотная последовательность и количество аминокислотных остатков (их всего 1025), входящих в состав ДПДГ; молекулярная масса фермента составляет 111 кД. Идентифицировали ген ДПДГ, локализованный в хромосоме 1 (локус 1р22). Цитоплазма клеток различных тканей и органов содержит ДПДГ, особенно большое количество фермента обнаруживают в клетках печени, в моноцитах, лимфоцитах, гранулоцитах, тромбоцитах. Однако в эритроцитах активность ДПДГ не отмечена (Van Kuilenburg и соавт., 1999). С середины 80-х годов появились сообщения о серьёзных осложнениях, возникающих при применении 5-фторурацила (причина осложнений - наследственно обусловленная низкая активность ДПДГ). Как показали Diasio и соавт. (1988), низкая активность ДПДГ наследуется по аутосомнорецессивному типу. Таким образом, ДПДГ - фермент, обладающий генетическим полиморфизмом. В будущем, по-видимому, произой- дёт внедрение методов фенотипирования и генотипирования ДПДГ в онкологическую практику для обеспечения безопасности химиотерапии 5-фторурацилом.
5.4. ФЕРМЕНТЫ II ФАЗЫ БИОТРАНСФОРМАЦИИ ЛЕКАРСТВЕННЫХ СРЕДСТВ
Глюкуронилтрансферазы
Глюкуронирование - наиболее важная реакция II фазы метаболизма лекарственных веществ. Глюкуронирование представляет присоединение (конъюгацию) к субстрату уридиндифосфат-глюкуроновой кислоты (УДФ-глюкуроновой кислоты). Эта реакция катализируется надсемейством ферментов, называемых «УДФ-глюкуронилтрансферазы» и обозначаемых как UGT. Надсемейство УДФ-глюкуронилтрансфераз включает два семейства и более двадцати изоферментов, локализованных в эндоплазматической системе клеток. Они катализируют глюкуронирование большого количества ксенобиотиков, включая ЛС и их метаболиты, пестициды и канцерогены. К соединениям, подвергающимся глюкуронированию, относят простые и сложные эфиры; соединения, содержащие карбоксильные, карбомоильные, тиольные и карбонильные группы, а также нитрогруппы. Глюкуронирование
приводит к увеличению полярности химических соединений, что облегчает их растворимость в воде и элиминацию. УДФ-глюкуронилтрансферазы обнаруживают у всех позвоночных животных: от рыб до человека. В организме новорождённых регистрируют низкую активность УДФ-глюкуронилтрансфераз, однако через 1-3 мес жизни активность данных ферментов можно сравнить с таковой у взрослых. УДФ-глюкуронилтрансферазы содержатся в печени, кишечнике, лёг- ких, головном мозге, обонятельном эпителии, почках, но печень - главный орган, в котором происходит глюкуронирование. Степень экспрессии различных изоферментов УДФ-глюкуронилтрансферазы в органах неодинакова. Так, изофермент УДФ-глюкуронилтрансферазы UGT1A1, катализирующий реакцию глюкуронирования билирубина, экспрессируется главным образом в печени, но не в почках. Изоферменты УДФ-глюкуронилтрансферазы UGT1A6 и UGT1A9, ответственные за глюкуронирование фенола, экспрессируются и в печени, и в почках одинаково. Как указывалось выше, по идентичности аминокислотного состава надсемейство УДФ-глюкуронилтрансфераз подразделяют на два семейства: UGT1 и UGT2. Изоферменты, семейства UGT1 сходны по аминокислотному составу на 62-80%, а изоферменты семейства UGT2 - на 57-93%. Изоферменты, входящие в состав семейств УДФ-глюкуронилтрансферазы человека, а также локализация генов и маркёрные субстраты изоферментов для фенотипирования представлены в таблице (табл. 5-7).
Физиологическая функция УДФ-глюкуронилтрансфераз - глюкуронирование эндогенных соединений. Продукт катаболизма гема - билирубин - наиболее хорошо изученный эндогенный субстрат УДФ-глюкуронилтрансферазы. Глюкуронирование билирубина предотвращает накопление токсичного свободного билирубина. При этом билирубин выделяется с желчью в виде моноглюкуронидов и диглюкуронидов. Другая физиологическая функция УДФ-глюкуронилтрансферазы - участие в метаболизме гормонов. Так, тироксин и трийодтиронин подвергаются глюкуронированию в печени и выводятся в виде глюкуронидов с желчью. УДФ-глюкуронилтрансферазы также участвуют в метаболизме стероидных гормонов, желчных кислот, ретиноидов, однако эти реакции в настоящее время изучены недостаточно.
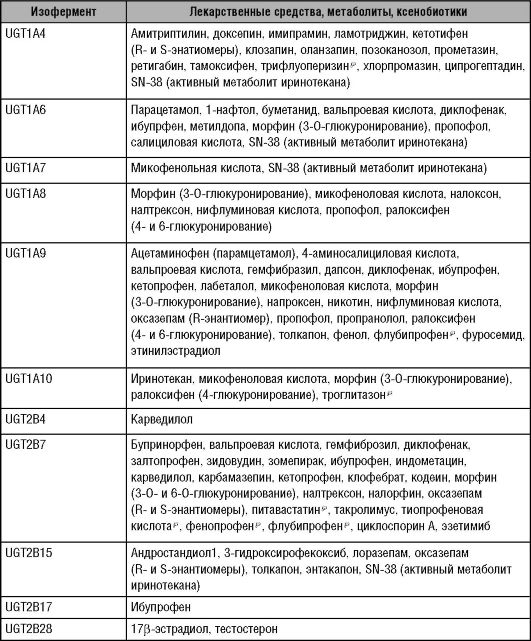
Глюкуронированию подвергаются ЛС разных классов, многие из них имеют узкую терапевтическую широту, например, морфин и хлорамфеникол (табл. 5-8).
Таблица 5-7. Состав семейств УДФ-глюкуронилтрансферазы человека, локализация генов и маркёрные субстраты изоферментов
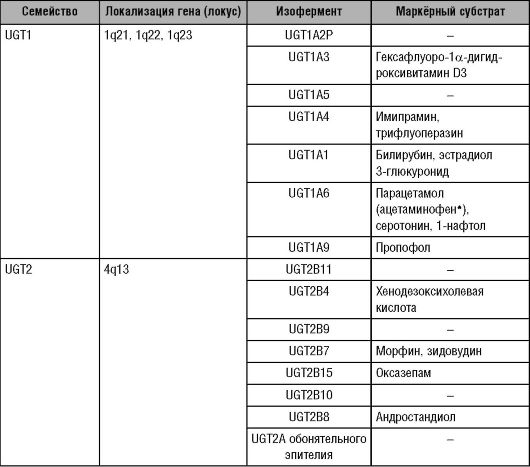 Таблица 5-8. Лекарственные
средства, метаболиты и ксенобиотики, подвергающиеся глюкоуронированию
различными изоферментами УДФ-глюкуронилтрансферазы
Таблица 5-8. Лекарственные
средства, метаболиты и ксенобиотики, подвергающиеся глюкоуронированию
различными изоферментами УДФ-глюкуронилтрансферазы
 Окончание таблицы 5-8
Окончание таблицы 5-8
 Лекарственные средства (представители разных химических групп), подвергающиеся глюкуронированию
Лекарственные средства (представители разных химических групп), подвергающиеся глюкуронированию
• Фенолы: пропофол, ацетаминофен, налоксон.
• Спирты: хлорамфеникол, кодеин, оксазепам.
• Алифатические амины: циклопироксоламинp, ламотриджин, амитриптилин.
• Карбоновые кислоты: ферпазонp, фенилбутазон, сульфинпиразон.
• Карбоксильные кислоты: напроксен, зомепиралp, кетопрофен. Таким образом, глюкуронированию подвергаются соединения,
содержащие разные функциональные группы, выполняющие функции акцепторов для УДФ-глюкуроновой кислоты. Как указывалось выше, в результате глюкуронирования образуются полярные неактивные метаболиты, легко выводящиеся из организма. Однако существует пример, когда в результате глюкуронирования образуется активный метаболит. Глюкуронирование морфина приводит к образованию морфин-6-глюкуронида, обладающего значительным анальгезирующим эффектом и реже, чем морфин, вызывающего тошноту и рвоту. Также глюкуронирование может способствовать биологической активации канцерогенов. К канцерогенным глюкуронидам относят N-глюкуронид 4-аминобифенила, N-глюкуронид N-ацетил- бензидина,О-глюкуронид-4-((гидрокисметил)-нитрозоамино)-1-(3-пи- ридил)-1-бутанона.
Давно известно о существовании наследственных нарушений глюкуронирования билирубина. К ним относят синдром Жильбера и синдром Криглера-Найяра. Синдром Жильбера - наследственное заболевание, наследуемое по аутосомно-рецессивному типу. Распространённость синдрома Жильбера среди населения составляет 1-5%. Причина развития данного заболевания - точечные мутации (как правило, замены в нуклеотидной последовательности) в гене UGT1. При этом образуется УДФ-глюкуронилтрансфераза, характеризующаяся малой активностью (25-30% от нормального уровня). Изменение глюкуронирования лекарственных веществ у больных с синдромом Жильбера изучено мало. Существуют данные о снижении клиренса толбутамида, парацетамола (ацетаминофена♠) и рифампинаp у больных с синдромом Жильбера. Изучали частоту побочных эффектов нового цитостатического препарата иринотекана у пациентов, страдающих одновременно колоректальным раком и синдромом Жильбера и у больных колоректальным раком. Иринотекан (СТР-11) - новый высокоэффективный препарат, обладающий цитостатическим действием, ингибирующий топоизомеразу I и применяемый при колоректальном раке при наличии резистентности к фторурацилу. Иринотекан в печени под действием карбоксиэстераз превращает-
ся в активный метаболит 7-этил-10-гидроксикамптотекин (SN-38). Главный путь метаболизма SN-38 - глюкуронирование с помощью UGT1A1. В ходе исследований побочные эффекты иринотекана (в частности, диарею) достоверно чаще регистрировали у больных, имеющих синдром Жильбера. Учёные доказали: носительство аллельных вариантов UGT1A1x1B, UGT1A1x26, UGT1A1x60 ассоциируется с более частым развитием гипербилирубинемии при применении иринотекана, при этом регистрировали низкие значения площади под фармакокинетической кривой глюкуронида SN-38. В настоящее время Американским управлением по контролю над пищевыми продуктами и медикаментами (Food and drug administration - FDA) одобрено определение аллельных вариантов гена UGT1A1 для выбора режима дозирования иринотекана. Существуют данные о влиянии носительства аллельных вариантов генов, кодирующих и другие изоформы UGT, на фармакокинетику и фармакодинамику различных ЛС.
Ацетилтрансферазы
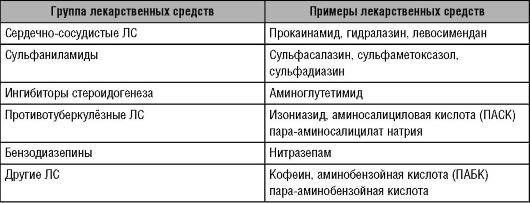
Ацетилирование эволюционно представляет один из самых ранних механизмов адаптации. Реакция ацетилирования необходима для синтеза жирных кислот, стероидов, функционирования цикла Кребса. Важная функция ацетилирования - метаболизм (биотрансформация) ксенобиотиков: ЛС, бытовых и промышленных ядов. На процессы ацетилирования влияет N-ацетилтрансфераза, а также кофермент А. Контроль интенсивности ацетилирования в организме человека происходит при участии β2-адренорецепторов и зависит от метаболических резервов (пантотеновой кислоты, пиридоксина, тиамина, липоевой кислоты*) и генотипа. Кроме того, интенсивность ацетилирования зависит от функционального состояния печени и других органов, содержащих N-ацетилтрансферазу (хотя ацетилирование, как и другие реакции II фазы, мало изменяется при заболеваниях печени). Между тем ацетилирование ЛС и других ксенобиотиков происходит преимущественно в печени. Выделено два изофермента N-ацетилтрансферазы: N-ацетилтрансфераза 1 (NAT1) и N-ацетилтрансфераза 2 (NAT2). NAT1 ацетилирует небольшое количество ариламинов и не обладает генетическим полиморфизмом. Таким образом, основной фермент ацетилирования - NAT2 . Ген NAT2 расположен в хромосоме 8 (локусы 8р23.1, 8р23.2 и 8р23.3). NAT2 ацетилирует различные ЛС, в том числе изониазид и сульфаниламиды (табл. 5-9).
Таблица 5-9. Лекарственные средства, подвергающиеся ацетилированию
 Наиболее
важным свойством NAT2 считают генетический полиморфизм. Впервые
полиморфизм ацетилирования описал в 1960-е годы Evans; он выделил
медленные и быстрые ацетиляторы изониазида. Также отметили, что у
«медленных» ацетиляторов, в связи с накоплением (кумуляцией) изониазида,
чаще возникают полиневриты. Так, у «медленных» ацетиляторов период
полувыведения изониазида составляет 3 ч, в то время как у «быстрых»
ацетиляторов - 1,5 ч. Развитие полиневритов обусловлено влиянием
изониазида: препарат тормозит переход пиридоксина (витамина В6)
в активный кофермент дипиридоксинфосфат, необходимый для синтеза
миелина. Предполагали, что у «быстрых» ацетиляторов применение
изониазида с большей вероятностью приведёт к развитию гепатотоксического
эффекта из-за более интенсивного образования ацетилгидразина, однако
практического подтверждения это предположение не получило.
Индивидуальная скорость ацетилирования существенно не влияет на режим
дозирования препарата при ежедневном приёме, но может уменьшать
эффективность терапии при периодическом использовании изониазида.
Проанализировав результаты лечения изониазидом 744 больных туберкулёзом,
выяснили, что у «медленных» ацетиляторов закрытие полостей в лёгких
происходит быстрее. Как показало исследование, проведённое Sunahara в
1963 году, «медленные» ацетиляторы - гомозиготы по «медленной» аллели
NAT2, а «быстрые» метаболизаторы - гомозиготы либо гетерозиготы по
«быстрой» аллели NAT2. В 1964 году Evans опубликовал данные о том, что
полиморфизм ацетилирования характерен не только для изониазида, но и для
гидралазина и сульфаниламидов. Затем наличие полиморфизма ацетили-
Наиболее
важным свойством NAT2 считают генетический полиморфизм. Впервые
полиморфизм ацетилирования описал в 1960-е годы Evans; он выделил
медленные и быстрые ацетиляторы изониазида. Также отметили, что у
«медленных» ацетиляторов, в связи с накоплением (кумуляцией) изониазида,
чаще возникают полиневриты. Так, у «медленных» ацетиляторов период
полувыведения изониазида составляет 3 ч, в то время как у «быстрых»
ацетиляторов - 1,5 ч. Развитие полиневритов обусловлено влиянием
изониазида: препарат тормозит переход пиридоксина (витамина В6)
в активный кофермент дипиридоксинфосфат, необходимый для синтеза
миелина. Предполагали, что у «быстрых» ацетиляторов применение
изониазида с большей вероятностью приведёт к развитию гепатотоксического
эффекта из-за более интенсивного образования ацетилгидразина, однако
практического подтверждения это предположение не получило.
Индивидуальная скорость ацетилирования существенно не влияет на режим
дозирования препарата при ежедневном приёме, но может уменьшать
эффективность терапии при периодическом использовании изониазида.
Проанализировав результаты лечения изониазидом 744 больных туберкулёзом,
выяснили, что у «медленных» ацетиляторов закрытие полостей в лёгких
происходит быстрее. Как показало исследование, проведённое Sunahara в
1963 году, «медленные» ацетиляторы - гомозиготы по «медленной» аллели
NAT2, а «быстрые» метаболизаторы - гомозиготы либо гетерозиготы по
«быстрой» аллели NAT2. В 1964 году Evans опубликовал данные о том, что
полиморфизм ацетилирования характерен не только для изониазида, но и для
гидралазина и сульфаниламидов. Затем наличие полиморфизма ацетили-
рования доказали и для других лекарственных средств. Применение прокаинамида и гидралазина у «медленных» ацетиляторов гораздо чаще вызывает поражение печени (гепатотоксичность), таким образом, и для этих препаратов характерен полиморфизм ацетилирования. Однако в случае с дапсоном (также подвергается ацетилированию) не удалось обнаружить различий в частоте возникновения волчаночноподобного синдрома при применении данного препарата «медленными» и «быстрыми» ацетиляторами. Распространённость «медленных» ацетиляторов варьирует: от 10-15% среди японцев и китайцев до 50% среди представителей европеоидной расы. Только в конце 80-х годов начали идентифицировать аллельные варианты гена NAT2, носительство которых обусловливает медленное ацетилирование. В настоящее время известно около 20 мутантных аллелей гена NAT2. Все указанные аллельные варианты наследуются по аутосомно-рецессивному типу.
Тип ацетилирования определяют, используя методы фенотипирования и генотипирования NAT2. В качестве маркёрных субстратов ацетилирования используют дапсон, изониазид и сульфадимин (сульфадимезин*). Отношение концентрации моноацетилдапсона к концентрации дапсона менее 0,35 в плазме крови через 6 ч после введения препарата характерно для «медленных» ацетиляторов, а более 0,35 - для «быстрых» ацетиляторов. Если в качестве маркерного субстрата используют сульфадимин, то наличие менее 25% сульфадимина в плазме крови (анализ выполняют через 6 ч) и менее 70% в моче (собранной через 5-6 ч после введения препарата) свидетельствует о фенотипе «медленного» ацетилирования.
Тиопурин S-метилтрансфераза
Тиопурин S-метилтрансфераза (ТРМТ) - фермент, катализирующий реакцию S-метилирования производных тиопурина - основной путь метаболизма цитостатических веществ из группы антагонистов пурина: 6-меркаптопурина, 6-тиогуанина, азатиоприна. 6-мер- каптопурин используют в составе комбинированной химиотерапии миелобластного и лимфобластного лейкоза, хронического миелолейкоза, лимфосаркомы, саркомы мягких тканей. При острых лейкозах применяют, как правило, 6-тиогуанин. В настоящее время известна аминокислотная последовательность и количество аминокислотных остатков, входящих в состав ТРМТ, - 245. Молекулярная масса ТРМТ составляет 28 кДа. Также идентифицировали ген ТРМТ, локализованный в хромосоме 6 (локус 6q22.3). ТРМТ располагается в цитоплазме кроветворных клеток.
В 1980 году Weinshiboum изучал активность ТРМТ у 298 здоровых добровольцев и обнаружил значительные различия активности ТРМТ у людей: 88,6% обследованных имели высокую активность ТРМТ, 11,1% - промежуточную. Низкую активность ТРМТ (или полное отсутствие активности фермента) зарегистрировали у 0,3% обследованных добровольцев. Так впервые описали генетический полиморфизм ТРМТ. Как показали более поздние исследования, для людей с низкой активностью ТРМТ характерна повышенная чувствительность к 6-меркаптопурину, 6-тиогуанину и азатиоприну; при этом развиваются опасные для жизни гематотоксические (лейкопения, тромбоцитопения, анемия) и гепатотоксические осложнения. В условиях низкой активности ТРМТ, метаболизм 6-меркаптопурина идёт по альтернативному пути - до высокотоксичного соединения 6-тио- гуанина нуклеотида. Lennard и соавт. (1990) изучали концентрацию 6-тиогуанина нуклеотида в плазме крови и активность ТРМТ в эритроцитах 95 детей, получавших 6-меркаптопурин по поводу острого лимфобластного лейкоза. Авторы выяснили: чем меньше активность ТРМТ, тем выше концентрации 6-TGN в плазме крови и тем более выражены побочные эффекты 6-меркаптопурина. В настоящее время доказано, что низкая активность ТРМТ наследуется по аутосомнорецессивному типу, причём у гомозигот регистрируют низкую активность ТРМТ, а у гетерозигот - промежуточную. Генетические исследования последних лет, выполненные методом полимеразной цепной реакции, позволили обнаружить мутации гена ТРМТ, определяющие низкую активность данного фермента. Безопасные дозы 6-меркапто- пурина: при высокой активности ТРМТ (нормальный генотип) назначают по 500 мг/(м2×сутки), при промежуточной активности ТРМТ (гетерозиготы) - по 400 мг/(м2×сутки), при медленной активности ТРМТ (гомозиготы) - по 50 мг/(м2×сутки).
Сульфотрансферазы
Сульфатирование - реакция присоединения (конъюгации) к субстрату остатка серной кислоты, при этом образуются сложные эфиры серной кислоты или сульфоматы. Сульфатированию в организме человека подвергаются экзогенные соединения (в основном фенолы) и эндогенные соединения (гормоны щитовидной железы, катехоламины, некоторые стероидные гормоны). В качестве кофермента реакции сульфатирования выступает 3'-фосфоаденилсульфат. Затем происходит превращение 3'-фосфоаденилсульфата в аденозин- 3',5'-бифосфонат. Реакция сульфатирования катализируется над-
семейством ферментов, называемых «сульфотрансферазы» (SULT). Сульфотрансферазы локализуются в цитозоле. В организме человека обнаружили три семейства. В настоящее время идентифицировали около 40 изоферментов сульфотрансферазы. Изоферменты сульфотрансферазы в организме человека кодируются, по крайней мере, 10 генами. Наибольшая роль в сульфатировании лекарственных веществ и их метаболитов принадлежит изоферментам сульфотрансферазы семейства 1 (SULT1). SULT1A1 и SULT1A3 - самые важные изоферменты данного семейства. Изоферменты SULT1 локализованы главным образом в печени, а также толстой и тонкой кишке, лёгких, головном мозге, селезёнке, плаценте, лейкоцитах. Изоферменты SULT1 имеют молекулярную массу около 34 кДа и состоят из 295 аминокислотных остатков, ген изоферментов SULT1 локализован в 16 хромосоме (локус 16р11.2). SULT1A1 (термостабильная сульфотрансфераза) катализирует сульфатирование «простых фенолов», в том числе лекарственных веществ фенольной структуры (миноксидилр, ацетаминофен, морфин, салициламид, изопреналин и некоторые другие). Следует отметить, что сульфатирование миноксидилар приводит к образованию его активного метаболита - миноксидила сульфата. SULT1A1 сульфатирует метаболиты лидокаина: 4-гидрокси-2,6-ксилидин(4-гидроксил) и ропивакаина: 3-гид- роксиропивакаин, 4-гидроксиропивакаин, 2-гидроксиметилропива- каин. Кроме того, SULT1A1 сульфатирует 17β-эстрадиол. Маркёрный субстрат SULT1A1 - 4-нитрофенол. SULT1A3 (термолабильная сульфотрансфераза) катализирует реакции сульфатирования фенольных моноаминов: дофамина, норадреналина, серотонина. Маркёрный субстрат SULT1A3 - дофамин. Изоферменты сульфотрансферазы семейства 2 (SULT2) обеспечивают сульфатирование дигидроэпиандростерона, эпиандростерона, андростерона. Изоферменты SULT2 участвуют в биологической активации канцерогенов, например, ПАУ (5-гидроксиметилхризен, 7,12-дигидроксиметилбенз[а]антрацен), N-гидрокси-2-ацетиламинофлуорен. Изоферменты сульфотрансферазы семейства 3 (SULT3) катализируют N-сульфатирование ациклических ариламинов.
Эпоксидгидролаза
Водная конъюгация играет важную роль в детоксикации и биологической активации большого количества ксенобиотиков, таких, как арены, алифатические эпоксиды, ПАУ, афлотоксин В1. Реакции водной конъюгации катализирует особый фермент - эпоксидгидролаза
(ЕРНХ). Наибольшее количество данного фермента обнаружено в печени. Учёные выделили две изоформы эпоксидгидролазы: ЕРНХ1 и ЕРНХ2. ЕРНХ2 состоит из 534 аминокислотных остатков, имеет молекулярную массу 62 кДа; ген ЕРНХ2 располагается в хромосоме 8 (локус 8р21-р12). ЕРНХ2 локализуется в цитоплазме и пероксисомах; данная изоформа эпоксидгидролазы играет небольшую роль в метаболизме ксенобиотиков. Большую часть реакций водной конъюгации катализирует ЕРНХ1. ЕРНХ1 состоит из 455 аминокислотных остатков, имеет молекулярную массу 52 кДа. Ген ЕРНХ1 располагается в хромосоме 1 (локус 1q42.1). Велико значение ЕРНХ1 в водной конъюгации токсических метаболитов лекарственных веществ. Противосудорожное средство фенитоин окисляется цитохромом Р-450 до двух метаболитов: парагидроксилатеда и дигидродиола. Указанные метаболиты - активные электрофильные соединения, способные ковалентно связываться с макромолекулами клеток; это приводит к гибели клетки, формированию мутаций, озлокачествлению, дефектам митоза. Кроме того, парагидроксилатед и дигидродиол, действуя как гаптены, могут вызывать и иммунологические реакции. Гиперплазия дёсен, а также тератогенные эффекты - токсические реакции фенитоина зарегистрированы у животных. Доказано, что эти эффекты обусловлены действием метаболитов фенитоина: парагидроксилатеда и дигидродиола. Как показали Buecher и соавт. (1990), низкая активность ЕРНХ1 (меньше 30% от нормы) в амниоцитах - серьёзный фактор риска развития врождённых аномалий плода у женщин, принимающих во время беременности фенитоин. Доказано также, что основная причина снижения активности ЕРНХ1 - точечная мутация в экзоне 3 гена ЕРНХ1; в результате синтезируется дефектный фермент (тирозин в 113 положении заменён на гистидин). Мутация наследуется по аутосомнорецессивному типу. Снижение активности ЕРНХ1 наблюдают только у гомозигот по этой мутантной аллели. Данные о распространённости гомозигот и гетерозигот по этой мутации отсутствуют.
Глутатионтрансферазы
Конъюгации с глутатионом подвергаются ксенобиотики с различной химической структурой: эпоксиды, ареноксиды, гидроксиламины (некоторые из них обладают канцерогенным действием). Среди лекарственных веществ конъюгации с глутатионом подвергаются этакриновая кислота (урегит♠) и гепатотоксичный метаболит парацетамола (ацетаминофена♠) - N-ацетилбензохинонимин, превраща-
ющийся при этом в нетоксичное соединение. В результате реакции конъюгации с глутатионом образуются цистеиновые конъюгаты, называемые «тиоэстеры». Конъюгацию с глутатионом катализируют ферменты глутатион SH-S-трансферазы (GST). Эта группа ферментов локализована в цитозоле, хотя описана и микросомальная GST (однако её роль в метаболизме ксенобиотиков изучена мало). Активность GST в эритроцитах человека у различных индивидуумов различается в 6 раз, однако зависимость активности фермента от пола при этом отсутствует). Тем не менее, как показали исследования, существует чёткая корреляция активности GST у детей и их родителей. По идентичности аминокислотного состава у млекопитающих выделяют 6 классов GST: α- (альфа-), μ- (мю-), κ- (каппа-), θ- (тета-), π- (пи-) и σ- (сигма-) GST. В организме человека в основном экспрессируются GST классов μ (GSTM), θ (GSTT и π (GSTР). Среди них наибольшее значение в метаболизме ксенобиотиков имеют GST класса μ, обозначаемые как GSTM. В настоящее время выделено 5 изоферментов GSTM: GSTM1, GSTM2, GSTM3, GSTM4 и GSTM5. Ген GSTM локализован в хромосоме 1 (локус 1р13.3). GSTM1 экспрессируется в печени, почках, надпочечниках, желудке; слабая экспрессия данного изофермента найдена в скелетных мышцах, миокарде. GSTM1 не экспрессируется в плодной печени, фибробластах, эритроцитах, лимфоцитах и тромбоцитах. GSTM2 («мышечная» GSTM) экспрессируется во всех вышеперечисленных тканях (особенно в мышечной), кроме фибробластов, эритроцитов, лимфоцитов, тромбоцитов и фетальной печени. Экспрессия GSTM3 («мозговая» GSTM) осуществляется во всех тканях организма, особенно в ЦНС. Важная роль в инактивации канцерогенов принадлежит GSTM1. Косвенным подтверждением этого считают достоверное увеличение частоты злокачественных заболеваний среди носителей нулевых аллелей гена GSTM1, у которых отсутствует экспрессия GSTM1. Harada и соавт. (1987), изучив образцы печени, изъятые у 168 трупов, обнаружили, что нулевая аллель гена GSTM1 достоверно чаще встречается у больных с гепатокарциномой. Board и соавт. (1987) впервые выдвинули гипотезу: в организме носителей нулевых аллелей GSTM1 не происходит инактивация некоторых электрофильных канцерогенов. По данным Board и соавт. (1990), распро- странённость нулевой аллели GSTM1 среди европейского населения составляет 40-45%, в то время как у представителей негроидной расы - 60%. Существуют данные о более высокой частоте рака лёгких у носителей нулевой аллели GSTM1. Как показали Zhong и соавт. (1993),
70% больных раком ободочной кишки - носители нулевой аллели GSTM1. Другой изофермент GST, принадлежащий к классу π, - GSTР1 (локализуется главным образом в печени и структурах гематоэнцефалического барьера) участвует в инактивации пестицидов и гербицидов, широко используемых в сельском хозяйстве.
5.5. ФАКТОРЫ, ВЛИЯЮЩИЕ НА БИОТРАНСФОРМАЦИЮ ЛЕКАРСТВЕННЫХ СРЕДСТВ
Генетические факторы, влияющие на систему биотрансформации и транспортёры лекарственных средств
Генетические факторы, представляющие однонуклеотидные полиморфизмы генов, кодирующих ферменты биотрансформации и транс- портёры ЛС, могут в значительной степени влиять на фармакокинетику ЛС. Межиндивидуальные различия в скорости метаболизма ЛС, которые можно оценить по отношению концентрации ЛС-субстрата к концентрации его метаболита в плазме крови или в моче (метаболическое отношение), позволяют выделить группы индивидуумов, различающиеся по активности того или иного изофермента метаболизма.
• «Экстенсивные» метаболизаторы (extensive metabolism, ЕМ) - лица с «нормальной» скоростью метаболизма определённых ЛС, как правило, гомозиготы по «дикой» аллели гена соответствующего фермента. К группе «экстенсивных» метаболизаторов принадлежит большинство населения.
• «Медленные» метаболизаторы (poor metabolism, РМ) - лица со сниженной скоростью метаболизма определённых ЛС, как правило, гомозиготы (при аутосомно-рецессивном типе наследования) или гетерозиготы (при аутосомно-доминантном типе наследования) по «медленной» аллели гена соответствующего фермента. У этих индивидуумов происходит синтез «дефектного» фермента, либо вообще отсутствует синтез фермента метаболизма. В результате происходит снижение ферментативной активности. Нередко обнаруживают полное отсутствие ферментативной активности. У этой категории лиц регистрируют высокие показатели отношения концентрации ЛС к концентрации его метаболита. Следовательно, у «медленных» метаболизаторов ЛС накапливается в организме в высоких концентрациях; это приводит к разви-
• тию выраженных нежелательных лекарственных реакций, вплоть до интоксикации. Именно поэтому таким пациентам (медленным метаболизаторам) необходимо выполнять тщательный подбор дозы ЛС. «Медленным» метаболизаторам назначают меньшие дозы ЛС, чем «активным». «Сверхактивные», или «быстрые» метаболизаторы (ultraextensive metabolism, UM) - лица с повышенной скоростью метаболизма определённых ЛС, как правило, гомозиготы (при аутосомнорецессивном типе наследования) или гетерозиготы (при аутосомно-доминантном типе наследования) по «быстрой» аллели гена соответствующего фермента или, что наблюдают чаще, несущие копии функциональных аллелей. У этой категории лиц регистрируют низкие значения отношения концентрации ЛС к концентрации его метаболита. В результате концентрация ЛС в плазме крови недостаточна для достижения терапевтического эффекта. Таким пациентам («сверхактивным» метаболизаторам) назначают более высокие дозы ЛС, чем «активным» метаболизаторам. Если присутствует генетический полиморфизм того или иного фермента биотрансформации, то распределение индивидуумов по скорости метаболизма ЛС-субстратов данного фермента приобретает бимодальный (если существует 2 типа метаболизаторов) или тримодальный (если существует 3 типа метаболизаторов) характер.
Полиморфизм характерен и для генов, кодирующих транспортёры ЛС, при этом фармакокинетика ЛС может изменяться в зависимости от функции данного транспортёра. Клиническое значение наиболее значимых ферментов биотрансформации и транспортёров рассмотрено ниже.
Индукция и ингибирование системы биотрансформации и транспортёров
Под индукцией фермента биотрансформации или транспортёра понимают абсолютное увеличение его количества и (или) активности вследствие воздействия определённого химического агента, в частности, ЛС. В случае с ферментами биотрансформации это сопровождается гипертрофией ЭПР. Индукции могут подвергаться как ферменты I фазы (изоферменты цитохрома Р-450) и II фазы биотрансформации (УДФ-глюкуронилтрансфераза и др.), так и транспортёры ЛС (гликопротеин-Р, транспортёры органических анионов и катионов). ЛС, индуцирующие ферменты биотрансформации и транспортёры, не обладают очевидным структурным сходством, однако для них харак-
терны некоторые общие признаки. Такие вещества растворимы в жирах (липофильны); служат субстратами ферментов (которые они индуцируют) и имеют, чаще всего, длительный период полувыведения. Индукция ферментов биотрансформации ведёт к ускорению биотрансформации и, как правило, к снижению фармакологической активности, а следовательно, и к эффективности совместно применяемых с индуктором ЛС. Индукция транспортёров ЛС может приводить к различным изменениям концентрации ЛС в плазме крови, в зависимости от функций данного транспортёра. Различные субстраты способны индуцировать ферменты биотрансформации ЛС и транспортёры ЛС с неодинаковыми молекулярной массой, субстратной специфичностью, иммунохимическими и спектральными характеристиками. Кроме того, существуют значительные межиндивидуальные различия в интенсивности индукции ферментов биотрансформации и транспортёров ЛС. Один и тот же индуктор может повышать активность фермента или транспортёра у различных индивидуумов в 15-100 раз.
Основные типы индукции
• «Фенобарбиталовый» тип индукции - непосредственное воздействие молекулы-индуктора на регуляторную область гена; это приводит к индукции фермента биотрансформации или транс- портёра ЛС. Такой механизм наиболее характерен для аутоиндукции. Под аутоиндукцией понимают увеличение активности фермента, метаболизирующего ксенобиотик, под действием самого ксенобиотика. Аутоиндукцию рассматривают как адаптивный механизм, выработанный в процессе эволюции для инактивации ксенобиотиков, в том числе растительного происхождения. Так, аутоиндукцией по отношению к цитохромам подсемейства IIВ обладает фитонцид чеснока - диалил сульфид. Барбитураты (индукторы изоферментов цитохрома Р-450 3А4, 2С9, подсемейства IIВ) - типичные аутоиндукторы (среди лекарственных веществ). Именно поэтому данный тип индукции получил название «фенобарбиталовый».
• «Рифампицин-дексаметазоновый» тип - индукция изоферментов цитохрома Р-450 1А1, 3А4, 2В6 и гликопротеина-Р опосредована взаимодействием молекулы индуктора со специфическими рецепторами, их относят к классу белков-регуляторов транскрипции: прегнан-Х-рецептор (PXR), Ah-рецептор, CAR-рецеп- тор. Соединяясь с этими рецепторами, ЛС-индукторы образуют комплекс, который, проникая в ядро клетки, воздействует на
• регуляторную область гена. В результате происходит индукция фермента биотрансформации ЛС, или транспортёра. По этому механизму рифампины, глюкокортикоиды, препараты зверобоя и некоторые другие вещества индуцируют изоферменты цитохрома Р-450 и гликопротеин-Р. «Этаноловый» тип - стабилизация молекулы фермента биотрансформации ЛС вследствие образования комплекса с некоторыми ксенобиотиками (этанол, ацетон). Например, этанол индуцирует изофермент 2Е1 цитохрома Р-450 на всех этапах его образования: от транскрипции до трансляции. Полагают, что стабилизирующий эффект этанола связан с его способностью активировать систему фосфорилирования в гепатоцитах через ЦАМФ. По данному механизму изониазид индуцирует изофермент 2Е1 цитохрома Р-450. С «этаноловым» механизмом связывают процесс индукции изофермента 2Е1 цитохрома Р-450 при голодании и сахарном диабете, в данном случае в качестве индукторов изофермента 2Е1 цитохрома Р-450 выступают кетоновые тела. Индукция ведёт к ускорению биотрансформации ЛС-субстратов соответствующих ферментов, и, как правило, к снижению их фармакологической активности. Среди индукторов наиболее часто применяют в клинической практике рифампицин (индуктор изоферментов 1А2, 2С9, 2С19, 3A4, 3А5, 3А6, 3А7 цитохрома Р-450; гликопротеин-Р) и барбитураты (индукторы изоферментов 1A2, 2В6, 2C8, 2С9, 2С19, 3A4, 3А5, 3А6, 3А7 цитохрома Р-450). Для развития индуцирующего эффекта барбитуратов требуется несколько недель. В отличие от барбитуратов, рифампицин, как индуктор, действует быстро. Эффект рифампицина можно обнаружить уже через 2-4 дня. Максимальный эффект препарата регистрируют через 6-10 дней. Индукция ферментов, или транспортёров ЛС, вызванная рифампицином и барбитуратами, иногда приводит к снижению фармакологической эффективности непрямых антикоагулянтов (варфарина, аценокумарола), циклоспорина, глюкокортикоидов, кетоконазола, теофиллина, хинидина, дигоксина, фексофенадина и верапамила (это требует коррекции режима дозирования данных ЛС т.е. увеличения дозы). Следует подчеркнуть, что при отмене индуктора ферментов биотрансформации ЛС дозу сочетаемого ЛС следует снижать, так как происходит увеличение его концентрации в плазме крови. Примером такого взаимодействия можно считать комбинацию антикоагулянтов непрямого действия и фенобарбитала. Как показали исследования, в 14% случаев кровотечения при лечении
непрямыми антикоагулянтами развиваются вследствие отмены ЛС, индуцирующих ферменты биотрансформации.
Некоторые соединения могут ингибировать активность ферментов биотрансформации и транспортёров ЛС. Причём при снижении активности ферментов, метаболизирующих ЛС, возможно развитие побочных эффектов, связанных с длительной циркуляцией этих соединений в организме. Ингибирование транспортёров ЛС может приводить к различным изменениям концентрации ЛС в плазме крови в зависимости от функций данного транспортёра. Некоторые лекарственные вещества способны ингибировать как ферменты I фазы биотрансформации (изоферменты цитохрома Р-450) и II фазы биотрансформации (N-ацетилтрансфераза и др.), так и транспортёры ЛС.
Основные механизмы ингибирования
• Связывание с регуляторной областью гена фермента биотрансформации или транспортёра ЛС. По данному механизму происходит ингибирование ферментов биотрансформации ЛС под действием большого количества препарата (циметидин, флуоксетин, омепразол, фторхинолоны, макролиды, сульфаниламиды и т.д.).
• Некоторые препараты, обладающие высоким аффинитетом (сродством) к определённым изоферферментам цитохрома Р-450 (верапамил, нифедипин, исрадипин, хинидин), ингибируют биотрансформацию ЛС с более низким аффинитетом к этим изоферментам. Подобный механизм называют конкурентным метаболическим взаимодействием.
• Прямая инактивация изоферментов цитохрома Р-450 (гастоденр). Угнетение взаимодействия цитохрома Р-450 с НАДФ-Н-цитохром Р-450 редуктазой (фумарокумарины сока грейпфрута и лайма).
Снижение активности ферментов биотрансформации ЛС под действием соответствующих ингибиторов ведёт к повышению концентрации в плазме крови этих ЛС (субстратов для ферментов). При этом происходит удлинение периода полувыведения лекарственных веществ. Всё это служит причиной развития побочных эффектов. Некоторые ингибиторы влияют на несколько изоферментов биотрансформации одновременно. Для угнетения нескольких изоформ ферментов могут потребоваться большие концентрации ингибитора. Так, флуконазол (противогрибковый препарат) в дозе 100 мг в сутки угнетает активность изофермента 2С9 цитохрома Р-450. При повышении дозы данного ЛС до 400 мг отмечают также угнетение
активности изофермента 3А4. Кроме того, чем выше доза ингибитора, тем быстрее развивается (и тем выше) его эффект. Ингибирование вообще развивается быстрее, чем индукция, обычно его можно зарегистрировать уже через 24 ч с момента назначения ингибиторов. На скорость ингибирования активности фермента влияет также путь введения ЛС-ингибитора: если ингибитор вводят внутривенно, то процесс взаимодействия произойдёт быстрее.
Ингибиторами и индукторами ферментов биотрансформации и транспортёров ЛС могут служить не только ЛС, но и фруктовые соки (табл. 5-10), и фитопрепараты (приложение 2) - всё это имеет клиническое значение при применении ЛС, выполняющих функции субстратов для данных ферментов и транспортёров.
Таблица 5-10. Влияние фруктовых соков на активность системы биотрансформации и транспортёров лекарственных средств
 5.6. ЭКСТРАГЕПАТИЧЕСКАЯ БИОТРАНСФОРМАЦИЯ
5.6. ЭКСТРАГЕПАТИЧЕСКАЯ БИОТРАНСФОРМАЦИЯ
Роль кишечника в биотрансформации лекарственных средств
Кишечник считают вторым по значимости органом (после печени), выполняющим биотрансформацию ЛС. В стенке кишечника осуществляются как реакции I фазы, так и реакции II фазы биотрансформации. Биотрансформация ЛС в стенке кишечника имеет большое значение в эффекте первого прохождения (пресистемной биотрансформации). Уже доказана существенная роль биотрансформации в стенке кишечника в эффекте первого прохождения таких ЛС, как циклоспорин А, нифедипин, мидазолам, верапамил.
Ферменты I фазы биотрансформации лекарственных средств в стенке кишечника
Среди ферментов I фазы биотрансформации ЛС, в стенке кишечника в основном локализованы изоферменты цитохрома Р-450. Среднее содержание изоферментов цитохрома Р-450 в стенке кишечника человека составляет 20 пмоль/мг микросомального белка (в печени - 300 пмоль/мг микросомального белка). Установлена чёткая закономерность: содержание изоферментов цитохрома Р-450 уменьшается от проксимальных отделов кишечника к дистальным (табл. 5-11). Кроме того, содержание изоферментов цитохрома Р-450 максимально на вершине ворсинок кишечника и минимально - в криптах. Преобладающийвкишечникеизоферментцитохрома Р-450 - CYP3А4 составляет 70% от всех изоферментов цитохрома Р-450 кишечника. По данным разных авторов, содержание CYP3А4 в стенке кишечника варьирует, что объясняют межиндивидульными различиями цитохрома Р-450. Также имеют значение и способы очистки энтероцитов.
Таблица 5-11. Содержание изофермента 3А4 цитохрома Р-450 в стенке кишечника и печени человека
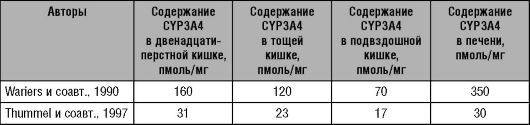 В
стенке кишечника идентифицированы также другие изоферменты: CYP2C9 и
CYP2D6. Однако, по сравнению с печенью, содержание указанных ферментов в
стенке кишечника незначительно (в 100-200 раз меньше). Проведённые
исследования продемонстрировали незначительную, по сравнению с печенью,
метаболическую активность изоферментов цитохрома Р-450 стенки кишечника
(табл. 5-12). Как показали исследования, посвященные изучению индукции
изоферментов цитохрома Р-450 стенки кишечника, индуцибельность
изоферментов стенки кишечника ниже, чем у изоферментов цитохрома Р-450
печени.
В
стенке кишечника идентифицированы также другие изоферменты: CYP2C9 и
CYP2D6. Однако, по сравнению с печенью, содержание указанных ферментов в
стенке кишечника незначительно (в 100-200 раз меньше). Проведённые
исследования продемонстрировали незначительную, по сравнению с печенью,
метаболическую активность изоферментов цитохрома Р-450 стенки кишечника
(табл. 5-12). Как показали исследования, посвященные изучению индукции
изоферментов цитохрома Р-450 стенки кишечника, индуцибельность
изоферментов стенки кишечника ниже, чем у изоферментов цитохрома Р-450
печени.
Таблица 5-12. Метаболическая активность изоферментов цитохромов Р-450 стенки кишечника и печени
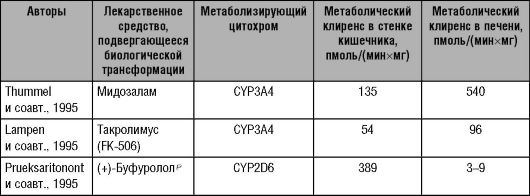 Ферменты II фазы биотрансформации лекарственных средств в стенке кишечника
Ферменты II фазы биотрансформации лекарственных средств в стенке кишечника
УДФ-глюкуронилтрансфераза и сульфотрансфераза - наиболее хорошо изученные ферменты II фазы биотрансформации ЛС, расположенные в стенке кишечника. Распределение этих ферментов в кишечнике аналогично изоферментам цитохрома Р-450. Cappiello и соавт. (1991) изучали активность УДФ-глюкуронилтрансферазы в стенке кишечника и печени человека по метаболическому клиренсу 1-нафтола, морфина и этинилэстрадиола (табл. 5-13). Как показали исследования, метаболическая активность УДФ-глюкуронилтрансферазы стенки кишечника ниже УДФ-глюкуронилтрансферазы печени. Подобная закономерность характерна и для глюкуронирования билирубина.
Таблица 5-13. Метаболическая активность УДФ-глюкуронилтрансферазы в стенке кишечника и в печени
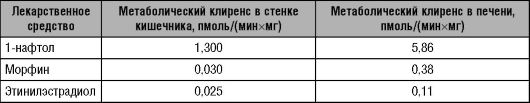 Cappiello
и соавт. (1987) изучали также активность сульфотрансферазы стенки
кишечника и печени по метаболическому клиренсу 2-нафтола. Полученные
данные свидетельствуют о наличии различий показателей метаболического
клиренса (причём клиренс 2-нафтола в стенке кишечника ниже, чем в
печени). В подвздошной кишке величина данного показателя составляет 0,64
нмоль/(минхмг), в сигмовидной кишке - 0,4 нмоль/(минхмг), в печени -
1,82 нмоль/(минхмг). Однако существуют препараты, сульфатирование
которых происходит в основном в стенке кишечника. К ним относят,
например, β2-адреномиметики: тербуталин и изопреналин (табл. 5-14).
Cappiello
и соавт. (1987) изучали также активность сульфотрансферазы стенки
кишечника и печени по метаболическому клиренсу 2-нафтола. Полученные
данные свидетельствуют о наличии различий показателей метаболического
клиренса (причём клиренс 2-нафтола в стенке кишечника ниже, чем в
печени). В подвздошной кишке величина данного показателя составляет 0,64
нмоль/(минхмг), в сигмовидной кишке - 0,4 нмоль/(минхмг), в печени -
1,82 нмоль/(минхмг). Однако существуют препараты, сульфатирование
которых происходит в основном в стенке кишечника. К ним относят,
например, β2-адреномиметики: тербуталин и изопреналин (табл. 5-14).
Таким образом, несмотря на определённый вклад в биотрансформацию лекарственных веществ, стенка кишечника по своей метаболической способности значительно уступает печени.
Таблица 5-14. Метаболический клиренс тербуталина и изопреналина в стенке кишечника и печени
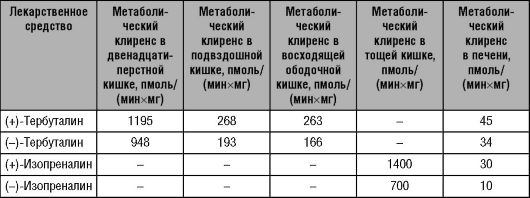 Роль лёгких в биотрансформации лекарственных средств
Роль лёгких в биотрансформации лекарственных средств
В лёгких человека присутствуют как ферменты I фазы биотрансформации (изоферменты цитохрома Р-450), так и ферменты II фазы
(эпоксидгидролаза, УДФ-глюкуронилтрансфераза и др.). В лёгочной ткани человека удалось идентифицировать различные изоферменты цитохрома Р-450: CYP1A1, CYP1B1, CYP2А, CYP2A10, CYP2A11, CYP2В, CYP2E1, CYP2F1, CYP2F3. Общее содержание цитохрома Р-450 в лёгких человека составляет 0,01 нмоль/мг микросомального белка (это в 10 раз меньше, чем в печени). Существуют изоферменты цитохрома Р-450, которые экспрессируются преимущественно в лёг- ких. К ним относят CYP1A1 (найден у человека), CYP2В (у мыши), CYP4В1 (у крысы) и CYP4В2 (у крупного рогатого скота). Эти изоферменты имеют большое значение в биологической активации ряда канцерогенов и пульмонотоксичных соединений. Информация об участии CYP1A1 в биологической активации ПАУ изложена выше. У мышей окисление бутилированного гидрокситолуена изоферментом CYP2В приводит к образованию пневмотоксичного электрофильного метаболита. Изоферменты CYP4В1 крыс и CYP4В2 крупного рогатого скота способствуют биологической активации 4-ипоменола (4-ипоме- нол - сильнодействующий пневмотоксичный фуранотерпеноид грибка сырого картофеля). Именно 4-импоменол стал причиной массового падежа крупного рогатого скота в 70-е годы в США и Англии. При этом 4-ипоменол, окисленный изоферментом CYP4В2, вызывал интерстициальную пневмонию, приводившую к летальному исходу.
Таким образом, экспрессия в лёгких специфичных изоферментов объясняет избирательную пульмонотоксичность некоторых ксенобиотиков. Несмотря на наличие в лёгких и других отделах дыхательных путей ферментов, их роль в биотрансформации лекарственных веществ ничтожна. В таблице приведены ферменты биотрансформации ЛС, обнаруженные в дыхательных путях человека (табл. 5-15). Определение локализации ферментов биотрансформации в дыхательных путях затруднено из-за использования в исследованиях гомогенизата лёгких.
Таблица 5-15. Ферменты биотрансформации, обнаруженные в дыхательных путях человека
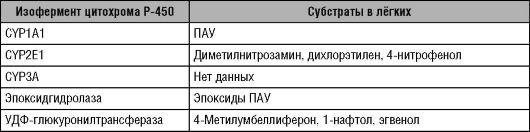 Роль почек в биотрансформации лекарственных средств
Роль почек в биотрансформации лекарственных средств
Исследования, выполненные в течение последних 20 лет, показали, что почки принимают участие в метаболизме ксенобиотиков и лекарственных веществ. При этом, как правило, происходит снижение биологической и фармакологической активности, однако в некоторых случаях возможен и процесс биологической активации (в частности, биоактивации канцерогенов).
В почках обнаружены как ферменты I фазы биотрансформации, так и ферменты II фазы. Причём ферменты биотрансформации локализованы и в корковом, и в мозговом веществе почек (табл. 5-16). Однако, как показали исследования, большее количество изоферментов цитохрома Р-450 содержит именно корковый слой почек, а не мозговой. Максимальное содержание изоферментов цитохрома Р-450 обнаружили в проксимальных почечных канальцах. Так, почки содержат изофермент CYP1A1, ранее считавшийся специфичным для лёгких, и CYP1A2. Причём указанные изоферменты в почках подвергаются индукции ПАУ (например, β-нафтовлавоном, 2-ацетиламино- флурином) так же, как и в печени. В почках обнаружили активность CYP2B1, в частности, описали окисление парацетамола (ацетаминофена♠) в почках под действием этого изофермента. Позднее продемонстрировали, что именно образование токсичного метаболита N-ацетибензахинонимина в почках под действием CYP2E1 (по аналогии с печенью) - основная причина нефротоксического действия данного препарата. При совместном применении парацетамола с индукторами CYP2E1 (этанолом, тестостероном и т.д.) риск поражения почек возрастает в несколько раз. Активность CYP3A4 в почках регистрируют не всегда (только в80% случаев). Следует отметить: вклад изоферментов цитохрома Р-450 почек в биотрансформацию лекарственных веществ скромен и, видимо, в большинстве случаев не имеет клинического значения. Однако для некоторых ЛС биохимическое преобразование в почках - основной путь биотрансформации. Как показали исследования, тропизетронр (противорвотное ЛС), главным образом, окисляется в почках под действием изоферментов CYP1A2 и CYP2E1.
Среди ферментов II фазы биотрансформации в почках наиболее часто определяют УДФ-глюкуронилтрансферазу и β-лиазу. Следует отметить, что активность β-лиазы в почках выше, чем в печени. Обнаружение этой особенности позволило разработать некоторые «пролекарства», при активации которых образуются активные мета-
болиты, селективно действующие на почки. Так, создали цитостатический препарат для лечения хронического гломерулонефрита - S-(6-пуринил)-L-цистеин. Это соединение - изначально неактивное, в почках под действием β-лиазы превращается в активный 6-мер- каптопурин. Таким образом, 6-меркуптопурин производит эффект исключительно в почках; это значительно снижает частоту и выраженность нежелательных лекарственных реакций.
Глюкуронированию в почках подвергаются такие ЛС, как парацетамол (ацетаминофен♠), зидовудин (азидотимидин♠), морфин, сульфаметазонр, фуросемид (лазикс♠) и хлорамфеникол (левомицетин♠).
Таблица 5-16. Распределение ферментов биотрансформации лекарственных средств в почках (Lohr и соавт., 1998)
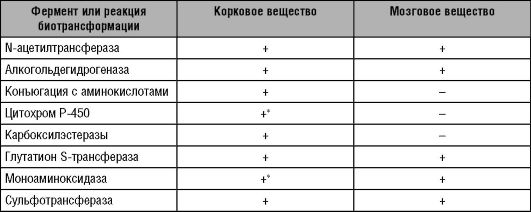 * - содержание фермента достоверно выше.
* - содержание фермента достоверно выше.
Литература
Кукес В.Г. Метаболизм лекарственных средств: клинико-фармакологические аспекты. - М.: Реафарм, 2004. - С. 113-120.
Середенин С.Б. Лекции по фармакогенетике. - М.: МИА, 2004. -
С. 303.
Diasio R.B., Beavers T.L., Carpenter J.T. Familial deficiency of dihydropyrimidine dehydrogenase: biochemical basis for familial pyrimidinemia and severe 5-fluorouracil-induced toxicity // J. Clin. Invest. - 1988. - Vol. 81. -
P. 47-51.
Lemoine A., Daniel A., Dennison A., Kiffel L. et al. FK 506 renal toxicity and lack of detectable cytochrome P-450 3A in the liver graft of a patient undergoing liver transplantation // Hepatology. - 1994. - Vol. 20. - P. 1472-1477.
Lewis D.F.V., Dickins M., Eddershaw P.J. et al. Cytochrome-P450 Substrate Specificities, Substrate structural Templates and Enzyme Active Site Geometries // Drug Metabol. Drug Interact. - 1999. - Vol. 15. - P. 1-51.
Nebert D.W., Adesnik M., Coon M.J. et al. The P450 gene superfamily: recommended nomenclature // DNA. - 1987. - Vol. 6 - P. 1-11.
Rau T., Wohileben G., Wuttke H. et al. CYP2D6 genotype: impact on adverse effects and nonresponse during treatment with antidepressants - a
pilot study // Clin. Pharmacol. Ther. - 2004. - Vol. 75 - P. 386-393.
Saxena R., Shaw G.L., RellingM.V. et al. Identification of a new variant CYP2D6 allele with a single base deletion in exon 3 and its association with the poor metabolizer phenotype // Hum. Mol. Genet. - 1994. - Vol. 3. -
P. 923-926.
Stamer U.M., Lehnen K., Hothker F. et al. Impact of CYP2D6 genotype on postoperative tramadol analgesia // Pain. - 2003. - Vol. 105. - P. 231-238. WoolfT.F. Handbook of Drug Metabolism. - 1999. - P. 153-169.