Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 9. МЫШЕЧНЫЕ ТКАНИ
Мышечными тканями (textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма (сердце, язык, кишечник и др.).
Свойством сокращаться с изменением формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
9.1. ОБЩАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА И КЛАССИФИКАЦИЯ
Основные морфологические признаки элементов мышечных тканей - удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов - специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы - миофиламенты или миофи-бриллы, обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков - актина и миозина, при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин - белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко снижается).
Классификация. В основу классификации мышечных тканей положены два принципа - морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы.
Первая подгруппа - поперечнополосатые (исчерченные) мышечные ткани (textus muscularis striatus). В цитоплазме их элементов миозиновые филамен-
ты постоянно полимеризованы, образуют с актиновыми нитями постоянно существующие миофибриллы. Последние организованы в характерные комплексы - саркомеры. В соседних миофибриллах структурные субъединицы саркомеров расположены на одном уровне и создают поперечную исчер-ченность.
Вторая подгруппа - гладкие (неисчерченные) мышечные ткани (textus muscularis nonstriatus). Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине (гладкими) нитями.
В соответствии с гистогенетическим принципом в зависимости от источников развития (эмбриональных зачатков) мышечные ткани и мышечные элементы подразделяются на: соматические (миотомные), целомические (из миоэпикардиальной пластинки висцерального листка спланхнотома), мезенхимные (из десмального зачатка в составе мезенхимы), нейральные (из нервной трубки), эпидермальные (из кожной эктодермы и из прехор-дальной пластинки).
9.2. ПОПЕРЕЧНОПОЛОСАТЫЕ МЫШЕЧНЫЕ ТКАНИ
Имеется две основные разновидности поперечнополосатых (исчерченных) тканей - скелетная (миотомная) и сердечная (целомическая).
9.2.1. Скелетная мышечная ткань
Гистогенез. Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани (textus muscularis striatus sceletalis) являются стволовые клетки миотомов - промиобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтон-ных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты - мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл - миофибрилл (рис. 9.1). В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндо-
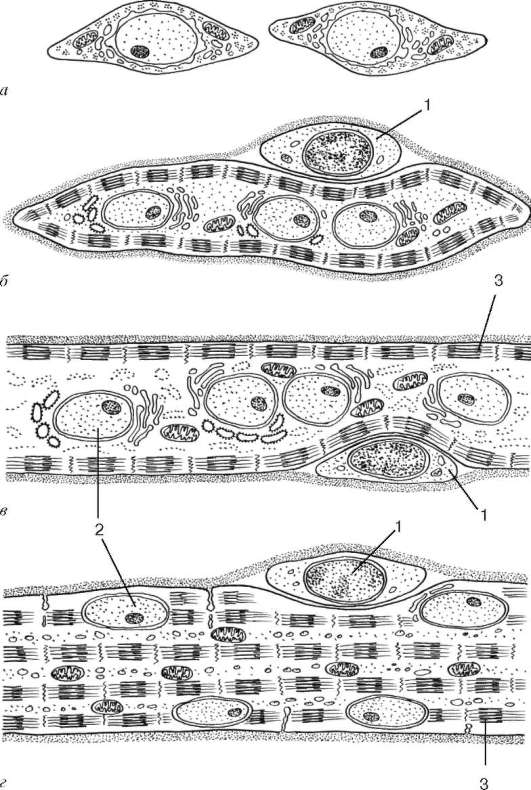
Рис. 9.1. Гистогенез скелетной мышечной ткани (по А. А. Клишову):
а - промиобласты; б - миосимпласт; в - мышечная трубочка; г - зрелое мышечное
волокно. 1 - миосателлитоцит; 2 - ядро миосимпласта; 3 - миофибриллы
плазматическая сеть в значительной степени редуцируется. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты. Эти клетки располагаются на поверхности миосимпластов. Миосателлитоциты, размножаясь, сливаются с миосимпластами, участвуя таким образом в создании оптимального ядерно-
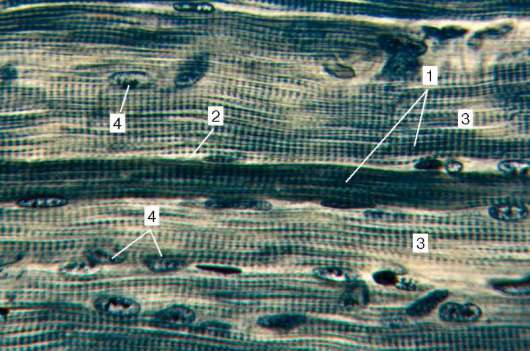
Рис. 9.2. Строение поперечнополосатой мышечной ткани (микрофотография):
1 - мышечные волокна; 2 - сарколемма; 3 - саркоплазма и миофибриллы; 4 - ядра
миосимпласта. Окраска - железный гематоксилин
саркоплазменного отношения, необходимого для синтеза специфических белков симпласта.
Строение. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосател-литоцитов, покрытых общей базальной мембраной (рис. 9.2-9.4). Длина всего волокна может измеряться сантиметрами при толщине 50-100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Строение миосимпласта. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под плазмолеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч (см. рис. 9.2). У полюсов ядер располагаются органеллы общего значения - комплекс Гольджи и небольшие фрагменты агранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно (см. рис. 9.3).
Саркомер - структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски с неодинаковым лучепреломлением (анизотропные А-диски и изотропные I-диски). Каждая миофибрил-ла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети - саркоплазмати-ческой сети. Соседние саркомеры имеют общую пограничную структуру - Z-линию (рис. 9.5). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы актиновых филаментов. От соседних Z-линий акти-новые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина
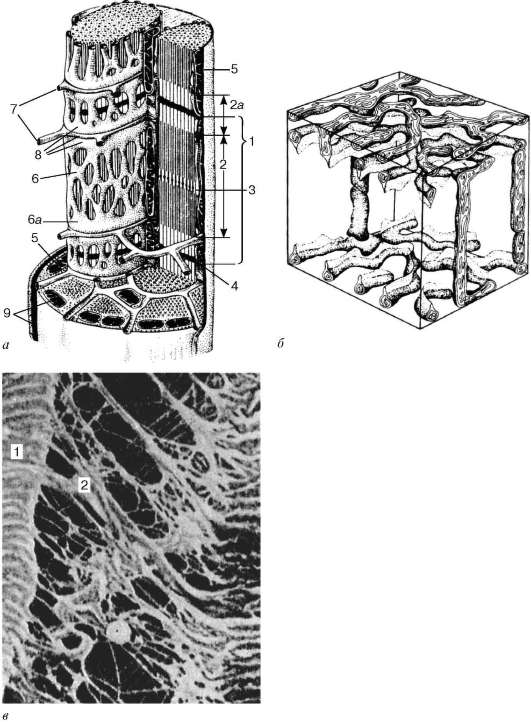
Рис. 9.3. Схема ультрамикроскопического строения миосимпласта (по Р. В. Крстичу, с изменениями) (а): 1 - саркомер; 2 - анизотропный диск (полоса А); 2а - изотропный диск (полоса I); 3 - линия М (мезофрагма) в середине анизотропного диска; 4 - линия Z (телофрагма) в середине изотропного диска; 5 - митохондрии; 6 - сар-коплазматическая сеть; 6а - конечная цистерна; 7 - поперечная трубочка (Т-трубочка); 8 - триада; 9 - сарколемма; б - схема пространственного расположения митохондрий в симпласте. Верхняя и нижняя плоскости рисунка ограничиваютанизотропныйдисксарко-мера (по Л. Е. Бакеевой, В. П. Скулачеву, Ю. С. Ченцову); в - эндомизий. Сканирующая электронная микрофотография, увеличение 2600 (препарат Ю. А. Хорошкова): 1 - мышечные волокна; 2 - коллагеновые фибриллы
фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомиозина. Она образует в сечении М-линию. В узлах этой М-линии закреплены концы мио-зиновых филаментов. Другие их концы направляются в сторону Z-линий и
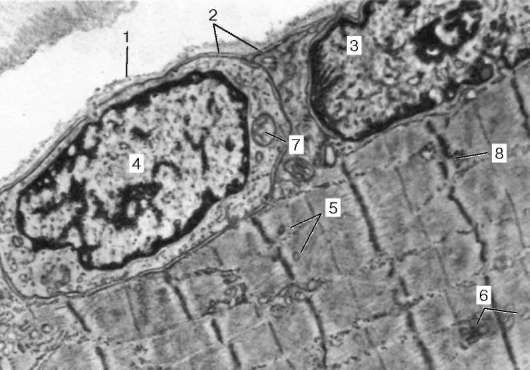
Рис. 9.4. Поверхностный участок миосимпласта и миосателлитоцит. Электронная микрофотография, увеличение 10 000 (препарат В. Л. Горячкиной и С. Л. Кузнецова): 1 - базальная мембрана; 2 - плазмолемма; 3 - ядро миосимпласта; 4 - ядро миоса-теллитоцита; 5 - миофибриллы; 6 - канальцы агранулярной эндоплазматической (саркоплазматической) сети; 7 - митохондрии; 8 - гликоген
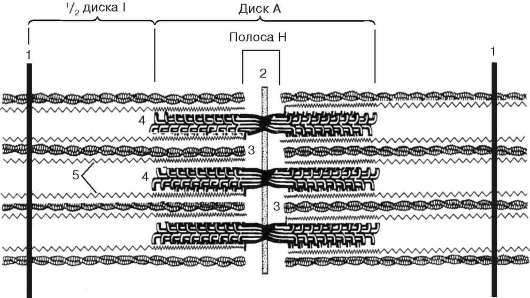
Рис. 9.5. Саркомер (схема):
1 - линия Z; 2 - линия М; 3 - филаменты актина; 4 - филаменты миозина; 5 - фибриллярные молекулы титина (по Б. Албертс, Д. Брей, Дж. Льюис и др., с изменениями)
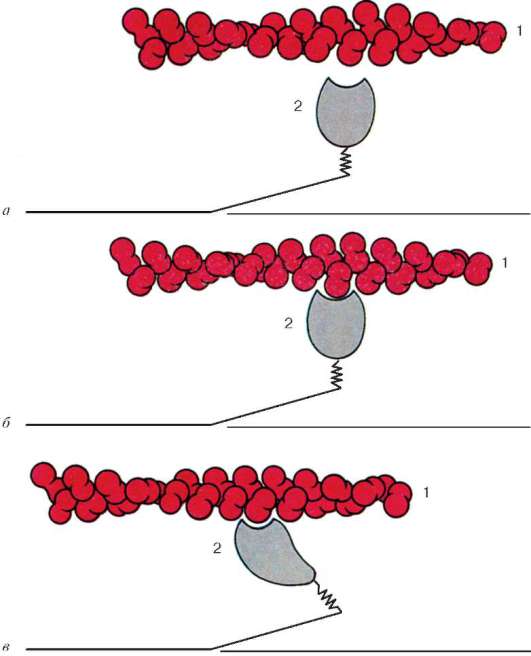
Рис. 9.6. Конформационные изменения, влекущие за собой взаимное смещение филаментов актина и миозина:
а-в - последовательные изменения пространственных отношений. 1 - актин; 2 - головка молекулы миозина (по Б. Албертс, Д. Брей, Дж. Льюис и соавт., с изменениями)
располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Молекулы миозина имеют длинный хвост и на одном из его концов две головки. При повышении концентрации ионов кальция в области присоединения головок (шарнирный участок) молекула изменяет свою конфигурацию (рис. 9.6). При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков - тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в ее кортикальном слое, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна.
Источником ионов кальция служат цистерны агранулярной эндоплазма-тической сети. Они вытянуты вдоль миофибрилл около каждого саркоме-ра и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или латеральные (L) цистерны.
С поверхности в глубину миосимпласта плазмолемма образует длинные трубочки, идущие поперечно (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда миосимпласт получает сигнал о начале сокращения, он перемещается по плазмолемме в виде потенциала действия и распространяется на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в канальцах сети и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет превращения АТФ в АДФ. Роль АТФ-азы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается и участвует в биохимических реакциях.
Миосателлитоциты. Эти малодифференцированные клетки являются источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются (см. рис. 9.1, 9.4). Миосателлитоциты одноядерны, их темные ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Активность ферментов в них различна, и представлены они в различных изомерных формах. Различно в них и содержание дыхательных ферментов - гликолитических и окислительных.
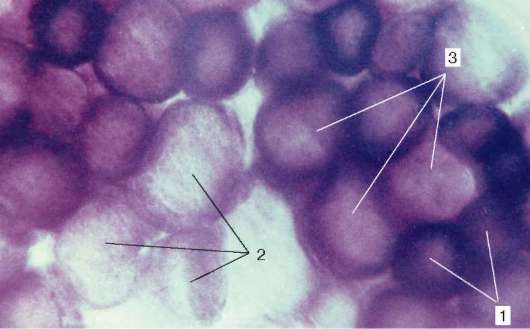
Рис. 9.7. Активность сукцинатдегидрогеназы в мышечных волокнах разного типа (препарат В. Ф. Четвергова, обработка по Нахласу и соавт.): 1 - высокая; 2 - низкая; 3 - средняя
По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные, что определяется молекулярной организацией миозина. Среди его изоформ существуют две основные - «быстрая» и «медленная». При постановке гистохимических реакций их идентифицируют по АТФ-азной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они богаты гликогеном, в них меньше миоглобина, поэтому их называют белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными.
Наряду с белыми и красными существуют и промежуточные волокна. В составе большинства скелетных мышц волокна разного гистохимического типа располагаются мозаично (рис. 9.7).
Свойства мышечных волокон меняются при изменении нагрузок - спортивных, профессиональных, а также в экстремальных условиях (невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
Регенерация. Ядра миосимпластов делиться не могут, так как в саркоплазме отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки сливаются с миосимпластами. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагмен-
ты фагоцитируются макрофагами. Восстановление тканей осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролиферации миосателлитоцитов. В симпласте активизируются гранулярная эндоплазматическая сеть и комплекс Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмо-леммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Миосателлитоциты, сохранившиеся рядом с повреждением, делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют новые миотубы, которые развиваются в мышечных волокна.
9.2.2. Скелетная мышца как орган
Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани - эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него (см. рис. 9.3, в), что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по несколько мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Васкуляризация. Артерии вступают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви пятого-шестого порядка образуют в перимизии артериолы. В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тучных клеток, принимающих участие в регуляции проницаемости сосудистой стенки.
Иннервация. В мышцах выявлены миелинизированные эфферентные (двигательные), афферентные (чувствительные), а также немиелинизиро-ванные вегетативные нервные волокна. Отросток нервной клетки, приносящий импульс от мотонейрона спинного мозга, ветвится в перимизии. Каждая его ветвь проникает сквозь базальную мембрану и у поверхности симпласта на плазмолемме образует терминали, участвуя в организации так называемой моторной бляшки (см. главу 10, рис. 10.18). При поступлении
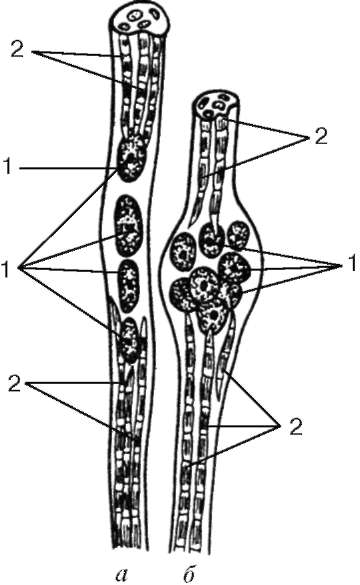
Рис. 9.8. Фрагмент мышечного веретена, содержащего мышечные волокна с ядерной цепочкой (а) и с ядерной сумкой (б) (схема по Г. С. Катинасу): 1 - ядра; 2 - миофибриллы (органеллы общего значения не показаны)
нервного импульса из терминалей выделяется ацетилхолин - медиатор, который вызывает возбуждение (потенциал действия), распространяющееся отсюда по плазмолемме миосимпласта.
Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом.
Группа мышечных волокон, ин-нервируемых одним мотонейроном, называется нервно-мышечной единицей. Мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Чувствительные нервные окончания располагаются не на рабочих (экстрафузальных) мышечных волокнах, а связаны со специализированными мышечными волокнами в так называемых мышечных вере-
тенах (с интрафузальными мышечными волокнами), которые расположены в перимизии.
Интрафузальные мышечные волокна. Интрафузальные мышечные волокна веретен значительно тоньше рабочих. Существует два их вида - волокна с ядерной сумкой и волокна с ядерной цепочкой (рис. 9.8). Ядра в тех и в других округлые и расположены в толще симпласта, а не у его поверхности. В волокнах с ядерной сумкой ядра симпласта образуют скопления в его утолщенной средней части. В волокнах с ядерной цепочкой в средней части симпласта утолщение не образуется, ядра лежат здесь продольно одно за другим. Рядом со скоплениями ядер расположены органеллы общего значения.
Миофибриллы находятся в концах симпластов. Сарколемма волокна соединяется с капсулой нервно-мышечного веретена, состоящей из плотной волокнистой соединительной ткани. Каждое мышечное волокно веретена спирально обвито терминалью чувствительного нервного волокна. В результате сокращения или расслабления рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена, соответственно изменяется тонус интрафузальных мышечных волокон. Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Именно поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом.
9.2.3. Сердечная мышечная ткань
Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани (textus muscularis striatus cardiacus) - симметричные участки висцерального листка спланхнотома в шейной части зародыша - миоэпикардиальные пластинки. Из них дифференцируются также клетки мезо-телия эпикарда. Исходные клетки сердечной мышечной ткани - кардиомио-бласты - характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндо-плазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролифератив-ным потенциалом.
После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез (рис. 9.9). В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постна-тальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов.
Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофи-бриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру. Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных мио-цитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином, одним-двумя ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронно-плотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза.
В целом в ходе гистогенеза возникает пять видов кардиомиоцитов - рабочие (сократительные), синусные (пейсмекерные), переходные, проводящие, а также секреторные. Рабочие (сократительные) кардиомиоциты образуют свои цепочки (рис. 9.10). Именно они, укорачиваясь, обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны
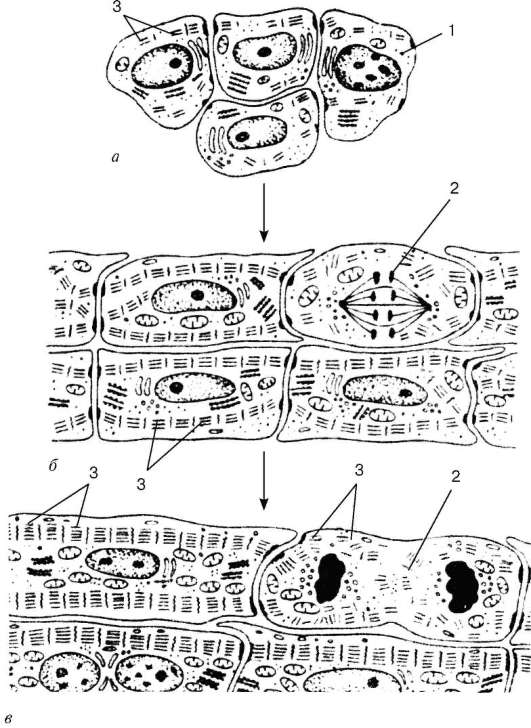
Рис. 9.9. Гистогенез сердечной мышечной ткани (схема по П. П. Румянцеву): а - кардиомиоциты в стенке сердечной трубки; б - кардиомиоциты в позднем эмбриогенезе; в - кардиомиоциты в постанатальном периоде. 1 - кардиомиоцит; 2 - митотически делящийся кардиомиоцит; 3 - миофиламенты и миофибриллы
передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Клетки воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние - проводящим и рабочим кардиомиоцитам. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами, и располагаются под эндо-
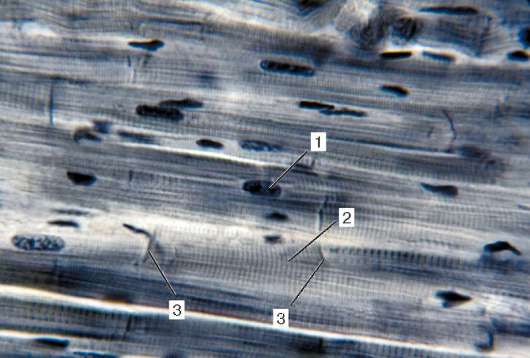
Рис. 9.10. Строение сердечной мышечной ткани (микрофотография). Окраска - железный гематоксилин:
1 - ядро кардиомиоцита; 2 - цепочка кардиомиоцитов; 3 - вставочные диски
кардом. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее - другим проводящим кар-диомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают пептидный гормон кардиодилатин, который циркулирует в крови в виде кардионатрина, вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия. Все кардиомиоциты покрыты базальной мембраной.
Строение сократительных (рабочих) кардиомиоцитов. Клетки имеют удлиненную (100-150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски (рис. 9.10). Кардиомиоциты могут ветвиться и образуют пространственную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения, за исключением агранулярной эндоплазматической сети и митохондрий.
Специальные органеллы, которые обеспечивают сокращение, называются миофибриллами. Они слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. Каждая митохондрия располагается на протяжении всего саркомера. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены,
контактируют с мембранами гладкой эндоплазматической (саркоплазмати-ческой) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные расширения (L-системы), формирующие вместе с Т-трубочками триады или диады (рис. 9.11, а). В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта.
Организация кардиомиоцитов в ткань. Кардиомиоциты соединяются друг с другом по типу «конец в конец». Здесь образуются вставочные диски: эти участки выглядят как тонкие пластинки при среднем увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмо-сомами (рис. 9.11, б).
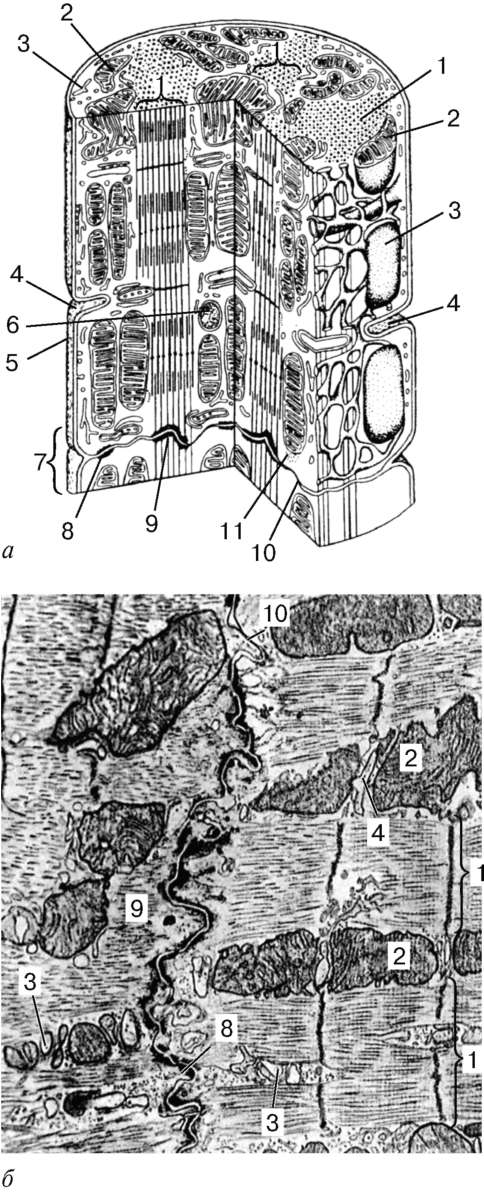
Рис. 9.11. Строение кардиомиоцита: а - схема (по Ю. И. Афанасьеву и В. Л. Горячкиной); б - электронная микрофотография вставочного диска. Увеличение 20 000. 1 - миофибриллы; 2 - митохондрии; 3 - саркотубулярная сеть; 4 - Т-трубочки; 5 - базальная мембрана; 6 - лизосома; 7 - вставочный диск; 8 - десмосома; 9 - зона прикрепления миофибрилл; 10 - щелевые контакты; 11 - гликоген
К каждой десмосоме со стороны цитоплазмы подходит миофибрил-ла, закрепляющаяся концом в десмо-плакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Регенерация. В гистогенезе сердечной мышечной ткани камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоиди-
зации. Например, у обезьян ядра до 50 % терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности. В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и др.) важную роль в компенсации повреждений кардиомиоцитов играют внутриклеточная регенерация, полиплоидизация ядер, возникновение многоядерных кардиомиоцитов.
9.3. ГЛАДКИЕ МЫШЕЧНЫЕ ТКАНИ
Различают три группы гладких (неисчерченных) мышечных тканей (textus muscularis nonstriatus) и клеток: мезенхимные, нейральные и миоэпи-телиальные клетки.
9.3.1. Мышечная ткань мезенхимного происхождения
Гистогенез. Эта ткань делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоци-тов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их в цитоплазме становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние. Дифференцируясь, они синтезируют компоненты межклеточного матрикса, коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Строение и функционирование клеток. Гладкий миоцит - веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается (рис. 9.12-9.14).
Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерии обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм. Наибольшей длины гладкие мио-
циты достигают в стенке матки - до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактильные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность.
Секреторные миоциты по своей ультраструктуре напоминают фиб-робласты, однако содержат в своей цитоплазме пучки тонких миофи-ламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцирован-ным. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца. Мономеры миозина располагаются рядом с филаментами актина. Плазмолемма образует впя-чивания - кавеолы, в которых концентрируются ионы кальция. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их термина-лей, изменяет проницаемость плаз-молеммы. Происходит высвобождение ионов кальция, что влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином.
Происходит втягивание актино-вых миофиламентов между миози-
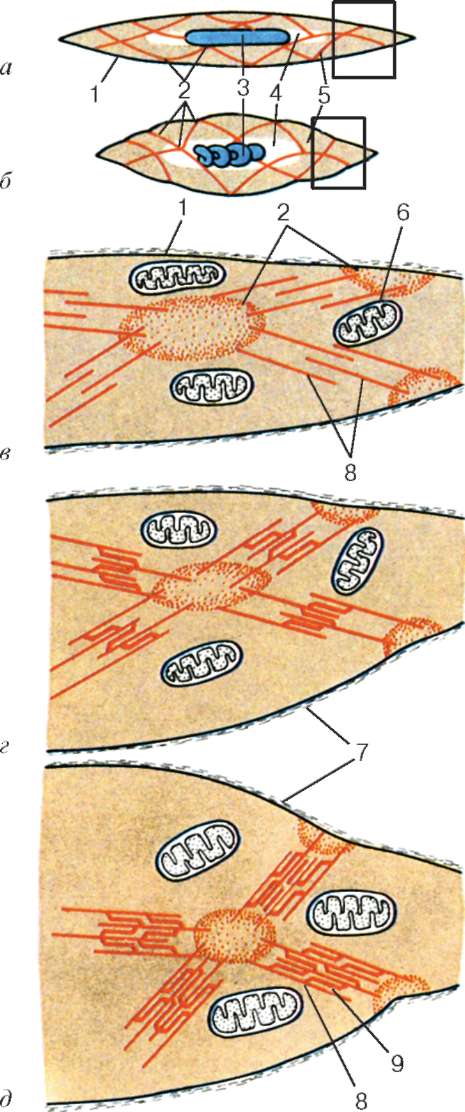
Рис. 9.12. Строение гладкого миоцита (схема):
а, в - при расслаблении; б, д - при наибольшем сокращении; г - при неполном сокращении; в-д - увеличенные изображения участков, обведенных рамками на фрагментах а и б. 1 - плазмолемма; 2 - плотные тельца; 3 - ядро; 4 - эндоплазма; 5 - сократительные комплексы; 6 - митохондрии; 7 - базальная мембрана; 8 - актиновые (тонкие) мио-филаменты; 9 - миозиновые (толстые) миофиламенты
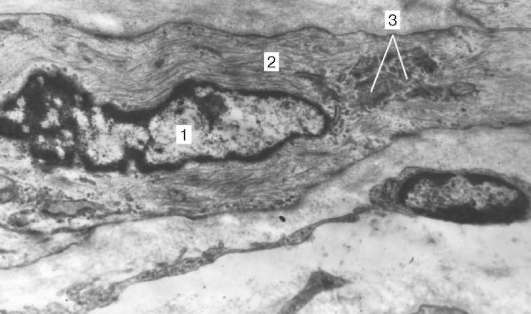
Рис. 9.13. Ультраструктура дифференцирующегося гладкого миоцита в стенке бронха:
1 - ядро; 2 - цитоплазма с миофиламентами; 3 - комплекс Гольджи, увеличение 35 000 (препарат А. Л. Зашихина)
новыми, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается (см. рис. 9.12). Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция перемещаются из цитоплазмы в кавеолы и в канальцы эндоплазматической сети, миозин деполимеризуется и «миофибриллы» распадаются. Сокращение прекращается. Таким образом, актиномиозиновые комплексы существуют в гладких миоцитах только в период сокращения при наличии в цитоплазме свободных ионов кальция.
Миоциты окружены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и аморфный компонент межклеточного матрикса. Взаимодействие миоцитов осуществляется при помощи цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом, участков мембранных контактов поверхностей миоцитов.
Регенерация. Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов. В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазматического отношения, увеличение количества свободных рибосом и полисом, актива-

Рис. 9.14. Строение гладкой мышечной ткани (объемная схема) (по Р. В. Крстичу, с изменениями):
1 - веретеновидные гладкие миоциты; 2 - цитоплазма миоцита; 3 - ядра миоци-тов; 4 - плазмолемма; 5 - базальная мембрана; 6 - поверхностные пиноцитозные пузырьки; 7 - межклеточные соединения; 8 - нервное окончание; 9 - коллагеновые фибриллы; 10 - микрофиламенты
ция ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема). При действии ряда повреждающих факторов отмечается феноти-пическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании интимальной гиперплазии при развитии атеросклероза.
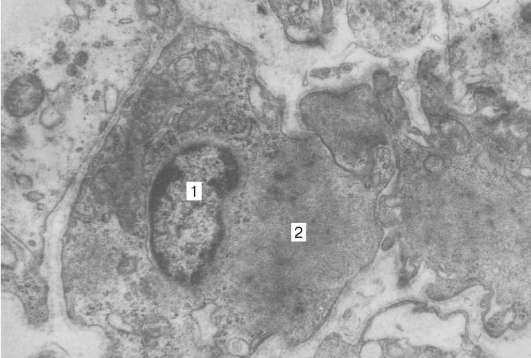
Рис. 9.15. Ультраструктура миопигментоцита (препарат Н. Н. Сарбаевой): 1 - ядро; 2 - миофиламенты, увеличение 6000
9.3.2. Мышечная ткань мезенхимного типа в составе органов
Миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффуз-но, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимно-го происхождения представлена главным образом в стенках кровеносных сосудов и многих полых внутренних органов.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы конкретных биологически активных веществ. Поэтому и на многие лекарственные препараты их реакция неодинакова. Возможно, разные функциональные свойства тканей связаны и с конкретной молекулярной организацией актиновых филаментов.
9.3.3. Мышечная ткань нейрального происхождения
Мышечная ткань радужки и цилиарного тела относится к четвертому типу сократимых тканей. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. В ряду
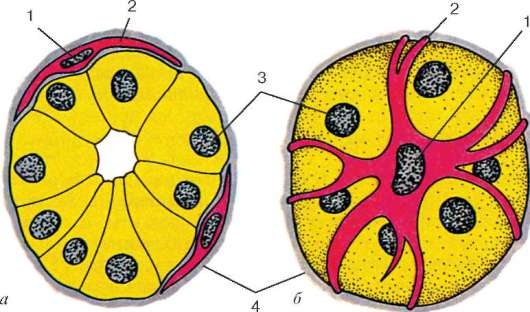
Рис. 9.16. Миоэпителиальные клетки в концевом отделе слюнной железы (схема по Г. С. Катинасу):
а - поперечный срез; б - вид с поверхности. 1 - ядра миоэпителиоцитов; 2 - отростки миоэпителиоцитов; 3 - ядра секреторных эпителиоцитов; 4 - базальная мембрана
позвоночных мышечные элементы радужки обнаруживают разнообразную дивергентную дифференцировку. Так, мионейральная ткань у рептилий и птиц представлена исчерченными многоядерными волокнами, имеющими большое сходство с мускулатурой скелетного типа. У млекопитающих и человека основной структурно-функциональной единицей мышц радужки является гладкий одноядерный миоцит, или миопигментоцит. Последние имеют пигментированное тело, содержащее одно ядро, вынесенное за пределы веретеновидной сократимой части (рис. 9.15).
Цитоплазма клеток содержит большое число митохондрий и пигментные гранулы, которые сходны по размерам и форме с гранулами пигментного эпителия. Миофиламенты в миопигментоцитах делятся на тонкие (7 нм) и толстые (1,5 нм), по размерам и расположению напоминают миофиламен-ты гладких миоцитов. Каждый миопигментоцит окружен базальной мембраной. Возле цитоплазматических отростков миоцитов обнаруживаются безмиелиновые нервные волокна. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы - суживающую и расширяющую зрачок.
Регенерация. В немногочисленных работах показана низкая регенераци-онная активность после повреждения или ее отсутствие.
9.3.4. Мышечные клетки эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные
клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются тоже из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез (рис. 9.16). В теле клетки располагаются ядро и органеллы общего значения, а в отростках - сократительный аппарат, организованный как и в клетках мышечной ткани мезенхимного типа.
Контрольные вопросы
1. Генетическая классификация мышечных тканей. Структурно-функциональные единицы разных типов мышечной ткани.
2. Поперечнополосатая скелетная мышечная ткань: развитие, строение, морфологические основы мышечного сокращения. Регенерация.
3. Поперечнополосатая сердечная мышечная ткань: развитие, специфика строения различных видов кардиомиоцитов, регенерация.
4. Разновидности гладких миоцитов: источники развития, топография в организме, регенерация.