Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 11. НЕРВНАЯ СИСТЕМА
Нервная система обеспечивает регуляцию всех жизненных процессов в организме и его взаимодействие с внешней средой. Анатомически нервную систему делят на центральную и периферическую. К первой относят головной и спинной мозг, вторая объединяет периферические нервные узлы, стволы и окончания.
Такое деление нервной системы условно и допускается лишь из методических соображений. Морфологическим субстратом рефлекторной деятельности нервной системы являются рефлекторные дуги, представляющие собой цепь нейронов различного функционального назначения, тела которых рас-
положены как в периферических узлах, так и в сером веществе центральной нервной системы.
С физиологической точки зрения нервную систему делят на соматическую, иннервирующую все тело, кроме внутренних органов, сосудов и желез, и автономную, или вегетативную, регулирующую деятельность перечисленных органов.
11.1. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ
Нервную систему формируют следующие эмбриональные источники: нервная трубка, нервный гребень (ганглиозная пластинка) и эмбриональные плакоды. Тканевые элементы оболочек являются мезенхимными производными. На стадии замыкания нейропоров передний конец трубки значительно расширяется, боковые стенки утолщаются, образуя зачатки трех мозговых пузырей. Лежащий краниально пузырь образует передний мозг, средний пузырь - средний мозг, а из третьего пузыря, который переходит в закладку спинного мозга, развивается задний (ромбовидный) мозг. Вскоре после этого нервная трубка изгибается почти под прямым углом, и посредством борозд-сужений первый пузырь разделяется на конечный и промежуточный отделы, а третий мозговой пузырь - на продолговатый и задний отделы мозга. Производные среднего и заднего мозговых пузырей образуют ствол мозга и являются древними образованиями; в них сохраняется сегментарный принцип строения, который исчезает в производных промежуточного и конечного мозга. В последних концентрируются интегративные функции. Так формируются пять отделов мозга: конечный и промежуточный мозг, средний, продолговатый и задний мозг (у человека это происходит примерно в конце 4-й нед эмбрионального развития). Конечный мозг формирует два полушария большого мозга.
В эмбриональном гисто- и органогенезе нервной системы развитие разных отделов мозга происходит с разной скоростью (гетерохронно). Раньше формируются каудальные отделы центральной нервной системы (спинной мозг, ствол мозга); время окончательного формирования структур головного мозга сильно варьирует. В ряде отделов головного мозга это происходит после рождения (мозжечок, гиппокамп, обонятельная луковица); в каждом отделе мозга существуют пространственно-временные градиенты формирования нейронных популяций, которые образуют уникальную структуру нервного центра.
Спинной мозг представляет собой часть центральной нервной системы, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегмен-тарность. В боковых отделах нервной трубки быстро возрастает масса клеток, тогда как дорсальная и вентральная ее части не увеличиваются в объеме и сохраняют эпендимный характер. Утолщенные боковые стенки нервной трубки делятся продольной бороздой на дорсальную - крыльную, и вентраль-
ную - основную пластинку. На этой стадии развития в боковых стенках нервной трубки можно выделить три зоны: эпендиму, выстилающую центральный канал, промежуточную (плащевой слой) и маргинальную (краевую вуаль). Из плащевого слоя в дальнейшем развивается серое вещество спинного мозга, а из краевой вуали - его белое вещество. Нейробласты передних столбов дифференцируются в мотонейроны (двигательные нейроны) ядер передних рогов. Их аксоны выходят из спинного мозга и образуют передние корешки спинномозговых нервов. В задних столбах и промежуточной зоне развиваются различные ядра вставочных (ассоциативных) клеток. Их аксоны, поступая в белое вещество спинного мозга, входят в состав различных проводящих пучков. В задние рога входят центральные отростки чувствительных нейронов спинномозговых узлов.
Одновременно с развитием спинного мозга закладываются спинномозговые и периферические узлы автономной нервной системы. Исходным материалом для них служат стволовые клеточные элементы нервного гребня, которые путем дивергентной дифференцировки развиваются в нейробла-стическом и глиобластическом направлениях (см. главу 10). Часть клеток нервного гребня мигрирует на периферию в места локализации узлов автономной нервной системы, параганглиев, нейроэндокринных клеток APUD-серии и хромаффинной ткани.
11.2.1. Нерв
Нервы (nervus) - периферические нервные стволы - состоят из миелино-вых и безмиелиновых волокон и соединительнотканных оболочек. В автономной нервной системе, как правило, в нервах встречаются одиночные нервные клетки и мелкие ганглии. На поперечном срезе нерва видны сечения осевых цилиндров нервных волокон и одевающие их глиальные оболочки. Между нервными волокнами в составе нервного ствола располагаются тонкие прослойки рыхлой волокнистой соединительной ткани - эндоневрий (endoneurium). Пучки нервных волокон одеты периневрием (perineurium). Периневрий состоит из чередующихся слоев плотно расположенных плоских клеток эпенди-моглиального дифферона и тонких слоев соединительной ткани. Таких слоев в периневрии толстых нервов несколько (5-6). Между пучками нервных волокон снаружи периневрия находится соединительная ткань с кровеносными сосудами. Периневрий образуется из оболочек спинного мозга при выходе из него корешков. Наружная оболочка нервного ствола - эпинев-рий (epineurium) - представляет собой плотную волокнистую соединительную ткань, богатую фибробластами, макрофагами и жировыми клетками. Соединительнотканные оболочки нерва содержат кровеносные и лимфатические сосуды и нервные окончания. В эпиневрий по всей длине нерва посту-
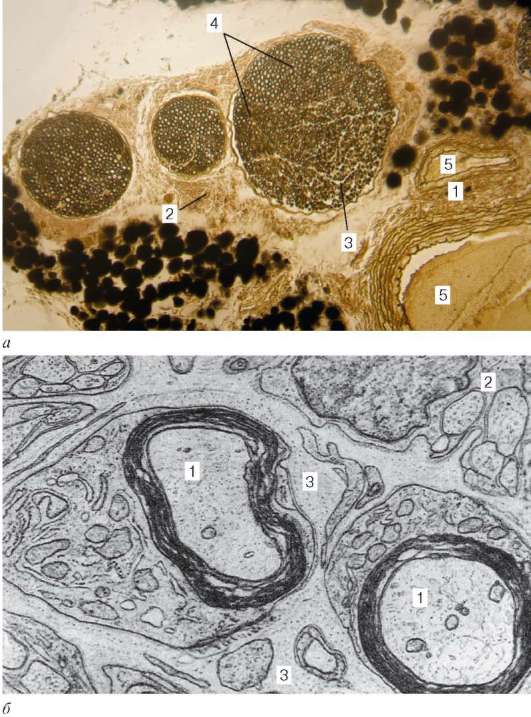
Рис. 11.1. Строение нерва:
а - микрофотография (обработка осмиевой кислотой): 1 - эпиневрий; 2 - перинев-рий; 3 - эндоневрий; 4 - нервные волокна миелиновые и безмиелиновые; 5 - кровеносные сосуды; б - электронная микрофотография (препарат А. И. Радостиной): 1 - миелиновые нервные волокна; 2 - безмиелиновые нервные волокна; 3 - прослойки соединительной ткани (эндоневрий)
пает большое количество анастомозирующих между собой кровеносных сосудов. Из эпиневрия артерии проникают в периневрий и эндоневрий (рис. 11.1). В эндоневрии артериолы формируют узкопетлистую капиллярную сеть, оплетающую нервные волокна.
Нервные волокна в составе периферического нерва способны к регенерации и формированию нервно-мышечных синапсов с клетками-мишенями (см. главу 10).
11.2.2. Чувствительные узлы
Чувствительные узлы (ganglii sensorii) лежат по ходу спинномозговых нервов либо черепных нервов. Спинномозговой узел
Спинномозговой узел (ganglion spinale) окружен соединительнотканной капсулой. От капсулы внутрь узла проникают тонкие прослойки соединительной ткани, в которой расположены кровеносные сосуды.
Нейроны спинномозгового узла имеют грушевидную или округлую форму диаметром 30-120 мкм. Они располагаются группами, преимущественно по периферии органа, тогда как его центр состоит главным образом из отростков этих клеток. Дендриты идут в составе чувствительной части смешанных спинномозговых нервов на периферию и заканчиваются там рецепторами. Аксоны в совокупности образуют задние корешки, несущие нервные импульсы, или в серое вещество спинного мозга, или по его заднему канатику в продолговатый мозг. В спинномозговых узлах высших позвоночных животных и человека биполярные нейроны в процессе созревания становятся псевдоуниполярными. Отростки клеток постепенно сближаются, и их основания сливаются (рис. 11.2). Вначале удлиненная часть тела (основание отростков) имеет небольшую длину, но со временем, разрастаясь, она многократно обвивает клетку и часто образует клубок.
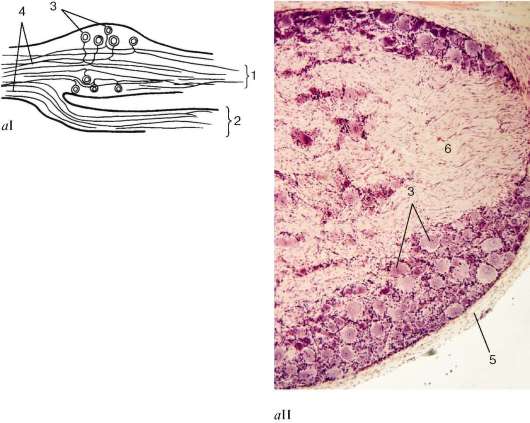
Рис. 11.2. Строение спинномозгового (чувствительного) узла: аI - схема; аII, б - микрофотографии (б - импрегнация нитратом серебра); в - ультраструктурная организация псевдоуниполярного нейрона и его микроокружения (по Р. В. Крстичу, с изменениями). 1 - задний корешок; 2 - передний корешок; 3 - псевдоуниполярные нейроны; 4 - нервные волокна; 5 - соединительнотканная капсула; 6 - узел (ганглий); 7 - нейролеммоцит; 8 - Т-образное разветвление отростка; 9 - ганглионар-ные глиоциты (сателлитные глиальные клетки); 10 - базальная мембрана; 11 - гемокапилляры; 12 - миелиновые и без-миелиновые волокна
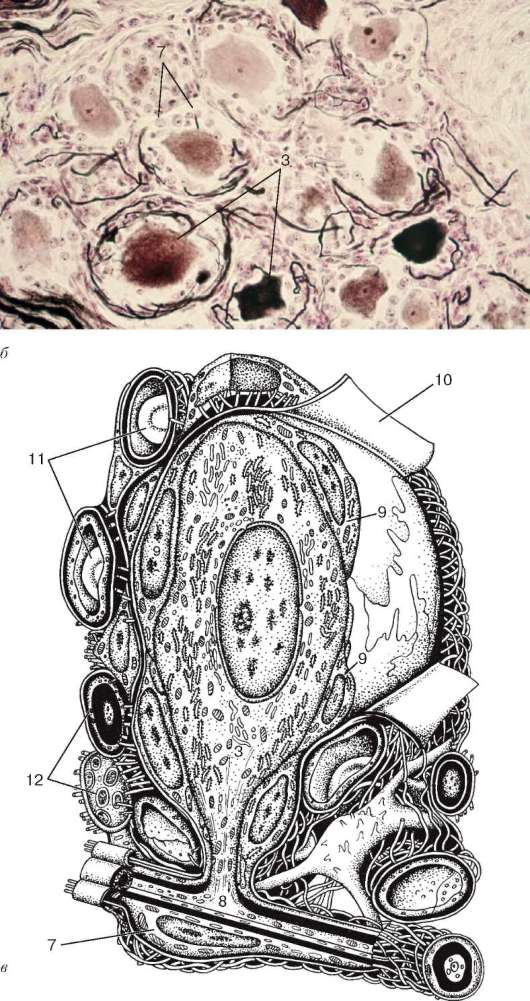
Рис. 11.2. Продолжение (расшифровку обозначений см. с. 297)
Существует и другая точка зрения на процесс формирования псевдоуниполярных нейронов: аксон отрастает от удлиненной части тела нейрона после формирования дендрита. Биполярные нейроны у низших позвоночных сохраняются в течение всей жизни. Биполярными являются и афферентные нейроны некоторых черепных нервов (gangl. spirale cochleare).
Дендриты и аксоны клеток в узле и за его пределами покрыты оболочками из нейролеммоцитов. Тела нервных клеток спинномозговых узлов окружены слоем клеток глии, которые здесь называются ганглионарными глиоцитами (gliocyti ganglii) (см. рис. 11.2, б, в). Они расположены вокруг тела нейрона и имеют округлые ядра. Снаружи глиальная оболочка тела нейрона покрыта тонковолокнистой соединительнотканной оболочкой. Клетки этой оболочки отличаются овальной формой ядер.
Нейроны в составе спинномозгового узла гетероморфны. Среди них различают малые, отвечающие за болевую и температурную чувствительность; большие нейроны специализированы на проприорецепции; промежуточные нейроны относятся к тактильным.
11.3. ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА 11.3.1. Спинной мозг
Спинной мозг (medulla spinalis) состоит из двух симметричных половин, отграниченных друг от друга спереди глубокой срединной щелью, а сзади - соединительнотканной перегородкой (рис. 11.3). В спинном мозге различают центральный канал, темное серое вещество (substantia grisea) и светлое белое вещество (substantia alba). Центральный канал выстлан эпендимоцитами. Он остается открытым от рождения до 17 лет жизни человека. В дальнейшем с LI по S в спинном мозге формируется интраспинальный орган. В его составе находятся олигодендроциты и эндокриноциты, вырабатывающие вазоактив-ный пептид. В последующем (с 36 лет) данный орган замещается глиально-волокнистыми структурами.
Серое вещество на поперечном сечении мозга представлено в виде буквы «Н» или бабочки. Выступы серого вещества принято называть рогами. Различают передние, или вентральные, задние, или дорсальные, и боковые, или латеральные, рога (cornu ventrale, cornu dorsale, cornu laterale). В процессе развития спинного мозга из нервной трубки образуются нейроны, которые группируются в 10 слоях, или в пластинах (пластины Рекседа). Для всех млекопитающих и человека характерна следующая архитектоника указанных пластин: I-V пластины соответствуют задним рогам, VI и VII пластины - промежуточной зоне, VIII и IX пластины - передним рогам, X пластина - зона околоцентрального канала (см. рис. 11.3, б). Такое деление на пластины дополняет организацию структуры серого вещества спинного мозга, основывающейся на локализации ядер. На поперечных срезах более отчетливо
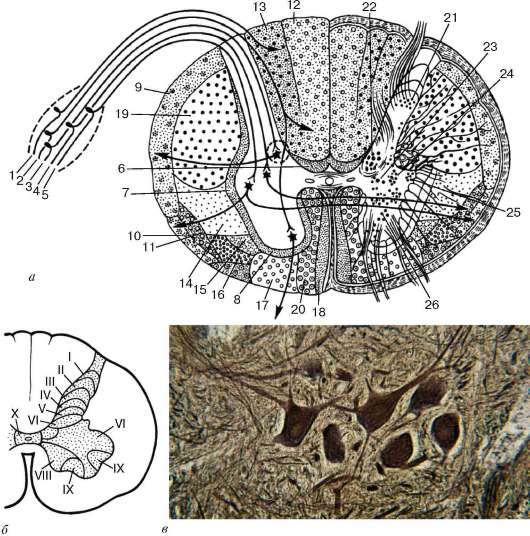
Рис. 11.3. Строение спинного мозга:
а - разрез спинного мозга и спинномозгового ганглия (схема по Т. Н. Радостиной и Л. С. Румянцевой): 1, 2 - рефлекторные пути сознательных проприоцептивных ощущений и осязания; 3, 4 - рефлекторные пути проприоцептивных импульсов; 5 - рефлекторные пути температурной и болевой чувствительности; 6 - задний собственный пучок; 7 - боковой собственный пучок; 8 - передний собственный пучок; 9 - задний спинно-мозжечковый путь; 10 - передний спинно-мозжечковый путь; 11 - спиноталамический путь; 12 - нежный пучок (пучок Голля); 13 - клиновидный пучок (пучок Бурдаха); 14 - руброспинальный путь; 15 - таламоспинальный путь; 16 - вестибулоспинальный путь; 17 - ретикулоспинальный путь; 18 - текто-спинальный путь; 19 - кортикоспинальный (пирамидный) боковой путь; 20 - кор-тикоспинальный пирамидный передний путь; 21 - собственное ядро заднего рога; 22 - грудное ядро; 23, 24 - ядра промежуточной зоны; 25 - боковое ядро (симпатическое); 26 - ядра переднего рога; б - схема расположения пластин Рекседа, соответствующих топографии ядер: I-V - задние рога; VI-VII - промежуточная зона; VIII-IX - передние рога; X - зона около центрального канала; в - микрофотография мотонейронов ядер переднего рога (импрегнация нитратом серебра)
видны ядерные группы нейронов, а на сагиттальных - лучше видно пластинчатое строение, где нейроны группируются в колонки. Каждая колонка нейронов соответствует определенной области на периферии тела. В пластинах нейроны группируются в модули. Каждый из них соответствует определенной территории на периферии (соматотопическая организация).
Серое вещество спинного мозга состоит из тел нейронов, безмиелиновых и тонких миелиновых волокон, макро- и микроглии и кровеносных сосудов. Основной составной частью серого вещества, отличающей его от белого, являются мультиполярные нейроны.
Белое вещество спинного мозга представляет собой совокупность продольно ориентированных преимущественно миелиновых волокон, гетеро-топических нейронов, фибриллярных астроцитов и гемокапилляров. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга.
Нейроны. Клетки, сходные по размерам, ультрамикроскопическому строению и функциональному значению, лежат в сером веществе группами, которые называются ядрами. Среди нейронов спинного мозга можно выделить следующие виды клеток: корешковые клетки (neurocytus radiculatus), нейриты которых покидают спинной мозг в составе его передних корешков, внутренние клетки, или интернейроны (neurocytus internus), отростки которых заканчиваются синапсами в пределах серого вещества спинного мозга, и пучковые клетки (neurocytus funicularis), аксоны которых проходят в белом веществе обособленными пучками волокон, несущими нервные импульсы от определенных ядер спинного мозга в его другие сегменты или в соответствующие отделы головного мозга, образуя проводящие пути. Отдельные участки серого вещества спинного мозга значительно отличаются друг от друга по составу нейронов, нервных волокон и нейроглии.
Серое вещество мозга состоит из мультиполярных нейронов трех типов. Первый тип нейронов является филогенетически более древним и характеризуется немногочисленными длинными, прямыми и слабо ветвящимися дендритами (изодендри-тический тип). Такие нейроны преобладают в промежуточной зоне и встречаются в передних и задних рогах. Второй тип нейронов имеет большое число сильно ветвящихся дендритов, которые переплетаются, образуя «клубки» (идиодендритический тип). Они характерны для ядер передних рогов, а также для задних рогов (ядра студневидного вещества, ядро Кларка). Третий тип нейронов по степени развития ден-дритов занимает промежуточное положение между первым и вторым типами. Они расположены в передних (дорсальная часть) и задних (вентральная часть) рогах, типичны для собственного ядра заднего рога.
В задних рогах различают губчатый слой, желатинозное вещество, собственное ядро заднего рога и грудное ядро. Между задними и боковыми рогами серое вещество вдается тяжами в белое, вследствие чего образуется его сетеобразное разрыхление, получившее название сетчатого образования.
Губчатый слой задних рогов характеризуется широкопетлистым глиаль-ным остовом, в котором содержится большое количество мелких вставочных нейронов.
В желатинозном веществе преобладают глиальные элементы. Нервные клетки здесь мелкие, и количество их незначительно.
Задние рога богаты диффузно расположенными вставочными клетками. Это мелкие мультиполярные ассоциативные и комиссуральные клетки, аксоны которых заканчиваются в пределах серого вещества спинного мозга той же стороны (ассоциативные клетки) или противоположной стороны (комиссуральные клетки).
Нейроны губчатой зоны, желатинозного вещества и вставочные клетки осуществляют связь между чувствительными клетками спинномозговых узлов и мотонейронами передних рогов, замыкая местные рефлекторные дуги.
Из структур заднего рога особый интерес представляют студневидное вещество, которое тянется непрерывно вдоль спинного мозга в I-IV пластинах. Нейроны продуцируют энкефалин - пептид опиоидного типа, инги-бирующий болевые эффекты. Нейроны IV пластины отвечают на слабые тактильные сигналы, являются ГАМК-эргическими. Студневидное вещество оказывает тормозное действие на функции спинного мозга путем контроля за поступающей в него сенсорной информацией - кожной и частично висцеральной и проприоцептивной.
Нейроны I и III пластин, выделяя метэнкефалин и нейротензин, снимают или уменьшают болевые эффекты, которые индуцируются импульсами с тонких корешковых волокон с веществом Р. В середине заднего рога располагается собственное ядро заднего рога. Оно состоит из вставочных нейронов, аксоны которых переходят через переднюю белую спайку на противоположную сторону спинного мозга в боковой канатик белого вещества, где они входят в состав вентрального спинно-мозжечкового и спинно-таламического путей и направляются в мозжечок и таламус (зрительный бугор).
V пластина. Грудное ядро (ядро Кларка) состоит из крупных вставочных нейронов с характерным расположением хроматофильного вещества (мелкая зернистость сосредоточена на периферии, а распыленная - в центре цитоплазмы. Клетки имеют сильно разветвленные дендриты. Их аксоны выходят в боковой канатик белого вещества той же стороны. Нейроны ядра Кларка получают информацию от рецепторов мышц, сухожилий и суставов (проприоцептивная чувствительность) по самым толстым корешковым волокнам и передают ее в мозжечок по заднему спинно-мозжечковому пути. Второй путь в головной мозг образует таламический канал, связанный с корой большого мозга (задняя центральная извилина).
Пластины IV-VI содержат крупные мультиполярные нейроны, к которым поступает большой объем тактильной и болевой сенсорной информации. Аксоны нейронов формируют путь общей чувствительности, заканчиваются на нейронах таламуса.
В VII пластине сосредоточены тормозные интернейроны Реншоу, связанные с коллатералями мотонейронов.
В промежуточной зоне (латеральные отделы VII пластины) расположены центры автономной нервной системы - преганглионарные холинергические нейроны ее симпатического и парасимпатического отделов. На уровне ThI-LII находится центр
симпатического отдела, куда от псевдоуниполярных клеток спинномозговых узлов входят аксоны с веществом Р и глутаминовой кислотой (висцеральная чувствительность), а также волокна из медиального ядра промежуточной зоны с холецистоки-нином, соматостатином и ВИП (вазоинтестинальным пептидом), волокна с нор-адреналином и серотонином (из голубоватого места и ядер шва). Парасимпатические преганглионарные нейроны находятся на уровне СIII-SII спинного мозга.
Таким образом, сенсорная чувствительность в спинном мозге имеет пространственную ориентацию. Экстероцептивная чувствительность (болевая, температурная, тактильная) связана с нейронами студневидного вещества и собственного ядра заднего рога, висцеральная - с нейронами промежуточной зоны, проприоцептив-ная - с ядром Кларка и тонким клиновидным ядром.
В VIII пластине расположено интерстициальное ядро Кахаля с интернейронами, переключающими информацию от псевдоуниполярных нейронов спинномозговых узлов на мотонейроны передних рогов спинного мозга.
IX пластина. В передних рогах расположены самые крупные нейроны спинного мозга, которые имеют диаметр тела 100-150 мкм и образуют значительные по объему ядра. Это корешковые клетки, поскольку их нейриты составляют основную массу волокон передних корешков. В составе смешанных спинномозговых нервов они поступают на периферию и образуют нейромышечные синапсы в скелетной мускулатуре. Следовательно, эти ядра представляют собой моторные соматические центры. В передних рогах наиболее выражены медиальная и латеральная группы моторных клеток. Первая иннервирует мышцы туловища и развита хорошо на всем протяжении спинного мозга. Вторая находится в области шейного и поясничного утолщений и иннервирует мышцы конечностей. Центральная группа мотонейронов иннервирует мышцы тазового и плечевого пояса.
Мотонейроны объединены в пять групп двигательных ядер - латеральные (переднее и заднее), медиальные (переднее и заднее), центральная. В ядрах нейроны образуют колонки. Различают крупные альфа-мотонейроны, аксоны которых образуют нейромышечные синапсы на экстрафузальных мышечных волокнах, вызывают быстрые фазические сокращения; альфа-мотонейроны малые - поддерживают тонус скелетной мускулатуры, мелкие гамма-мотонейроны иннервируют интрафу-зальные мышечные волокна нервно-мышечных веретен.
В терминалях аксона имеются синаптические пузырьки с ацетилхолином, на теле нейрона и дендритах - многочисленные синапсы - до 1000 и более аксосо-матических терминалей, образованных центрифугальными волокнами, аксонами интернейронов VI-VIII пластин, афферентными волокнами задних корешков.
X пластина - здесь располагаются интернейроны, связывающие центральное серое вещество с другими участками серого вещества спинного мозга.
В сером веществе спинного мозга много рассеянных пучковых нейронов. Аксоны этих клеток выходят в белое вещество и сразу же делятся на более длинную восходящую и более короткую нисходящую ветви. В совокупности эти волокна образуют собственные, или основные, пучки белого вещества, непосредственно прилегающие к серому веществу. По своему ходу они дают много коллатералей, которые, как и сами ветви, заканчива-
ются синапсами на мотонейронах передних рогов 4-5 смежных сегментов спинного мозга.
Глиоциты спинного мозга. Спинномозговой канал выстлан эпендимоцитами. Это цилиндрические клетки. Апикальная часть клетки имеет микроворсинки и реснички (киноцилии). Посредством микрофиламентов, десмосом, щелевых контактов плазмолеммы боковой поверхности клетки объединяются в эпендиму - эпителиоподобную выстилку. Боковые поверхности соседних клеток образуют множество взаимных интердигитаций. Овальное ядро располагается у основания клетки, а органеллы - в апикальной половине эпендимоцита. Базальный отросток клетки содержит большое количество глиофиламентов. Эпендимоциты осуществляют трансцеллюлярный транспорт веществ и в той или иной степени секреторную функцию.
Основную часть остова серого вещества составляют протоплазматиче-ские и фиброзные астроциты. Отростки волокнистых астроцитов выходят за пределы серого вещества и вместе с элементами соединительной ткани принимают участие в образовании перегородок в белом веществе и глиальных мембран вокруг кровеносных сосудов и на поверхности спинного мозга. Олигодендроциты входят в состав оболочек нервных волокон.
Микроглия поступает в спинной мозг по мере врастания в него кровеносных сосудов и распределяется в сером и белом веществе. Клетки микро-глии - наиболее мелкие из всех глиальных элементов и реже встречаются в центральной нервной системе. Микроглиоциты составляют около 3 % всех клеток центральной нервной системы, располагаются в сером и белом веществе мозга и часто сопровождают нервные клетки. Показано, что около 50 % клеток микроглии - это макрофаги мозга, происходящие от моноцитов крови. Другая половина микроглиальных клеток - это «покоящиеся астроциты», способные при различных условиях к активной пролиферации и дифференцировке в астроциты.
11.3.2. Головной мозг
В головном мозге различают серое и белое вещество, но распределение этих двух составных частей здесь значительно сложнее, чем в спинном мозге. Большая часть серого вещества головного мозга располагается на поверхности большого мозга и в мозжечке, образуя их кору. Меньшая часть образует многочисленные ядра ствола мозга. Каждое ядро представляет собой нервный центр, где переключаются и качественно изменяются нервные импульсы. Ядра образуются в результате миграции клеток и формирования между ними связей. Каждый нейрон в составе ядра посылает сигналы одновременно нескольким десяткам тысяч клеток, и сам получает не меньшую по объему информацию. Проводящие пути интегрируют ядра в системы, регулирующие сенсорную, двигательную, нейроэндокринную активность, управляют сном и бодрствованием, другими физиологическими ритмами.
Ствол мозга
Проводящие пути и детали строения ствола мозга подробно описаны в курсах нормальной анатомии и неврологии. В состав ствола мозга входят продолговатый мозг, мост, мозжечок и структуры среднего и промежуточного мозга. Все ядра серого вещества ствола мозга состоят из мультиполяр-ных нейронов.
Ядра ствола мозга подразделяются на чувствительные, двигательные и переключательные (ассоциативные).
В чувствительных ядрах, несмотря на такое название, однако, сосредоточены вставочные по функции нейроны, на которых оканчиваются аксоны псевдоуниполярных или биполярных нейронов чувствительных ганглиев (см. выше). Эти ядра занимают дорсальную сторону ствола мозга и, по сути, аналогичны нейронам задних рогов спинного мозга. Чувствительные ядра формируют латерально расположенную соматосенсорную зону и лежащую медиально висцеросенсорную зону. Нейроны ядра воспринимают сенсорную информацию от экстерорецепторов головы, болевую, температурную информацию, импульсы от рецепторов слизистых оболочек носовой и ротовой полости, рецепторов гравитации и др.
В двигательных ядрах, которые занимают вентральную сторону ствола мозга, находятся как мотонейроны, иннервирующие соматическую мускулатуру головы, глаза, языка, глотки, гортани и другое, так и нейроны, аксоны которых формируют преганглионарные волокна парасимпатического отдела автономной нервной системы.
В ассоциативных (переключательных) ядрах много нейронов, которые обеспечивают переключение импульсов из спинного мозга и ствола мозга на нейроны коры и в обратном направлении.
Ствол головного мозга связан с периферическими органами афферентными и эфферентными волокнами черепных нервов.
К черепным нервам относят ядра подъязычного, добавочного, блуждающего, языкоглоточного, преддверно-улиткового нервов продолговатого мозга; отводящего, лицевого, тройничного нервов моста.
Аппарат черепных нервов вполне аналогичен собственному аппарату спинного мозга. Собственный внутренний аппарат ствола головного мозга обеспечивает связи между его частями.
Рефлекторные дуги построены не из простой цепочки нейронов, а включают группы нейронов, в которых возможны осуществление пре- и постси-наптического торможения и модуляция с помощью нейропептидов интернейронов потока афферентных и эфферентных сигналов. Группировки нейронов являются общим принципом внутренней организации корковых и ядерных структур мозга. Объединять нейроны в единую функциональную систему могут коллатерали афферентного отростка и интернейроны (продуцирующие нейропептиды), модифицирующие проведение нервного импульса.
Различные части ствола головного мозга тесно взаимосвязаны благодаря наличию внутреннего собственного рефлекторного аппарата. Важную роль в установлении этих взаимосвязей играет также аппарат двусторонних свя-
зей спинного мозга и стволовой части головного мозга, который включает восходящие и нисходящие пути.
Продолговатый мозг. Продолговатый мозг характеризуется присутствием выше перечисленных ядер черепных нервов, которые концентрируются преимущественно в его дорсальной части, образующей дно IV желудочка. Они содержат крупные мультиполярные нервные клетки, нейриты которых образуют синаптические связи с клетками мозжечка и таламуса. В нижние оливы поступают волокна от мозжечка, красного ядра, ретикулярной формации и спинного мозга, с которыми нейроны нижних олив связаны особыми волокнами. В центральной области продолговатого мозга располагается ретикулярная формация. Это сетчатое образование начинается в верхней части спинного мозга и тянется через продолговатый мозг, мост, средний мозг, центральные части таламуса, гипоталамус и другие области, соседние с таламусом. Ретикулярная формация - восходящая диффузно активирующая система головного мозга - располагается между чувствительными и двигательными ядрами, занимает около 9 % объема ствола мозга. В ее сети находятся мультиполярные нейроны (размером от 5 до 120 мкм). В ней преобладают изодендритические нейроны с редкими и мало ветвящимися отростками, имеющими высокую концентрацию синаптических окончаний на всем своем протяжении. Более крупные нейроны, которые образуют длинные восходящие и нисходящие связи, располагаются в ретикулярной формации медиально, а мелкие (ассоциативные) - латерально. Ретикулярная формация получает импульсы через афферентные пути, но сами импульсы проходят через нее в 4-5 раз медленнее, чем через прямые пути. Отростки нейронов ретикулярной формации направляются в кору большого мозга, мозжечка, в ядра ствола мозга, где формируют синапсы (холин-, адрен-, дофаминергические и др.) с нейронами. Так осуществляется интегративная функция ретикулярной формации. Нисходящие волокна нейронов ретикулярной формации взаимодействуют с моторными нейронами спинного мозга. При этом они тормозят их активность. Функции ретикулярной формации связаны с активацией поведения, сменой фаз сна и бодрствования, облегчения или торможения нервных процессов в мозге и др.
Белое вещество в продолговатом мозге занимает преимущественно вен-тролатеральное положение. Основные пучки миелиновых нервных волокон представлены кортико-спинальными пучками (пирамиды продолговатого мозга), лежащими в его вентральной части. В боковых областях располагаются веревчатые тела, образованные волокнами спинно-мозжечковых путей. Отсюда эти волокна поступают в мозжечок. Отростки нейронов ядер клиновидного и тонкого пучков в виде внутренних дуговых волокон пересекают ретикулярную формацию, перекрещиваются по средней линии, образуя шов, и направляются к таламусу.
Мост делится на дорсальную (покрышковую) и вентральную части. Дорсальная часть содержит волокна проводящих путей продолговатого мозга, ядра V-VIII черепных нервов, ретикулярную формацию моста. В вентральной части располагаются собственные ядра моста и волокна пирамидных путей, идущие продольно. Ядра моста построены из мультиполярных нейронов, размеры и форма которых в различных ядрах неодинаковы.
К переключательным ядрам задней части моста относятся верхнее оливное ядро, ядра трапециевидного тела и ядро латеральной петли. Центральные отростки нейронов улиткового узла заканчиваются на переднем и заднем улитковых ядрах продолговатого мозга. Аксоны нейронов переднего улиткового ядра заканчиваются в верхнем оливном ядре и ядрах трапециевидного тела. Аксоны верхнего оливного ядра, заднего улиткового ядра и ядер трапециевидного тела образуют латеральную петлю. В состав последней входят также клетки ядра латеральной петли и их отростки. Латеральная петля заканчивается в первичных слуховых центрах - нижнем холмике крыши среднего мозга и медиальном коленчатом теле.
Средний мозг состоит из крыши среднего мозга (четверохолмия), покрышки среднего мозга, черного вещества и ножек мозга. Четверохолмие состоит из пластинки крыши, двух ростральных (верхних) и двух каудальных (нижних) холмиков. Ростральные холмики (звено зрительного анализатора) характеризуются послойным расположением нейронов, каудальные (часть слухового анализатора) построены по ядерному принципу. В покрышке среднего мозга находится до 30 ядер, и в том числе красное ядро. Красное ядро состоит из крупноклеточной и мелкоклеточной частей. Крупноклеточная часть получает импульсы из базальных ганглиев конечного мозга и передает сигналы по руброспинальному тракту в спинной мозг, а по колла-тералям руброспинального тракта - в ретикулярную формацию. Мелкие нейроны красного ядра возбуждаются импульсами из мозжечка по церебел-лорубральному тракту и посылают импульсы в ретикулярную формацию. Черное вещество получило свое название в связи с тем, что в его мелких веретенообразных нейронах содержится меланин. Ножки мозга образованы миелиновыми волокнами, идущими от коры большого мозга.
Промежуточный мозг. Филогенетически новой надстройкой в ростральной части ствола является промежуточный мозг (таламический мозг и гипо-таламическая область). В промежуточном мозге преобладает по объему таламус, который состоит практически из серого вещества, разделенного прослойками белого вещества на ядра. К ним подходят восходящие (афферентные) проводящие пути и формируются синапсы с нейронами таламуса. Отростки последних образуют связи (лучистый венец) с нейронами гипоталамуса, нейронами коры большого мозга и нейронами различных центров промежуточного мозга. Нервные импульсы к таламусу идут по экстрапирамидному двигательному пути.
В каудальной группе ядер (подушка таламуса) заканчиваются волокна зрительного пути. Вентрально от него располагается богатая мелкими ядрами гипоталамическая (подбугорная) область.
Гипоталамическая область ствола мозга - высший вегетативный центр, содержащий комплекс ядер (более 40), тесно связанных с центральной нервной системой с помощью афферентных и эфферентных путей. Гипоталамус участвует в регуляции температуры, кровяного давления, водного, жирового обмена и др. Среди ядер гипоталамуса особо выделяются те, которые синтезируют нейрогормоны (см. рис. 10.5, в). В отличие от нейронов, например, коры большого мозга, нейроны гипоталамических ядер обладают двойной
чувствительностью: к действию нейромедиаторов, выделяемых в области межнейрональных синапсов; к действию физико-химических факторов окружающей среды (температура, осмотическое давление, концентрация глюкозы и др.).
Мозжечок
Мозжечок представляет собой центральный орган равновесия и координации движений. Он связан со стволом мозга афферентными и эфферентными проводящими пучками, образующими в совокупности три пары ножек мозжечка. Двусторонние связи мозжечка со стволом и корой большого мозга позволяют мозжечку также участвовать в организации внимания, долгосрочной памяти, речевой деятельности мозга и др. На поверхности мозжечка много извилин и борозд, которые значительно увеличивают ее площадь (у взрослых людей 975-1500 см2). Борозды и извилины создают на разрезе характерную
для мозжечка картину «древа жизни». Основная масса серого вещества в мозжечке располагается на поверхности и образует его кору. Меньшая часть серого вещества лежит глубоко в белом веществе в виде центральных ядер. В центре каждой извилины имеется тонкая прослойка белого вещества, покрытая слоем серого вещества - корой (рис. 11.4, I, II).
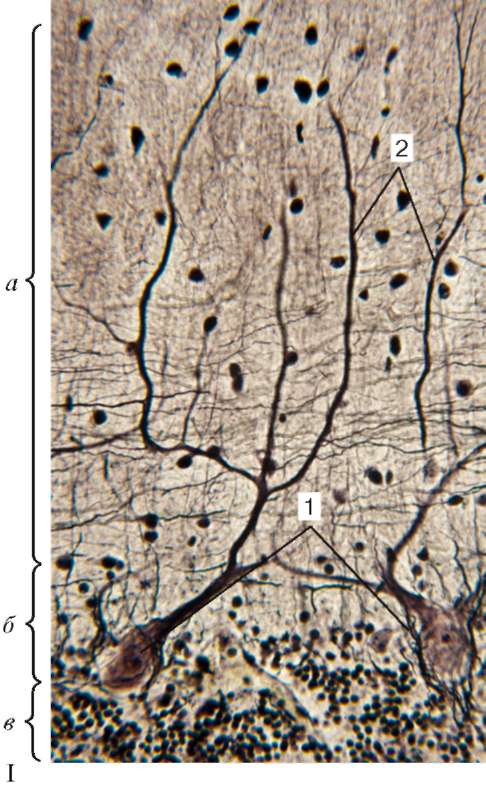
Рис. 11.4. Строение мозжечка (импрегнация нитратом серебра):
I - микрофотография: а - молекулярный слой; б - ганглионарный слой (слой грушевидных нейронов); в - зернистый слой; 1 - тела грушевидных нейронов;
2 - разветвления дендритов грушевидных нейронов.
Развитие. Мозжечок образуется за счет разрастания дорсолатераль-ной стенки нервной трубки в области заднего мозга. В первые недели развития человека миграция нейро-бластов матричной зоны приводит к закладке ядер и клеток Пуркинье. На 9-11-й нед матричные стволовые клетки отделяются от эпендим-ного слоя и мигрируют (первичная миграция) на поверхность зачатка мозжечка. Там они образуют наружный герминативный слой (к 21-й нед развития его толщина составляет 6-9 клеточных слоев). Отсюда клетки, дифференцируясь в нейробла-сты, мигрируют в обратном направлении. Продвижение клеток через слой клеток Пуркинье направляет радиальная (бергмановская) глия. Скапливаясь в глубине коры ниже слоя клеток Пуркинье, нейробласты
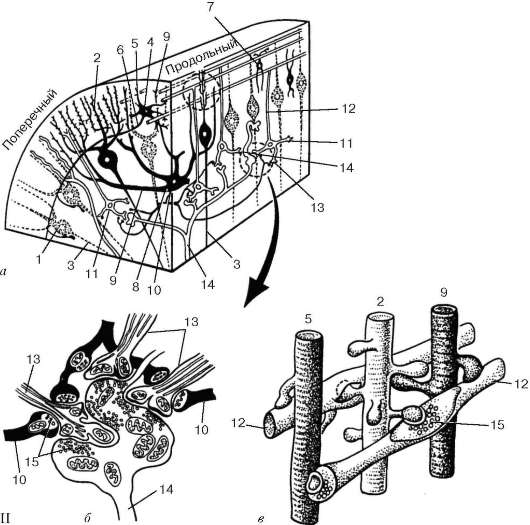
Рис. 11.4. Продолжение
II - схема синаптических связей нейронов в коре мозжечка (по Сентагатаи): а - извилина мозжечка (пунктирной линией на продольном срезе ограничены участки, показанные на рис. б и в); б - клубочек зернистого слоя; в - синапсы в молекулярном слое. 1 - клетка Пуркинье (грушевидный нейрон); 2 - дендриты грушевидных нейронов; 3 - аксон грушевидного нейрона; 4 - корзинчатые нейроны; 5 - дендриты и 6 - аксон корзинчатого нейрона; 7 - звездчатый нейрон; 8 - большие звездчатые нейроны; 9 - дендриты большого звездчатого нейрона; 10 - аксон большого звездчатого нейрона; 11 - клетки-зерна (зернистые нейроны); 12 - аксон клетки-зерна; 13 - дендриты клеток-зерен; 14 - моховидные нервные волокна; 15 - синаптические пузырьки. В черный цвет окрашены тормозящие нейроны
формируют внутренний зернистый слой коры. Такая миграция нейробла-стов способствует по мере их созревания формированию послойной структуры коры мозжечка и образованию характерных для нее внутренних меж-нейрональных связей. Процессы дифференциации в коре продолжаются и после рождения. Так, поверхностный герминативный слой сохраняется в
мозжечке человека в течение первого года жизни. До 5 лет индивидуальная вариабельность строения коры мозжечка выражена слабо. Однако продолжающиеся гистогенетические процессы, которые заключаются в росте и усложнении ветвлений отростков нейронов коры, процессы их специализации к 20 годам жизни приводят к отчетливой индивидуальной вариабельности строения гистологических элементов коры.
Строение. В коре мозжечка различают три слоя: наружный - молекулярный (stratum moleculare), средний - слой клеток Пуркинье (грушевидных нейронов, stratum Purkinjense) и внутренний - зернистый (stratum granulosum). Слой клеток Пуркинье содержит нейроны грушевидной формы. В мозжечке взрослого человека среднее количество клеток Пуркинье составляет 30,5?106. Аксоны клеток Пуркинье отходят от оснований их тел, направляются через зернистый слой в белое вещество и заканчиваются синапсами на нейронах ядер мозжечка и латерального вестибулярного ядра.
Грушевидные нейроны располагаются строго в один ряд. От их крупного (70X35 мкм) тела в молекулярный слой отходят 2-3 дендрита, которые, обильно ветвясь, пронизывают всю толщу молекулярного слоя. Дендриты покрыты огромным количеством шипиков. Все ветви дендритов располагаются только в одной плоскости, перпендикулярной к направлению извилин, поэтому при поперечном и продольном сечении извилин дендриты грушевидных нейронов выглядят различно. На долю клеток Пуркинье приходится до 73 % всех синапсов на нейронах ядер мозжечка. Среди грушевидных нейронов располагаются еще несколько видов нейронов - клетки-канделябры и разновидность клеток Гольджи - нейроны Пензы. Клетки-канделябры относятся к разновидностям тормозных ГАМК-эргических интернейронов, оказывающих свое влияние на дендриты клеток Пуркинье, тогда как клетки Пензы влияют на клетки-зерна. В клетках Пуркинье присутствует несколько специфических нейрохимических веществ. В них обнаружены: кальбиндин, ГАМК и NO-синтаза.
Молекулярный слой содержит два основных вида нейронов: корзинчатые и звездчатые. Корзинчатые нейроны (neuron corbiferum) располагаются на нескольких уровня: в глубине слоя находятся самые крупные, в верхних - более мелкие клетки. Тела корзинчатых нейронов имеют размеры от 8 до 20 мкм. Они составляют до 20 % клеток молекулярного слоя. Гистохимически в нейронах выявляются ГАМК, парвальбумин, кальбин-дин, соматостатин, NO-синтаза. Наиболее характерная черта этих нейронов - их аксонные системы: горизонтально расположенный аксон распространяется более чем на 500 мкм параллельно поверхности извилины мозжечка. По ходу он дает многочисленные восходящие и нисходящие кол-латерали. Одна корзинчатая клетка через свою аксонную систему связана примерно с 250 клетками Пуркинье. Коллатерали, спускающиеся к телам грушевидных нейронов, и совместно с другими волокнами, густо оплетая эти нейроны, формируют на них характерную структуру корзинок нервных волокон (corbis neurofibrarum). К клетке Пуркинье сходятся аксоны примерно 20-30 корзинчатых клеток, охватывая все тело нейрона и начальный сегмент аксона, образуя ГАМКергические тормозные синапсы. В свою очередь,
аксонные коллатерали оказывают тормозное влияние и на корзинчатые клетки молекулярного слоя, которые, как указывалось выше, оказывают, в свою очередь, тормозные влияния на клетки Пуркинье, формируя таким образом своеобразные локальные тормозные «петли» в пределах слоя грушевидных нейронов и молекулярного слоя коры мозжечка.
Звездчатые нейроны (neuron stellatum) имеют размеры 15-20 мкм в диаметре, от их тела радиально отходят дендриты и аксоны. Различают коротко- и длинноаксонные звездчатые нейроны. Медиатором в образуемых ими синапсах, по-видимому, является таурин и ГАМК. Звездчатые нейроны лежат выше корзинчатых. Мелкие звездчатые нейроны снабжены тонкими короткими дендритами и слаборазветвленными нейритами, образующими синапсы на дендритах грушевидных нейронов. Крупные звездчатые нейроны, в отличие от мелких, имеют длинные и сильно разветвленные дендри-ты и аксоны. Ветви их аксонов соединяются с дендритами грушевидных нейронов, но некоторые из них достигают тел грушевидных нейронов и входят в состав так называемых корзинок. Корзинчатые и звездчатые нейроны молекулярного слоя представляют собой единую систему вставочных нейронов, передающую тормозные нервные импульсы на дендриты и тела клеток Пуркинье.
Очень богат нейронами зернистый слой. Первым типом клеток этого слоя можно считать клетки-зерна, или зернистые нейроны (neuron granulosum). Они имеют диаметр 7-10 мкм, круглое крупное ядро. Короткие дендриты (3-4) заканчиваются в этом же слое миниатюрной бифуркацией в виде «коготка». Вступая в синаптическую связь с окончаниями приходящих в мозжечок возбуждающих афферентных (моховидных) волокон, дендриты зернистых нейронов образуют характерные структуры, именуемые клубочком (гломеру-лой) мозжечка (glomerulus cerebellari).
Аксоны клеток-зерен проходят в молекулярный слой и в нем Т-образно делятся на две ветви, ориентированные параллельно поверхности коры вдоль извилин мозжечка. Преодолевая большие расстояния, эти параллельные волокна пересекают ветвления дендритов многих клеток Пуркинье и образуют с ними и дендритами корзинчатых и звездчатых нейронов синапсы. Таким образом, аксоны клеток-зерен передают возбуждение, полученное ими от моховидных волокон, на значительное расстояние многим грушевидным нейронам.
Вторым типом клеток зернистого слоя мозжечка являются тормозные большие звездчатые нейроны (neuron stellatum magnum), или клетки Гольджи. Различают два вида таких клеток: с короткими и длинными аксонами. Нейроны с короткими аксонами (neuronum stellatum breviaxonicum) лежат вблизи слоя грушевидных нейронов. Их разветвленные дендриты распространяются в молекулярном слое и образуют синапсы с параллельными волокнами - аксонами клеток-зерен. Аксоны направляются в зернистый слой к клубочкам мозжечка и заканчиваются тормозными ГАМКергическими синапсами на концевых ветвлениях дендритов клеток-зерен. Полагают, что таким образом клетки Гольджи регулируют количество активных клеток-зерен в микрокомплексах коры мозжечка. В клетках Гольджи выявляются
ацетилхолин и окись азота (NO), энкефалин и соматостатин. Таким образом, клетки Гольджи осуществляют тормозное влияние на уровне зернистого слоя.
Немногочисленные звездчатые нейроны с длинными аксонами (neuronum stellatum longiaxonicum) имеют обильно ветвящиеся в зернистом слое дендри-ты и аксоны, выходящие в белое вещество. Предполагают, что эти клетки обеспечивают связь между различными областями коры мозжечка.
Третьим типом являются крупные мультиполярные нейроны - клетки Лугаро. Клетки Лугаро находятся в верхней части зернистого слоя, непосредственно под слоем клеток Пуркинье, имеют длинные горизонтальные дендриты, идущие в листках мозжечка в сагиттальном направлении. Аксон клетки Лугаро имеет богатые разветвления, проникающие в молекулярный слой. Там он образует множественные тормозные ГАМК-глицинергические синапсы на телах и дендритах корзинчатых и звездчатых нейронов. Гистохимически в клетках Лугаро обнаружен NO. Обнаружены контакты клеток Лугаро с апикальными дендритами клеток Гольджи. Клетки Лугаро имеют множественные связи с возвратными коллатера-лями аксонов клеток Пуркинье, аксонов корзинчатых нейронов и клеток Гольджи. Уникальность клеток Лугаро заключается в их физиологических свойствах - это так называемые молчащие клетки. Они активируются в присутствии серотонина, поступающего по серотонинергическим афферентным волокнам из ядер шва. Их активность связана с регуляцией тормозной активности клеток Пуркинье.
В зернистом слое выявлена популяция мелких клеток, которые были названы униполярными кисточковыми нейронами. Это небольшие клетки (8-12 мкм) с одиночным дендритом, образующим на конце кисточки. Тонкие веточки кисточки тесно переплетаются с терминалью моховидного волокна. Здесь формируется возбуждающий глутаматный синапс. Аксон клетки распространяется только в пределах зернистого слоя и образует возбуждающие глутаматергические синаптические контакты как с дендритами соседних униполярных нейронов, так и с дендритами клеток-зерен, входя в состав клубочков мозжечка. Таким образом, получая возбуждающие влияния от моховидных волокон, они сами оказывают дополнительное возбуждающее воздействие на клетки-зерна.
Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами - моховидными и так называемыми лазящими волокнами. Источниками моховидных волокон являются нейроны нижней оливы, ядра моста, ряд ядер ретикулярной формации и в меньшей степени нейроны ядер мозжечка. В зернистом слое терминали моховидного волокна образуют специализированный тип пресинаптического окончания - розетку. В последней возникают вогнутые участки, которые охватывают пальцевидные утолщения дендритов клеток-зерен. Возникает синаптический комплекс (клубочек мозжечка) радиусом около 2,5 мм, где каждое окончание моховидного волокна является пресинаптической мембраной для дендритов нескольких клеток-зерен. Каждая клетка-зерно получает от моховидных волокон 4-5 возбуждающих синапсов. Сами моховидные волокна обра-
зуют возбуждающие синапсы с 400-600 зернистыми нейронами в пределах нескольких листков мозжечка. В состав клубочка также входят аксоны униполярных кисточковых (brush) нейронов и ГАМКергических клеток Гольджи. ГАМК действует как типичный тормозный медиатор для ГАМК-рецепторов розеток моховидных волокон и клеток-зерен. Клубочек мозжечка рассматривается как структурно-функциональная единица, в которой сочетаются и возбуждающие, и тормозные влияния из разных источников. Это обеспечивает регуляцию деятельности клеток-зерен и соответственно регулирует их влияние на клетки Пуркинье.
Система лазящих волокон (лиановидные, восходящие волокна) является уникальной во всей центральной нервной системе - каждая клетка Пуркинье иннервируется одним волокном. Это следствие постнатальной элиминации мультиволоконной иннервации клеток Пуркинье, в результате гибели части нейронов нижней оливы, достигающей максимума на 1-2-й нед постнатального развития. Каждое волокно от одной клетки нижней оливы иннервирует до семи клеток Пуркинье, образуя на каждой клетке многочисленные аксошипиковые аспартатергические контакты. Коллатерали волокон, проникающие в молекулярный слой, образуют возбуждающие контакты с корзинчатыми и звездчатыми клетками. Кроме лазящих и моховидных волокон, в кору мозжечка входят другие афферентные волокна. Одни их них содержат разнообразные амины, в основном серотонин, а также дофамин, норадреналин и нейропептиды (вещество Р, нейротензин, ангиотензин, галланин, орексин и др.). Основным источником этих волокон являются нейроны ядер шва и голубоватого места. Волокна широко распределяются по слоям мозжечка, образуя по своему ходу многочисленные варикозности. Эти волокна оказывают модулирующее воздействие на нейроны мозжечка, что выражается в регуляции циклов бодрствования и сна, управлении эмоциональным поведением. Также в кору мозжечка входят волокна из гипоталамуса. Полагают, что они функционально связаны с участием мозжечка в регуляции эмоционального поведения человека (например, реакции на страх).
На примере нейронной организации мозжечка успешно разрабатываются модели модульной организации. Например, концепция мозжечкового микрокомплекса позволяет объяснить роль мозжечка в связях с системами спинного мозга, ствола и полушарий конечного мозга. В основе этой концепции лежат данные о закономерном распределении афферентных и эфферентных волокон в продольных зонах мозжечка, а также данные о нейрохимической неоднородности группировок клеток в коре мозжечка. Так, получая сенсорную информацию через афферентные волокна от соответствующих рецептивных полей, группировки клеток Пуркинье и нейронов ядер мозжечка, связанные с соответствующими нейронами красного ядра среднего мозга, могут осуществлять контроль над двигательными реакциями в ответ на раздражение только определенных рецептивных зон. Это обеспечивает четкую топическую организацию афферентных и эфферентных связей мозжечка на уровне микрозон рецепции и определенных моторных единиц.
Кора мозжечка содержит различные глиальные элементы. В зернистом слое имеются волокнистые и протоплазматические астроциты. Ножки отростков волокнистых астроцитов образуют периваскулярные мембраны. Во всех слоях в мозжечке имеются олигодендроциты. Особенно богаты этими клетками зернистый слой и белое вещество мозжечка. В слое клеток Пуркинье между ними лежат глиальные клетки с темными ядрами. Отростки этих клеток направляются к поверхности коры и образуют гли-альные волокна молекулярного слоя мозжечка, поддерживающие ветвления дендритов клеток Пуркинье (gliofibra sustentans). Микроглия в большом количестве содержится в молекулярном слое и в слое клеток Пуркинье.
Кора большого мозга
Кора большого мозга вместе с подкорковым центром (стриатум) входит в состав конечного мозга, который развивается из дорсальных отделов переднего мозгового пузыря. К первому месяцу эмбриогенеза человека возникают основные структуры конечного мозга и начинается формирование слоев неокортекса, закладка борозд, извилин, долей и полей.
Развитие коры полушарий большого мозга (неокортекса) человека в эмбриогенезе происходит из вентрикулярной герминативной зоны конечного мозга (см. главу 10). Первые нейроны, выселяясь из вентрикулярной зоны, формируют «предпластинку» под поверхностью мозгового пузыря (миграция осуществляется изнутри кнаружи). На 7-8-й нед из предпла-стинки возникает поверхностная кортикальная пластинка за счет миграции малодифференцированных нейронов сверху вниз. Эти нейроны образуют 2-6-й слои коры. Миграция нейронов осуществляется двумя способами. На ранних стадиях нейрогенеза - за счет ядерной миграции - перемещения ядра по цитоплазме отростка нейрона. В дальнейшем ведущая роль в процессах миграции принадлежит клеткам Кахаля-Ретциуса, которые вырабатывают секреторный гликопротеин, действующий как аттрактант на нейроны кортикальной пластинки. Это способствует перемещению нейронов по радиальным глиоцитам через ранее возникшие слои нейронов. Кроме того, миграция нейронов может происходить и в тангенциальном направлении.
Дифференцировка и миграция глиоцитов коры происходит за счет миграции клеток из вентрикулярной зоны в вышележащие слои развивающейся коры.
С 15-20-й нед развития человека возникают основные извилины, и далее (с 25-30-й нед) формируются вторичные извилины. Продолжается генерация нейробластов и глиобластов. Этот процесс осуществляется за счет образования клеток в небольших участках вентрикулярной зоны в различные периоды эмбриогенеза (гетерохронно). В каждом из этих участков образуются группы нейронов, последовательно выстраивающихся вдоль одного или нескольких волокон радиальной глии в виде колонки (рис. 11.5, I, II). Подобные, так называемые онтогенетические, колонки в дальнейшем служат основой для формирования функциональных интегративных единиц нео-кортекса: мини- и макроколонок. Для установления сроков формирования
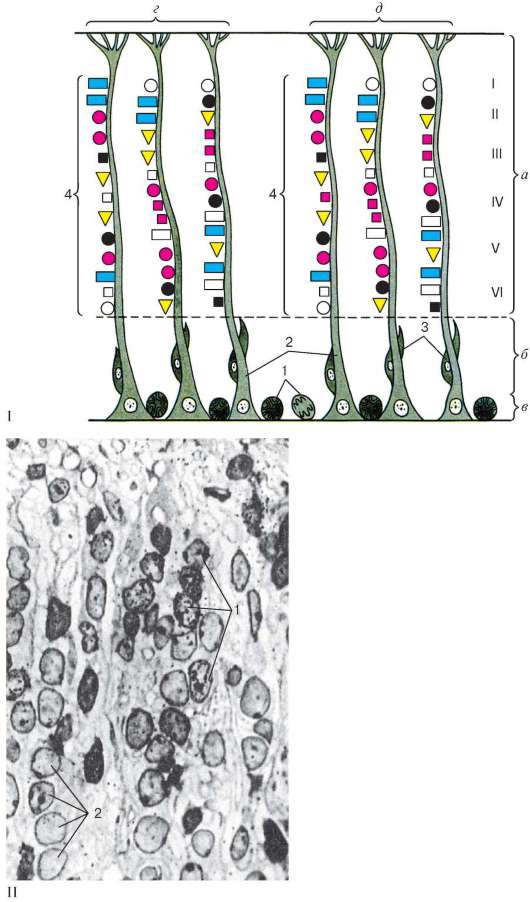
Рис. 11.5. Эмбриогенез коры полушарий большого мозга (неокортекса): I - схема пространственно-временного образования нейронов в неокортек-се млекопитающих (представлены две соседние колонки коры): а - кора (корковая пластинка); б - белое вещество; в - вентрикулярная герминативная зона; г, д - две рядом лежащие колонки неокортекса; I-VI формирующиеся слои коры: 1 - малодифференцирован-ные делящиеся клетки; 2 - радиальные эмбриональные глиоциты; 3 - молодые нейроны, мигрирующие в корковую пластинку; 4 - группы нейронов, последовательно образующиеся в различные сроки эмбриогенеза (по К. Ю. Резникову); II - радиоавтограф неокортекса новорожденной мыши, получившей 3Н-тимидин на 16-е сут эмбриогенеза. Полутонкий срез, окрашенный толуидиновым синим, увеличение 2000: 1 - группы меченных изотопом нейронов, образовавшихся на 16-е сут эмбриогенеза; 2 - нейроны без метки, образовавшиеся в другие сроки эмбриогенеза (препарат К. Ю. Резникова, Г. Д. Назаревской)
в эмбриогенезе различных групп нейронов применяют радиоизотопный метод (см. рис. 11.5, II).
Строение. Кора большого мозга представлена слоем серого вещества толщиной около 3 мм. Наиболее сильно развита она в передней центральной извилине, где ее толщина достигает 5 мм. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга. В коре содержится около 10-14 млрд нервных клеток. Различные участки ее, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток (цитоархитектоника), расположения волокон (миелоархитектоника) и функциональным значением, называются полями. Они представляют собой места высшего анализа и синтеза нервных импульсов. Резко очерченные границы между ними отсутствуют. Для коры характерно расположение клеток и волокон пластинками (слоями) (рис. 11.6).
Цитоархитектоника коры большого мозга. Мультиполярные нейроны коры весьма разнообразны по форме. Среди них можно выделить пирамидные, звездчатые, веретенообразные, паукообразные и горизонтальные нейроны.
Пирамидные нейроны (50-90 %) составляют основную и наиболее специфическую для коры большого мозга форму (см. рис. 11.6, а). Размеры их варьируют от 10 до 140 мкм. Они имеют вытянутое треугольное тело, вершина которого обращена к поверхности коры. От вершины и боковых поверхностей тела отходят дендриты, заканчивающиеся в различных слоях серого вещества. От основания пирамидных клеток берут начало аксоны, в одних клетках короткие, образующие ветвления в пределах данного участка коры, в других - длинные, поступающие в белое вещество.
Пирамидные нейроны различных пластинок коры отличаются размерами и имеют разное функциональное значение. Мелкие клетки представляют собой вставочные нейроны, нейриты которых связывают отдельные участки коры одного полушария (ассоциативные нейроны) или двух полушарий (комиссуральные нейроны). Эти клетки встречаются в разных количествах во всех слоях коры. Особенно богата ими кора большого мозга человека. Аксоны крупных пирамид принимают участие в образовании пирамидных путей, проецирующих импульсы в соответствующие центры ствола и спинного мозга.
Нейроны коры расположены нерезко отграниченными пластинками. Каждая пластинка характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных пластинок: I - молекулярная (lamina molecularis), II - наружная зернистая (lamina granularis externa), III - наружная пирамидная (lamina piramidalis externa), IV - внутренная зернистая (lamina granularis interna), V - внутренняя пирамидная (ганглионарная) (lamina piramidalis interna), VI - мультиформная (lamina multiformis) (см. рис. 11.6, а).
Молекулярная пластинка коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы (см. рис. 11.6, а). Их аксоны проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярной пластинки. Основной объем пластинки занимают терминальные ветвления аксонов клеток нижележащих пластинок:
апикальные (дистальные) дендриты пирамидных нейронов, на которых оканчиваются ГАМКергические аксоны корково-корковых связей; волокна от нейронов неспецифических подкорковых систем мозга (модуляторные норадренергические, дофа-минергические и серотонинергиче-ские волокна из ствола мозга).
Наружная зернистая пластинка образована мелкими пирамидными и звездчатыми нейронами диаметром около 10 мкм. Дендриты этих клеток поднимаются в молекулярный слой. Аксоны или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя. Основная масса синап-
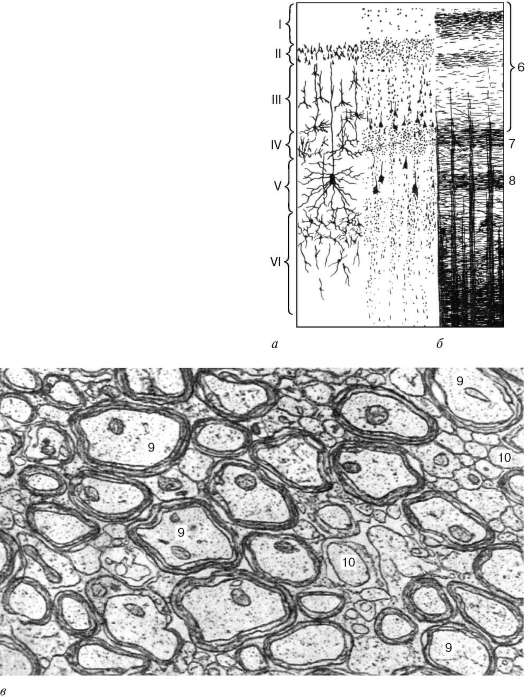
Рис. 11.6. Цито- и миелоархитектоника коры полушарий большого мозга человека (схема):
а - расположение клеток (цитоархитектоника); б - расположение волокон (миелоархитектоника); в - электронная микрофотография белого вещества полушарий большого мозга (препарат И. Г. Павловой); г - схема межнейрональных связей в коре полушарий большого мозга. I, II, III, IV, V, VI - пластинки (слои) коры
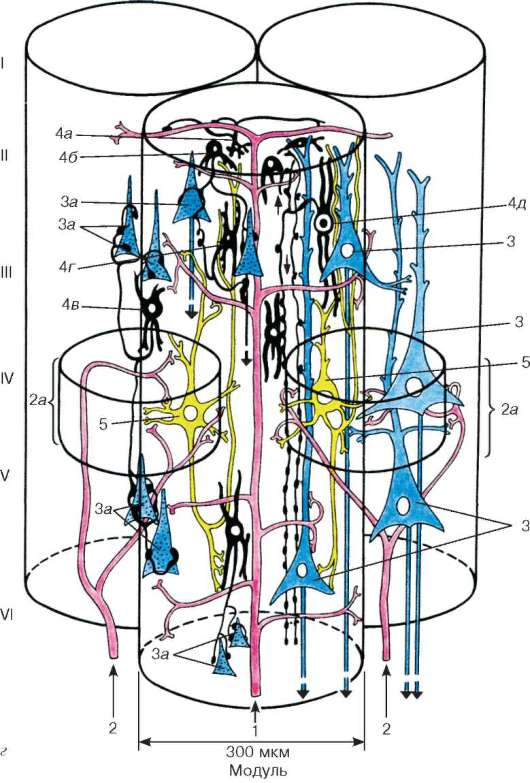
Рис. 11.6. Продолжение Афферентные волокна (обозначены розовым): 1 - кортико-кортикальные; 2 - специфические; 2а - зона распространения специфических афферентных волокон; 3 - пирамидные нейроны (обозначены голубым); 3а - заторможенные пирамидные нейроны (обозначены голубым с черным пунктиром); 4 - тормозные нейроны и их синапсы (обозначены черным цветом); 4а - клетки с аксональной кисточкой; 4б - малые корзинчатые клетки; 4в - большие корзинчатые клетки; 4г - клетки, формирующие аксо-аксональные синапсы; 4д - клетки с двойным букетом дендритов (тормозные нейроны); 5 - шипиковые звездчатые клетки (окрашены в желтый цвет), возбуждающие пирамидные нейроны непосредственно и путем стимуляции клеток с двойным букетом дендритов; 6 - наружный главный слой волокон; 7 - полоска внутренней зернистой пластинки; 8 - полоска внутренней пирамидной пластинки; 9 - миелиновые волокна; 10 - безмиелиновые волокна
сов представлена аксодендритными и аксосоматическими тормозными ГАМКергическими контактами.
Самая толстая пластинка коры большого мозга - наружная пирамидная. Она особенно хорошо развита в прецентральной извилине. Величина пирамидных клеток последовательно увеличивается в пределах 10-40 мкм от наружной зоны пластинки к внутренней. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярной пластинке. Дендриты, берущие начало от боковых поверхностей пирамиды и ее основания, имеют незначительную длину и образуют синапсы со смежными клетками этой пластинки. Аксон пирамидной клетки всегда отходит от ее основания. В мелких клетках он остается в пределах коры; аксон же, принадлежащий крупной пирамидной клетке, обычно формирует миелино-вое ассоциативное или комиссуральное волокно, идущее в белое вещество. Здесь преобладают аксошипиковые контакты, аксосоматические контакты, образованные аксонами тормозных звездчатых нейронов.
Внутренняя зернистая пластинка в некоторых полях коры развита очень сильно (например, в зрительной зоне коры). Однако она может почти отсутствовать (в прецентральной извилине). Пластинка образована мелкими звездчатыми интернейронами. В ее состав входит большое количество горизонтальных волокон. В пластинке преобладают аксодендритные контакты.
Внутренняя пирамидная пластинка (ганглионарный слой) коры образована крупными пирамидными клетками и небольшим числом звездчатых интернейронов. Область прецентральной извилины содержит гигантские пирамидные нейроны, описанные впервые киевским анатомом В. А. Бецем в 1874 г. (клетки Беца). Это очень большие клетки, достигающие в высоту 120 мкм и в ширину 80 мкм. В отличие от других пирамидных клеток коры они характеризуются наличием крупных глыбок хроматофильного вещества. Аксоны клеток этого слоя образуют главную часть кортикоспиналь-ных и кортиконуклеарных путей и оканчиваются синапсами на клетках двигательных ядер.
Прежде чем пирамидный путь покинет кору, от него отходит множество коллатералей. Аксоны клеток Беца дают коллатерали, посылающие тормозящие импульсы в саму кору. Коллатерали волокон пирамидного пути идут в полосатое тело, красное ядро, ретикулярную формацию, ядра моста и нижних олив. Ядра моста и нижних олив передают сигнал в мозжечок. Таким образом, когда пирамидный путь передает сигнал, вызывающий моторную активность, в спинной мозг, одновременно сигналы получают базальные ганглии, ствол мозга и мозжечок. Помимо коллатералей пирамидных путей, существуют волокна, которые идут непосредственно от коры к промежуточным ядрам: хвостатому телу, красному ядру, ядрам ретикулярной формации ствола мозга и др.
Мультиформная пластинка образована нейронами преимущественно веретенообразной формы, а также мелкими полигональными нейронами. Внешняя зона содержит более крупные клетки. Нейроны внутренней зоны мельче и лежат на большом расстоянии друг от друга. Нейриты клеток уходят в белое вещество в составе эфферентных путей головного мозга.
Дендриты достигают молекулярного слоя коры. В этой пластинке преобладают аксодендритные синапсы.
Крупные пирамидные клетки являются основными нейронами, к которым по центрифугальным волокнам приходят импульсы из других отделов центральной нервной системы и передаются через синапсы на их дендриты и тела. От этих клеток импульс уходит по аксонам, формирующим центри-петальные эфферентные пути. Внутри коры между нейронами формируются сложные связи (см. рис. 11.6, б). Медиатором в аксонных окончаниях пирамидных нейронов является аспартат/глутамат, оказывающий возбуждающее действие на клетки-мишени. Кроме того, обнаружены ко-медиаторы и нейропепдиды (энкефалин, ацетилхолин, АТФ).
Исследуя ассоциативную кору, составляющую 90 % неокортек-са, Сентаготаи и представители его школы установили, что структурно-функциональной единицей неокортекса является модуль - вертикальная колонка диаметром около 300 мкм. Модуль организован вокруг кортико-кортикального волокна (см. рис. 11.6, г), представляющего собой волокно, идущее либо от пирамидных клеток того же полушария (ассоциативное волокно), либо от противоположного (комиссуральное). В модуль входят два таламокортикальных волокна (2) - специфических афферентных волокна, оканчивающихся в IV слое коры на шипиковых звездчатых нейронах и базальных дендритах пирамидных нейронов. Каждый модуль, по мнению Сентаготаи, подразделяется на два микромодуля диаметром менее 100 мкм. Всего в неокортексе человека примерно 3 млн модулей.
Аксоны пирамидных нейронов модуля проецируются на три модуля той же стороны и через мозолистое тело на два модуля противоположного полушария. В отличие от специфических афферентных волокон, оканчивающихся в IV слое коры, кортико-кортикальные волокна образуют окончания во всех слоях коры и, достигая I слоя, дают горизонтальные ветви, выходящие далеко за пределы модуля. Помимо специфических афферентных волокон, на пирамидные нейроны (см. рис. 11.6, г) возбуждающее влияние оказывают шипиковые звездчатые нейроны. Различают два вида шипиковых звездчатых клеток: 1) шипиковые звездчатые нейроны фокального типа, образующие множественные синапсы на апикальных дендритах пирамидного нейрона (правая сторона рисунка), и 2) шипиковые звездчатые нейроны диффузного типа, аксоны которых широко ветвятся в IV слое и возбуждают базальные дендриты пирамидных нейронов. Коллатерали аксонов пирамидных нейронов (не показанные на схеме) вызывают диффузное возбуждение соседних пирамидных клеток.
Тормозная система модуля представлена следующими типами нейронов:
1) клетки с аксональной кисточкой образуют в I слое множественные тормозные синапсы на горизонтальных ветвях кортико-кортикальных волокон;
2) корзинчатые нейроны - тормозные нейроны, образующие тормозящие синапсы на телах практически всех пирамидных нейронов. Они подразделяются на малые корзинчатые нейроны, оказывающие тормозящее влияние на пирамиды II, III и V слоев модуля, и большие корзинчатые клетки, располагающиеся на периферии модуля и имеющие тенденцию подавлять
пирамидные нейроны соседних модулей; 3) аксо-аксональные нейроны, тормозящие пирамидные нейроны II и III слоев. Каждая такая клетка образует тормозящие синапсы на начальных участках аксонов сотен нейронов II и III слоев. Они тормозят, таким образом, кортико-кортикальные волокна, но не проекционные волокна нейронов V слоя; 4) клетки с двойным букетом дендритов располагаются во II и III слоях и, тормозя практически все тормозные нейроны, производят вторичное возбуждающее действие на пирамидные нейроны. Ветви их аксонов направлены вверх и вниз и распространяются в узкой колонке (50 мкм). Таким образом, клетка с двойным букетом дендритов растормаживает пирамидные нейроны в микромодуле (в колонке диаметром 50-100 мкм). Мощный возбуждающий эффект фокальных шипиковых звездчатых клеток объясняется тем, что они одновременно возбуждают пирамидные нейроны и клетку с двойным букетом дендри-тов. Таким образом, первые три тормозных нейрона тормозят пирамидные клетки, а клетки с двойным букетом дендритов возбуждают их, угнетая тормозные нейроны.
Система тормозных нейронов играет роль фильтра, тормозящего часть пирамидных нейронов коры.
Различные поля коры характеризуются преимущественным развитием тех или других ее пластинок. Так, в моторных центрах коры, например в передней центральной извилине, сильно развиты III, V и VI и плохо выражены II и IV пластинки. Это так называемый агранулярный тип коры. Из этих областей берут начало нисходящие проводящие пути центральной нервной системы. В чувствительных корковых центрах, где заканчиваются афферентные проводники, идущие от органов обоняния, слуха и зрения, слабо развиты пластинки, содержащие крупные и средние пирамидные нейроны, тогда как зернистые пластинки (II и IV) достигают своего максимального развития. Это гранулярный тип коры.
Миелоархитектоника коры. Среди нервных волокон коры полушарий большого мозга можно выделить ассоциативные волокна, связывающие отдельные участки коры одного полушария, комиссуральные, соединяющие кору различных полушарий, и проекционные волокна, как афферентные, так и эфферентные, которые связывают кору с ядрами низших отделов центральной нервной системы (см. рис. 11.6, б). Эти волокна в коре полушарий образуют радиальные лучи, заканчивающиеся в пирамидном слое. Кроме уже описанного тангенциального сплетения молекулярного слоя, на уровне внутреннего зернистого и ганглионарного слоев расположены два тангенциальных слоя миелиновых нервных волокон - внешняя и внутренняя полосы, которые, очевидно, образуются концевыми ветвлениями афферентных волокон и коллатералей нейритов клеток коры, таких как пирамидные нейроны. Вступая в синаптические связи с нейронами коры, горизонтальные волокна обеспечивают широкое распространение в ней нервного импульса. Строение коры в различных отделах большого мозга сильно варьирует, поэтому детальное изучение ее клеточного состава и хода волокон является предметом специального курса. Кора полушарий большого мозга содержит мощный нейроглиальный аппарат, выполняющий трофическую, защитную, опорную и разграничительную функции.
11.4. АВТОНОМНАЯ НЕРВНАЯ СИСТЕМА
Часть нервной системы, контролирующая висцеральные функции организма, такие как моторика и секреция органов пищеварительной системы, кровяное давление, потоотделение, температура тела, обменные процессы и другое, называется автономной (вегетативной) нервной системой. По своим физиологическим особенностям и морфологическим признакам автономная нервная система делится на симпатическую и парасимпатическую. В большинстве случаев обе системы одновременно принимают участие в иннервации органов.
Автономная нервная система состоит из центральных отделов, представленных ядрами головного и спинного мозга, и периферических: нервных стволов, узлов (ганглиев) и нервных сплетений.
Ядра центрального отдела автономной нервной системы находятся в среднем и продолговатом мозге, а также в боковых рогах грудных, поясничных и крестцовых сегментов спинного мозга. К симпатической нервной системе относятся автономные ядра боковых рогов грудного и верхнепоясничного отделов спинного мозга, к парасимпатической - автономные ядра III, VII, IX и X пар черепных нервов и автономные ядра крестцового отдела спинного мозга. Мультиполярные нейроны ядер центрального отдела представляют собой ассоциативные нейроны рефлекторных друг автономной нервной системы (рис. 11.7). Их нейриты покидают центральную нервную систему через передние корешки спинномозговых нервов или черепные нервы и оканчиваются синапсами на нейронах одного из периферических автономных узлов. Это преганглионарные волокна автономной нервной системы, обычно миелиновые. Преганглионарные волокна симпатической и парасимпатической автономной нервной системы - холинер-гические. Их терминали содержат мелкие светлые синаптические пузырьки (40-60 нм) и одиночные крупные темные везикулы (60-150 нм).
Периферические узлы автономной нервной системы лежат как вне органов (симпатические паравертебральные и превертебральные узлы, парасимпатические узлы головы), так и в стенке органов в составе интрамуральных нервных сплетений пищеварительного тракта, сердца, матки, мочевого пузыря и др.
Паравертебральные узлы расположены по обе стороны позвоночника и со своими соединительными стволами образуют симпатические цепочки.
Превертебральные узлы образуют кпереди от брюшной аорты и ее главных ветвей брюшное сплетение, в состав которого входят чревный, верхний брыжеечный и нижний брыжеечный ганглии.
Автономные узлы снаружи покрыты соединительнотканной капсулой. Прослойки соединительной ткани проникают внутрь узла, образуя его остов. Узлы состоят из мультиполярных нервных клеток, весьма разнообразных по форме и величине. Дендриты нейронов многочисленны и сильно ветвятся. Аксоны в составе постганглионарных (обычно безмиелиновых) волокон поступают в соответствующие внутренние органы. Каждый нейрон и его отростки окружены глиальной оболочкой. Наружная поверхность
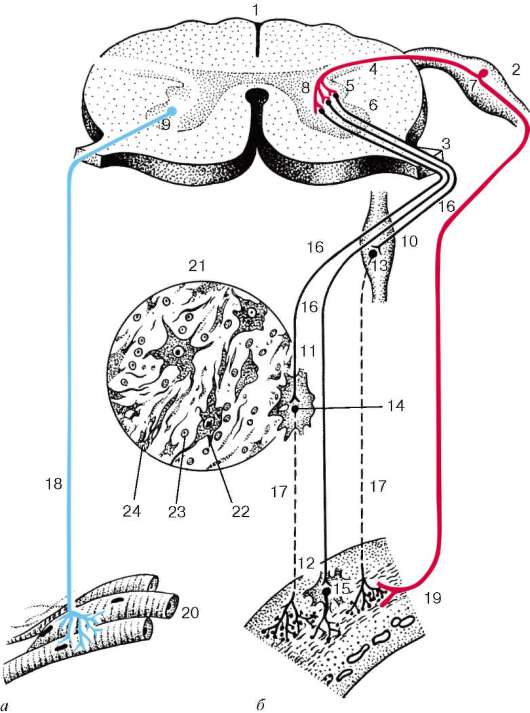
Рис. 11.7. Рефлекторные дуги соматической (а) и автономной (б) нервной системы (схема по В. Г. Елисееву, Ю. И. Афанасьеву, Е. Ф. Котовскому):
1 - спинной мозг; 2 - спинномозговой узел; 3 - передний корешок; 4 - задний рог; 5 - боковой рог; 6 - передний рог; 7 - чувствительный (афферентный) нейрон соматической и симпатической нервной системы; 8 - центральный (афферентный) нейрон автономной нервной системы; 9 - двигательный (эфферентный) нейрон передних рогов; 10 - узел симпатического ствола; 11 - узел чревного (солнечного) сплетения; 12 - интрамуральный узел (узел нервного сплетения пищевода), 13-15 - периферические (эфферентные) нейроны автономной нервной системы;
16 - преганглионарные волокна эфферентного пути автономной нервной системы;
17 - постганглионарные волокна эфферентного пути автономной нервной системы;
18 - эфферентный путь соматической нервной системы; 19 - стенка пищевода; 20 - поперечнополосатая скелетная мышечная ткань; 21 - микроскопическое строение периферических узлов автономной нервной системы; 22 - мультиполярная нервная клетка; 23 - клетка глии; 24 - нервное волокно
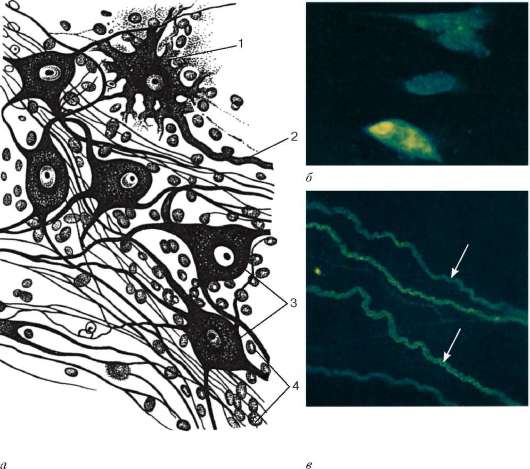
Рис. 11.8. Нейроны и нервные волокна автономной нервной системы: а - нейроны автономного узла: 1 - длинноаксонный нейрон; 2 - аксон; 3 - равно-отростчатые нейроны; 4 - ядра глиоцитов; б - адренергические нейроны (препарат В. Л. Арбузова); в - адренергические нервные волокна (препарат Т. Н. Радостиной). Метод Фалька
глиальной оболочки покрыта базальной мембраной, кнаружи от которой расположена тонкая соединительнотканная оболочка. Преганглионарные волокна, вступая в соответствующий ганглий, заканчиваются на дендри-тах или перикарионах нейронов аксодендритными либо аксосоматическими синапсами. Синапсы микроскопически выявляются в виде утолщений по ходу волокна или терминальных утолщений. Электронно-микроскопически пресинаптическая часть характеризуется типичными для холинергических синапсов прозрачными мелкими синаптическими (40-60 нм) и одиночными крупными (80-150 нм) темными пузырьками.
Цитоплазма нейронов симпатического ганглия содержит катехоламины, о чем свидетельствуют наличие мелких гранулярных пузырьков и различная степень флюоресценции на препаратах, обработанных формальдегидом по методу Фалька, их перикарионов и отростков, в том числе аксонов,
поступающих в виде постганглионарных волокон в соответствующие органы (рис. 11.8).
В составе симпатических ганглиев имеются небольшие группы гра-нулосодержащих, мелких интенсивно флюоресцирующих клеток (МИФ-клетки). Они характеризуются короткими отростками и обилием в цитоплазме гранулярных пузырьков, соответствующих по флюоресценции и электронно-микроскопической характеристике пузырькам клеток мозгового вещества надпочечника. МИФ-клетки окружены глиальной оболочкой. На телах МИФ-клеток, реже на их отростках, видны холинергические синапсы, образованные терминалями преганглионарных волокон. МИФ-клетки рассматриваются как внутриганглионарная тормозная система. Они, возбуждаясь преганглионарными холинергическими волокнами, выделяют катехоламины. Последние, распространяясь диффузно или по сосудам ганглия, оказывают тормозящее влияние на синаптическую передачу с преган-глионарных волокон на периферические нейроны ганглия.
Узлы парасимпатического отдела автономной нервной системы лежат или вблизи иннервируемого органа, или в его интрамуральных нервных сплетениях. Преганглионарные волокна заканчиваются на телах нейронов, а чаще на их дендритах холинергическими синапсами. Аксоны этих клеток (постганглионарные волокна) следуют в мышечной ткани иннервируемых органов в виде варикозных терминалей и образуют нейро-мышечные холи-нергические синапсы.
Интрамуральные сплетения. Значительное количество нейронов автономной нервной системы сосредоточено в нервных сплетениях самих иннервируемых органов: в пищеварительном тракте, сердце, мочевом пузыре и др.
Узлы интрамуральных сплетений, как и другие автономные узлы, содержат, кроме эфферентных нейронов, рецепторные и ассоциативные клетки местных рефлекторных дуг. Морфологически в интрамуральных нервных сплетениях различают три типа клеток, описанных Догелем. Длинноаксонные эфферентные нейроны (клетки I типа) имеют много коротких ветвящихся дендритов и длинный нейрит, уходящий за пределы ганглия (см. рис. 11.8, а). Равноотростчатые (афферентные) нейроны (клетки II типа) содержат несколько отростков. По морфологическим признакам нельзя определить, какой из них аксон, так как отростки, не разветвляясь, уходят далеко от тела клетки. Экспериментально установлено, что их нейриты образуют синапсы на клетках I типа. Клетки III типа (ассоциативные) посылают свои отростки в соседние ганглии, где они заканчиваются на дендритах их нейронов.
Специфическими особенностями отличается интрамуральная система пищеварительного тракта (энтеральная система). В стенке пищеварительной трубки расположены три нервных сплетения: подсерозное, мышечно-кишечное и под-слизистое, - содержащие скопления нервных клеток, связанные пучками нервных волокон (рис. 11.9). Наиболее массивное нервное сплетение - мышечно-кишечное - расположено между продольным и циркулярным мышечными слоями. Электронно-микроскопически и гистохимически в межмышечном сплетении выяв-
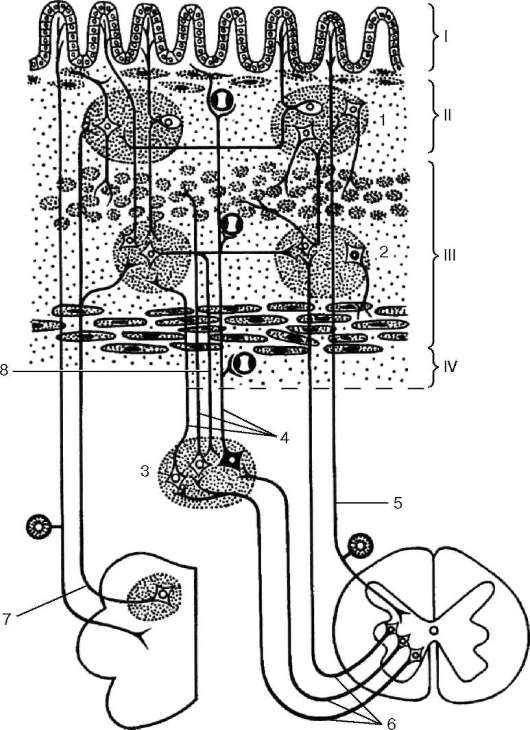
Рис. 11.9. Интрамуральные нервные сплетения пищеварительного тракта (схема по А. Д. Ноздрачеву):
I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная оболочка. 1 - подслизистое сплетение; 2 - мышечно-кишечное сплетение; 3 - симпатический ганглий; 4 - постганглионарные симпатические волокна; 5 - афферентные волокна (дендриты нейронов спинномозгового ганглия); 6 - симпатические преганглионарные волокна; 7 - афферентные волокна блуждающего нерва; 8 - афферентные волокна местных рефлекторных путей
лены холинергические нейроны, возбуждающие двигательную активность кишечника, и тормозные нейроны, представленные адренергическими и неадренер-гическими (пуринергическими) нейронами. Морфологически пуринергические
нейроны характеризуются содержанием в перикарионе и отростках крупных (размером 80-120 нм) электронно-плотных гранул. В составе интрамуральных вегетативных ганглиев содержатся и пептидергические нейроны, выделяющие ряд гормонов (вазоинтестинальный пептид, вещество Р, соматостатин и др.). Считается, что эти нейроны осуществляют нервные и эндокринные функции, а также модулируют функциональную деятельность эндокринных аппаратов различных органов.
Постганглионарные волокна нейронов интрамуральных сплетений в мышечной ткани органа образуют терминальное сплетение, тонкие стволы которого содержат несколько варикозно-расширенных аксонов. Варикозные расширения (0,5-2 мкм в диаметре) содержат синаптические пузырьки и митохондрии. Межварикозные участки (шириной 0,1-0,5 мкм) заполнены нейротрубочками и нейрофиламента-ми. Синаптические пузырьки холинергических нейро-мышечных синапсов мелкие светлые (размером 30-60 нм), адренергических - мелкие гранулярные (размером
50-60 нм).
11.5. ОБОЛОЧКИ ГОЛОВНОГО И СПИННОГО МОЗГА
Головной и спинной мозг покрыт тремя оболочками: мягкой, непосредственно прилегающей к тканям мозга, паутинной и твердой, которая граничит с костной тканью черепа и позвоночника.
Мягкая мозговая оболочка непосредственно прилежит к ткани мозга и отграничена от нее краевой глиальной мембраной. В рыхлой соединительной ткани оболочки имеются большое количество кровеносных сосудов, питающих мозг, многочисленные нервные волокна, концевые аппараты и одиночные нервные клетки.
Паутинная оболочка представлена тонким слоем рыхлой волокнистой соединительной ткани. Между ней и мягкой мозговой оболочкой находится сеть перекладин, состоящих из тонких пучков коллагеновых и тонких эластических волокон. Эта сеть связывает оболочки между собой. Между мягкой мозговой оболочкой, повторяющей рельеф поверхности мозга, и паутинной, проходящей по возвышенным участкам, не заходя в углубления, располагается подпаутинное (субарахноидальное) пространство, пронизанное тонкими коллагеновыми и эластическими волокнами, связывающими оболочки между собой. Субарахноидальное пространство сообщается с желудочками мозга и содержит цереброспинальную жидкость.
Твердая мозговая оболочка образована плотной волокнистой соединительной тканью, содержащей много эластических волокон. В полости черепа она плотно сращена с надкостницей. В спинномозговом канале твердая мозговая оболочка отграничена от периоста позвонков эпидуральным пространством, заполненным слоем рыхлой волокнистой соединительной ткани, что обеспечивает ей некоторую подвижность. Между твердой мозговой и паутинной оболочками располагается субдуральное пространство. В субдураль-ном пространстве содержится небольшое количество жидкости. Оболочки со стороны субдурального и субарахноидального пространства покрыты слоем плоских клеток глиальной природы.
11.6. ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ НЕРВНОЙ СИСТЕМЫ
Изменения в центральной нервной системе в раннем постнатальном онтогенезе связаны с ее созреванием. У новорожденных для корковых нейронов очень характерно высокое ядерно-цитоплазматическое отношение. С возрастом размеры нейронов увеличиваются за счет увеличения объема цитоплазмы. При этом наиболее быстро (в первые 3 мес жизни) увеличиваются размеры пирамидных нейронов II и IV слоев. Более медленное увеличение характерно для клеток-зерен и малых пирамидных нейронов IV слоя. Увеличивается число синаптических контактов.
Возрастные изменения коры большого мозга. В процессе развития коры большого мозга человека в онтогенезе отмечаются изменения в распределении и структуре основных ее компонентов - нейронов и глиоцитов, а также кровеносных сосудов. Уже к моменту рождения представлены ней-роглиососудистые ансамбли коры полушарий большого мозга. Однако большинство нейронов имеют неопределенную форму со слабовыражен-ными отростками и небольшие размеры. Группировки нейронов, особенно «гнездного типа», как и локальные волокнистые сети, выражены крайне слабо. Глиальные клетки мелкие. Кровеносные сосуды мягкой оболочки тонкие, капиллярная сеть редкая, одинаковая по плотности во всех слоях коры (рис. 11.10).
На первом году жизни наблюдаются типизация формы пирамидных и звездчатых нейронов, их увеличение, развитие дендритных и аксонных арборизаций, внутриансамблевых связей по вертикали. К 3 годам в ансамблях выявляются «гнездные» группировки нейронов, более четко сформированные вертикальные дендритные пучки и пучки радиальных волокон. Увеличиваются веретенообразные звездчатые нейроны, распределяющие свои аксонные коллатерали в вертикальном направлении. К 5-6 годам нарастает полиморфизм нейронов, отражающий их функциональную специализацию; усложняется система внутриансамблевых связей по горизонтали за счет роста в длину и разветвлений боковых и базальных дендритов пирамидных нейронов и развития боковых терминалей их апикальных ден-дритов. К 9-10 годам увеличиваются клеточные группировки, значительно усложняется структура короткоаксонных нейронов и расширяется сеть аксонных коллатералей всех форм интернейронов, образующих в ансамблях различных корковых зон отчетливо структурированные вертикальные колонки. К 12-14 годам в ансамблях четко обозначаются специализированные формы пирамидных нейронов, все типы интернейронов достигают высокого уровня дифференцировки; во всех ансамблях удельный объем волокон значительно выше удельного объема клеточных элементов; значительно увеличиваются диаметр и толщина стенок внутрикорковых артерий. К 18 годам ансамблевая организация коры по основным параметрам своей архитектоники достигает уровня таковой у взрослых.
У взрослых людей по сравнению с новорожденными уменьшается число нейронов в единице объема коры. Уменьшение зависит от гибели части
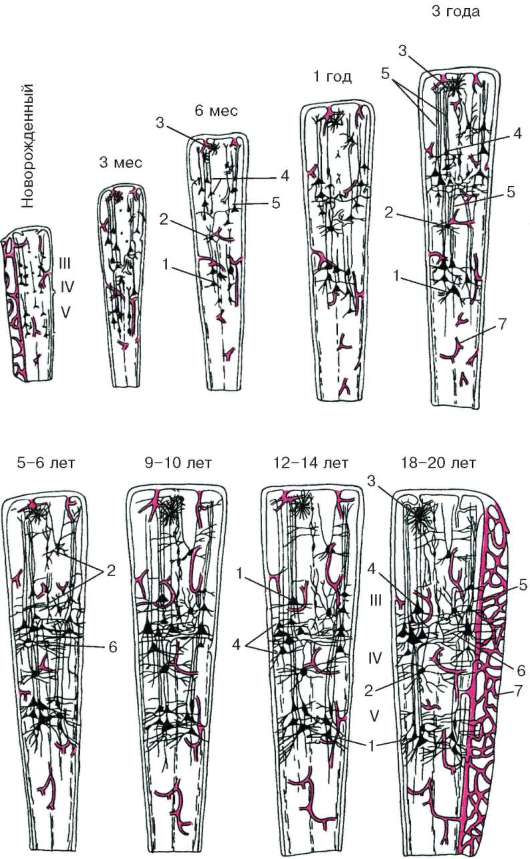
Рис. 11.10. Возрастные изменения коры большого мозга человека (нейроглиососуди-стые ансамбли) (по Л. К. Семеновой, В. А. Васильевой, Т. А. Цехмистренко): III, IV, V - пластинки (слои) коры большого мозга. 1 - группы нейронов; 2 - глио-циты; 3 - звездчатые клетки; 4 - пирамидные нейроны; 5 - дендритные и аксонные арборизации; 6 - вертикальные дендритные пучки и пучки радиальных волокон; 7 - кровеносные сосуды
нейронов, но главным образом от разрастания нервных волокон и нейро-глии, что приводит к увеличению толщины коры и механическому «раздвиганию» нейронов. У новорожденных в нейронах средней лобной извилины отсутствует хроматофильное вещество, его количество в нейронах увеличивается у ребенка к 3-6 мес, а в двухлетнем возрасте становится таким же, как и у взрослых. Формирование миелиновых оболочек вокруг аксонов в ряде областей коры (средняя и нижняя лобные извилины, средняя и нижняя височные извилины и др.) происходит после рождения ребенка.
Изменения в центральной нервной системе в старческом возрасте связаны прежде всего со склеротическими изменениями сосудов мозга. В старости мягкая и паутинная оболочки мозга утолщаются. В них могут появиться отложения извести. Наблюдается атрофия коры большого мозга, прежде всего лобной и теменной долей. Уменьшается число нейронов в единице объема коры, зависит это главным образом от гибели клеток. Нейроны уменьшаются в размере, частично теряют базофильное вещество, ядра уплотняются, их контур становится неровным. Быстрее других изменяются пирамидные нейроны V слоя двигательной зоны коры и грушевидные клетки коры мозжечка. В нейронах различных отделов нервной системы накапливаются гранулы липофусцина.
11.7. КРОВОСНАБЖЕНИЕ ЦЕНТРАЛЬНОЙ
НЕРВНОЙ СИСТЕМЫ
Кровоснабжение спинного мозга осуществляется через передние и задние корешковые артерии, входящие в него с передними и задними корешками и образующие артериальную сеть в мягкой мозговой оболочке. Здесь формируются продольные артерии, из которых главная - передняя спинальная артерия, проходящая в передней срединной щели.
Капиллярная сеть в сером веществе более густая, чем в белом. Вены спинного мозга не сопровождают артерии. Мелкие вены, идущие с периферии спинного мозга и из передней срединной щели, образуют сплетение в мягкой мозговой оболочке, особенно густое на дорсальной поверхности спинного мозга, откуда кровь оттекает в вены, сопровождающие вентральные и дорсальные корешки.
Артериальное кровоснабжение головного мозга осуществляется внутренними сонными и позвоночными артериями, которые сливаются в основании мозга в базилярную артерию. Ветви этих артерий проходят в мягкую мозговую оболочку, и отсюда мелкие веточки следуют в вещество мозга. Капиллярная сеть в сером веществе головного мозга также более густая, чем в белом. Капилляры мозга имеют непрерывную эндотелиальную выстилку и хорошо развитую базальную мембрану. Здесь происходит избирательный обмен веществ между нервной тканью и кровью, в котором принимает участие так называемый гематоэнцефалический барьер. Избирательность обмена веществ между тканью и кровью обеспечивается, помимо морфологиче-
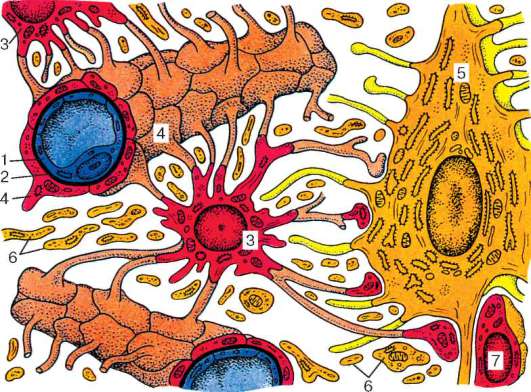
Рис. 11.11. Строение гематоэнцефалического барьера (схема):
1 - эндотелий гемокапилляра; 2 - базальная мембрана; 3 - тело астроцита; 4 - пластинчатые окончания отростков астроцитов; 5 - нейрон; 6 - отростки нейронов (нейропиль); 7 - олигодендроглиоцит
ских особенностей самих капилляров (сплошная эндотелиальная выстилка с хорошо развитыми десмосомами, плотная базальная мембрана), также и тем, что отростки глиоцитов, прежде всего астроцитов, образуют на поверхности капилляров слой, отграничивающий нейроны от непосредственного соприкосновения с сосудистой стенкой (рис. 11.11).
Контрольные вопросы
1. Какие виды нервных волокон входят в состав нерва?
2. Кора большого мозга: фило- и онтогенетические этапы развития.
3. Модульный принцип организации коры большого мозга как основа деятельности центральной нервной системы человека.
4. Развитие, строение коры и ядер мозжечка. Характеристика нервных связей в коре мозжечка.
5. Рефлекторные дуги (соматическая и вегетативные). Нейронный состав, топография.