Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 18. КОЖА И ЕЕ ПРОИЗВОДНЫЕ
18.1. КОЖА
Кожа (cutis) образует внешний покров организма, площадь которого у взрослого человека достигает 1,5-2 м2. Кожа состоит из эпидермиса (эпителиальная ткань) и дермы (соединительнотканная основа). Кожа располагается на подкожной основе (клетчатке), или гиподерме, с помощью которой она соединяется с подлежащими частями организма (рис. 18.1). Толщина кожи в различных частях тела варьирует от 0,5 до 5 мм.
Функции. Кожа защищает подлежащие части организма от повреждений. Здоровая кожа непроницаема для микроорганизмов, многих ядовитых и вредных веществ, за исключением растворенных в жирах.
Кожа участвует в водно-солевом, а также в тепловом обмене с внешней средой. В течение суток через кожу человека выделяется около 500 мл воды, что составляет 1 % всего ее количества в организме. Кроме воды, через кожу вместе с потом выводятся различные соли, главным образом хлориды, а также молочная кислота и продукты азотистого обмена. Около 80 % всех тепловых потерь организма происходит через кожную поверхность. В случаях нарушения этой функции (например, при длительной работе в резиновом комбинезоне) могут возникнуть перегревание организма и тепловой удар.
В коже под действием ультрафиолетовых лучей из предшественников синтезируется витамин D3. Отсутствие его в организме вызывает тяжелое заболевание - рахит.
Наличие в коже обильной сосудистой сети и артериоловенулярных анастомозов определяет значение ее как депо крови. У взрослого человека в сосудах кожи может задерживаться до 1 л крови.
Кожа активно участвует в иммунных процессах. В ней происходят распознавание антигенов и их элиминация.
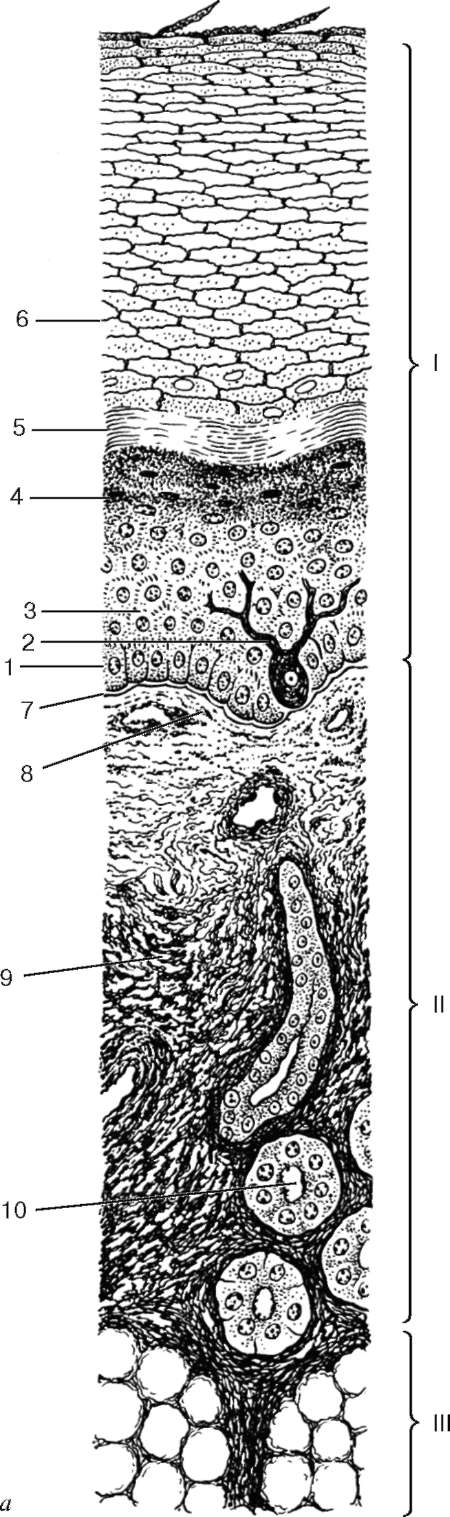
Рис. 18.1. Кожа пальца: а: I - эпидермис; II - дерма; III - гиподерма; 1 - базальный слой; 2 - меланоцит; 3 - шиповатый слой; 4 - зернистый слой; 5 - блестящий слой; 6 - роговой слой; 7 - базальная мембрана; 8 - сосочковый слой; 9 - сетчатый слой; 10 - концевой отдел потовой железы
Благодаря обильной иннервации кожный покров представляет собой огромное рецепторное поле, в котором сосредоточены осязательные, температурные и болевые нервные окончания. В некоторых участках кожи, например на голове и кистях, на 1 см2 ее поверхности насчитывается до 300 чувствительных точек. Кожа образует производные, или придатки, в виде желез (потовых, сальных, молочных, волос и ногтей).
Развитие. Кожа развивается из двух эмбриональных зачатков. Эпителиальный покров (эпидермис) ее образуется из кожной эктодермы, а подлежащие соединительнотканные слои - из мезенхимы дерматомов (производных сомитов). В первые недели развития зародыша эпителий кожи состоит всего из одного слоя плоских клеток. Постепенно эти клетки становятся все более высокими. В конце 2-го мес над ними появляется второй слой клеток, а на 3-м мес эпителий становится многослойным. Одновременно в наружных его слоях (в первую очередь на ладонях и подошвах) начинаются процессы ороговения. На 3-м мес внутриутробного периода в коже закладываются эпителиальные зачатки волос, желез и ногтей. В эпидермис проникают пигментные клетки, клетки Лангерганса, клет-
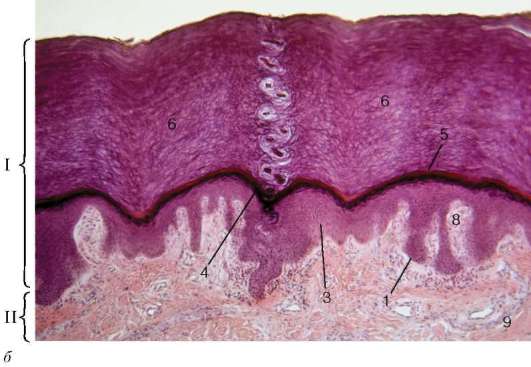
Рис. 18.1. Продолжение
б - микрофотография (обозначения см. выше)
ки Меркеля. В соединительнотканной основе кожи в этот период начинают образовываться волокна и густая сеть кровеносных сосудов. В глубоких слоях этой сети местами появляются очаги кроветворения. Лишь на 5-м мес внутриутробного развития образование кровяных элементов в них прекращается и на их месте формируется жировая ткань. Происходит иннервация кожи. В целом гисто- и органогенез кожи и ее производных характеризуется индуктивным взаимодействием эпидермиса и дермы. В качестве индуктора вначале выступает мезенхима, затем усиливается роль эпидермиса, что приводит к усложнению структуры и функции кожи как органа.
18.1.1. Эпидермис
Эпидермис (epidermis) - это полидифферонная ткань, ведущий эпителиальный дифферон которой формирует многослойный плоский ороговевающий эпителий. В последнем постоянно происходят обновление и специфическая дифференцировка клеток (кератинизация). Толщина его колеблется от 0,03 до 1,5 мм и более. По сравнению с кожей ладоней и подошв стоп эпидермис других участков кожи значительно тоньше. Толщина его, например, на волосистой части не превышает 170 мкм. Блестящий слой в нем отсутствует, а роговой представлен лишь 2-3 рядами ороговевших клеток - чешуек.
На основании различной толщины эпидермиса кожа подразделяется на толстую и тонкую. Толстая кожа покрывает небольшие участки тела (ладони, подошвы), тогда как тонкая выстилает остальные обширные его поверхности.
На ладонях и подошвах эпидермис состоит из многих десятков слоев клеток, которые объединены в пять основных слоев: базальный, шиповатый,
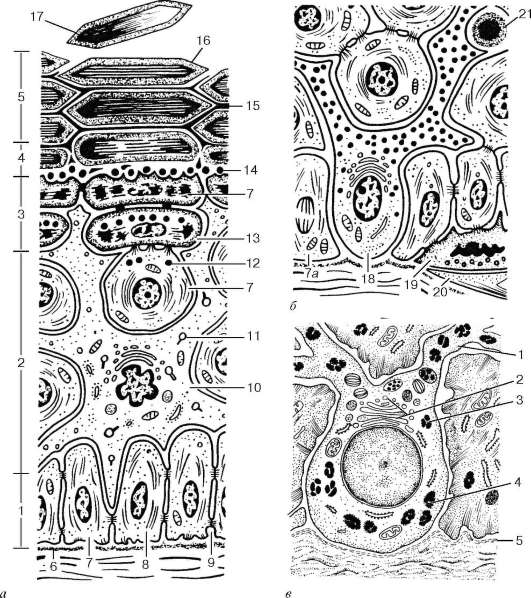
Рис. 18.2. Строение эпидермиса (по Е. Ф. Котовскому):
а - пролиферативная единица; б - меланоцит и клетка Меркеля: 1 - базальный слой; 2 - шиповатый слой; 3 - зернистый слой; 4 - блестящий слой; 5 - роговой слой; 6 - базальная мембрана; 7 - кератиноцит; 7а - митотическое деление клетки; 8 - кератиновые тонофиламенты; 9 - десмосомы; 10 - внутриэпидермальный макрофаг (клетка Лангерганса); 11 - гранулы Бирбека; 12 - кератиносомы; 13 - слой кератолинина; 14 - образование межклеточного цемента; 15 - межклеточный цемент; 16 - кератиноцит (роговая чешуйка); 17 - отпадающая роговая чешуйка; 18 - меланоцит с гранулами пигмента; 19 - клетка Меркеля с секреторными гранулами; 20 - афферентное нервное волокно; 21 - Т-лимфоцит; в - меланоцит: 1 - отростки меланоцита; 2 - комплекс Гольджи; 3 - премеланосомы; 4 - меланосомы; 5 - базальная мембрана эпидермиса
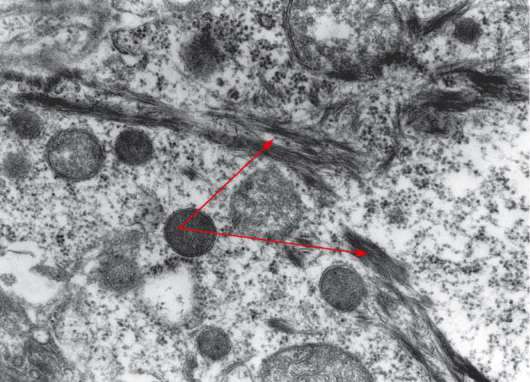
Рис. 18.3. Ультраструктура цитоплазмы кератиноцита: стрелками обозначены пучки тонофиламентов; увеличение 54 000 (препарат В. И. Семкина)
зернистый, блестящий и роговой (рис. 18.2). В остальных участках кожи четыре слоя (отсутствует блестящий слой). В них различают 5 типов клеток: кератиноциты (эпителиоциты), клетки Лангерганса (внутриэпидермальные макрофаги), лимфоциты, меланоциты, клетки Меркеля (см. рис. 18.2).
Из этих клеток в эпидермисе и каждом из его слоев (свыше 85 %) основу составляют кератиноциты. Они непосредственно участвуют в ороговении (кератинизации) эпидермиса. При этом в кератиноцитах происходит синтез специальных белков - кислых и щелочных типов кератинов, филаггри-на, инволюкрина, кератолинина и других, устойчивых к механическим и химическим воздействиям, и формируются кератиновые тонофиламенты и кератиносомы (рис. 18.3). Затем в них разрушаются органеллы и ядра, а между ними образуется межклеточное цементирующее вещество, богатое липидами - церамидами (керамидами) и поэтому непроницаемое для воды. Одновременно кератиноциты постепенно перемещаются из нижнего слоя в поверхностный, где завершается их дифференцировка и они получают название роговых чешуек (эпидермальных корнеоцитов). Весь процесс кератинизации продолжается 3-4 нед (на подошвах стоп - быстрее).
Базальный слой (stratum basale) образован кератиноцитами, меланоци-тами, клетками Меркеля, Лангерганса и камбиальными (стволовыми). Кератиноциты соединяются с базальной мембраной полуили гемидес-мосомами, а между собой и с клетками Меркеля - с помощью десмо-сом. Кератиноциты имеют столбчатую или кубическую форму, округлое богатое хроматином ядро и базофильную цитоплазму. В ней выявляются органеллы, кератиновые промежуточные тонофиламенты и в некоторых клетках гранулы черного пигмента меланина. Меланин фагоцитируется
кератиноцитами из меланоцитов, в которых образуется. В базальном слое кератиноциты размножаются путем митотического деления, и новообразованные клетки включаются в процесс кератинизации (дифференциров-ки). В базальном слое встречаются клетки в С0-периоде жизненного цикла. Среди них - стволовые клетки дифферона кератиноцитов (см. рис. 18.2), которые способны возвращаться в митотический цикл.
Таким образом, базальный слой, включающий стволовые клетки и делящиеся малодифференцированные кератиноциты, является ростковым, за счет которого постоянно (каждые 3-4 нед) происходит обновление (физиологическая регенерация) эпидермиса.
Меланоциты - пигментные клетки нейроглиальной природы, не связанные десмосомами с соседними клетками. Они имеют несколько ветвящихся отростков, достигающих зернистого слоя. В их цитоплазме отсутствуют тонофибриллы, но много рибосом и меланосом. Меланосомы - структуры овальной формы, состоящие из плотных пигментных гранул и фибриллярного каркаса, окруженных общей мембраной. Они оформляются в комплексе Гольджи, где к ним присоединяются ферменты тирозиназа и ДОФА-оксидаза. Эти ферменты участвуют в образовании из аминокислоты тирозина кожного пигмента меланина, содержащегося в меланосомах (от лат. melas - черный).
Положительная реакция на ДОФА-оксидазу является специфической для мела-ноцитов. В среднем на 10 кератиноцитов приходится один меланоцит (эпидермаль-ная меланиновая единица). Пигмент меланин обладает способностью задерживать ультрафиолетовые лучи и поэтому не позволяет им проникать в глубь организма, где они могут вызвать повреждение внутренних органов. Синтез пигмента возрастает под действием ультрафиолетовой радиации и меланоцитстимулирующего гормона гипофиза. В самом эпидермисе УФ-лучи оказывают влияние также на кера-тиноциты, стимулируя в них синтез витамина D, участвующего в минерализации костной ткани.
Клетки Меркеля (осязательные) нейроглиальные по происхождению. Они наиболее многочисленны в сенсорных областях кожи (пальцы, кончик носа и др.). К их основанию подходят афферентные нервные волокна. Клетки Меркеля и афферентные нервные волокна образуют в эпидермисе осязательные механорецепторы, реагирующие на прикосновение. В цитоплазме клеток выявляются гранулы с плотной сердцевиной, содержащие бомбе-зин, ВИП, энкефалин, эндорфин и другие пептиды. В связи с этим клетки Меркеля относятся к дисперсной эндокринной системе (APUD-серии). Эти клетки участвуют в регуляции регенерации эпидермиса, а также тонуса и проницаемости кровеносных сосудов дермы с помощью ВИП и гистамина, высвобождающегося под их влиянием из тучных клеток.
Клетки Лангерганса - внутриэпидермальные антигенпредставляющие клетки костномозгового происхождения, выполняют иммунологические функции макрофагов эпидермиса.
Эти клетки способны мигрировать из эпидермиса в дерму и в регионарные лимфатические узлы, а также могут воспринимать антигены в эпидермисе и
«представлять» их внутриэпидермальным лимфоцитам и лимфоцитам регионарных лимфатических узлов, запуская таким образом иммунологические реакции. Лимфоциты, относящиеся к Т-популяции, проникают в базальный и шиповатый слои эпидермиса из дермы. Здесь может происходить их пролиферация под влиянием интерлейкина-1 (ИЛ-1), выделяемого клетками Лангерганса, а также под влиянием факторов типа тимозина и тимопоэтина, вырабатываемых кератиноцитами. Таким образом, внутриэпидермальные макрофаги (клетки Лангерганса) и лимфоциты участвуют в построении иммунологического защитного барьера кожи, относящегося к периферической части иммунной системы организма.
Клетки Лангерганса составляют от 2 до 8 % клеток эпидермиса. Они не связаны десмосомами с окружающими кератиноцитами. Для них характерны отростчатая форма, лопастного вида ядро и присутствие в цитоплазме аргентаффинных гранул (гранулы Бирбека), имеющих вид теннисных ракеток. Своими отростками клетки Лангерганса объединяют окружающие их кератиноциты в эпидермальные пролиферативные единицы (ЭПЕ), из которых состоит эпидермис в некоторых участках кожи. Пролиферативная единица имеет форму вертикальной колонки, занимает всю толщу эпидермиса, возникает в результате пролиферации и дифференцировки одной стволовой клетки. Она состоит из центрально расположенной клетки Лангерганса и кератиноцитов (примерно из 20 в «тонкой» коже и из 50 - в «толстой» коже) всех слоев эпидермиса. В ЭПЕ клетки Лангерганса оказывают регулирующее влияние на пролиферацию и дифференцировку кератиноцитов, возможно, с помощью цитокинов, обнаруженных в их гранулах.
Другая разновидность внутриэпидермальных макрофагов - клетки Гринстейна. Эти клетки при взаимодействии с внутриэпидермальными Т-лимфоцитами-супрессорами также участвуют в регуляции иммунных реакций.
Шиповатый слой (stratum spinosum) включает кератиноциты и клетки Лангерганса. Кератиноциты, образующие 5-10 слоев, имеют полигональную форму. Они соединяются между собой и с находящимися в базальном слое кератиноцитами с помощью многочисленных десмосом, имеющих вид шипов на поверхности клеток (после мацерации). В их цитоплазме усиливаются синтез кератина и образование из него тонофиламент, которые соединяются в пучки - тонофибриллы (см. рис. 18.3). Также формируются новые структуры - кератиносомы, или ламеллярные гранулы (гранулы Одланда). Они представляют собой ограниченные мембраной скопления пластин, содержащих липиды (холестеринсульфат, церамиды и др.) и гидролитические ферменты.
Зернистый слой (stratum granulosum) состоит из 3-4 слоев кератиноци-тов овальной формы, в которых синтезируются белки - кератин, филаг-грин, инволюкрин и кератолинин. Филаггрин участвует в агрегации кера-тиновых тонофиламент, образуя между ними аморфный матрикс. К ним присоединяются белки, полисахариды, липиды, аминокислоты, которые высвобождаются при начинающемся здесь (под влиянием гидролитических ферментов кератиносом и лизосом) распаде ядер и органелл. В результате образуется сложное по составу соединение - кератогиалин. Последний при
световой микроскопии выявляется в виде крупных, не ограниченных мембраной базофильных гранул. Они заполняют цитоплазму кератиноцитов и придают им зернистый вид. Инволюкрин и кератолинин образуют белковый слой под плазмолеммой, защищающий ее от действия гидролитических ферментов кератиносом и лизосом, которые активизируются под воздействием клеток Лангерганса. При этом количество кератиносом в кератино-цитах увеличивается, и они выделяются путем экзоцитоза в межклеточные щели, где содержащиеся в них липиды (церамиды, холестеринсульфат и др.) образуют цементирующее вещество. Последнее соединяет кератиноциты между собой и создает в эпидермисе водонепроницаемый барьер, который предохраняет кожу от высыхания.
Блестящий слой (stratum lucidum) образуют плоские кератиноциты (эпи-дермальные корнеоциты), в которых полностью разрушаются ядро и орга-неллы, кератогиалиновые гранулы сливаются в светопреломляющую массу, состоящую из агрегированных кератиновых фибрилл и аморфного матрик-са, включающего филаггрин, и более толстым становится слой кератоли-нина под плазмолеммой. Между клетками почти исчезают десмосомы, но увеличивается количество цементирующего вещества, богатого липидами. Постепенно кератиноциты полностью заполняются продольно расположенными кератиновыми фибриллами, спаянными аморфным матриксом из филаггрина. Одновременно кератиноциты смещаются в наружный роговой слой.
Роговой слой (stratum corneum), толщина которого на ладонях и подошвах стоп достигает 600 мкм и более, состоит из закончивших дифференцировку кератиноцитов (корнеоцитов), получивших название роговых чешуек. Они имеют форму плоских многогранников, расположенных друг на друге в виде колонок. Чешуйки имеют толстую прочную оболочку, содержащую белок кератолинин. Вся внутренняя часть чешуек заполнена продольно расположенными кератиновыми фибриллами, связанными дисульфидными мостиками. Фибриллы упакованы в аморфном матриксе, состоящем из другого вида кератина. Филаггрин при этом расщепляется на аминокислоты, которые включаются в кератин фибрилл. Чешуйки связаны между собой с помощью межклеточного цементирующего вещества, богатого липидами, что делает его непроницаемым для воды. В процессе жизнедеятельности постоянно происходит десквамация (отторжение) роговых чешуек с поверхности эпидермиса. Важную роль в этом, вероятно, играют липолитические ферменты (стероидили холестеринсульфатазы), найденные в лизосомах клеток Лангерганса. Под их влиянием может происходить расщепление межклеточного вещества - холестеринсульфата, цементирующего роговые чешуйки.
Таким образом, в эпидермисе кожи происходят постоянно взаимосвязанные процессы пролиферации и дифференциации кератиноцитов. Значение этих процессов заключается в том, что они приводят к образованию в эпидермисе регулярно обновляющегося рогового слоя, который отличается механической и химической устойчивостью, высокой гидроизолирующей способностью, плохой теплопроводимостью и непроницаемостью для бактерий и их токсинов.
Электронно-микроскопически блестящий слой как отдельный слой не определяется.
Процессы пролиферации и дифференциации в эпидермисе регулируются при участии нервной системы, эндокринных желез (надпочечников и др.), а также регу-ляторных веществ - кейлонов, простагландинов, эпителиального фактора роста, вырабатываемых в самом эпидермисе клетками Лангерганса и кератиноцитами. Нарушение этих регуляторных механизмов приводит к изменению процессов пролиферации и кератинизации в эпидермисе, что лежит в основе некоторых кожных болезней (псориаз и др.).
Под влиянием некоторых внешних и внутренних факторов характер эпидермиса может существенно изменяться. Так, например, при сильных механических воздействиях (трении), при А-авитаминозе, под влиянием гидрокортизона резко усиливаются процессы ороговения.
Между эпидермисом и подлежащей дермой располагается базальная мембрана (см. рис. 18.1, 18.2).
18.1.2. Собственно кожа, дерма
Собственно кожа (corium), или дерма, имеет толщину от 0,5 до 5 мм, наибольшую - на спине, плечах, бедрах. Дерма делится на два слоя - сосочковый и сетчатый (см. рис. 18.1), которые не имеют между собой четкой границы.
Сосочковый слой (stratum papillare) располагается непосредственно под эпидермисом, состоит из рыхлой соединительной ткани, выполняющей трофическую функцию. Свое название этот слой получил от многочисленных сосочков, вдающихся в эпителий. Их величина и количество в коже различных частей тела неодинаковы. Наибольшее количество сосочков высотой до 0,2 мм находится в коже ладоней и подошв. В коже лица сосочки развиты слабо, а с возрастом могут совсем исчезнуть. Сосочковый слой дермы определяет индивидуальный характер расположения борозд и линий на поверхности кожи. Это имеет судебно-медицинское значение для идентификации личности.
Соединительная ткань сосочкового слоя состоит из тонких коллагено-вых, эластических и ретикулярных волокон, а также из клеток, среди которых наиболее часто встречаются фибробласты, макрофаги, тучные клетки и др. Здесь также находятся гладкие мышечные клетки, местами собранные в небольшие пучки, связанные с корнем волоса. Это мышца, поднимающая волосы. Однако имеются мышечные пучки, не связанные с волосами. Больше всего их в коже головы, щек, лба и тыльной поверхности конечностей. Сокращение мышечных клеток обусловливает появление так называемой гусиной кожи. При этом сжимаются мелкие кровеносные сосуды и уменьшается приток крови к коже, вследствие чего понижается теплоотдача организма.
Сетчатый слой (stratum reticulare), обеспечивающий прочность кожи, образован плотной неоформленной соединительной тканью с мощными пучками коллагеновых волокон и сетью эластических волокон. Пучки колла-
геновых волокон проходят в основном в двух направлениях: одни из них лежат параллельно поверхности кожи, другие - косо. Вместе они образуют сеть, строение которой определяется функциональной нагрузкой на кожу. В участках кожи, испытывающих сильное давление (кожа стопы, подушечек пальцев, локтей и др.), хорошо развита широкопетлистая, грубая сеть коллагеновых волокон. Наоборот, в тех участках, где кожа подвергается значительному растяжению (область суставов, тыльная сторона стопы, лицо и т. п.), в сетчатом слое обнаруживается узкопетлистая более нежная колла-геновая сеть. Эластические волокна в основном повторяют ход коллагено-вых пучков. Их значительно больше в участках кожи, часто испытывающих растяжение (в коже лица, суставов и т. п.). Клеточные элементы сетчатого слоя представлены главным образом фибробластами.
В дерме вокруг сосудов микроциркуляторного русла - лимфатических капилляров и посткапиллярных венул присутствуют периваскулярные лимфо-идные узелки, схожие с узелками селезенки.
Периваскулярные лимфоидные узелки имеют центральную и мантийную зоны, в которых происходят пролиферация и дифференцировка лимфоцитов. Полагают, что благодаря этим узелкам в коже может быстро развиваться иммунная защитная реакция при поступлении в нее антигенов. Постоянное присутствие в эпидермисе и дерме иммунокомпетентных клеток, а также лимфоидных узелков свидетельствует о том, что кожа является не только местом реализации иммунологических процессов, но и активно участвует в них, играя роль одного из органов иммуногенеза.
В большинстве участков кожи человека в ее сетчатом слое располагаются кожные железы - потовые и сальные, а также корни волос.
В дерме некоторых участков кожи имеется пигмент, который располагается в цитоплазме меланодесмоцитов - клеток подлежащей к эпидермису соединительной ткани. Они имеют отростчатую форму и в отличие от меланоцитов эпидермиса не дают положительной ДОФА-реакции, т. е. они содержат, но не синтезируют пигмент. Предполагают, что пигмент поступает из эпидермиса и фагоцитируется меланодесмоцитами.
Дермальные меланоциты встречаются лишь в определенных участках - в области анального отверстия и в околососковых кружках.
Пигментный обмен в коже тесно связан с содержанием в ней витаминов (А, С, РР), а также зависит от эндокринных факторов - гормонов гипофиза, надпочечников и др.
Пучки коллагеновых волокон из сетчатого слоя дермы продолжаются в слое подкожной основы.
Подкожная основа (tela subcutanea), или гиподерма, не является частью кожи. Она богата жировой тканью и смягчает действие на кожу различных механических факторов. Поэтому гиподерма особенно хорошо развита в тех участках кожи, которые подвергаются сильным механическим воздействиям (подушечки пальцев, ступни и т. п.). Здесь подкожная основа полностью сохраняется даже при крайней степени истощения организма. Гиподерма обеспечивает некоторую подвижность кожи по отношению к нижележащим частям, что в значительной мере предохраняет ее от разрывов и других
механических повреждений. Скопление жировой ткани в гиподерме ограничивает теплоотдачу.
Васкуляризация. Кровеносные сосуды образуют в коже несколько сплетений, от которых отходят веточки, питающие различные ее части. Сосудистые сплетения залегают в коже на разных уровнях.
Различают глубокое и поверхностное артериальные сплетения, а также одно глубокое и два поверхностных венозных сплетения.
Артерии кожи берут начало из широкопетлистой сосудистой сети, расположенной между мышечными фасциями и подкожной основой (фасциальная артериальная сеть). От этой сети отходят сосуды, которые, пройдя слой подкожной жировой ткани, на границе ее с дермой разветвляются и образуют глубокую кожную артериальную сеть. От нее идут веточки, снабжающие кровью жировые дольки, потовые железы и волосы. Из глубокой кожной артериальной сети начинаются артерии, которые проходят сетчатый слой дермы и в основании сосочкового слоя распадаются на артериолы, образующие подсосочковую (поверхностную) артериальную сеть. От этой сети, в свою очередь, отходят более тонкие короткие веточки - терминальные артериолы, распадающиеся в сосочках на капилляры, имеющие форму шпилек, длина которых не превышает 0,4 мм. Терминальные артериолы, отходящие от подсосочковой сети, снабжают кровью группы сосочков. Характерно, что они не анастомозируют друг с другом. Этим можно объяснить, почему иногда покраснение или побледнение кожи происходит «пятнами». От подсосочковой сети отходят также артериальные сосуды к сальным железам и корням волос.
Капилляры сосочкового слоя, сальных желез и корней волос собираются в вены, впадающие в подсосочковые венозные сплетения. Различают два подсосочковых сплетения, лежащих одно за другим. Из них кровь поступает в кожное (глубокое) венозное сплетение, лежащее между дермой и гиподермой. В это же сплетение отводится кровь от жировых долек и потовых желез. Кожное сплетение соединяется с фасциальным венозным сплетением, от которого отходят более крупные венозные стволы.
В коже многочисленны артериоловенулярные анастомозы, особенно на кончиках пальцев рук и ног, в области ногтевого ложа. Они имеют прямое отношение к процессу терморегуляции.
Лимфатические сосуды кожи образуют два сплетения: поверхностное, лежащее ниже подсосочковых венозных сплетений, и глубокое, расположенное на границе с гиподермой.
Иннервация. Кожа иннервируется как ветвями цереброспинальных нервов, так и нервами автономной нервной системы. К цереброспинальной нервной системе принадлежат многочисленные чувствительные нервы, образующие в коже огромное количество чувствительных нервных сплетений. Нервы автономной нервной системы иннервируют в коже сосуды, гладкие миоциты и потовые железы.
Нервы в подкожной основе образуют основное нервное сплетение кожи, от которого отходят многочисленные стволики, дающие начало новым сплетениям, расположенным вокруг корней волос, потовых желез, жировых долек и в сосочковом слое дермы. Густое нервное сплетение сосоч-кового слоя посылает миелиновые и безмиелиновые нервные волокна в соединительную ткань и в эпидермис, где они образуют большое число чувствительных нервных окончаний. Нервные окончания распределены в
коже неравномерно. Они особенно многочисленны вокруг корней волос и в участках кожи с повышенной чувствительностью, например, на ладонях и подошвах, на лице, в области половых органов. К ним относятся свободные и несвободные нервные окончания: пластинчатые нервные тельца (тельца Фатера-Пачини), концевые колбы, осязательные тельца и осязательные клетки Меркеля (см. главу 10). Считается, что чувство боли передается расположенными в эпидермисе свободными нервными окончаниями, где они предположительно доходят до зернистого слоя, а также нервными окончаниями, лежащими в сосочковом слое дермы.
Вероятно, что свободные окончания одновременно являются терморецепторами. Чувство прикосновения (осязание) воспринимается осязательными тельцами и клетками Меркеля, а также нервными сплетениями вокруг корней волос. Осязательные тельца находятся в сосочковом слое дермы, осязательные клетки Меркеля - в базальном слое эпидермиса.
Чувство давления связано с наличием пластинчатых нервных телец Фатера-Пачини, лежащих глубоко в коже. К механорецепторам относятся и концевые колбы, расположенные, в частности, в коже наружных половых органов.
18.2. ПРОИЗВОДНЫЕ КОЖИ 18.2.1. Железы кожи
В коже человека находятся три вида желез: молочные, потовые и сальные. Поверхность железистого эпителия потовых и сальных желез примерно в 600 раз превышает поверхность эпидермиса. Эти кожные железы обеспечивают терморегуляцию (около 20 % тепла отдается организмом путем испарения пота), защиту кожи от повреждений (жировая смазка предохраняет кожу от высыхания, а также от мацерации водой и влажным воздухом), выделение из организма некоторых ферментов и продуктов обмена веществ (мочевина, мочевая кислота, аммиак и др.).
Потовые железы (gll. sudoriferae) встречаются почти во всех участках кожного покрова. Их количество достигает более 2,5 млн. Наиболее богата потовыми железами кожа лба, лица, ладоней и подошв, подмышечных и паховых складок. В этих местах на 1 см2 поверхности кожи открывается свыше 300 желез, тогда как в других участках кожи - 120-200 желез. Секрет потовых желез - пот - содержит 98 % воды и 2 % плотного остатка, который состоит из органических и неорганических веществ. Кроме воды, вместе с потом организм выделяет продукты белкового обмена и некоторые соли, например хлорид натрия. За сутки выделяется около 500-600 мл пота.
Потовые железы подразделяются на эккринные (мерокринные) и апокрин-ные. Эккринные железы распространены в коже повсеместно. Апокринные железы находятся лишь в определенных местах кожного покрова, напри-
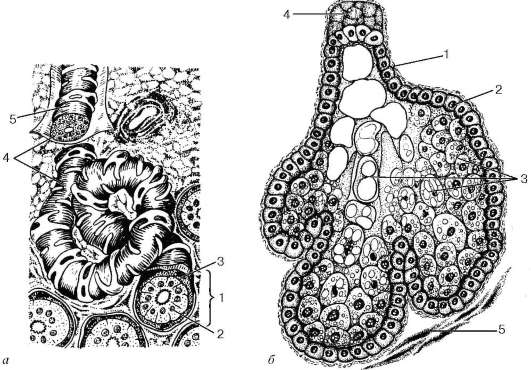
Рис. 18.4. Строение желез кожи (схема):
а - потовая железа: 1 - концевой отдел; 2 - железистые клетки; 3 - веретеновид-ные миоэпителиоциты; 4 - выводной проток; 5 - базальная мембрана; б - сальная железа: 1 - базальная мембрана; 2 - базальные себоциты; 3 - себоциты на разных стадиях жирового перерождения; 4 - выводной проток; 5 - гладкие миоциты мышцы, поднимающей волос
мер, в подмышечных впадинах, области заднего прохода, коже лба, больших половых губах. Развиваются они в период полового созревания организма и отличаются несколько большими размерами. Секрет их богаче органическими веществами, которые при разложении на поверхности кожи придают ему особенный, резкий запах. Разновидностью апокринных потовых желез являются железы век и железы, выделяющие ушную серу.
Потовые железы по строению являются простыми трубчатыми. Они состоят из длинного выводного протока и не менее длинного концевого отдела, закрученного в виде клубочка (рис. 18.4). Диаметр клубочка около 0,3-0,4 мм. Концевые отделы располагаются в глубоких частях сетчатого слоя на границе его с подкожной основой, а выводные протоки эккринных желез, пройдя через оба слоя дермы и эпидермис, открываются на поверхности кожи так называемой потовой порой. Выводные протоки многих апо-кринных желез не заходят в эпидермис и не образуют потовых пор, а впадают вместе с выводными протоками сальных желез в волосяные воронки.
Концевые отделы эккринных потовых желез имеют диаметр около 30-35 мкм. Они выстланы железистым эпителием, клетки которого бывают кубической или цилиндрической формы. Среди них различают светлые и
темные железистые клетки. В слабобазофильной цитоплазме светлых клеток постоянно встречаются капли жира, гранулы гликогена и пигмента. Обычно в них содержится высокоактивная щелочная фосфатаза. Светлые клетки выделяют воду и ионы металлов, темные клетки - органические макромолекулы (сиаломуцины). Кроме того, при потоотделении секреторные клетки выделяют вазоактивный пептид брадикинин, обладающий сосудорасширяющим действием, что способствует отдаче тепла организмом. Помимо секреторных клеток, на базальной мембране концевых отделов располагаются миоэпителио-циты. Сокращаясь, они способствуют выделению секрета.
Концевые отделы апокринных желез значительно более крупные: их диаметр достигает 150-200 мкм. Они состоят из железистых и миоэпителиаль-ных клеток. Железистые клетки имеют оксифильную цитоплазму с фосфо-липидными гранулами и не отличаются высокой активностью щелочной фосфатазы. В процессе секреции апикальные концы клеток разрушаются (апокриния) и входят в состав секрета. Функция апокринных потовых желез связана с функцией половых желез: в предменструальный и менструальный периоды и во время беременности секреция апокринных желез возрастает.
Переход концевого отдела в выводной проток совершается резко. Стенка выводного протока состоит из двухслойного кубического эпителия, клетки которого окрашиваются более интенсивно. Проходя через эпидермис, выводной проток эккринных потовых желез приобретает штопорообразный ход. Здесь его стенка образована плоскими клетками. Клетки выводных протоков потовых желез обладают слабо выраженной способностью реаб-сорбировать компоненты секрета.
Сальные железы (gll. sebaceae) наибольшего развития достигают в период полового созревания под влиянием половых гормонов: у мужчин - тестостерона, у женщин - прогестерона. В отличие от потовых желез сальные железы почти всегда связаны с волосами своим выводным протоком. Только там, где нет волос (губы, соски, слизистые оболочки наружных половых органов и др.), они располагаются самостоятельно. Больше всего сальных желез на голове, лице и верхней части спины. На ладонях и подошвах они отсутствуют. Секрет сальных желез (кожное сало) обладает бактерицидными свойствами и служит жировой смазкой для волос и эпидермиса кожи.
За сутки сальные железы человека выделяют около 20 г кожного сала. Секрет смягчает кожу, придает ей эластичность и облегчает трение соприкасающихся поверхностей кожи, а также препятствует развитию в ней микроорганизмов. В отличие от потовых сальные железы располагаются более поверхностно - в пограничных отделах сосочкового и сетчатого слоев дермы. Около одного корня волоса можно встретить 1-3 железы. Сальные железы являются простыми альвеолярными с разветвленными концевыми отделами (см. рис. 18.4, б). Секретируют они по голокринному типу.
Концевые отделы, диаметр которых колеблется от 0,2 до 2 мм, состоят из клеток - себоцитов, среди которых различают малодифференцированные, дифференцированные и погибающие формы. Малодифференцированные клетки, способные к пролиферации, составляют наружный ростковый (герминативный) слой концевого отдела, лежащий на базальной мембране. Вступившие
на путь дифференцировки клетки из этого слоя перемещаются внутрь концевого отдела. При этом они увеличиваются в объеме, и в них развивается агранулярная эндоплазматическая сеть, в которой синтезируются липиды, постепенно заполняющие цитоплазму. Затем происходит гибель себоцитов по механизму апоптоза. Клетки, распадаясь, превращаются в секрет - кожное сало. Последнее по выводному протоку поступает в воронку волоса и далее на поверхность его стержня и эпидермиса кожи. Выводной проток короткий. Стенка его состоит из многослойного плоского эпителия. Ближе к концевому отделу количество слоев в стенке протока уменьшается, эпителий становится кубическим и переходит в наружный ростковый слой концевого отдела.
18.2.2. Волосы
Волосы (pili) покрывают почти всю поверхность кожи. Наиболее плотно они расположены на голове. Длина волос колеблется от нескольких миллиметров до 1,5-2 м (редко), толщина - от 0,005 до 0,6 мм.
Различают три вида волос: длинные, к которым относятся волосы головы, бороды, усов, а также располагающиеся в подмышечных впадинах и на лобке; щетинистые - волосы бровей, ресниц, а также растущие в наружном слуховом проходе и в преддверии полости носа; пушковые, покрывающие остальные участки кожного покрова.
Развитие. Волосы развиваются на 3-м мес эмбриогенеза. Группы клеток эпидермиса в виде тяжей врастают в дерму, образуя волосяные фолликулы, из которых и происходит рост волос. Прежде всего волосы появляются в области бровей, подбородка и верхней губы. Несколько позднее они образуются в коже других участков тела.
Перед рождением или вскоре после него эти первые волосы (lanugo) выпадают (за исключением области бровей, век и головы) и заменяются новыми, пушковыми волосами (villus). В области бровей, век и головы позднее также происходит смена волос на более грубые - длинные и щетинистые.
В период полового созревания грубые волосы появляются в подмышечных впадинах, на лобке, а у мужчин, кроме того, на лице, иногда на груди, спине, бедрах и т. п. Волосы, образовавшиеся в период полового созревания, по характеру строения являются окончательными. В дальнейшем они подвергаются периодической смене.
Строение. Волосы являются эпителиальными придатками кожи. В волосе различают две части: стержень и корень. Стержень волоса находится над поверхностью кожи. Корень волоса скрыт в толще кожи и доходит до подкожной основы. Стержень длинных и щетинистых волос состоит из коркового, мозгового вещества и кутикулы; в пушковых волосах имеются только корковое вещество и кутикула. Корень волоса состоит из эпителиоцитов, находящихся на разных стадиях формирования коркового, мозгового вещества и кутикулы волоса.
Корень волоса располагается в волосяном мешке, стенка которого состоит из внутреннего и наружного корневых эпителиальных влагалищ. Все
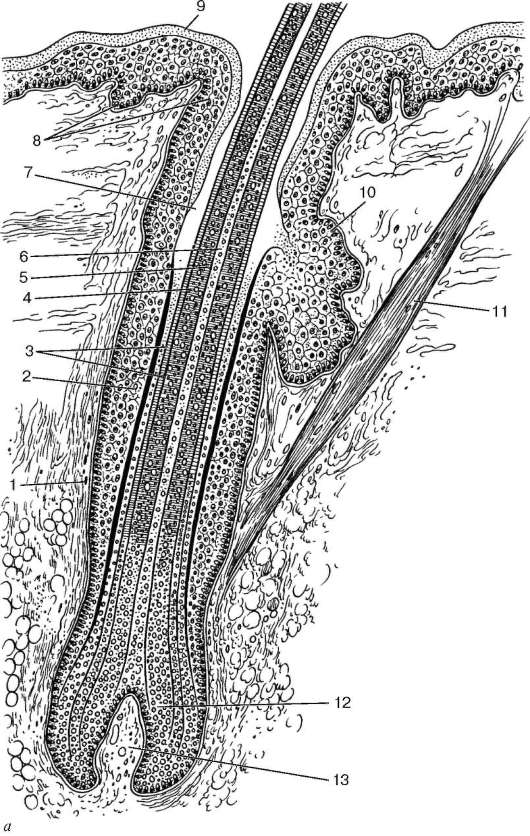
Рис. 18.5. Строение волоса:
а - схема (по А. Хэму и Д. Кормаку, с изменениями): 1 - дермальное корневое влагалище (волосяная сумка); 2 - наружное эпителиальное корневое влагалище; 3 - два слоя внутреннего эпителиального корневого влагалища; 4 - кутикула волоса; 5 - корковое вещество волоса; 6 - мозговое вещество волоса; 7 - волосяная воронка; 8 - базальный и шиповатый слои эпидермиса; 9 - роговой слой эпидермиса; 10 - сальная железа; 11 - мышца, поднимающая волос; 12 - волосяная луковица; 13 - волосяной сосочек
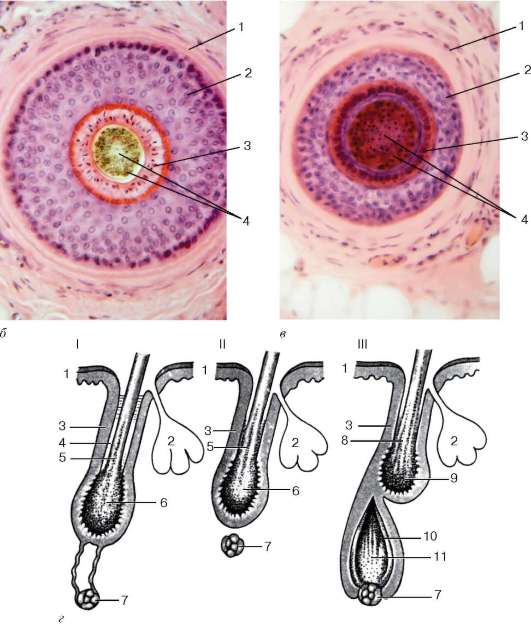
Рис. 18.5. Продолжение
б - поперечный срез корня волоса (микрофотография); в - поперечный срез волоса, удаленный участок от корня (микрофотография): 1 - дермальное корневое влагалище (волосяная сумка); 2 - наружное эпителиальное корневое влагалище; 3 - два слоя внутреннего эпителиального корневого влагалища; 4 - корковое и мозговое вещество волоса; г - схема фаз цикла волосяного фолликула: I - фаза катагена; II - фаза телогена; III - начало фазы анагена: 1 - эпидермис; 2 - сальная железа; 3 - наружное корневое эпителиальное влагалище; 4 - разрушающееся внутреннее корневое влагалище; 5 - вещество волоса; 6 - колба волоса; 7 - волосяной сосочек; 8 - вещество волоса в фазе телогена; 9 - колба волоса в фазе телогена; 10 - матрикс новообразующегося внутреннего эпителиального влагалища и волоса в фазе анаге-на; 11 - матрикс новообразующегося волоса
вместе они составляют волосяной фолликул. Фолликул окружен соединительнотканным дермальным корневым влагалищем (волосяной сумкой)
(рис. 18.5).
Корень волоса заканчивается расширением - волосяной луковицей. С ней сливаются оба эпителиальных корневых влагалища фолликула. Снизу в волосяную луковицу вдается соединительная ткань с капиллярами в виде дермального сосочка волоса. В месте перехода корня волоса в стержень эпидермис кожи образует углубление - волосяную воронку. Волос, выйдя из воронки, появляется над поверхностью кожи. Базальный и шиповатый слои эпидермиса воронки переходят в наружное корневое эпителиальное влагалище. Внутреннее корневое эпителиальное влагалище на этом уровне заканчивается. В волосяную воронку открывается проток одной или нескольких сальных желез. Ниже сальных желез в косом направлении проходит мышца, поднимающая волос (m. arrector pili).
Волосяная луковица (bulbus pili) является ростковой частью волоса (эпителиальным матриксом). Она состоит из эпителиальных клеток матрик-са, способных к размножению. Размножаясь, клетки волосяной луковицы передвигаются в мозговое и корковое вещество корня волоса, его кутикулу и во внутреннее эпителиальное влагалище. Таким образом, за счет клеток волосяной луковицы происходит рост самого волоса и его внутреннего корневого эпителиального влагалища. Питание волосяной луковицы осуществляется сосудами, расположенными в дермальном сосочке волоса (papilla dermalis pili). По мере того как клетки волосяной луковицы переходят в мозговое и корковое вещество, в кутикулу волоса и внутреннее корневое эпителиальное влагалище, в них усиливаются процессы ороговения. В более удаленных от волосяной луковицы участках клетки погибают и превращаются в роговые чешуйки. Поэтому строение корня волоса, его кутикулы и внутреннего эпителиального влагалища на разных уровнях неодинаковое. Наиболее интенсивно процесс ороговения клеток происходит в корковом веществе и кутикуле волоса. В результате в них образуется «твердый» кератин, который отличается по физическим и химическим свойствам от «мягкого» кератина эпидермиса кожи.
Твердый кератин более прочный. У человека из него, кроме того, построены ногти, а у животных - копыта, клювы, перья.
Твердый кератин плохо растворяется в воде, кислотах и щелочах; в его составе особенно много серосодержащей аминокислоты цистина. При образовании твердого кератина отсутствуют промежуточные стадии - накопление в клетках зерен кератогиалина.
Во внутреннем корневом эпителиальном влагалище и в мозговом веществе волоса процессы ороговения протекают так же, как в эпидермисе кожи, т. е. в клетках появляются зерна кератогиалина (трихогиалина). Среди кератиноцитов матрикса (базальных эпителиоцитов) волосяной луковицы и наружного корневого влагалища располагаются пигментные клетки - меланоциты. Они синтезируют пигмент меланин в двух формах, от которых зависит цвет волос, - в форме эумеланина, имеющего цвет от коричневого до черного, и в форме феомеланина - желтого и рыжего цвета.
Мозговое вещество волоса (medulla pili) хорошо выражено только в длинных и щетинистых волосах. Мозговое вещество состоит из клеток полигональной формы, лежащих друг на друге в виде монетных столбиков. Они содержат ацидофильные, блестящие гранулы трихогиалина, мелкие пузырьки газа и небольшое количество зерен пигмента. Процессы ороговения в мозговом веществе протекают медленно, поэтому примерно до уровня протоков сальных желез мозговое вещество состоит из не полностью ороговевших эпителиоцитов, в которых обнаруживаются пикнотические ядра или их остатки. Только выше указанного уровня клетки подвергаются полному ороговению. С возрастом процессы ороговения в мозговом веществе волоса усиливаются, при этом в клетках снижается количество пигмента - волосы седеют.
Корковое вещество волоса (cortex pili) составляет основную его массу. Процессы ороговения в корковом веществе протекают интенсивно и без промежуточных стадий. На протяжении большей части корня и всего стержня волоса корковое вещество состоит из плоских роговых чешуек. Только в области шейки волосяной луковицы в этом веществе встречаются не полностью ороговевшие клетки с овальными ядрами. В роговых чешуйках содержатся твердый кератин, зерна пигмента и пузырьки газа. Чем лучше в волосе развито корковое вещество, тем он прочнее, эластичнее.
Кутикула волоса (cuticula pili) непосредственно прилежит к корковому веществу. Ближе к волосяной луковице она представлена цилиндрическими клетками - кутикулярными эпителиоцитами, лежащими перпендикулярно к поверхности коркового вещества. В более отдаленных от луковицы участках эти клетки приобретают наклонное положение и превращаются в роговые чешуйки, накладывающиеся друг на друга в виде черепицы. Эти чешуйки содержат твердый кератин, но полностью лишены пигмента.
Внутреннее корневое эпителиальное влагалище (vagina radicularis epithelialis interna) является производным волосяной луковицы. В нижних отделах корня волоса оно переходит в вещество волосяной луковицы, а в верхних отделах на уровне протоков сальных желез исчезает. В нижних отделах во внутреннем эпителиальном влагалище различают три слоя: кутикулу, внутренний (гранулосодержащий) эпителиальный слой Хаксли и наружный (бледный) эпителиальный слой Хенле. В средних и верхних отделах корня волоса все эти три слоя сливаются, и здесь внутреннее корневое эпителиальное влагалище состоит только из полностью ороговевших клеток, содержащих мягкий кератин.
Наружное корневое эпителиальное влагалище (vagina radicularis epithelialis externa) образуется из базального и шиповатого слоев эпидермиса кожи, которые продолжаются вплоть до волосяной луковицы. Клетки его богаты гликогеном. Это влагалище постепенно истончается и переходит в волосяную луковицу.
Дермальное корневое влагалище (vagina radicularis dermalis), или волосяная сумка, - соединительнотканная оболочка волоса. В ней различают наружный - продольный слой волокон и внутренний - циркулярный слой во локон.
Мышца, поднимающая волос (m. arrector pili), состоит из гладких мышечных клеток. У щетинковых, пушковых волос, волос бороды и подмышечных впадин она отсутствует или развита слабо. Мышца залегает в косом направлении и одним концом вплетается в волосяную сумку, а другим - в сосочковый слой дермы. При ее сокращении корень принимает перпендикулярное направление к поверхности кожи, и в результате этого стержень волоса несколько приподнимается над кожей (волосы «встают дыбом»), что наблюдается у многих животных.
Смена волос. Волосяные фолликулы в процессе своей жизнедеятельности проходят через повторяющиеся циклы. Каждый из них включает период гибели старого волоса и периоды образования и роста нового волоса, что обеспечивает смену волос. У человека в зависимости от возраста, области кожи, а также от воздействия различных внешних факторов цикл продолжается от 2 до 5 лет. В нем различают три фазы - катагена, телогена и анаге-на. Фаза катагена, продолжающаяся 1-2 нед, характеризуется прекращением митотической активности в эпителиальном матриксе (луковице) волоса, которая превращается в так называемую волосяную колбу, состоящую из подвергшихся ороговению клеток. Колба отделяется от волосяного сосочка и вместе с волосом поднимается кверху, в результате чего волосяной фолликул становится короче. В нем разрушается внутреннее эпителиальное влагалище, тогда как наружное эпителиальное влагалище сохраняется, образуя мешочек, в основании которого остаются стволовые клетки, дающие в дальнейшем начало развитию и росту нового волоса. Фаза телогена продолжается 2-4 мес. Это период покоя, когда оставшаяся часть корня волоса и колба могут сохраняться в фолликуле до следующего цикла. Фаза анагена составляет основную часть времени цикла волосяного фолликула - около 2- 5 лет. Она включает период формирования нового фолликула и рост волоса, что напоминает морфогенез фолликулов в эмбриогенезе в коже плода. При этом стволовые клетки в основании наружного эпителиального влагалища начинают интенсивно делиться, образуя конусообразный эпителиальный матрикс, который постепенно обрастает нижерасположенный волосяной сосочек. Пролиферация клеток матрикса приводит к образованию нового волоса, кутикулы и внутреннего эпителиального влагалища. По мере роста нового волоса старый волос вытесняется наружу. Процесс заканчивается выпадением старого и появлением на поверхности кожи нового волоса.
18.2.3. Ногти
Ноготь (ungus) - это роговой эпителиальный придаток кожи. Развитие ногтей начинается на 3-м мес внутриутробного развития. Прежде чем появиться ногтю, на месте его будущей закладки образуется так называемое ногтевое ложе. При этом эпителий, покрывающий дорсальные поверхности терминальных фаланг пальцев рук и ног, утолщается и несколько погружается в подлежащую соединительную ткань. В более поздней стадии из эпителия проксимальной части ногтевого ложа начинает вырастать сам ноготь.
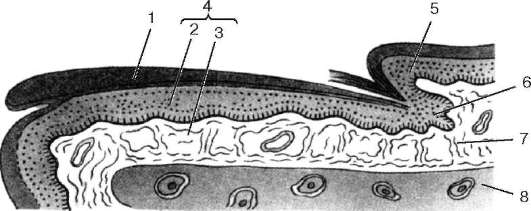
Рис. 18.6. Строение ногтя (по Е. Ф. Котовскому):
1 - ногтевая пластинка; 2 - подногтевая пластинка (ростковый и шиповатый слой эпидермиса); 3 - сосочковый слой дермы; 4 - ногтевое ложе; 5 - задний ногтевой валик; 6 - матрикс ногтя; 7 - соединительнотканные волокна; 8 - кость фаланги пальца
Вследствие медленного роста (около 0,25-1 мм в неделю) только к последнему месяцу беременности ноготь достигает кончика пальца.
Ноготь - роговая пластинка, лежащая на ногтевом ложе (рис. 18.6). Ногтевое ложе состоит из эпителия и соединительной ткани. Эпителий ногтевого ложа - подногтевая пластинка (hyponychium), представлен ростковым слоем эпидермиса. Лежащая на нем ногтевая пластинка является его роговым слоем. Ногтевое ложе с боков и у основания ограничено кожными складками - ногтевыми валиками (задним и боковыми). Ростковый слой их эпидермиса переходит в эпителий ногтевого ложа, а роговой слой надвигается на ноготь сверху (особенно на его основание), образуя так называемую надногтевую пластинку, или кожицу (eponychium). Между ногтевым ложем и ногтевыми валиками имеются ногтевые щели (задняя и боковые). Ногтевая (роговая) пластинка своими краями вдается в эти щели. Она образована плотно прилегающими друг к другу роговыми чешуйками, в которых содержится твердый кератин. Ногтевая (роговая) пластинка подразделяется на корень, тело и край. Корнем ногтя называется задняя часть ногтевой пластинки, лежащей в задней ногтевой щели. Лишь небольшая часть корня выступает из-за задней ногтевой щели (из-под заднего ногтевого валика) в виде беловатого участка полулунной формы - луночки ногтя. Остальная часть ногтевой пластинки, расположенная на ногтевом ложе, составляет тело ногтя. Свободный конец ногтевой пластинки - край - выступает за пределы ногтевого ложа.
Соединительная ткань ногтевого ложа содержит большое количество волокон, часть которых располагается параллельно ногтевой пластинке, а часть - перпендикулярно к ней. Последние достигают кости конечной фаланги пальца и соединяются с ее надкостницей. Соединительная ткань ногтевого ложа образует продольные складки, в которых проходят кровеносные сосуды. Участок эпителия ногтевого ложа, на котором лежит корень ногтя, является местом его роста и носит название матрикс ногтя. В последнем происходит процесс размножения и ороговения клеток. Образующиеся
роговые чешуйки (онихоциты) смещаются в ногтевую (роговую) пластинку, которая в результате этого увеличивается в длине, т. е. происходит рост ногтя. Соединительная ткань ногтевой матрицы образует сосочки, в которых располагаются многочисленные кровеносные сосуды.
Реактивность и регенерация кожи. Реактивные свойства кожи отчетливо проявляются при действии факторов внешней и внутренней среды. В эпидермисе хорошо выражены процессы клеточного обновления (физиологическая регенерация). Высокий уровень пролиферации эпителиоцитов отмечается в области матрикса волоса, а также в концевых отделах потовых и сальных желез. Выделяются две разновидности стволовых клеток эпителиального дифферона. Одни из них располагаются в базальном слое эпидермиса и являются источником развития дифферона ороговевающих эпи-телиоцитов эпителиально-пролиферативной единицы в физиологических условиях. Эти клетки обеспечивают также регенерацию при поверхностных повреждениях эпителиального покрова кожи.
Вторая разновидность стволовых клеток эпителия локализуется в наружном корневом эпителиальном влагалище, непосредственно под устьями протоков сальных желез. Эти клетки являются источником развития поверхностных эпителиоцитов, эпителиоцитов волоса и его эпителиальных влагалищ, эпителиоцитов желез, а также участвуют в регенерации кожи при ее глубоких и обширных повреждениях.
При травматических и ожоговых повреждениях кожи репаративная регенерация происходит путем стягивания краев раны за счет развития грануляционной ткани и эпителизации раневой поверхности. Исходом регенерации кожи является, как правило, образование регенератов в виде соединительнотканного рубца, не содержащего производных кожи (желез, волос) - регенерата рубцового типа. Реже образуется регенерат кожного типа, по строению приближающийся к интактной коже. Важную камбиальную роль в эпителизации раневой поверхности играют малодифференцированные клетки концевых отделов потовых желез и наружных корневых эпителиальных влагалищ волоса. Это учитывается при взятии кожных лоскутов для трансплантации - после срезания тонкого лоскута кожи возникшая раневая поверхность достаточно быстро покрывается новым эпидермисом.
18.2.4. Молочные железы
Традиционно гистофизиологию молочных желез рассматривают при изучении женской репродуктивной системы, так как железы обеспечивают питание ребенка после его рождения, когда прекращается получение им питательных веществ через плацентарную кровь. Однако молочные железы являются производными кожи. Они представляют собой видоизмененные кожные потовые железы.
Развитие. Молочные железы закладываются у зародыша на 6-7-й нед в виде двух уплотнений эпидермиса («молочные линии»), тянущихся вдоль туловища. Из этих утолщений формируются так называемые «молочные
точки», из которых в подлежащую мезенхиму врастают плотные эпителиальные тяжи. Затем они, разветвляющиеся на своих дистальных концах, формируют эпителиальные зачатки молочных желез, в которых находятся стволовые клетки для концевых секреторных отделов.
Закладка желез у мальчиков и девочек происходит одинаково. Несмотря на неполное развитие желез, у новорожденных обнаруживается секреторная деятельность, которая продолжается обычно в течение недели и затем прекращается у мальчиков навсегда, а у девочек железы до наступления половой зрелости находятся в покоящемся состоянии. В течение детского возраста у обоих полов происходит развитие млечных ходов.
С наступлением половой зрелости возникают резкие половые различия в темпах развития молочных желез. У мальчиков образование новых ходов замедляется и затем прекращается. У девочек развитие железистых трубок значительно ускоряется, и к началу менструаций появляются первые концевые секреторные отделы. В течение овариально-менструального цикла происходят циклические изменения концевых отделов: в первые дни после менструации они подвергаются обратному развитию, а во второй половине цикла вновь появляются. Однако у девушек молочная железа не достигает окончательного развития, которое наблюдается только при беременности, в период лактации.
Строение. У половозрелой женщины каждая молочная железа состоит из 15-20 отдельных железок, разделенных прослойками рыхлой соединительной и жировой ткани. Эти железы по своему строению являются сложными альвеолярными, и их выводные протоки открываются на вершине соска. Выводные протоки переходят в расширенные млечные синусы (sinus lactiferi), служащие резервуарами, в которых накапливается молоко, продуцируемое в альвеолах. В млечные синусы впадают многочисленные ветвящиеся и анастомозирующие млечные протоки (ductus lactiferi), заканчивающиеся до наступления периода лактации тонкими слепыми эпителиальными трубочками - альвеолярными млечными ходами (ductuli alveolares lactiferi). Во время беременности и в лактирующей молочной железе стволовые клетки в составе этих терминальных ветвлений являются источником развития клеточных дифферонов, входящих в состав секреторного отдела - альвеол (рис. 18.7, 18.8).
Млечные синусы открываются на верхушке соска, представляющего собой утолщение кожи. Его эпидермис сильно пигментирован, в базаль-ную часть эпителиального слоя вдаются длинные и часто разветвляющиеся сосочки дермы, в которых располагаются многочисленные инкапсулированные нервные окончания. Обилие этих чувствительных рецепторных окончаний связано с тем, что выделение молока обусловлено рефлексом на раздражение этих окончаний актом сосания. Полного развития молочная железа достигает во время беременности.
Выработка молока происходит в альвеолах, имеющих вид округлых или слегка вытянутых пузырьков. Стенку альвеолы образуют диффероны молочных экзокриноцитов (галактоцитов) и миоэпителиоцитов. Молочные экзо-криноциты соединяются с помощью замыкающих пластинок и десмосом,
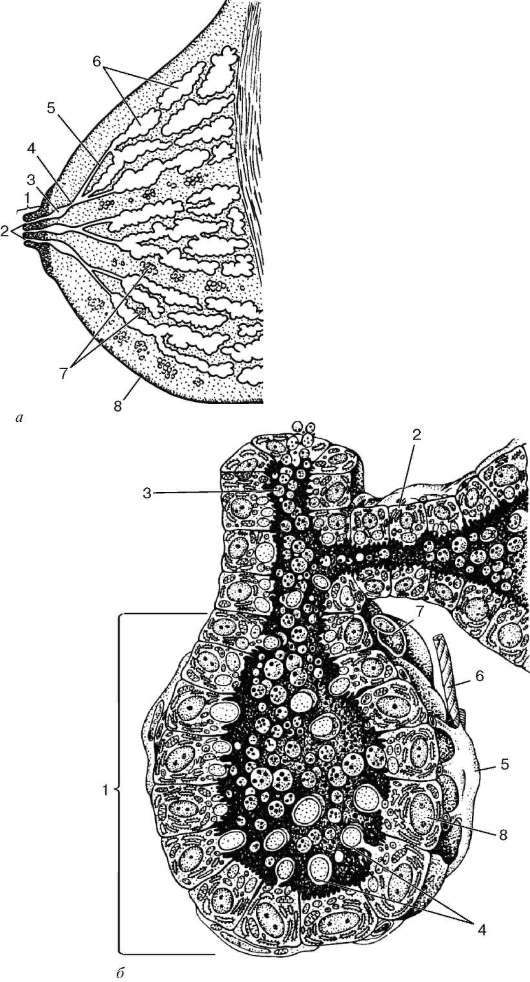
Рис. 18.7. Молочная железа: а - схема строения железы: 1 - сосок; 2 - выводные протоки; 3 - млечные синусы; 4 - млечные протоки; 5 - млечные ходы; 6 - альвеолы; 7 - прослойки рыхлой соединительной ткани с жировыми клетками; 8 - кожа; б - схема ультрамикроскопического строения альвеолы лактирующей железы (по Р. Крстичу; с изменениями): 1 - альвеола; 2 - млечный ход; 3 - внутридольковый проток; 4 - апокринная секреция; 5 - мио-эпителиоцит; 6 - нервное волокно; 7 - капилляр; 8 - галактоцит
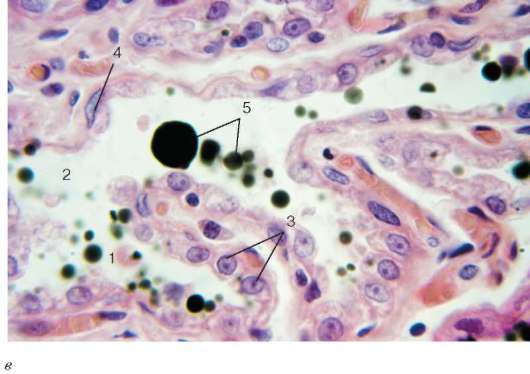
Рис. 18.7. Продолжение
в - молочная железа в стадии лактации (микропрепарат): 1 - альвеола; 2 - млечный ход; 3 - галактоцит; 4 - миоэпителиоцит; 5 - секрет
располагаются в один слой на базальной мембране. На апикальной поверхности клеток находятся небольшие микроворсинки. В молочных экзокри-ноцитах хорошо развита гранулярная и агранулярная эндоплазматиче-ская сеть, образованная канальцами и цистернами. В комплексе Гольджи, хорошо развитом в галактоцитах, завершаются образование и конденсация казеина, а также синтез лактозы, чему способствует наличие специального фермента - лактосинтетазы. Кроме того, в экзокриноцитах обнаруживаются микротрубочки и микрофиламенты, особенно в апикальных частях цитоплазмы, которые способствуют транспортировке секреторных продуктов к апикальному краю молочного экзокриноцита.
При выделении синтезированных компонентов молока жировые капли, достигающие больших размеров, перемещаются к апикальной мембране и, облекаясь ею, вытягиваются над краем галактоцита. По мере экструзии жировая капля вместе с окружающей ее частью апикальной мембраны отрывается и попадает в полость альвеолы (см. рис. 18.7). В полости альвеолы капельки жира, раздробляясь, превращаются в тонкую эмульсию, к которой примешиваются казеин, лактоза и соль, т. е. формируется молоко, которым заполняется полость альвеолы. Молоко - сложная водная эмульсия, в состав которой входят жировые капельки (триглицериды молока, а также жирные кислоты, являющиеся предшественниками триглицеридов), белки (из них специфическими для молока являются казеин, а также лактоглобулины и лактоальбумины), углеводы (в том числе специфический для молока дисаха-рид - лактоза, или молочный сахар), соли и вода. У основания лактоцитов (как и в других эктодермальных железах, например в потовых или слюнных) обнаруживаются миоэпителиальные клетки, охватывающие своими отрост-
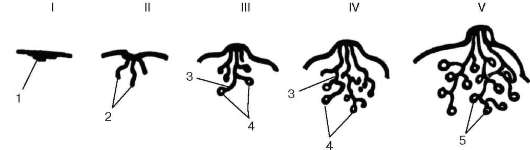
Рис. 18.8. Морфогенез молочной железы (схема):
I - закладка железы в эмбриогенезе; II - развитие выводных протоков в эмбриогенезе; III - период наступления половой зрелости (начало развития секреторных отделов); IV - нелактирующая молочная железа взрослой женщины; V - лактирующая молочная железа. 1 - утолщение эпидермиса («молочная точка»); 2 - эпителиальные тяжи; 3 - формирующиеся выводные протоки; 4 - альвеолы; 5 - альвеолы лакти-рующей железы
ками альвеолу снаружи. Цитоплазма содержит актомиозиновые комплексы, темное ядро. Опорожнению альвеолы и переходу молока в млечные ходы способствует сокращение миоэпителиальных клеток.
По окончании периода лактации молочная железа претерпевает инволюцию, однако часть альвеол, образовавшихся во время предыдущей беременности, сохраняется.
Строение выводных протоков. Стенки внутридольковых протоков образованы кубическим эпителием и содержат миоэпителиоциты. Млечные протоки выстланы двухслойным кубическим эпителием и миоэпителиоцитами. Эпителий в синусах - трехслойный, а на вершине соска становится многослойным плоским. Вдоль млечных протоков и синусов в соединительной и жировой ткани проходят пучки гладкой мышечной ткани, сокращения которой способствуют молокоотдаче.
Возрастные изменения. У девочек в период полового созревания начинается интенсивное развитие молочных желез. Из разветвленных железистых трубок дифференцируются секреторные отделы - альвеолы. В течение полового цикла секреторная активность железистых клеток повышается в период овуляции и снижается во время менструации.
После прекращения образования овариальных гормонов с наступлением климактерического периода или после кастрации молочная железа претерпевает обратное развитие.
Регуляция функции молочных желез. В онтогенезе зачатки молочных желез начинают интенсивно развиваться после наступления полового созревания, когда в результате значительного увеличения образования эстрогенов устанавливаются менструальные циклы и происходит развитие вторичных признаков женского пола. Но полного развития и окончательной дифференцировки молочные железы достигают только во время беременности. С момента имплантации зародыша в эндометрий матки в дольках молочной железы разрастаются млечные ходы, на концах которых форми-
руются альвеолы. Во второй половине беременности в альвеолах начинается секреция молозива (colostrum). Интенсивная секреция полноценного молока устанавливается в первые дни после рождения ребенка.
Регуляция деятельности функционирующей молочной железы осуществляется двумя основными гормонами - аденогипофизарным пролактином (лактотропный гормон), который стимулирует секреторную функцию железистых клеток альвеол (галактоцитов), и окситоцином - гипоталамическим нейрогормоном, который способствует выделению молока по системе выводных протоков. В свою очередь, секреция пролактина возбуждается тем же гипоталамическим тиролиберином, который активирует тиротропную функцию гипофиза, а угнетается дофамином - нейроамином, образующимся в туберальных ядрах медиобазального гипоталамуса и переносимым с током портальной крови в переднюю долю гипофиза, где этот нейроамин действует прямо на лактотропоциты, блокируя секрецию пролактина.
Контрольные вопросы
1. Источники и основные этапы развития кожи и ее производных.
2. Клеточные слои эпидермиса. Роль клеток разных дифферонов в гисто-физиологии кожи.
3. Источники и характеристика физиологической и репаративной регенерации кожи и ее производных.
4. Структурно-функциональная единица молочной железы: клеточные диффероны, гистофизиология.