Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 20. ПОЛОВАЯ СИСТЕМА
Половая система объединяет органы, которые обеспечивают воспроизводство позвоночных и человека, и включает гонады, где происходит образование половых клеток и синтез половых гормонов, и добавочные органы полового тракта.
В мужском и женском организмах органы половой системы имеют выраженные морфофункциональные особенности, определяющие вторичные половые признаки. В мужском организме гонады представлены яичками, а добавочные органы - семявыносящими путями, семенными пузырьками, предстательной и бульбо-уретральными железами и половым членом. В женском организме гонады представлены яичниками, а добавочные органы - маткой, маточньми трубами (яйцеводами), влагалищем, наружными половыми органами. В женском организме с половым созреванием тесно связана гистофизиология молочной железы (см. главу 18).
Различия между полами предопределяются генетически через половые хромосомы (XY у мужчин и XX у женщин). Существенной особенностью женской половой системы является цикличность и периодичность деятельности. При этом созревание женской половой клетки и изменения активности секреции женских половых гормонов регулярно повторяются, тогда как мужская половая система функционирует непрерывно с момента достижения организмом половой зрелости до начала возрастного увядания.
Развитие. Закладка половой системы в начальных стадиях эмбриогенеза протекает у обоих полов одинаково (индифферентная стадия) и во взаимодействии с развитием выделительной системы (рис. 20.1). Гонада становится заметна у 4-недельного зародыша в виде половых валиков - утолщений цело-мического эпителия на вентромедиальной поверхности обеих первичных почек (mesonephros). Первичные половые клетки у зародышей обоего пола - гоноци-ты - появляются в пресомитных стадиях эмбриогенеза (во 2-й фазе гастру-ляции). Однако клетки четко выявляются при формировании желточного пузырька. В стенке последнего гоноциты характеризуются большими размерами, крупным ядром, повышенным содержанием гликогена и высокой активностью щелочной фосфатазы в цитоплазме. Здесь клетки размножаются, затем,
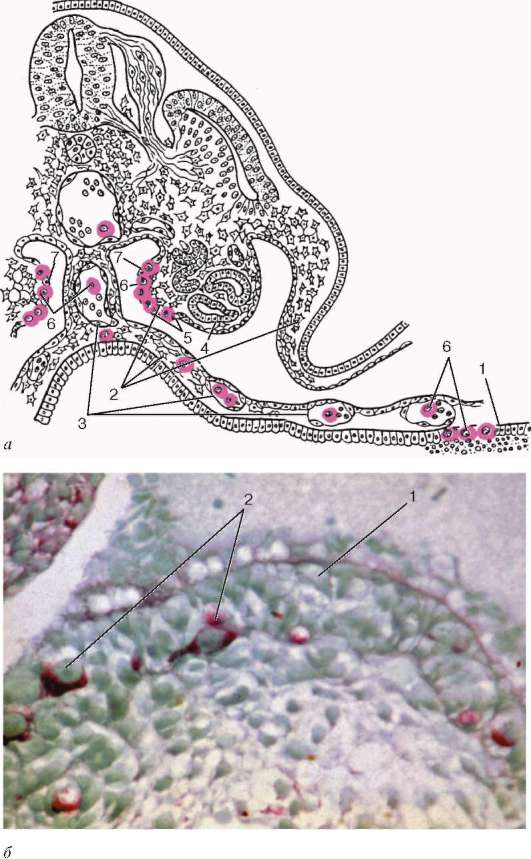
Рис. 20.1. Развитие гонад в эмбриогенезе:
а - схема первичной локализации гоноцитов (окрашены) в желточном мешке зародыша и их последующей миграции в зачаток гонад (по Пэттену, с изменениями А. Г. Кнорре): 1 - эпителий желточного пузырька; 2 - мезенхима; 3 - сосуды; 4 - первичная почка (мезонефрос); 5 - зачаток гонады; 6 - первичные половые клетки; 7 - поверхностный эпителий; б - половой валик зародыша человека 31-32 сут развития (препарат В. Г. Кожухаря): 1 - эпителий полового валика; 2 - гоноциты
продолжая деление, мигрируют по мезенхиме желточного пузырька, задней кишки и с кровотоком в толщу половых валиков. С 33-35 сут из клеток цело-мического эпителия формируются половые тяжи, которые врастают в подлежащую мезенхиму. Тяжи в своем составе содержат гоноциты. Объем гонад увеличивается, они выступают в целомическую полость, обособляются, но остаются связанными с первичной почкой. Клетки последней подвергаются апоптозу, однако часть клеток мезонефроса выселяется в окружающую мезенхиму и вступает в контакт с эпителиоцитами половых тяжей. На этой стадии развития происходит формирование гонадной бластемы, в составе которой находятся гоноциты, клетки целомического происхождения, клетки мезонефрального происхождения и клетки мезенхимы. До 7-й нед гонада не дифференцирована по полу и называется индифферентной.
В процессе развития индифферентной гонады из мезонефрального протока первичной почки, тянущегося от ее тела к клоаке, отщепляется параллельно идущий парамезонефральный проток.
Половые различия в строении индифферентной гонады регистрируются на 6-7-й нед эмбриогенеза человека, причем мужская гонада развивается раньше женской. Среди факторов дифференцировки мужских гонад большую роль играет Y-хромосома, на коротком плече которой локализуется ген половой детерминации (ГПД) и ряд других генов-участников детерминации пола. Экспрессия последних влияет на развитие из клеток целомическо-го происхождения поддерживающих эпителиоцитов (сустентоцитов, клеток Сертоли). Клетки Сертоли, в свою очередь, оказывают влияние на диффе-ренцировку интерстициальных эндокриноцитов (клеток Лейдига). Эти клетки обнаруживаются между половыми тяжами. Эмбриональные источники развития клеток точно не выявлены. К вероятным источникам относят клетки мезонефроса или клетки нейрального происхождения.
Начало выработки клетками Лейдига гормона тестостерона вызывает преобразование мезонефральных протоков в систему мужских половых протоков (выносящие канальцы яичка, проток придатка, семявыносящий проток, семенные пузырьки, семяизвергающий проток). В свою очередь, выработка клетками Сертоли гормона регрессии парамезонефрального протока вызывает апоптоз клеток парамезонефрального протока. На 3-м мес внутриутробного развития в срезах яичек отчетливо видны извитые тяжи, в составе которых гоноциты дифференцируются в сперматогонии.
20.1. МУЖСКАЯ ПОЛОВАЯ СИСТЕМА 20.1.1. Яички
Яички, или семенники (testes), - мужские гонады, в которых образуются мужские половые клетки и мужской половой гормон - тестостерон.
Развитие. При развитии яичка по верхнему краю первичной почки формируется будущая соединительнотканная капсула семенника - белочная
оболочка (tunica albuginea), которая отделяет половые тяжи от полового валика, давшего им начало. В дальнейшем половые тяжи развиваются в семенные канальцы (tubuli seminiferi). Семенные канальцы сливаются с канальцами семявыносящей системы, формирующейся путем перестройки эпителиальной выстилки канальцев мезонефроса. Так, канальцы сети (rete testis), приближаясь к белочной оболочке средостения, сливаются в выносящие канальцы (ductuli efferentes). Выносящие канальцы яичка, собираясь, переходят далее в канал придатка яичка (ductus epididymis), проксимальный отдел которого, многократно извиваясь, образует придаток семенника (epididymis), тогда как его дистальная часть становится семявыносящим протоком (ductus defferes). Парамезонефральный проток в мужском организме атрофируется и сохраняется только краниальный конец (формирует гидатиды, которые прикрепляются к соединительнотканной структуре яичка) и дистальный, превращающийся в мужскую маточку (utriculus prostaticus). Последняя у взрослого мужчины располагается в толще предстательной железы (рис. 20.2).
К концу 3-го мес завершается миграция яичек в малый таз. Опускание яичек в мошонку происходит между 6-м и 8-м мес развития.
В онтогенезе эндокринная функция яичка устанавливается раньше, чем генеративная. Мужской половой гормон - тестостерон начинает вырабатываться у зародыша человека приблизительно с 8-10-й нед внутриутробного периода. На 3-м мес эмбриогенеза клетки Лейдига в яичке достаточно многочисленны и образуют околососудистые скопления. С 6-го мес количество клеток уменьшается и остается неизменным до 2-го мес постнаталь-ной жизни.
Строение. Снаружи большая часть семенника покрыта серозной оболочкой - брюшиной, под которой располагается плотная соединительнотканная оболочка, получившая название белочной (tunica albuginea) (рис. 20.3). На задней поверхности яичка белочная оболочка утолщается, формируя средостение (mediastinum testis), от которого в глубь железы отходят соединительнотканные перегородки (septula testis), разделяющие железу на дольки (около 250 долек), в каждой из которых находится 1-4 извитых семенных канальца (tubuli seminiferi convoluti). Каждый семенной каналец имеет диаметр от 150 до 250 мкм и длину от 30 до 70 см. Приближаясь к средостению, канальцы (300-450 в каждом семеннике) сливаются и становятся прямыми, а в толще средостения соединяются с канальцами сети семенника. Из сети выходит 10-12 выносящих канальцев (ductuli efferens), впадающих в проток придатка (ductus epididymis). В дольках яичка между петлями извитых семенных канальцев находится интерстициальная (соединительная) ткань с гемо- и лимфатическими сосудами. В составе этой ткани, кроме фибробластов, обнаруживаются макрофаги, тучные клетки, а около кровеносных капилляров (преимущественно фенестрированного типа) группами располагаются гормонсинтезирующие клетки Лейдига (интерстици-альные эндокриноциты).
Внутреннюю выстилку канальца образует эпителиосперматогенный слой, расположенный на базальной мембране. Собственная оболочка (tunica propria) канальца представлена базальным слоем (stratum basale), миоидным слоем (stratum myoideum) и волокнистым слоем (stratum fibrosum). Кнаружи от базальной
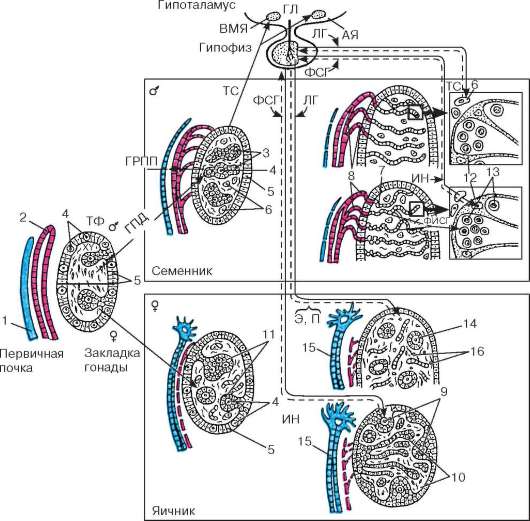
Рис. 20.2. Стадии развития гонад и становления их гормональной регуляции в онтогенезе (по Б. В. Алешину, Ю. И. Афанасьеву, О. И. Бриндаку, Н. А. Юриной): ТФ - телоферрон; ГПД - ген половой детерминации; ГРПП - гормон регрессии парамезонефрального протока; ТС - тестостерон; Э - эстрадиол; П - прогестерон; ФСГ - фолликулостимулирующий гормон; ФИСГ - фактор, ингибирующий спер-матогонии; ЛГ - лютеинизирующий гормон; ИН - ингибин; ГЛ - гонадолиберин; АЯ - аркуатное ядро; ВМЯ - вентромедиальное ядро. 1 - парамезонефральный проток; 2 - мезонефральный проток; 3 - половые тяжи; 4 - гоноциты; 5 - эпителий; 6 - клетки Лейдига; 7 - сеть семенника; 8 - выносящие канальцы семенника; 9 - корковое вещество яичника; 10 - мозговое вещество яичника; 11 - примор-диальные фолликулы; 12 - клетки Сертоли; 13 - сперматогонии; 14 - первичные фолликулы; 15 - маточная труба; 16 - интерстициальные клетки
мембраны эпителия находится сеть коллагеновых волокон базального слоя. Миоидный слой образован миоидными клетками, содержащими актино-вые филаменты. Миоидные клетки обеспечивают ритмические сокращения стенки канальцев. Наружный волокнистый слой состоит из двух частей.
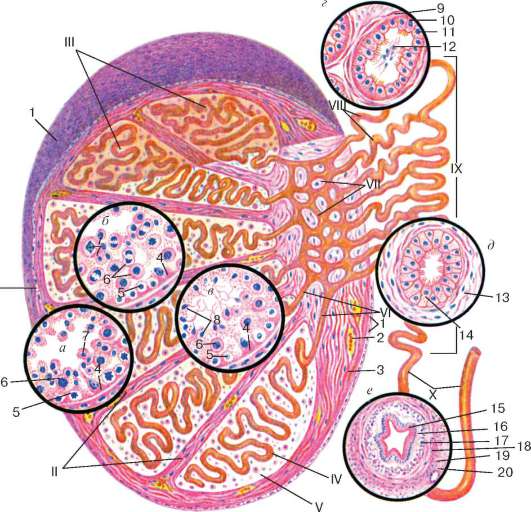
Рис. 20.3. Строение яичка (по Е. Ф. Котовскому):
а - эпителиосперматогенный слой в фазе размножения сперматогоний и в начале фазы роста сперматоцитов; б - эпителиосперматогенный слой в конце фазы роста и в фазе созревания сперматоцитов; в - фаза формирования; г - строение семявыно-сящего канальца яичка; д - строение канала придатка; е - строение семявынося-щего канала. I - оболочки яичка; II - перегородки яичка; III - дольки яичка; IV - извитой семенной каналец; V - интерстициальная ткань; VI - прямые канальцы яичка; VII - сеть яичка; VIII - выносящие канальцы яичка; IX - канал придатка; X - семявыносящий канал. 1 - мезотелий; 2 - кровеносный сосуд; 3 - соединительнотканные клетки; 4 - поддерживающие эпителиоциты (клетки Сертоли); 5 - сперматогонии; 6 - сперматоциты; 7 - сперматиды; 8 - сперматозоиды в просвете извитого семенного канальца; 9 - мышечно-волокнистая оболочка семявыносящего канальца; 10 - реснитчатые эпителиоциты; 11 - кубические эпителиоциты; 12 - сперматозоиды в семявыносящем канальце яичка; 13 - мышечно-фиброзная оболочка канала придатка яичка; 14 - двухрядный реснитчатый эпителий семявыносящего канала; 15 - двухрядный реснитчатый эпителий; 16 - собственная пластинка слизистой оболочки; 17 - внутренний продольный слой мышечной оболочки; 18 - средний циркулярный слой мышечной оболочки; 19 - наружный продольный слой мышечной оболочки; 20 - адвентициальная оболочка
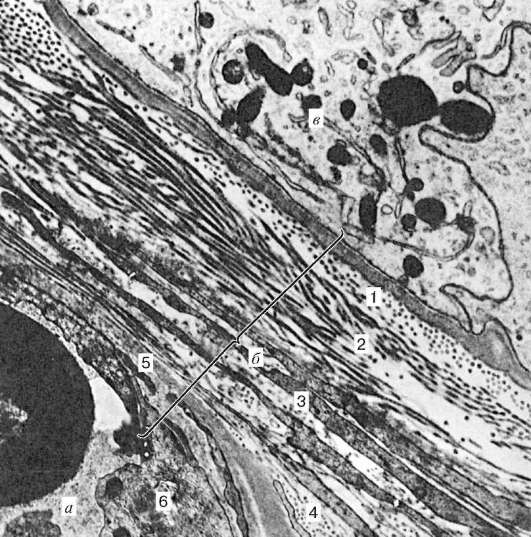
Рис. 20.4. Гематотестикулярный барьер яичка человека. Электронная микрофотография, ув. 24 000 (по А. Ф. Астраханцеву):
а - капилляр; б - гематотестикулярный барьер; в - поддерживающий эпителиоцит. 1 - базальная мембрана; 2 - внутренний волокнистый (базальный) слой; 3 - мио-идный слой; 4 - наружный волокнистый слой; 5 - базальная мембрана эндотелио-цитов; 6 - эндотелий
Непосредственно к миоидному слою примыкает неклеточный слой, образованный базальной мембраной миоидных клеток и коллагеновыми волокнами. За ними расположен слой, состоящий из фибробластоподобных клеток, прилежащий к базальной мембране эндотелиоцитов гемокапилляра.
Избирательность поступления веществ из крови в эпителиосперматоген-ный слой и различия в химическом составе плазмы крови и жидкости из семенных канальцев позволили сформулировать представление о гематоте-стикулярном барьере. Гематотестикулярным барьером называется совокупность структур, располагающихся между просветами капилляров и семенных канальцев (рис. 20.4).
Эпителиосперматогенный слой (epithelium spermatogenicum) формируется двумя клеточными дифферонами: сперматогенными клетками (cellulae spermatogenicae), которые находятся на различных стадиях дифференциров-ки (стволовые клетки, сперматогонии, сперматоциты, сперматиды и сперматозоиды) и поддерживающими эпителиоцитами (клетками Сертоли), или
сустентоцитами (epitheliocytus sustentans). Гистологические элементы двух клеточных дифферонов находятся в тесной морфофункциональной связи.
Поддерживающие эпителиоциты лежат на базальной мембране, имеют пирамидальную форму и достигают своей вершиной просвета извитого семенного канальца. Ядра клеток имеют неправильную форму с инвагинациями, ядрышко (ядрышко и две группы околоядрышкового хроматина). В цитоплазме особенно хорошо развита агранулярная эндоплазматическая сеть, комплекс Гольджи. Присутствуют также микротрубочки, микрофила-менты, лизосомы и особые кристаллоидные включения. Обнаруживаются включения липидов, углеводов, липофусцина. На боковых поверхностях сустентоциты образуют бухтообразные углубления, в которых располагаются дифференцирующиеся сперматогонии, сперматоциты и сперматиды. Между соседними поддерживающими клетками формируются зоны плотных контактов, которые подразделяют весь слой на два отдела - наружный базальный и внутренний адлюминальный. В базальном отделе расположены сперматогонии, имеющие максимальный доступ к питательным веществам, поступающим из кровеносных капилляров. В адлюминальном отделе находятся сперматоциты на стадии мейоза, а также сперматиды и сперматозоиды, которые не имеют доступа к тканевой жидкости и получают питательные вещества непосредственно от поддерживающих эпителиоцитов.
Клетки Сертоли создают микросреду, необходимую для дифференцирующихся половых клеток, изолируют формирующиеся половые клетки от токсических веществ и различных антигенов, препятствуют развитию иммунных реакций. Кроме того, они способны к фагоцитозу дегенерирующих половых клеток и последующему лизису с помощью своего лизо-сомального аппарата. Клетки синтезируют андрогенсвязывающий белок (АСБ), который транспортирует мужской половой гормон к сперматидам. Секреция АСБ усиливается под влиянием ФСГ. Поддерживающие эпителиоциты имеют поверхностные рецепторы ФСГ, а также рецепторы тестостерона и его метаболитов.
Различают два вида клеток Сертоли - светлые клетки, продуцирующие ингибин, тормозящий секрецию ФСГ аденогипофизом, и темные клетки, продуцирующие факторы, стимулирующие деление половых клеток.
Генеративная функция. Сперматогенез
Образование мужских половых клеток (сперматогенез) протекает в извитых семенных канальцах и включает четыре последовательные стадии, или фазы: размножение, рост, созревание и формирование (рис. 20.5).
Начальной фазой сперматогенеза является размножение сперматогоний, занимающих наиболее периферическое (базальное) положение в эпителио-сперматогенном слое. Среди сперматогоний выделяются два типа клеток: 1) стволовые клетки типа А; 2) клетки-предшественники типа В.
Морфологически в популяции стволовых А-сперматогоний различают светлые и темные клетки (см. рис. 20.5). Для обоих видов клеток характерно преобладание в ядрах деконденсированного хроматина и расположение ядрышек около ядерной оболочки. Однако в темных клетках типа А степень
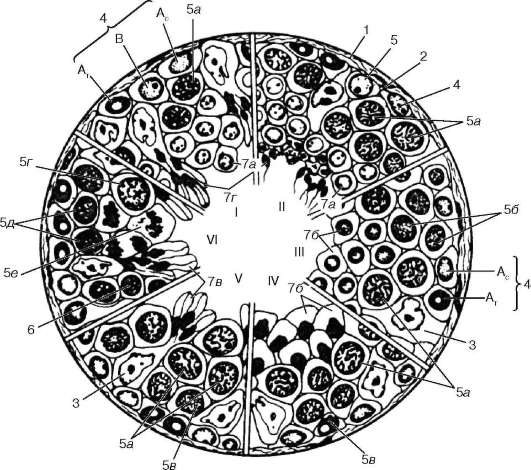
Рис. 20.5. Сперматогенез (по И. Г. Клермону, с изменениями):
I-VI - стадии цикла развития мужских половых клеток в семенных канальцах человека. 1 - соединительнотканная капсула канальца; 2 - базальная мембрана; 3 - поддерживающие клетки; 4 - сперматогонии; тип Ас - светлый; тип АТ - темный; В - тип В; 5 - сперматоциты 1-го порядка: 5а - в пахитене; 5б - в прелептотене; 5в - в лептотене; 5г - в диплотене; 5д - в зиготене; 5е - делящиеся сперматоциты 1-го порядка; 6 - сперматоциты 2-го порядка с интерфазными ядрами; 7 - сперма-тиды на различных стадиях развития (а, б, в, г)
конденсации хроматина больше, чем в светлых. Темные клетки относят к «резервным» медленно обновляющимся стволовым клеткам, а светлые - к быстро обновляющимся клеткам. Для стволовых клеток характерно наличие овальных ядер с диффузно распределенным хроматином, одного или двух ядрышек, большое содержание в цитоплазме рибосом и полисом, малое количество других органелл. Клетки типа В имеют более крупные ядра, хроматин в них не дисперсный, а собран в глыбки.
Часть стволовых клеток типа А после ряда митотических циклов становится источником развития В-сперматогоний - клеток-предшественников первичных сперматоцитов. Сперматогонии типа В после митотического деления не завершают цитокинез и остаются соединенными цитоплазмати-
ческими мостиками. Появление таких спаренных сперматогоний свидетельствует о начале процессов дифференцировки мужских половых клеток. Дальнейшее деление таких клеток приводит к образованию цепочек или групп сперматогоний, соединенных цитоплазматическими мостиками.
В следующей фазе (роста) сперматогонии перестают делиться и дифференцируются в сперматоциты 1-го порядка (первичные сперматоциты). Син-цитиальные группы сперматогоний перемещаются в адлюминальную зону эпителиосперматогенного слоя. В фазе роста сперматогонии увеличиваются в объеме и вступают в первое деление мейоза (редукционное деление). Профаза первого деления длинная и состоит из лептотены, зиготены, пахи-тены, диплотены, диакинеза.
Перед профазой в S-периоде сперматоцита 1-го порядка происходит удвоение количества ДНК. Сперматоцит находится в прелептотене. В леп-тотене хромосомы становятся видимыми в виде тонких нитей. В зиготе-не гомологичные хромосомы расположены парами (конъюгируют), образуя биваленты, между конъюгирующими хромосомами происходит обмен генами. В пахитене (от лат. pachys - толстый) пары конъюгирующих хромосом продолжают укорачиваться и одновременно утолщаться. Гомологичные хромосомы находятся в тесном контакте по всей своей длине. С помощью электронного микроскопа в сперматоцитах 1-го порядка обнаружены синаптонемные комплексы в местах соприкосновения гомологичных хромосом - парные параллельно расположенные ленты шириной около 60 нм, разделенные светлым промежутком шириной около 100 нм. В светлом промежутке видны срединная электронно-плотная линия и пересекающие его тонкие филаменты. Оба конца комплекса присоединены к ядерной оболочке. У человека формируется 23 синаптонемных комплекса. В диплотене гомологичные хромосомы, образующие бивалент, отходят друг от друга, так что становится видна каждая в отдельности, но сохраняют связь в перекрестках хромосом. Одновременно можно видеть, что каждая хромосома состоит из двух хроматид. Дальнейшая спирализация приводит к тому, что пары конъюгирующих хромосом приобретают вид коротких телец разнообразной формы - так называемых тетрад. Поскольку каждая тетрада образована двумя конъюгировавшими хромосомами, число тетрад оказывается вдвое меньше, чем исходное число хромосом, т. е. гаплоидным, - у человека 23 тетрады. В диакинезе хромосомы еще более утолщаются, после чего клетка вступает в метафазу первого деления мейоза (или первого деления созревания) и хромосомы располагаются в экваториальной плоскости. В анафазе обе хромосомы каждого бивалента расходятся к полюсам клетки - по одной к каждому полюсу. Таким образом, в каждой из двух дочерних клеток - сперматоцитов 2-го порядка (вторичных сперматоцитов) - содержится гаплоидное число хромосом (23 у человека), но каждая хромосома представлена диадой.
Второе деление созревания начинается сразу вслед за первым, и происходит как обычный митоз без репликации хромосом. В анафазе второго деления созревания диады сперматоцитов 2-го порядка разъединяются на монады, или одиночные хроматиды, расходящиеся к полюсам. В результате сперма-
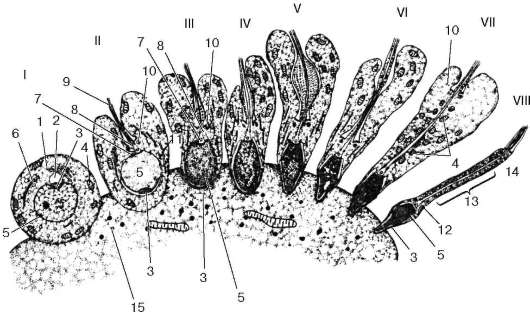
Рис. 20.6. Сперматогенез (дифференцировка сперматиды в сперматозоид) (по Б. В. Алешину):
I - сперматида, внедрившаяся в верхушку поддерживающей клетки; II-VIII - последовательные стадии формирования сперматозоида. 1 - комплекс Гольджи;
2 - акробласт; 3 - зачаток акросомы; 4 - митохондрия; 5 - ядро; 6 - центриоль; 7 - проксимальная центриоль; 8 - дистальная центриоль; 9 - трубки акронемы; 10 - кольцо; 11 - микротрубочки; 12 - шейка; 13 - митохондриальное влагалище; 14 - хвост; 15 - клетка Сертоли
тиды получают столько же монад, сколько было диад в ядрах сперматоцитов 2-го порядка, т. е. гаплоидное число. Сперматоциты 2-го порядка имеют меньшие размеры, чем сперматоциты 1-го порядка, и располагаются в средних и более поверхностных частях эпителиосперматогенного слоя.
Таким образом, каждая исходная сперматогония дает начало 4 спермати-дам с гаплоидным набором хромосом. Сперматиды больше не делятся, но путем сложной перестройки превращаются в зрелые сперматозоиды. Эта трансформация составляет суть фазы формирования (рис. 20.6).
Сперматиды представляют собой небольшие округлые клетки со сравнительно крупными ядрами. Скапливаясь около верхушек поддерживающих клеток, сперматиды частично погружаются в их цитоплазму, что создает условия для формирования сперматозоидов из сперматид. Ядро сперматиды постепенно уплотняется и уплощается.
В сперматидах около ядра располагаются комплекс Гольджи, центросома и скапливаются мелкие митохондрии. Процесс формирования сперматозоида начинается с образования в зоне комплекса Гольджи уплотненной гранулы - акробласта, прилежащего к поверхности ядра. В дальнейшем акробласт, увеличиваясь в размерах, в виде шапочки охватывает ядро, а в середине акробласта дифференцируется уплотненное тельце. Такую структуру называют акросомой. Центросома, состоящая из двух центрио-
лей, перемещается в противоположный конец сперматиды. Проксимальная центриоль прилегает к поверхности ядра, а дистальная разделяется на две части. От передней части дистальной центриоли начинает формироваться жгутик (flagellum), который затем становится осевой нитью развивающегося сперматозоида. Задняя же половина дистальной центриоли принимает вид колечка. Смещаясь вдоль жгутика, это колечко определяет заднюю границу средней, или связующей, части сперматозоида.
Цитоплазма по мере роста жгутика сползает с ядра и сосредоточивается в связующей части. Митохондрии располагаются спиралеобразно между проксимальной центриолью и колечком.
Цитоплазма сперматиды во время ее превращения в спермий сильно редуцируется. В области головки она сохраняется только в виде тонкого слоя, покрывающего акросому; небольшое количество цитоплазмы остается в области связующей части и, наконец, она очень тонким слоем покрывает жгутик. Часть цитоплазмы сбрасывается и распадается в просвете семенного канальца либо поглощается клетками Сертоли. Клетки Сертоли вырабатывают жидкость, накапливающуюся в просвете извитого семенного канальца. В эту жидкость попадают, высвобождаясь из верхушек поддерживающих клеток, сформировавшиеся сперматозоиды и вместе с ней уходят в дистальные части канальца.
Сперматогенез у человека длится около 64-75 сут и протекает на протяжении извитого семенного канальца волнообразно. Поэтому набор клеток в составе сперматогенного дифферона по ходу канальца меняется в соответствии с фазой сперматогенеза.
Реактивность и регенерация. Сперматогенез чрезвычайно чувствителен к повреждающим воздействиям. При различных интоксикациях, авитаминозах, недостаточности питания и других условиях (особенно при воздействии ионизирующим излучением) сперматогенез ослабевает и даже прекращается. Аналогичные деструктивные процессы развиваются при крипторхизме (когда семенники не опускаются в мошонку, а остаются в брюшной полости), длительном пребывании организма в среде с высокой температурой, лихорадочных состояний и особенно после перевязки или перерезки семя-выводящих каналов. Деструктивный процесс при этом поражает в первую очередь формирующиеся сперматозоиды и сперматиды. Последние набухают, нередко сливаются в характерные округлые массы - так называемые семенные шары, плавающие в просвете канальца. Так как сперматогонии и сперматоциты 1-го порядка при этом сохраняются более длительно, то восстановление сперматогенеза после прекращения действия повреждающего агента иногда оказывается возможным.
Клетки Сертоли в указанных обстоятельствах сохраняются и даже гипертрофируются, а клетки Лейдига часто увеличиваются в количестве и образуют большие скопления между запустевающими семенными канальцами.
Эндокринные функции
В рыхлой соединительной ткани между петлями извитых канальцев располагаются интерстициальные эндокриноциты (гландулоциты, клет-
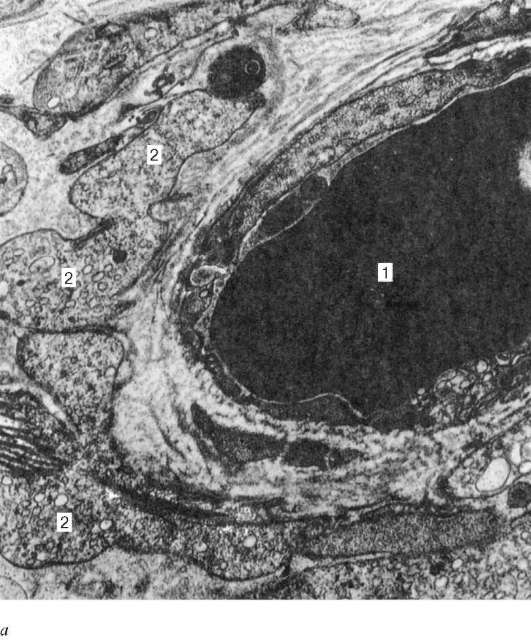
Рис. 20.7. Интерстициальные эндокриноциты (клетки Лейдига) яичка человека (по А. Ф. Астраханцеву):
а - капилляр интерстициальной соединительной ткани с прилежащими эндокри-ноцитами, увеличение 22 000; б - эндокриноцит, увеличение 10 000; в - фрагмент эндокриноцита, увеличение 26 000. 1 - капилляр; 2 - фрагменты цитоплазмы эндокриноцитов; 3 - ядро эндокриноцита; 4 - липидная капля; 5 - агранулярная эндоплазматическая сеть; 6 - строма
ки Лейдига), скапливающиеся здесь вокруг кровеносных капилляров (рис. 20.7). Эти клетки сравнительно крупные, округлой или многоугольной формы, с ацидофильной цитоплазмой, вакуолизированной по периферии, содержащей гликопротеидные включения, а также глыбки гликогена и белковые кристаллоиды в виде палочек или лент. С возрастом в цитоплазме клеток Лейдига начинает откладываться пигмент. Хорошо развитая гладкая эндоплазматическая сеть, многочисленные митохондрии с трубчатыми кри-стами указывают на способность клеток Лейдига к выработке стероидных веществ, в данном случае - мужского полового гормона.
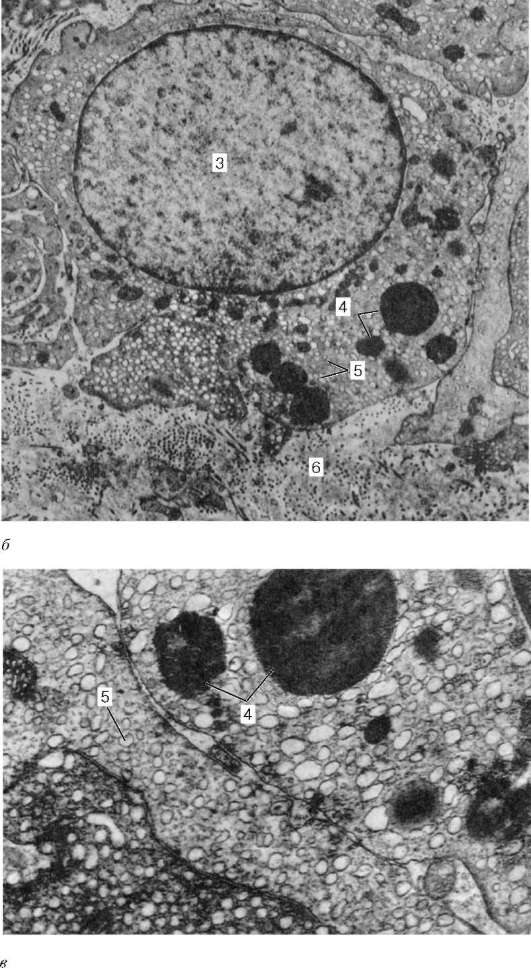
Рис. 20.7. Продолжение (обозначения см. выше)
20.1.2. Семявыносящие пути
Семявыносящие пути составляют систему канальцев (см. рис. 20.3) яичка и его придатков, по которым сперма (сперматозоиды и семенная жидкость) продвигается в мочеиспускательный канал.
Отводящие пути начинаются прямыми канальцами яичка (tubuli seminiferi recti), впадающими в сеть яичка (rete testis), располагающуюся в средостении (mediastinum). От сети отходят 12-15 извитых выносящих канальцев (ductuli effe-rentes testis), которые открываются в единый проток придатка (ductus epi-didymidis) в области головки придатка. Этот проток, многократно извиваясь, формирует тело придатка и в нижней хвостовой части становится прямым семявыносящим протоком (ductus deferens). Последний формирует ампулу семя-выносящего протока. За ампулой в проток открывается выносящий проток семенного пузырька, после чего семявыносящий проток продолжается в семя-извергающий проток. Семяизвергающий проток (ductus ejaculatorius) проникает в предстательную железу и открывается в простатическую часть мочеиспускательного канала.
Все семявыводящие пути построены по общему плану и состоят из слизистой, мышечной и адвентициальной оболочек. Эпителий, выстилающий эти канальцы, обнаруживает признаки железистой деятельности, особенно выраженной в головке придатка.
В прямых канальцах яичка эпителий образован клетками призматической формы. В канальцах сети семенника в эпителии преобладают кубические и плоские клетки. В эпителии семявыводящих канальцев чередуются группы реснитчатых эпителиоцитов с железистыми клетками, секретирую-щими по апокринному типу.
В придатке яичка эпителий протока становится двухрядным. В его составе находятся столбчатые эпителиоциты, несущие на своих апикальных верхушках стереоцилии, а между базальными частями этих клеток располагаются вставочные эпителиоциты. Эпителий протока придатка принимает участие в выработке жидкости, разбавляющей сперму во время прохождения сперматозоидов, а также в образовании гликокаликса - тонкого слоя, которым покрываются сперматозоиды. Удаление гликокаликса при эякуляции приводит к активизации сперматозоидов (капацитация). Одновременно придаток семенника оказывается резервуаром для накапливающейся спермы.
Продвижение спермы по семявыводящим путям обеспечивается сокращением мышечной оболочки, образованной циркулярным слоем гладких мышечных клеток.
Проток придатка далее переходит в семявыносящий проток (ductus deferens). Слизистая оболочка протока представлена эпителием и собственной пластинкой слизистой оболочки. Эпителий - многорядный столбчатый - включает базальные (малодифференцированные) клетки, столбчатые клетки со стереоцилиями, а также клетки, богатые митохондриями. Собственная пластинка слизистой оболочки содержит много эластических волокон. Мышечная оболочка состоит из трех слоев - внутреннего продольно-
го, среднего циркулярного и наружного продольного. В толще мышечной оболочки располагается нервное сплетение, образованное скоплением ганглиозных клеток, иннервирующих пучки гладких мышечных клеток. Сокращениями их обеспечивается эякуляция спермы. В связи со значительным развитием мышечной оболочки слизистая оболочка семявынося-щего протока собирается в продольные складки (см. рис. 20.3). Дистальный конец этого протока ампулообразно расширен. Снаружи семявыводящие пути на всем протяжении покрыты соединительнотканной адвентициаль-ной оболочкой.
Ниже места соединения семявыносящего протока и семенных пузырьков начинается семявыбрасывающий проток. Он проникает через предстательную железу и открывается в мочеиспускательный канал. В дистальной части протока эпителий становится многослойным переходным. В отличие от семявыносящего протока семявыбрасывающий проток не имеет столь выраженной мышечной оболочки. Наружная оболочка его срастается с соединительнотканной стромой предстательной железы.
Васкуляризация. Кровоснабжение семенника обеспечивается через ветвь внутренней семенной артерии, входящей в состав семенного канатика в средостение, где она разветвляется на сеть капилляров, проникающих по соединительнотканным перегородкам внутрь долек и оплетающих извитые семенные канальцы. Вокруг этих капилляров скапливаются интерстици-альные клетки.
Лимфатические капилляры также образуют сеть между канальцами семенника, а далее образуют выносящие лимфатические сосуды.
Иннервация. Нервные волокна, как симпатические, так и парасимпатические, проникают в семенник вместе с кровеносными сосудами. В паренхиме семенника рассеяны многочисленные чувствительные нервные окончания. Нервные импульсы, поступающие в семенник, способны оказывать некоторое влияние на его генеративную и эндокринную функции, но основная регуляция его деятельности осуществляется гуморальными влияниями гонадотропных гормонов аденогипофиза.
Возрастные изменения. Генеративная функция семенника начинается еще в препубертатном возрасте, но в этот период сперматогенез останавливается в начальных стадиях. Полное завершение сперматогенеза (образование сперматозоидов) наступает лишь после достижения половой зрелости - пубертатного периода. У новорожденного семенные канальцы еще имеют вид сплошных клеточных тяжей, состоящих из поддерживающих эпителиоцитов и сперматогоний. Такое строение семенные канальцы сохраняют в течение первых 4 лет постнатального периода развития мальчика. Просвет в семенных канальцах появляется лишь к 7-8 годам жизни. В это время количество сперматогоний значительно увеличивается, а к 9 годам среди них появляются одиночные сперматоциты 1-го порядка, что указывает на начало второй стадии сперматогенеза - стадии роста. Между 10 и 15 годами семенные канальцы становятся извитыми: в их просветах обнаруживаются сперматоциты 1-го и 2-го порядка и даже сперматиды, а клетки Сертоли достигают полной зрелости. К 12-14 годам заметно усиливаются
рост и развитие выводящих протоков и придатка семенника, что свидетельствует о поступлении в циркуляцию мужского полового гормона в достаточно высокой концентрации. В соответствии с этим в семенниках отмечается большое количество крупных клеток Лейдига. Возрастная инволюция семенника у мужчин происходит между 50 и 80 годами. Она проявляется в нарастающем ослаблении сперматогенеза, разрастании соединительной ткани. Однако даже в пожилом возрасте в некоторых семенных канальцах сохраняется сперматогенез и их строение остается нормальным.
Параллельно прогрессирующей атрофии эпителиосперматогенного слоя увеличивается деструкция клеток Лейдига, вследствие чего ослабевает продукция мужского полового гормона, а это в свою очередь оказывается причиной возрастной атрофии предстательной железы и частично наружных половых органов. С возрастом в цитоплазме клеток Лейдига начинает откладываться пигмент.
20.1.3. Добавочные железы мужской половой системы
К добавочным железам мужской половой системы относятся семенные пузырьки, предстательная железа, бульбоуретральные железы.
Семенные пузырьки
Семенные пузырьки - парные мешковидные структуры, развиваются как выпячивания стенки семявыносящего протока в его дистальной (верхней) части. Эти железистые органы, вырабатывающие жидкий слизистый секрет, слабощелочной реакции, богатый фруктозой, который примешивается к сперме и разжижает ее и простагландины. В стенке пузырьков имеются оболочки, границы между которыми выражены нечетко: слизистая, мышечная, адвентициальная (рис. 20.8). Слизистая оболочка собрана в многочисленные разветвленные складки, местами срастающиеся между собой, вследствие чего она приобретает ячеистый вид. Слизистая оболочка покрыта однослойным столбчатым эпителием, лежащим на тонкой базальной мембране. В составе эпителия различают столбчатые и базаль-ные эпителиоциты. В собственной пластинке слизистой оболочки много эластических волокон. В слизистой оболочке расположены терминальные отделы желез альвеолярного типа, состоящие из слизистых экзокриноцитов (exocrinocytus mucosus).
Мышечная оболочка хорошо выражена и состоит из двух слоев гладких мышечных клеток - внутреннего циркулярного и наружного продольного. Адвентициальная оболочка состоит из плотной волокнистой соединительной ткани с большим содержанием эластических волокон.
Предстательная железа
Предстательная железа, или простата (prostata), - мышечно-железистый орган, охватывающий верхнюю часть мочеиспускательного канала (уре-
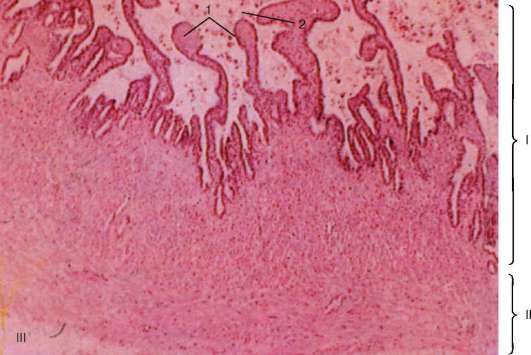
Рис. 20.8. Семенной пузырек:
I - слизистая оболочка; II - мышечная оболочка; III - наружная соединительнотканная оболочка. 1 - складки слизистой оболочки; 2 - секрет в просвете железы
тры), в которую открываются протоки многочисленных простатических желез.
Развитие. У человека формирование предстательной железы начинается на 11-12-й нед внутриутробного развития, при этом из эпителия мочеиспускательного канала в окружающую мезенхиму врастают 5-6 тяжей. В первой половине пренатального эмбриогенеза из разрастающихся эпителиальных тяжей развиваются преимущественно альвеолярно-трубчатые простатические железы. В процессе развития многослойный эпителий желез под влиянием андрогенов становится многорядным, в составе которого возникают диффероны секреторных, слизистых и эндокринных клеток. Базальные эпителиоциты являются камбиальными. Со второй половины эмбриогенеза преобладает рост гладкой мышечной ткани и соединительнотканных прослоек предстательной железы. Просветы в эпителиальных тяжах появляются в конце предплодного периода развития зародыша. Обособленно от указанных желез из эпителия уретры возникают небольшие железы, располагающиеся между предстательной маточкой и семявы-брасывающим протоком.
Строение. Предстательная железа - дольчатая железа, покрытая тонкой соединительнотканной капсулой. Ее паренхима состоит из многочисленных отдельных желез, выводные протоки которых открываются в мочеиспускательный канал. Различают слизистые (периуретральныё), подслизистые
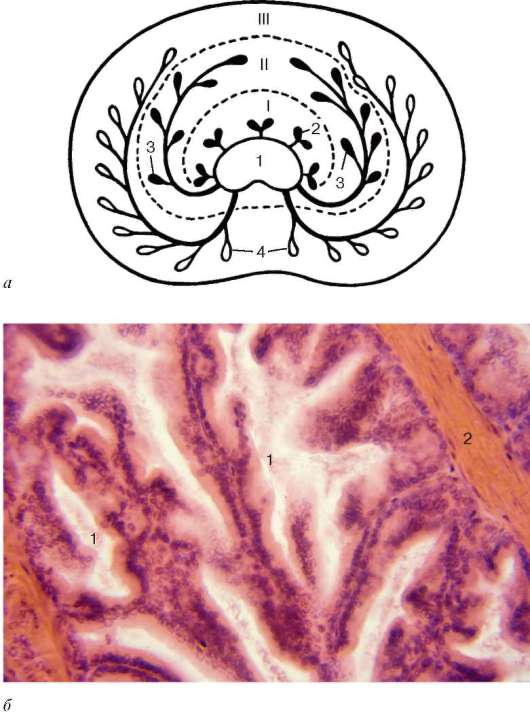
Рис. 20.9. Предстательная железа:
а - схема строения железы (по Дж. Гранту, с изменениями): I - периуретральная железистая зона (слизистая оболочка); II - промежуточная зона (подслизистая основа); III - периферическая зона; 1 - мочеиспускательный канал; 2 - мелкие железы периуретральной зоны; 3 - железы промежуточной зоны; 4 - железы периферической зоны (главные железы); б - микрофотография: 1 - концевые отделы желез; 2 - гладкие миоциты и соединительнотканная строма
(промежуточные) и главные железы, которые располагаются вокруг мочеиспускательного канала тремя перечисленными выше группами.
В периуретральной железистой зоне в составе слизистой оболочки непосредственно вокруг мочеиспускательного канала располагаются мелкие слизистые железы. В переходной зоне в соединительной ткани подслизистой основы в виде кольца залегают подслизистые железы. Главные железы зани-
мают остальную, большую часть органа. Концевые отделы альвеолярно-трубчатых предстательных желез образованы высокими экзокриноцитами предстательной железы (exocrinocytus prostaticus), или простатоцитами (prosta-tocytus), между основаниями которых располагаются мелкие базальные эпителиоциты (рис. 20.9). Кроме того, в эпителии желез и выводных протоков встречаются эндокриноциты предстательной железы, относящиеся к дисперсной эндокринной системе (APUD-серии клеток), по механизму паракрин-ной регуляции действующие на секреторную и сократительную активность тканей предстательной железы. Выводные протоки перед впадением в уретру расширяются в виде ампул неправильной формы, выстланных многорядным столбчатым эпителием. Мышечно-эластическую строму железы (stroma myoelasticum) образуют рыхлая соединительная ткань и мощные пучки гладких мышечных клеток, радиально расходящиеся от центра предстательной железы и разделяющие ее на дольки. Каждая долька и каждая железа окружены продольными и циркулярными слоями гладких мышечных клеток, которые, сокращаясь, выбрасывают секрет из предстательных желез в момент эякуляции.
В месте впадения семявыносящего канала в мочеиспускательный канал в предстательной железе расположен семенной бугорок (colliculus seminalis). С поверхности он выстлан переходным эпителием, а его основу составляют соединительная ткань, богатая эластическими волокнами, и гладкие мышечные клетки. Благодаря наличию многочисленных нервных окончаний семенной бугорок отличается наибольшей чувствительностью. Возбуждение семенного бугорка вызывает его эрекцию, благодаря чему предотвращается попадание эякулята в мочевой пузырь.
Позади семенного бугорка располагается предстательная маточка (utriculus prostaticus), открывающаяся на поверхность семенного бугорка.
Функции предстательной железы многообразны. Вырабатываемый простатой секрет, выбрасываемый во время эякуляции, содержит иммуноглобулины, ферменты, витамины, лимонную кислоту, ионы цинка и др. Секрет участвует в разжижении эякулята.
Структуру и функции железы контролируют гормоны гипофиза, андро-гены, эстрогены. Предстательная железа чувствительна к гормонам яичка. Она находится в зависимости от тестостерона семенников и атрофируется после кастрации. Тестостерон проникает в клетки путем диффузии, где подвергается активному метаболизму и превращению в дегидротестостерон (ДГТ). После связывания в клетке со специфическим рецептором андрогена ДГТ проникает в ядро, где активирует образование специфических ферментов и белков простаты. Кроме того, железа оказывает влияние на половую дифференцировку гипоталамуса (участвует в предопределении его диффе-ренцировки по мужскому типу), а также вырабатывает фактор, стимулирующий рост нервных волокон.
Васкуляризация. Кровоснабжение простаты осуществляется за счет ветвей артерии прямой кишки и мочевого пузыря. Венозная система состоит из многочисленных анастомозирующих вен, образуя мочепузырное предстательное венозное сплетение.
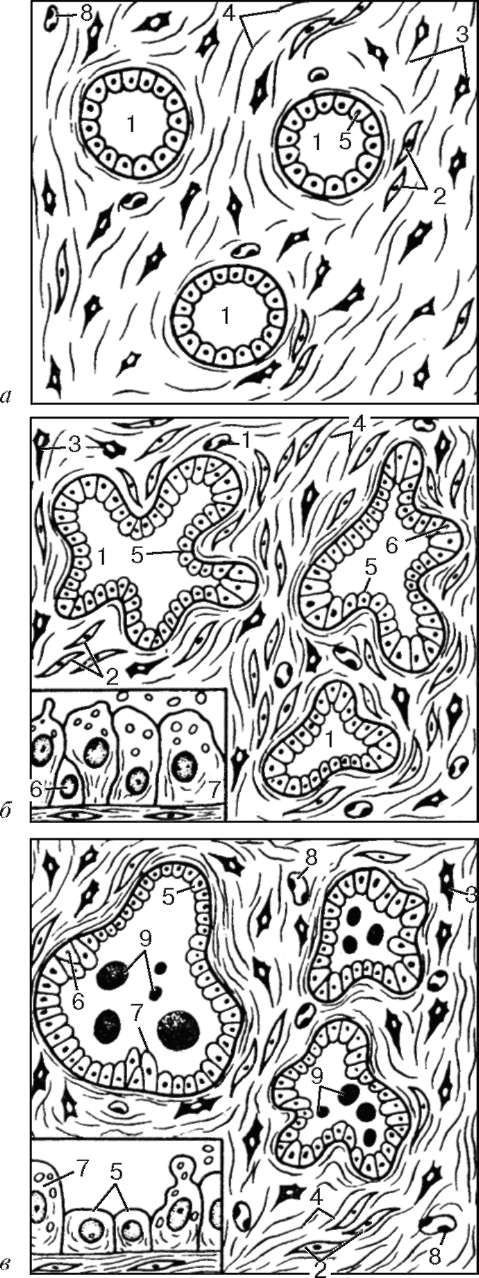
Рис. 20.10. Возрастные изменения предстательной железы (по Б. В. Троценко): а - срез предстательной железы ребенка; б - срез предстательной железы в зрелом возрасте; в - срез предстательной железы в пожилом возрасте. 1 - концевые отделы желез; 2 - гладкие миоциты; 3 - фибробласты; 4 - волокна соединительной ткани; 5 - кубические клетки концевых отделов; 6 - базальные эпителиоциты; 7 - эпителиальные клетки столбчатой формы; 8 - капилляры; 9 - конкреции (крахмальные тельца) в секреторных отделах предстательной железы
Возрастные изменения. Предстательная железа в течение всей жизни человека претерпевает возрастную перестройку, связанную со снижением образования половых гормонов и проявляющуюся сдвигами в соотношении между железистым эпителием, соединительнотканными и гладкомышечными клетками этого органа.
Секреторные отделы предстательной железы ребенка имеют эпителий, состоящий из двух видов клеток - столбчатых и базальных эпителио-цитов (рис. 20.10). Соединительная ткань образует массивные пучки по ходу выводных протоков и значительно уплотняется вокруг секреторных отделов. В ней преобладают фибробласты, макрофаги и колла-геновые волокна. В строме сравнительно мало гладких мышечных клеток.
В период полового созревания в цитоплазме железистых клеток концевых отделов усиливаются секреторные процессы. Эпителий становится высоким. В период наибольшей функциональной активности (в возрасте 20-35 лет) в предстательной железе секреторные элементы преобладают над соединительнотканными, возрастает синтез гликогена, гликозаминогликанов и гликопротеидов. В дальнейшем (в 35-60 лет) некоторые железистые дольки начинают атрофироваться, а соединительная ткань разрастается
и уплотняется. Железистый эпителий постепенно становится низким (см. рис. 20.10). В полости секреторных отделов образуются и скапливаются простатические конкреции, которые особенно часто встречаются в старческом возрасте.
Бульбоуретральные железы
Бульбоуретральные (куперовы) железы - парные железы, расположенные по обеим сторонам основания полового члена по краям луковицы мочеиспускательного канала. По своему строению являются альвеолярно-трубчатыми, открывающимися своими протоками в верхней части мочеиспускательного канала. Их концевые отделы и выводные протоки имеют неправильную форму. Концевые трубчато-альвеолярные отделы местами соединяются друг с другом и состоят из слизистых экзокриноцитов (exocrinocytus bulboure-tralis). Снаружи располагаются миоэпителиоциты. В расширенных альвеолах этих желез эпителий бывает чаще всего уплощенным, в остальных отделах железы - кубическим или столбчатым. Клетки эпителия заполнены капельками мукоида и своеобразными палочковидными включениями. Между концевыми отделами располагаются прослойки рыхлой волокнистой неоформленной соединительной ткани, содержащей пучки гладких мышечных клеток.
20.1.4. Половой член
Половой член (penis) - копулятивный орган. Его основная масса образована тремя пещеристыми (кавернозными) телами, которые, переполняясь кровью, становятся ригидными и обеспечивают эрекцию. Снаружи пещеристые тела окружены белочной оболочкой (tunica albuginea), образованной плотной волокнистой соединительной тканью. Эта ткань изобилует эластическими волокнами и содержит значительное количество гладких мышечных клеток. В середине нижнего кавернозного тела проходит мочеиспускательный канал, по которому выделяется сперма. Он разделяется на предстательную часть (pars prostatica), перепончатую часть (pars membranacea) и губчатую часть ( pars spongiosa).
Мочеиспускательный канал имеет хорошо выраженную слизистую оболочку. Ее эпителий в предстательной железе переходный, в мембранозной части - многорядный призматический, а начиная с области ладьевидной ямки в губчатой части эпителий уретры становится многослойным плоским и обнаруживает признаки ороговения (рис. 20.11). В многорядном эпителии встречаются многочисленные бокаловидные и немногочисленные эндокринные клетки. Под эпителием располагается собственная пластинка слизистой оболочки, богатая эластическими волокнами. В рыхлой волокнистой ткани этого слоя проходит сеть венозных сосудов, имеющая связь с полостями кавернозного тела уретры. В губчатой части мочеиспускательного канала в слизистой оболочке располагаются трубчато-альвеолярные железы мочеиспускательного канала (уретральные). Эпителий желез состоит из столбча-
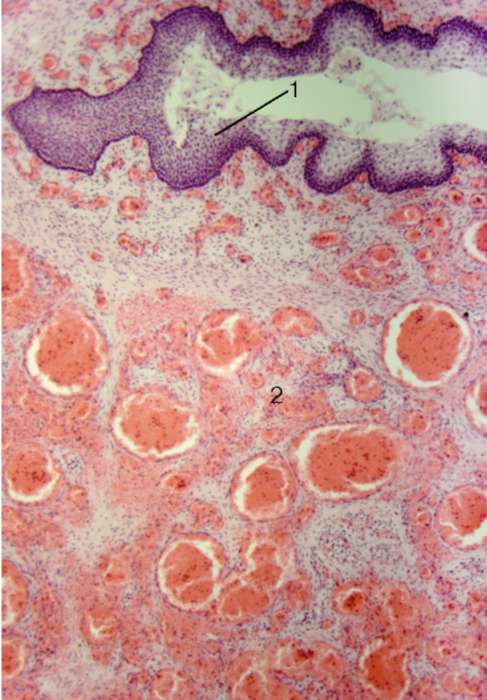
Рис. 20.11. Строение мочеиспускательного канала:
1 - многослойный плоский эпителий;
2 - кавернозное тело
тых, базальных и эндокринных клеток. В подслизистой основе имеется сеть широких венозных сосудов.
Мышечная оболочка мочеиспускательного канала хорошо развита в его простатической части, где она состоит из внутреннего продольного и наружного циркулярного слоев гладких миоцитов. При переходе перепончатой части мочеиспускательного канала в его пещеристую часть мышечные слои постепенно истончаются и сохраняются только одиночные пучки мышечных клеток.
Основа головки полового члена состоит из плотной волокнистой соединительной ткани, в которой заложена сеть анастомозирующих вен, переполняющихся кровью во время эрекции. В толстой стенке их располагаются продольные и циркулярно расположенные пучки гладких мышечных клеток. Кожа, покрывающая головку полового члена, тонкая. В ней расположены сальные (препуциальные) железы (gll. sebacea preputiales). Васкуляризация. Артерии, приносящие кровь к кавернозным телам, имеют толстую мышечную оболочку и широкий просвет. Артерия полового члена, снабжающая его кровью, распадается на несколько крупных ветвей, которые проходят по перегородкам пещеристой ткани. При спокойном состоянии полового члена они спиралевидно закручены и поэтому называются завитыми, или улитковыми (аа. helicinae). Во внутренней оболочке этих артерий имеются утолщения, состоящие из пучков гладких мышечных клеток, а также коллагеновых волокон. Эти утолщения оказываются своего рода клапанами, закрывающими просвет сосуда. Вены тоже отличаются толстой стенкой, хорошо выраженным мышечным слоем во всех оболочках: продольным - во внутренней оболочке, циркулярным - в средней и продольным - в наружной адвентициальной оболочке. Сосудистые полости кавернозных тел, сеть которых находится между артериями и венами, имеют очень тонкие стенки, выстланные эндотелием. Кровь из полостей уходит по небольшим тонкостенным сосудам, впадающим в глубокие вены. Эти сосуды играют роль клапанов или шлюзов, так как во время эрекции стенка вены сокращается и зажимает их просвет, что препятствует оттоку крови из полостей. В сосудистой системе полового члена обнаружены также типичные артериоловенулярные анастомозы.
Иннервация. Симпатические безмиелиновые волокна в половом члене образуют сплетение, иннервирующее пучки гладких мышечных клеток в стенках сосудов и в перегородках между сосудистыми полостями пещеристых тел. В коже полового члена и слизистой оболочке мочеиспускательного канала рассеяны многочисленные рецепторы. Среди них имеются свободные ветвящиеся окончания, залегающие в эпителии головки полового члена и крайней плоти, а также в субэпителиальной ткани.
Особенно многочисленны и разнообразны в тканях полового члена несвободные инкапсулированные окончания. К ним относятся осязательные тельца в сосочковом слое крайней плоти и головке полового члена, генитальные тельца, пластинчатые в глубоких слоях соединительнотканной части полового члена и в белочной оболочке пещеристых тел.
Гормональная регуляция деятельности мужской половой системы
Обе функции гонад (генеративная и гормонообразовательная) активируются аденогипофизарными гонадотропинами - фоллитропином (фолликулостимулирующий гормон) и лютропином (лютеинизирующий гормон). Фоллитропин влияет преимущественно на эпителиоспермато-генный слой, герминативную функцию семенника, а функции клеток Лейдига регулируются лютропином. Однако в действительности взаимодействия гонадотропинов более сложны. Доказано, что регуляция герминативной функции семенника осуществляется совместным влиянием фоллитропина и лютропина. Пептидные ингибины угнетают фоллику-лостимулирующую функцию гипофиза (по механизму отрицательной обратной связи), что приводит к ослаблению влияния, оказываемого на семенники фоллитропином, но не препятствует действию на него лютро-пина. Тем самым ингибин регулирует взаимодействие обоих аденогипо-физарных гонадотропинов, проявляющееся в регуляции ими деятельности семенника (рис. 20.12).
20.2. ЖЕНСКАЯ ПОЛОВАЯ СИСТЕМА
Женская половая система включает половые железы - яичники и органы полового тракта (маточные трубы, матка, влагалище, наружные половые органы).
20.2.1. Яичники
Яичники (парный орган) выполняют генеративную (развитие женских половых клеток) и эндокринную (выработка половых гормонов) функции.
Развитие. Индифферентная гонадная бластема, в составе которой находятся гоноциты, тяжи клеток целомического происхождения (половые шнуры), канальцы первичной почки (мезонефроса) и клетки мезенхимы,
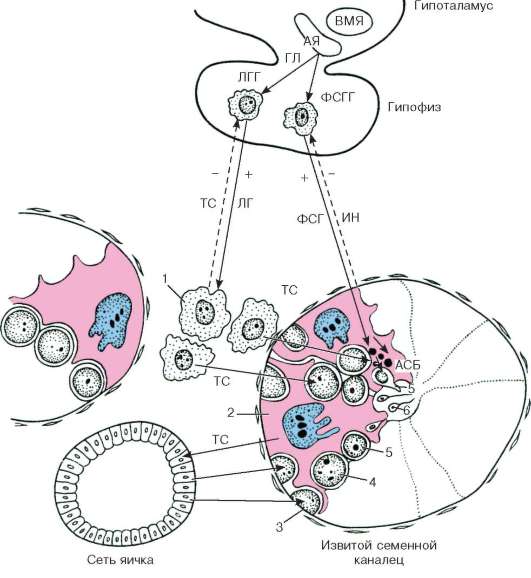
Рис. 20.12. Гормональная регуляция сперматогенеза (схема Б. В. Алешина, Ю. И. Афанасьева, О. И. Бриндака, Н. А. Юриной):
АСБ - андрогенсвязывающий белок; АЯ - аркуатное ядро; ВМЯ - вентромедиаль-ное ядро; ГЛ - гонадолиберин; ИН - ингибин; ТС - тестостерон; ЛГ - лютеини-зирующий гормон; ЛГГ - ЛГ-гонадотропоциты; ФСГ - фолликулостимулирующий гормон; ФСГГ - ФСГ-гонадотропоциты. 1 - клетка Лейдига; 2 - клетка Сертоли; 3 - сперматогонии; 4 - сперматоциты; 5 - сперматиды; 6 - сперматозоиды. Сплошные и прерывистые стрелки - обратные связи («+» - взаимодействия)
развивается в яичник с 6-й нед эмбриогенеза. При этом мезонефральные протоки атрофируются, а клетки канальцев первичной почки формируют клеточные тяжи и канальцы внутрияичниковой сети (rete ovarii). Пара-мезонефральные (мюллеровы) протоки развиваются в маточные трубы, концы которых расширяются в воронки, охватывающие яичники. Нижние части
парамезонефральных протоков, сливаясь, приводят к образованию матки и влагалища.
К началу 7-й нед развития яичник отделяется от мезонефроса углубляющимися бороздами, и начинают формироваться ворота органа, через которые проходят кровеносные и лимфатические сосуды и нервы. У 7-8-недельных эмбрионов заметно формирование коркового вещества яичника. Между половыми шнурами постепенно прорастает мезенхима, разделяющая их на отдельные островки клеток. В результате размножения овогоний, особенно на 3-4-м мес эмбриогенеза, количество половых клеток прогрессивно увеличивается. Для этого периода развития характерна неполная цитотомия овогоний, что необходимо для синхронизации митотических циклов групп клеток. В последующем каждая половая клетка окружается одним слоем плоских эпителиальных клеток и называется примордиальным фолликулом. С 3-го мес развития около половины овогоний вступают в малый рост и в профазу 1-го деления мейоза и именуются овоцитами 1-го порядка, или первичными овоцитами. Остальные овогонии продолжают размножаться. Однако к моменту рождения остается лишь 4-5 % общего числа овогоний вследствие их гибели. Сохранившиеся в яичнике половые клетки переходят в профазу 1-го деления мейоза, но останавливаются на стадии диплотены. В этом состоянии половые клетки (примордиальные фолликулы) сохраняются до периода полового созревания. В целом к моменту рождения число половых клеток составляет около 300 000-400 000.
Мозговое вещество яичника развивается из разрастающейся мезенхимы. Эндокринная функция яичников начинает проявляться по достижении женским организмом половой зрелости. Первичный малый рост фолликулов не зависит от гормонов гипофиза.
Яичник взрослой женщины. С поверхности орган окружен белочной оболочкой (tunica albuginea), образованной плотной волокнистой соединительной тканью, покрытой мезотелием (рис. 20.13). Свободная поверхность мезотелия снабжена микроворсинками. В цитоплазме определяются умеренно развитая гранулярная эндоплазматическая сеть, митохондрии и другие органеллы. Под белочной оболочкой располагается корковое вещество, а глубже - мозговое вещество.
Корковое вещество (cortex ovarii) образовано так называемыми овариаль-ными фолликулами различной степени зрелости, расположенными в соединительнотканной строме. Термином «овариальный фолликул» обозначается клеточно-тканевый комплекс, состоящий из половой клетки и окружающего ее эпителия, который претерпевает изменения в процессе поступательного развития примордиального фолликула в преовуляторный фолликул. Примордиальные фолликулы состоят из овоцита в диплотене профазы 1-го деления мейоза, окруженного одним слоем плоских эпителиальных клеток и базальной мембраной (см. рис. 20.13). Ядра эпителиальных клеток удлиненные, с инвагинациями. По мере роста фолликулов увеличивается размер половой клетки. Вокруг плазмолеммы появляется неклеточная оболочка из гликозаминогликанов - прозрачная зона, или оболочка (zona seu capsula pellucida), снаружи от которой располагается слой фолликулярных эпите-
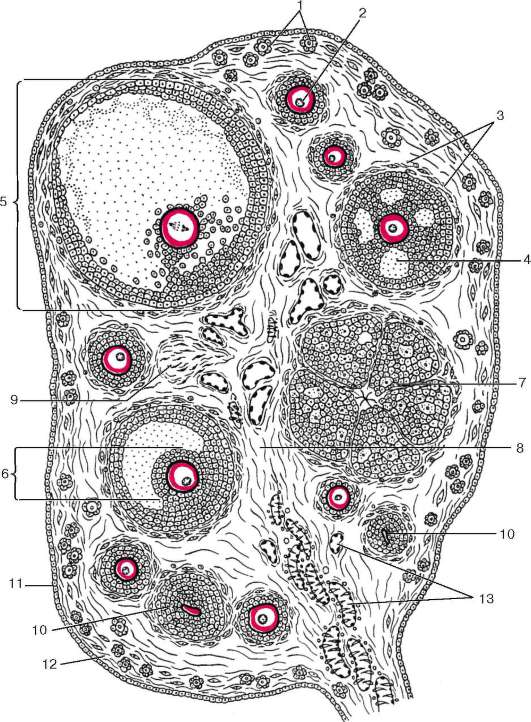
Рис. 20.13. Строение яичника (по Ю. И. Афанасьеву):
1 - примордиальные фолликулы в корковом веществе; 2 - растущий фолликул; 3 - соединительнотканная оболочка фолликула; 4 - фолликулярная жидкость; 5 - зрелый фолликул; 6 - яйценосный бугорок; 7 - желтое тело; 8 - интерстициаль-ная ткань; 9 - беловатое тело; 10 - атретический фолликул; 11 - поверхностный эпителий; 12 - белочная оболочка; 13 - кровеносные сосуды в мозговом веществе яичника
лиоцитов кубической или призматической формы на базальной мембране. В цитоплазме эпителиоцитов (на стороне, обращенной к овоциту) хорошо развиты комплекс Гольджи с секреторными включениями, рибосомы и полирибосомы. На поверхности клеток видны два вида микроворсинок: одни проникают в прозрачную зону, а другие обеспечивают контакт между фолликулярными эпителиоцитами. Подобные микроворсинки имеются и у овоцита. Такие фолликулы, состоящие из овоцита, формирующейся прозрачной зоны и кубических фолликулярных эпителиоцитов, называются растущими фолликулами (рис. 20.13, 20.14, б).
Дальнейший рост фолликула обусловлен продолжающейся пролиферацией клеток фолликулярного эпителия, увеличением числа его слоев и формированием снаружи (из клеток соединительной ткани яичника) так называемой покрышки фолликула (theca folliculi). По мере дальнейшего развития тека фолликула дифференцируется на внутреннюю (theca interna) и наружную (theca externa). В theca interna (вокруг разветвляющихся капилляров) располагаются интерстициальные эндокриноциты, соответствующие клеткам Лейдига семенника. Совместно с фолликулярными эпителиоцитами они начинают активную продукцию женских половых гормонов (эстрогенов), которая регулируется гонадотропинами гипофиза. Параллельно в фолликуле формируется полость в результате активной секреции фолликулярной жидкости. Эстрогены вместе с другими продуктами жизнедеятельности фолликула (органическими соединениями, ионами, многочисленными факторами роста) выделяются в полость фолликула. Наружная тека (theca externa) образована плотной соединительной тканью. Далее, по мере роста полостного фолликула и накопления в нем жидкости, овоцит смещается к одному из полюсов фолликула. Стенка фолликула постепенно истончается, однако в месте расположения овоцита остается многослойной - образует яйценосный бугорок, или кумулюс (cumulus oophorus).
Накапливающаяся в фолликуле жидкость приводит к выделению ово-цита из массы клеток яйценосного бугорка. Овоцит остается связанным с клетками кумулюса лишь тонкой клеточной ножкой. Со стороны фолликулярной полости поверхность овоцита покрыта 2-3 слоями клеток фолликулярного эпителия, что напоминает венец (поэтому данная оболочка овоцита получила название лучистого венца - corona radiata). Клетки лучистого венца имеют длинные ветвистые отростки, проникающие через прозрачную зону и достигающие поверхности овоцита. По этим отросткам к овоциту от фолликулярных эпителиоцитов поступают питательные вещества и факторы регуляции. Зрелый фолликул, достигший своего максимального развития, называется граафовым пузырьком по имени автора (R. de Graaf), впервые описавшего его. Зрелый фолликул, готовый к овуляции, имеет и другое название - преовуляторный фолликул (см. рис. 20.13, 20.14). Овоцит преовуляторного фолликула возобновляет мейоз - завершает первое деление мейоза и вступает во второе деление, однако деление блокируется в метафазе. В метафазе происходит овуляция - выход овоцита из яичника. Полное завершение овоцитом мейоза произойдет лишь в том случае, если половая клетка будет оплодотворена мужской половой клеткой.
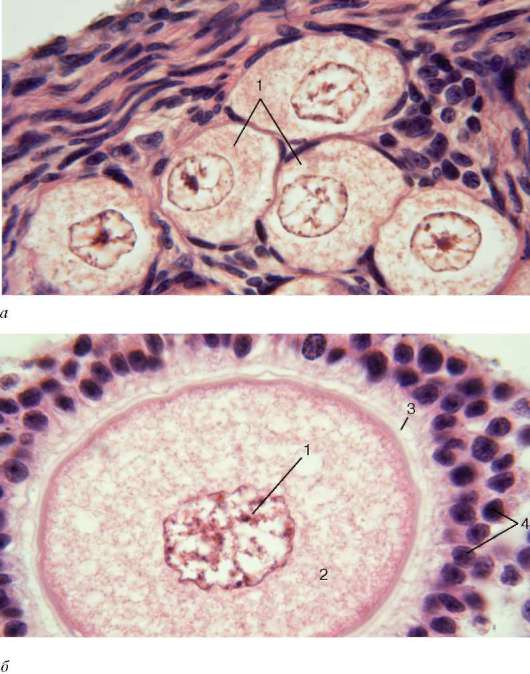
Рис. 20.14. Строение фолликулов, овоцитов и желтого тела яичника (микрофотографии):
а - примордиальные фолликулы: 1 - овоциты 1-го порядка (первичные); б - растущий фолликул: 1 - ядро; 2 - цитоплазма с равномерно распределенными желтковыми включениями; 3 - прозрачная зона; 4 - фолликулярные эпителиоциты; в - зрелый фолликул в начале овуляции: 1 - яйцеклетка; 2 - полость фолликула; 3 - стенка пузырька; 4 - поверхность яичника; г - желтое тело: 1 - лютеиновые клетки на разных стадиях дифференцировки; д - атретическое тело: 1 - прозрачная зона; 2 - фолликулярные эпителиоциты
В корковом веществе яичника среди развивающихся фолликулов находятся атретические фолликулы. Атретический фолликул (folliculus atreticus) - это фолликул с погибающей половой клеткой, не способный к продолжению развития. Гибель овоцитов начинается с лизиса органелл, кортикальных гранул и сморщивания ядра. При этом прозрачная зона утрачивает свою шаровидную форму и становится складчатой, утолщается и гиалинизируется.
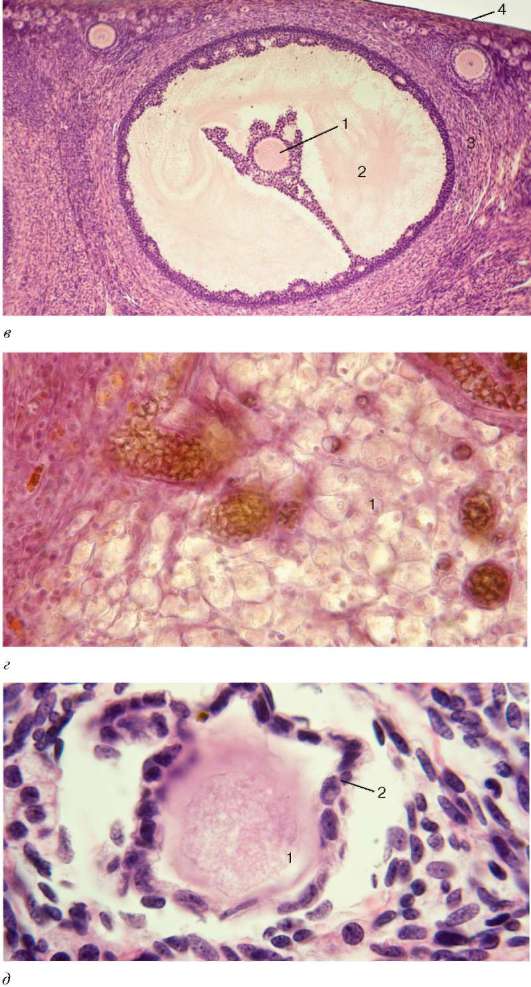
Рис. 20.14. Продолжение (обозначения см. выше)
В ходе дальнейшей инволюции атретических фолликулов на их месте остаются скопления отдельных клеток.
Причины атрезии до конца не выяснены, однако она признана ключевым фактором отбора фолликулов (и половых клеток) для овуляции (рис. 20.14, д). Атрезия примордиальных и растущих фолликулов небольшого размера протекает по типу дегенерационной - от таких фолликулов в яичниках остаются мелкие полости (микрокисты), которые затем бесследно исчезают. Атрезия крупных растущих фолликулов протекает по типу продуктивной (текогенный тип): по мере гибели фолликулярных эпителиоцитов существенно гипертрофируется внутренняя часть покрышки фолликула. Хорошая иннервация атретических фолликулов, а также увеличение содержания рибонуклеопротеидов и липидов в гипертрофирующихся клетках и возрастание активности их ферментов свидетельствуют о повышении метаболизма и высокой функциональной активности атретических фолликулов. В частности, интерстициальные клетки фолликула становятся активными продуцентами половых гормонов (преимущественно группы андрогенов и небольшого количества эстрогенов).
Мозговое вещество яичника (medulla ovarii) состоит из органоспецифиче-ской рыхлой соединительной ткани, в которой проходят магистральные кровеносные сосуды, лимфатические сосуды, нервы. В мозговом веществе находятся остатки канальцев первичной почки - сеть яичника (rete ovarii).
Генеративная функция. Овогенез
Овогенез отличается от сперматогенеза рядом особенностей и проходит в три стадии. Так, первая стадия - размножение овогоний - у человека осуществляется во внутриутробном периоде развития (у некоторых видов млекопитающих и в первые месяцы постнатальной жизни), когда в яичнике зародыша происходит деление овогоний и формирование примордиальных фолликулов (рис. 20.15).
Во второй стадии (роста) различают малый и большой рост. Первый протекает в эмбриогенезе, большой рост овоцитов - в репродуктивном возрасте (в функционирующем яичнике). Третья стадия - созревания. Эта стадия, как и при сперматогенезе, включает два деления мейоза, причем второе следует за первым без интеркинеза, что приводит к уменьшению (редукции) числа хромосом вдвое, и набор их становится гаплоидным. При первом делении созревания первичный овоцит (1-го порядка) делится, в результате чего образуются вторичный овоцит (2-го порядка) и небольшое первое полярное (редукционное) тельце. Вторичный ово-цит получает почти всю массу накопленного желтка и поэтому остается столь же крупным по объему, как и первичный овоцит. Полярное тельце (полоцит) представляет собой мелкую клетку с небольшим количеством цитоплазмы, получающую по одной диаде от каждой тетрады ядра первичного овоцита. При втором делении созревания в результате деления вторичного овоцита образуются одна гаплоидная яйцеклетка и второе полярное тельце. Первое полярное тельце иногда тоже делится на две мелкие клетки. В результате этих преобразований первичного овоцита
образуются одна яйцеклетка и три полярных тельца. Четвертая стадия - формирование - в овогенезе отсутствует.
Овуляция. Наступление овуляции - разрыв фолликула и выход вторичного овоцита в брюшную полость - вызывается действием лютеинизиру-ющего гормона (лютропин), когда выделение его гипофизом резко увеличивается. Перед овуляцией происходит выраженная гиперемия яичника,
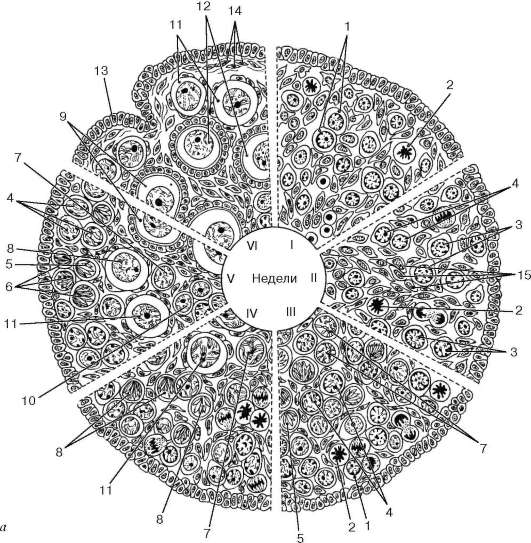
Рис. 20.15. Овогенез в антенатальном периоде развития (по Л. Ф. Курило): а - схема стадий овогенеза: I - 6-7 нед; II - 9-10 нед; III - 12-13 нед; IV - 16- 17 нед; V - 27-28 нед; VI - 38-40 нед. 1 - овогонии в интерфазе; 2 - овогонии в митозе; 3 - овоцит на стадии прелептотенной конденсации хромосом; 4 - овоцит на стадии прелептотенной деконденсации хромосом; 5 - овоцит в лептотене; 6 - овоцит в зиготене; 7 - овоцит в пахитене; 8 - овоцит в диплотене; 9 - овоцит в диктиотене; 10 - островки половых клеток на границе коркового и мозгового вещества; 11 - при-мордиальный фолликул; 12 - однослойный (первичный) фолликул; 13 - покровный эпителий; 14 - белочная оболочка яичника; 15 - тяжи соединительной ткани
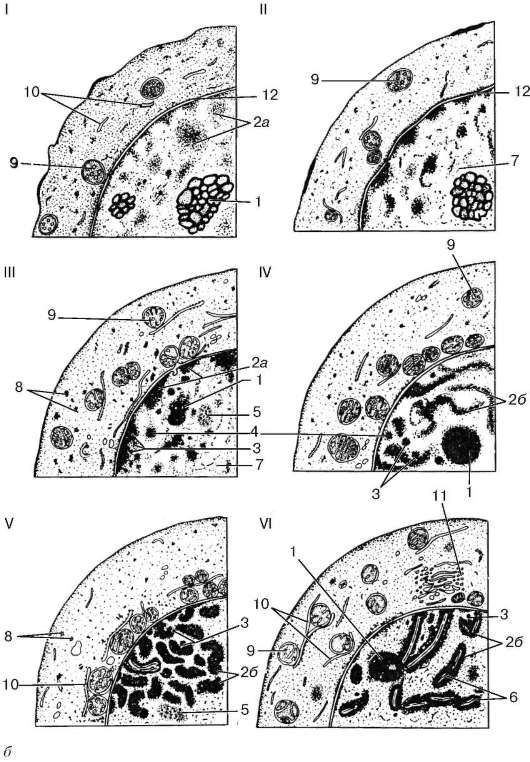
Рис. 20.15. Продолжение
б - схема ультраструктуры женских половых клеток дофолликулярных стадий овогенеза у плодов человека: I - гоноцит; II - овогония в интерфазе; III - овоцит в прелептотенной деконденсации хромосом; IV - овоцит в лептотене; V - овоцит в зиготене; VI - овоцит в пахитене. 1 - ядрышко; 2а - хроматин; 2б - хромосомы; 3 - перихроматиновые гранулы; 4 - сферы 90-120 нм; 5 - скопления интерхроматино-вых гранул; 6 - синаптонемный комплекс; 7 - элементарные хромосомные нити; 8 - рибосомы; 9 - митохондрии; 10 - эндоплазматическая сеть; 11 - комплекс Гольджи; 12 - ядерная оболочка
развитие интерстициального отека, инфильтрация стенки фолликула сег-ментоядерными гранулоцитами. Объем фолликула и давление в нем быстро возрастают, стенка его резко истончается. В нервных волокнах и терминалях обнаруживается наивысшая концентрация катехоламинов. Известную роль в овуляции может играть окситоцин. Перед наступлением овуляции секреция окситоцина увеличивается в ответ на раздражение нервных окончаний (располагающихся в theca interna), обусловленное повышением внутрифол-ликулярного давления. Кроме того, истончению и разрыхлению фолликула способствуют протеолитические ферменты, а также взаимодействие гиалу-роновой кислоты и гиалуронидазы, находящихся в его оболочке.
Вторичный овоцит, находящийся в блоке метафазы 2-го деления мейо-за, окруженный клетками лучистого венца, из брюшной полости попадает в воронку и далее в просвет маточной трубы. Здесь при встрече со спермием снимается блок деления и завершается второе деление мейоза.
Желтое тело (corpus luteum). Тканевые элементы стенки лопнувшего зрелого фолликула претерпевают изменения, приводящие к формированию желтого тела - временной добавочной эндокринной железы в составе яичника. При этом в полость запустевшего фолликула изливается кровь из сосудов внутренней части теки. Сгусток крови быстро замещается соединительной тканью в центре развивающегося желтого тела. В развитии желтого тела различают четыре стадии. В первой стадии - пролиферации и васкуляризации - происходит размножение фолликулярных эпителиоцитов, и между ними врастают капилляры из внутреннего слоя теки. Затем наступает вторая стадия - железистого метаморфоза, когда фолликулярные эпителиоциты гипертрофируются и в них накапливается желтый пигмент (лютеин), принадлежащий к группе липохромов. Такие клетки называются лютеоцитами (luteocyti). Объем новообразующе-гося желтого тела быстро увеличивается, и оно приобретает желтый цвет, хорошо различимый при жизни. С этого момента желтое тело начинает продуцировать свой гормон - прогестерон, переходя в третью стадию - расцвета (см. рис. 20.13, 20.14, г). Продолжительность этой стадии различна. Если оплодотворения не произошло, время расцвета желтого тела ограничивается 12-14 днями. В этом случае оно называется менструальным желтым телом (corpus luteum menstruationis). Более длительно желтое тело сохраняется, если наступила беременность - желтое тело беременности (corpus luteum graviditatis).
Разница между желтым телом беременности и менструальным ограничивается только длительностью стадии расцвета и размерами (1,5- 2 см в диаметре у менструального и более 5 см в диаметре у желтого тела беременности). После прекращения функционирования как желтое тело беременности, так и менструальное претерпевают инволюцию (стадию обратного развития). Железистые клетки атрофируются, а соединительная ткань центрального рубца разрастается. В результате на месте бывшего желтого тела формируется беловатое тело (corpus albicans) - соединительнотканный рубец. Оно сохраняется в яичнике на протяжении нескольких лет.
Эндокринные функции
В то время как яичко на протяжении своей активной деятельности непрерывно вырабатывает половой гормон, для яичника характерна циклическая (поочередная) продукция эстрогенов и гормона желтого тела - прогестерона.
Эстрогены (эстрадиол, эстрон и эстриол) обнаруживаются в жидкости, накапливающейся в полостях фолликулов. Поэтому эти гормоны ранее именовались фолликулярными, или фолликулинами. Яичник начинает интенсивно продуцировать эстрогены при достижении женским организмом половой зрелости, когда устанавливаются половые циклы, которые у низших млекопитающих проявляются регулярным наступлением течки (oestrus) - выделением пахучей слизи из влагалища, поэтому гормоны, под влиянием которых наступает течка, получили название эстрогенов.
Возрастное затухание деятельности яичников приводит к прекращению половых циклов.
Васкуляризация. Для яичника характерны спиралевидный ход артерий и вен и их обильное ветвление. Распределение сосудов в яичнике претерпевает изменения в связи с циклом фолликулов. В периоде роста фолликулов в развивающейся внутренней части теки формируется сосудистое сплетение, сложность которого возрастает ко времени овуляции и формирования желтого тела. В последующем по мере обратного развития желтого тела сосудистое сплетение редуцируется. Вены во всех частях яичника связаны многочисленными анастомозами, и емкость венозной сети значительно превышает емкость артериальной системы.
Иннервация. Нервные волокна, входящие в яичник, как симпатические, так и парасимпатические, образуют сети вокруг фолликулов и желтых тел, а также в мозговом веществе. Кроме того, в яичниках обнаруживаются многочисленные рецепторы, через которые афферентные сигналы поступают в ЦНС и достигают гипоталамуса.
20.2.2. Другие органы женской половой системы
Маточные трубы
Маточные трубы, или яйцеводы (tubae uterinae), - парные органы, по которым половые клетки из яичников проходят в матку.
Развитие. Маточные трубы развиваются из верхней части парамезонеф-ральных протоков.
Строение. Стенка яйцевода имеет три оболочки: слизистую (tunica mucosa), мышечную (tunica muscularis) и серозную (tunica serosa) (рис. 20.16). Слизистая оболочка собрана в крупные разветвленные продольные складки. Она покрыта однослойным столбчатым эпителием, который образован диф-феронами реснитчатых и секреторных эпителиоцитов.
Последние секретируют слизь, основными компонентами которой являются гликозаминогликаны, преальбумины, простагландины и др. Собственная пластинка слизистой оболочки представлена рыхлой соединительной тканью. Мышечная оболочка, следующая за слизистой, состоит из
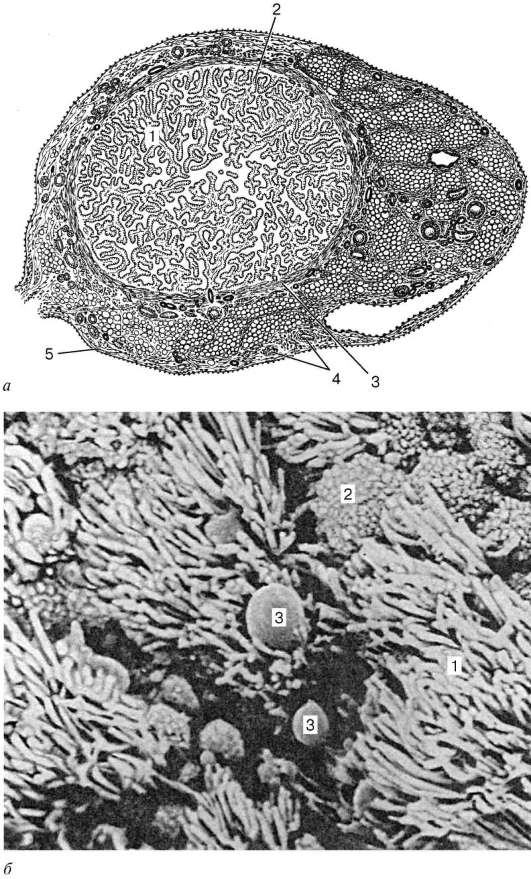
Рис. 20.16. Маточная труба:
а - строение (поперечный срез): 1 - складки слизистой оболочки; 2 - собственная пластинка слизистой оболочки; 3 - мышечная оболочка; 4 - кровеносный сосуд; 5 - серозная оболочка; б - сканирующая электронная микрофотография слизистой оболочки маточной трубы (по Савараги и Тонака): 1 - мерцательные реснички; 2 - апикальные поверхности секреторных эпителиоцитов; 3 - капли секрета
внутреннего циркулярного или спирального слоя и наружного продольного. Снаружи яйцеводы покрыты серозной оболочкой.
Дистальный конец яйцевода расширяется в воронку и заканчивается бахромкой (фимбриями). В момент овуляции сосуды фимбрий яйцеводов увеличиваются в объеме, воронка при этом плотно охватывает яичник. Передвижение половой клетки по яйцеводу обеспечивается не только движением ресничек эпителиальных клеток, выстилающих полость маточной трубы, но и перистальтическими сокращениями ее мышечной оболочки.
Матка
Матка (uterus) - мышечный орган, предназначенный для осуществления внутриутробного развития плода.
Развитие. Матка и влагалище развиваются у зародыша из дистального отдела левого и правого парамезонефральных протоков в месте их слияния. В связи с этим вначале тело матки характеризуется некоторой двурогостью, но к 4-му мес внутриутробного развития слияние заканчивается и матка приобретает грушевидную форму.
Строение. Стенка матки состоит из трех оболочек: слизистой, или эндометрия (endometrium), мышечной, или миометрия (myometrium), и серозной, или периметрия (perimetrium) (рис. 20.17). В эндометрии различают два слоя - функциональный и базальный. Строение функционального (поверхностного) слоя зависит от овариальных гормонов и претерпевает глубокую перестройку на протяжении менструального цикла. Слизистая оболочка матки выстлана однослойным столбчатым эпителием, образованным дифферона-ми реснитчатых и секреторных эпителиоцитов. Реснитчатые клетки располагаются преимущественно вокруг устьев маточных желез. Собственная пластинка слизистой оболочки матки образована рыхлой волокнистой со единительной тканью.
Некоторые клетки соединительной ткани развиваются в предециду-альные клетки крупного размера и округлой формы, содержащие в своей цитоплазме глыбки гликогена и липопротеиновые включения. Количество предецидуальных клеток возрастает (со времени менструации), особенно при формировании плаценты в период беременности.
В слизистой оболочке находятся многочисленные маточные железы, простирающиеся через всю толщу эндометрия. По форме маточные железы относятся к простым трубчатым.
Миометрий состоит из трех слоев гладких мышечных клеток - внутреннего подслизистого (stratum musculare submucosum), среднего сосудистого с косо-продольным расположением миоцитов (stratum musculare vasculosum), богатого сосудами, и наружного надсосудистого (stratum musculare supravasculosum) с косопродольным расположением мышечных клеток, но перекрестным по отношению к сосудистому слою. Такое расположение мышечных пучков имеет определенное значение в регуляции интенсивности циркуляции крови в течение менструального цикла.
Между пучками мышечных клеток имеются прослойки соединительной ткани, изобилующей эластическими волокнами. Гладкие мышечные
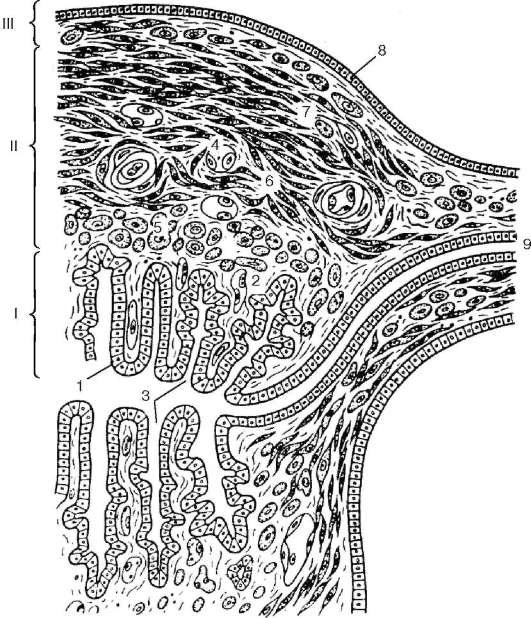
Рис. 20.17. Стенка матки (по Ю. И. Афанасьеву):
I - эндометрий; II - миометрий; III - периметрий. 1 - однослойный столбчатый эпителий; 2 - собственная пластинка слизистой оболочки; 3 - маточные железы (крипты); 4 - кровеносные сосуды; 5 - подслизистый мышечный слой; 6 - сосудистый мышечный слой; 7 - надсосудистый мышечный слой; 8 - мезотелий; 9 - маточная труба
клетки миометрия длиной около 50 мкм во время беременности сильно гипертрофируются, достигая иногда в длину 500 мкм. Они слегка ветвятся и соединяются отростками в сеть.
Периметрий покрывает большую часть поверхности матки. Не покрыты брюшиной лишь передняя и боковые поверхности надвлагалищной части шейки матки. В формировании периметрия принимают участие мезотелий, лежащий на поверхности органа, и рыхлая соединительная ткань, составляющие прослойку, примыкающую к мышечной оболочке матки. Однако
не во всех местах эта прослойка одинакова. Вокруг шейки матки, особенно с боков и спереди, находится большое скопление жировой ткани, которое получило название параметрия (parametrium). В других участках матки эта часть периметрия образована относительно тонким слоем рыхлой волокнистой соединительной ткани.
Шейка матки имеет вид цилиндра, в центре которого проходит шеечный канал. Слизистая оболочка выстилает полость канала и переходит на область внутреннего зева матки. В слизистой оболочке в составе однослойного столбчатого эпителия различают реснитчатые и слизистые эпи-телиоциты, секретирующие слизь. Но наибольшее количество секрета вырабатывается многочисленными сравнительно крупными разветвленными шеечными железами, располагающимися в строме складок слизистой оболочки.
Во влагалищной части шейки матки происходит стык эпителиев. Здесь начинается многослойный плоский неороговевающий эпителий, продолжающийся в эпителий влагалища. В области стыка двух эпителиев возникают атипический рост эпителиоцитов, формирование псевдоэрозий и развитие рака шейки матки.
Мышечная оболочка шейки представлена мощным циркулярным слоем гладких мышечных клеток, составляющим так называемый сфинктер матки, при сокращении которого выжимается слизь из шеечных желез. При расслаблении этого мышечного кольца возникает лишь своего рода аспирация (всасывание), способствующая втягиванию в матку спермы, попавшей во влагалище.
Васкуляризация. Система кровоснабжения матки хорошо развита. Артерии, несущие кровь к миометрию и эндометрию, в циркулярном слое миометрия спиралевидно закручиваются, что способствует их автоматическому сжатию при сокращении матки. Особенное значение это приобретает во время родов, так как предотвращается возможность сильного маточного кровотечения в связи с отделением плаценты. Входя в эндометрий, приносящие артерии дают начало мелким артериям двух видов, одни из них, прямые, не выходят за пределы базального слоя эндометрия, другие же, спиральные, снабжают кровью функциональный слой.
Лимфатические сосуды в эндометрии образуют глубокую сеть, которая через лимфатические сосуды миометрия соединяется с наружной сетью, располагающейся в периметрии.
Иннервация. Матка получает нервные волокна, преимущественно симпатические, от подчревного сплетения. На поверхности матки в периметрии эти симпатические волокна образуют хорошо развитое маточное сплетение. От этого поверхностного сплетения отходят веточки, снабжающие миометрий и проникающие в эндометрий. Около шейки матки в окружающей клетчатке располагается группа крупных ганглиев, в которых, кроме симпатических нервных клеток, имеются хромаффинные клетки. В толще миометрия ганглионарные клетки отсутствуют. В последнее время получены данные, свидетельствующие о том, что матка иннервируется как симпатическими, так и некоторым количеством парасимпатических волокон.
В то же время в эндометрии обнаружено большое количество рецептор-ных нервных окончаний различной структуры, раздражение которых не только вызывает сдвиги в функциональном состоянии самой матки, но и отражается на многих общих функциях организма: кровяном давлении, дыхании, общем обмене веществ, гормонообразователь-ной деятельности гипофиза и других эндокринных желез, наконец, на деятельности центральной нервной системы.
Влагалище
Стенка влагалища состоит из слизистой (tunica mucosa), мышечной (tunica musculare) и адвентициальной оболочек (tunica adventitia). В составе слизистой оболочки имеется многослойный плоский неороговевающий эпителий, в котором различают три слоя: базальный, парабазальный, промежуточный и поверхностный, или функциональный (рис. 20.18).
Эпителий слизистой оболочки влагалища претерпевает значительные ритмические (циклические) изменения в последовательных фазах менструального цикла. В клетках поверхностных слоев эпителия (в его функциональном слое) откладываются зерна кератогиалина, но полного ороговения клеток в норме не происходит. Клетки этого слоя эпителия богаты гликогеном. Распад гликогена под влиянием микробов, всегда обитающих во влагалище, приводит к образованию молочной кислоты, поэтому влагалищная слизь имеет кислую реакцию и обладает бактерицидными свойствами, что предохраняет влагалище от развития в нем патогенных микроорганизмов. Железы в стенке влагалища отсутствуют. Базальная граница эпителия неровная, так как собственная пластинка слизистой оболочки формирует сосочки неправильной формы, вдающиеся в эпителиальный пласт.
Основу собственной пластинки слизистой оболочки составляет рыхлая волокнистая соединительная ткань, эластические волокна которой образуют поверхностную и глубокую сети. Собственная пластинка часто инфильтрируется лимфоцитами, иногда в ней встречаются единичные лимфоидные узелки. Подслизистая основа во влагалище не выражена и собственная пластинка слизистой оболочки непосредственно переходит в прослойки соединительной ткани в мышечной оболочке, которая в основном состоит из продольно идущих пучков гладких мышечных клеток, между
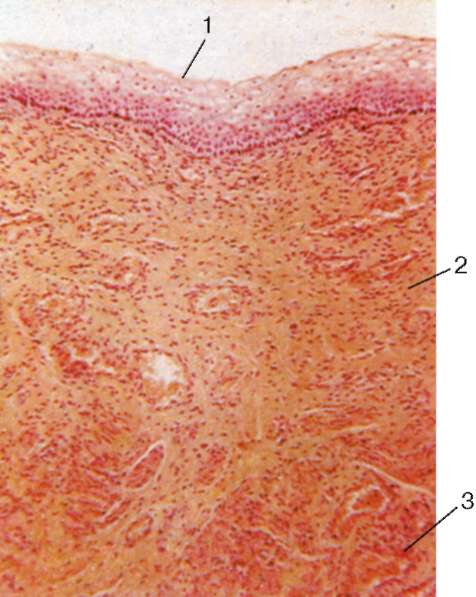
Рис. 20.18. Влагалище: 1 - многослойный плоский неороговевающий эпителий; 2 - собственная пластинка слизистой оболочки; 3 - пучки гладкой мышечной ткани
пучками которых в средней части мышечной оболочки имеется небольшое количество циркулярно расположенных мышечных элементов.
Адвентициальная оболочка влагалища состоит из рыхлой волокнистой неоформленной соединительной ткани, связывающей влагалище с соседними органами. В этой оболочке располагается венозное сплетение.
20.3.3. Овариально-менструальный цикл
Циклическая деятельность женской половой системы (яичники, маточные трубы, матка, влагалище), т. е. последовательные изменения ее функции и структуры - овариально-менструальный цикл -регулярно повторяется в одном и том же порядке. У женщин и самок человекообразных обезьян половые циклы характеризуются регулярными маточными кровотечениями (менструациями).
У большинства женщин, достигших половой зрелости, менструации повторяются регулярно через 28 дней. В овариально-менструальном цикле различают три периода, или фазы: менструальный (фаза десквама-ции эндометрия), которым заканчивается предыдущий менструальный цикл, постменструальный период (фаза пролиферации эндометрия) и, наконец, предменструальный период (функциональная фаза, или фаза секреции), во время которого происходит подготовка эндометрия к возможной имплантации оплодотворенной яйцеклетки, если наступило оплодотворение.
Менструальный период. Начало менструальной фазы определяется резким изменением кровоснабжения эндометрия. На протяжении предыдущей предменструальной (функциональной) фазы под влиянием прогестерона, интенсивно секретируемого желтым телом, вступившим в этот период в стадию расцвета, кровеносные сосуды эндометрия достигают максимального развития. Прямые артерии дают начало капиллярам, питающим базальный слой эндометрия, а спиралевидные артерии, разрастающиеся в этой фазе, закручиваются в клубочки и образуют густую сеть капилляров, ветвящихся в функциональном слое эндометрия. По мере того как к концу предменструального периода желтое тело в яичнике начинает атрофироваться (вступает в стадию обратного развития), прекращается поступление прогестерона в циркуляцию. Вследствие этого начинаются спазмы спиралевидных артерий, результатом чего оказывается значительное уменьшение притока крови к эндометрию (ишемическая фаза) и в нем развивается гипоксия, а в сосудах возникают тромбы. Стенки сосудов теряют эластичность и становятся ломкими. На прямые артерии указанные изменения не распространяются, и базальный слой эндометрия продолжает снабжаться кровью.
В функциональном слое эндометрия вследствие ишемии начинаются некротические изменения. После длительного спазма спиралевидные артерии вновь расширяются и приток крови к эндометрию увеличивается. Но так как стенки этих сосудов стали хрупкими, в них возникают многочисленные разрывы, и в строме эндометрия начинаются кровоизлияния, обра-
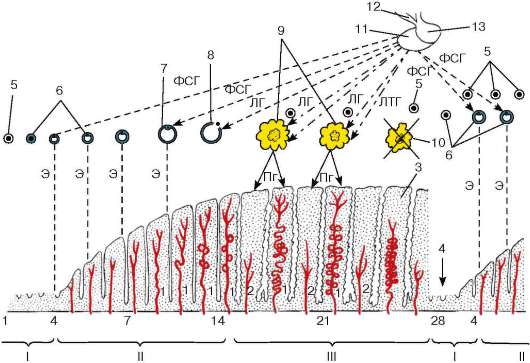
Рис. 20.19. Овариально-менструальный цикл (схема):
I - менструальная фаза; II - постменструальная фаза; III - предменструальная фаза. 1 - извитая артерия эндометрия; 2 - прямая артерия эндометрия; 3 - спазм и регрессия терминальных ветвей извитых артерий (ишемическая фаза); 4 - кровоизлияние в эндометрии; 5 - примордиальный фолликул в яичнике; 6 - растущие фолликулы; 7 - зрелый (граафов) фолликул; 8 - овуляция; 9 - желтое тело в стадии расцвета; 10 - обратное развитие желтого тела; 11 - передняя доля гипофиза; 12 - воронка промежуточного мозга; 13 - задняя доля гипофиза. ФСГ - действие фоллитропина на растущие фолликулы; ЛГ - действие лютеинизирующего гормона (лютропина) на овуляцию и образование желтых тел; ЛТГ - действие лактотропина (пролактина) на сформированное желтое тело; Э - действие эстрогена на матку, стимулирующее рост эндометрия (постменструальная или пролиферативная фаза); Пг - действие прогестерона на эндометрий (предменструальная фаза)
зуются гематомы. Некротизирующийся функциональный слой отторгается, расширенные кровеносные сосуды эндометрия вскрываются, и наступает маточное кровотечение.
В день менструации в организме женщины практически отсутствуют овариальные гормоны, так как секреция прогестерона прекращается, а секреция эстрогенов (которой препятствовало желтое тело, пока оно было в расцвете) еще не возобновилась. Но, поскольку начавшаяся регрессия желтого тела растормаживает рост очередной группы фолликулов, продукция эстрогенов становится возможной. Под их влиянием в матке активизируется регенерация эндометрия и усиливается пролиферация эпителия за счет донышек маточных желез, которые сохранились в базальном слое после десквамации функционального слоя. Через 2-3 дня пролиферации
Рис. 20.20. Строение матки женщины в репродуктивном периоде в разные фазы цикла (по О. В. Волковой).
I - фаза пролиферации; II - фаза секреции; III - фаза десквамации; а - эпителий; б - соединительнотканная основа; в - железы; г - гладкие мышцы; д - сосуды; е - гемостаз и диапедез элементов крови
менструальное кровотечение останавливается и начинается очередной постменструальный период. Таким образом, постменструальная фаза определяется влиянием эстрогенов, а предменструальная - влиянием прогестерона. Овуляция наступает в яичнике на 12-17-й день менструального цикла, т. е. приблизительно посередине между двумя очередными менструациями. В связи с участием гормонов яичника в регуляции перестройки, претерпеваемой маткой, описанный процесс обычно называют не менструальным, а овариально-менструальным циклом (рис. 20.19).
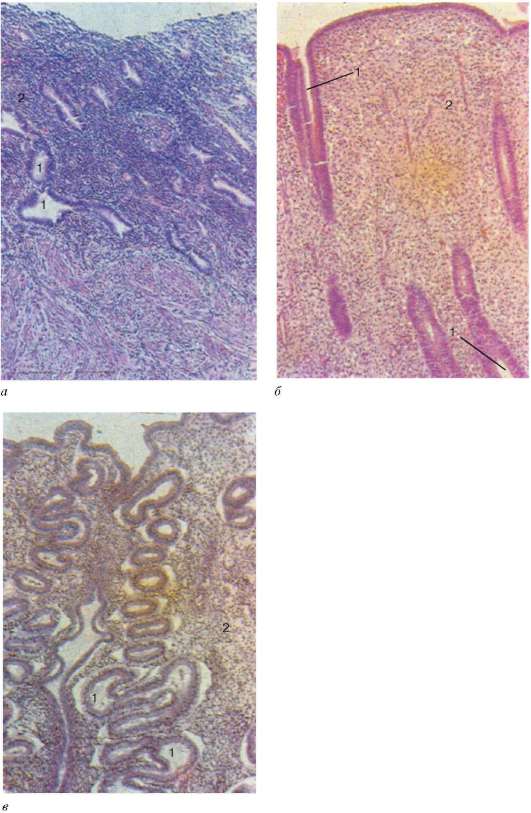
Рис. 20.21. Строение эндометрия матки женщины в различные фазы цикла. Микрофотографии (препараты Ю. И. Ухова):
а - менструальная фаза; б - постменструальная фаза пролиферации; в - предменструальная фаза секреции (20-е сут менструального цикла). 1 - маточные железы (крипты); 2 - собственная пластинка слизистой оболочки
Постменструальный период. Этот период начинается вслед за окончанием менструации (см. рис. 20.19). В этот момент эндометрий представлен только базальным слоем, в котором остались дистальные отделы маточных желез. Уже начавшаяся регенерация функционального слоя позволяет назвать данный период пролиферативной фазой (рис. 20.20, 20.21). Она продолжается с 5-го по 14-15-й день цикла. Пролиферация регенерирующего эндометрия наиболее интенсивна в начале данной фазы (5-11-й день цикла), затем темп регенерации замедляется и наступает период относительного покоя (11-14-й день). Маточные железы в постменструальном периоде растут быстро, но остаются узкими, прямыми и не секретируют. Как уже упоминалось, рост эндометрия стимулируется эстрогенами, которые продуцируются полостными (антральными) фолликулами. Следовательно, во время постменструального периода в яичнике происходит рост очередного фолликула, который достигает стадии зрелого к 14-му дню цикла.
Предменструальный период. В конце постменструального периода в яичнике наступает овуляция, а на месте лопнувшего зрелого фолликула образуется желтое тело, вырабатывающее прогестерон, который активирует маточные железы, начинающие секретировать. Они увеличиваются в размерах, становятся извитыми и часто разветвляются. Их клетки набухают, а просветы желез заполняются выделяемым секретом. В цитоплазме появляются вакуоли, содержащие гликоген и гликопротеиды, - сначала в базальной части, а затем смещающиеся к апикальному краю. Слизь, обильно выделяемая железами, становится густой. В участках эпителия, выстилающего полость матки между устьями маточных желез, клетки приобретают призматическую форму, а на верхушках многих из них развиваются реснички. Толщина эндометрия увеличивается по сравнению с предыдущим постменструальным периодом, что обусловливается гиперемией и накоплением отечной жидкости в собственной пластинке. В клетках соединительнотканной стромы тоже откладываются глыбки гликогена и капельки липидов. Некоторые из этих клеток дифференцируются в децидуальные (см. «Плацента» в главе 21).
Если произошло оплодотворение, то эндометрий участвует в формировании плаценты. Если же оплодотворение не состоялось, то функциональный слой эндометрия разрушается и отторгается при очередной менструации.
Циклические изменения влагалища. С началом пролиферации эндометрия (на 4-5-е сут после окончания менструации), т. е. в постменструальном периоде, во влагалище заметно набухают эпителиальные клетки. На 7-8-е сут в этом эпителии дифференцируется промежуточный слой уплотненных клеток, а к 12-14-м сут цикла (к концу постменструального периода) в базальном слое эпителия клетки сильно набухают и увеличиваются в объеме. В верхнем (функциональном) слое влагалищного эпителия клетки разрыхляются, и в них накапливаются глыбки кератогиалина. Однако процесс кератинизации не доходит до полного ороговения. В предменструальном периоде деформированные уплотненные клетки функционального слоя влагалищного эпителия продолжают отторгаться, а клетки базального слоя уплотняются.
Состояние эпителия влагалища зависит от уровня овариальных гормонов в крови, поэтому по картине мазка, полученного с поверхности влагалища, можно судить о фазе менструального цикла и о его нарушениях.
Во влагалищных мазках содержатся слущенные эпителиоциты, могут быть клетки крови - лейкоциты и эритроциты. Среди эпителиоцитов различают клетки, находящиеся на различных стадиях дифференциров-ки, - базофильные, ацидофильные и промежуточные. Соотношение числа вышеуказанных клеток варьирует в зависимости от фазы овариально-менструального цикла. В ранней, пролиферативной фазе (7-е сут цикла) преобладают поверхностные базофильные эпителиоциты, в овуляторной фазе (11-14-е сут цикла) преобладают поверхностные ацидофильные эпителио-циты, в лютеиновой фазе (21-е сут цикла) увеличивается содержание промежуточных эпителиоцитов с крупными ядрами и лейкоцитов; в менструальной фазе значительно возрастает число клеток крови - лейкоцитов и эритроцитов (рис. 20.22).
Во время менструации в мазке преобладают эритроциты и нейтрофилы, эпителиальные клетки встречаются в небольшом количестве. В начале постменструального периода (в пролиферативной фазе цикла) влагалищный эпителий сравнительно тонок, а в мазке содержание лейкоцитов быстро уменьшается и появляются эпителиоциты с пикнотическими ядрами. К моменту овуляции (в середине овариально-менструального цикла) такие клетки в мазке становятся преобладающими, а толщина влагалищного эпителия возрастает. Наконец, в предменструальной фазе цикла количество клеток с пикнотическим ядром уменьшается, но усиливается десквамация нижележащих слоев, клетки которых обнаруживаются в мазке. Перед наступлением менструации содержание эритроцитов в мазке начинает возрастать.
20.3.4. Возрастные изменения органов женской половой системы
Морфофункциональное состояние органов женской половой системы зависит от возраста и активности нейроэндокринной системы.
Матка. У новорожденной девочки длина матки не превышает 3 см и, постепенно увеличиваясь в течение препубертатного периода, достигает конечных размеров по достижении половой зрелости.
К концу детородного периода и в связи с приближением климакса, когда гормонообразовательная деятельность яичников ослабевает, в матке начинаются инволютивные изменения, прежде всего в эндометрии. Дефицит лютеинизирующего гормона в переходном (предклимактерическом) периоде проявляется тем, что маточные железы, сохраняя еще способность к росту, уже перестают функционировать. После установления менопаузы атрофия эндометрия быстро прогрессирует, особенно в функциональном слое. Параллельно в миометрии развивается атрофия мышечных клеток, сопровождающаяся развитием соединительной ткани. В связи с этим размеры и масса матки, претерпевающей возрастную инволюцию, значительно умень-
Рис. 20.22. Влагалищные мазки, взятые в различные фазы овариально-менструаль-ного цикла:
а - пролиферативная фаза; б - овуляторная фаза; в - лютеиновая фаза; г - менструальная фаза. 1 - поверхностные эпителиальные базофильные клетки; 2 - поверхностные эпителиальные ацидофильные клетки; 3 - промежуточные эпителиальные клетки; 4 - лейкоциты; 5 - эритроциты
шаются. Наступление климактерического периода характеризуется уменьшением размеров органа и количества миоцитов в нем, а в кровеносных сосудах возникают склеротические изменения. Это является следствием снижения гормонообразования в яичниках.
Рис. 20.22. Продолжение (обозначения см. выше)
Яичники. В первые годы жизни размеры яичников у девочки увеличиваются преимущественно за счет роста мозговой части. Атрезия фолликулов, прогрессирующая в детском возрасте, сопровождается разрастанием соединительной ткани, а после 30 лет разрастание соединительной ткани захватывает и корковое вещество яичника.
Затухание менструального цикла в климактерическом периоде характеризуется уменьшением размеров яичников и исчезновением фолликулов в них, склеротическими изменениями их кровеносных сосудов. Вследствие недостаточной продукции лютропина овуляции и образования желтых тел
не происходит, и поэтому овариально-менструальные циклы сначала становятся ановуляторными, а затем прекращаются, и наступает менопауза.
Влагалище. Морфогенетические и гистогенетические процессы, приводящие к формированию основных структурных элементов органа, завершаются к периоду полового созревания.
После наступления климактерического периода влагалище претерпевает атрофические изменения, его просвет суживается, складки слизистой оболочки сглаживаются, количество влагалищной слизи уменьшается. Слизистая оболочка редуцируется до 4-5 слоев клеток, не содержащих гликогена. Эти изменения создают условия для развития инфекции (сениль-ный вагинит).
Гормональная регуляция деятельности женской половой системы. Как
упоминалось, фолликулы начинают расти еще в яичниках зародыша. Малый рост овоцитов, как и малый рост фолликулов в яичниках зародыша, не зависит от гормонов гипофиза. В функционирующем яичнике под влиянием гонадотропинов передней доли гипофиза (фолли-тропина и лютропина) происходят пролиферация и дифференцировка фолликулярных эпителиоцитов и эндокриноцитов внутренней теки. Развитие фолликулов с полостью становится полностью зависимым от гонадотропинов.
К окончанию роста фолликула возрастающее содержание лютропина в крови вызывает овуляцию и образование желтого тела. Фаза же расцвета желтого тела, в течение которой оно продуцирует и секретирует прогестерон, усиливается и удлиняется благодаря добавочному влиянию адено-гипофизарного пролактина. Местом приложения прогестерона является слизистая оболочка матки, которая под его влиянием подготавливается к восприятию оплодотворенной яйцевой клетки (зиготы). В то же время прогестерон ингибирует рост новых фолликулов. Наряду с выработкой прогестерона в желтом теле сохраняется выработка небольшого количества эстрогенов. Поэтому в конце фазы расцвета желтого тела вновь отмечается поступление эстрогенов в циркуляцию.
Половая дифференцировка гипоталамуса. Непрерывность мужской половой функции и цикличность женской связаны с особенностями секреции лютропина гипофизом. В мужском организме фоллитропин и лютропин секретируются одновременно и равномерно. Цикличность же женской половой функции обусловливается тем, что выделение лютропина из гипофиза в циркуляцию происходит не равномерно, а периодически, когда гипофиз выбрасывает в кровь повышенное количество этого гормона, достаточное для вызова овуляции и развития желтого тела в яичнике (так называемая овуляционная квота лютропина). Гормонопоэтические функции аденогипофиза регулируются аденогипофизотропными нейрогормонами медиобазального гипоталамуса.
Гипоталамическая регуляция лютеинизирующей функции передней доли гипофиза осуществляется двумя центрами. Один из них («низший» центр), находящийся в туберальных ядрах (аркуатном и вентромедиальном) медиобазального гипоталамуса, активирует переднюю долю гипофиза к непрерывной тонической секреции
обоих гонадотропинов. При этом количество выделяемого лютропина обеспечивает лишь секрецию эстрогенов яичниками и тестостерона семенниками, но слишком мало для вызова овуляции и образования желтого тела в яичнике. Другой центр («высший», или «овуляторный») локализуется в преоптической области медиоба-зального гипоталамуса и модулирует деятельность низшего центра, в результате чего последний активирует гипофиз к массивному выбросу «овуляторной квоты» лютропина.
При отсутствии влияния андрогена преоптический овуляторный центр сохраняет способность периодически возбуждать деятельность «низшего центра», как это свойственно женскому полу. Но у зародыша мужского пола благодаря наличию в его организме мужского полового гормона данный овуляторный центр гипоталамуса маскулинизируется. Критический период, после которого овуляторный центр теряет способность модифицироваться по мужскому типу и окончательно закрепляется в качестве женского, ограничивается у человеческого плода концом внутриутробного периода.
20.3. НАРУЖНЫЕ ПОЛОВЫЕ ОРГАНЫ
Преддверие влагалища выстлано многослойным плоским эпителием. В преддверие влагалища открываются две большие железы преддверия (бар-толиновы железы). По форме эти железы альвеолярно-трубчатые, образованы экзокриноцитами, которые секретируют слизь. В малых губах покрывающий их многослойный эпителий слегка ороговевает, а базальный слой его пигментирован. Основу малых губ составляет рыхлая соединительная ткань, богатая эластическими волокнами и кровеносными сосудами. В ней заложены многочисленные сальные железы.
Большие губы влагалища представляют собой складки кожи с обильными прослойками жировой ткани. В больших губах много сальных и потовых желез.
Клитор по эмбриональному развитию и строению соответствует дорсальной части мужского полового члена. Он состоит из двух эректильных пещеристых тел, заканчивающихся головкой, которая покрыта многослойным плоским эпителием, слегка ороговевающим.
Иннервация. Наружные половые органы, особенно клитор, обильно снабжены различными рецепторами. В эпителии этих органов разветвляются свободные нервные окончания. В соединительнотканных сосочках собственной пластинки их слизистой оболочки находятся осязательные нервные тельца, а в дерме - инкапсулированные генитальные тельца. В больших губах и клиторе встречаются также пластинчатые тельца.
Контрольные вопросы
1. Эмбриональные источники развития органов мужской половой системы, роль первичной почки.
2. Строение семенника, гематотестикулярный барьер, семявыносящие пути.
3. Сперматогенез: последовательность и содержание фаз, центральная и интраорганная (пара- и аутокринная) регуляция.
4. Эмбриональные источники развития органов женской половой системы, роль целомического эпителия и первичной почки в органогенезе.
5. Морфогенетические и хронологические особенности овогенеза у человека.
6. Развитие, строение, функции органов женского репродуктивного тракта.