Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 13. СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Сердечно-сосудистая система включает органы (сердце, кровеносные и лимфатические сосуды), которые обеспечивают распространение по организму крови и лимфы, содержащих питательные и биологически активные вещества, газы, продукты метаболизма.
13.1. КРОВЕНОСНЫЕ СОСУДЫ
Кровеносные сосуды представляют собой систему замкнутых трубок различного диаметра, осуществляющих транспортную функцию, регуляцию кровоснабжения органов и обмен веществ между кровью и окружающими тканями.
Развитие. Первые кровеносные сосуды появляются в мезенхиме стенки желточного мешка на 2-3-й нед эмбриогенеза человека, а также в стенке хориона в составе так называемых кровяных островков. Клетки с ангиобластическими потенциями, расположенные по периферии островков, теряют связь с клетками, расположенными в центральной части, уплощаются и дифференцируются в эндотелиальные клетки первичных кровеносных сосудов (рис. 13.1). Клетки центральной части островка округляются и дифференцируются в клетки крови. Из мезенхимных клеток, окружающих сосуд, позднее возникают гладкие мышечные клетки, перициты и адвентициальные клетки стенки сосуда, а также фибробласты.
В теле зародыша из мезенхимы образуются первичные кровеносные сосуды, имеющие вид трубочек и щелевидных пространств. В конце 3-й нед внутриутробного развития сосуды тела зародыша начинают сообщаться с сосудами внезародышевых органов.
Дальнейшее развитие стенки сосудов происходит после начала циркуляции крови под влиянием тех гемодинамических условий (кровяное давление, скорость кровотока), которые создаются в различных частях тела, что обусловливает появление специфических особенностей строения стен-
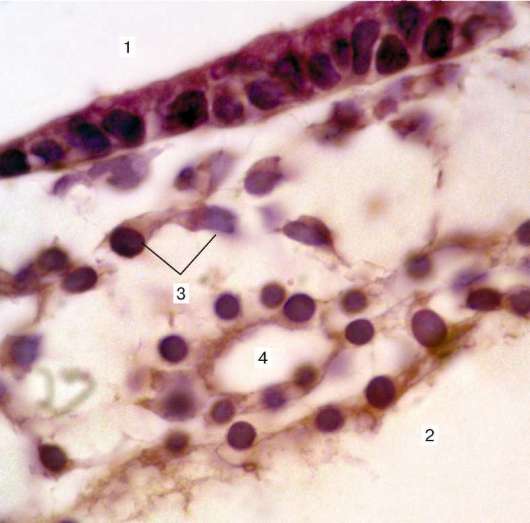
Рис. 13.1. Закладка первичных кровеносных сосудов у 17-дневного зародыша человека (зародыш «Крым»). Микрофотография (по Н. П. Барсукову):
1 - полость амниотического пузырька; 2 - полость желточного мешка; 3 - внезаро-дышевая мезенхима; 4 - первичные кровеносные сосуды
ки внутриорганных и внеорганных сосудов. В ходе перестроек первичных сосудов в эмбриогенезе часть из них редуцируется.
Классификация и общая характеристика сосудов. В кровеносной системе различают артерии, артериолы, капилляры, венулы, вены и артериоловенулярные анастомозы. Взаимосвязь между артериями и венами осуществляется системой сосудов микроциркуляторного русла.
По артериям кровь течет от сердца к органам. Как правило, эта кровь насыщена кислородом, за исключением легочной артерии, несущей венозную кровь. По венам кровь притекает к сердцу и содержит в отличие от крови легочных вен мало кислорода. Капилляры соединяют артериальное звено кровеносной системы с венозным, кроме так называемых чудесных сетей (rete mirabile), в которых капилляры находятся между двумя одноименными сосудами (например, между артериями в клубочках почки). Стенка всех артерий, так же как и вен, состоит из трех оболочек: внутренней (tunica intima, или interna), средней (tunica media) и наружной (tunica adventitia, или externa). Их толщина, тканевый состав и функциональные особенности неодинаковы в сосудах разных типов.
13.1.1. Артерии
Классификация. По особенностям строения их стенки артерии бывают трех типов: эластического, мышечного и смешанного (мышечно-эластического). Классификация основывается на соотношении количества мышечных клеток и эластических элементов в средней оболочке артерий.
Артерии эластического типа
Артерии эластического типа (arteriae elastotypica) характеризуются сильно выраженным развитием в их средней оболочке эластических структур (мембраны, волокна). К ним относятся сосуды крупного калибра, такие как аорта и легочная артерия, в которых кровь протекает под высоким давлением (120-130 мм рт. ст.) и с большой скоростью (0,5-1,3 м/с). В эти сосуды кровь поступает либо непосредственно из сердца, либо вблизи от него из дуги аорты. Артерии крупного калибра выполняют главным образом транспортную функцию. Наличие большого количества эластических элементов (волокон, мембран) позволяет этим сосудам растягиваться при систоле сердца и возвращаться в исходное положение во время диастолы. В качестве примера сосуда эластического типа рассматривается строение аорты (рис. 13.2).
Внутренняя оболочка аорты включает эндотелий (endothelium), субэндоте-лиальный слой (stratum subendotheliale) и сплетение эластических волокон ( plexus fibroelasticus).
Эндотелий аорты человека состоит из клеток, различных по форме и размерам, расположенных на базальной мембране. По протяженности сосуда размеры и форма клеток неодинаковы. Иногда клетки достигают 500 мкм в длину и 150 мкм в ширину. Чаще они бывают одноядерными, но встречаются и многоядерные. Размеры ядер также неодинаковы. В эндотелиаль-ных клетках слабо развита эндоплазматическая сеть гранулярного типа. Митохондрии весьма многочисленны (от 200 до 700), разнообразны по форме и величине, очень много микрофиламентов, образующих цитоскелет (см. главу 4).
Субэндотелиальный слой составляет примерно 15-20 % толщины стенки сосуда и состоит из рыхлой тонкофибриллярной соединительной ткани, богатой клетками звездчатой формы. В последних обнаруживается большое количество пиноцитозных пузырьков и микрофиламентов, а также эндоплазматическая сеть гранулярного типа. Эти клетки, как консоли, поддерживают эндотелий. В субэндотелиальном слое встречаются отдельные продольно направленные гладкие мышечные клетки (гладкие миоциты).
Глубже субэндотелиального слоя в составе внутренней оболочки расположено густое сплетение эластических волокон, соответствующее внутренней эластической мембране. В межклеточном веществе внутренней оболочки аорты содержится большое количество гликозаминогликанов, фосфолипиды. Основное аморфное вещество играет большую роль в трофике стенки сосуда. Физико-химическое состояние этого вещества обусловливает степень проницаемости стенки сосуда. У людей среднего и пожилого
возраста в межклеточном веществе обнаруживаются холестерин и жирные кислоты.
Внутренняя оболочка аорты в месте отхождения от сердца образует три карманоподобные створки («полулунные клапаны»).
Средняя оболочка аорты состоит из большого количества (50-70) эластических окончатых мембран (mem-branae elasticae fenestratae), связанных между собой эластическими волокнами и образующих единый эластический каркас вместе с эластическими элементами других оболочек (см. рис. 13.2).
При сканирующей электронной микроскопии выявляются три типа эластических мембран: гомогенные, волокнистые и смешанные. У человека в средней оболочке встречаются гомогенные и смешанные мембраны, состоящие из гомогенного слоя и одного или двух волокнистых слоев. Эластические волокна, с одной стороны, вплетаются в окончатые эластические мембраны, а с другой - контактируют с гладкими миоцитами, образуя вокруг них своеобразный чехлик из продольно расположенных эластических волокон. Коллагеновые волокна межмембранного пространства принимают участие в соединении соседних окончатых эластических мембран.
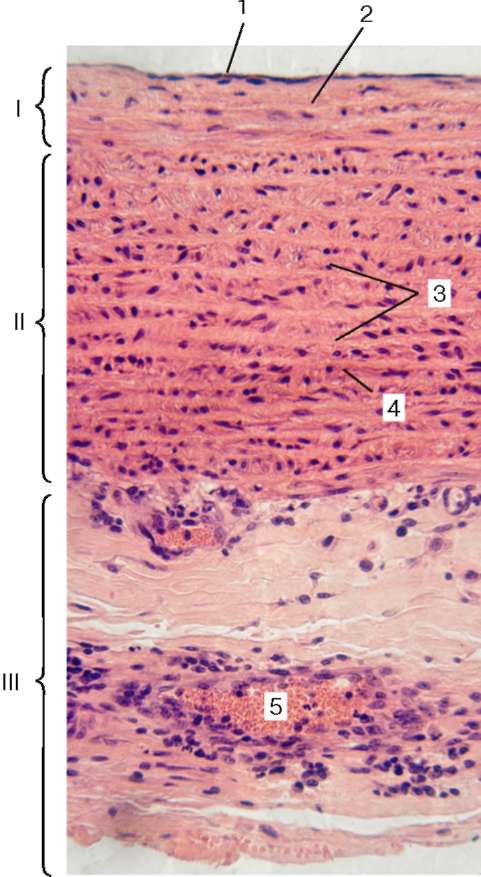
Рис. 13.2. Аорта человека; поперечный срез (микрофотография): I - внутренняя оболочка; II - средняя оболочка; III - наружная оболочка. 1 - эндотелий; 2 - субэндотелиальный слой; 3 - окончатые эластические мембраны; 4 - гладкие миоциты; 5 - сосуды сосудов
Между мембранами средней оболочки артерии эластического типа залегают гладкие мышечные клетки, косо расположенные по отношению к мембранам.
Одной из особенностей структурной организации гладких миоцитов аорты является наличие в их цитоплазме многочисленных промежуточных филаментов, состоящих из белка виментина, в то время как промежуточные филаменты гладких миоцитов других сосудов, способных более сильно сокращаться, состоят из вимен-тина и десмина. Помимо сократительной функции, гладкие миоциты выполняют секреторную функцию - синтезируют гликозаминогликаны, коллаген и эластин.
Окончатые эластические мембраны, эластические и коллагеновые волокна и гладкие миоциты погружены в аморфное вещество, богатое глико-
заминогликанами. Такое строение средней оболочки делает аорту высокоэластичной и смягчает толчки крови, выбрасываемой в сосуд во время сокращения левого желудочка сердца, а также обеспечивает поддержание тонуса сосудистой стенки во время диастолы.
Наружная оболочка аорты построена из рыхлой волокнистой соединительной ткани с большим количеством толстых эластических и колла-геновых волокон, имеющих главным образом продольное направление. В наружной оболочке проходят питающие сосуды (vasa vasorum) и нервные стволики (nervi vasorum). Наружная оболочка предохраняет сосуд от перерастяжения и разрывов.
Артерии мышечного типа
К артериям мышечного типа (аа. myotypicae) относятся преимущественно сосуды среднего и мелкого калибра, т. е. большинство артерий организма (артерии тела, конечностей и внутренних органов).
В стенках этих артерий имеется относительно большое количество гладких мышечных клеток, что обеспечивает дополнительную нагнетающую силу и регулирует приток крови к органам (рис. 13.3; 13.4, б).
В состав внутренней оболочки входят эндотелий с базальной мембраной, субэн-дотелиальный слой и внутренняя эластическая мембрана. Эндотелиальные клетки, расположенные на базальной мембране, вытянуты вдоль продольной оси сосуда. Субэндотелиальный слой состоит из тонких эластических и кол-лагеновых волокон, преимущественно продольно направленных, а также мало-дифференцированных соединительнотканных клеток.
Во внутренней оболочке некоторых артерий - сердца, почек, яичников, матки, пупочной артерии, легких - обнаруживаются продольно расположенные гладкие миоциты.
В основном веществе субэндотелиального слоя находятся гликозами-ногликаны. Субэндотелиальный слой лучше развит в артериях среднего и крупного калибра и слабее - в мелких артериях. Кнаружи от субэндоте-лиального слоя расположена тесно связанная с ним внутренняя эластическая мембрана (membrana elastica interna). В мелких артериях она очень тонкая. В более крупных артериях мышечного типа эластическая мембрана отчетливо выражена (на гистологических препаратах она имеет вид извитой блестящей эластической пластинки).
Средняя оболочка артерии содержит гладкие мышечные клетки, расположенные по пологой спирали, между которыми находятся в небольшом числе соединительнотканные клетки и волокна (коллагеновые и эластические). Коллагеновые волокна образуют опорный каркас для гладких миоцитов. В артериях обнаружен коллаген I, II, IV, V типа. Спиральное расположение мышечных клеток обеспечивает при сокращении уменьшение объема сосуда и проталкивание крови.
Эластические волокна стенки артерии на границе с наружной и внутренней оболочками сливаются с эластическими мембранами. Таким образом, создается единый эластический каркас, который, с одной стороны, придает сосуду эластичность при растяжении, а с другой - упругость при сдавлении
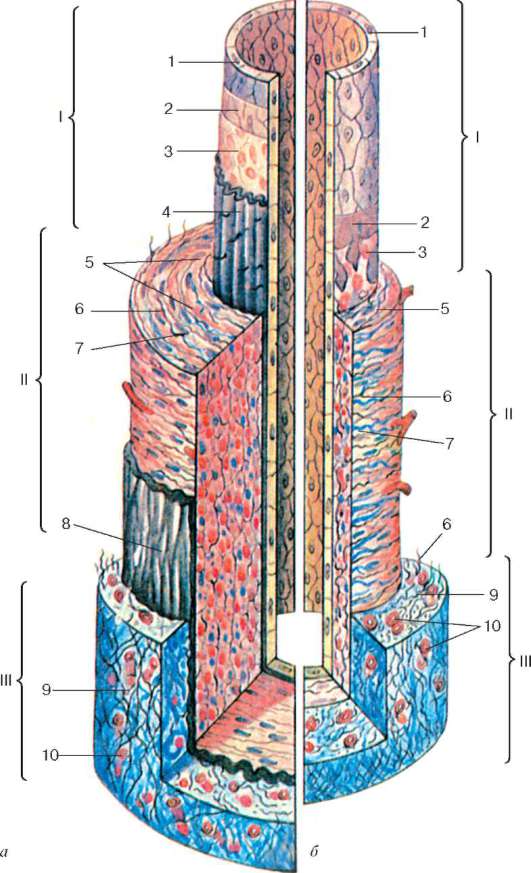
Рис. 13.3. Строение стенки артерии и вены среднего калибра (по Ю. И. Афанасьеву): а - артерия; б - вена; I - внутренняя оболочка: 1 - эндотелий; 2 - базальная мембрана; 3 - субэндотелиальный слой; 4 - внутренняя эластическая мембрана; II - средняя оболочка: 5 - гладкие миоциты; 6 - эластические волокна; 7 - коллагеновые волокна; III - наружная оболочка: 8 - наружная эластическая мембрана; 9 - волокнистая соединительная ткань; 10 - сосуды сосудов
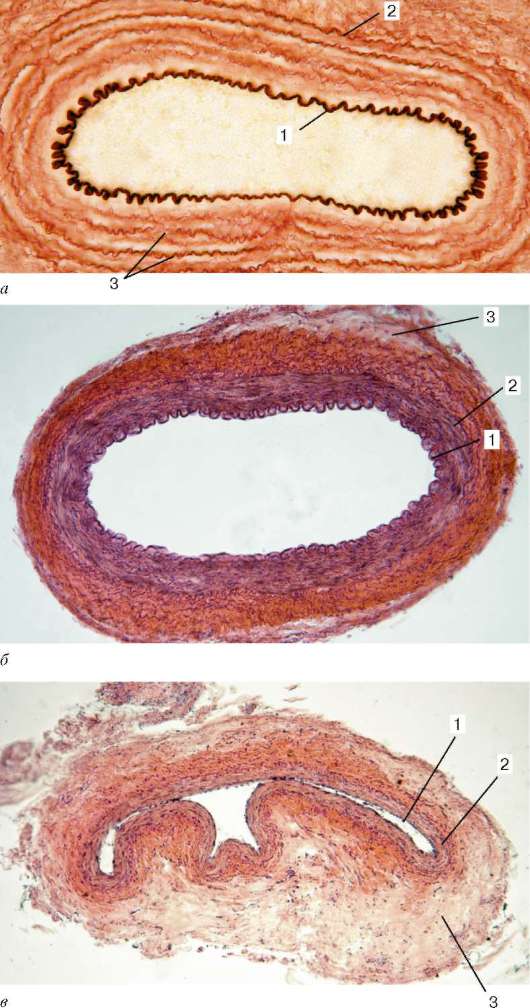
Рис. 13.4. Артерии и вены мышечного типа. Микрофотографии: а - эластический каркас артерии (поперечный срез, окраска - орсеин): 1 - внутренняя эластическая мембрана; 2 - наружная эластическая мембрана; 3 - эластические волокна; б - артерия и вена (поперечный срез): 1 - внутренняя оболочка; 2 - средняя оболочка; 3 - наружная оболочка; артерия зияет; вена в спавшемся состоянии; в - артерия и вена (поперечный срез): 1 - внутренняя оболочка; 2 - средняя оболочка; 3 - наружная оболочка; артерия зияет; вена в спавшемся состоянии
(см. рис. 13.3). Эластический каркас препятствует спадению артерий, что обусловливает их постоянное зияние и непрерывность в них тока крови (см. рис. 13.4).
Гладкие мышечные клетки средней оболочки артерий мышечного типа своими сокращениями поддерживают кровяное давление, регулируют приток крови в сосуды микроциркуляторного русла органов. На границе между средней и наружной оболочками располагается наружная эластическая мембрана (membrana elastica externa). Она состоит из продольно идущих толстых, густо переплетающихся эластических волокон, которые иногда приобретают вид сплошной эластической пластинки. Обычно наружная эластическая мембрана бывает тоньше внутренней и не у всех артерий достаточно хорошо выражена.
Наружная оболочка состоит из рыхлой волокнистой соединительной ткани, в которой соединительнотканные волокна имеют преимущественно косое и продольное направление. В этой оболочке постоянно встречаются нервы, кровеносные сосуды, питающие стенку, а также тучные клетки. Последние участвуют в регуляции местного кровотока.
По мере уменьшения диаметра артерии и их приближения к артериолам все оболочки артерии истончаются. Во внутренней оболочке резко уменьшается толщина субэндотелиального слоя и внутренней эластической мембраны. Количество мышечных клеток и эластических волокон в средней оболочке также постепенно убывает. В наружной оболочке уменьшается количество эластических волокон, исчезает наружная эластическая мембрана.
Артерии мышечно-эластического типа
По строению и функциональным особенностям артерии мышечно-эластического, или смешанного, типа (аа. mixtotypicae) занимают промежуточное положение между сосудами мышечного и эластического типов. К ним относятся, в частности, сонная и подключичная артерии. Внутренняя оболочка этих сосудов состоит из эндотелия, расположенного на базальной мембране, субэндотелиального слоя и внутренней эластической мембраны. Эта мембрана располагается на границе внутренней и средней оболочек и характеризуется отчетливой выраженностью и четкой отграниченностью от других элементов сосудистой стенки.
Средняя оболочка артерий смешанного типа состоит из примерно равного количества гладких мышечных клеток, спирально ориентированных эластических волокон и окончатых эластических мембран. Между гладкими мышечными клетками и эластическими элементами обнаруживается небольшое количество фибробластов и коллагеновых волокон (см. рис. 13.4, а).
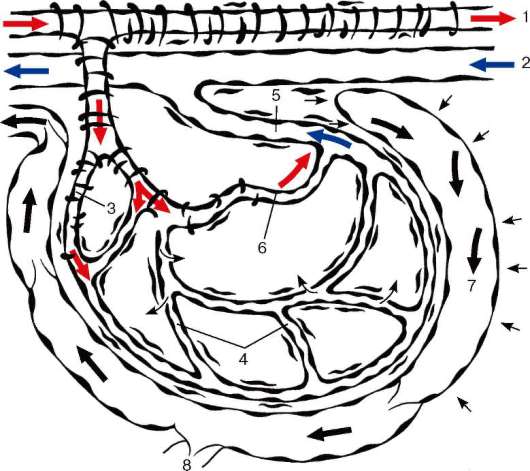
Рис. 13.5. Сосуды микроциркуляторного русла (схема по Ю. И. Афанасьеву): 1 - артерия; 2 - вена; 3 - артериолы; 4 - капилляры; 5 - венулы; 6 - артериоло-венулярный анастомоз; 7 - лимфатический капилляр; 8 - лимфатические сосуды. Толстыми стрелками обозначены направления движения крови и лимфы; тонкими стрелками - транскапиллярный обмен
В наружной оболочке артерий можно выделить два слоя: внутренний, содержащий отдельные пучки гладких мышечных клеток, и наружный, состоящий преимущественно из продольно и косо расположенных пучков коллагеновых и эластических волокон и соединительнотканных клеток. В ее составе присутствуют сосуды сосудов и нервные волокна. Занимая промежуточное положение между сосудами мышечного и эластического типов, артерии смешанного типа (например, подключичные) не только могут сильно сокращаться, но и обладают высокими эластическими свойствами, что особенно отчетливо проявляется при повышении кровяного давления.
13.1.2. Микроциркуляторное русло
Этим термином в ангиологии обозначается система мелких сосудов, включающая артериолы, капилляры, венулы, а также артериоловенулярные анастомозы. Этот функциональный комплекс кровеносных сосудов, окруженный лимфатическими капиллярами и лимфатическими сосудами, вместе с окружающей соединительной тканью обеспечивает регуляцию крове-
наполнения органов, транскапиллярный обмен и дренажно-депонирующую функцию (рис. 13.5). Чаще всего элементы микроциркуляторного русла образуют густую систему анастомозов прекапиллярных, капиллярных и посткапиллярных сосудов, но могут быть и другие варианты с выделением какого-либо основного, предпочтительного канала, например анастомоза прекапиллярной артериолы и посткапиллярной венулы и др. В каждом органе адекватно его функции существуют специфические особенности конфигурации, диаметра и плотности расположения сосудов микроцирку-ляторного русла.
Сосуды микроциркуляторного русла пластичны при изменении кровотока. Они могут депонировать форменные элементы или быть спазмиро-ваны и пропускать лишь плазму, изменять проницаемость для тканевой жидкости.
Артериолы
Это наиболее мелкие артериальные сосуды мышечного типа диаметром не более 50-100 мкм, которые, с одной стороны, связаны с артериями, а с другой - постепенно переходят в капилляры (рис. 13.6). В артериолах сохраняются три оболочки, характерные для артерий вообще, однако выражены они очень слабо.
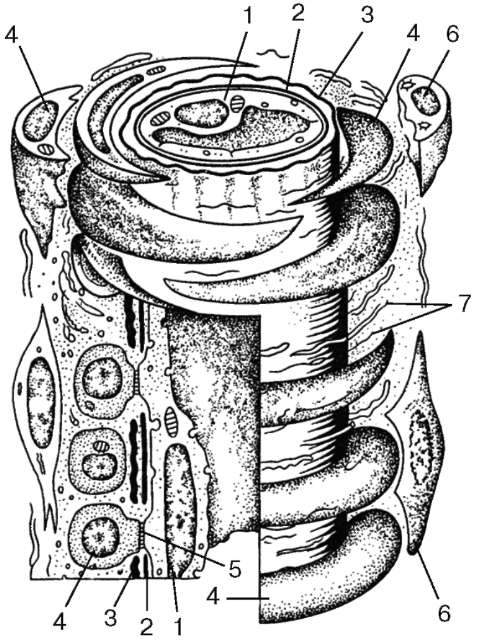
Рис. 13.6. Строение артериолы (схема по Ю. И. Афанасьеву):
1 - эндотелиальная клетка; 2 - базальная мембрана; 3 - внутренняя эластическая мембрана; 4 - гладкая мышечная клетка; 5 - контакт гладкого миоцита с эндоте-лиоцитом; 6 - адвентициальные клетки; 7 - соединительнотканные волокна
Внутренняя оболочка этих сосудов состоит из эндотелиальных клеток с базальной мембраной, тонкого субэндотелиального слоя и тонкой внутренней эластической мембраны. Средняя оболочка образована 1- 2 слоями гладких мышечных клеток, имеющих спиралевидное направление. В прекапиллярных артериолах (прекапиллярах) гладкие мышечные клетки располагаются поодиночке. Расстояние между ними увеличивается в дистальных отделах, однако они обязательно присутствуют в месте отхождения прекапилля-ров от артериолы и в месте разделения прекапилляра на капилляры. В артериолах обнаруживаются перфорации в базальной мембране эндотелия и внутренней эластической мембране, благодаря которым осуществляется непосредственный тесный контакт эндотелиоцитов и гладких мышечных клеток (см. рис. 13.6). Такие контакты создают
условия для передачи информации от эндотелия гладким мышечным клеткам. В частности, при выбросе в кровь адреналина надпочечников эндотелий синтезирует фактор, который вызывает сокращение гладких мышечных клеток. Между мышечными клетками артериол обнаруживается небольшое количество эластических волокон. Наружная эластическая мембрана отсутствует. Наружная оболочка представлена рыхлой волокнистой соединительной тканью.
В функциональном отношении артериолы являются, по выражению И. М. Сеченова, «кранами сосудистой системы», которые регулируют приток крови к органам благодаря сокращению спирально направленных гладких мышечных клеток, иннервируемых эфферентными нервными волокнами. В месте отхождения капилляра от прекапиллярных артериол имеется сужение, обусловленное циркулярно расположенными гладкими мышечными клетками в устье капилляров, выполняющих роль прекапиллярных сфинктеров.
Капилляры
Кровеносные капилляры (vasae haemocapillariae) наиболее многочисленные и самые тонкие сосуды, имеющие, однако, различный просвет (рис. 13.7). Это обусловлено как органными особенностями капилляров, так и функциональным состоянием сосудистой системы.
Например, наиболее узкие капилляры (диаметром от 4,5 до 6-7 мкм) находятся в поперечнополосатых мышцах, нервах, легких и т. п., более широкие капилляры (диаметром 8-11 мкм) - в коже и слизистых оболочках. В кроветворных органах, некоторых железах внутренней секреции, печени встречаются капилляры с широким, но меняющимся на протяжении сосуда диаметром (20-30 мкм и более). Такие капилляры называются синусо-идными. Специфические вместилища крови капиллярного типа - лакуны - имеются в пещеристых телах полового члена.
В большинстве случаев капилляры формируют сеть, однако они могут образовывать петли (в сосочках кожи, ворсинках кишки, синовиальных ворсинках суставов и др.), а также клубочки (сосудистые клубочки в почке). В капиллярах, образующих петли, выделяют артериальный и венозный отделы. Ширина артериального отдела в среднем равна диаметру эритроцита, а венозного - несколько больше.
Количество капилляров в разных органах неодинаково. Например, на поперечном разрезе в мышце человека на 1 мм2 насчитывается от 1400 до 2000 капилляров, а в коже на той же площади - 40. В любой ткани в обычных физиологических условиях находится до 50 % нефункционирую-щих капилляров. Просвет их, как правило, сильно уменьшен, но полного закрытия его при этом не происходит. Для форменных элементов крови эти капилляры оказываются непроходимыми, в то же время плазма продолжает по ним циркулировать. Число капилляров в определенном органе связано с его общими морфофункциональными особенностями, а количество

Рис. 13.7. Капилляр. Микрокиносъемка (по В. И. Козлову):
1 - эндотелий; 2 - перицит; 3 - адвентициальная клетка; 4 - эритроциты в просвете капилляра. Стрелкой обозначено направление движения крови
открытых капилляров зависит от интенсивности работы органа в данный момент.
Площадь поперечного сечения среза капиллярного русла в любой области во много раз превышает площадь поперечного сечения исходной артерии.
В стенке капилляров различают три тонких слоя (как аналоги трех оболочек рассмотренных выше сосудов). Внутренний слой представлен эндоте-лиальными клетками, расположенными на базальной мембране, средний состоит из перицитов, заключенных в базальную мембрану, а наружный - из редко расположенных адвентициальных клеток и тонких коллагеновых волокон, погруженных в аморфное вещество.
Эндотелиальный слой. Внутренняя выстилка капилляра представляет собой пласт лежащих на базальной мембране вытянутых, полигональной формы эндотелиальных клеток с извилистыми границами, которые хорошо выявляются при импрегнации серебром (рис. 13.8).
Ядра эндотелиальных клеток обычно уплощенные, овальной формы. Ядросодержащие части эндотелиоцитов, как правило, выбухают в просвет капилляра, располагаясь в шахматном порядке (I тип) или напротив друг друга (II тип). Наиболее благоприятные условия кровотока в капиллярах создаются при I типе расположения ядер, который встречается чаще. При сокращении эндотелиоцитов, ядра которых располагаются друг напротив друга, может произойти закрытие просвета капилляров.
Наиболее вытянутые эндотелиоциты длиной 75-175 мкм, а наиболее короткие - длиной 5-8 мкм. Толщина эндотелиальных клеток неодинакова. В различных капиллярах она колеблется от 200 нм до 1-2 мкм на периферии и 3-5 мкм в околоядерных участках. Клетки эндотелия обычно тесно прилежат друг к другу, часто обнаруживаются плотные и щелевые контакты. Поверхность эндотелиальных клеток, обращенная к току крови, покрыта слоем гликопротеидов (параплазмолеммальный слой), с которым связаны атромбогенная и барьерная функция эндотелия, а также участие эндотелия в регуляции сосудистого тонуса. Атромбогенная функция эндотелия обусловлена не только отрицательным зарядом гликокаликса, но также и способностью эндотелия синтезировать вещества, обладающие атромбо-генными свойствами, такие как простациклин, ингибирующий агрегацию тромбоцитов. Барьерная функция эндотелия связана с рецепторами, цито-
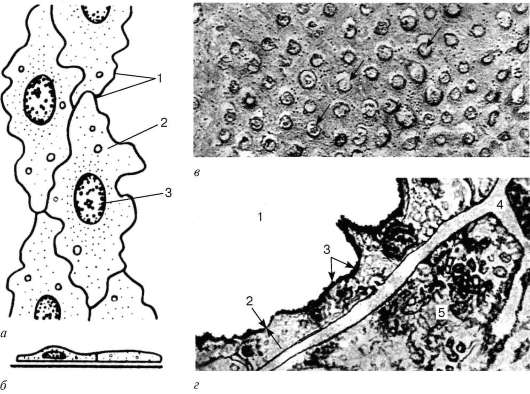
Рис. 13.8. Эндотелий капилляров:
а - плоскостное изображение; б - отвесный срез (схема по Ю. И. Афанасьеву): 1 - границы клеток; 2 - цитоплазма; 3 - ядро; в - фенестры в эндотелиоцитах пери-тубулярного капилляра почки. Электронная микрофотография, увеличение 20 000 (по А. А. Миронову); г - параплазмолеммальный слой эндотелиоцита гемокапил-ляра. Электронная микрофотография, увеличение 80 000 (по В. В. Куприянову, Я. Л. Караганову и В. И. Козлову): 1 - просвет капилляра; 2 - плазмолемма; 3 - параплазмолеммальный слой; 4 - базальная мембрана; 5 - цитоплазма перицита
скелетом эндотелиоцитов, базальной мембраной (см. ниже). Вдоль внутренней и наружной поверхностей эндотелиальных клеток располагаются пи-ноцитозные пузырьки и кавеолы, отображающие трансэндотелиальный транспорт различных веществ и метаболитов. В венозном отделе капилляра их больше, чем в артериальном. Органеллы, как правило, немногочисленны и расположены в околоядерной зоне.
Внутренняя поверхность эндотелия капилляра, обращенная к току крови, может иметь ультрамикроскопические выступы в виде отдельных микроворсинок, особенно в венозном отделе капилляра. В этих отделах капилляров цитоплазма эндотелиоцитов образует клапанообразные структуры. Эти цитоплазматические выросты увеличивают поверхность эндотелия и в зависимости от активности транспорта жидкости через эндотелий изменяют свои размеры.
Эндотелий участвует в образовании базальной мембраны. Одна из функций эндотелия - сосудообразующая (неоваскулогенез). Эндотелиальные клетки обра-
зуют между собой простые соединения, контакты типа замка и плотные контакты с локальным слиянием внешних пластинок плазмолеммы контактирующих эндо-телиоцитов и облитерацией межклеточной щели. Эндотелиоциты синтезируют и выделяют факторы, активирующие систему свертывания крови (тромбопластин, тромбоксан), и антикоагулянты (простациклин и др.). Участие эндотелия в регуляции сосудистого тонуса опосредуется также через рецепторы. При связывании вазоактивных веществ с рецепторами в эндотелиальных клетках синтезируется либо фактор расслабления, либо фактор сокращения гладких миоцитов. Эти факторы специфичны и действуют только на гладкие миоциты сосудов. Базальная мембрана эндотелия капилляров - это тонкофибриллярная, пористая, полупроницаемая пластина толщиной 30-35 нм, в состав которой входят коллаген IV и V типов, гликопротеины, а также фибронектин, ламинин и сульфатосодержащие протеогликаны. Базальная мембрана выполняет опорную, разграничительную и барьерную функции. Между эндотелиальными клетками и перицитами базальная мембрана местами истончается и прерывается, а сами клетки здесь связаны между собой посредством плотных контактов плазмолеммы. Эта область эндотелиопери-цитарных контактов служит местом передачи различных факторов от одной клетки другой.
Перициты. Эти соединительнотканные клетки имеют отростчатую форму и окружают кровеносные капилляры, располагаясь в расщеплениях базальной мембраны эндотелия. На перицитах некоторых капилляров обнаружены эфферентные нервные окончания, функциональное значение которых, по-видимому, связано с регуляцией изменения просвета капилляров.
Адвентициальные клетки. Это малодифференцированные клетки, расположенные снаружи от перицитов. Они окружены аморфным веществом соединительной ткани, в котором находятся тонкие коллагеновые волокна. Адвентициальные клетки являются камбиальными полипотентными предшественниками фибробластов, остеобластов и адипоцитов и др.
Классификация капилляров. В основу классификации капилляров положены результаты электронно-микроскопических исследований эндотелия и базальной мембраны.
Различают три типа капилляров (рис. 13.9). Наиболее распространенный тип капилляров - соматический, описанный выше (к этому типу относятся капилляры со сплошными эндотелиальной выстилкой и базальной мембраной); второй тип - фенестрированные капилляры с порами в эндотелио-цитах, затянутых диафрагмой (фенестрами), и третий тип - капилляры перфорированного типа со сквозными отверстиями в эндотелии и базальной мембране. Капилляры соматического типа находятся в сердечной и скелетной мышцах, в легких и других органах (рис. 13.10).
Фенестрированные капилляры встречаются в эндокринных органах, в собственной пластинке слизистой оболочки тонкой кишки, в бурой жировой ткани, в почке. Перфорированные капилляры характерны для органов кроветворения, в частности для селезенки, а также для печени.
Фенестры и в особенности щели облегчают проникновение различных макромолекул и корпускулярных частиц через стенку капилляров. Растяжимость эндотелия и проницаемость для коллоидных частиц в венозном отделе капилляра выше, чем в артериальном.
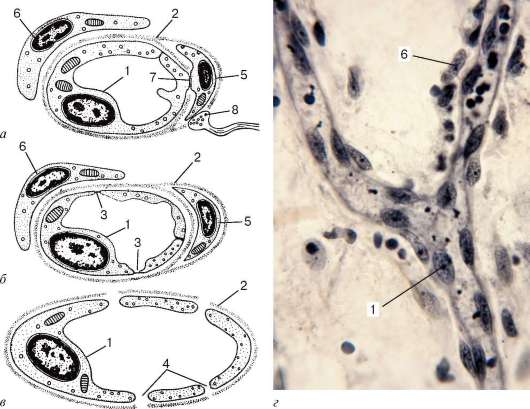
Рис. 13.9. Три типа капилляров (по Ю. И. Афанасьеву):
а - капилляр с непрерывными эндотелиальной выстилкой и базальной мембраной; б - капилляр с фенестрированным эндотелием и непрерывной базальной мембраной; в - капилляр с щелевидными отверстиями в эндотелии и прерывистой базальной мембраной. 1 - эндотелиоцит; 2 - базальная мембрана; 3 - фенестры; 4 - щели (поры); 5 - перицит; 6 - адвентициальная клетка; 7 - контакт эндотелиоцита и перицита; 8 - нервное окончание; г - капилляр, обозначения, общие с а-в (микрофотография, окраска - железный гематоксилин)
Кровеносные капилляры осуществляют основные обменные процессы между кровью и тканями, а в некоторых органах (легкие) участвуют в обеспечении газообмена между кровью и воздухом. Тонкость стенок капилляров, огромная площадь их соприкосновения с тканями (более 6000 м2), медленный кровоток (0,5 мм/с), низкое кровяное давление (20-30 мм рт. ст.) обеспечивают наилучшие условия для обменных процессов.
Стенка капилляров тесно связана функционально и морфологически с окружающей соединительной тканью (изменение состояния базальной мембраны и основного вещества соединительной ткани).
Изменение просвета капилляров при различных физиологических и патологических условиях в значительной мере зависит от давления крови в самих капиллярах, что связано с тонусом мышечных клеток артериол и мелких вен, прекапиллярных сфинктеров, а также артериоловенулярных анастомозов и состоянием перицитов.
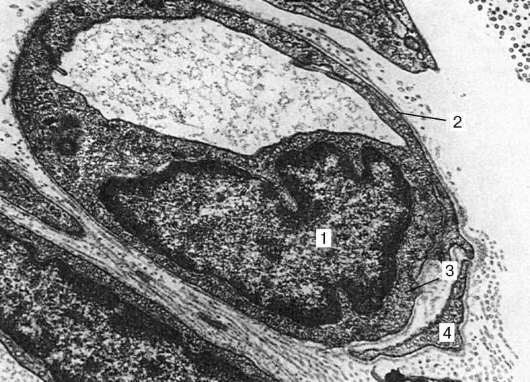
Рис. 13.10. Строение капилляра соматического типа. Электронная микрофотография, увеличение 13 000 (по Н. А. Юриной и А. И. Радостиной):
1 - ядро эндотелиоцита; 2 - базальная мембрана; 3 - везикулы в цитоплазме; 4 - перицит
Отводящий отдел микроциркуляторного русла начинается венозной частью капилляров, для которых характерны более крупные микроворсинки на люминальной поверхности эндотелия и складки, напоминающие створки клапанов, относительно большое число митохондрий и пиноци-тозных пузырьков. В эндотелии отводящего отдела чаще обнаруживаются фенестры. Диаметр венозного отдела капилляра может быть шире артериального в 1,5-2 раза.
Венулы
Различают три разновидности венул (venulae): посткапиллярные, собирательные и мышечные. Посткапиллярные венулы (диаметр 8-30 мкм) по своему строению напоминают венозный отдел капилляра, но в стенке этих венул отмечается больше перицитов, чем в капиллярах. Посткапиллярные вену-лы с высоким эндотелием служат местом выхода лимфоцитов из сосудов (в органах иммунной системы). В собирательных венулах (диаметр 30-50 мкм) появляются отдельные гладкие мышечные клетки и более отчетливо выражена наружная оболочка. Мышечные венулы (диаметр 50-100 мкм) имеют один-два слоя гладких мышечных клеток в средней оболочке и сравнительно хорошо развитую наружную оболочку.
Венозный отдел микроциркуляторного русла вместе с лимфатическими капиллярами выполняет дренажную функцию, регулируя гематолимфати-ческое равновесие между кровью и внесосудистой жидкостью, удаляя продукты метаболизма тканей. Через стенки венул, так же как через капилляры,
мигрируют лейкоциты. Медленный кровоток (не более 1-2 мм в секунду) и низкое кровяное давление (около 10 мм рт. ст.), а также растяжимость этих сосудов создают условия для депонирования крови.
Артериоловенулярные анастомозы
Артериоловенулярные анастомозы (ABA) - это соединения сосудов, несущие артериальную кровь в вены в обход капиллярного русла. Они обнаружены почти во всех органах, диаметр ABA колеблется от 30 до 500 мкм, а длина может достигать 4 мм. Объем кровотока в ABA во много раз больше, чем в капиллярах, скорость кровотока значительно увеличена. Так, если через капилляр 1 мл крови проходит в течение 6 ч, то такое же количество крови через ABA проходит за две секунды. ABA отличаются высокой реактивностью и способностью к ритмическим сокращениям с частотой до 12 раз в минуту.
Классификация (рис. 13.11). Различают две группы анастомозов: 1) истинные ABA (шунты), по которым сбрасывается чисто артериальная кровь; 2) атипичные ABA (полушунты), по которым течет смешанная кровь.
Первая группа истинных анастомозов (шунты) может иметь различную внешнюю форму - прямые короткие соустья, петли, ветвящиеся соединения. По своему строению они подразделяются на две подгруппы: а) простые ABA и б) ABA, снабженные специальными сократительными структурами.
В простых истинных анастомозах границы перехода одного сосуда в другой соответствуют участку, где заканчивается средняя оболочка артериолы. Регуляция кровотока осуществляется гладкими мышечными клетками средней оболочки самой артериолы, без специальных дополнительных сократительных аппаратов. Во второй подгруппе анастомозы могут иметь специальные сократительные устройства в виде валиков или подушек в субэндотелиальном слое, образованные продольно расположенными гладкомышечными клетками. Сокращение подушек, выступающих в просвет анастомоза, приводит к прекращению кровотока. К этой же подгруппе относятся ABA эпителиоидного типа (простые и сложные). Простые ABA эпителиоидного типа характеризуются наличием в средней оболочке внутреннего продольного и наружного циркулярного слоев гладких мышечных клеток, которые по мере приближения к венозному концу заменяются на короткие овальные светлые клетки (Е-клетки), похожие на эпителиальные. В венозном сегменте ABA стенка его резко истончается. Средняя оболочка здесь содержит лишь незначительное количество гладких мышечных клеток в виде циркулярно расположенных поясков. Наружная оболочка состоит из рыхлой соединительной ткани. Сложные, или клу-бочковые (гломерулярные), ABA эпителиоидного типа отличаются от простых тем, что приносящая (афферентная) артериола делится на 2-4 ветви, которые переходят в венозный сегмент. Эти ветви окружены одной общей соединительнотканной оболочкой. Такие анастомозы часто обнаруживаются в дерме кожи и гиподерме, а также в параганглиях.
Вторая группа - атипичные анастомозы (полушунты) - представляет собой соединения артериол и венул, по которым кровь протекает через короткий, но широкий, диаметром до 30 мкм, капилляр, поэтому сбрасываемая в венозное русло кровь является не полностью артериальной.
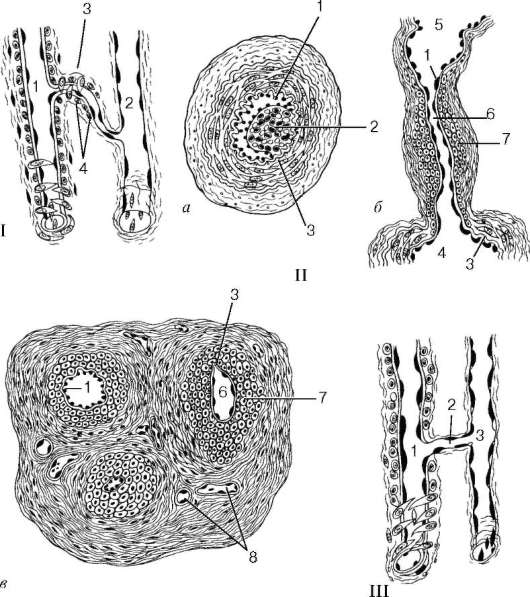
Рис. 13.11. Артериоловенулярные анастомозы (ABA) (по Ю. И. Афанасьеву): I - ABA без специального запирательного устройства: 1 - артериола; 2 - венула; 3 - анастомоз; 4 - гладкие миоциты анастомоза; II - ABA со специальным запиратель-ным устройством: а - анастомоз типа замыкающей артерии; б - простой анастомоз эпителиоидного типа; в - сложный анастомоз эпителиоидного типа (клубочковый): 1 - эндотелий; 2 - продольно расположенные пучки гладких миоцитов; 3 - внутренняя эластическая мембрана; 4 - артериола; 5 - венула; 6 - анастомоз; 7 - эпи-телиоидные клетки анастомоза; 8 - капилляры в соединительнотканной оболочке; III - атипичный анастомоз: 1 - артериола; 2 - короткий капилляр; 3 - венула
ABA, особенно клубочкового типа, богато иннервированы. ABA принимают участие в регуляции кровенаполнения органов, местного и общего давления крови, в мобилизации депонированной в венулах крови. Эти соединения играют определенную роль в стимуляции венозного кровотока, артериализации венозной крови, мобилизации депонированной крови и регуляции тока тканевой жидкости в венозное русло. Велика роль ABA в компенсаторных реакциях организма при нарушении кровообращения и развитии патологических процессов.
13.1.3. Вены
Вены большого круга кровообращения осуществляют отток крови от органов, участвуют в обменной и депонирующей функциях. Различают поверхностные и глубокие вены, причем последние в двойном количестве сопровождают артерии. Вены широко анастомозируют, образуя в органах сплетения.
Отток крови начинается по посткапиллярным венулам. Низкое кровяное давление (15-20 мм рт. ст.) и незначительная скорость (в органных венах около 10 мм/с) кровотока определяют сравнительно слабое развитие эластических элементов в стенках венах и их большую растяжимость. Количество же гладких мышечных клеток в стенке вен неодинаково и зависит от того, движется ли в них кровь к сердцу под действием силы тяжести или против нее. Необходимость преодоления силы тяжести крови в венах нижних конечностей приводит к сильному развитию гладких мышечных элементов в этих сосудах по сравнению с венами верхних конечностей, головы и шеи. Во многих венах (в подкожных и других) имеются клапаны (valvulae venosae), являющиеся производными внутренней оболочки. Вены головного мозга и его оболочек, внутренних органов, подчревные, подвздошные, полые и безымянные клапанов не имеют.
Клапаны в венах способствуют току венозной крови к сердцу, препятствуя ее обратному движению. Одновременно клапаны предохраняют сердце от излишней затраты энергии на преодоление колебательных движений крови, постоянно возникающих в венах под влиянием различных внешних воздействий (изменение атмосферного давления, мышечное сжатие и др.).
Одна из отличительных особенностей гистологической структуры вены - относительно слаборазвитый эластический каркас. Как правило, в венах отсутствуют внутренняя и наружная эластические мембраны. Эластические волокна, располагающиеся преимущественно в продольном направлении, немногочисленны. Низкое давление и слаборазвитый эластический каркас приводят к спадению стенки вен и возрастанию сопротивления току крови (см. рис. 13.4, в).
Классификация. По степени развития мышечных элементов в стенках вен они могут быть разделены на две группы: вены фиброзного (безмышечного) и вены мышечного типа. Вены мышечного типа в свою очередь подразделяются на вены со слабым, средним и сильным развитием мышечных элементов.
В венах, так же как и в артериях, различают три оболочки: внутреннюю, среднюю и наружную. Выраженность и строение этих оболочек в различных венах существенно различаются.
Вены фиброзного типа
Вены фиброзного типа (venae fibrotypicae) отличаются тонкостью стенок и отсутствием средней оболочки, в связи с чем их называют еще венами безмышечного типа. К венам этого типа относят безмышечные вены твер-
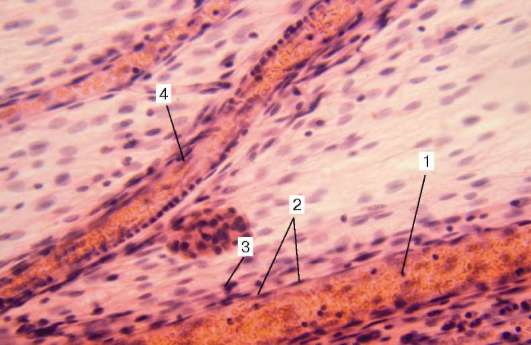
Рис. 13.12. Артериола и венула. Тотальный препарат мягкой мозговой оболочки (микрофотография):
1 - венула; 2 - эндотелий; 3 - адвентициальные клетки; 4 - артериола
дой и мягкой мозговых оболочек (рис. 13.12), вены сетчатки глаза, костей, селезенки и плаценты.
Вены мозговых оболочек и сетчатки глаза податливы при изменении кровяного давления, могут сильно растягиваться, но скопившаяся в них кровь сравнительно легко под действием собственной силы тяжести оттекает в более крупные венозные стволы. Вены костей, селезенки и плаценты также пассивны в продвижении по ним крови. Это объясняется тем, что все они сращены с плотными элементами соответствующих органов и не спадаются, поэтому отток крови по ним совершается легко. Эндотелиальные клетки, выстилающие эти вены, имеют более извилистые границы, чем в артериях. Снаружи к ним прилежит базальная мембрана, а затем тонкий слой рыхлой волокнистой соединительной ткани, срастающийся с окружающими тканями.
Вены мышечного типа
Вены мышечного типа (venae myotypicae) характеризуются наличием в их оболочках гладких мышечных клеток, количество и расположение которых в стенке вены обусловлены гемодинамическими факторами.
Выделяют вены со слабым, средним и сильным развитием мышечных элементов. Вены со слабым развитием мышечных элементов различны по диаметру. К ним относятся вены мелкого и среднего калибра (до 1-2 мм), сопровождающие артерии мышечного типа в верхней части туловища, шеи и лица, а также такие крупные, как, например, верхняя полая вена. В этих сосудах кровь в значительной мере продвигается пассивно вследствие своей тяжести. К этому же типу вен можно отнести и вены верхних конечностей. Стенки таких вен несколько тоньше соответствующих по калибру артерий,
содержат меньше мышечных элементов и на препаратах находятся обычно в спавшемся состоянии.
Вены мелкого и среднего калибра со слабым развитием мышечных элементов имеют плохо выраженный субэндотелиальный слой, а в средней оболочке содержится небольшое количество мышечных клеток. В некоторых мелких венах, например в венах пищеварительного тракта, гладкие мышечные клетки в средней оболочке образуют отдельные «пояски», располагающиеся далеко друг от друга. Благодаря такому строению вены могут сильно расширяться и выполнять депонирующую функцию. В наружной оболочке мелких вен встречаются единичные продольно направленные гладкие мышечные клетки.
Среди вен крупного калибра, в которых слабо развиты мышечные элементы, наиболее типична верхняя полая вена (рис. 13.13), в средней оболочке стенки которой отмечается небольшое количество гладких мышечных клеток. Это обусловлено отчасти прямохождением человека, в силу чего кровь по этой вене стекает к сердцу благодаря собственной тяжести, а также дыхательным движениям грудной клетки. В начале диастолы (расслабление мускулатуры) желудочков сердца в предсердиях появляется даже небольшое отрицательное кровяное давление, которое как бы подсасывает кровь из полых вен.
Примером вены среднего калибра со средним развитием мышечных элементов является плечевая вена. Эндотелиальные клетки ее внутренней оболочки короче, чем в соответствующей артерии. Субэндотелиальный слой состоит из соединительнотканных волокон и клеток, ориентированных в основном вдоль сосуда. Внутренняя оболочка этого сосуда формирует клапанный аппарат, а также имеет в своем составе отдельные продольно направленные гладкие мышечные клетки. Внутренняя эластическая мембрана в вене не выражена. На границе между внутренней и средней оболочками располагается только сеть эластических волокон. Эластические волокна внутренней оболочки плечевой вены, как и в артериях, связаны с эластическими волокнами средней и наружной оболочек и составляют единый каркас.
Средняя оболочка этой вены гораздо тоньше средней оболочки соответствующей артерии. Она обычно состоит из циркулярно расположенных пучков гладких миоцитов, разделенных прослойками волокнистой соединительной ткани. Наружная эластическая мембрана в этой вене отсутствует, поэтому соединительнотканные прослойки средней оболочки переходят непосредственно в рыхлую волокнистую соединительную ткань наружной оболочки. В плечевой вене она очень сильно развита: ее толщина в 2-3 раза превышает толщину средней оболочки. Коллагеновые и эластические волокна наружной оболочки направлены преимущественно продольно. Кроме того, в наружной оболочке встречаются отдельные гладкие мышечные клетки и небольшие их пучки, которые также расположены продольно.
К венам с сильным развитием мышечных элементов относятся крупные вены нижней половины туловища и ног. Для них характерно развитие пучков гладких мышечных клеток во всех трех их оболочках, причем во
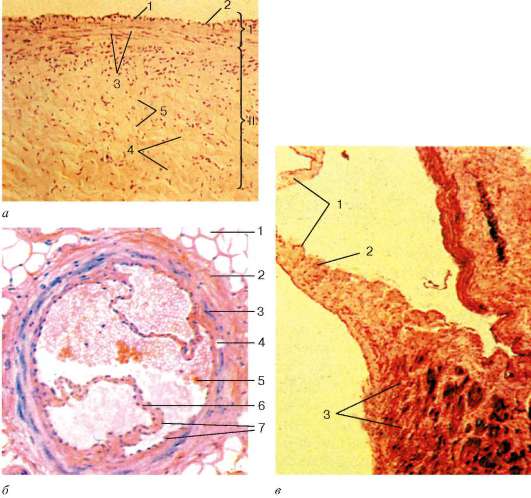
Рис. 13.13. Вены со слабым (а, б) и сильным (в) развитием мышечных элементов: а - верхняя полая вена (поперечный срез): I - внутренняя и средняя оболочки; II - наружная оболочка; 1 - просвет вены; 2 - эндотелий; 3 - гладкие миоциты; 4 - коллагеновые волокна; 5 - соединительнотканные клетки; б - поперечный срез вены в области прикрепления клапана: 1 - жировые клетки; 2 - наружная оболочка; 3 - средняя оболочка; 4 - внутренняя оболочка; 5 - кровь; 6 - створки клапана; 7 - эндотелий (по Ю. И. Афанасьеву); в - бедренная вена с клапаном (продольный срез): 1 - створка клапана; 2 - эндотелий; 3 - гладкие миоциты
внутренней и наружной оболочках они имеют продольное направление, а в средней - циркулярное.
Бедренная вена. Внутренняя оболочка ее состоит из эндотелия и субэн-дотелиального слоя, образованного рыхлой соединительной тканью, в которой продольно залегают пучки гладких мышечных клеток. Внутренняя эластическая мембрана отсутствует, однако на ее месте видны скопления эластических волокон.
Внутренняя оболочка бедренной вены формирует клапаны, представляющие собой ее тонкие складки (см. рис. 13.13, в). Эндотелиальные клетки,
покрывающие клапан со стороны, обращенной в просвет сосуда, имеют удлиненную форму и направлены вдоль створок клапана, а на противоположной стороне клапан покрыт эндотелиальными клетками полигональной формы, лежащими поперек створок. Основу клапана составляет волокнистая соединительная ткань. При этом на стороне, обращенной к просвету сосуда, под эндотелием залегают преимущественно эластические волокна, а на противоположной стороне - много коллагеновых волокон. В основании створки клапана может находиться некоторое количество гладких мышечных клеток.
Средняя оболочка бедренной вены содержит пучки циркулярно расположенных гладких мышечных клеток, окруженных коллагеновыми и эластическими волокнами. Выше основания клапана средняя оболочка истончается. Ниже места прикрепления клапана мышечные пучки перекрещиваются, создавая утолщение в стенке вены. В наружной оболочке, образованной рыхлой соединительной тканью, обнаруживаются пучки продольно расположенных гладких мышечных клеток, сосуды сосудов, нервные волокна.
Сходное строение (наличие циркулярного слоя гладких миоцитов в средней и их продольных пучков в наружной и внутренней оболочках) имеют и другие вены нижних конечностей (подколенные, большие и малые подкожные).
Нижняя полая вена также относится к венам с сильным развитием мышечных элементов (рис. 13.14). Внутренняя оболочка нижней полой вены представлена эндотелием, субэндотелиальным слоем и слоем эластических волокон. Во внутренней части средней оболочки наряду с гладкими мышечными клетками залегает подинтимальная сеть кровеносных и лимфатических капилляров, а в наружной части - артериолы и венулы. Капилляры в нижней полой вене отсутствуют.
Внутренняя и средняя оболочки нижней полой вены человека развиты относительно слабо. Во внутренней оболочке в субэндотелиальном слое находятся немногочисленные продольно расположенные гладкие мышечные клетки. В средней оболочке выявляется циркулярный мышечный слой, который в грудном участке нижней полой вены истончается. Наружная оболочка нижней полой вены имеет большое количество продольно расположенных пучков гладких мышечных клеток и по своей толщине превышает толщину внутренней и средней оболочек вместе взятых. Между пучками гладких мышечных клеток лежат прослойки рыхлой волокнистой соединительной ткани. Сокращение пучков гладких миоцитов в наружной оболочке не только способствует проталкиванию крови вверх (против силы тяжести), но и приводит к образованию поперечных складок, препятствующих обратному току крови. В устье нижней полой вены в ее наружную оболочку заходят пучки поперечнополосатых мышечных клеток миокарда. К наружной оболочке подходят сосудисто-нервные пучки (комплексы, состоящие из артерий, вен, лимфатических сосудов и нервов). В наружной оболочке из них образуются сплетения кровеносных и лимфатических сосудов (vasa vasorum и vasa lymphaticorum), многочисленные нервные волокна, а в ней также залегают пластинчатые нервные окончания (тельца Фатера-
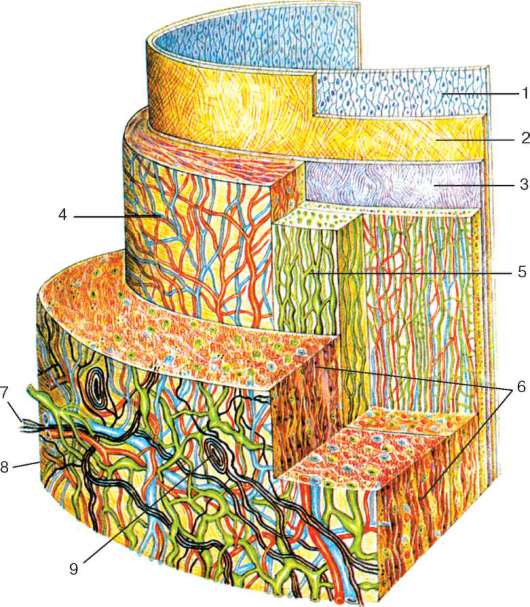
Рис. 13.14. Конструкция стенки нижней полой вены человека (схема): 1 - эндотелий; 2 - субэндотелиальный слой; 3 - слой эластических волокон внутренней оболочки; 4 - артериолы и венулы в средней оболочке; 5 - сеть лимфатических капилляров; 6 - пучки гладких мышечных клеток в наружной оболочке; 7 - сплетения кровеносных и лимфатических сосудов; 8 - нервные волокна (черные); 9 - пластинчатое нервное окончание (по В. Я. Бочарову)
Пачини). Подобную структурную организацию имеют вены брюшной полости (наружная и общая подвздошная, почечная и др.).
13.1.4. Органные особенности строения кровеносных сосудов
Некоторые отделы сосудистой системы имеют органные особенности строения артерий. Например, артерии черепа отличаются слабым развитием эластических элементов в средней и наружной оболочках; наружной эластической мембраны в них нет. Внутренняя эластическая мембрана, наоборот, выражена отчетливо. Такие же особенности существуют и у артерий головного мозга.
В пупочной артерии отсутствует внутренняя эластическая мембрана. В затылочной артерии сильно развиты пучки гладких мышечных клеток во внутренней оболочке. В почечной, брыжеечной, селезеночной и венечной артериях пучки продольно расположенных гладких мышечных клеток хорошо выражены в наружной оболочке. В артериях матки, полового члена, артериях сосочковых мышц сердца и пупочного канатика, особенно в месте его перехода в плаценту, пучки гладких мышечных клеток находятся и во внутренней, и в наружной оболочках. Некоторые вены, как и артерии, имеют ярко выраженные органные особенности строения. Так, в легочной и пупочной венах, в отличие от всех других вен, очень хорошо развит циркулярный мышечный слой в средней оболочке, вследствие чего они напоминают по своему строению артерии. Вены сердца в средней оболочке содержат продольно направленные пучки гладких мышечных клеток. В воротной вене средняя оболочка состоит из двух слоев: внутреннего - кольцевого и наружного - продольного. В некоторых венах, например сердечных, обнаруживаются эластические мембраны, которые способствуют большей упругости и эластичности этих сосудов, расположенных в постоянно сокращающемся органе. В стенках глубоких вен желудочков сердца нет ни мышечных клеток, ни эластических мембран. Эти вены построены по типу синусоидов, имеющих на дистальном конце вместо клапанов сфинктеры. Стенки вен наружной оболочки сердца содержат продольно направленные пучки гладких мышечных клеток. В надпочечниках есть вены, которые имеют продольные мышечные пучки во внутренней оболочке, выступающие в виде подушечек в просвет вены, особенно в устье. Вены печени, подслизистой основы кишечника, слизистой оболочки носа, вены полового члена и другие снабжены сфинктерами, регулирующими отток крови.
13.2. ЛИМФАТИЧЕСКИЕ СОСУДЫ
Лимфатические сосуды - часть лимфатической системы, включающей еще и лимфатические узлы. В функциональном отношении лимфатические сосуды тесно связаны с кровеносными, особенно в области расположения сосудов микроциркуляторного русла. Именно здесь происходят образование тканевой жидкости и проникновение ее в лимфатическое русло. Через мелкие лимфоносные пути осуществляются постоянная миграция лимфоцитов из кровотока и их рециркуляция из лимфатических узлов в кровь.
Классификация. Среди лимфатических сосудов различают лимфатические капилляры, интра- и экстраорганные лимфатические сосуды, отводящие лимфу от органов, и главные лимфатические стволы тела - грудной проток и правый лимфатический проток, впадающие в крупные вены шеи. По строению различают лимфатические сосуды безмышечного (фиброзного) и мышечного типов.
Лимфатические капилляры. Лимфатические капилляры - начальные отделы лимфатической системы, в которые из тканей поступает тканевая жидкость вместе с продуктами обмена веществ, а в патологических случаях - инородные частицы и микроорганизмы. По лимфатическому руслу могут распространяться и клетки злокачественных опухолей.
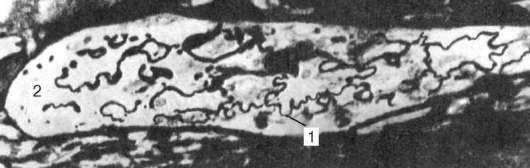
Рис. 13.15. Лимфатический капилляр. Тотальный препарат (импрегнация нитратом серебра):
1 - границы эндотелиальных клеток; 2 - замкнутый конец капилляра (по Янчо)
Лимфатические капилляры представляют собой систему замкнутых с одного конца, уплощенных эндотелиальных трубок, анастомозирую-щих друг с другом и пронизывающих органы (рис. 13.15; см. рис. 13.5). Диаметр лимфатических капилляров в несколько раз больше, чем кровеносных. В лимфатической системе, как и в кровеносной, почти всегда имеются резервные капилляры, наполняющиеся лишь при усилении лимфообразования.
Стенка лимфатических капилляров состоит из эндотелиальных клеток, которые в 3-4 раза крупнее, чем клетки кровеносных капилляров. Базальная мембрана и перициты в лимфатических капиллярах отсутствуют. Эндотелиальная выстилка лимфатического капилляра тесно связана с окружающей соединительной тканью с помощью так называемых стропных, или фиксирующих, филаментов, которые вплетаются в коллагеновые волокна, расположенные вдоль лимфатических капилляров (рис. 13.16). Лимфатические капилляры и начальные отделы отводящих лимфатических сосудов обеспечивают гема-толимфатическое равновесие как необходимое условие микроциркуляции в здоровом организме.
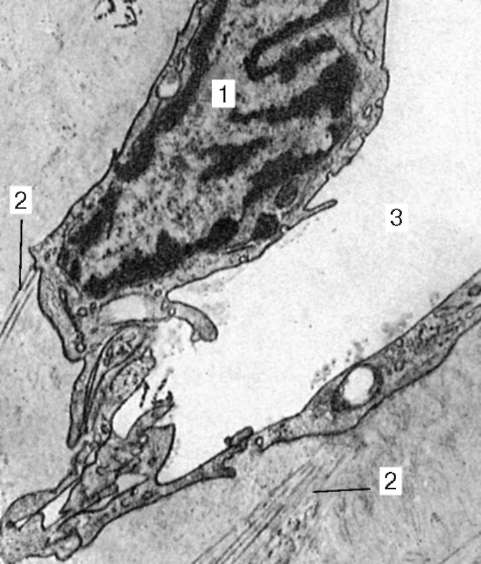
Рис. 13.16. Лимфатический капилляр перикарда крысы. Электронная микрофотография, увеличение 6300: 1 - эндотелиоцит; 2 - фиксирующие фи-ламенты; 3 - просвет капилляра (по Г. В. Булановой)
Отводящие лимфатические сосуды. Основной отличительной особенностью строения лимфатических сосудов является наличие в них клапанов и хорошо развитой наружной оболочки. В местах расположения клапанов лимфатические сосуды колбовидно расширяются. В строении стенок лимфатические сосуды имеют много общего с венами. Это объясняется сход-
ством лимфо- и гемодинамических условий этих сосудов: наличием низкого давления и направлением тока жидкости от органов к сердцу.
Лимфатические сосуды в зависимости от диаметра подразделяются на мелкие, средние и крупные. Как и вены, эти сосуды по своему строению могут быть безмышечными и мышечными. В мелких сосудах диаметром 30-40 мкм, которые являются главным образом внутриорган-ными лимфатическими сосудами, мышечные элементы отсутствуют и их стенка состоит из эндотелия и соединительнотканной оболочки.
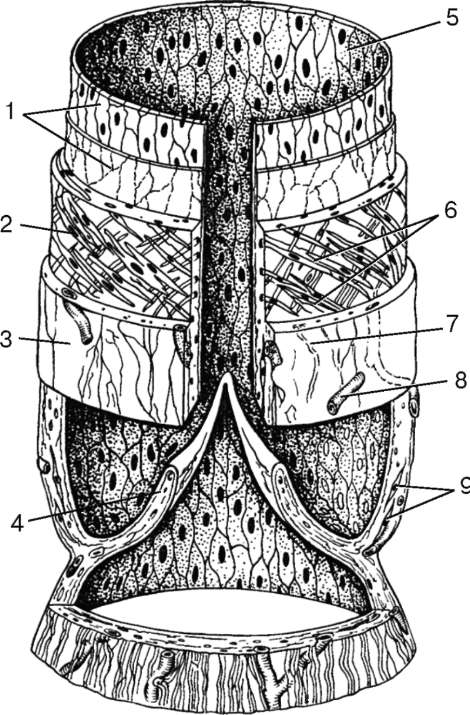
Средние и крупные лимфатические сосуды (диаметром более 0,2 мм) имеют три хорошо развитые оболочки: внутреннюю, среднюю и наружную. Во внутренней оболочке под эндотелием находятся продольно и косо ориентированные пучки коллагеновых и эластических волокон. Дупликатура внутренней оболочки формирует многочисленные клапаны. Участки, расположенные между двумя соседними клапанами, называются клапанным сегментом, или лимфангионом. В лимфангио-не выделяют мышечную манжетку, стенку клапанного синуса и область прикрепления клапана (рис. 13.17). Клапаны состоят из центральной соединительнотканной пластинки, покрытой с внутренней и наружной поверхностей эндотелием. Под эндотелием створки клапана, обращенной к стенке сосуда, располагается эластическая мембрана. В толще центральной соединительнотканной пластинки клапана обнаруживаются пучки гладких мышечных клеток. На границе внутренней и средней оболочек лежит не всегда хорошо выраженная внутренняя эластическая мембрана.
Средняя оболочка лимфатических сосудов слабо развита в сосудах головы, верхней части туловища и верхних конечностей. В лимфатических сосудах нижних конечностей она, наоборот, выражена отчетливо. В стенке этих сосудов находятся пучки гладких мышечных клеток, имеющие циркулярное и косое направление. Большого развития достигает мышечный слой в средней оболочке коллекторов подвздошного лимфатического сплетения,
Рис. 13.17. Лимфангион (схема по А. В. Борисову):
1 - внутренняя оболочка; 2 - средняя оболочка; 3 - наружная оболочка; 4 - створка клапана; 5 - эндотелиоциты; 6 - миоциты мышечной манжетки, залегающие в два слоя под углом 45° к продольной оси лимфангиона; 7 - пучки коллагеновых волокон наружной оболочки; 8 - кровеносные капилляры наружной оболочки; 9 - гладкие мышечные клетки в основании клапана
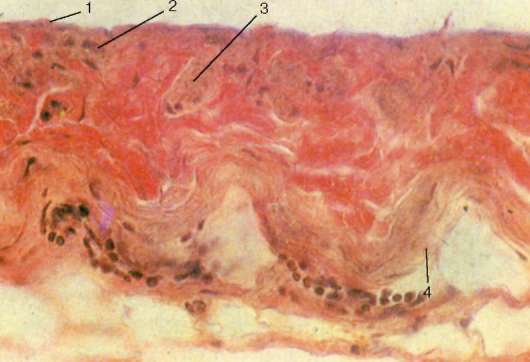
Рис. 13.18. Грудной лимфатический проток. Продольный срез (препарат В. А. Куд-ряшовой):
1 - эндотелий; 2 - продольно ориентированные гладкие миоциты внутренней оболочки; 3 - циркулярно ориентированные гладкие миоциты средней оболочки; 4 - адвентициальная оболочка
околоаортальных лимфатических сосудов и шейных лимфатических стволов, сопровождающих яремные вены. Эластические волокна в средней оболочке могут различаться по количеству, толщине и направлению.
Наружная оболочка лимфатических сосудов образована рыхлой соединительной тканью, которая без резкой границы переходит в окружающую соединительную ткань. Иногда в наружной оболочке встречаются отдельные продольно направленные гладкие мышечные клетки.
В качестве примера строения крупного лимфатического сосуда рассмотрим один из главных лимфатических стволов - грудной лимфатический проток. Его стенка имеет неодинаковое строение на различных уровнях. Наиболее сильного развития она достигает на уровне диафрагмы (рис. 13.18). На этом месте в стенке сосуда выделяются три оболочки, напоминающие по своему строению оболочки нижней полой вены. Внутренняя и средняя оболочки выражены относительно слабо. Цитоплазма эндотелиаль-ных клеток богата пиноцитозными пузырьками. Это указывает на активный трансэндотелиальный транспорт жидкости. Базальная часть клеток неровная. Сплошной базальной мембраны нет.
В субэндотелиальном слое рыхло залегают пучки коллагеновых фибрилл. Несколько глубже находятся единичные гладкие мышечные клетки, имеющие во внутренней оболочке продольное, а в средней - косое и циркулярное направление. На границе внутренней и средней оболочек иногда встречается плотное сплетение тонких эластических волокон, которое сравнивают с внутренней эластической мембраной. Как и в кровеносных сосу-
дах, эти эластические волокна связаны с подобными элементами других оболочек грудного протока в единый эластический каркас.
В средней оболочке расположение эластических волокон в основном совпадает с циркулярным и косым направлением пучков гладких мышечных клеток. Наружная оболочка грудного лимфатического протока в 3-4 раза толще двух других оболочек и содержит мощные продольно лежащие пучки гладких мышечных клеток, разделенные прослойками соединительной ткани. Толщина мышечных слоев грудного лимфатического протока, особенно в наружной его оболочке, уменьшается в направлении тока лимфы. При этом стенка лимфатического протока в его устье оказывается в 2-3 раза тоньше, чем на уровне диафрагмы. На протяжении трудного протока встречается до 9 полулунных клапанов. Створки клапанов состоят из тех же элементов, что и внутренняя оболочка протока. У основания клапана в стенке протока наблюдается утолщение, образованное скоплением соединительной ткани и гладких мышечных клеток, направленных циркулярно. В створках клапанов имеются единичные мышечные клетки, расположенные поперечно.
Васкуляризация сосудов. Все крупные и средние кровеносные сосуды имеют для своего питания собственную систему, носящую название «сосуды сосудов». Они приносят артериальную кровь к стенке сосудов из артерий, проходящих в окружающей соединительной ткани. В артериях сосуды сосудов проникают до глубоких слоев средней оболочки. Внутренняя оболочка артерий получает питательные вещества непосредственно из крови, протекающей в данной артерии. В диффузии питательных веществ через внутреннюю оболочку артерий большую роль играют белково-гликозаминогликановые комплексы, входящие в состав основного вещества стенок этих сосудов. Кровеносные капилляры стенок артерий собираются в вены, которые чаще всего попарно сопровождают соответствующую артерию и открываются в близлежащую вену. В венах сосуды сосудов снабжают артериальной кровью все три оболочки. Капилляры стенок вен открываются в просвет той же вены. В крупных лимфатических сосудах артерии и вены, питающие их стенки, идут раздельно.
Кроме кровеносных сосудов, в стенке артерий, вен и лимфатических стволов находятся лимфатические сосуды.
Возрастные изменения. Строение сосудов непрерывно меняется в течение всей жизни человека. Развитие сосудов под влиянием функциональной нагрузки заканчивается примерно к 30 годам. В дальнейшем в стенках артерий происходит разрастание соединительной ткани, что ведет к их уплотнению. В артериях эластического типа этот процесс выражен сильнее, чем в остальных артериях. После 60-70 лет во внутренней оболочке всех артерий обнаруживаются очаговые утолщения коллагеновых волокон, в результате чего в крупных артериях внутренняя оболочка по толщине приближается к средней. В мелких и средних артериях внутренняя оболочка разрастается слабее. Внутренняя эластическая мембрана с возрастом постепенно истончается и расщепляется. Мышечные клетки средней оболочки атрофируются. Эластические волокна подвергаются зернистому распаду и фрагмен-
тации, в то время как коллагеновые волокна разрастаются. Одновременно с этим во внутренней и средней оболочках у пожилых людей появляются известковые и липидные отложения, которые прогрессируют с возрастом. В наружной оболочке у людей старше 60-70 лет появляются продольно лежащие пучки гладких мышечных клеток.
Возрастные изменения в венах сходны с таковыми в артериях. Однако перестройка стенки вены человека начинается еще на первом году жизни. Так, к моменту рождения человека в средней оболочке стенок бедренной и подкожных вен нижних конечностей имеются лишь пучки циркулярно ориентированных мышечных клеток. Только к моменту вставания на ноги (к концу первого года) и повышения дистального гидростатического давления развиваются продольные мышечные пучки. Просвет вены по отношению к просвету артерии у взрослых (2:1) больше, чем у детей (1:1). Расширение просвета вен обусловлено меньшей эластичностью стенки вен, возрастанием у взрослых кровяного давления.
Сосуды сосудов до возраста 50-60 лет, как правило, бывают умеренно спазмированными, после 65-70 лет просвет их расширяется.
Лимфатические сосуды многих органов у людей старческого возраста характеризуются многочисленными мелкими варикозными вздутиями и выпячиваниями. Во внутренней оболочке стенок крупных лимфатических стволов и грудного протока у людей старше 35 лет увеличивается количество коллагеновых волокон. Этот процесс значительно прогрессирует к 60-70 годам. Одновременно количество мышечных клеток и эластических волокон уменьшается.
Регенерация. Мелкие кровеносные и лимфатические сосуды обладают способностью к регенерации. Восстановление дефектов сосудистой стенки после ее повреждения начинается с регенерации и роста ее эндотелия. Уже к концу первых - началу вторых суток на месте нанесенного повреждения наблюдается многочисленное деление эндотелиальных клеток. Мышечные клетки поврежденного сосуда, как правило, восстанавливаются более медленно и неполно по сравнению с другими тканевыми элементами сосуда. Восстановление их происходит частично путем деления миоцитов, а также в результате дифференцировки перицитов. Эластические элементы развиваются слабо. В случае полного разрыва среднего и крупного сосудов регенерации его стенки без оперативного вмешательства, как правило, не наступает, хотя восстановление циркуляции крови в соответствующей области может наблюдаться очень рано. Это происходит, с одной стороны, благодаря компенсаторной перестройке коллатеральных сосудов, а с другой - вследствие развития и роста новых мелких сосудов - капилляров. Новообразование капилляров начинается с того, что цитоплазма эндоте-лиальных клеток артериол и венул набухает в виде почки, затем эндотели-альные клетки подвергаются делению. По мере роста эндотелиальной почки в ней появляется полость. В развитии и росте эндотелиальной почки участвуют перициты, которые своими факторами оказывают влияние на пролиферацию эндотелиоцитов. Такие слепо заканчивающиеся трубки растут навстречу друг другу и смыкаются концами. Цитоплазматические перего-
родки между ними истончаются и прорываются, и во вновь образованном капилляре устанавливается циркуляция крови.
Лимфатические сосуды после их повреждения регенерируют несколько медленнее, чем кровеносные. Регенерация лимфатических сосудов может происходить за счет или почкования дистальных концов эндотелиальных трубок, или перестройки лимфатических капилляров в отводящие сосуды.
13.3. СЕРДЦЕ
Сердце (соr) - основной орган, приводящий в движение кровь.
Развитие. Первая закладка сердца появляется в начале 3-й нед развития у эмбриона длиной 1,5 мм в виде парного скопления мезенхимных клеток, которые расположены под висцеральным листком спланхнотома. Позднее эти скопления превращаются в две удлиненные трубочки, впадающие вместе с прилегающими висцеральными листками спланхнотома мезодермы в цело-мическую полость тела (рис. 13.19). В дальнейшем мезенхимные трубки сливаются и из их стенок образуются тканевые элементы эндокарда. Та область висцеральных листков спланхнотома мезодермы, которая прилежит к этим трубкам, получила название миоэпикардиальных пластинок. Последние приближаются к закладке эндокарда, окружают ее снаружи и сливаются друг с другом. Этот процесс идет в краниокаудальном направлении. Вначале возникают желудочковые, затем предсердные и синусно-предсердные зоны будущего сердца. Миоэпикардиальные пластинки дифференцируются на две части: во внутренней, прилежащей к мезенхимной трубке, находятся стволовые кардио-миобласты, а в наружной - тканевые элементы эпикарда.
Клетки зачатка миокарда - кардиомиобласты - делятся и дифференцируются в кардиомиоциты (см. главу 9). Их объем увеличивается, и на 2-м мес развития зародыша в них появляются миофибриллы с поперечной исчерченностью. Z-полоски появляются одновременно с саркотубулярной сетью и поперечными инвагинациями клеточной мембраны (Т-системы). На плазмолеммах контактирующих кардиомиоцитов местами отмечаются десмосомоподобные структуры. Формирующиеся в кардиомиоцитах мио-фибриллы также прикрепляются к плазмолеммам, где позднее образуются вставочные диски.
В конце 2-го мес появляются признаки формирования проводящей системы, кардиомиоциты которой отличаются многоядерностью, замедленной дифференцировкой миофибриллярного аппарата. К 4-му мес заканчивается образование всех отделов проводящей системы сердца. Развитие мышечной ткани левого желудочка происходит быстрее, чем правого.
Клапаны сердца - предсердно-желудочковые и желудочково-сосудистые - развиваются в основном как дупликатура эндокарда.
Левый предсердно-желудочковый клапан появляется в виде эндокардиального валика, в который позднее (у эмбриона 2,5 мес) начинает врастать соединительная
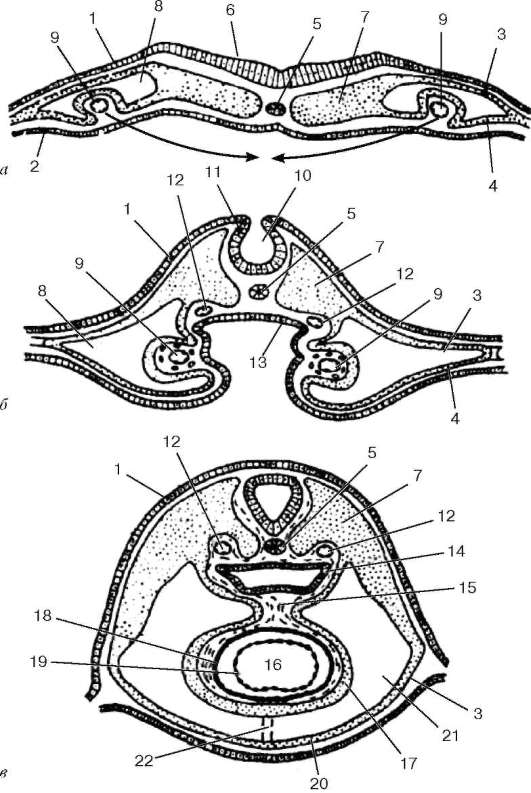
Рис. 13.19. Развитие сердца. Поперечные разрезы зародышей на трех последовательных стадиях формирования сердца (по Штралю, Гису и Борну):
а - две парные закладки сердца; б - их сближение; в - слияние в одну непарную закладку. 1 - эктодерма; 2 - энтодерма; 3 - париетальный листок спланхнотома; 4 - висцеральный листок спланхнотома; 5 - хорда; 6 - нервная пластинка; 7 - сомит; 8 - вторичная полость тела; 9 - эндотелиальная закладка сердца (парная); 10 - нервный желобок; 11 - нервные валики; 12 - нисходящая аорта (парная); 13 - образующаяся головная кишка; 14 - головная кишка; 15 - спинная сердечная брыжейка; 16 - полость сердца; 17 - эпикард; 18 - миокард; 19 - эндокард; 20 - околосердечная сумка; 21 - перикардиальная полость; 22 - редуцирующаяся брюшная сердечная брыжейка
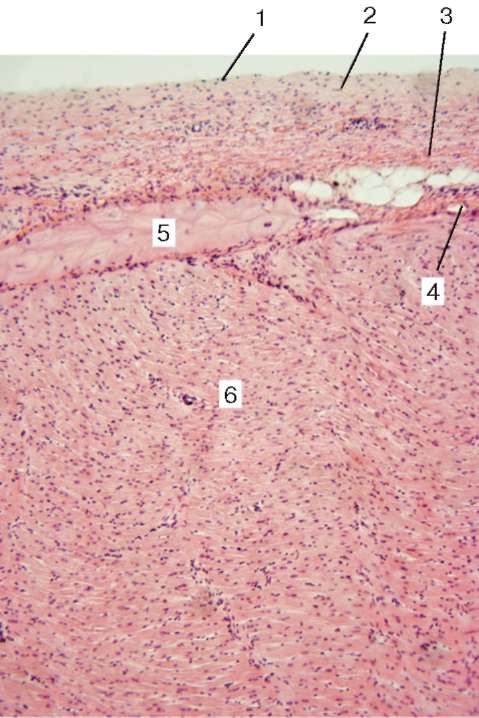
Рис. 13.20. Строение стенки сердца: 1 - эндотелий; 2 - субэндотелиальный слой; 3 - мышечно-эластический слой; 4 - капилляры; 5 - атипичные мышечные клетки (проводящие миоциты); 6 - типичные кардиомиоциты миокарда (микрофотография, малое увеличение)
ткань из эпикарда. На 4-м мес внутриутробного периода из эпикарда в створку клапана врастает пучок коллагеновых волокон, образующий в будущем фиброзную пластинку. Правый предсердно-желудочковый клапан закладывается как мышечно-эндокардиальный валик. С 3-го мес развития зародыша мышечная ткань правого атриовентрикуляр-ного клапана уступает место соединительной ткани, врастающей со стороны миокарда и эпикарда. У взрослого человека мышечная ткань сохраняется в виде рудимента только с предсердной стороны в основании клапана. Таким образом, предсердно-желудочковые клапаны являются производными не только эндокарда, но и соединительной ткани миокарда и эпикарда. Аортальные клапаны имеют двойное происхождение: синусная сторона их образуется из соединительной ткани фиброзного кольца, которая покрывается эндотелием, а желудочковая - из эндокарда. Первые нервные терминали выявляются в предсердиях 5,5-недельных эмбрионов человека, а на 8-й нед в предсердиях обнаруживаются ганглии, состоящие из 4-10 нейробластов. Из клеток нервного гребня, мигрировавших в зачаток предсердий, образуются холинергические нейроны, глиоциты и мелкие грануляр-
ные клетки. Холинергический и адренергический нервные аппараты сердца развиваются почти одновременно. Врастание нервных волокон в развивающееся сердце идет поэтапно. Сначала появляются нервные волокна в правом, затем в левом предсердии, позже - в правом, затем в левом желудочке. При этом вначале в предсердиях выявляются веточки от симпатических стволов, а позднее - ветви грудных симпатических волокон.
Строение. В стенке сердца различают три оболочки: внутреннюю - эндокард, среднюю, или мышечную, - миокард, и наружную, или серозную, - эпикард (рис. 13.20).
13.3.1. Эндокард
Эндокард (endocardium) выстилает изнутри камеры сердца, сосочковые мышцы, сухожильные нити, а также клапаны сердца. Толщина эндокарда в различных участках неодинакова. Он толще в левых камерах сердца, особенно на межжелудочковой перегородке и у устья крупных артериальных стволов - аорты и легочной артерии, а на сухожильных нитях зна-
чительно тоньше. Поверхность эндокарда, обращенная в полость сердца, выстлана эндотелием, состоящим из полигональных клеток, лежащих на толстой базальной мембране (см. рис. 13.20). За ним следует субэндоте-лиальный слой, образованный соединительной тканью, богатой мало-дифференцированными соединительнотканными клетками. Глубже располагается мышечно-эластический слой, в котором эластические волокна переплетаются с гладкими мышечными клетками. Эластические волокна гораздо лучше выражены в эндокарде предсердий, чем в эндокарде желудочек. Гладкие мышечные клетки сильнее всего развиты в эндокарде у места выхода аорты и могут иметь многоотростчатую форму. Самый глубокий слой эндокарда - наружный соединительнотканный - лежит на границе с миокардом. Он состоит из соединительной ткани, содержащей толстые эластические, коллагеновые и ретикулярные волокна.
Питание эндокарда осуществляется главным образом диффузно за счет крови, находящейся в камерах сердца. Кровеносные сосуды имеются лишь в наружном соединительнотканном слое эндокарда.
Клапаны
Между предсердиями и желудочками сердца, а также желудочками и крупными сосудами располагаются клапаны. Предсердно-желудочковый (атриовен-трикулярный) клапан в левой половине сердца двустворчатый, в правой - трехстворчатый. Они представляют собой покрытые эндотелием тонкие фиброзные пластинки из плотной волокнистой соединительной ткани с небольшим количеством клеток (рис. 13.21). Эндотелиальные клетки, покрывающие клапан, частично перекрывают друг друга в виде черепицы или образуют пальцевидные вдавливания цитоплазмы одной клетки в другую. Кровеносных сосудов створки клапанов не имеют. В субэндотелиальном слое выявлены тонкие кол-лагеновые волокна, которые постепенно переходят в фиброзную пластинку створки клапана, а в месте прикрепления дву- и трехстворчатого клапанов - в фиброзные кольца. В основном веществе створок клапанов обнаружено большое количество гликозаминогликанов.
Строение предсердных и желудочковых частей створок клапанов неодинаково.
Предсердная сторона их имеет гладкую поверхность, здесь в субэндотелиаль-ном слое располагаются густое сплетение эластических волокон и пучки гладких мышечных клеток. Количество мышечных пучков увеличивается в основании клапана. Желудочковая сторона имеет неровную поверхность. Она снабжена выростами, от которых начинаются сухожильные нити (chordae tendineae). В этой области под эндотелием располагается лишь небольшое количество эластических волокон. На границе между восходящей частью дуги аорты и левым желудочком сердца локализуются аортальные клапаны. По своему строению они имеют много общего с предсердно-желудочковыми клапанами и клапанами легочной артерии. На вертикальном разрезе в створке клапана можно различить три слоя: внутренний, средний и наружный. Внутренний слой, обращенный к желудочку сердца, представляет собой продолжение эндокарда. Эндотелий этого слоя характеризуется наличием пучков филаментов толщиной 5-8 нм и многочисленных пиноцитозных
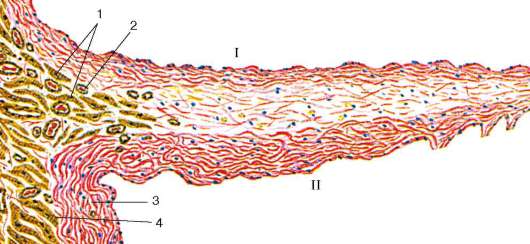
Рис. 13.21. Предсердно-желудочковый (атриовентрикулярный) клапан сердца человека (по В. Я. Бочарову):
I - предсердная сторона; II - желудочковая сторона; 1 - сердечная мышечная ткань в основании створки клапана; 2 - кровеносные сосуды; 3 - эндокард левого желудочка; 4 - миокард левого желудочка
пузырьков. В субэндотелиальном слое содержатся фибробласты с длинными тонкими отростками, которые в виде консолей поддерживают эндотелиальные клетки. К субэндотелиальному слою прилежат плотные пучки коллагеновых фибрилл, идущих продольно и поперечно, за которым следует смешанная эластикоколлагеновая прослойка. Средний слой тонкий, состоит из рыхлой волокнистой соединительной ткани, богатой клеточными элементами.
Наружный слой, обращенный к аорте, кроме эндотелия, содержит коллагеновые волокна, которые берут начало от фиброзного кольца вокруг аорты. Опорный скелет сердца образован фиброзными кольцами между предсердиями и желудочками и плотной соединительной тканью в устьях крупных сосудов. Кроме плотных пучков коллагеновых волокон, в составе «скелета» сердца имеются эластические волокна, а иногда бывают даже хрящевые пластинки.
13.3.2. Миокард
Многотканевая мышечная оболочка сердца (myocardium) состоит из тесно связанных между собой поперечнополосатых мышечных клеток - кардио-миоцитов (см. главу 9). Между мышечными элементами располагаются прослойки рыхлой соединительной ткани, сосуды, нервы. Различают сократительные (рабочие) сердечные миоциты (myociti cardiaci), проводящие сердечные миоциты (myocyti cardiacus conducens), входящие в состав так называемой проводящей системы сердца, и секреторные предсердные кардиомиоциты (cardiomyocyti atrialis secretans).
Сердечные сократительные (рабочие) миоциты характеризуются рядом структурных и цитохимических особенностей. На продольных срезах они почти прямоугольной формы, длина колеблется от 50 до 120 мкм, ширина составляет 15-20 мкм. Клетки покрыты сарколеммой, состоящей из плазмо-
леммы и базальной мембраны, в которую вплетаются тонкие коллагеновые и эластические волокна, образующие «наружный скелет» кардиомиоцитов. Базальная мембрана кардиомиоцитов, содержащая большое количество гликопротеинов, способных связывать Са2+, может принимать участие наряду с саркотубулярной сетью и митохондриями в перераспределении Са2+ в цикле сокращение - расслабление. Базальная мембрана латеральных сторон - кардиомиоцитов инвагинирует в канальцы Т-системы (в отличие от соматических мышечных волокон).
Кардиомиоциты желудочков значительно интенсивнее пронизаны канальцами Т-системы, чем соматические мышечные волокна. Канальцы L-системы (латеральные расширения саркоплазматической сети) и Т-системы образуют диады (один каналец L-системы и один - Т-системы), реже триады (два канальца L-системы и один - Т-системы). В центральной части миоцита расположены одно-два ядра овальной или удлиненной формы. Между миофибриллами находятся многочисленные митохондрии.
В отличие от желудочковых кардиомиоцитов, форма которых близка к цилиндрической, предсердные миоциты чаще отростчатые, их размеры меньше. В миоцитах предсердий меньше митохондрий, миофибрилл саркоплазматической сети. В пред-сердных кардиомиоцитах менее выражена активность сукцинатдегидрогеназы, но более высока активность ферментов, связанных с метаболизмом гликогена (фосфо-рилаза, гликогенсинтетаза и др.). Отличительными особенностями этих кардиомио-цитов являются относительно хорошо развитая гранулярная эндоплазматическая сеть и значительное развитие комплекса Гольджи. Указанные выше морфологические признаки связаны с наличием в предсердных кардиомиоцитах специфических предсердных гранул, содержащих гормоноподобные пептиды (атриопептин, натрийуретический фактор типа С). Секреторные сократительные предсердные миоциты (эндокринные предсердные миоциты) располагаются преимущественно в правом предсердии и ушках сердца. При растяжении предсердий секрет поступает в кровь и воздействует на собирательные трубочки почки, клетки клубочковой зоны коры надпочечников, участвующие в регуляции объема внеклеточной жидкости и уровня артериального давления.
Еще одной отличительной чертой предсердных миоцитов у многих млекопитающих является слабое развитие Т-системы канальцев. В тех предсердных мио-цитах, где нет Т-системы, на периферии клеток, под сарколеммой, располагаются многочисленные пиноцитозные пузырьки и кавеолы. Полагают, что эти пузырьки и кавеолы являются функциональными аналогами Т-канальцев.
Энергия, необходимая для сокращения сердечной мышцы, образуется главным образом за счет взаимодействия АДФ с креатинфосфатом, в результате чего возникают креатин и АТФ. Главным субстратом дыхания в сердечной мышце являются жирные кислоты и в меньшей степени - углеводы. Процессы анаэробного расщепления углеводов (гликолиз) в миокарде (кроме проводящей системы) человека практического значения не имеют.
Кардиомиоциты сообщаются между собой в области вставочных дисков (disci intercalati). В гистологических препаратах они имеют вид темных полосок. Строение вставочного диска на его протяжении неодинаково (см. рис. 9.10 и 9.11). Различают десмосомы, места вплетения миофибрилл в плазмо-лемму (промежуточные контакты) и щелевые контакты - нексусы. Если первые два участка диска выполняют механическую функцию, то третий
осуществляет электрическую связь кардиомиоцитов. Нексусы обеспечивают быстрое проведение импульсов от клетки к клетке. Зоны прикрепления миофибрилл всегда располагаются на уровне, соответствующем очередной Z-линии.
Иммуноцитохимически в этих участках показано наличие L-актинина и винку-лина. Как и в скелетных мышцах, в кардиомиоцитах цитоскелет представлен промежуточными филаментами диаметром 10 нм. Эти филаменты, состоящие из белка десмина или скелетина, располагаются как вдоль длинной оси, так и поперек. При этом промежуточные нити проходят поперек через М- и Z-линии миофибрилл, скрепляя их и поддерживая соседние саркомеры на одном уровне.
С помощью вставочных дисков кардиомиоциты соединяются в мышечные «волокна». Продольные и боковые связи (анастомозы) кардиомиоцитов обеспечивают функциональное единство миокарда.
Между кардиомиоцитами находится интерстициальная соединительная ткань, содержащая большое количество кровеносных и лимфатических капилляров. Каждый миоцит контактирует с двумя-тремя капиллярами.
Проводящая система сердца
Проводящая система сердца (systema conducens cardiacum) - мышечные клетки, формирующие и проводящие импульсы к сократительным клеткам сердца. В состав проводящей системы входят синусно-предсердный (синусный) узел, предсердно-желудочковый (атриовентрикулярный) узел, предсердно-желудочковый пучок (пучок Гиса) и их разветвления (волокна Пуркинье), передающие импульсы на сократительные мышечные клетки.
Различают несколько типов мышечных клеток, которые в неодинаковых соотношениях находятся в различных отделах этой системы (рис. 13.22).
Клетки узла проводящей системы. Формирование импульса происходит в синусном узле, центральную часть которого занимают возбуждающие кар-диомиоциты - водители ритма, или пейсмекерные клетки (Р-клетки), способные к самопроизвольным сокращениям (см. рис. 13.22). Они отличаются небольшими размерами, многоугольной формой с максимальным диаметром 8-10 мкм, небольшим количеством миофибрилл, не имеющих упорядоченной ориентировки.
Миофиламенты в составе миофибрилл расположены рыхло. А- и I-диски различаются нечетко. Митохондрии небольшие, округлой или овальной формы, немногочисленные. Саркоплазматическая сеть развита слабо. Т-система отсутствует, но вдоль плазмолеммы находится много пиноцитозных пузырьков и кавеол, которые в 2 раза увеличивают мембранную поверхность клеток. Высокое содержание свободного кальция в цитоплазме этих клеток при слабом развитии саркоплазматической сети обусловливает способность клеток синусного узла генерировать импульсы к сокращению. Поступление необходимой энергии обеспечивается преимущественно процессами гликолиза. Между клетками встречаются единичные десмосомы и нексусы.
По периферии узла располагаются переходные кардиомиоциты. Это тонкие, вытянутые клетки, поперечное сечение которых меньше поперечного сечения типичных сократительных кардиомиоцитов. Миофибриллы более
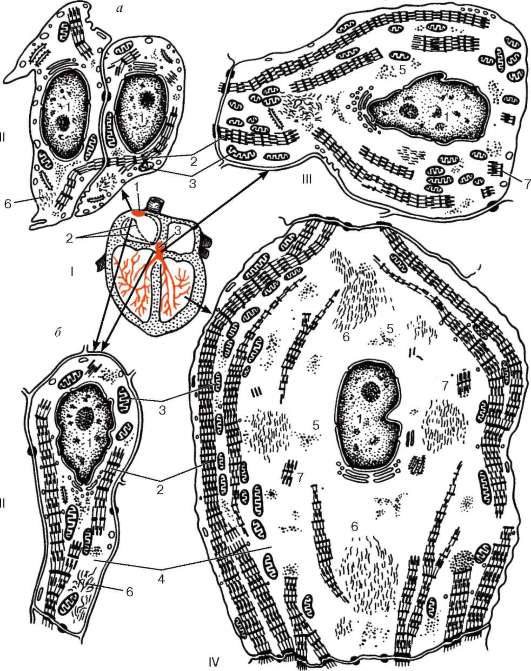
Рис. 13.22. Кардиомиоциты проводящей системы сердца (по П. П. Румянцеву): I - схема расположения элементов проводящей системы сердца; II - кардиомиоциты синусного и предсердно-желудочкового узлов: а - Р-клетки; б - переходные клетки; III - кардиомиоцит из пучка Гиса; IV - кардиомиоцит из ножек пучка Гиса (волокна Пуркинье). 1 - ядра; 2 - миофибриллы; 3 - митохондрии; 4 - саркоплазма; 5 - глыбки гликогена; 6 - промежуточные филаменты; 7 - миофиламентные комплексы
развиты, ориентированы параллельно друг другу, но не всегда. Отдельные переходные клетки могут содержать короткие Т-трубочки. Переходные клетки сообщаются между собой как с помощью простых контактов, так и путем образования более сложных соединений типа вставочных дисков. Функциональное значение этих клеток состоит в передаче возбуждения от Р-клеток к клеткам пучка и рабочему миокарду.
Кардиомиоциты предсердно-желудочкового пучка проводящей системы (пучка Гиса) и его ножек (волокон Пуркинье) содержат относительно длинные миофибриллы, имеющие спиралевидный ход. В функциональном отношении они являются передатчиками возбуждения от переходных клеток к клеткам рабочего миокарда желудочков.
Мышечные клетки проводящей системы в стволе и разветвлениях ножек ствола проводящей системы располагаются небольшими пучками, они окружены прослойками рыхлой волокнистой соединительной ткани. Ножки пучка разветвляются под эндокардом, а также в толще миокарда желудочков. Кардиомиоциты проводящей системы разветвляются в миокарде и проникают в сосочковые мышцы. Это обусловливает натяжение сосочковыми мышцами створок клапанов (левого и правого) еще до того, как начнется сокращение миокарда желудочков.
По строению кардиомиоциты пучка отличаются большим диаметром (15 мкм и более), почти полным отсутствием Т-систем, тонкостью миофи-брилл, которые без определенного порядка располагаются главным образом по периферии клетки. Ядра, как правило, расположены эксцентрично. Эти клетки в совокупности образуют предсердно-желудочковый пучок и ножки пучка (волокна Пуркинье). Кардиомиоциты в составе этих волокон самые крупные не только в проводящей системе, но и во всем миокарде. В них много гликогена, редкая сеть миофибрилл, нет Т-трубочек. Клетки связаны между собой нексусами и десмосомами.
В проводящей системе сердца преобладают энзимы, принимающие участие в анаэробном гликолизе (фосфорилаза, дегидрогеназа молочной кислоты). Понижена активность аэробных ферментов цикла трикарбоновых кислот и митохондриальной цепи переноса электронов (цитохромоксидаза). В проводящих волокнах содержание калия ниже, а кальция и натрия выше, чем в сократительных кардиомиоцитах.
В миокарде много афферентных и эфферентных нервных волокон (рис. 13.23, а, б). Типичных нервно-мышечных синапсов здесь нет. Раздражение нервных волокон, окружающих проводящую систему, а также нервов, подходящих к сердцу, вызывает изменение ритма сердечных сокращений. Это указывает на решающую роль нервной системы в ритме сердечной деятельности, а следовательно, и в передаче импульсов по проводящей системе.
13.3.3. Эпикард и перикард
Наружная оболочка сердца, или эпикард (epicardium), представляет собой висцеральный листок перикарда (pericardium). Эпикард образован тонкой (не более 0,3-0,4 мм) пластинкой соединительной ткани, плотно срастающейся с миокардом. Свободная поверхность ее покрыта мезотелием.
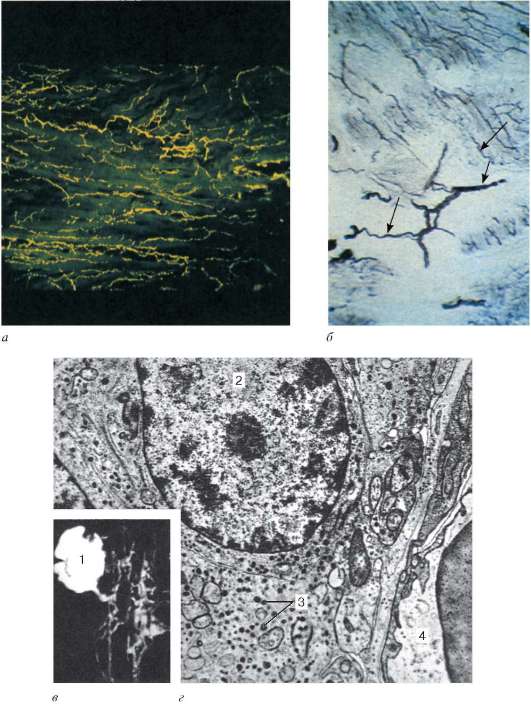
Рис. 13.23. Адренергические (а) и холинергические (б) нервные волокна и малые интенсивно флюоресцирующие клетки - МИФ-клетки (в, г):
а - алюминий-формальдегидный метод (препарат Р. А. Стропуса); б - по методу М. Карновского; в - флюоресцентная микроскопия; г - электронная микрофотография, увеличение 10 000 (препарат А. А. Сосунова и В. Н. Швалева): 1 - малая интенсивно флюоресцирующая клетка; 2 - ядро; 3 - гранулы секрета; 4 - капилляр
В соединительнотканной основе эпикарда различают поверхностный слой коллагеновых волокон, слой эластических волокон, глубокий слой кол-лагеновых волокон и глубокий коллагеново-эластический слой, который составляет до 50 % всей толщи эпикарда. На предсердиях и некоторых участках желудочков последний слой отсутствует или сильно разрыхлен. Здесь же иногда отсутствует и поверхностный коллагеновый слой.
В париетальном листке перикарда соединительнотканная основа развита сильнее, чем в эпикарде. В ней много эластических волокон, особенно в глубоком его слое. Поверхность перикарда, обращенная к перикардиальной полости, тоже покрыта мезотелием. По ходу кровеносных сосудов встречаются скопления жировых клеток. Перикард имеет многочисленные нервные окончания, преимущественно свободного типа.
Васкуляризация. Венечные (коронарные) артерии имеют плотный эластический каркас, в котором четко выделяются внутренняя и наружная эластические мембраны. Гладкие мышечные клетки в артериях обнаруживаются в виде продольных пучков во внутренней и наружной оболочках. В основании клапанов сердца кровеносные сосуды у места прикрепления створок разветвляются на капилляры. Кровь из капилляров собирается в коронарные вены, впадающие в правое предсердие или венозный синус (строение вен - см. «Органные особенности строения сосудов»). Проводящая система, особенно ее узлы, обильно снабжена кровеносными сосудами. Лимфатические сосуды в эпикарде сопровождают кровеносные. В миокарде и эндокарде они проходят самостоятельно и образуют густые сети. Лимфатические капилляры обнаружены также в предсердно-желудочковых и аортальных клапанах. Из капилляров лимфа, оттекающая от сердца, направляется в парааортальные и парабронхиальные лимфатические узлы. В эпикарде и перикарде находятся сплетения сосудов микро-циркуляторного русла.
Иннервация. В стенке сердца обнаруживается несколько нервных сплетений (в основном из безмиелиновых волокон адренергической и холи-нергической природы) и ганглиев. Наибольшая плотность расположения нервных сплетений отмечается в стенке правого предсердия и синусно-предсердного узла проводящей системы. Рецепторные окончания в стенке сердца (свободные и инкапсулированные) образованы нейронами ганглиев блуждающих нервов и нейронами спинномозговых узлов (C-T) и, кроме того, ветвлениями дендритов равноотростчатых нейронов внутриорганных ганглиев (афферентные нейроны). Эффекторная часть рефлекторной дуги в стенке сердца представлена расположенными среди кардиомиоцитов и по ходу сосудов органа нервными волокнами холинергической природы, образованными аксонами находящихся в сердечных ганглиях длинноаксонных нейронов (эфферентные нейроны). Последние получают импульсы по пре-ганглионарным волокнам из нейронов ядер продолговатого мозга, приходящих сюда в составе блуждающих нервов. Эффекторные адренергические нервные волокна образованы ветвлениями аксонов нейронов ганглиев симпатической нервной цепочки. На этих нейронах также синапсами заканчиваются преганглионарные волокна - аксоны нейронов симпатических
ядер боковых рогов спинного мозга. Эффекторы представляют собой варикозные утолщения по ходу адренергических нервных волокон, содержащие синаптические пузырьки. В состав нервных ганглиев сердца входят богатые катехоламинами так называемые малые интенсивно флюоресцирующие клетки - МИФ-клетки (см. рис. 13.23). Это небольшие клетки (длиной 10-20 мкм), содержащие в цитоплазме много крупных гранулярных пузырьков (до 200 нм) с катехоламинами. Эндоплазматическая сеть в них развита слабо. На плазмолемме этих клеток обнаруживаются нервные окончания адренергических и холинергических нервов. Они рассматриваются как вставочные нейроны, выделяющие свои медиаторы в кровеносное русло.
Возрастные изменения. В течение онтогенеза можно выделить три периода изменения гистологической структуры сердца: период дифференциров-ки, период стабилизации и период инволюции. Дифференцировка гистологических элементов сердца, начавшаяся еще в зародышевом периоде, заканчивается к 16-20 годам. Существенное влияние на процессы диф-ференцировки кардиомиоцитов и морфогенез желудочков оказывает зара-щение овального отверстия и артериального протока, которое приводит к изменению гемодинамических условий - уменьшению давления и сопротивления в малом круге и увеличению давления в большом. Одновременно отмечаются физиологическая атрофия миокарда правого желудочка и физиологическая гипертрофия миокарда левого желудочка. В ходе дифферен-цировки сердечные миоциты обогащаются саркоплазмой, в результате чего их ядерно-цитоплазматическое отношение уменьшается. Количество миофибрилл прогрессивно увеличивается. Мышечные клетки проводящей системы при этом дифференцируются быстрее, чем сократительные. При дифференцировке соединительной ткани стромы сердца наблюдаются постепенное уменьшение количества ретикулярных волокон и замена их зрелыми коллагеновыми волокнами.
В период между 20 и 30 годами при обычной функциональной нагрузке сердце человека находится в стадии относительной стабилизации. В возрасте старше 30-40 лет в миокарде обычно начинается некоторое разрастание соединительнотканной стромы. При этом в стенке сердца, особенно в эпикарде, появляются адипоциты.
Степень иннервации сердца также изменяется с возрастом. Максимальная плотность внутрисердечных сплетений на единицу площади и высокая активность медиаторов отмечаются в период полового созревания. После 30-летнего возраста неуклонно уменьшаются плотность адренергических нервных сплетений и содержание медиаторов в них, а плотность холинер-гических сплетений и количество медиаторов в них сохраняются почти на исходном уровне. Нарушение равновесия в автономной иннервации сердца предрасполагает к развитию патологических состояний. В старческом возрасте уменьшается активность медиаторов и в холинергических сплетениях сердца.
Регенерация. У новорожденных, а возможно, и в раннем детском возрасте, когда способные к делению кардиомиоциты еще сохраняются, регенераторные процессы сопровождаются увеличением количества клеток.
У взрослых физиологическая регенерация кардиомиоцитов осуществляется главным образом путем внутриклеточной регенерации, без увеличения количества клеток. Клетки соединительной ткани всех оболочек пролифе-рируют, как в любом другом органе.
При повышенных систематических функциональных нагрузках общее количество клеток не возрастает, но в цитоплазме увеличиваются содержание органелл общего значения и миофибрилл, а также размер клеток (происходит функциональная гипертрофия кардиомиоцитов); соответственно возрастает и степень плоидности ядер.
Контрольные вопросы
1. Общий план строения сосудистой стенки; классификация и особенности строения артерий в зависимости от условий гемодинамики.
2. Эмбриональные источники развития сосудов, особенности строения вен в зависимости от условий гемодинамики.
3. Строение сосудов микроциркуляторного русла в функциональном аспекте.
4. Эмбриональные источники развития сердца, сократительные (рабочие) и атипические кардиомиоциты. Строение стенок предсердий и желудочков сердца, сердечных клапанов.