Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 5. ОСНОВНЫЕ ПОНЯТИЯ ОБЩЕЙ ГИСТОЛОГИИ
Ткань - это возникшая в ходе эволюции частная система организма, состоящая из одного или нескольких дифферонов клеток и их производных, обладающая специфическими функциями благодаря кооперативной деятельности всех ее элементов.
5.1. ТКАНЬ КАК СИСТЕМА
Любая ткань - сложная система, элементы которой - клетки и их производные. Сами ткани тоже являются элементами морфофункциональных единиц, а последние выступают в роли элементов органов. Поскольку по отношению к системе высшего ранга (в нашем случае - организму) системы более низких рангов рассматриваются как частные, то и о тканях следует говорить как о частных системах.
В любой системе все элементы упорядочены в пространстве и функционируют согласованно друг с другом; система в целом обладает при этом свойствами, не присущими ни одному из ее элементов, взятому в отдельности. Соответственно и в каждой ткани ее строение и функции несводимы к простой сумме свойств отдельных входящих в нее клеток и их производных. Ведущими элементами тканевой системы являются клетки. Кроме клеток, различают клеточные производные (постклеточные структуры и симпла-сты) и межклеточное вещество (схема 5.1).
Среди клеточных структур целесообразно различать те, которые, будучи рассматриваемы и вне ткани, полностью обладают свойствами живого (например, способностью к размножению, регенерации при повреждениях и т. п.), и те, которые не обладают полнотой свойств живого. Постклеточные (послеклеточные) структуры относятся к последним.
Клеточные структуры, прежде всего, могут быть представлены индивидуально существующими клетками, каждая из которых имеет собственное ядро и собственную цитоплазму. Такие клетки могут быть либо одноядер-
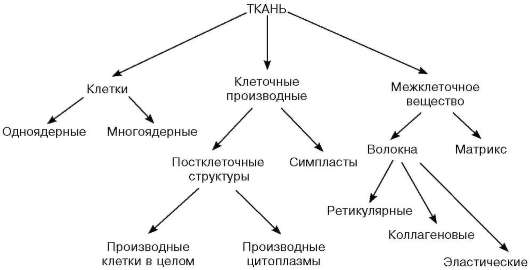
Схема 5.1. Основные структурные элементы тканей
ными, либо многоядерными (если на каком-то этапе произошла нуклеото-мия без цитотомии). Если клетки по достижении какого-либо этапа развития сливаются друг с другом, то возникают симпласты. Примерами их могут служить симпластотрофобласт, остеокласты и симпластическая часть мышечного волокна скелетной мышечной ткани. Симпласты имеют совершенно иной принцип возникновения, нежели многоядерные клетки, так что эти понятия смешивать нецелесообразно.
Особо следует упомянуть случай, когда при делении клеток цитотомия остается незавершенной и отдельные из них остаются соединенными тонкими цитоплазматическими мостиками. Это - синцитий. Такая структура у млекопитающих встречается только в ходе развития мужских половых клеток, однако, поскольку эти клетки не относятся к соматическим, данную структуру не приходится причислять к тканевым.
Постклеточными структурами называют те производные клеток, которые утратили (частично или полностью) свойства, присущие клеткам как живым системам. Несмотря на это, постклеточные структуры выполняют важные физиологические функции, их нельзя расценивать просто как отмирающие или погибшие клетки. Среди постклеточных структур различают производные клеток в целом и производные их цитоплазмы. К первым относятся эритроциты большинства млекопитающих (форменные элементы крови, утратившие ядро на одном из этапов своего развития), роговые чешуйки эпидермиса, волосы, ногти. Примером вторых могут служить тромбоциты (производные цитоплазмы мегакариоцитов).
Межклеточное вещество - продукты синтеза в клетках. Его подразделяют на основное («аморфное», матрикс) и на волокна. Основное вещество может существовать в формах жидкости, золя, геля или быть минерализованным. Среди волокон различают обычно три вида: ретикулярные, коллагеновые и эластические.
Клетки всегда находятся во взаимодействии друг с другом и с межклеточным веществом. При этом формируются различные структурные объединения. Клетки могут лежать в межклеточном веществе на расстоянии друг от друга и взаимодействовать через него без непосредственных контактов (например, в рыхлой волокнистой соединительной ткани), либо соприкасаясь отростками (ретикулярная ткань) или образуя сплошные клеточные массы, или пласты (эпителий, эндотелий).
Дистантно клетки могут взаимодействовать с помощью химических соединений, которые клетки синтезируют и выделяют в процессе своей жизнедеятельности. Такие вещества служат не в качестве внешних секретов, как, например, слизь или пищевые ферменты, а выполняют регуля-торные функции, действуя на другие клетки, стимулируя или тормозя их активность. На этой основе формируется система положительных и отрицательных обратных связей, образуя управляющие контуры. Для реализации каждой из связей требуется некоторое время. Поэтому в тканях активность их жизнедеятельности не остается строго постоянной, а колеблется вокруг некоего среднего состояния. Такие регулярные колебания являются проявлением биологических ритмов на тканевом уровне.
Среди регуляторных веществ (иногда их называют биологически активными веществами) различают гормоны и интеркины. Гормоны поступают в кровь и способны действовать на значительных расстояниях от места их выработки. Интеркины действуют местно. К их числу относят вещества, угнетающие и стимулирующие клеточное размножение, определяющие направления дифференцировки клеток-предшественников, а также регулирующие запрограммированную клеточную гибель (апоптоз).
Таким образом, все межклеточные взаимодействия, как непосредственные, так и через межклеточное вещество, обеспечивают функционирование ткани как единой системы. Только на основе системного подхода возможно изучение тканей, понимание общей гистологии.
5.2. РАЗВИТИЕ ТКАНЕЙ (ЭМБРИОНАЛЬНЫЙ ГИСТОГЕНЕЗ)
В эмбриогенезе человека наблюдаются все процессы, характерные для позвоночных животных: оплодотворение, образование зиготы, дробление, гаструляция, формирование трех зародышевых листков, обособление комплекса эмбриональных зачатков тканей и органов, а также мезенхимы, заполняющей пространства между зародышевыми листками.
Геном зиготы не активен. По мере дробления в клетках - бластомерах - отдельные части генома активизируются, причем в разных бластомерах - разные. Этот путь развития генетически запрограммирован и обозначается как детерминация. В результате появляются стойкие различия их биохимических (а также и морфологических) свойств - дифференцировка. Одновременно дифференцировка сужает потенции дальнейшей активации
генома, которая возможна теперь за счет его оставшейся неактивированной части - происходит ограничение возможностей развития - комми-тирование.
По времени дифференцировка не всегда совпадает с детерминацией: детерминация в клетках может уже совершиться, а специфические функции и морфологические особенности проявятся позже. Подчеркнем, что все эти процессы совершаются на уровне генома, но без изменения набора генов как целого: гены не исчезают из клетки, хотя они могут быть и не активными. Такие изменения называют эпигеномными, или эпигенетическими.
Вопрос о том, насколько возможен возврат активной части генома вновь в неактивное состояние (дедифференцировка) в естественных условиях, остается неясным (это не исключает таких возможностей при генно-инженерных экспериментах).
Дифференцировка и коммитирование в эмбриогенезе появляются не сразу. Они совершаются последовательно: сначала преобразуются крупные участки генома, детерминирующие наиболее общие свойства клеток, а позднее - более частные свойства. В развивающемся организме дифферен-цировка сопровождается специфической организацией или размещением специализирующихся клеток, что выражается в установлении определенного плана строения в ходе онтогенеза - морфогенеза.
В результате дробления зародыш разделяется на внезародышевую и зародышевую части, причем становление тканей идет и в той, и в другой. В результате гаструляции в зародышевой части формируются гипобласт и эпибласт, а далее - образуются три зародышевых листка. В составе последних вследствие детерминации обособляются эмбриональные зачатки (еще не ткани). Их клетки обладают такой детерминацией и, в то же время, коммитированием, что в естественных условиях они не могут превратиться в клетки другого эмбрионального зачатка. Эмбриональные зачатки в свою очередь представлены стволовыми клетками - источниками дифферонов, формирующих ткани в эмбриональном гистогенезе (рис. 5.1). Межклеточного вещества зачатки не имеют.
В процессе образования трех зародышевых листков часть клеток мезодермы выселяется в промежутки между зародышевыми листками и формирует сетевидную структуру - мезенхиму, заполняющую пространство между зародышевыми листками. В последующем дифференцировка зародышевых листков и мезенхимы, приводящая к появлению эмбриональных зачатков тканей и органов, происходит неодновременно (гетерохронно), но взаимосвязанно (интегративно).
На понятии «мезенхима» следует остановиться особо. Содержание, которое вкладывают в него, весьма разнообразно. Часто ее определяют как эмбриональную соединительную ткань либо как эмбриональный зачаток. В последнем случае говорят о развитии из мезенхимы конкретных тканей, на основе чего даже делают выводы о родственности этих тканей. Мезенхиму считают источником развития клеток фибробластического ряда и клеток крови, эндотелиоцитов и гладких миоцитов, клеток мозгового вещества надпочечников. В частности, такая концепция долгое время «обосновывала» принадлежность эндотелия к соединительной ткани с отрицани-
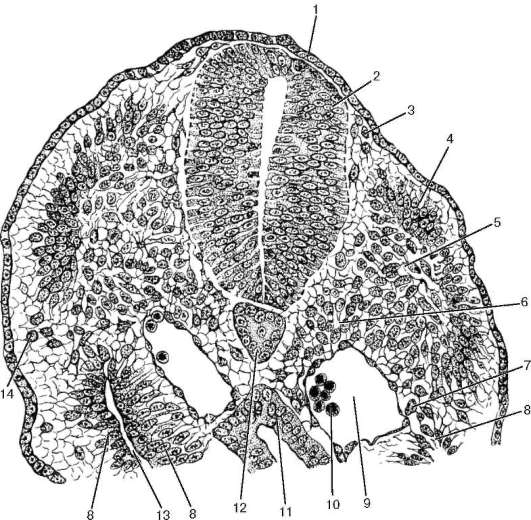
Рис. 5.1. Локализация эмбриональных зачатков тканей и органов в теле зародыша (срез зародыша в стадии 12 сомитов, по А. А. Максимову, с изменениями): 1 - кожная эктодерма; 2 - нервная трубка; 3 - нейральный гребень; 4 - дерматом; 5 - миотом; 6 - склеротом; 7 - сегментная ножка; 8 - выстилка целома; 9 - аорта, выстланная эндотелием; 10 - клетки крови; 11 - кишечная трубка; 12 - хорда; 13 - полость целома; 14 - мигрирующие клетки, образующие мезенхиму
ем его тканевой специфичности. В некоторых учебниках анатомии до сих пор можно встретить классификацию мышц (как органов) на основании их развития либо из миотомов, либо из мезенхимы.
Признание мезенхимы в качестве эмбриональной соединительной ткани вряд ли состоятельно, хотя бы потому, что клетки ее еще не обладают одним из основных свойств ткани - специфической функцией. Они не синтезируют коллаген, эластин, гликозаминогликаны, как это свойственно фибробластам соединительной ткани, они не сокращаются, как миоциты, не обеспечивают двустороннего транспорта веществ, как эндотелиоциты. Морфологически они неотличимы друг от друга. Вряд ли можно считать мезенхиму и единым эмбриональным зачатком: в ходе развития зародыша клетки многих из них выселяются в нее, будучи уже соответственно детерминированными.
В составе мезенхимы совершается, в частности, миграция промиобластов и миобластов (выселившихся из сомитов), предшественников меланоцитов и клеток мозгового вещества надпочечников, клеток АПУД-серии (высе-
лившихся из сегментов нейрального гребня), клеток-предшественников эндотелия (скорее всего, выселившихся из спланхнотомов) и другие. Можно полагать, что, мигрируя и вступая друг с другом в контактные или химические взаимоотношения, клетки могут детализировать свою детерминацию.
Во всяком случае, считать мезенхиму единым эмбриональным зачатком не приходится. В рамках эпигеномных представлений ее надо рассматривать как гетерогенное образование. Клетки мезенхимы, хотя и сходны по морфологическим признакам, вовсе не безлики и не однолики в эпигеном-ном смысле. Поскольку клетки мезенхимы дают начало многим тканям, ее называют также плюриили полипотентным зачатком. Такое понимание противоречит представлению о зачатках как клеточных группировках, в которых клетки уже достигли значительной степени коммитированности. Признание мезенхимы единым зачатком означало бы отнесение к одному типу таких тканей, как скелетная, мышечная, кровь, железистый эпителий мозгового вещества надпочечников и многих других.
Как уже было отмечено, говорить о происхождении какой-либо ткани из зародышевого листка совершенно недостаточно для характеристики свойств и принадлежности к гистогенетическому типу. Столь же малозначаще и постулирование развития какой-либо ткани из мезенхимы. Судьба клеток мезенхимы по завершении их миграции - дифференциация в клетки конкретных тканей в составе конкретных органов. После этого мезенхимы как таковой не остается. Поэтому концепции о так называемом мезенхимном резерве неправомерны. В составе дефинитивных тканей, безусловно, могут оставаться либо стволовые клетки, либо клетки-предшественники, но это - клетки с уже детерминированными гистиотипическими свойствами.
Диффероны. Совокупность клеток, ведущих свое начало от общей пред-ковой формы, можно рассматривать как ветвящееся дерево последовательных процессов детерминации, сопровождающихся при этом коммитиро-ванием путей развития. От клеток, у которых эти процессы совершаются на уровне эмбриональных зачатков, можно проследить отдельные ветви, ведущие к различным конкретным дефинитивным (зрелым) клеточным видам. Такие исходные клетки называют стволовыми, а совокупность ветвей их потомков объединяют в диффероны. В составе дифферона происходят дальнейшая детерминация и коммитирование потенций развития стволовой клетки, в результате чего возникают так называемые клетки-предшественники. В каждой из таких ветвей, в свою очередь, возникают уже зрелые дифференцированные клетки, которые затем стареют и отмирают (рис. 5.2). Стволовые клетки и клетки-предшественники способны к размножению и в совокупности могут быть названы камбиальными.
Так, в системе крови от единой стволовой клетки всех форменных элементов (см. более подробно в главе 7 «Кровь» и «Кроветворение») возникают общая ветвь гранулоцитов и моноцитов, общая ветвь различных видов лимфоцитов, а также не ветвящаяся эритроидная линия (иногда такие ветви и линии тоже рассматривают как отдельные диффероны).
Хотя стволовые клетки детерминируются еще в составе эмбриональных зачатков, они могут сохраняться и в тканях взрослых организмов, но их
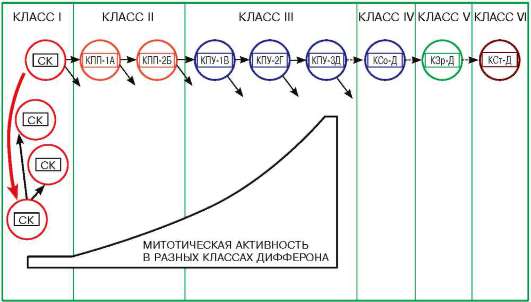
Рис. 5.2. Схема организации клеточного дифферона:
Классы клеток в диффероне: I - стволовые клетки; II - полипотентные клетки-предшественники; III - унипотентные клетки-предшественники; IV - созревающие клетки; V - зрелые клетки; выполняющие специфические функции; VI - стареющие и гибнущие клетки. В классах I-III происходит размножение клеток, это отображено на схеме двумя стрелками, отходящими от клетки вправо. Митотическая активность при этом нарастает. Клетки классов IV-VI не делятся (вправо отходит лишь одна стрелка).
СК - стволовые клетки; КПП - клетки-предшественники полипотентные; КПУ - клетки-предшественники унипотентные; КСо - клетки созревающие (уже не делящиеся, но еще не имеющие окончательных специфических функций); КЗр - зрелые клетки (обладающие специфическими функциями); КСт - стареющие клетки (утрачивающие полноту специфических функций).
Цифры после указания на класс клеток условно означают номер поколения в данном классе, следующие за ними буквы - свойства клеток. Обратите внимание, что дочерние клетки, возникшие в результате последовательных делений (классы I-III), имеют разную детерминацию, но сохраняют ее свойства в классах IV-VI. Толстая стрелка слева, направленная вниз, - сигнал для деления стволовой клетки, после того как одна из них вышла из популяции и вступила на путь дифференци-ровки
собственных предков уже не остается. Поэтому в организме нет таких клеточных форм, которые могли бы восполнить убыль стволовых, если она по какой-либо причине произошла, поэтому важнейшее свойство стволовых клеток - самоподдержание их популяции. Это означает, что в естественных условиях, если одна из стволовых клеток вступает на путь дифференциации, и, таким образом, общая их численность снижается на одну, восстановление популяции происходит только за счет деления аналогичной стволовой клетки из той же популяции. При этом она полностью сохраняет свои исходные свойства. В диффероне самоподдерживающуюся клеточную
популяцию выделяют в класс I. Наряду с этим определяющим признаком, стволовые клетки обладают и более частными, но существенными, с медицинской точки зрения, свойствами: стволовые клетки делятся очень редко, следовательно, они наиболее устойчивы к повреждающим воздействиям. Поэтому в случае чрезвычайных ситуаций они гибнут в последнюю очередь. Пока стволовые клетки сохраняются в организме, клеточная форма регенерации тканей возможна после устранения вредоносных воздействий. Если пораженными оказались и стволовые клетки, то клеточная форма регенерации не происходит.
В отличие от стволовых клеток, численность популяции клеток-предшественников может пополняться не только за счет деления клеток, себе подобных, но и за счет менее дифференцированных форм. Чем далее заходит дифференцировка, тем меньшую роль играет самоподдержание, поэтому пополнение популяции дефинитивных клеток происходит, в основном, за счет деления предшественников на промежуточных этапах развития, а стволовые клетки включаются в размножение только тогда, когда активности промежуточных предшественников для пополнения популяции недостаточно.
Клетки-предшественники (иногда их называют полустволовыми) составляют следующую часть гистогенетического древа. Они коммитированы и могут дифференцироваться, но не по всем возможным, а лишь по некоторым направлениям. Если таких путей несколько, клетки называют полипо-тентными (класс II), если же они способны дать начало лишь одному виду клеток - унипотентными (класс III). Пролиферативная активность клеток-предшественников выше, чем у стволовых, и именно они пополняют ткань новыми клеточными элементами.
На следующем этапе развития деления прекращаются, но морфологические и функциональные свойства клеток продолжают изменяться. Такие клетки называют созревающими и относят к классу IV. По достижению окончательной дифференцировки зрелые клетки (класс V) начинают активно функционировать. На последнем этапе их специфические функции угасают и клетки гибнут путем апоптоза (стареющие клетки, класс VI). Направление развития клеток в диффероне зависит от многих факторов: в первую очередь, от интеркинов микроокружения и от гормональных.
Соотношение клеток различной степени зрелости в дифферонах разных тканей организма неодинаково. Клетки различных дифферонов в процессе гистогенеза могут объединяться, причем количество дифферонов в каждом виде тканей может быть различным. Клетки дифферонов, входящих в ткань, участвуют в синтезе ее общего межклеточного вещества. Результатом гистоге-нетических процессов является формирование тканей с их специфическими функциями, не сводимыми к сумме свойств отдельных дифферонов.
Итак, под тканями целесообразно понимать частные системы организма, относящиеся к особому уровню его иерархической организации и включающие в качестве ведущих элементов клетки. Клетки тканей могут относиться к единому или к нескольким стволовым дифферонам. Клетки
одного из дифферонов могут преобладать и быть функционально ведущими. Все элементы ткани (клетки и их производные) равно необходимы для ее жизнедеятельности.
5.3. КЛАССИФИКАЦИИ ТКАНЕЙ
Существенное место среди вопросов общей гистологии занимают проблемы классификации тканей. В отличие от формальных классификаций, отталкивающихся от удобных для наблюдения признаков, естественные классификации призваны учитывать глубокие природные связи между объектами. Именно поэтому структура любой естественной классификации отражает реальную структуру природы.
Время от времени классификационные схемы меняются. Это означает, что в изучении природы сделан еще один шаг, и закономерности исследованы более полно и точно. Разносторонность подходов к характеристикам предметов классификации определяет и многомерность классификационных схем.
С позиции филогенеза предполагается, что в процессе эволюции как у беспозвоночных, так и позвоночных образуются четыре тканевые системы, или группы. Они обеспечивают основные функции организма: 1 - покровные, отграничивающие его от внешней среды и разграничивающие среды внутри организма; 2 - внутренней среды, поддерживающие динамическое постоянство состава организма; 3 - мышечные, отвечающие за движение; и 4 - нервные (или нейральные), координирующие восприятие сигналов внешней и внутренней среды, их анализ и обеспечивающие адекватные ответы на них.
Объяснение этому феномену дали А. А. Заварзин и Н. Г. Хлопин, которые заложили основы учения об эволюционной и онтогенетической детерминации тканей. Так, было выдвинуто положение о том, что ткани образуются в связи с основными функциями, обеспечивающими существование организма во внешней среде. Поэтому изменения тканей в филогенезе идут параллельными путями (теория параллелизмов А. А. Заварзина). При этом дивергентный путь эволюции организмов ведет к возникновению все большего разнообразия тканей (теория дивергентной эволюции тканей Н. Г. Хлопина). Из этого следует, что ткани в филогенезе развиваются и параллельными рядами, и дивергентно. Дивергентная дифференциация клеток в каждой из четырех тканевых систем в конечном итоге и привела к большому разнообразию видов тканей.
Позже выяснилось, что в ходе дивергентной эволюции конкретные ткани могут развиваться не только из одного, а из нескольких источников. Выделение основного из них, дающего начало ведущему клеточному типу в составе ткани, создает возможности для классификации тканей по генетическому признаку, единство же структуры и функции - по морфо-физиологическому. Большинство гистологов сейчас опираются именно на
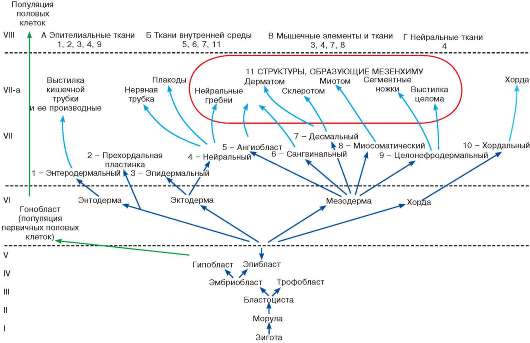
Схема 5.2. Развитие эмбриональных зачатков и тканей:
Арабские цифры - эмбриональные зачатки; римские цифры - этапы развития зародыша и гистогенеза; А-Г - группы тканей.
В основании схемы (I уровень) лежит зигота. На II уровень поставлена морула - форма строения зародыша, которая возникает на этапе дробления. На III уровне отмечена бластоциста. В ней выделяются эмбриобласт и трофобласт (IV уровень). С этого времени развитие идет дивергентно. В эмбриобласте выделяются два листка - эпибласт и гипобласт, показанные на V уровне.
Возникновение и развитие половых клеток выделено особым стилем линии. Они остаются недетерминированными вплоть до взрослого состояния организма и, соответственно, не коммитируются. Поэтому если эмбриональные зачатки определять как совокупность клеток с соответствующей детерминацией и коммитированием, то понятие зачатка к совокупности первичных половых клеток неприменимо. На втором этапе гаструляции возникают три зародышевых листка (VI уровень). Именно в зародышевых листках в конце гаструляции и происходит детерминация (и соответствующее коммитирование) эмбриональных зачатков (VII уровень). Локализация зачатков в теле зародыша отмечена на VII уровне добавлением буквы «а». В энтодерме детерминируется энтеродермальный зачаток (1 - источник эпителиев кишечника и органов, связанных с ним).
В зародышевой эктодерме детерминируются эпидермальный и нейральный зачатки (3 и 4). Механизм детерминации прехордальной пластинки (2) до сих пор вызывает дискуссии, поэтому на схеме она отмечена как особая ветвь, возникающая при диф-ференцировке эпибласта, но не входящая в какой-либо определенный зародышевый листок.
В мезодерме детерминируются следующие зачатки: ангиобласт (5 - источник сосудистого эндотелия), сангвинальный (6 - источник форменных элементов крови), десмальный (7 - от греческого «десмос» - соединять, связывать, источник соединительных тканей и стромы гемопоэтических тканей), миосоматический (8 - источник поперечнополосатой скелетной мышечной ткани), целонефродермальный (9 - источник выстилки целома, эпителиев почек и половых органов, а также сердечной мышечной ткани). С мезодермой рассматривается и хорда, где детерминируется хордальный зачаток (10).
Клетки, мигрирующие и образующие мезенхиму (11), обозначены стрелками, выделенными цветом.
В соответствии с ведущими функциями тканей, последние представлены четырьмя основными морфофункциональными группами (VIII уровень схемы). В каждой группе присутствуют клетки, берущие начало из разных эмбриональных зачатков. Они обозначены соответствующими арабскими цифрами
сочетание морфофункциональной классификации А. А. Заварзина с генетической системой тканей Н. Г. Хлопина (однако из этого не следует, что удалось построить совершенную классификацию, которая была бы общепризнанной).
В настоящее время можно представить следующую схему классификации тканей (схема 5.2). На ней римскими цифрами показаны основные узлы, отражающие развитие зародыша от зиготы через уровень становления зародышевых листков и, далее, - эмбриональных зачатков. Заглавными буквами обозначены основные ткани, относящиеся к главным четырем морфо-функциональным группам. Эмбриональные зачатки обозначены арабскими
цифрами. Каждая группа может быть образована несколькими дифферона-ми, относящимися к разным гистогенетическим типам, однако существуют и монодифферонные ткани.
Очень часто при описании тканей среди прочих их функций выделяют так называемую «защитную», хотя это, по сути дела, отражает лишь сугубо утилитарный медицинский, но не общебиологический подход. В действительности же все функции тканей обеспечивают, прежде всего, нормальное динамическое равновесие всех систем организма в обычных постоянно меняющихся условиях существования. Лишь иногда воздействие факторов, нарушающих равновесие, переходит допустимые границы. В таких случаях обычные реакции, действительно, интенсифицируются и мобилизуются для восстановления нарушенного равновесия, и, как следствие, их качественные взаимоотношения меняются. Именно в подобных случаях на базе физиологических реакций возникают защитные. Они направлены на нейтрализацию и на ликвидацию агента, ставшего из нормального раздражителя угрожающим. Таким образом, понятие защиты целесообразно применять лишь в условиях патологии, применительно же к норме стоит говорить о поддержании равновесных соотношений. В норме нет факторов, с которыми надо бороться и от которых следует защищаться, в нормальных условиях ткани работают, будучи уравновешенными между собой и с окружающей средой.
В соответствии с морфофункциональным принципом, целесообразно в рамках группы выделять подгруппы, например группу тканей внутренней среды подразделить на подгруппы кровь и лимфу с кроветворными тканями, волокнистые соединительные ткани и скелетные ткани. В группе нейральных тканей в одну подгруппу целесообразно выделить собственно нервную ткань (совокупность нейронов как систему, непосредственно обусловливающую ее функции) и глию (как совокупность тканей, непосредственно «обслуживающих» нейроны), а также микроглию. В группе мышечных тканей выделяют подгруппы гладких и поперечнополосатых (неисчерченных и исчерченных).
5.4. РЕГЕНЕРАЦИЯ ТКАНЕЙ
Знание основ эмбрионального гистогенеза необходимо для понимания теории регенерации, т. е. восстановления структуры биологического объекта после утраты части ее элементов. Соответственно уровням организации живого различают внутриклеточную, клеточную, тканевую, органную формы регенерации. Предметом общей гистологии является регенерация на тканевом уровне. У разных тканей возможности регенерации неодинаковы. Различают физиологическую и репаративную регенерацию. Физиологическая регенерация генетически запрограммирована. Репаративная регенерация происходит после случайной гибели клеток, например, в результате интоксикации (в том числе и алкогольной), воздействий постоянного природного радиационного фона, космических лучей на организм.
Таблица 5.1. Регенерационные возможности тканей
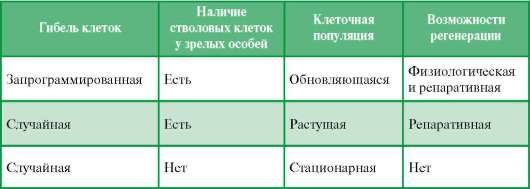
При физиологической регенерации популяция клеток обновляется постоянно. Дифференцированные зрелые клетки имеют ограниченный срок жизни и, выполнив свои функции, гибнут путем апоптоза. Убыль популяции клеток восполняется за счет деления клеток-предшественников, а последних - за счет стволовых клеток. Такие ткани называют обновляющимися. Примерами таких тканей (среди многих других) могут служить многослойный кожный эпителий и кровь.
В некоторых тканях активное размножение клеток идет до тех пор, пока не закончился рост организма. Далее физиологической регенерации в них не происходит, хотя и после завершения роста в них остаются малодиффе-ренцированные клетки. В ответ на случайную гибель специализированных клеток возникает размножение малодифференцированных клеток, и популяция восстанавливается. После восстановления клеточной популяции размножение клеток снова угасает. Такие ткани относят к растущим. Отдельными примерами их могут служить эндотелий сосудов, нейроглия, эпителий печени.
Есть и такие ткани, в которых после окончания роста размножение клеток не наблюдается. В этих случаях ни физиологическая, ни репаративная регенерация невозможна. Такие ткани называют стационарными. Примерами могут служить сердечная мышечная ткань и собственно нервная ткань (совокупность нейронов). У взрослого человека в таких тканях регенерация происходит лишь на внутриклеточном уровне.
Изложенное кратко иллюстрирует табл. 5.1.
Контрольные вопросы
1. Перечислите основные структурные элементы тканей.
2. Охарактеризуйте понятия зародышевый листок, эмбриональный зачаток, дифферон.
3. Дайте определение ткани с позиции клеточно-дифферонной организации.
4. Назовите формы регенерации тканей.