Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
ЦИТОЛОГИЯ. Глава 4. УЧЕНИЕ О КЛЕТКЕ (ОСНОВЫ ОБЩЕЙ ЦИТОЛОГИИ)
Основой строения эукариотических организмов является наименьшая единица живого - клетка (cellula).
Клетка - это ограниченная активной мембраной, упорядоченная структурированная система биополимеров (нуклеиновые кислоты, белки, полисахариды, липиды) и их макромолекулярных комплексов, образующих ядро и цитоплазму, осуществляющих поддержание и воспроизведение всей системы в целом.
Кроме клеток, в организме находятся их производные: симпласт, синцитий, межклеточное вещество (см. главу 5).
Содержимое клетки отделено от внешней среды плазматической мембраной (плазмолеммой). Все эукариотические клетки состоят из двух основных компонентов: ядра и цитоплазмы. В ядре различают хроматин (хромосомы), ядрышки, ядерную оболочку, нуклеоплазму (кариоплазму) и ядерный белковый остов (матрикс). Цитоплазма неоднородна по своему составу и строению и включает гиалоплазму (или основную плазму), в которой находятся органеллы; каждая из них выполняет обязательную функцию. Часть органелл имеет мембранное строение: эндоплазматическая сеть, комплекс Тольджи, лизосомы, пероксисомы и митохондрии. Немембранные органеллы цитоплазмы представлены рибосомами, клеточным центром, ресничками, жгутиками и компонентами цитоскелета. Кроме того, в гиалоплазме могут встретиться и иные необязательные структуры, или включения (жировые капли, пигментные гранулы и др.). Такое разделение клетки на отдельные компоненты не означает их структурной и функциональной обособленности. Все эти компоненты выполняют отдельные внутриклеточные функции, необходимые для существования клетки как целого, как элементарной единицы живого. Изучением общих черт строения и функционирования клеток занимается наука цитология, или, как ее теперь называют, биология клетки. Она исследует отдельные клеточные структуры, их участие в общеклеточных физиологических процессах, пути регуляции этих процессов, воспроизведение клеток и их органелл, приспособление клеток к условиям среды, реакции на действие различных
факторов. Изучение цитологии имеет большое значение для медицины, так как практически все болезни человека являются результатом различных клеточных поражений или нарушений функций клеток в составе тканей различных органов.
4.1. КЛЕТОЧНАЯ ТЕОРИЯ
Клеточная теория - это обобщенное представление о строении клеток как единиц живого, об их воспроизведении и роли в формировании многоклеточных организмов.
Появлению и формулированию отдельных положений клеточной теории предшествовал довольно длительный (более 300 лет) период накопления знаний о строении различных одноклеточных и многоклеточных организмов, растений и позвоночных животных. Все это легло в основу сформулированной Т. Шванном (1838) клеточной теории строения организмов (см. главу 3). Большую роль в развитии клеточной теории сыграли работы немецкого патолога Р. Вирхова.
В книге «Целлюлярная патология как учение, основанное на физиологической и патологической гистологии» (1855-1859) он обосновал фундаментальное положение о преемственности клеточного развития. Р. Вирхов, в противоположность Т. Шванну и М. Шлейдену, отстаивал взгляд на образование новых клеток не из «цитобластемы» - бесструктурной живой субстанции, а путем деления предсуществующих клеток (omnis cellula e cellula).
Создание клеточной теории и ее дальнейшее развитие стало важнейшим событием в биологии, одним из решающих доказательств единства происхождения всей живой природы. Клеточная теория оказала значительное влияние на развитие биологии и медицины, послужила главным фундаментом для становления таких дисциплин, как эмбриология, гистология. Принятие принципа клеточного строения организма оказало огромное влияние на физиологию, переведя ее на изучение реально функционирующих единиц - клеток. Она дала основы для научного понимания жизни, для понимания индивидуального развития и возникновения патологических изменений организмов.
Клеточная теория сохраняет свое значение до настоящего времени. Основные положения клеточной теории изложены ниже.

Р. Вирхов (1821-1902)
1. Клетка - наименьшая единица живого. Согласно одному из современных определений, живые организмы представляют собой открытые (т. е. обменивающиеся с окружающей средой веществами и энергией), саморегулирующиеся и самовоспроизводящиеся системы, важнейшими функционирующими компонентами которых являются белки и нуклеиновые кислоты. Все проявления жизни связаны с белками. Белки - функционирующие молекулы, обладающие сложной организацией и строгой функциональной специфичностью, которая определяется нуклеиновыми кислотами, несущими в себе информацию о строении тех или других белков. Живому свойствен ряд совокупных признаков: генетическая индивидуальность, способность к воспроизведению (репродукции), использование и трансформация энергии, метаболизм, реактивность и раздражимость, адаптивная изменчивость. Такую совокупность этих признаков впервые можно обнаружить только на клеточном уровне. Именно клетка как таковая является наименьшей единицей, обладающей всеми свойствами, отвечающими определению «живое».
2. Сходство клеток разных организмов по строению. Клетки могут иметь разнообразную внешнюю форму: шаровидную (лейкоциты), многогранную (клетки железистого эпителия), звездчатую и разветвленно-отростчатую (нервные и костные клетки), веретеновидную (гладкие мышечные клетки, фибробласты), столбчатую (кишечный эпителиоцит), уплощенную (эндо-телиоцит, мезотелиоцит) и др. Однако при изучении клеток тканей различных растений или животных обращает на себя внимание существование общего плана их организации (рис. 4.1). Такое сходство в строении клеток определяется общеклеточными функциями, связанными с поддержанием самой живой системы (синтез нуклеиновых кислот и белков, биоэнергетика клетки и др.). Одновременно это сходство указывает на общность происхождения всех эукариотических организмов.
Различие клеток в многоклеточном организме, обусловленное специализацией их функций, связано с развитием органелл специального значения. Так, если рассматривать мышечную клетку, то в ней, кроме общеклеточных структур (мембранные системы, рибосомы и др.), встречаются в большом количестве фибриллярные компоненты - миофиламенты и миофибриллы, обеспечивающие движение, сокращение. В нервной клетке, кроме общеклеточных компонентов, можно увидеть большое количество микротрубочек и промежуточных филаментов в клеточных отростках. Вся совокупность этих отличительных черт нервной клетки связана с ее специализацией - генерацией и передачей нервного импульса (подробно эти вопросы рассматриваются в разделе «Учение о тканях»).
3. Размножение клеток путем деления исходной клетки. Размножение прокариотических и эукариотических клеток происходит только путем деления исходной клетки, которому предшествует воспроизведение ее генетического материала (репликация ДНК). У эукариотических клеток единственно полноценным способом деления является митоз, или непрямое деление. При этом по двум дочерним клеткам распределяется равное количество хромосом, которые до этого удвоились в числе.
Митоз наблюдается у всех эукариотических (растительных и животных) клеток. Современная наука отвергает иные пути образования клеток и увеличения их числа в норме.
4. Клетки имеют одинаковый объем генетической информации. Это положение основано на том, что все клетки произошли от зиготы - одноклеточного зародыша. Однако морфологически и функционально клетки разных тканей значительно отличаются друг от друга. Несмотря на то, что потомки одноклеточного зародыша должны обладать одинаковыми генетическими потенциями, по мере развития зародыша его клетки все больше и больше отличаются друг от друга как по свойствам, так и по строению. Это связано с тем, что в разных клетках развивающегося организма одинаковая по объему генетическая информация реализуется не полностью (вследствие их детерминации и дифференциальной активности генов).
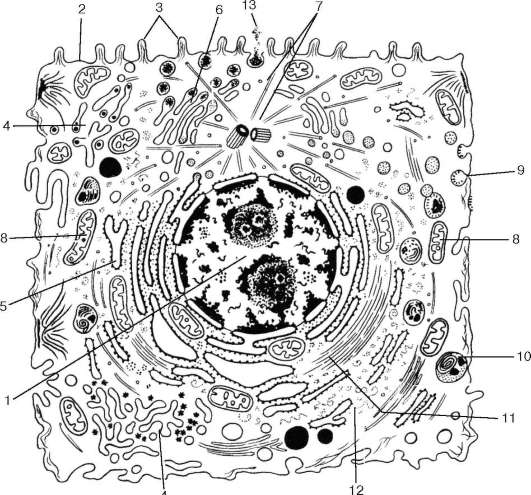
Рис. 4.1. Ультрамикроскопическое строение клетки животных организмов (схема): 1 - ядро; 2 - плазмолемма; 3 - микроворсинки; 4 - агранулярная эндоплазма-тическая сеть; 5 - гранулярная эндоплазматическая сеть; 6 - комплекс Гольджи; 7 - центриоль и микротрубочки клеточного центра; 8 - митохондрии; 9 - цито-плазматические пузырьки; 10 - лизосомы; 11 - микрофиламенты; 12 - рибосомы; 13 - выделение гранул секрета
Индивидуальное развитие от одной клетки до многоклеточного зрелого организма - результат последовательного, избирательного включения работы разных генов в различных клетках. Это приводит к появлению клеток со специфическими для них структурами и особыми функциями, к процессу, называемому дифференцировкой. Дифференцировка обусловлена активностью разных генов в разных клетках, проявляемой по мере развития многоклеточного организма. Другими словами, сходство в строении клеток как данного организма, так и разных организмов определяется сходством общеклеточных функций, направленных на поддержание жизни самих клеток и их размножение. Разнообразие же в строении клеток - это результат их функциональной специализации, дифференцировки в процессе развития.
5. Клетки как части целостного организма. Каждое проявление деятельности целого организма, будь то реакция на раздражение или движение, иммунные реакции и многое другое, осуществляется специализированными клетками различных тканей. Однако, хотя клетка и является единицей функционирования в многоклеточном организме, деятельность ее не обособлена от других клеток и от межклеточного вещества. Специализированные клетки объединяются в системы тканей и органов, подчиненные и связанные межклеточными, тканевыми, гуморальными и нервными формами регуляции. Вот почему мы говорим об организме как о целом, а о клетках - как об элементарных единицах живого, специализированных на выполнении строго определенных функций, осуществляющих их в комплексе со всеми элементами, входящими в состав сложно организованной системы многоклеточного организма.
4.2. СТРУКТУРНЫЕ КОМПОНЕНТЫ КЛЕТКИ 4.2.1. Цитоплазма
Цитоплазма (cytoplasma), часть клетки, отделенная от окружающей среды плазмолеммой, включает гиалоплазму и находящиеся в ней обязательные клеточные компоненты - органеллы, а также различные непостоянные структуры - включения.
Гиалоплазма
Гиалоплазма (от греч. hyalinos - прозрачный), или матрикс цитоплазмы, представляет собой очень важную часть клетки, ее истинную внутреннюю
среду.
В электронном микроскопе матрикс цитоплазмы имеет вид гомогенного или тонкозернистого вещества с низкой электронной плотностью. Гиалоплазма - студнеобразная коллоидная система. Эта система способна переходить из золеобразного (жидкого) состояния в гелеобразное и обратно.
В организованной, упорядоченной многокомпонентной системе гиалоплаз-мы отдельные зоны могут менять свое агрегатное состояние в зависимости от условий или от функциональной задачи; в бесструктурной на взгляд гиа-лоплазме могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых молекул. В состав гиалоплазмы входят главным образом различные глобулярные белки. Они составляют 20-25 % общего содержания белков в эукариотической клетке. К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и других важных соединений. В гиалоплаз-ме располагаются ферменты активации аминокислот при синтезе белков, транспортные (трансферные) рибонуклеиновые кислоты (тРНК). В гиало-плазме при участии рибосом и полирибосом (полисом) происходит синтез белков, необходимых для собственно клеточных нужд, для поддержания и обеспечения жизни данной клетки. Осмотические и буферные свойства клетки в значительной степени определяются составом и структурой гиало-плазмы. Важнейшая роль гиалоплазмы заключается в том, что эта полужидкая среда объединяет все клеточные структуры и обеспечивает химическое взаимодействие их друг с другом. Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов: перенос аминокислот, жирных кислот, нуклеотидов, сахаров. В ней идет постоянный поток ионов к плазматической мембране и от нее к митохондриям, к ядру и вакуолям. В гиалоплазме происходит анаэробный синтез аденозинтрифосфата (АТФ) - гликолиз. Она является основным вместилищем и зоной перемещения массы молекул АТФ. В гиалоплазме происходит отложение запасных продуктов: гликогена, жировых капель, некоторых пигментов.
4.2.2. Органеллы
Органеллы - постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции.
Классификация органелл. Различают мембранные и немембранные органеллы. Мембранные органеллы представлены цитоплазматической сетью (эндо-плазматическим ретикулумом), комплексом Гольджи (аппаратом Гольджи), митохондриями, лизосомами, пероксисомами. К немембранным органеллам относят рибосомы (полирибосомы), клеточный центр и элементы цитоскелета (микротрубочки, микрофиламенты и промежуточные филаменты).
Мембранные органеллы
Структурно-химическая характеристика мембран клеток
К клеточным мембранам относятся плазмолемма, ядерная оболочка, мембраны митохондрий, эндоплазматической сети, комплекса Гольджи, лизосом и перок-сисом. Общей чертой всех мембран клетки является то, что они представляют собой тонкие (6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками) (рис. 4.2).
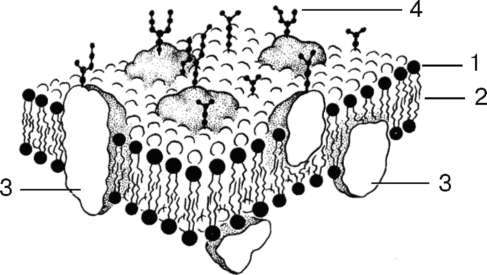
Рис. 4.2. Строение клеточной мембраны (схема);
1 - липиды; 2 - гидрофобная зона би-слоя липидных молекул; 3 - интегральные белки мембраны; 4 - полисахариды гликокаликса
Основными химическими компонентами клеточных мембран являются липиды (40 %) и белки (60 %); кроме того, во многих мембранах обнаружены углеводы (5-10 %).
К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях и жирах (липофиль-ность). Состав липидов в разных мембранах неодинаков. Например, плазматическая мембрана в отличие от мембран эндоплазматической сети и митохондрий обогащена холестерином. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфо-липиды (глицерофосфатиды), сфингомиелины и из стероидных липидов - холестерин.
Особенностью липидов является разделение их молекул на две функционально различные части: гидрофобные неполярные, не несущие зарядов («хвосты»), состоящие из жирных кислот, и гидрофильные, заряженные полярные «головки». Это определяет способность липидов самопроизвольно образовывать двухслойные (билипидные) мембранные структуры толщиной 5-7 нм.
Мембраны различаются и набором белковых молекул. Многие мембранные белки состоят из двух частей - участков, богатых полярными (несущими заряд) аминокислотами, и участков, обогащенных неполярными аминокислотами: глицином, аланином, валином, лейцином. Такие белки в липидных слоях мембран располагаются так, что их неполярные участки погружены в «жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная (гидрофильная) же часть этих белков взаимодействует с головками липидов и обращена в сторону водной фазы. Эти белки пронизывают мембрану, их называют интегральными белками мембран. Кроме интегральных белков, существуют белки, частично встроенные в мембрану, - полуинтегральные и примембранные, не встроенные в билипидный слой. По биологической роли белки мембран можно разделить на белки-ферменты, белки-переносчики, рецепторные и структурные белки.
Углеводы мембран входят в их состав не в свободном состоянии, они связаны с молекулами липидов или белков. Такие вещества называются соответственно гликолипидами и гликопротеидами. Как бы ни было велико различие между мембранами по количеству и составу их липидов, белков и углеводов, мембраны обладают рядом общих свойств, определяемых их основной структурой. Все мембраны являются барьерными структурами,
резко ограничивающими свободную диффузию веществ между цитоплазмой и средой, с одной стороны, и между матриксом и содержимым мембранных органелл - с другой.
Особенность же специфических функциональных нагрузок каждой мембраны определяется свойствами и особенностями белковых компонентов, большая часть которых представляет собой ферменты или ферментные системы. Важную роль в функционировании мембран играют гликолипиды и гликопротеиды надмембранного слоя.
Плазматическая мембрана. Барьерно-рецепторная и транспортная системы клетки
Плазматическая мембрана, или плазмолемма (membrana cellularis), среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку.
Химический состав плазматической мембраны. Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран.
Снаружи от плазмолеммы располагается надмембранный слой - глико-каликс (glycocalyx). Толщина этого слоя около 3-4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гли-копротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы (см. рис. 4.2). При использовании специальных методов выявления полисахаридов (краситель рутениевый красный) показано, что они образуют структуру, подобную «чехлу» поверх плазматической мембраны.
В гликокаликсе могут располагаться белки, не связанные с билипидным слоем. Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др.
Функции плазматической мембраны - это разграничение цитоплазмы с внешней средой, рецепции и транспорта различных веществ как внутрь клетки, так и из нее.
Рецепторные функции связаны с локализацией на плазмолемме специальных структур, участвующих в специфическом «узнавании» химических и физических факторов. Клеточная поверхность обладает большим набором компонентов - рецепторов, определяющих возможность специфических реакций с различными агентами. Рецепторами на поверхности клетки могут служить гликопротеиды и гликолипиды мембран (см. рис. 4.2). Считается, что такие чувствительные к отдельным веществам участки могут быть разбросаны по всей поверхности клетки или собраны в небольшие зоны. Существуют рецепторы биологически активных веществ - гормонов, медиаторов, специфических антигенов разных клеток или белков и др.
С плазмолеммой связана локализация специфических рецепторов, отвечающих за такие важные процессы, как взаимное распознавание клеток и развитие иммунитета. Так, в составе плазмолеммы всех клеток находится молекула гистосовместимости I класса (гликопротеин), которая состоит из: а) интегрального трансмембранного белка, часть которого находится в цитоплазме, другая - пронизывает плазмолемму, и последняя, наиболее длинная часть молекулы, находится в гликокаликсе; б) периферического мембранного белка с малой молекулярной массой; в) короткой молекулы белка, которая нековалентно связывается с петлями внеклеточной части интегрального трансмембранного белка. Именно последняя часть молекулы (пептид из 9 аминокислот) является фрагментом нормального белка клетки данной особи. Он и распознается как «свой» клетками иммунной системы человека. В случае мутации на месте белка гистосовместимости появляется белок с иной структурой молекулы (например, кодируемый вирусом), и в ответ на это возникает иммунная реакция со стороны организма, направленная на уничтожение данной клетки. Этот механизм сохраняет генетическую индивидуальность клеток, следовательно, и организма.
В плазмолемме светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что, в свою очередь, приводит к генерации электрического импульса.
Выполняя транспортную функцию, плазмолемма обеспечивает пассивный перенос ряда веществ, например воды, ряда ионов и некоторых низкомолекулярных соединений. Другие вещества проникают через мембрану путем активного переноса против градиента концентрации с затратой энергии за счет расщепления АТФ. Так транспортируются многие органические молекулы (сахара, аминокислоты и др.). Эти процессы могут быть сопряжены с транспортом ионов, в них участвуют белки-переносчики.
Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процесса эндо-цитоза (рис. 4.3). Эндоцитоз формально разделяют на фагоцитоз (захват и
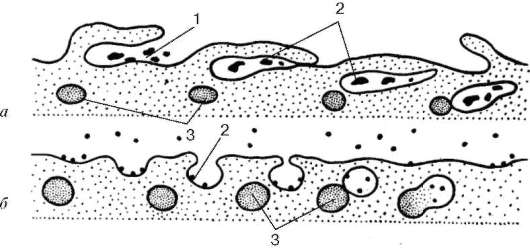
Рис. 4.3. Эндоцитоз. Разные типы образования пиноцитозных пузырьков (а, б):
1 - сорбция частиц на поверхности плазматической мембраны; 2 - погружение
частиц в цитоплазму; 3 - первичные лизосомы
поглощение клеткой крупных частиц, например, бактерий или фрагментов других клеток) и пиноцитоз (захват отдельных молекул и макромолекуляр-ных соединений).
Пиноцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
В дальнейшем эндоцитозные пузырьки, или эндосомы, могут сливаться друг с другом, расти, и в их внутренней полости, кроме поглощенных веществ, начинают обнаруживаться гидролитические ферменты (гидролазы), поступающие сюда из лизосом (см. ниже). Эти ферменты расщепляют биополимеры до мономеров, которые в результате активного транспорта через мембрану пузырька переходят в гиалоплазму. Таким образом, поглощенные молекулы внутри мембранных вакуолей, образовавшихся из элементов плазмолеммы, подвергаются внутриклеточному пищеварению.
При фагоцитозе клетка, например макрофаг, после того, как к ее плазмо-лемме прикрепляется бактерия, формирует длинные цитоплазматические отростки, которые охватывают бактерию, и макрофаг постепенно поглощает бактерию с образованием фагосомы.
Плазмолемма принимает участие в выведении веществ из клетки (экзоци-тоз). В этом случае внутриклеточные продукты (белки, мукополисахариды, липопротеиды и др.), заключенные в вакуоли или пузырьки и отграниченные от гиалоплазмы мембраной, подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются, и содержимое вакуоли поступает в окружающую среду.
Процесс эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы, таких как микротрубочки и сократимые микрофиламенты. Последние, соединяясь с определенными участками плазмолеммы, могут, изменяя свою длину, втягивать мембрану внутрь клетки, что приводит к отделению от плазмолеммы эндоцитозных вакуолей. Часто, непосредственно примыкая к плазмолемме, микрофиламенты образуют сплошной кортикальный слой.
Плазмолемма многих клеток животных может образовывать выросты. У ряда клеток такие выросты включают в свой состав специальные компоненты цитоплазмы (микротрубочки, фибриллы), что приводит к развитию органелл движения - ресничек, жгутиков и др.
Наиболее часто встречаются на поверхности многих животных клеток микроворсинки. Это выросты цитоплазмы, ограниченные плазмолеммой, имеющие форму цилиндра с закругленной вершиной. Микроворсинки характерны для клеток эпителия, но обнаруживаются и у клеток других тканей. Диаметр микроворсинок около 100 нм. Число и длина их различны у разных типов клеток. Возрастание числа микроворсинок приводит к резкому увеличению площади клеточной поверхности. Это особенно важно для клеток, участвующих во всасывании. Так, в кишечном эпителии на
1 мм2 поверхности насчитывается до 2?108 микроворсинок.
Межклеточные соединения
Плазмолемма принимает активное участие в образовании специальных структур - межклеточных контактов, или соединений (junc-tiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 4.4).
Общим для этих клеток является то, что на их поверхности располагаются специальные углеводные части интегральных белков, глико-протеидов, которые специфически взаимодействуют и соединяются с соответствующими белками на поверхности соседних клеток.
Межклеточные соединения делятся на простые и сложные.
Простое межклеточное соединение ( junctio intercellularis simplex) - сближение плазмолемм соседних клеток на расстояние 15-20 нм (рис. 4.5). При этом происходит взаимодействие слоев гликокалик-са соседних клеток. С помощью
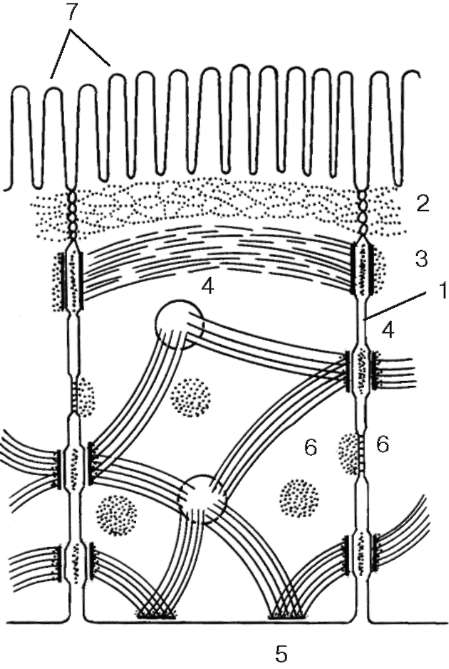
Рис. 4.4. Расположение различных межклеточных соединений в клетках кишечного эпителия (схема):
1 - простое соединение; 2 - плотное соединение (замыкающая зона); 3 - адгезивный поясок (поясок слипания); 4 - десмосома (пятно сцепления); 5 - полудес-мосома; 6 - щелевое (коммуникационное) соединение; 7 - микроворсинки
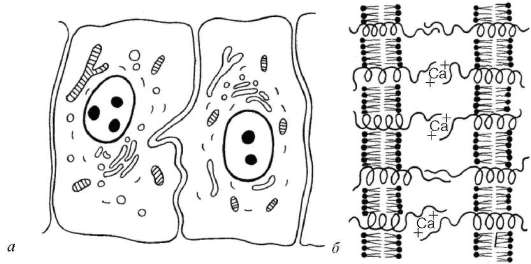
Рис. 4.5. Простое межклеточное соединение (схема):
а - простое соединение двух эпителиальных клеток; б - связывание интегральными гликопротеидами (интегринами и кадгеринами) плазматических мембран соседних клеток
белков-рецепторов, входящих в состав гликокаликса, происходит распознавание клеток, возникших из общего зачатка, и объединение их в пласты. Например, Е-кадгерины участвуют в образовании контактов только между эпителиальными клетками, обеспечивая их соединение практически по всей поверхности контактирующих клеток.
Сложные межклеточные соединения представляют собой небольшие парные специализированные участки плазматических мембран двух соседних клеток. Они подразделяются на замыкающие (изолирующие), сцепляющие (заякоривающие) и коммуникационные (объединяющие) соединения.
К замыкающим относится плотное соединение (замыкающая зона - zonu-la occuludens). В этом соединении принимают участие специальные интегральные белки, расположенные на поверхности соседних клеток, образующие подобие ячеистой сети (рис. 4.6).
Эта ячеистая сеть окружает в виде пояска весь периметр клетки, соединяясь с такой же сетью на поверхности соседних клеток. Данная область непроницаема для макромолекул и ионов, и, следовательно, она замыкает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Этот тип соединений характерен для клеток однослойных эпителиев и эндотелия некоторых сосудов.
К сцепляющим, или заякоривающим, соединениям относятся адгезивный поясок (поясок слипания) и десмосома. Общим для этой группы соединений является то, что к участкам плазматических мембран со стороны цитоплазмы подходят фибриллярные элементы цитоскелета (актиновые филаменты,
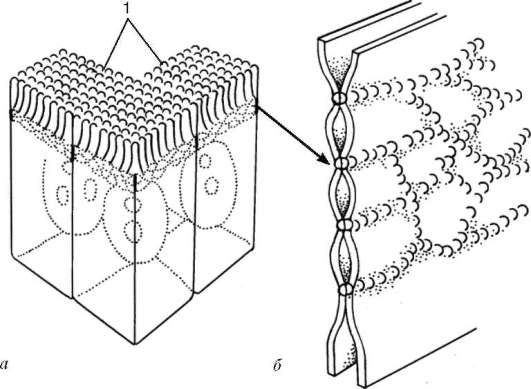
Рис. 4.6. Плотное соединение (замыкающая зона):
а - расположение плотного соединения (вставочная пластинка) на клетках кишечного эпителия; б - трехмерная схема участка плотного соединения. 1 - микроворсинки
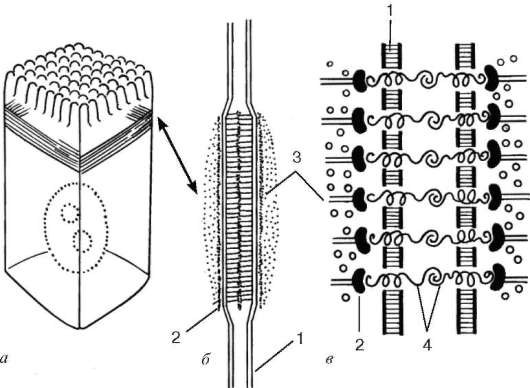
Рис. 4.7. Адгезивный поясок (поясок слипания):
а - расположение его в клетке; б - вид на срезе; в - схема молекулярной организации. 1 - плазмолемма; 2 - слой белков сцепления; 3 - актиновые микрофиламенты; 4 - связующие гликопротеиды
промежуточные филаменты и спектрин) и связываются с мембранами в области соединения соседних клеток.
Адгезивный поясок, или поясок слипания (zonula adherens), - парное образование в виде лент, каждая из которых опоясывает апикальные части соседних клеток и обеспечивает в этой области их прилипание друг к другу (рис. 4.7). Здесь клетки связаны друг с другом интегральными гликопротеи-дами, к которым со стороны цитоплазмы и той и другой клетки примыкает слой примембранных белков, включающих характерный белок винкулин. К этому слою подходит и связывается с ним пучок актиновых микрофила-ментов. Взаимодействие актиновых микрофиламентов с актинсвязывающи-ми белками во многих соседствующих клетках может привести к изменению рельефа всего эпителиального пласта.
К сцепляющим соединениям может быть отнесен фокальный контакт, характерный для фибробластов. В этом случае клетка соединяется не с соседней клеткой, а с элементами внеклеточного субстрата. В образовании фокального контакта также принимают участие актиновые микрофила-менты. К сцепляющим межклеточным соединениям относятся и десмосомы (рис. 4.8).
Десмосома, или пятно слипания (macula adherens). Это парные структуры, представляющие собой небольшую площадку или пятно диаметром около 0,5 мкм. Со стороны цитоплазмы к плазматической мембране прилежит слой белков, в состав которого входят десмоплакины. В этот слой со стороны цитоплазмы внедряются пучки промежуточных филаментов. С внешней стороны плазмолеммы соседних клеток в области десмосом соединяются
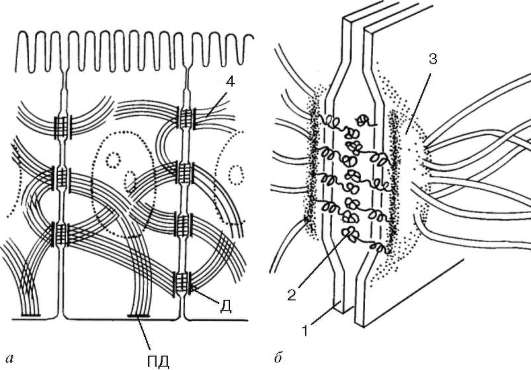
Рис. 4.8. Десмосома:
а - расположение в клетке; б - схема ультраструктуры. 1 - плазмолемма; 2 - десмо-глеиновый слой; 3 - слой десмоплакина; 4 - промежуточные филаменты. Д - десмосома; ПД - полудесмосома
с помощью трансмембранных белков - десмоглеинов. Например, каждая клетка эпидермиса кожи может иметь до нескольких сотен десмосом.
Функциональная роль десмосом заключается главным образом в механической связи между клетками. Десмосомы связывают друг с другом клетки в различных эпителиях, в сердечной и гладких мышцах. Полудесмосомы связывают эпителиальные клетки с базальной мембраной.
Коммуникационные соединения в клетках животных представлены щелевыми соединениями и синапсами (рис. 4.9).
Щелевое соединение, или нексус (nexus), представляет собой область протяженностью 0,5-3 мкм, где плазмолеммы разделены промежутком в 2-3 нм (см. рис. 4.9). Со стороны цитоплазмы никаких специальных примембран-ных структур в данной области не обнаруживается, но в структуре плазмо-лемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей.
Функциональная роль щелевого соединения заключается в переносе ионов и мелких молекул (молекулярная масса 2?103) от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексусы.
Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы).
Синапсы - участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому (см. главу 10).
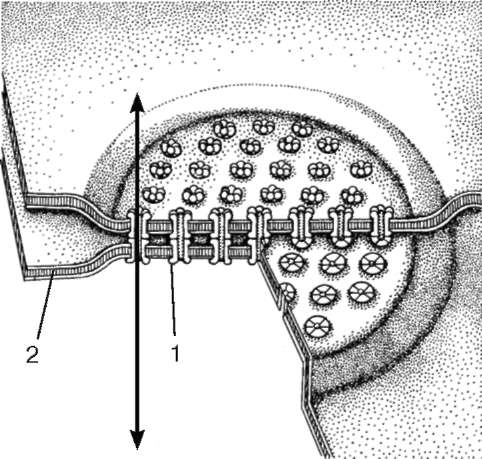
Рис. 4.9. Щелевое (коммуникационное) соединение:
1 - коннексон; 2 - плазмолемма
Вакуолярная система
Эндоплазматическая сеть
Эндоплазматическая сеть (эндо-плазматический ретикулум) была открыта К. Р. Портером в 1945 г. Этот компонент вакуолярной системы клетки представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих трехмерную мембранную сеть. В состав сети входят гранулярные и агранулярные участки, которые могут чередоваться.
Гранулярная эндоплазматическая сеть (reticulum endoplasmicum granulosum) на ультратонких срезах представлена замкнутыми мембранами, которые на сечениях образуют уплощенные мешки, цистерны, трубочки.
Диаметр цистерн значительно варьирует и в зависимости от функциональной активности клетки колеблется от 20 нм до несколько микрометров. Отличительной чертой мембран гранулярной эндоплазматической сети является то, что они со стороны гиалоплазмы покрыты многочисленными рибосомами (рис. 4.10).
Гранулярная эндоплазматическая сеть имеет разное строение. Для малоспециализированных клеток или для клеток с низкой метаболической активностью характерно наличие редких и разрозненных цистерн. Если возникают локальные скопления гранулярной эндоплазматической сети, то это свидетельствует об активном синтезе секреторных белков. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазмати-ческая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков - ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. Внутри канальцев или вакуолей гранулярной эндоплазматической
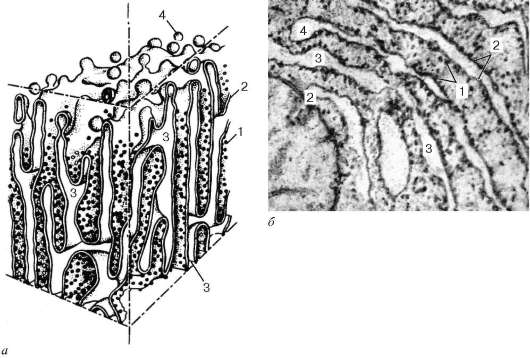
Рис. 4.10. Строение гранулярной эндоплазматической сети: а - схема; б - электронная микрофотография участка среза эпителиальной клетки печени. 1 - рибосомы; 2 - пластинки; 3 - внутренние полости цистерн; 4 - отщепляющиеся мембранные пузырьки, лишенные рибосом
сети происходит модификация белков, например, связывание их с сахарами (первичное глюкозилирование).
В гранулярной эндоплазматической сети на ее рибосомах происходит синтез мембранных интегральных белков, которые встраиваются в толщу мембраны. Здесь же со стороны гиалоплазмы идет синтез липидов и их встраивание в мембрану. В результате этих двух процессов наращиваются сами мембраны эндоплазматической сети и другие компоненты вакуоляр-ной системы клетки.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли, трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50-100 нм. Гладкая эндоплаз-матическая сеть возникает и развивается на основе гранулярной эндоплаз-матической сети. В отдельных участках гранулярной эндоплазматической сети образуются новые липопротеидные мембранные участки, лишенные рибосом. Эти участки могут разрастаться, отщепляться от гранулярных мембран и функционировать как самостоятельная вакуолярная система.
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды, например, в эндокринных клетках коркового вещества надпочечников, в эпителиальных клетках извитых семенных канальцев.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматиче-ская сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани (см. главу 9).
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при некоторых отравлениях в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь заполненные гладким эндо-плазматическим ретикулумом.
Комплекс Гольджи
Комплекс Гольджи (пластинчатый комплекс) был открыт в 1898 г. К. Гольджи. Автор, используя свойства связывания тяжелых металлов (осмия или серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом (apparatus reticularis internus). В дальнейшем его стали называть аппаратом, или комплексом Гольджи (complexus Golgiensis). Подобные структуры затем были описаны во всех клетках эукариот.
При рассмотрении в электронном микроскопе комплекс Гольджи представлен мембранными структурами, собранными вместе в небольших зонах
(рис. 4.11).
Отдельная зона скопления этих мембран называется диктиосомой (стопкой Гольджи). Таких зон в клетке может быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми находятся тонкие прослойки гиалоплазмы. Каждая цистерна имеет переменную толщину: в центре ее мембраны могут быть сближены (до 25 нм), а на периферии иметь расширения - ампулы, ширина которых непостоянна. Кроме плотно расположенных плоских цистерн, в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул), которые встречаются главным образом в его периферических участках. Иногда они отшнуровываются от ампулярных расширений на краях плоских цистерн. В зоне диктиосомы различают проксимальную (cis) и дистальную (trans) поверхности. В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная поверхность обращена к ядру, в то время как дистальная - к поверхности клетки.
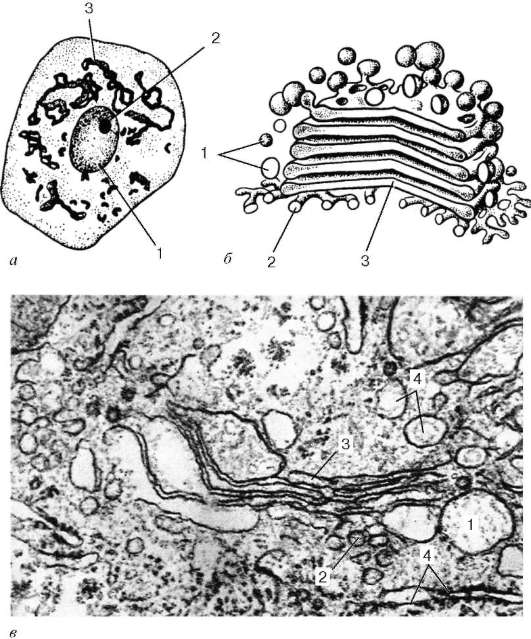
Рис. 4.11. Комплекс Гольджи:
а - нервная клетка спинного мозга, импрегнация серебром по методу Гольджи: 1 - ядро; 2 - ядрышко; 3 - комплекс Гольджи; б - схема ультрамикроскопического строения (трехмерная реконструкция); в - комплекс Гольджи на ультратонком срезе (печеночная клетка): 1 - пузырьки; 2 - трубочки; 3 - уплощенные мешочки (цистерны); 4 - фрагменты гранулярной эндоплазматической сети
В клетках отдельные диктиосомы могут быть связаны друг с другом системой везикул и цистерн, примыкающих к дистальной поверхности, так что образуется рыхлая трехмерная сеть, выявляемая в световом и электронном микроскопах («транс-сеть» комплекса Гольджи).
Комплекс Гольджи участвует в сегрегации и накоплении продуктов, синтезированных в эндоплазматической сети, в их химической перестройке, созревании; в его цистернах происходят синтез полисахаридов, их ком-плексирование с белками, что приводит к образованию пептидоглика-нов. С помощью комплекса Гольджи осуществляется процесс выведения готовых секретов за пределы секреторной клетки. Кроме того, комплекс
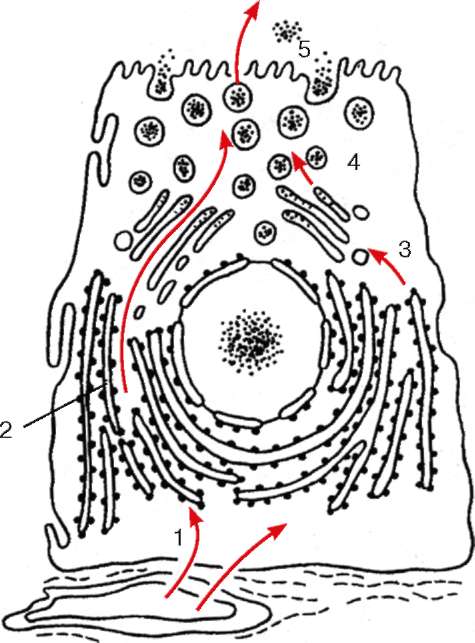
Рис. 4.12. Участие клеточных структур в белковой секреции (схема): 1 - поступление аминокислот из гемо-капилляра к рибосомам гранулярной эндоплазматической сети; 2 - синтез и сегрегация белков; 3 - переход белков в вакуоли комплекса Гольджи; 4 - отщепление от комплекса Гольджи пузырьков с секреторными продуктами; 5 - выход секрета из клетки
Гольджи обеспечивает формирование лизосом. Мембраны комплекса образуются путем отщепления мелких вакуолей от гранулярного эндоплазматического ретикулума. Эти вакуоли поступают в проксимальный отдел комплекса Гольджи, где и сливаются с его мембранами. Следовательно, в комплекс Гольджи поступают новые порции мембран и продуктов, синтезированных в гранулярном эндоплазма-тическом ретикулуме. В мембранных цистернах комплекса Гольджи происходят вторичные изменения в структуре белков, синтезированных в гранулярном эндоплазматическом ретикулуме. Эти изменения (модификации) связаны с перестройкой олигосахаридных цепочек синтезированных гликопротеидов. Внутри полостей комплекса Гольджи с помощью различных ферментов (транс-глюкозидаз) по разному модифицируются лизосомные белки и белки секретов: происходят последовательная замена и наращивание олигосахаридных цепочек. Модифицирующиеся белки переходят от цистерны проксимальной цис-поверхности в цистерны дистальной поверхности путем эстафетного переноса мелких вакуолей, содержащих транспортируемый белок.
В цистернах дистальной (trans) поверхности происходит сортировка белков: на внутренних поверхностях мембран цистерн располагаются рецепторы, узнающие или секреторные белки, или белки, входящие в состав лизосом (гидролазы). В результате от цистерн дистальной поверхности дик-тиосом отщепляются два типа мелких вакуолей: а) содержащие гидролазы - лизосомы (первичные); б) секреторные белки.
Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах белок, накапливающийся внутри цистерн эндо-плазматической сети, транспортируется далее в вакуоли комплекса Гольджи (рис. 4.12).
Затем накопленный белок может конденсироваться, образуя секреторные белковые продукты (как это, например, наблюдается в поджелудочной, молочной и других железах). От ампулярных расширений цистерн комплек-
са Гольджи отщепляются пузырьки, содержащие эти белки. В дальнейшем они могут сливаться друг с другом и эндосомами и увеличиваться в размерах, образуя секреторные гранулы. После этого секреторные гранулы начинают двигаться к поверхности клетки, соприкасаются с плазмолеммой, с которой сливаются их собственные мембраны, и таким образом содержимое гранул оказывается за пределами клетки. Морфологически этот процесс называется экструзией (выбрасывание, экзоцитоз) и напоминает пиноцитоз только с обратной последовательностью стадий.
Нужно отметить, что с самого момента образования до выведения из клеток секретируемые продукты отделены мембраной от гиалоплазмы. Следовательно, мембраны комплекса Гольджи выполняют сегрегирующую роль при образовании клеточных секретов. В вакуолях комплекса Гольджи иногда происходят накопление ресинтезированных молекул липидов и образование сложных белков - липопротеидов, которые могут транспортироваться вакуолями за пределы клетки. Вакуоли комплекса Гольджи дают начало лизосомам.
Лизосомы
Лизосомы (lysosomae) - это разнообразный класс вакуолей размером 0,2- 0,4 мкм, ограниченных одиночной мембраной. Характерным признаком лизосом является наличие в них гидролитических ферментов - гидролаз (протеиназы, нуклеазы, фосфатазы, липазы и др.), расщепляющих различные биополимеры при кислом значении рН. Лизосомы были открыты в 1949 г. де Дювом.
Кроме собственно лизосом (первичных) различают аутофаголизосомы, или гетеролизосомы (вторичные лизосомы), и телолизосомы (остаточные тельца) (рис. 4.13).
Разнообразие морфологии лизосом объясняется тем, что эти частицы участвуют в процессах внутриклеточного переваривания, образуя сложные пищеварительные вакуоли как экзогенного (внеклеточного), так и эндогенного (внутриклеточного) происхождения.
Лизосомы (первичные) представляют собой мелкие мембранные пузырьки размером около 0,2-0,5 мкм, заполненные бесструктурным веществом, содержащим гидролазы, в том числе активную кислую фосфатазу, которая является маркерным ферментом для лизосом. Эти мелкие пузырьки практически очень трудно отличить от мелких везикул на периферии зоны комплекса Гольджи, которые также содержат кислую фосфатазу. Местом ее синтеза является гранулярная эндоплазматическая сеть. Затем этот фермент появляется в цистернах проксимальной поверхности диктиосомы, а затем в мелких везикулах по периферии диктиосомы и, наконец, в лизосомах. Таким образом, весь путь образования лизосом очень сходен с образованием секреторных (зимогенных) гранул в клетках поджелудочной железы, за исключением последнего этапа.
Гетерофаголизосомы (вторичные лизосомы), или внутриклеточные пищеварительные вакуоли, формируются при слиянии лизосом с фагоцитарными или пиноцитозными вакуолями. Если происходит слияние лизосомы
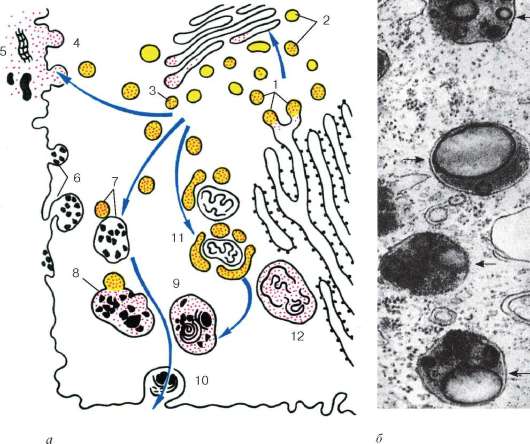
Рис. 4.13. Строение лизосом:
а - схема участия структур клетки в образовании лизосом и во внутриклеточном пищеварении: 1 - образование из гранулярной эндоплазматической сети мелких пузырьков, содержащих гидролитические ферменты; 2 - перенос ферментов в комплекс Гольджи; 3 - образование первичных лизосом; 4 - выделение и использование (5) гидролаз при внеклеточном расщеплении; 6 - эндоцитозные пузырьки; 7 - слияние первичных лизосом и эндоцитозных пузырьков; 8 - образование вторичных лизосом; 9 - телолизосомы; 10 - экскреция остаточных телец; 11 - слияние первичных лизосом с разрушающимися структурами клетки; 12 - аутофаголизосома; б - электронная микрофотография среза гетерофаголизо-сом (обозначены стрелками)
с измененными органеллами самой клетки, то такая структура называется аутофаголизосома. При этом ферменты лизосомы получают доступ к субстратам, которые они и начинают расщеплять. Вещества, попавшие в состав гетероили аутофаголизосом (вторичных лизосом), расщепляются гидролазами до мономеров, которые транспортируются через мембрану лизосо-мы в гиалоплазму, где они реутилизируются, т. е. включаются в различные обменные процессы.
Однако расщепление, переваривание макромолекул лизосомой может идти в ряде клеток не до конца. В этом случае в вакуолях лизосом накапли-
ваются непереваренные продукты. Такая органелла носит название тело-лизосома, или остаточное тельце (corpusculum residuale). Остаточные тельца содержат меньше гидролитических ферментов, в них происходит уплотнение содержимого, его перестройка. Часто в остаточных тельцах наблюдается вторичная структуризация неперевариваемых липидов, которые образуют слоистые структуры. Там же откладываются пигментные вещества. Например, у человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигмента старения» - липофусцина.
При участии лизосом (аутофаголизосом) может происходить модификация продуктов, которые синтезированы самой клеткой. Так, с помощью лизосомальных ферментов в клетках щитовидной железы гидролизуется тиреоглобулин, что приводит к образованию тиреоидных гормонов, которые затем выводятся в кровеносное русло путем экзоцитоза.
В аутофаголизосомах обнаруживаются фрагменты или даже целые цито-плазматические структуры, например митохондрии, элементы эндоплаз-матической сети, рибосомы, гранулы гликогена и другие, что является доказательством их определяющей роли в процессах внутриклеточного пищеварения.
Функциональное значение аутофагоцитоза еще неясно. Есть предположение, что этот процесс связан с отбором и уничтожением измененных, поврежденных клеточных компонентов. В этом случае лизосомы играют роль внутриклеточных «чистильщиков», убирающих дефектные структуры. Интересно, что в нормальных условиях число аутофаголизосом увеличивается при метаболических стрессах, например при гормональной индукции активности клеток печени. Значительно возрастает число аутофаголизосом при различных повреждениях клеток, в этом случае аутофагоцитозу могут подвергаться целые зоны внутри клеток.
Увеличение числа аутофаголизосом в клетках при патологических процессах - обычное явление.
Пероксисомы
Пероксисомы (peroxysomae) в клетках тканей человека - это небольшие (размером 0,3-1,5 мкм) овальной формы тельца, ограниченные мембраной, содержащие гранулярный матрикс, в центре которого часто видны кри-сталлоподобные структуры, состоящие из фибрилл и трубок (сердцевина). Пероксисомы особенно характерны для клеток печени, почек. Во фракции пероксисом обнаруживаются ферменты окисления аминокислот, при работе которых образуется перекись водорода, а также выявляется фермент катала-за, разрушающий ее. Каталаза пероксисом играет важную защитную роль, так как Н2О2 является токсичным веществом для клетки.
Таким образом, одномембранные органеллы клетки, составляющие вакуолярную систему, обеспечивают синтез и транспорт внутриклеточных биополимеров, продуктов секреции, выводимых из клетки, что сопровождается биосинтезом всех мембран этой системы. Лизосомы и пероксисомы участвуют в деградации экзогенных и эндогенных субстратов клетки.
Митохондрии
Митохондрии (mitochondrial) - энергетическая система клетки, органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Исходя из этого, митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания.
Термин «митохондрия» был введен Бенда в 1897 г. для обозначения зернистых и нитчатых структур в цитоплазме разных клеток. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина - от 1 до 10 мкм. Подсчеты показывают, что количество их в клетках сильно варьирует - от единичных элементов до сотен. Так, в клетке печени они составляют более 20 % общего объема цитоплазмы и содержат около 30-35 % общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны.
Во многих случаях отдельные митохондрии могут иметь гигантские размеры и представлять собой разветвленную сеть - митохондриальный рети-кулум. Так, например, в скелетных мышцах митохондриальный ретикулум представлен множеством разветвленных и гигантских митохондриальных тяжей. Гигантские разветвленные митохондрии встречаются в клетках проксимальных отделов нефронов и др.
Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в сердечной мышце митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика и т. п. Увеличение числа митохондрий в клетках происходит путем деления, или почкования, исходных митохондрий.
Митохондрии ограничены двумя мембранами толщиной около 7 нм (рис. 4.14).
Наружная митохондриальная мембрана (membrana mitochondrialis externa) отделяет их от гиалоплазмы. Обычно она имеет ровные контуры и замкнута, так что представляет собой мембранный мешок. Внешнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя митохондриальная мембрана (membrana mitochondrialis interna) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс (matrix mitochondrialis). Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные выпячивания внутрь митохондрий. Такие выпячивания чаще всего имеют вид плоских гребней, или крист.
Матрикс митохондрий имеет тонкозернистое строение (см. рис. 4.14, б), в нем иногда выявляются тонкие нити (толщиной около 2-3 нм) и гранулы размером около 15-20 нм. Нити матрикса митохондрий представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы.
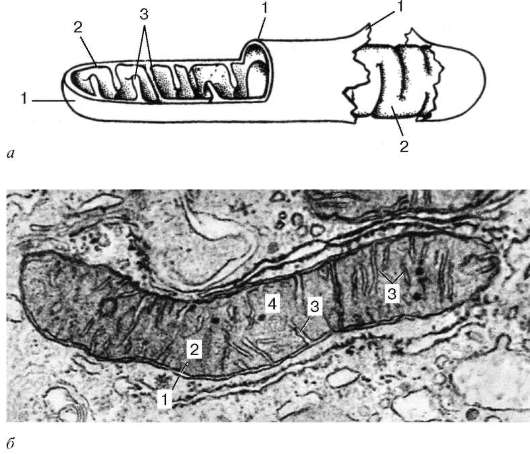
Рис. 4.14. Ультрамикроскопическое строение митохондрии:
а - схема; б - электронная микрофотография среза митохондрии печеночной клетки. 1 - наружная митохондриальная мембрана; 2 - внутренняя митохондриальная мембрана; 3 - кристы; 4 - митохондриальный матрикс
Основная функция митохондрий - синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования аденозиндифосфата (АДФ).
Начальные этапы этих сложных процессов совершаются в гиалоплазме. Здесь происходит первичное окисление субстратов (например, сахаров) до пировиноградной кислоты (пирувата) с одновременным синтезом небольшого количества АТФ. Эти процессы совершаются в отсутствие кислорода (анаэробное окисление, гликолиз). Все последующие этапы выработки энергии - аэробное окисление и синтез основной массы АТФ - осуществляются с потреблением кислорода и локализуются внутри митохондрий. При этом происходит дальнейшее окисление пирувата и других субстратов энергетического обмена с выделением СО2 и переносом протонов на их акцепторы. Эти реакции осуществляются с помощью ряда ферментов так называемого цикла трикарбоновых кислот, которые локализованы в матриксе митохондрии.
В мембранах крист митохондрии располагаются системы дальнейшего переноса электронов и сопряженного с ним фосфорилирования АДФ (окислительное фосфорилирование). При этом происходит перенос электронов от одного белка-акцептора электронов к другому и, наконец, связывание их с кислородом, вследствие чего образуется вода. Одновременно с этим часть
энергии, выделяемой при таком окислении в цепи переноса электронов, запасается в виде макроэргической связи при фосфорилировании АДФ, что приводит к образованию большого числа молекул АТФ - основного внутриклеточного энергетического эквивалента. Именно на мембранах крист митохондрии происходит процесс окислительного фосфорилирования с помощью расположенных здесь белков цепи окисления и фермента фосфо-рилирования АДФ, АТФ-синтетазы.
Выявлено, что в матриксе митохондрии локализуется автономная система митохондриального белкового синтеза. Она представлена молекулами ДНК, свободными от гистонов, что сближает их с ДНК бактериальных клеток. На этих ДНК происходит синтез молекул РНК разных типов: информационных, трансферных (транспортных) и рибосомных. В матрик-се митохондрий наблюдается образование рибосом, отличных от рибосом цитоплазмы. Эти рибосомы участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром. Однако такая система белкового синтеза не обеспечивает всех функций митохондрии, поэтому автономию митохондрий можно считать ограниченной, относительной. Малые размеры молекул митохондриальных ДНК не могут определить синтез всех белков митохондрий. Показано, что подавляющее большинство белков митохондрий находится под генетическим контролем клеточного ядра и синтезируется в цитоплазме. Митохондриальная ДНК кодирует лишь 13 митохондриальных белков, которые локализованы в мембранах и представляют собой структурные белки, ответственные за правильную интеграцию в митохондриальных мембранах отдельных функциональных белковых комплексов.
Митохондрии в клетках могут увеличиваться в размерах и числе. В последнем случае происходит деление перетяжкой или фрагментация исходных крупных митохондрий на более мелкие, которые в свою оче-
редь могут расти и снова делиться. Митохондрии очень чувствительны к изменениям проницаемости мембран, что может приводить к их обратимому набуханию.
Немембранные органеллы
Рибосомы
Рибосомы (ribosomae) - элементарные аппараты синтеза белковых, полипептидных молекул - обнаруживаются во всех клетках (рис. 4.15). Рибосомы - это сложные рибону-клеопротеиды, в состав которых входят белки и молекулы рибосо-мальных РНК (рРНК) примерно в равных весовых отношениях. Размер функционирующей рибосомы эука-
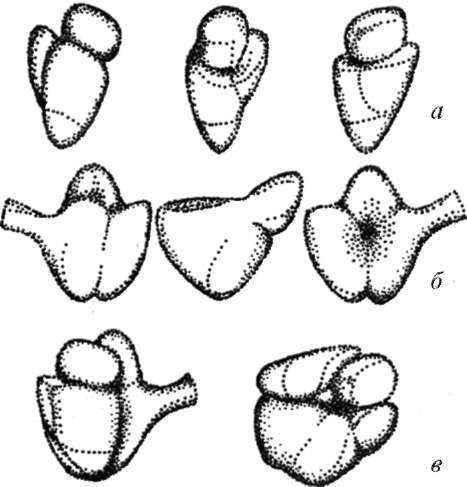
Рис. 4.15. Строение рибосом:
а - малая субъединица; б - большая
субъединица; в - полная рибосома
риотических клеток 25x20x20 нм. Такая рибосома состоит из большой и малой субъединиц. Каждая из субъединиц построена из рибонуклеопро-теидного тяжа, где рРНК взаимодействует с разными белками и образует тело рибосомы.
Различают единичные рибосомы и комплексы рибосом (полисомы). Рибосомы могут располагаться свободно в гиалоплазме или быть связанными с мембранами эндоплазматической сети. В малоспециализированных и быстрорастущих клетках в основном обнаруживаются свободные рибосомы. В специализированных клетках рибосомы располагаются в составе гранулярной эндоплазматической сети. Синтетическая деятельность свободных рибосом направлена в основном на собственные нужды клетки. Связанные рибосомы обеспечивают синтез белков «на экспорт», т. е. на обеспечение нужд организма. Содержание РНК и соответственно степень белковых синтезов коррелируют с интенсивностью базофилии цитоплазмы, т. е. со способностью окрашиваться основными красителями.
Цитоскелет
Цитоскелет (cytoskeleton) - опорно-двигательная система клетки, включающая немембранные белковые нитчатые органеллы, выполняющие как каркасную, так и двигательную функции в клетке. Эти структуры являются динамическими образованиями, они могут быстро возникать в результате полимеризации их элементарных молекул и так же быстро разбираться, исчезать при деполимеризации. К этой системе относятся фибриллярные структуры и микротрубочки.
Фибриллярные структуры цитоплазмы. К фибриллярным компонентам цитоплазмы эукариотических клеток относятся микрофиламенты (micro-filamenti) толщиной 5-7 нм и так называемые промежуточные филаменты (filamenti intermedii) толщиной около 10 нм (рис. 4.16).
Микрофиламенты встречаются практически во всех типах клеток. Они располагаются в кортикальном слое цитоплазмы, непосредственно под плазмолеммой, пучками или слоями. Их можно видеть в псевдоподиях амеб или в движущихся отростках фибробластов, в микроворсинках кишечного эпителия. Микрофиламенты часто образуют пучки, направляющиеся в клеточные отростки.
С помощью иммунофлюоресцентных методов показано, что в состав микрофиламентов кортикального слоя и пучков входят белки: актин, миозин, тропомиозин, альфа-актинин. Следовательно, микрофиламенты не что иное, как внутриклеточный сократительный аппарат, обеспечивающий не только подвижность клеток при активном амебоидном их перемещении, но, вероятно, и большинство внутриклеточных движений, таких как токи цитоплазмы, движение вакуолей, митохондрий, деление клетки. Кроме того, актиновые микрофиламенты играют и каркасную роль. Соединяясь с рядом стабилизирующих белков, они могут образовывать временные или постоянные (как в микроворсинках кишечного эпителия) пучки или сети, играющие большую роль в структурировании цитоплазмы.
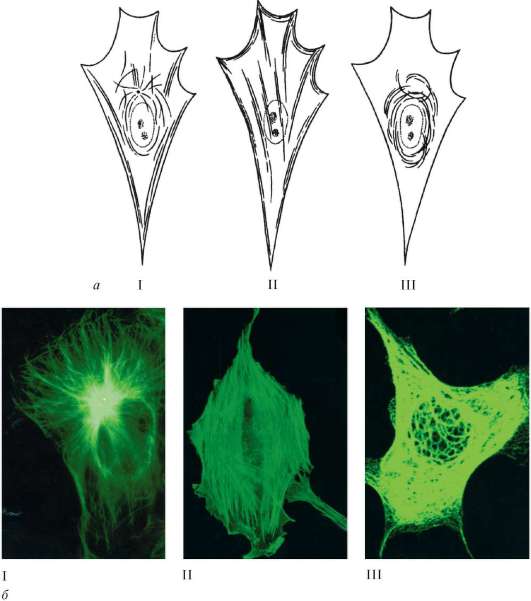
Рис. 4.16. Микрофиламенты и микротрубочки:
а - схема; б - микрофотографии (иммунофлюоресцентный анализ); бI - микротрубочки в культуре клеток фибробластов мыши (тубулин); бII - актиновые микрофиламенты в культуре клеток; бIII - промежуточные филаменты в культуре клеток эмбриональной почки свиньи
Промежуточные филаменты. Это тонкие (10 нм) неветвящиеся, часто располагающиеся пучками нити. Характерно, что в клетках разных тканей их белковый состав различен. Например, в эпителии кожного типа в состав промежуточных филаментов входит кератин. Пучки кератиновых промежуточных филаментов в эпителиальных клетках образуют тонофиламен-ты, которые подходят к десмосомам. В состав промежуточных филаментов
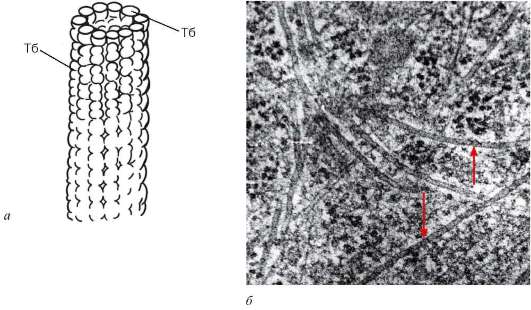
Рис. 4.17. Строение микротрубочек: а - Тб-субъединица, димер тубу-лина в составе микротрубочек; б - микротрубочки в цитоплазме клетки (стрелки)
клеток, производных мезенхимы (например, фибробластов), входит другой белок - виментин; в мышечных клетках обнаруживается десмин; в нервных клетках в состав нейрофиламентов также входит особый белок. Роль промежуточных микрофиламентов, скорее всего, опорно-каркасная; эти фибриллярные структуры не так лабильны, как микротрубочки и микро-филаменты.
В клинике с помощью иммуноморфологических методов тканевое происхождение тех или иных опухолей определяется именно по белкам их промежуточных филаментов. Это очень важно для диагностики и правильного выбора типа химиотерапевтических противоопухолевых препаратов.
Микротрубочки (microtubuli). В клетках микротрубочки принимают участие в создании ряда временных (цитоскелет интерфазных клеток, веретено деления) или постоянных (центриоли, реснички, жгутики) структур.
Микротрубочки представляют собой прямые, неветвящиеся длинные полые цилиндры (рис. 4.17). Их внешний диаметр составляет около 24 нм, внутренний просвет имеет ширину 15 нм, а толщина стенки - 5 нм. Стенка микротрубочек построена за счет плотно уложенных округлых субъединиц диаметром около 5 нм. В электронном микроскопе на поперечных сечениях микротрубочек видны большей частью 13 субъединиц, выстроенных в виде однослойного кольца. Микротрубочки, выделенные из разных источников (реснички простейших, клетки нервной ткани, веретено деления), имеют сходный состав и содержат белки - тубулины.
Очищенные тубулины способны при определенных условиях собираться в микротрубочки с такими же параметрами, какие характерны для микротрубочек внутри клеток. Добавление алкалоида колхицина предотвращает самосборку микротрубочек или приводит к разборке уже существующих. Деполимеризация тубулинов или торможение их полимеризации также вызывается понижением температуры, но после повышения температуры до
37 °С снова происходит самосборка микротрубочек. Деполимеризация тубу-линов и исчезновение микротрубочек происходят и при действии на живую клетку колхицина или охлаждения.
Микротрубочки (цитоскелет) интерфазных клеток. Практически во всех эукариотических клетках в гиалоплазме можно видеть длинные невет-вящиеся микротрубочки. В больших количествах они обнаруживаются в цитоплазматических отростках нервных клеток, фибробластов и других изменяющих свою форму клеток (см. рис. 4.16). Одно из функциональных значений микротрубочек цитоплазмы заключается в создании эластичного, но одновременно устойчивого внутриклеточного каркаса (цитоскелета), необходимого для поддержания формы клетки.
При действии колхицина, вызывающего деполимеризацию тубу-линов, сильно меняется форма клеток. Если отростчатую и плоскую клетку в культуре фибробластов обработать колхицином, то она теряет полярность и сжимается. Точно так же ведут себя другие клетки: колхицин прекращает рост клеток хрусталика, отростков нервных клеток.
Создавая внутриклеточный скелет, микротрубочки могут быть факторами ориентированного движения клетки в целом и ее внутриклеточных компонентов, задавать своим расположением векторы для направленных потоков разных веществ и для перемещения крупных структур. Разрушение микротрубочек колхицином нарушает транспорт веществ в аксонах нервных клеток, приводит к блокаде секреции и т. п.
В аксоне нервной клетки по интерфазным микротрубочкам, как по рельсам, могут передвигаться различные мелкие вакуоли, например синапти-ческие пузырьки, содержащие нейромедиаторы, или митохондрии. Эти перемещения основываются на связи микротрубочек со специальными белками - транслокаторами (динеины и кинезины), которые, в свою очередь, связываются с транспортируемыми структурами. Микротрубочки являются составной частью клеточного центра, ресничек и жгутиков. О роли микротрубочек в делящихся клетках будет сказано далее. Система микротрубочек развивается в связи с центриолью, которая является местом, где происходят начальная полимеризация тубулинов и рост микротрубочек цитоскелета.
Клеточный центр
Клеточный центр (центросома) состоит из центриолей и связанных с ними микротрубочек - центросферы. Термин «центриоли» был предложен Т. Бо-вери в 1895 г. для обозначения очень мелких телец, размер которых находится на границе разрешающей способности светового микроскопа. В некоторых объектах удавалось видеть, что мелкие плотные тельца - центриоли (centriolum) окружены зоной более светлой цитоплазмы, от которой ради-ально отходят тонкие фибриллы. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центриоли часто определяют полярность клеток эпителия и располагаются вблизи комплекса Гольджи.
Тонкое строение центриолей удалось изучить только с помощью электронного микроскопа. Основой строения центриоли являются расположенные по окружности 9 триплетов микротрубочек, образующих таким образом полый цилиндр. Его диаметр составляет около 0,2 мкм, а длина - 0,3- 0,5 мкм (хотя встречаются центриоли, достигающие в длину нескольких микрометров) (рис. 4.18).
Системы микротрубочек центриоли можно описать формулой: (9х3)+0, подчеркивая отсутствие микротрубочек в ее центральной части.
Обычно в интерфазных клетках присутствуют две центриоли - рядом друг с другом, образующие диплосому (diplosoma). В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают материнскую и дочернюю. Обе центриоли сближены, конец дочерней центриоли направлен к поверхности материнской цен-триоли.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый, матрикс. Часто можно обнаружить несколько дополнительных структур, связанных с центриолями: спутники (сателлиты), фокусы схождения микротрубочек, дополнительные микротрубочки, образующие особую зону - центросферу вокруг центриоли.
При подготовке клеток к митотическому делению происходит удвоение центриолей. Этот процесс у различных объектов осуществляется в разное время - в течение синтеза ДНК или после него. Он заключается в том, что две центриоли в диплосоме расходятся и около каждой из них возникает заново по одной новой дочерней, так что в клетке перед делением обнаруживаются две диплосомы, т. е. четыре попарно связанные центриоли. Этот способ увеличения числа центриолей был назван дупликацией. Увеличение
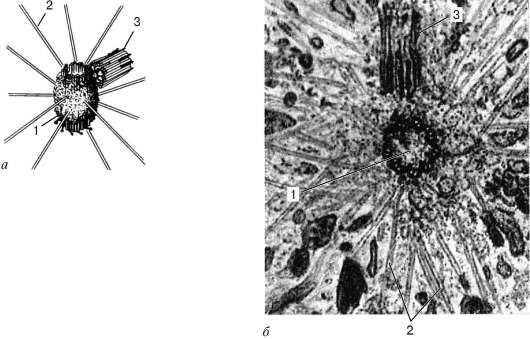
Рис. 4.18. Строение клеточного центра в полюсе митотического веретена клетки:
а - схема; б - электронная микрофотография. 1 - активная материнская центриоль, окруженная тонкофибриллярным матриксом, от которого отходят микротрубочки полярной лучистости (2); 3 - неактивная дочерняя центриоль
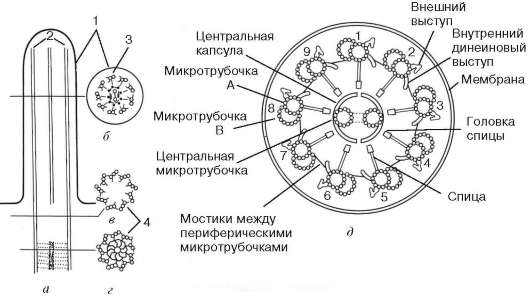
Рис. 4.19. Общее строение реснички:
а - продольный срез; б - поперечный срез тела реснички; в, г - срезы базального тельца. 1 - плазматическая мембрана; 2 - микротрубочки; 3 - дуплеты микротрубочек (А и В); 4 - триплеты микротрубочек базального тельца; д - схема поперечного среза реснички
числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования зачатка, процентриоли, вблизи и перпендикулярно к исходной центриоли.
Центриоли участвуют в индукции полимеризации тубулина при образовании микротрубочек в интерфазе. Перед митозом центриоль является центром полимеризации микротрубочек веретена клеточного деления. Центриоль - центр роста микротрубочек аксонемы ресничек или жгутиков. Наконец, она сама индуцирует полимеризацию тубулинов новой процентриоли, возникающей при ее дупликации.
Реснички и жгутики
Это специальные органеллы движения. В световом микроскопе эти структуры выглядят как тонкие выросты клетки. В основании реснички (fla-gellum) в цитоплазме видны хорошо красящиеся мелкие гранулы - базаль-ные тельца. Длина ресничек составляет 5-10 мкм, а длина жгутиков может достигать 150 мкм (рис. 4.19).
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема («осевая нить») - сложная структура, состоящая в основном из микротрубочек. Проксимальная часть реснички (базальное тельце) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм).
Базальное тельце по своей структуре очень сходно с центриолью. Оно также состоит из 9 триплетов микротрубочек. Часто в основании реснички лежит пара базальных телец, располагающихся под прямым углом друг к другу, подобно диплосоме.
Аксонема (axonema) в своем составе имеет 9 дублетов аксонемальных микротрубочек, образующих стенку цилиндра аксонемы и связанных друг с другом с помощью белковых выростов - «ручек» (см. рис. 4.19). Кроме периферических дублетов микротрубочек, в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9х2)+2 в отличие от (9х3)+0 системы центриолей и базальных телец. Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: два триплета микротрубочек базального тельца, расположенные у апикального полюса клетки под плазмолеммой, связываются с микротрубочками дублетов аксонемы.
Свободные клетки, имеющие реснички и жгутики, обладают способностью двигаться, а неподвижные клетки движением ресничек могут перемещать жидкость и корпускулярные частицы. При движении ресничек и жгутиков длина их не уменьшается, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна. В различных клетках это движение может быть маятникообразным, крючкообразным или волнообразным.
Основной белок ресничек - тубулин - не способен к сокращению и укорочению. Движение ресничек осуществляется за счет активности белка динеина, локализованного в «динеиновых ручках» дублетов микротрубочек. Незначительные смещения дублетов микротрубочек относительно друг друга вызывают изгиб всей реснички. Если такое локальное смещение будет происходить вдоль жгутика, то возникает волнообразное его движение.
Дефекты ресничек могут приводить к различным видам патологии, например, к наследственному рецидивирующему бронхиту и хроническому синуситу, возникающим в результате нарушений функции ресничного эпителия воздухоносных путей и полостей. Дефекты жгутиков встречаются при различных формах наследственного мужского бесплодия.
4.2.3. Включения
Включения цитоплазмы - необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток. Различают трофические, секреторные, экскреторные и пигментные включения. К трофическим включениям относятся капельки нейтральных жиров, которые могут накапливаться в гиалоплазме. В случае недостатка субстратов для жизнедеятельности клетки эти капельки могут постепенно исчезать, включаясь в обменные процессы. Другим видом включений резервного характера является гликоген - полисахарид, откладывающийся также в гиалоплазме (рис. 4.20). Отложение запасных белковых гранул обычно связано с активностью эндоплазматической сети. Так, запасы белка
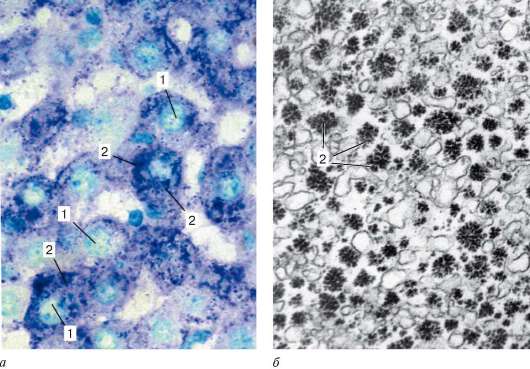
Рис. 4.20. Включения гликогена в клетках печени:
а - окраска - ШИК-реакция: 1 - ядро; 2 - гликоген; б - электронная микрофотография: гликоген в клетках печени
вителлина в яйцеклетках амфибии накапливаются в вакуолях эндоплазма-тической сети.
Секреторные включения - обычно округлые образования различных размеров, содержащие биологически активные вещества, образующиеся в клетках в процессе синтетической деятельности.
Экскреторные включения не содержат каких-либо ферментов или других активных веществ. Обычно это продукты метаболизма, подлежащие удалению из клетки.
Пигментные включения могут быть экзогенными (каротин, пылевые частицы, красители и др.) и эндогенными (гемоглобин, гемосидерин, билирубин, меланин, липофусцин). Наличие их в клетках может изменять цвет ткани и органа временно или постоянно. Нередко пигментация ткани служит одним из диагностических признаков некоторых заболеваний человека или характеризует возрастные изменения тканей и др.
4.2.4. Ядро
Ядро (nucleus) клетки - структура, обеспечивающая хранение и реализацию наследственной (генетической) информации, регуляцию синтеза белков.
Главными структурами, определяющими эти свойства, являются хромосомы, в ДНК которых содержится вся генетическая информация клеток. Хромосомы могут находиться в двух структурно-функциональных состояниях. В неделящихся, интерфазных клетках они находятся в различной степени деконденсации, или в рабочем состоянии, и представляют собой хроматин ядер интерфазных клеток. При делении клеток хроматин максимально уплотняется, конденсируется и образует собственно митотическую хромосому. Интерфазные хромосомы (хроматин) и митотические хромосомы представляют собой в химическом отношении идентичные образования.
Роль ядерных структур в жизнедеятельности клеток
Ядро обеспечивает две группы общих функций: а) хранение и передача генетической информации дочерним клеткам при делении; б) использование генетической информации в процессе синтеза белков.
Хранение и поддержание наследственной информации в виде неизменной структуры ДНК связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекул ДНК. В ядре происходит воспроизведение, или репликация молекул ДНК, что дает возможность при митозе двум дочерним клеткам получить совершенно одинаковые в качественном и количественном отношении объемы генетической информации.
Другой группой клеточных процессов, обеспечиваемых активностью ядра, является создание собственно аппарата белкового синтеза (рис. 4.21). Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК (иРНК), но и транскрипция всех видов транспортных и рибосомных РНК (тРНК, рРНК). В ядре происходит также образование субъединиц рибосом путем комплексирования синтезированных в ядрышке рРНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро.
Таким образом, ядро является не только вместилищем генетического материала, но и местом, где этот материал функционирует и воспроизводится. Вот почему нарушение любой из перечисленных выше функций ядра ведет к гибели клетки.
Структура и химический состав клеточного ядра
Ядро неделящейся (интерфазной) клетки обычно одно на клетку (хотя встречаются и многоядерные клетки). Ядро состоит из хроматина (хромосом), ядрышка, ядерного белкового остова (матрикса), нуклеоплазмы (кариоплазмы) и ядерной оболочки, отделяющей ядро от цитоплазмы (рис. 4.22). Электронно-микроскопически также различают перихроматиновые, меж-хроматиновые, интерхроматиновые гранулы и фибриллы.
Хроматин
При наблюдении живых или фиксированных клеток внутри ядра выявляются зоны плотного вещества, которые хорошо воспринимают разные
красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра получил название «хроматин» (от греч. chroma - цвет, краска). Такими же свойствами хроматина обладают и хромосомы, которые отчетливо видны как плотные окрашивающиеся тельца во время митотического деления клеток. В состав хроматина входит ДНК в комплексе с белками. В неделящихся (интерфазных) клетках хроматин, выявляемый в световом микроскопе, может более или менее равномерно заполнять объем ядра или же располагаться отдельными глыбками. Это связано с тем, что в интерфазном состоянии хромосомы теряют свою компактную форму, разрыхляются, или декон-денсируются. Степень такой декон-
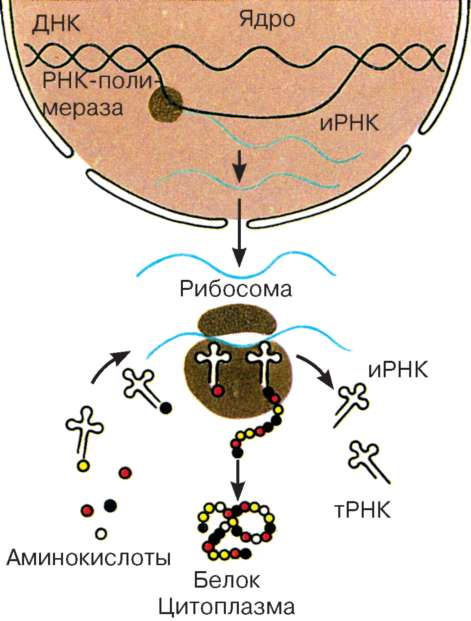
Рис. 4.21. Белковый синтез в клетке (схема)

Рис. 4.22. Ультрамикроскопическое строение ядра интерфазной клетки: 1 - ядерная оболочка (наружная и внутренняя мембраны, перинуклеарное пространство); 2 - комплекс ядерной поры; 3 - гетерохроматин (конденсированный хроматин); 4 - эухроматин (диффузный хроматин); 5 - ядрышко (гранулярная и фибриллярная части); 6 - межхроматиновые гранулы РНК; 7 - перихроматиновые гранулы; 8 - кариоплазма
денсации хромосом может быть различной. Зоны полной деконденсации хромосом и их участков морфологи называют эухроматином (euchromatinum). При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина, называемого гетерохроматином (heterochro-matinum). Степень деконденсации хромосомного материала - хроматина в интерфазе отражает функциональное состояние ядра клетки. Чем больший объем ядра занимает эухроматин, тем интенсивнее в нем протекают синтетические процессы.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных телец - хромосом.
Таким образом, хроматин (хромосомы) клеток может находиться в двух структурно-функциональных состояниях: в активном, рабочем, частично или полностью деконденсированном, когда с его участием в интерфазном ядре происходят процессы транскрипции и репликации ДНК, и в неактивном, в состоянии метаболического покоя и при максимальной их конденсированности, когда они выполняют функцию распределения и переноса генетического материала в дочерние клетки во время деления клеток.
Наблюдения за структурой хроматина с помощью электронного микроскопа показали, что как в препаратах выделенного интерфазного хроматина или выделенных митотических хромосом, так и в составе ядра на ультратонких срезах всегда видны элементарные хромосомные фибриллы толщиной 30 нм.
В химическом отношении фибриллы хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов (ДНП), в состав которых входят ДНК и специальные хромосомные белки - гистоновые и негисто-новые. В составе хроматина обнаруживается также РНК. Количественные отношения ДНК, белка и РНК составляют 1:1,3:0,2. Обнаружено, что длина индивидуальных линейных молекул ДНК может достигнуть сотен микрометров и даже нескольких сантиметров. Среди хромосом человека самая большая первая хромосома содержит молекулу ДНК длиной до 4 см. Суммарная длина молекул ДНК во всех хромосомах одной клетки человека составляет около 170 см, что соответствует массе 6?10-12 г.
В хромосомах существует множество мест независимой репликации, т. е. удвоения ДНК, - репликонов. ДНК эукариотических хромосом представляют собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. В составе генома человека должно встречаться более 50 000 репли-конов или участков ДНК, которые удваиваются как независимые единицы. Синтез ДНК как на участках отдельной хромосомы, так и среди разных хромосом идет неодновременно, асинхронно. Так, например, в некоторых хромосомах человека (1, 3, 16) репликация наиболее интенсивно начинается в плечах хромосом и заканчивается (при высокой интенсивности включения метки) в центромерном районе (см. ниже). Наиболее поздно репликация заканчивается в хромосомах или в их участках, находящихся в компактном (конденсированном) состоянии. Например, поздно реплицируется ДНК
инактивированной Х-хромосомы, которая формирует в ядре клеток женской особи тельце полового хроматина.
На долю белков хроматина приходится 60-70 % сухой массы. К ним относятся гистоны и негистоновые белки. Негистоновые белки составляют лишь 20 % от количества гистонов. Гистоны - щелочные белки, обогащенные основными аминокислотами (главным образом лизином и аргинином). Они обеспечивают специфическую укладку хромосомной ДНК и участвуют в регуляции транскрипции. Гистоны расположены по длине молекулы ДНК в виде блоков (глобул). В один такой блок входят 8 молекул гистонов. Нить ДНК делает около двух оборотов вокруг гистоновых молекул. Весь этот комплекс (ДНК-гистоны) образует нуклеосому. Размер нуклеосомы около 10 нм. При образовании нуклеосом происходит компактизация, или сверхспира-лизация, ДНК, что приводит к укорочению длины хромосомной фибриллы примерно в 7 раз. Между соседними нуклеосомами располагается связующий (линкерный) участок ДНК, который также соединен с молекулой гистона. Таким образом хромосомная фибрилла приобретает вид нити бус или четок, где каждая бусина (нуклеосома) - гистоны, связанные с участком ДНК. Такие нуклеосомные нити толщиной 10 нм дополнительно скручиваются вокруг оси и образуют основную элементарную фибриллу хроматина толщиной 30 нм (рис. 4.23).
В интерфазе фибриллы хроматина образуют петли. Эти петли собраны в розетки, где основания нескольких петель связаны друг с другом негистоно-выми белками ядерного матрикса. Такие петлевые группы (петлевые домены) при падении активности хроматина могут конденсироваться, уплотняться, образуя хромомеры, или хромоцентры, интерфазных ядер. Хромомеры выявляются также в составе митотических хромосом. Хромомеры тесно
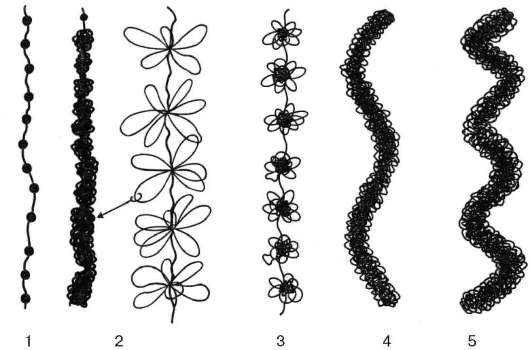
Рис. 4.23. Схема различных уровней компактизации хроматина:
1 - нуклеосомы; 2 - фибрилла толщиной 30 нм; 3 - хромомер, петлевой домен; 4 -
хромонема; 5 - хроматида
располагаются друг за другом и образуют новый фибриллярный уровень компактизации - хромонему. Последняя, далее конденсируясь, формирует основу хроматиды (хромосомы).
Негистоновые белки интерфазных ядер образуют ядерный матрикс, представляющий собой основу, определяющую морфологию и метаболизм ядра. После извлечения ДНК, гистонов, РНК и других растворимых компонентов ядра остается фиброзная ядерная пластинка (ламина), подстилающая ядерную оболочку, и внутриядерная сеть, к которой крепятся фибриллы хроматина.
Функциональная роль ядерного матрикса заключается в поддержании общей формы ядра, в организации не только пространственного расположения в ядре многочисленных и деконденсированных хромосом, но и в организации их активности. На элементах ядерного матрикса располагаются ферменты синтеза РНК и ДНК. Белки ядерного матрикса участвуют в дальнейшей компактизации ДНК в интерфазных и митотических хромосомах.
Хроматин - хромосомы во время митоза
Во время деления клеток интерфазное ядро претерпевает ряд существенных изменений: ядерная оболочка распадается на мелкие вакуоли, а хроматин конденсируется и образует митотические хромосомы.
Морфология митотических хромосом. Каждая хромосома представляет собой фибриллу ДНП, сложно уложенную в относительно короткое тельце - собственно митотическую хромосому. Фибриллы хроматина в митотиче-ской хромосоме образуют многочисленные розетковидные петлевые домены (хромомеры), которые при дальнейшей конденсации хроматина образуют видимую в светооптическом микроскопе митотическую хромосому.
Морфологию митотических хромосом лучше всего изучать в момент их наибольшей конденсации, а именно, в метафазе и в начале анафазы. Хромосомы в этом состоянии представляют собой палочковидные структуры разной длины с довольно постоянной толщиной. У большинства хромосом удается найти зону первичной перетяжки (центромера), которая делит хромосому на два плеча (рис. 4.24).
Хромосомы с равными или почти равными плечами называют мета-центрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом называют акроцентрическими. В зоне первичного сужения расположен кинетохор - сложная белковая структура, имеющая форму овальной пластинки, связанной с ДНК центромерного района хромосомы. К кине-тохору во время митоза подходят микротрубочки клеточного веретена, связанные с перемещением хромосом при делении клетки. Некоторые хромосомы имеют, кроме того, вторичную перетяжку, располагающуюся вблизи одного из концов хромосомы и отделяющую маленький участок - спутник хромосомы. Вторичные перетяжки называют, кроме того, ядрышковыми организаторами, так как именно на этих участках хромосом в интерфазе происходит образование ядрышка. В этих местах локализована ДНК, ответственная за синтез рибосомных РНК.
Рис. 4.24. Строение хромосомы:
хромосома в световом микроскопе (а) и ее схематическое изображение (б); хромосома при дифференциальной окраске (в) и ее схематическое изображение (г); д - хромосома в сканирующем электронном микроскопе; е - хромосома в трансмиссионном мегавольтном электронном микроскопе. 1 - теломеры; 2 - центромеры; 3 - плечи хромосомы
Плечи хромосом оканчиваются теломерами - конечными участками. Размеры хромосом, как и их число, у разных организмов варьируют в широких пределах.
Совокупность числа, размеров и особенностей строения хромосом называется кариотипом данного вида. Кариотип не зависит ни от вида клеток, ни от возраста данного организма.
При специальных методах окраски хромосомы неравномерно воспринимают красители: вдоль их длины наблюдается чередование окрашенных и неокрашенных участков - дифференциальная неоднородность хромосомы. Важно то, что каждая хромосома имеет свой, неповторимый рисунок такой дифференциальной окраски. Применение методов дифференциаль-
ной окраски позволило детально изучить строение хромосом. Хромосомы человека принято подразделять по их размерам на 7 групп (А, В, С, D, Е, F, G). Если при этом легко отличить крупные (1, 2) хромосомы от мелких (19, 20), метацентрические от акроцентрических (13), то внутри групп трудно отличить одну хромосому от другой. Так, в группе С6 и С7 хромосомы схожи между собой, как и с Х-хромосомой. Только дифференциальное окрашивание позволяет четко отличить эти хромосомы друг от друга.
После митоза хромосомы деконденсируются, образуя хроматин интерфазного ядра, однако каждая хромосома сохраняет свою индивидуальность и занимает в интерфазном ядре отдельную область (рис. 4.25).
Ядрышко
Практически во всех живых клетках эукариотических организмов в ядре видно одно или несколько обычно округлой формы телец величиной 1-5 мкм, сильно преломляющих свет, - это ядрышко, или нуклеола (nucleolus). К общим свойствам ядрышка относится способность хорошо окрашиваться различными красителями, особенно основными. Такая базофилия определяется тем, что ядрышки богаты РНК. Ядрышко - самая плотная структура ядра - является участком хромосомы, одним из ее локусов с наиболее высокой концентрацией и активностью синтеза РНК в интерфазе. Оно не является самостоятельной структурой или органеллой. Образование ядрышек и их число связаны с активностью и числом определенных участков хромосом - ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек; количество ядрышек в клетках данного типа может изменяться за счет слияния ядрышек или за счет изменения числа хромосом с ядрышковыми организаторами. ДНК ядрышкового организатора представлена множественными (несколько сотен) копиями генов рРНК: на каждом из этих генов синтезируется высокомолекулярный предшественник РНК, который превращается в более короткие молекулы РНК, входящие в состав субъединиц рибосомы.
Схему участия ядрышек в синтезе цитоплазматических белков можно представить следующим образом: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком, здесь происходит сборка рибонуклеопротеидных частиц - субъединиц
рибосом; субъединицы, выходя из ядрышка в цитоплазму, организуются в рибосомы и участвуют в процессе синтеза белка.
Ядрышко неоднородно по своему строению: в световом микроскопе можно видеть его тонковолокнистую организацию. В электронном микроскопе выявляются две части: гранулярная и фибриллярная (см. рис. 4.22, б). Диаметр гранул около 15-20 нм, толщина фибрилл 6-8 нм.
Рис. 4.25. Хромосомные территории в интерфазном ядре
В составе ядрышек обнаруживаются фибриллярные центры, содержащие ДНК ядрышковых организаторов хромосом, вокруг которых расположена плотная фибриллярная часть, синтезирующая предшественники рибосомных РНК (рРНК). Гранулярная часть представлена строящимися и зрелыми субъединицами рибосом, которые по мере их организации транспортируются в цитоплазму, где образуют функционирующие рибосомы, участвующие в синтезе белков.
Ультраструктура ядрышек зависит от активности синтеза РНК: при высоком уровне синтеза рРНК в ядрышке выявляется большое число гранул, при прекращении синтеза количество гранул снижается, ядрышки превращаются в плотные фибриллярные тельца базофильной природы.
Действие многих веществ (актиномицин, митомицин, ряд канцерогенных углеводородов, циклогексимид, гидрооксимочевина и др.) вызывает в клетках падение интенсивности ряда синтезов и в первую очередь активности ядрышек. При этом возникают изменения в структуре ядрышек: их сжатие, обособление фибриллярных и гранулярных зон, потеря гранулярного компонента, распад всей структуры. Эти изменения отражают степень повреждения ядрышковых структур, связанных главным образом с подавлением синтеза рРНК.
Ядерная оболочка
Ядерная оболочка (tegmentum nucleare), или кариолемма, состоит из внешней ядерной мембраны (m. nuclearis externa) и внутренней мембраны оболочки (m. nucle-aris interna), разделенных перинуклеарным пространством (рис. 4.26). Ядерная оболочка содержит многочисленные ядерные поры (pori nucleares).
Из многих свойств и функциональных нагрузок ядерной оболочки следует подчеркнуть ее роль как барьера, отделяющего содержимое ядра от цитоплазмы, ограничивающего свободный доступ в ядро крупных агрегатов биополимеров, регулирующего транспорт макромолекул между ядром и цитоплазмой.
Мембраны оболочки ядра в морфологическом отношении не отличаются от остальных внутриклеточных мембран. В общем виде оболочка ядра может быть представлена как полый двухслойный мешок, отделяющий содержимое ядра от цитоплазмы.
Наружная мембрана оболочки ядра, непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматической сети: на ней со стороны гиалоплазмы расположены многочисленные полирибосомы, а сама наружная мембрана может прямо переходить в мембраны эндоплаз-матической сети. Одной из важных функций оболочки ядра следует считать ее участие в создании внутриядерного порядка - в фиксации хромосомного материала в трехмерном пространстве ядра. В интерфазе часть хроматина структурно связана с внутренней мембраной оболочки ядра. Эта связь опосредуется с помощью фиброзной ядерной пластинки (ламины), с которой связываются фибриллы хроматина.
Наиболее характерными структурами оболочки ядра являются ядерные поры. Они образуются за счет слияния наружной и внутренней мембран обо-
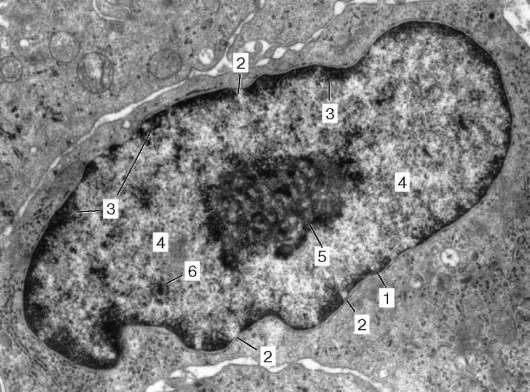
Рис. 4.26. Строение ядра интерфазной клетки:
1 - оболочка ядра (наружная и внутренняя мембраны, перинуклеарное пространство); 2 - комплекс ядерной поры; 3 - гетерохроматин; 4 - эухроматин; 5 - ядрышко; 6 - межхроматиновые гранулы РНК. Электронная микрофотография, увеличение 12 000
лочки ядра. Формирующиеся при этом округлые сквозные отверстия поры имеют диаметр около 90 нм. Эти отверстия в ядерной оболочке заполнены сложно организованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом ядерной поры (complexus pori nuclearis). Последний имеет октагональную симметрию. По границе отверстия в наружной и внутренней мембранах оболочки ядра располагаются по 8 белковых субъединиц, которые составляют белковые кольца ядерной поры (наружное и внутреннее). От наружного кольца поры в сторону цитоплазмы отходят длинные филаменты. От внутреннего кольца поры в глубь ядра также отходят филаменты, образуя структуру, подобную корзинке.
Комплекс ядерной поры в функциональном отношении представляет собою сложную систему, которая активно участвует не только в рецепции транспортируемых макромолекул (белков и нуклеопротеидов), но и собственно в актах их переноса (транслокации), при которых используется АТФ. В состав каждого комплекса ядерной поры входит несколько сотен различных белков.
Число ядерных пор зависит от метаболической активности клеток: чем интенсивнее синтетические процессы в клетках, тем больше пор в оболочке ядра. Так, у эритробластов (клеток-предшественников ядерных эритроцитов) низших позвоночных животных во время интенсивного синтеза и накопления гемоглобина в оболочке ядра обнаруживаются около 30 пор на
1 мкм2 поверхности. После того как эти процессы заканчиваются, в ядрах зрелых клеток - эритроцитов - прекращается синтез ДНК и РНК и количество пор в оболочке ядра снижается до 5 на 1 мкм2 поверхности. В оболочке ядра зрелых сперматозоидов поры не обнаруживаются. В среднем в оболочке ядра соматической клетки обнаруживается несколько тысяч поро-вых комплексов.
4.3. ВОСПРОИЗВЕДЕНИЕ КЛЕТОК 4.3.1. Клеточный цикл и его регуляция
Делению клеток предшествует редупликация хромосом вследствие синтеза ДНК. Это правило является общим для про- и эукариотических клеток. Время существования клетки от одного деления до другого называется клеточным циклом (cyclus cellularis).
Во взрослом организме высших позвоночных клетки различных тканей и органов имеют неодинаковую способность к делению. Встречаются популяции клеток, полностью потерявшие свойство делиться. Это большей частью специализированные, дифференцированные клетки (например, зернистые лейкоциты крови). В организме есть постоянно обновляющиеся ткани - различные эпителии, кроветворные ткани. В таких тканях существует часть клеток, которые постоянно делятся, заменяя стареющие или погибающие клетки (например, клетки базального слоя покровного эпителия, клетки крипт кишечника, кроветворные клетки костного мозга). Многие клетки, не размножающиеся в обычных условиях, приобретают вновь это свойство при процессах репаратив-ной регенерации органов и тканей. В гистогенезе большинство клеток после определенного числа делений переходит в гетеросинтетическую интерфазу, которая включает время роста, дифференцировки, функционирования, старения и смерти. В целом это характеризует жизненный цикл клетки.
При изучении клеточного цикла встречаются как диплоидные (2 с), так и тетраплоидные (4 с) и интерфазные клетки с промежуточным количеством ДНК. Это объясняется особенностями цикла размножения клеток. Весь клеточный цикл состоит из четырех отрезков времени: собственно митоза (М), пресинтети-ческого (G1), синтетического (S) и постсинтетического (G2) периодов интерфазы (рис. 4.27).
Рис. 4.27. Клеточный цикл (схема). Пояснения в тексте
В G1-периоде, наступающем сразу после деления, клетка имеет диплоидное содержание ДНК в ядре (2 с). После деления в периоде G1 в дочер-
них клетках общее содержание белков и РНК вдвое меньше, чем в исходной родительской клетке. В периоде G1 наблюдается рост клеток главным образом за счет накопления клеточных белков, что обусловлено увеличением количества РНК в клетке, и подготовка клетки к синтезу ДНК.
Обнаружено, что подавление синтеза белка или иРНК в G1-периоде предотвращает наступление S-периода, так как в течение G1-периода происходят синтезы ферментов, необходимых для образования предшественников ДНК (например, нуклеотидфосфокиназ), ферментов метаболизма РНК и белка. При этом резко повышается активность ферментов, участвующих в энергетическом обмене.
В следующем, S-периоде, происходит удвоение количества ДНК в ядре и соответственно удваивается число хромосом. В ядрах разных клеток, находящихся в S-периоде, можно обнаружить разные количества ДНК - от 2 до 4 с, что отражает постепенное накопление ДНК по мере прохождения клеткой синтетического периода клеточного цикла. S-период является узловым в клеточном цикле. Без синтеза ДНК неизвестно ни одного случая вступления клеток в митотическое деление.
Единственным исключением является второе деление созревания половых клеток в мейозе, когда между двумя делениями нет синтеза ДНК.
В S-периоде уровень синтеза РНК возрастает соответственно увеличению количества ДНК, достигая своего максимума в С2-периоде.
Постсинтетический (G2) период называется также премитотическим. В данном периоде синтезируется иРНК, необходимая для митоза. Среди синтезирующихся в это время белков особое место занимают тубулины - белки митотического веретена.
В конце G2-периода или в митозе по мере конденсации митотических хромосом синтез РНК резко снижается и полностью прекращается во время митоза. Синтез белка во время митоза понижается до 25 % исходного уровня и затем в последующих периодах достигает своего максимума в G2-периоде, в общем повторяя характер синтеза РНК.
В растущих тканях растений и животных всегда есть клетки, которые находятся вне цикла. Такие клетки принято называть клетками G0-периода. Это клетки, которые после митоза не вступают в пресинтетический период (G1). Именно они представляют собой так называемые покоящиеся, временно или окончательно переставшие размножаться клетки. В некоторых тканях такие клетки могут находиться длительное время, не изменяя особенно своих морфологических свойств: они сохраняют способность к делению. Это, например, камбиальные клетки (стволовые в кроветворной ткани). Чаще потеря (хотя бы и временная) способности делиться сопровождается специализацией и дифференцировкой. Такие дифференцирующиеся клетки выходят из цикла, но в особых условиях могут снова входить в цикл. Например, большинство клеток печени находится в G0-периоде; они не синтезируют ДНК и не делятся. Однако при удалении части печени у экспериментальных животных многие клетки начинают подготовку к митозу, переходят к синтезу ДНК и могут митотически делиться. В других случаях, например в эпидермисе кожи, после выхода клетки из цикла
размножения они дифференцируются, выполняют свои защитные функции, а затем погибают (ороговевшие клетки покровного эпителия). Многие клетки теряют полностью способность возвращаться в митотический цикл. Так, например, нейроны головного мозга и кардио-миоциты постоянно находятся в G0-периоде клеточного цикла (до смерти организма).
Регуляция вхождения и выхода клеток из клеточного цикла находится под контролем особой системы белковых факторов. Было обнаружено множество факторов роста (ФР), стимулирующих клетки к размножению, к пролиферации. Так, например, ФР из тромбоцитов стимулирует размножение клеток соединительной ткани, гормон эритро-поэтин вызывает размножение пред-
шественников эритроцитов, гормон прогестерон стимулирует пролиферацию клеток молочной железы и т. п.
Разные ФР передают сигналы на синтез специальных внутриклеточных белков, образующих каскад протеинкиназ (фосфорилаз), связанных с запуском клеточного цикла.
В состав этих белков, факторов, стимулирующих митоз, входит комплекс, состоящий из двух субъединиц: регуляторной (белок циклин) и каталитической (циклин-зависимая протеиназа).
У млекопитающих в реализации всего клеточного цикла участвуют 9 различных циклинов и 7 циклин-зависимых киназ (ЦЗК). При этом для перехода от одного периода клеточного цикла к другому используются разные циклины (D, E, A, B и др.) и разные ЦЗК (рис. 4.28). Так, например, переход от интерфазного ядра в G2-периоде непосредственно к митозу определяется фактором, состоящим из циклинов А/В и протеин-зависимой киназы 1.
Рис. 4.28. Участие различных циклинов и циклин-зависимых киназ в клеточном цикле млекопитающих: 1 - циклин D+ЦЗК 4, ЦЗК 6; 2 - циклин Е+ЦЗК 2; 3 - циклин А+ЦЗК 2; 4 - циклин В/А+ЦЗК 1
Деление клеток: митоз
Митоз (mitosis), кариокинез, или непрямое деление, - универсальный способ деления любых эукариотических клеток. При этом редуплицирован-ные и конденсированные хромосомы переходят в компактную форму мито-тических хромосом, образуется веретено деления, участвующее в сегрегации и переносе хромосом (ахроматиновый митотический аппарат), происходят расхождение хромосом к противоположным полюсам клетки и деление тела клетки (цитокинез, цитотомия). Процесс непрямого деления клеток приня-
Рис. 4.29. Митоз клетки (схема):
1 - интерфаза; 2 - профаза; 3 - метафаза; 4 - анафаза; 5 - телофаза; 6 - ранняя интерфаза
то подразделять на несколько основных фаз: профаза, метафаза, анафаза, телофаза (рис. 4.29).
Профаза. После окончания S-периода количество ДНК в интерфазном ядре равно 4 с, так как произошло удвоение хромосомного материала. Однако морфологически регистрировать удвоение числа хромосом в этом периоде не всегда удается. Это связано с тем, что в профазе сестринские хромосомы тесно соприкасаются и взаимно спирализуются одна относительно другой. Тем не менее в профазе каждая из хромосом двойная, что является результатом их редупликации в S-периоде клеточного цикла. Позднее хромосомы в каждой такой паре начинают обособляться, раскручиваться. Сестринские хромосомы в митозе отчетливо выявляются в конце профазы, когда видно, что общее их число в начинающей делиться клетке равно 4 n. Следовательно, уже в начале профазы хромосомы состояли из двух сестринских хромосом, или хроматид. Число их (4 n) в профазе точно соответствует количеству ДНК (4 с).
Параллельно конденсации хромосом в профазе происходят исчезновение и дезинтеграция ядрышек в результате инактивации рибосомных генов в зоне ядрышковых организаторов.
Одновременно с этим в середине профазы начинается разрушение ядерной оболочки: исчезают ядерные поры, оболочка распадается сначала на фрагменты, а затем на мелкие мембранные пузырьки.
В это время меняются и структуры, связанные с синтезом белка. Происходит уменьшение количества гранулярной эндоплазматической сети, она распадается на короткие цистерны и вакуоли, число рибосом на ее мембранах резко падает. Значительно (до 25 %) редуцируется число полисом как на мембранах, так и в гиалоплазме, что является признаком общего снижения уровня синтеза белка в делящихся клетках.
Второе важнейшее событие при митозе тоже происходит во время профазы - это образование веретена деления. В профазе репродуцировавшиеся в S-периоде центриоли начинают расходиться к противоположным полюсам клетки. К каждому полюсу отходит по двойной центриоли, или диплосоме. По мере расхождения диплосом начинают формироваться микротрубоч-
ки, отходящие от периферических участков одной из центриолей каждой диплосомы.
Сформированный в метафазе аппарат деления в животных клетках имеет веретеновидную форму и состоит из нескольких зон: двух зон центросфер с центриолями внутри них и промежуточной между ними зоны волокон веретена. Во всех этих зонах имеется большое число микротрубочек (рис. 4.30).
Микротрубочки в центральной части этого аппарата, в собственном веретене деления, так же как микротрубочки центросфер, возникают в результате полимеризации тубулинов в зоне центриолей. Эти микротрубочки достигают кине-тохоров, расположенных в области центромерных перетяжек хромосом, и связываются с ними. В веретене деления различают два типа микротрубочек: идущие от полюса к центру веретена и хромосомные, соединяющие хромосомы с одним из полюсов.
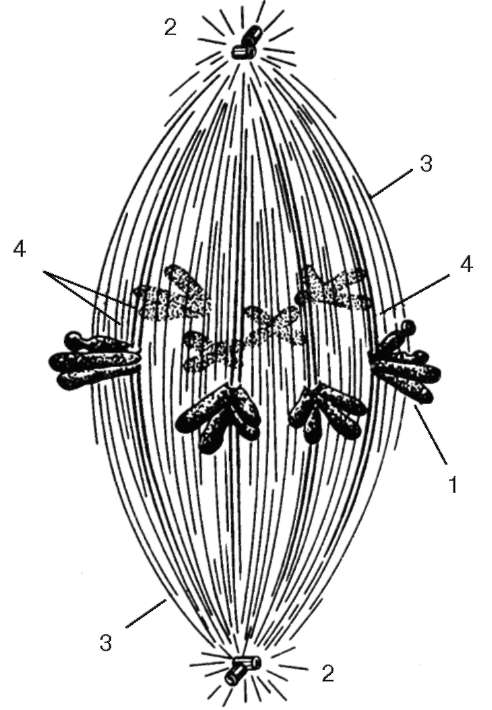
Рис. 4.30. Строение митотического веретена (схема):
1 - хромосомы; 2 - клеточный центр; 3 - центриолярные микротрубочки; 4 - кинетохорные микротрубочки
Метафаза занимает около трети времени всего митоза. Во время метафа-зы заканчивается образование веретена деления, а хромосомы выстраиваются в экваториальной плоскости веретена, образуя так называемую экваториальную (метафазную) пластинку хромосом, или материнскую звезду. К концу метафазы завершается процесс обособления друг от друга сестринских хроматид. Их плечи лежат параллельно друг другу, между ними хорошо видна разделяющая их щель. Последним местом, где контакт между хрома-тидами сохраняется, является центромер (первичная перетяжка).
Анафаза. Хромосомы все одновременно теряют связь друг с другом в области центромер и синхронно начинают удаляться друг от друга по направлению к противоположным полюсам клетки. Скорость движения хромосом равномерная, она может достигать 0,2-0,5 мкм/мин. Анафаза - самая короткая стадия митоза (несколько процентов от всего времени), но за это время происходит ряд событий. Главными из них являются обособление двух идентичных наборов хромосом и перемещение их в противоположные концы клетки. Расхождение хромосом по направлению к полюсам происходит одновременно с расхождением самих полюсов.
Показано, что расхождение хромосом связано с укорочением (деполимеризацией) микротрубочек в районе кинетохоров хромосом и с работой
белков-транслокаторов, перемещающих хромосомы. Дополнительное расхождение полюсов в анафазе обеспечивается за счет скольжения относительно друг друга межполюсных микротрубочек, которое обеспечивается работой другой группы белков-транслокаторов.
Телофаза начинается с остановки разошедшихся диплоидных (2 n) наборов хромосом (ранняя телофаза) и завершается, когда возникает реконструкция нового интерфазного ядра (поздняя телофаза, ранний G1-период) и происходит разделение исходной клетки на две дочерние (цитокинез, цитотомия). В ранней телофазе хромосомы, не меняя своей ориентации (центромерные участки - к полюсу, теломерные - к центру веретена), начинают деконденсироваться и увеличиваться в объеме. В местах их контактов с мембранными пузырьками цитоплазмы образуется новая ядерная оболочка. После замыкания ядерной оболочки начинается формирование новых ядрышек. Клетка переходит в новый G1-период клеточного цикла.
Важное событие телофазы - разделение клеточного тела - цитото-мия, или цитокинез, который происходит путем образования перетяжки в результате впячивания плазматической мембраны внутрь клетки. При этом в подмембранном слое цитоплазмы располагаются сократимые элементы типа актиновых миофиламентов, ориентированные циркулярно в зоне экватора клетки. Сокращение филаментов приводит к впячиванию плазматической мембраны в области этого кольца, что завершается разделением клетки на две.
Аномалии клеточного деления
При повреждении митотического аппарата (действие холода или агентов, вызывающих деполимеризацию тубулинов) может произойти или задержка митоза в метафазе, или рассеивание хромосом. При нарушениях репродукции центриолей могут возникать многополюсные и асимметричные митозы и т. п. Нарушения цитотомии приводят к появлению клеток с гигантскими ядрами или многоядерных полиплоидных клеток. Это связано с подавлением образования актиновых микрофиламентов, участвующих в формировании клеточной перетяжки в конце телофазы.
Полиплоидия - образование клеток с повышенным содержанием ДНК. Такие полиплоидные клетки появляются в результате полного отсутствия или незавершенности отдельных этапов митоза. Появление полиплоидных соматических клеток может наблюдаться в норме при блокаде деления клеточного тела. В печени взрослых млекопитающих встречаются, кроме диплоидных, тетра- и октаплоидные (4 n и 8 n) клетки, а также двуядерные клетки разной степени плоидности.
Процесс полиплоидизации этих клеток происходит следующим образом. После S-периода клетки, обладающие 4 с количеством ДНК, вступают в митотическое деление, проходят все его стадии, включая телофазу, но не приступают к цитотомии. Таким образом, образуется двуядерная клетка (2x2 n). Если она снова проходит S-период, то оба ядра в такой клетке будут содержать по 4 с ДНК и 4 n хромосом. Такая двуядерная клетка входит в митоз, на стадии метафазы происходит объединение хромосомных
наборов (общее число хромосом равно 8 n), а затем - нормальное деление, в результате которого образуются две тетраплоидные клетки. Этот процесс попеременного появления двуядерных и одноядерных клеток приводит к появлению ядер с 8 n, 16 n и даже 32 n количеством хромосом. Подобным способом образуются полиплоидные клетки в печени, в эпителии мочевого пузыря, в пигментном эпителии сетчатки, в ацинарных отделах слюнных и поджелудочной желез, мегакариоциты костного мозга.
Необходимо отметить, что полиплоидизация соматических клеток характерна для специализированных, дифференцированных клеток и не встречается при генеративных процессах, таких как эмбриогенез (исключая провизорные органы) и образование половых клеток; нет полиплоидии среди стволовых клеток.
Процесс митотического деления клеток очень чувствителен к действию самых разнообразных факторов. Наиболее часто встречается остановка митоза на стадии метафазы. Это происходит в результате изменений веретена деления. Многие вещества, останавливающие митоз, например, такие цитостатики, как колхицин и колцемид, препятствуют полимеризации тубулинов. В результате этого новые микротрубочки веретена не образуются, а готовые полностью разбираются. При этом митотические хромосомы собираются в центре клетки, но не образуют метафазную пластинку, а располагаются без всякого порядка (К-митоз). К сходным результатам приводит действие на клетку ингибиторов синтеза АТФ (динитрофенол, олигомицин) и ряда ядовитых веществ (мер-каптоэтанол). Если действие этих факторов кратковременное, то возможны восстановление микротрубочек веретена и клеточное деление. При умеренных воздействиях клетки могут не погибнуть, а без митоза вступать в следующий клеточный цикл. В этом случае неразошедшие-ся хромосомы деконденсируются, образуются новая ядерная оболочка и новое, но уже тетраплоидное ядро, которое переходит в G1-фазу. Так возникают полиплоидные клетки при действии колхицина.
К аномалиям деления клеток относятся и многополюсные митозы. В этом случае в метафазе образуется не биполярное веретено, а веретено с тремя или четырьмя полюсами. Такая аномалия связана с нарушениями функций центриолей: диплосома распадается на две активные моноцентриоли. Эти изменения могут происходить спонтанно (что характерно для опухолевых клеток) или после воздействия различных ингибиторов митотического деления. Такие аномальные трех- и четы-рехполюсные митотические фигуры могут вступать в анафазу и участвовать в расхождении хромосом к полюсам, вслед за чем может наступить цитотомия с образованием 3 или 4 клеток. В этих случаях не происходит равномерного распределения хромосом, а образовавшиеся клетки содержат случайные и уменьшенные наборы хромосом. Клетки с ненормальным числом хромосом называют анэуплоидными. Эти клетки обычно быстро погибают.
Нарушения митотического деления могут быть связаны со структурными изменениями самих хромосом. Так, воздействие различными формами лучистой энергии (ультрафиолетовый свет, рентгеновские лучи и т. п.) или разными алкилирующими соединениями (иприт, цитоста-тики) может привести к нарушениям структуры хромосом и изменениям хода митоза. В результате таких воздействий возникают так называемые хромосомные аберрации. Это могут быть делеции - потери участков хромосом, инверсии - перестановка участков хромосом, транслокации - перенос участков с одной хромосомы на другие.
При разрыве хромосомы та ее часть, которая не несет центромеры, не участвует в хромосомном делении, отстает от основной массы хромосом и случайно оказывается в одной из дочерних клеток. Такой фрагмент хромосомы в интерфазе покрывается собственной ядерной оболочкой (возникает дополнительное микроядро). Ясно, что при этом обе дочерние клетки будут анэуплоидными.
В других случаях в результате объединения двух поврежденных хромосом возникает одна хромосома, но с двумя центромерами, которые растягиваются к противоположным полюсам. При этом между двумя группами хромосом в анафазе и в телофазе виден «мост», возникает растянутая аберрантная хромосома.
4.4. РЕАКЦИЯ КЛЕТОК НА ВНЕШНИЕ ВОЗДЕЙСТВИЯ
Организм и его клетки постоянно подвергаются воздействию самых разнообразных химических, физических или биогенных факторов. Эти факторы могут вызывать первичное нарушение одной или нескольких клеточных структур, что в свою очередь приводит к функциональным нарушениям. В зависимости от интенсивности поражения, его длительности и характера судьба клетки может быть различна. Измененные в результате повреждения клетки могут адаптироваться, приспособиться к воздействующему фактору, восстанавливаться, реактивироваться после снятия повреждающего воздействия или измениться необратимо и погибнуть. Именно поэтому функциональные и морфологические картины клеток в этих состояниях очень разнообразны. На различные факторы при обратимом повреждении клетки отвечают рядом изменений. Одним из проявлений общеклеточной реакции на повреждение является изменение способности клетки связывать различные красители. Так, нормальные клетки, поглощая из внеклеточной среды растворенные в ней красители, откладывают их в виде гранул. Такое грану-лообразование происходит в цитоплазме, ядро при этом остается бесцветным. При повреждении клеток многими физическими (нагревание, давление) или химическими факторами (изменение рН среды, добавление спирта или какого-либо иного денатурирующего агента) гранулообразование пре-
кращается, цитоплазма и ядро диффузно окрашиваются проникшим в клетку красителем. Если действие фактора обратимо и при устранении его клетка возвращается к норме, то снова восстанавливается ее способность к гранулообразованию. При различных повреждениях клеток значительно падает окислительное фосфорилирование: прекращается синтез АТФ и возрастает потребление кислорода. Для поврежденных клеток характерны усиление гликолитических процессов, снижение количества АТФ, активация протеолиза. Совокупность неспецифических обратимых изменений цитоплазмы, возникающих под воздействием различных агентов, была обозначена термином «паранекроз» (Насонов Д. Н., Александров В. Я., 1940).
При различных воздействиях на клетку наиболее частым изменением структуры ядра является конденсация хроматина, что может отражать падение ядерных синтетических процессов. При гибели клетки происходит агрегация хроматина, возникают грубые сгустки внутри ядра (пикноз), что часто завершается распадом на части (кариорексис) или растворением ядра (кариолизис). Ядрышки при подавлении синтеза рРНК уменьшаются в размерах, теряют гранулы, фрагментируются.
К наиболее часто встречающимся изменениям ядерной оболочки относятся расширение (отечность) перинуклеарного пространства, извитость контура ядерной оболочки, что нередко сочетается с пикнозом ядра. На ранних этапах повреждения клетки часто приобретают шаровидную форму и теряют многочисленные клеточные выросты и микроворсинки. В дальнейшем, наоборот, изменения плазмолеммы сводятся к появлению на поверхности клеток различных выростов или мелких пузырей.
На начальных стадиях нарушения окислительного фосфорилирования происходят сжатие митохондриального матрикса и некоторое расширение межмембранного пространства. В дальнейшем этот тип реакции митохондрий может смениться их набуханием, что особенно часто встречается при самых различных патологических изменениях клеток. Митохондрии при этом принимают сферическую форму и увеличиваются в размерах, происходит обводнение матрикса, он становится светлым. Набухание митохондрий, как правило, сопровождается редукцией числа и размера крист. При необратимом повреждении митохондрий происходит разрыв их мембран, матрикс смешивается с гиалоплазмой.
Эндоплазматическая сеть чаще всего подвергается вакуолизации и распаду на мелкие пузырьки. При этом на мембранах сети уменьшается число рибосом, что однозначно указывает на падение белкового синтеза. Цистерны комплекса Гольджи могут увеличиваться в объеме или распадаться на мелкие вакуоли. В поврежденных клетках происходит активация лизосом, увеличивается число аутофаголизосом. При тяжелых клеточных повреждениях мембраны лизосом разрываются и лизосомные гидролазы начинают разрушать сами клетки - происходит лизис клеток.
При повреждении клетки резко снижается ее митотическая активность. Клетки часто задерживаются на разных стадиях митоза, главным образом из-за нарушения митотического аппарата, очень чувствительного к изменениям внутриклеточной среды.
Если изменения в клетке не зашли слишком далеко, происходят репарация клеточных повреждений, возврат клетки к нормальному функциональному уровню. Процессы восстановления внутриклеточных структур называются внутриклеточной регенерацией.
Репарация клеток бывает полной, когда восстанавливаются все свойства данных клеток, или неполной. В последнем случае после снятия действия повреждающего фактора нормализуется ряд функций клеток, но через некоторое время клетки погибают без всякого воздействия. Особенно часто это наблюдается при поражениях клеточного ядра.
Повреждение клеток внешними и внутриорганизменными факторами может привести к нарушениям регуляции их метаболизма. При этом происходит интенсивное отложение или же, наоборот, резорбция ряда клеточных включений. Кроме того, наблюдается нарушение регуляции проницаемости клеточных мембран, что приводит к вакуолизации мембранных органелл. В патологической анатомии такие изменения в структуре клеток называют дистрофиями. Так, например, при жировой дистрофии в клетках накапливаются жировые включения. Часто в цитоплазме измененных клеток обнаруживаются скопления липопротеидных комплексов, имеющих вид многослойных мембранных пластов. Нарушение регуляторных процессов метаболизма сахаров приводит к патологическому отложению и накоплению гликогена (углеводная дистрофия), что, вероятно, связано с недостаточностью фермента, расщепляющего гликоген (глюкозо-6-фосфатазы). Часто в измененных клетках животных происходит отложение различных пигментов, белковых гранул (белковая дистрофия) и др.
Особой формой патологического нарушения регуляторных процессов могут быть нарушения специализации, одним из которых является злокачественный опухолевый рост. Опухолевые клетки характеризуются безудержностью, неограниченностью размножения, нарушением уровня дифференцировки, изменениями строения клеток, относительной автономностью от регуляторных влияний со стороны организма, способностью к метастазированию. Все эти свойства опухолевые клетки сохраняют от поколения к поколению, т. е. свойства злокачественности являются наследственной особенностью таких клеток. Поэтому раковые клетки относят к мутантам, обладающим измененной генетической структурой; именно изменением генотипа клетки можно объяснить непрерывную передачу дочерним клеткам дефектной (в отношении регуляции) информации.
При необратимом повреждении клетки гибнут. Дать определение момента клеточной смерти очень трудно (так же, как и при смерти целого организма), так как умирание - это не одномоментное явление, а процесс.
4.5. ГИБЕЛЬ КЛЕТОК
Различают две основные морфологические формы гибели клеток - некроз и апоптоз (рис. 4.31).
Некроз вызывается главным образом различными внешними факторами, химическими или физическими, которые прямо или опосредованно влияют на проницаемость мембран или на клеточную энергетику. Во всех этих случаях наблюдается достаточно монотонная последовательность нарушения клеточных функций и структур. Общим является то, что в клетке происходит изменение ионного состава, наблюдаются набухание орга-нелл, прекращение синтеза АТФ, белков, нуклеиновых кислот, деградация ДНК, активация лизосомных ферментов, что в конечном итоге приводит к растворению клетки - лизису.
Апоптоз может происходить без первичного нарушения клеточного метаболизма. При этом в результате воздействия различных стимулов происходит активация в ядре некоторых генов, ответственных за самоуничтожение клетки. Программа такого самоуничтожения (запрограммированной гибели клетки) может включаться в результате воздействия на клетку сигнальных молекул (часто это различные белковые факторы или различные гормоны). Так, некоторые лейкоциты погибают при действии на них глюкокортикои-дов. К активации генов самоуничтожения может приводить прекращение выработки регулирующего сигнала. Например, после удаления семенников полностью погибают клетки предстательной железы. Апоптоз наблюдается при нормальном эмбриональном развитии организма. Так, клетки тканей головастиков погибают в результате активации этого процесса гормонами. Гибнут клетки эмбриональных закладок, например клетки протока первичной почки, нейробласты периферических ганглиев и др. Во взрослом орга-
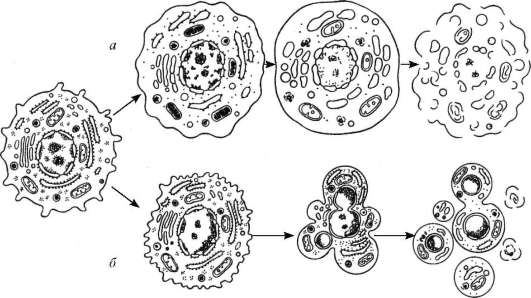
Рис. 4.31. Пути клеточной гибели:
а - некроз; б - апоптоз. Пояснения в тексте
низме апоптозу подвергаются клетки молочной железы при ее инволюции, клетки желтого тела яичника.
Причиной гибели клеток при апоптозе является активация каскада латентных протеиназ - каспаз. Существуют инициирующие и эффектор-ные каспазы. Субстратами для действия активированных каспаз служат более 60 различных белков. Это, например, киназа фокальных адгезивных структур, инактивация которой приводит к отделению апоптотиче-ских клеток от соседних в эпителии; это ламины, которые при действии каспаз разбираются, что приводит к почкованию ядер; это цитоскелетные белки, деградация которых вызывает изменение формы клеток и их распад на фрагменты - апоптотические тельца; это активируемая эндонуклеаза, вызывающая фрагментацию ДНК, и др.
Морфологически процесс апоптоза значительно отличается от некроза. На ранних его стадиях происходит возрастание уровня кальция в цитоплазме, но при этом мембранные органеллы не изменяются, синтез РНК и белка не снижается. Позднее в ядре хроматин конденсируется, образуя грубые скопления по периферии ядра. Ядра начинают фрагментиро-ваться, распадаться на «микроядра», каждое из которых покрыто ядерной оболочкой. Затем или одновременно с этим цитоплазма также начинает фрагментироваться. От клетки отшнуровываются крупные фрагменты, часто содержащие «микроядра». Это так называемые апоптотические тельца. Апоптотические тельца в норме поглощаются соседними клетками или фагоцитами, а также претерпевают вторичные некротические изменения и в конце концов растворяются.
Контрольные вопросы
1. Охарактеризуйте предмет и задачи цитологии, ее значение для практической медицины.
2. Перечислите основные положения клеточной теории. Каково теоретическое и практическое значение клеточной теории.
3. Какую роль в жизнедеятельности клетки играет ядро? Какие структурные компоненты входят в состав ядра?
4. Цитоплазма, ее постоянные и непостоянные структурные компоненты. Роль органелл в жизнедеятельности клетки.
5. Какие изменения происходят в ядре и цитоплазме клетки при подготовке к делению и собственно митозе?
6. Какие морфофункциональные изменения происходят в клетке при действии неблагоприятных внешних и внутриорганизменных факторов?