Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 16. ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Пищеварительная система человека состоит из пищеварительной трубки и расположенных вне ее желез (слюнные железы, печень и поджелудочная железа), секрет которых участвует в процессе пищеварения. Пищеварением называются процессы химической и механической обработки пищи с последующим всасыванием продуктов ее расщепления, происходящие последовательно в различных отделах пищеварительной трубки.
В пищеварительной системе условно выделяют три основных отдела: передний, средний и задний.
Передний отдел включает органы полости рта, глотку и пищевод. В переднем отделе происходит главным образом механическая обработка пищи. Средний отдел состоит из желудка, тонкой и толстой кишки, печени и поджелудочной железы. В этом отделе осуществляются преимущественно химическая обработка пищи, всасывание продуктов ее расщепления и формирование каловых масс. Задний отдел представлен каудальной частью прямой кишки и обеспечивает функцию эвакуации непереваренных остатков пищи из пищеварительного канала.
Для понимания гистологического строения и гистофизиологии различных отделов пищеварительной системы вначале необходимо ознакомиться с особенностями гистогенеза и общими принципами строения пищеварительной трубки.
Гистогенез. Эпителиальная выстилка пищеварительной трубки и железы развиваются из эктодермы и энтодермы. Из энтодермы формируются однослойный столбчатый эпителий слизистой оболочки желудка, тонкой и большей части толстой кишки, а также железистая паренхима печени и поджелудочной железы. Из эктодермы ротовой и анальной бухт эмбриона образуется многослойный плоский эпителий полости рта, слюнных желез и каудального отдела прямой кишки. Мезенхима является источником развития соединительной ткани и сосудов, а также гладкой мускулатуры пищеварительных органов. Из висцерального листка спланхнотома развивается однослойный плоский эпителий (мезотелий) серозной оболочки - висцерального листка брюшины.
16.1. ОБЩИЙ ПЛАН МИКРОСКОПИЧЕСКОГО СТРОЕНИЯ ПИЩЕВАРИТЕЛЬНОЙ ТРУБКИ
Пищеварительная трубка в любом ее отделе состоит из внутренней слизистой оболочки (tunica mucosa), подслизистой основы (tela submucosa), мышечной оболочки (tunica muscularis) и наружной оболочки, которая представлена либо серозной оболочкой (tunica serosa), либо адвентициальной оболочкой (tunica adventitia) (рис. 16.1).
Слизистая оболочка. Свое название она получила в связи с тем, что ее поверхность постоянно увлажняется выделяемой железами слизью. Эта оболочка состоит, как правило, из трех пластинок: эпителия, собственной пластинки слизистой оболочки (lamina propria mucosae) и мышечной пластинки слизистой оболочки (lamina muscularis mucosae).
Эпителий в переднем и заднем отделах пищеварительной трубки - многослойный плоский, а в среднем отделе - однослойный столбчатый. Железы
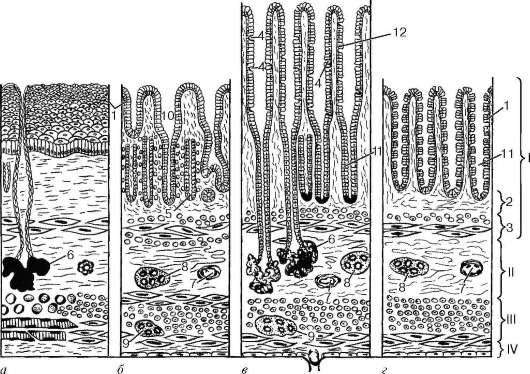
Рис. 16.1. Общий план строения пищеварительной трубки:
а - пищевод; б - желудок; в - тонкая кишка; г - толстая кишка: I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная (адвентициальная) оболочка. 1 - эпителий слизистой оболочки; 2 - собственная пластинка слизистой оболочки; 3 - мышечная пластинка слизистой оболочки; 4 - бокаловидный экзокриноцит; 5 - железы в собственной пластинке слизистой оболочки; 6 - железы, расположенные в подслизистой основе; 7 - сосудистые сплетения; 8 - подслизистое нервное сплетение; 9 - межмышечное нервное сплетение; 10 - желудочные ямочки; 11 - крипты; 12 - ворсинки
расположены либо эндоэпителиально (например, бокаловидные клетки в кишечнике), либо экзоэпителиально в собственной пластинке слизистой оболочки (пищевод, желудок) и в подслизистой основе (пищевод, двенадцатиперстная кишка) или за пределами пищеварительного канала (печень, поджелудочная железа).
Собственная пластинка слизистой оболочки лежит под эпителием, отделена от него базальной мембраной и представлена рыхлой волокнистой соединительной тканью. Здесь находятся кровеносные и лимфатические сосуды, нервные элементы, скопления лимфоидной ткани. В некоторых отделах (пищевод, желудок) могут располагаться простые железы.
Мышечная пластинка слизистой оболочки расположена на границе с под-слизистой основой и состоит из одного-трех слоев, образованных гладкими мышечными клетками. В некоторых отделах (язык, кроме корня языка, где располагаются сосочки, окруженные валом, десны) гладкие мышечные клетки отсутствуют.
Рельеф слизистой оболочки на протяжении всего пищеварительного канала неоднороден. Поверхность ее может быть гладкой (губы, щеки), образовывать углубления (ямочки в желудке, крипты в кишечнике), складки (во всех отделах), ворсинки (в тонкой кишке).
Подслизистая основа. Состоит из рыхлой волокнистой соединительной ткани. Наличие подслизистой основы обеспечивает подвижность слизистой оболочки, образование складок. В подслизистой основе имеются сплетения кровеносных и лимфатических сосудов, скопления лимфоидной ткани и подслизистое нервное сплетение (plexus nervorum submucosus). В некоторых отделах (пищевод, двенадцатиперстная кишка) расположены железы.
Мышечная оболочка. Состоит, как правило, из двух слоев мышечных элементов - внутреннего циркулярного и наружного продольного. В переднем и заднем отделах пищеварительного канала мышечная ткань преимущественно поперечнополосатая, а в среднем - гладкая. Мышечные слои разделены соединительной тканью, в которой находятся кровеносные и лимфатические сосуды и межмышечное нервное сплетение (plexus nervorum intermuscularis). Сокращения мышечной оболочки способствуют перемешиванию и продвижению пищи в процессе пищеварения.
Серозная оболочка. Большая часть пищеварительной трубки покрыта серозной оболочкой - висцеральным листком брюшины. Брюшина состоит из соединительнотканной основы, в которой расположены сосуды и нервные элементы, и из мезотелия. В некоторых отделах (пищевод, часть прямой кишки) серозная оболочка отсутствует. В таких местах пищеварительная трубка покрыта снаружи адвентициальной оболочкой, состоящей только из соединительной ткани.
Васкуляризация. Стенка пищеварительной трубки на всем протяжении обильно снабжена кровеносными и лимфатическими сосудами (рис. 16.2, а, б). Артерии образуют наиболее мощные сплетения в подслизистой основе, которые тесно связаны с артериальными сплетениями, лежащими в собственной пластинке слизистой оболочки. В тонкой кишке артериальные сплетения формируются также в мышечной оболочке. Сети кровеносных
капилляров располагаются под эпителием слизистой оболочки, вокруг желез, крипт, желудочных ямочек, внутри ворсинок, сосочков языка и в мышечных слоях. Вены также формируют сплетения подслизистой основы и слизистой оболочки.
Наличие артериоловенулярных анастомозов обеспечивает регуляцию притока крови в различные участки пищеварительного тракта в зависимости от фазы пищеварения.
Лимфатические капилляры образуют сети под эпителием, вокруг желез и в мышечной оболочке (см. рис. 16.2, а, б). Лимфатические сосуды формируют сплетения подслизи-стой основы и мышечной оболочки, а иногда и наружной оболочки (пищевод). Наиболее крупные сплетения сосудов располагаются в под-слизистой основе.
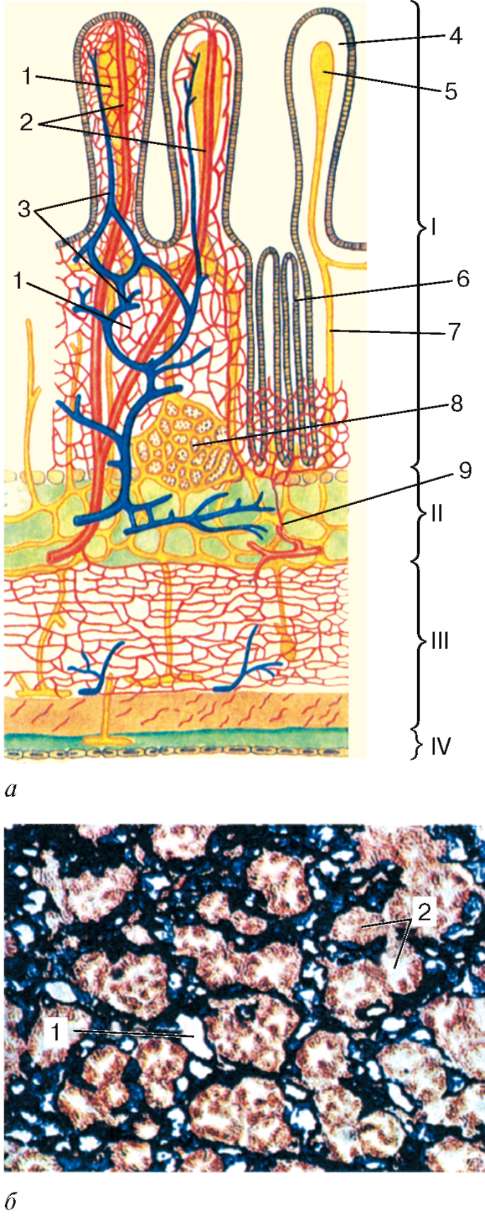
Рис. 16.2. Васкуляризация стенки пищеварительной трубки (по Н. А. Юриной, Л. С. Румянцевой):
а - расположение кровеносных и лимфатических сосудов в стенке пищеварительной трубки (на примере тонкой кишки): I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная оболочка; 1 - кровеносные капилляры; 2 - артерии; 3 - вены; 4 - кишечная ворсинка; 5 - лимфатический капилляр; 6 - кишечная железа (крипта); 7 - лимфатический сосуд; 8 - лимфоидный узелок; 9 - сосудистые сплетения в подслизистой основе; б - лимфатические капилляры слизистой оболочки желудка человека (препарат В. С. Ревазова): 1 - лимфатические капилляры; 2 - железы желудка, инъекция синей массы Герата
Иннервация. Эфферентную иннервацию обеспечивают ганглии автономной нервной системы, расположенные либо вне пищеварительной трубки (экстрамуральные симпатические ганглии), либо в толще ее (интрамуральные парасимпатические ганглии) (рис. 16.3). К экстрамураль-ным ганглиям относятся верхний шейный, звездчатый и другие узлы симпатической цепочки, иннервиру-ющие пищевод, ганглии солнечного (чревного) и тазового сплетений, иннервирующие желудок и кишечник. Интрамуральными являются ганглии
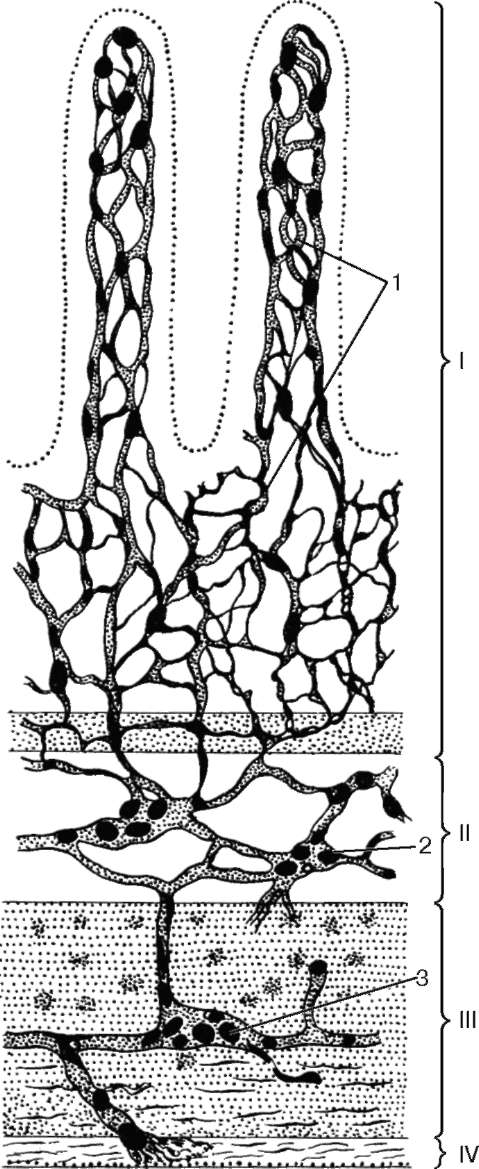
Рис. 16.3. Расположение нервных элементов в стенке тонкой кишки (по Рамон-и-Кахалю):
I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная оболочка; 1 - разветвление нервных волокон в слизистой оболочке; 2 - подслизистое нервное сплетение; 3 - межмышечное кишечное нервное сплетение
межмышечного (ауэрбахова), подсли-зистого (мейсснерова) и подсерозно-го или адвентициального, сплетений. Нейронный состав сплетений описан в главе 11. Аксоны эфферентных нейронов симпатических и парасимпатических сплетений иннервируют мышцы и железы.
Афферентная иннервация осуществляется окончаниями дендритов чувствительных нервных клеток, находящихся в составе интрамуральных ганглиев, и окончаниями дендритов чувствительных клеток чувствительных (спинномозговых) ганглиев. Чувствительные нервные окончания располагаются в мышцах, эпителии, волокнистой соединительной ткани. Афферентные окончания в стенке пищеварительного канала могут быть поливалентными, т. е. одновременно иннервировать различные ткани - эпителиальную, мышечную, соединительную, а также кровеносные сосуды.
В эпителии слизистой оболочки и железах всех отделов пищеварительной системы, но особенно в ее среднем отделе, имеются одиночные эндокринные клетки. Выделяемые ими биологически активные вещества - нейро-трансмиттеры и гормоны оказывают как местное действие, регулируя функцию желез и гладких мышц сосудов, так и общее влияние на организм.
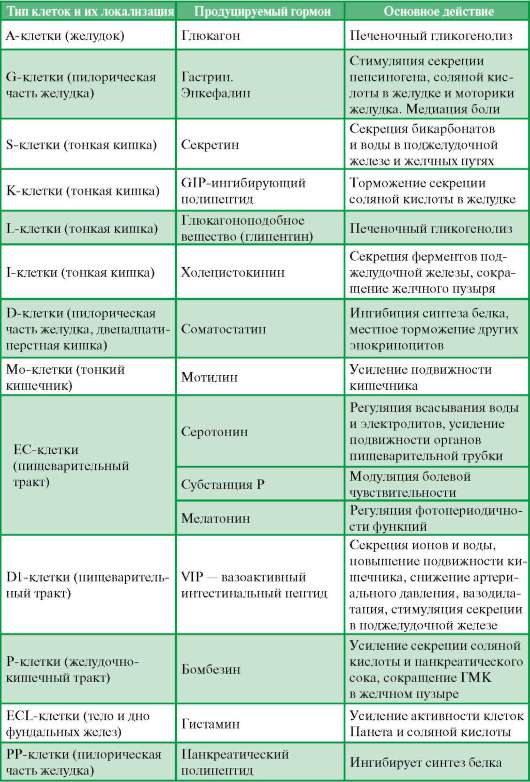
Классификация, источники развития и особенности строения клеток дисперсной эндокринной системы описаны в главе 15. В органах пищеварения их совокупность иногда называют гастроэнтеропанкреатической системой (ГЭП-системой). Главных клеток в этой системе пищеварительного тракта насчитывается более 10 типов. Данные о их локализации, структуре, продуцируемых ими гормонах и их основном действии представлены в табл. 16.1.
Таблица 16.1. Главные эндокринные клетки желудочно-кишечного тракта
Ниже, при рассмотрении структуры и функции конкретных органов пищеварительной системы более подробно описаны различные виды эндокринных клеток.
16.2. ПЕРЕДНИЙ ОТДЕЛ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Передний отдел включает полость рта со всеми ее структурными образованиями, глотку и пищевод. К производным полости рта относятся губы, щеки, десны, твердое и мягкое нёбо, язык, миндалины, слюнные железы, зубы. Здесь находится также орган вкуса.
Основная механическая функция переднего отдела определяет особенности его строения. В частности, для переднего отдела характерно наличие в слизистой оболочке многослойного плоского эпителия, предохраняющего от повреждений подлежащую соединительную ткань. Кроме того, в переднем отделе пищеварительной системы может происходить начальная химическая обработка углеводов пищи амилазой и мальтазой слюны. Некоторые органы переднего отдела пищеварительной трубки участвуют в обеспечении защитной функции.
16.2.1. Полость рта
Слизистая оболочка полости рта характеризуется следующими особенностями: наличием многослойного плоского эпителия (толщина 180- 600 мкм), отсутствием или слабым развитием мышечной пластинки слизистой оболочки и отсутствием в некоторых участках подслизистой основы. В последнем случае она твердо сращена с подлежащими тканями и лежит непосредственно на мышцах (например, в языке) или на кости (в деснах и твердом нёбе). Слизистая оболочка в местах расположения лимфоидной ткани (миндалины) образует складки.
Наличие множества поверхностно лежащих мелких кровеносных сосудов, просвечивающих через эпителий, придает слизистой оболочке характерный розовый цвет. Хорошо увлажненный эпителий способен пропускать многие вещества в эти сосуды, что часто используется во врачебной практике для введения таких лекарств, как нитроглицерин, валидол и других, через слизистую оболочку рта.
Губы
В губе различают три части: кожную (pars cutanea), промежуточную (pars intermedia) и слизистую (pars mucosa). В толще губы находятся поперечнополосатые мышцы (рис. 16.4).
Кожная часть губы имеет строение кожи. Она покрыта многослойным плоским ороговевающим эпителием и снабжена сальными, потовыми желе-
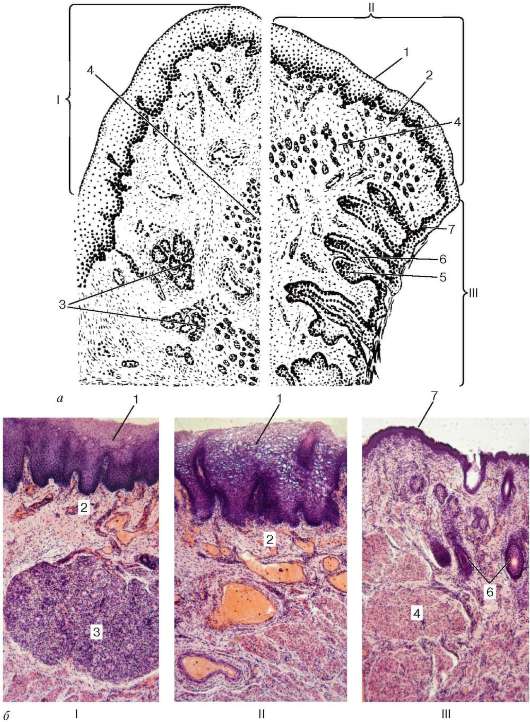
Рис. 16.4. Микроскопическое строение губы:
а - схема (по В. Г. Елисееву и др.); б - микрофотографии: I - слизистая часть; II - переходная часть; III - кожная часть. 1 - многослойный плоский эпителий; 2 - собственная пластинка слизистой оболочки с капиллярами; 3 - слюнные железы; 4 - мышцы; 5 - сальные железы; 6 - волосяной мешок; 7 - эпидермис;
зами и волосами. Эпителий этой части расположен на базальной мембране; под мембраной лежит рыхлая волокнистая соединительная ткань, образующая высокие сосочки, которые вдаются в эпителий.
Промежуточная часть губы состоит из двух зон: наружной (гладкой) и внутренней (ворсинчатой). В наружной зоне роговой слой эпителия сохраняется, но становится тоньше и прозрачнее. В этой зоне нет волос, постепенно исчезают потовые железы, а сохраняются только сальные железы, открывающие свои протоки на поверхность эпителия. Сальных желез больше в верхней губе, особенно в области угла рта. Собственная пластинка слизистой оболочки является продолжением соединительнотканной основы кожи; сосочки ее в этой зоне невысокие. Внутренняя зона у новорожденных покрыта эпителиальными сосочками, которые иногда называют ворсинками. Эти эпителиальные сосочки по мере роста организма постепенно сглаживаются и становятся малозаметными. Эпителий внутренней зоны переходной части губы взрослого человека в 3-4 раза толще, чем в наружной зоне, лишен рогового слоя. Сальные железы здесь, как правило, отсутствуют. Лежащая под эпителием рыхлая волокнистая соединительная ткань, вдаваясь в эпителий, образует очень высокие сосочки, в которых находятся многочисленные капилляры. Циркулирующая в них кровь просвечивает через эпителий и обусловливает красный цвет губы. Сосочки содержат огромное количество нервных окончаний, поэтому красный край губы очень чувствителен.
Слизистая часть губы покрыта многослойным плоским неороговевающим эпителием. Однако в клетках поверхностного слоя эпителия все же можно обнаружить небольшое количество зерен кератина. Эпителиальный пласт в слизистой части губы значительно толще, чем в кожной. Собственная пластинка слизистой оболочки здесь образует сосочки, однако они менее высокие, чем в лежащей рядом переходной части. Мышечная пластинка слизистой оболочки отсутствует, и поэтому собственная пластинка без резкой границы переходит в подслизистую основу, примыкающую непосредственно к поперечнополосатым мышцам.
В подслизистой основе располагаются секреторные отделы слюнных губных желез (gll. labiales). Железы довольно крупные, иногда достигают величины горошины. По строению это сложные альвеолярно-трубчатые железы. По характеру секрета они относятся к смешанным слизисто-белковым железам. Выводные протоки их выстланы многослойным плоским неоро-говевающим эпителием и открываются на поверхности губы.
В подслизистой основе слизистой части губы расположены артерии и обширное венозное сплетение, распространяющееся и в красную часть губы.
Щеки
Щеки - это мышечные образования, покрытые снаружи кожей, а изнутри - слизистой оболочкой. В слизистой оболочке щеки различают три зоны: верхнюю, или максиллярную (zona maxillaris), нижнюю, или мандибу-лярную (zona mandibularis), и среднюю, или промежуточную (zona intermedia). В слизистой оболочке отсутствует мышечная пластинка.
Максиллярная и мандибулярная зоны щеки имеют строение, сходное со строением слизистой части губы. Эпителий здесь многослойный плоский неороговевающий, сосочки собственной пластинки слизистой оболочки небольших размеров. В этих областях хорошо выражена подслизистая основа, в которой находится большое количество щечных желез (gll. buccales). Наиболее крупные из них лежат в области коренных зубов (gll. molares). По мере удаления от ротового отверстия железы располагаются в более глубоких слоях, т. е. в толще щечных мышц и даже снаружи от них.
Средняя, или промежуточная, зона щеки занимает область шириной около 10 мм, тянущуюся от угла рта до ветви нижней челюсти. В эмбриональном периоде и в течение первого года жизни ребенка на этом участке имеются эпителиальные ворсинки, такие же, как на переходной части губы. Сосочки собственной пластинки слизистой оболочки здесь, как и в переходной части губы, больших размеров. Слюнные железы отсутствуют. Для промежуточной зоны щеки характерно наличие нескольких редуцированных желез, таких же, как в промежуточной части губы. Промежуточная зона щеки, как и промежуточная часть губы, является зоной контакта кожи и слизистой оболочки полости рта, формирующейся вследствие срастания эмбриональных закладок при формировании ротового отверстия.
С подлежащими тканями слизистая оболочка щеки соединена при помощи подслизистой основы, которая представлена жировой клетчаткой. Здесь располагается много кровеносных сосудов и нервов. Мышечная оболочка щеки образована щечной мышцей, в толще которой лежат щечные слюнные железы с белково-слизистыми и чисто слизистыми секреторными отделами.
Десны. Твердое нёбо
Десны являются частью слизистой оболочки полости рта, которая плотно сращена с надкостницей верхней и нижней челюстей. Слизистая оболочка выстлана многослойным плоским эпителием, который иногда оро-говевает. Собственная пластинка слизистой оболочки образует длинные сосочки, глубоко вдающиеся в эпителий. Сосочки становятся ниже в той части десны, которая непосредственно прилежит к зубам. В соединительной ткани собственной пластинки слизистой оболочки местами имеются большие скопления тучных клеток. Основное вещество ее резко метахро-матично, т. е. содержит значительное количество гликозаминогликанов. Мышечная пластинка слизистой оболочки отсутствует. В области прилегания десны к зубу образуется десневая борозда. Здесь эпителий прилегает к поверхности зуба и срастается с кутикулой эмали, образуя зубо-десневое соединение. Десна богато иннервирована. В эпителии находятся свободные нервные окончания, а в собственной пластинке слизистой оболочки - инкапсулированные и неинкапсулированные нервные окончания.
Твердое нёбо состоит из костной основы, покрытой слизистой оболочкой. Подслизистая основа отсутствует, поэтому слизистая оболочка плотно сращена с надкостницей. Слизистая оболочка выстлана многослойным плоским неороговевающим эпителием. В области шва твердого нёба
эпителий слизистой оболочки иногда образует утолщения, имеющие вид характерных тяжей. У новорожденных они представлены эпителиальными тельцами, состоящими из концентрически наслоенных эпителио-цитов и достигающими величины булавочных головок. Собственная пластинка слизистой оболочки образует сосочки, вдающиеся в эпителий. В ней расположены мощные пучки коллагеновых волокон, переплетающиеся между собой и вплетающиеся в надкостницу (фиброзная зона). Эта особенность строения твердого нёба лучше выражена в местах плотных сращений слизистой оболочки с костью (например, область шва, зона перехода в десны). В остальных местах по бокам от срединного шва между собственной пластинкой слизистой оболочки и надкостницей располагаются следующие структуры: в передней части твердого нёба - жировая ткань (жировая зона); далее - скопления слизистых желез (gll. palatinae) - железистая зона. Нёбные слюнные железы альвеолярно-трубчатые, разветвленные.
Мягкое нёбо. Язычок
Мягкое нёбо и язычок состоят из сухожильно-мышечной основы, покрытой слизистой оболочкой. В мягком нёбе и язычке различают ротоглоточную (переднюю) и носоглоточную (заднюю) поверхности. У плодов и новорожденных гистологическая граница между ними проходит приблизительно по линии перегиба слизистой оболочки с ротовой поверхности на носовую на дужках мягкого нёба и на язычке. У взрослых эта граница смещается на заднюю (носовую) поверхность, так что весь язычок оказывается одетым слизистой оболочкой, характерной для полости рта.
Слизистая оболочка ротовой поверхности мягкого нёба и язычка покрыта многослойным плоским неороговевающим эпителием. Собственная пластинка слизистой оболочки образует высокие узкие сосочки, глубоко вдающиеся в эпителий. За ней располагается развитый слой эластических волокон. Мышечная пластинка слизистой оболочки отсутствует.
Далее следует хорошо подслизистая основа, образованная рыхлой соединительной тканью, с множеством адипоцитов, в которой расположены нёбные (слизистые слюнные) железы. Выводные протоки этих желез открываются на ротовой поверхности мягкого нёба и язычка.
В язычке скопления желез находятся и внутри мышечного слоя. Поперечнополосатая мышечная ткань, составляющая основу язычка, имеет ряд особенностей. Мышечные волокна ее ветвятся и образуют между собой анастомозы.
Слизистая оболочка носовой поверхности мягкого нёба покрыта многорядным реснитчатым эпителием, содержащим бокаловидные клетки. Собственная пластинка слизистой оболочки здесь лишена сосочков и отделена от эпителия хорошо выраженной базальной мембраной. На поверхности эпителия открываются мелкие слюнные железы слизистого типа. За собственной пластинкой следует слой эластических волокон. Мышечная пластинка слизистой оболочки и подслизистая основа отсутствуют. В месте перехода слизистой оболочки ротовой поверхности нёба в носовую эпите-
лий становится сначала многослойным призматическим, а затем многорядным реснитчатым.
Язык
Язык человека, помимо участия во вкусовом восприятии, механической обработке пищи и акте глотания, является органом речи. Основу языка составляет поперечнополосатая мышечная ткань соматического типа.
Язык покрыт слизистой оболочкой. Рельеф ее различен на нижней, боковых и верхней поверхностях языка. Наиболее простое строение имеет слизистая оболочка на его нижней поверхности. Эпителий здесь многослойный плоский неороговевающий. Собственная пластинка слизистой оболочки вдается в эпителий, образуя короткие сосочки. За собственной пластинкой следует подслизистая основа, которая прилежит непосредственно к мышцам. Благодаря наличию подслизистой основы слизистая оболочка нижней поверхности языка легко смещается.
Слизистая оболочка верхней и боковых поверхностей языка неподвижно сращена с его мышечным телом и снабжена особыми образованиями - сосочками. Подслизистая основа отсутствует. В языке человека имеется четыре вида сосочков: нитевидные (papillae filiformes), грибовидные (papillae fungiformes), желобоватые, или окруженные валом (papillae vallatae), и листовидные (papillae foliatae). Все сосочки языка являются производными слизистой оболочки и построены по общему плану (рис. 16.5). Поверхность сосочков образована многослойным плоским неороговевающим или частично оро-говевающим (в нитевидных сосочках) эпителием, лежащим на базальной мембране. Основу каждого сосочка составляет вырост (первичный сосочек) собственной пластинки слизистой оболочки. От вершины первичного сосочка отходит от 5 до 20 и более тонких соединительнотканных вторичных сосочков, вдающихся в эпителий. В соединительнотканной основе сосочков языка расположены многочисленные кровеносные капилляры, просвечивающиеся через эпителий (кроме нитевидных) и придающие сосочкам характерный красный цвет.
Нитевидные сосочки самые многочисленные, равномерно покрывают верхнюю поверхность языка, концентрируясь особенно в углу, образованном сосочками, окруженными валом. По размерам они наиболее мелкие среди сосочков языка. Длина их около 0,3 мм. Наряду с нитевидными сосочками встречаются конические (papillae conicae). При ряде заболеваний процесс отторжения поверхностных ороговевающих эпителиоцитов может замедляться, а эпителиальные клетки, накапливаясь в больших количествах на вершинах сосочков, образуют мощные роговые пласты. Эти массы, покрывая беловатой пленкой поверхность сосочков, создают картину языка, обложенного белым налетом.
Грибовидные сосочки немногочисленны и располагаются на спинке языка среди нитевидных сосочков. Наибольшее их количество сосредоточено на кончике языка и по его краям. Они крупнее нитевидных сосочков, их длина составляет 0,7-1,8 мм, диаметр - 0,4-1 мм. Основная масса этих сосочков имеет форму гриба с узким основанием и широкой вершиной. Среди них
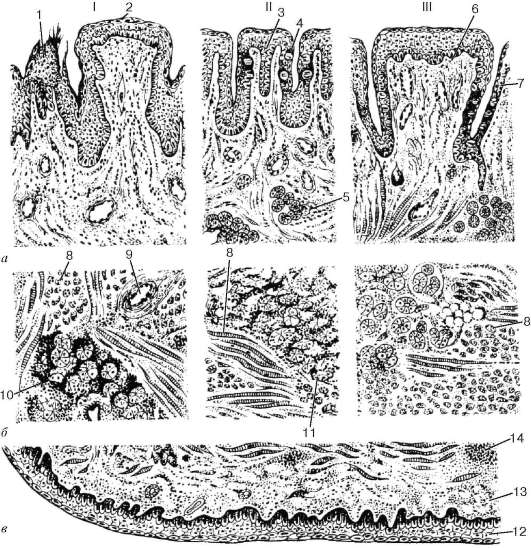
Рис. 16.5. Микроскопическое строение языка человека, продольный разрез на разных уровнях (схема по В. Г. Елисееву и др.):
а - верхняя поверхность языка - спинка языка; б - средняя часть языка; в - нижняя поверхность языка. I - кончик языка; II - боковая поверхность языка; III - корень языка. 1 - нитевидный сосочек; 2 - грибовидный сосочек; 3 - листовидный сосочек; 4 - вкусовые почки; 5 - серозные железы; 6 - желобоватый сосочек; 7 - эпителий валика желобоватого сосочка; 8 - поперечнополосатая мышца; 9 - кровеносные сосуды; 10 - смешанная слюнная железа; 11 - слизистая слюнная железа; 12 - многослойный плоский эпителий; 13 - собственная пластинка слизистой оболочки; 14 - лимфоидный узелок
встречаются конические и линзовидные формы. В толще эпителия находятся вкусовые почки (gemmae gustatoriae), располагающиеся чаще всего в области «шляпки» грибовидного сосочка. В срезах через эту зону в каждом грибовидном сосочке обнаруживается до 3-4 вкусовых почек. В некоторых сосочках вкусовые почки отсутствуют.
Желобоватые сосочки (сосочки языка, окруженные валом) встречаются на верхней поверхности корня языка в количестве от 6 до 12. Расположены они между телом и корнем языка вдоль пограничной линии. Они хорошо различимы даже невооруженным глазом. Их длина около 1-1,5 мм, диаметр 1-3 мм. В отличие от нитевидных и грибовидных сосочков, отчетливо возвышающихся над уровнем слизистой оболочки, верхняя поверхность этих сосочков лежит почти на одном уровне с нею. Они имеют узкое основание и широкую, уплощенную свободную часть. Вокруг сосочка располагается узкая, глубокая борозда - желобок (отсюда название - желобоватый сосочек). Желоб отделяет сосочек от валика - утолщения слизистой оболочки, окружающего сосочек. Наличие этой детали в строении сосочка послужило поводом к возникновению другого названия - «сосочек, окруженный валом». В толще эпителия боковых поверхностей этого сосочка и окружающего его валика расположены многочисленные вкусовые почки. В соединительной ткани сосочков и валиков часто встречаются пучки гладких мышечных клеток, расположенных продольно, косо или циркулярно. Сокращение этих пучков обеспечивает сближение сосочка с валиком. Это способствует наиболее полному соприкосновению пищевых веществ, попадающих в борозду сосочка, со вкусовыми почками, заложенными в эпителии сосочка и валика. В рыхлой волокнистой соединительной ткани основания сосочка и между прилежащими к нему пучками поперечнополосатых волокон находятся концевые отделы слюнных белковых желез, выводные протоки которых открываются в борозду сосочка. Секрет этих желез промывает и очищает борозду сосочка от пищевых частиц, слущивающегося эпителия и микробов.
Листовидные сосочки языка хорошо развиты только у детей. Они представлены двумя группами, расположенными по правому и левому краям языка. Каждая группа включает 4-8 параллельно расположенных сосочков, разделенных узкими пространствами. Длина одного сосочка около 2-5 мм. В эпителии боковых поверхностей сосочка заключены вкусовые почки. В пространства, разделяющие листовидные сосочки, открываются выводные протоки слюнных белковых желез. Их концевые отделы расположены между мышцами языка. Секрет этих желез промывает узкие пространства между сосочками. У взрослого человека листовидные сосочки редуцируются, а на местах, где ранее располагались белковые железы, развиваются жировая и лимфоидная ткани.
Слизистая оболочка корня языка характеризуется отсутствием сосочков. Поверхность имеет ряд возвышений и углублений. Возвышения образуются за счет скопления в собственной пластинке слизистой оболочки лимфоидных узелков, достигающих иногда 0,5 см в диаметре. Здесь же слизистая оболочка образует углубления - крипты, в которые открываются протоки многочисленных слюнных слизистых желез. Совокупность скоплений лимфоидной ткани в корне языка называется язычной миндалиной.
Мышцы языка образуют тело этого органа. Пучки поперечнополосатых мышц языка располагаются в трех взаимно перпендикулярных направле-
ниях: одни из них лежат вертикально, другие продольно, третьи поперечно. Мускулатура языка разделена на правую и левую половины плотной соединительнотканной перегородкой. Рыхлая соединительная ткань, лежащая между отдельными мышечными волокнами и пучками, содержит много жировых долек. Здесь же расположены концевые отделы слюнных желез языка. На границе между мышечным телом и собственной пластинкой слизистой оболочки верхней поверхности языка имеется мощная соединительнотканная пластинка, состоящая из переплетающихся наподобие решетки пучков коллагеновых и эластических волокон. Она образует так называемый сетчатый слой. Это своеобразный апоневроз языка, который особенно сильно развит в области желобоватых сосочков. На конце и у краев языка толщина его уменьшается. Поперечнополосатые мышечные волокна, проходя через отверстия сетчатого слоя, прикрепляются к маленьким сухожилиям, образованным пучками коллагеновых волокон, лежащими в собственной пластинке слизистой оболочки.
Слюнные железы языка (gll. lingualis) подразделяются на три вида: белковые, слизистые и смешанные.
Белковые слюнные железы расположены поблизости от желобоватых и листовидных сосочков в толще языка. Это простые трубчатые разветвленные железы. Их выводные протоки открываются в борозды сосочков, окруженных валом, или между листовидными сосочками и выстланы многослойным плоским эпителием. Концевые отделы состоят из клеток конической формы, выделяющих белковый секрет.
Слизистые железы расположены главным образом в корне языка и вдоль его боковых краев. Это одиночные простые альвеолярно-трубчатые разветвленные железы. Их протоки выстланы многослойным плоским эпителием. На корне языка они открываются в крипты язычной миндалины. Трубчатые концевые отделы этих желез состоят из слизистых клеток.
Смешанные железы располагаются в его переднем отделе. Их протоки (около 6 млн) открываются вдоль складок слизистой оболочки под языком. Секреторные отделы смешанных желез расположены в толще языка.
Кровоснабжение языка осуществляется входящими в него язычными артериями, которые обильно разветвляются и образуют широкую сеть в мышцах языка. Они же дают ветви к поверхностным частям языка. В сетчатом слое языка сосуды располагаются горизонтально, а затем от них отходят вертикальные конечные ветви к сосочкам слизистой оболочки. Конечные ветви формируют в соединительнотканных сосочках капиллярную сеть, от которой в каждый более мелкий сосочек входит одна петля кровеносных капилляров. Кровь от поверхностных слоев языка оттекает в венозное сплетение, располагающееся в собственной пластинке слизистой оболочки. Более крупное венозное сплетение находится в основании языка. Лимфатические сосуды также образуют сеть в собственной пластинке слизистой оболочки. Эта сеть связана с более крупной сетью, имеющейся в подслизистой основе нижней поверхности языка.
Лимфатические сосуды в большом количестве находятся и в области миндалины языка.
Иннервация. Разветвления подъязычного нерва и барабанной струны образуют многочисленные двигательные нервные окончания на поперечнополосатых мышечных волокнах. Чувствительная иннервация передних 2/3 языка осуществляется ветвями тройничного нерва, задней 1/3 - ветвями языкоглоточного нерва. В собственной пластинке слизистой оболочки языка имеется хорошо выраженное нервное сплетение, от которого отходят нервные волокна к вкусовым почкам, эпителию, железам и сосудам. Нервные волокна, входящие в эпителий, разветвляются среди эпителиальных клеток и заканчиваются свободными нервными окончаниями. Иннервация вкусовых почек описана в главе 12.
16.2.2. Лимфоэпителиальное глоточное кольцо Пирогова. Миндалины
На границе полости рта и глотки в слизистой оболочке располагаются большие скопления лимфоидной ткани. В совокупности они образуют лимфоэпителиальное глоточное кольцо, окружающее вход в дыхательные и пищеварительные пути. Наиболее крупные скопления этого кольца носят название миндалин. По месту их расположения различают нёбные миндалины, глоточную миндалину, язычную миндалину. Кроме перечисленных миндалин, в слизистой оболочке переднего отдела пищеварительной трубки существует ряд скоплений лимфоидной ткани, из которых наиболее крупными являются скопления в области слуховых труб - трубные миндалины и в желудочке гортани - гортанные миндалины.
Миндалины выполняют в организме важную защитную функцию, обезвреживая микробы, постоянно попадающие из внешней среды в организм через носовые и ротовое отверстия. Наряду с другими органами, содержащими лимфоидную ткань, они обеспечивают образование лимфоцитов, участвующих в реакциях гуморального и клеточного иммунитета (см. главу 14).
Развитие. Нёбные миндалины закладываются на 9-й нед эмбриогенеза в виде углубления многорядного реснитчатого эпителия латеральной стенки глотки, под которым лежат компактно расположенные мезен-химные клетки и многочисленные кровеносные сосуды. На 11-12-й нед формируется тонзиллярный синус, эпителий которого перестраивается в многослойный плоский, а из мезенхимы развивается ретикулярная ткань; появляются сосуды, в том числе посткапиллярные венулы с высокими эндотелиоцитами. Происходит заселение органа лимфоцитами. На 14-й нед среди лимфоцитов определяются главным образом Т-лимфоциты (21 %) и немного В-лимфоцитов (1 %). На 17-18-й нед появляются первые лимфо-идные узелки. К 19-й нед содержание Т-лимфоцитов возрастает до 60 %, а В-лимфоцитов - до 3 %. Глоточная миндалина развивается на 4-м мес внутриутробного периода из эпителия и подлежащей мезенхимы дорсальной стенки глотки. У плода она покрыта многорядным мерцательным эпителием. Язычная миндалина закладывается на 5-м мес.
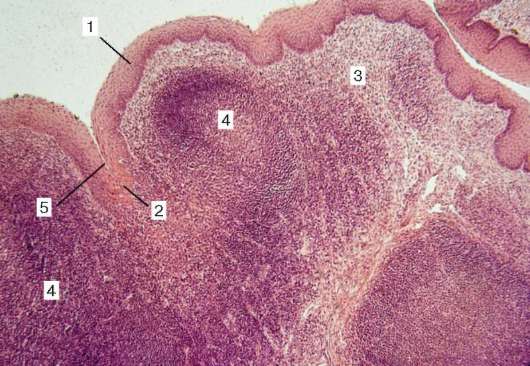
Рис. 16.6. Миндалина (микрофотография):
1 - многослойный плоский неороговевающий эпителий; 2 - многослойный плоский эпителий, инфильтрированный лимфоцитами; 3 - собственная пластинка слизистой оболочки; 4 - лимфоидные узелки; 5 - крипта
Миндалины достигают максимального развития в детском возрасте. Начало инволюции миндалин совпадает с периодом полового созревания.
Строение. Нёбные миндалины во взрослом организме представлены двумя телами овальной формы, расположенными по обеим сторонам глотки между нёбными дужками. Каждая миндалина состоит из нескольких складок слизистой оболочки, в собственной пластинке которой расположены многочисленные лимфоидные узелки (noduli lymphathici) (рис. 16.6). От поверхности миндалины в глубь органа отходят 10-20 крипт (criptae tonsillares), которые разветвляются и образуют вторичные крипты. Слизистая оболочка покрыта многослойным плоским неороговевающим эпителием. Во многих местах, особенно в криптах, эпителий часто бывает инфильтрирован (заселен) лимфоцитами и зернистыми лейкоцитами. Лейкоциты, проникающие в толщу эпителия, обычно в большем или меньшем количестве выходят на его поверхность и передвигаются навстречу бактериям, попадающим в полость рта вместе с пищей и воздухом. Микробы в миндалине активно фагоцитируются лейкоцитами, при этом часть лейкоцитов погибает. Под влиянием микробов и различных ферментов, выделяемых лейкоцитами, эпителий миндалины часто бывает разрушен. Однако через некоторое время за счет размножения камбиальных клеток эпителиального пласта эти участки восстанавливаются.
Собственная пластинка слизистой оболочки образует небольшие сосочки, вдающиеся в эпителий. В рыхлой соединительной ткани этого слоя расположены многочисленные лимфоидные узелки. В центрах некоторых узелков хорошо выражены более светлые участки - герминативные центры.
Лимфоидные узелки миндалин чаще всего отделены друг от друга тонкими прослойками соединительной ткани. Однако некоторые узелки могут сливаться. Мышечная пластинка слизистой оболочки не выражена.
Подслизистая основа, располагающаяся под скоплением лимфоидных узелков, образует вокруг миндалины капсулу, от которой в глубь миндалины отходят соединительнотканные перегородки. В этом слое сосредоточены основные кровеносные и лимфатические сосуды миндалины и ветви языкоглоточного нерва, осуществляющие ее иннервацию. Здесь же находятся и секреторные отделы небольших слюнных желез. Протоки этих желез открываются на поверхности слизистой оболочки, расположенной вокруг миндалины. Снаружи от подслизистой основы лежат поперечнополосатые мышцы глотки.
Глоточная миндалина расположена в участке дорсальной стенки глотки, лежащем между отверстиями слуховых труб. Строение ее сходно с другими миндалинами. Во взрослом организме она выстлана многослойным плоским неороговевающим эпителием. Однако в криптах глоточной миндалины и у взрослого иногда встречаются участки многорядного реснитчатого эпителия, характерного для эмбрионального периода развития.
При некоторых патологических состояниях глоточная миндалина может быть очень сильно увеличена (аденоиды).
Язычная миндалина расположена в слизистой оболочке корня языка. Эпителий, покрывающий поверхность миндалины и выстилающий крипты, многослойный плоский неороговевающий. Эпителий и подлежащая собственная пластинка слизистой оболочки инфильтрированы лимфоцитами, проникающими сюда из лимфоидных узелков. На дне многих крипт открываются выводные протоки слюнных желез языка. Их секрет способствует промыванию и очищению крипт.
16.2.3. Слюнные железы
Общая морфофункциональная характеристика. В полость рта открываются выводные протоки трех пар больших слюнных желез: околоушных, под-нижнечелюстных и подъязычных, лежащих за пределами слизистой оболочки. Кроме того, в толще слизистой оболочки полости рта находятся многочисленные мелкие слюнные железы: губные, щечные, язычные, нёбные.
Эпителиальные структуры всех слюнных желез развиваются из эктодермы, как и многослойный плоский эпителий, выстилающий полость рта. Поэтому для строения их выводных протоков и секреторных отделов характерна многослойность.
Слюнные железы представляют собой сложные альвеолярные или альвеолярно-трубчатые железы. Они состоят из концевых отделов и протоков, выводящих секрет (рис. 16.7).
Концевые отделы (portio terminalis) по строению и характеру выделяемого секрета бывают трех типов: белковые (серозные), слизистые и смешанные (т. е. белково-слизистые).
Выводные протоки слюнных желез подразделяются на внутридольковые (ductus intralobularis), включающие вставочные (ductus intercalates) и исчерченные (ductus striatus), междольковые (ductus interlobularis) выводные протоки и главный выводной проток железы (ductus excretorius principalis) (см. рис. 16.7).
Белковые железы выделяют жидкий секрет, богатый ферментами. Слизистые железы образуют более густой, вязкий секрет с большим содержанием муцина - вещества, в состав которого входят гликопротеины. По механизму отделения секрета из клеток все слюнные железы являются мерокринными.
Слюнные железы выполняют экзокринные и эндокринные функции. Экзокринная функция заключается в регулярном отделении в полость рта слюны. В ее состав входят вода (около 99 %), белковые вещества, в том числе ферменты, неорганические вещества, а также клеточные элементы (клетки эпителия, лейкоциты).
Слюна увлажняет пищу, придает ей полужидкую консистенцию, что облегчает процессы жевания и глотания. Постоянное смачивание слизистой оболочки щек и губ слюной способствует акту артикуляции. Одной из важных функций слюны является ферментативная обработка пищи. Ферменты слюны могут участвовать в расщеплении полисахаридов (амилаза, мальтаза, гиалуронидаза), нуклеиновых кислот и нуклеопротеидов (нуклеазы и кал-ликреин), белков (калликреиноподобные протеазы, пепсиноген, трипсино-подобные ферменты), клеточных оболочек (лизоцим). Помимо секреторной функции, слюнные железы выполняют экскреторную функцию. Со слюной во внешнюю среду выделяются различные органические и неорганические вещества: мочевая кислота, креатин, железо, йод и др. Защитная функция слюнных желез состоит в выделении бактерицидного вещества - лизоци-ма.
Эндокринная функция слюнных желез заключается в выработке биологически активных веществ, подобных гормонам - инсулина, паротина, фактора роста нервов (ФРН), фактора роста эпителия (ФРЭ), тимоциттранс-формирующего фактора (ТТФ), фактора летальности и др. Слюнные железы активно участвуют в регуляции водно-солевого гомеостаза.
Развитие. Закладка околоушных желез происходит на 8-й нед эмбриогенеза, когда из эпителия полости рта в подлежащую мезенхиму начинают расти эпителиальные тяжи по направлению к правому и левому ушным отверстиям. От этих тяжей отпочковываются многочисленные выросты, формирующие сначала выводные протоки, а затем концевые отделы. На 10-12-й нед наблюдаются система разветвленных эпителиальных тяжей, представленных малодифференцированными клетками, врастание нервных волокон. На 4-6-м мес развития в результате дивергентной дифференци-ровки клеток эпителиальных тяжей появляются секреторные эпителиоциты и миоэпителиальные клетки, входящие в состав концевых отделов желез, а к 8-9-му мес в них появляются просветы. Вставочные протоки и концевые отделы у плодов и детей до 2-летнего возраста представлены типичными слизистыми клетками. Из мезенхимы на 5-5,5 мес внутриутробного развития формируются соединительнотканная капсула и прослойки междоль-
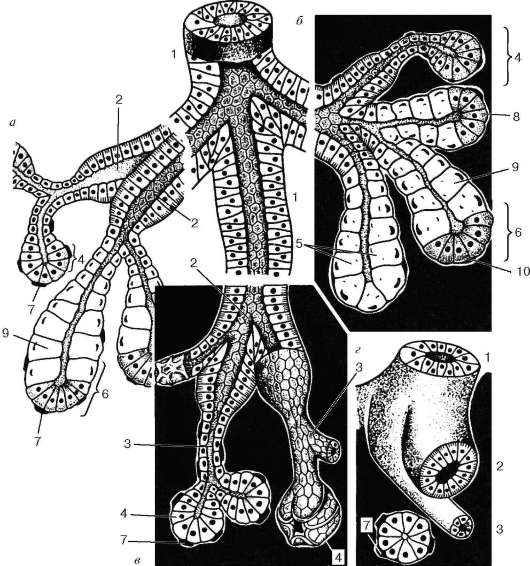
Рис. 16.7. Строение слюнных желез (по В. Г. Елисееву и др.):
а - долька поднижнечелюстной железы; б - долька подъязычной железы; в - долька околоушной железы; г - поперечное сечение различных отделов околоушной железы. 1 - выводной проток железы; 2 - исчерченный проток; 3 - вставочные протоки; 4 - белковый концевой отдел; 5 - слизистый концевой отдел; 6 - смешанный концевой отдел (белково-слизистый); 7 - миоэпителиоциты; 8 - серозное полулуние; 9 - слизистые клетки (мукоциты); 10 - белковые клетки (сероциты)
ковой соединительной ткани. Сначала секрет имеет слизистый характер. В последние месяцы развития плода в его слюне проявляют ферменты, расщепляющие полисахариды.
Поднижнечелюстные железы закладываются на 6-й нед эмбриогенеза. На 8-й нед в эпителиальных тяжах образуются просветы. Эпителий первичных выводных протоков сначала двухслойный, затем многослойный.
Концевые отделы формируются на 16-й нед. Слизистые клетки концевых отделов образуются в процессе ослизнения клеток вставочных протоков. Процесс дифференцировки концевых отделов и внутридольковых протоков на вставочные отделы и слюнные трубки продолжается в постнатальном периоде развития. У новорожденных в составе концевых отделов находятся железистые клетки кубической и призматической формы, вырабатывающие белковый секрет (полулуния Джиануцци), и миоэпителиальный клетки. Секреция в концевых отделах начинается у 4-месячных плодов. Состав секрета отличается от секрета взрослого человека.
Подъязычные железы закладываются на 8-й нед эмбриогенеза в виде выростов поднижнечелюстных желез. На 12-й нед отмечаются почкование и ветвление эпителиального зачатка.
Околоушная железа
Околоушная железа (gl. parotis) - сложная альвеолярная разветвленная железа, выделяющая белковый секрет в полость рта, а также обладающая эндокринной функцией. Снаружи она покрыта плотной соединительнотканной капсулой. Железа имеет дольчатое строение. В прослойках соединительной ткани между дольками расположены междольковые протоки и кровеносные сосуды (рис. 16.8).
Концевые отделы околоушной железы белковые (серозные) (см. рис. 16.8). Они состоят из секреторных клеток конической формы - белковых клеток, или сероцитов (serocyti), и миоэпителиальных клеток. Белковые клетки имеют узкую апикальную часть, выступающую в просвет концевого отдела. В ней содержатся ацидофильные секреторные гранулы, количество которых изменяется в зависимости от фазы секреции. Базальная часть клетки более широкая, содержит ядро.
В фазе накопления секрета размеры клеток значительно увеличиваются, а после выделения его уменьшаются, ядро округляется. В секрете околоушных желез преобладает белковый компонент, но часто содержатся и мукополисахариды, поэтому такие железы могут быть названы серо-мукозными. В секреторных гранулах выявляются альфа-амилаза, ДНК-аза. Цитохимически и электронно-микроскопически различают несколько типов гранул - ШИК-положительные с электронно-плотным ободком, ШИК-отрицательные и мелкие гомогенные сферической формы. В концевых отделах между сероцитами располагаются межклеточные секреторные канальцы, просвет которых имеет диаметр около 1 мкм. В эти канальцы выделяется из клеток секрет, который далее поступает в просвет концевого секреторного отдела. Общая секреторная площадь концевых отделов обеих желез достигает почти 1,5 м2.
Миоэпителиальные клетки (миоэпителиоциты) составляют второй слой клеток в концевых секреторных отделах. По происхождению это эпителиальные клетки, по функции - сократительные элементы, напоминающие мышечные. Их называют также звездчатыми миоэпителиоцитами, так как они имеют звездчатую форму и своими отростками охватывают концевые секреторные отделы наподобие корзинок (см. рис. 16.7). Миоэпителиальные
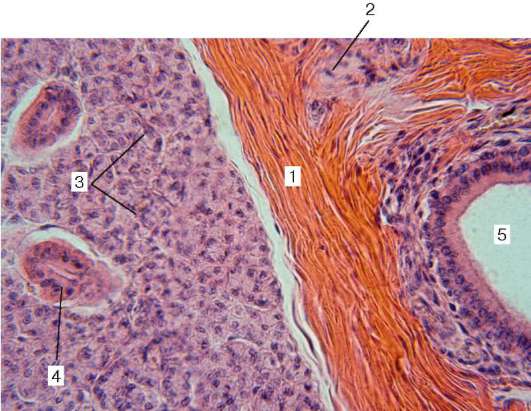
Рис. 16.8. Околоушная железа (микрофотография):
1 - междольковая рыхлая соединительная ткань; 2 - кровеносный сосуд; 3 - концевые секреторные отделы; 4 - исчерченный проток; 5 - междольковый проток
клетки всегда располагаются между базальной мембраной и основанием эпителиальных клеток. Своими сокращениями они способствуют выделению секрета из концевых отделов.
Внутридольковые вставочные протоки околоушной железы начинаются непосредственно от ее концевых отделов. Они обычно сильно разветвлены (см. рис. 16.7). Вставочные протоки выстланы кубическим или плоским эпителием, в составе которого находятся малодифференцированные камбиальные клетки. Второй слой в них образуют миоэпителиоциты.
Исчерченные слюнные протоки являются продолжением вставочных и располагаются также внутри долек. Диаметр их значительно больший, чем вставочных протоков, просвет хорошо выражен. Исчерченные протоки ветвятся и часто образуют ампулярные расширения. Они выстланы однослойным призматическим эпителием. Цитоплазма эпителиоцитов ацидофильна. В апикальной части клеток видны микроворсинки, секреторные гранулы с содержимым различной электронной плотности, комплекс Гольджи. В базальных частях эпителиальных клеток отчетливо выявляется базальная исчерченность, образованная митохондриями, расположенными в цитоплазме между складками плазмолеммы перпендикулярно к базальной мембране. Эти клетки осуществляют транспорт воды и ионов. В исчерченных протоках выявлены циклические изменения, не связанные с ритмом пищеварительного процесса.
Междольковые выводные протоки выстланы двухслойным эпителием. По мере укрупнения протоков эпителий их постепенно становится много-
слойным. Выводные протоки окружены прослойками рыхлой волокнистой соединительной ткани.
Главный проток околоушной железы, начинающийся в ее теле, проходит через жевательную мышцу, а его устье расположено на поверхности слизистой оболочки щеки на уровне второго верхнего большого коренного зуба. Проток выстлан многослойным кубическим, а в устье - многослойным плоским эпителием.
Поднижнечелюстная железа
Поднижнечелюстная железа (gl. submaxillare) - сложная альвеолярная (местами альвеолярно-трубчатая) разветвленная железа. По характеру отделяемого секрета она смешанная, т. е. белково-слизистая. С поверхности железа окружена соединительнотканной капсулой.
В составе поднижнечелюстной железы находятся концевые секреторные отделы (ацинусы) двух типов: белковые и белково-слизистые, т. е. смешанные (рис. 16.9), но преобладают в ней белковые концевые отделы. Секреторные гранулы белковых клеток (сероцитов) обладают невысокой электронной плотностью. Часто внутри гранул содержится электронно-плотная сердцевина. Ацинус состоит из 10-18 секреторных клеток, из которых лишь 4- 6 расположены вокруг его просвета. Секреторные гранулы содержат глико-липиды и гликопротеиды.
Смешанные концевые отделы более крупные, чем белковые, и состоят из двух видов клеток - слизистых и белковых. Слизистые клетки (mucocyti) более крупные по сравнению с белковыми и занимают центральную часть концевого отдела. Ядра слизистых клеток располагаются всегда у их основания, они сильно уплощены и уплотнены. Цитоплазма этих клеток имеет ячеистую структуру благодаря наличию в ней слизистого секрета, который избирательно окрашивается муцикармином и электронно-прозрачен. Небольшое количество белковых клеток охватывает слизистые клетки в виде серозного полулуния (semilunium serosum). Серозные полулуния (Джиануцци) являются характерными структурами смешанных желез. Между железистыми клетками расположены межклеточные секреторные канальцы. Снаружи от клеток полулуния лежат миоэпителиальные клетки.
Вставочные протоки поднижнечелюстной железы менее разветвленные и более короткие, чем в околоушной железе, что объясняется ослизнением части этих отделов в процессе развития. Клетки этих отделов содержат мелкие секреторные гранулы, часто с плотными сердцевинами.
Исчерченные протоки в поднижнечелюстной железе очень хорошо развиты, длинные и сильно ветвятся (см. рис. 16.7). В них часто встречаются сужения и расширения. Выстилающий их призматический эпителий с хорошо выраженной базальной исчерченностью содержит желтый пигмент.
Среди клеток исчерченных протоков при электронной микроскопии различают несколько типов - широкие темные, высокие светлые, мелкие треугольной формы (малодифференированные) и клетки, имеющие форму бокала. В базальной части высоких клеток на боковых поверхностях расположены многочисленные цитоплазматические выросты. У некоторых
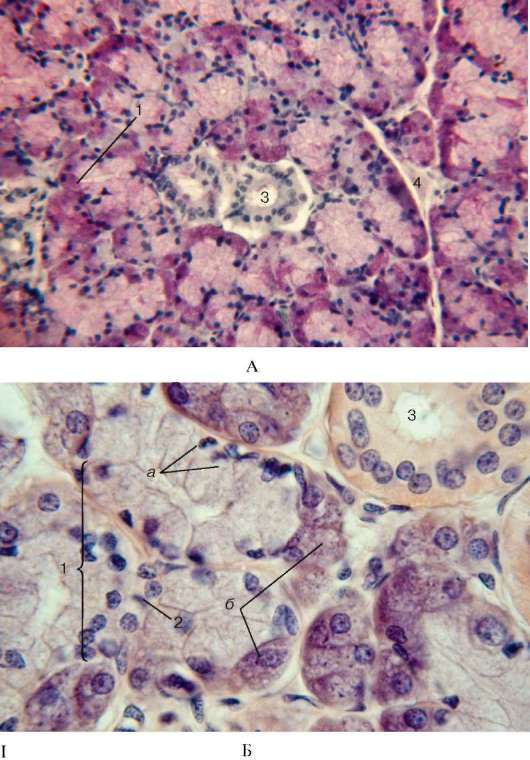
Рис. 16.9. Строение поднижнечелюстной железы:
I - микрофотографии: малое (А) и большое (Б) увеличение. 1 - смешанный концевой отдел: а - мукоциты; б - сероциты, образующие серозное полулуние; 2 - миоэпителиальная клетка; 3 - исчерченный проток; 4 - междольковая рыхлая соединительная ткань
животных (грызуны), кроме исчерченных протоков, имеются гранулярные отделы, в клетках которых нередко обнаруживаются хорошо развитый комплекс Гольджи и гранулы, содержащие трипсиноподобные протеазы, а также ряд гормональных и ростстимулирующих факторов. Показано, что эндокринные функции слюнных желез (выделение инсулиноподобного и других веществ) связаны с этими отделами.
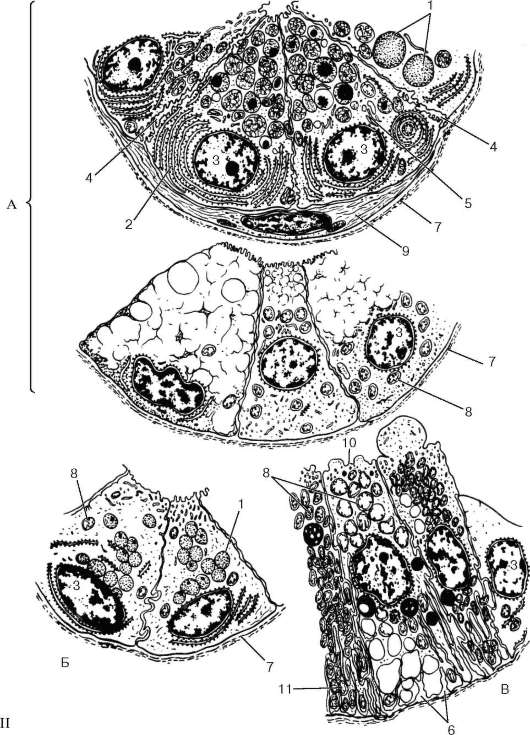
Рис. 16.9. Продолжение
II - схема ультрамикроскопического строения клеток ацинуса (А), вставочного протока (Б), исчерченного протока (В) (по Е. А. Шубниковой): 1 - секреторные гранулы; 2 - гранулярная эндоплазматическая сеть; 3 - ядро; 4 - межклеточный каналец; 5 - комплекс Гольджи; 6 - складка базальной плазмолеммы; 7 - базальная мембрана; 8 - митохондрии; 9 - миоэпителиальная клетка; 10 - светлая клетка; 11 - темная клетка
Междольковые выводные протоки поднижнечелюстной железы, располагающиеся в соединительнотканных перегородках, выстланы сначала двухслойным, а затем многослойным эпителием. Главный проток поднижнече-люстной железы открывается рядом с протоком подъязычной железы на переднем крае уздечки языка. Его устье выстлано многослойным плоским эпителием. Проток поднижнечелюстной железы более разветвлен, чем проток околоушной железы.
Подъязычная железа
Подъязычная железа (gl. sublinguale) - сложная альвеолярно-трубчатая разветвленная железа. По характеру отделяемого секрета - смешанная, слизисто-белковая, с преобладанием слизистой секреции. В ней имеются концевые секреторные отделы трех типов: белковые, смешанные и слизистые.
Белковые концевые отделы очень немногочисленны (см. рис. 16.7). Смешанные концевые отделы составляют основную массу железы. Полулуния, образованные секреторными клетками, в них выражены лучше, чем в под-нижнечелюстной железе. Клетки, формирующие полулуния, в подъязычной железе значительно отличаются от таковых в околоушной и подниж-нечелюстной железах. Их секреторные гранулы дают реакцию на муцин. Эти клетки выделяют одновременно белковый и слизистый секрет и потому носят название серомукозных клеток (серомукоцитов). Последние образуют серомукозное полулуние. В серомукоцитах сильно развита гранулярная эндоплазматическая сеть. Они снабжены межклеточными секреторными канальцами. Чисто слизистые концевые отделы этой железы состоят из характерных слизистых клеток, содержащих хондроитинсульфат В и гликопротеиды. Миоэпителиоциты образуют наружный слой концевых отделов.
В подъязычной железе общая площадь вставочных протоков очень мала, так как они еще в процессе эмбрионального развития почти целиком ослиз-няются, формируя слизистые части концевых отделов. Исчерченные протоки в этой железе развиты слабо: они очень короткие, а в некоторых местах отсутствуют. Эти протоки выстланы призматическим или кубическим эпителием, в котором также видна базальная исчерченность, как и в соответствующих протоках других слюнных желез.
В цитоплазме эпителиальных клеток, выстилающих исчерченные протоки, содержатся мелкие пузырьки, которые рассматриваются как показатель экскреции.
Внутридольковые и междольковые выводные протоки подъязычной железы образованы двухслойным призматическим, а в устье - многослойным плоским эпителием. Соединительнотканные внутридольковые и междоль-ковые перегородки в этих железах развиты лучше, чем в околоушных или поднижнечелюстных железах.
Васкуляризация. Все слюнные железы богато снабжены сосудами. Артерии, входящие в железы, сопровождают разветвления выводных протоков. От них отходят веточки, питающие стенки протоков. У концевых
отделов мелкие артерии распадаются на капиллярную сеть, густо оплетающую каждый из этих отделов. Из кровеносных капилляров кровь собирается в вены, которые следуют по ходу артерий.
Для кровеносной системы слюнных желез характерно наличие значительного количества артериоловенулярных анастомозов. Они располагаются в воротах железы, при входе сосудов в дольку и перед капиллярными сетями концевых отделов. Анастомозы в слюнных железах обусловливают возможность значительного изменения интенсивности кровоснабжения отдельных концевых отделов, долек и даже всей железы, а следовательно, изменения секреции в слюнных железах.
Иннервация. Эфферентные, или секреторные, волокна крупных слюнных желез происходят из двух источников: отделов парасимпатической и симпатической нервной системы. Гистологически в железах обнаруживаются миелиновые и безмиелиновые нервы, следующие по ходу сосудов и протоков. Они образуют нервные окончания в стенках сосудов, на концевых отделах и в выводных протоках желез. Морфологические различия между секреторными и сосудистыми нервами не всегда можно определить. В экспериментах на поднижнечелюстной железе животных было показано, что вовлечение в рефлекс симпатических эфферентных путей ведет к образованию вязкой слюны, содержащей большое количество слизи. При раздражении парасимпатических эфферентных путей образуется жидкий белковый секрет. Закрытие и открытие просвета артериоловенулярных анастомозов и конечных вен также определяется нервными импульсами.
Возрастные изменения. После рождения процессы морфогенеза в околоушных слюнных железах продолжаются до 16-20 лет; при этом железистая ткань преобладает над соединительной тканью. После 40 лет происходит уменьшение объема железистой ткани, увеличение жировой ткани, разрастание соединительной ткани. В течение первых двух лет жизни в околоушных железах вырабатывается в основном слизистый секрет, с 3-го года до глубокой старости - белковый, а к 80-м годам опять преимущественно слизистый.
В поднижнечелюстных железах полное развитие серозных и слизистых секреторных отделов наблюдается у 5-месячных детей. Максимальное развитие их отмечается к 25 годам. После 50 лет начинается инволюция железы.
Рост подъязычных желез, как и других, наиболее интенсивно происходит в течение первых двух лет жизни.
Регенерация. Функционирование слюнных желез неизбежно сопровождается частичным разрушением эпителиальных железистых клеток. Отмирающие клетки характеризуются крупными размерами, пикнотиче-скими ядрами и плотной зернистой цитоплазмой, сильно окрашивающейся кислыми красителями. Такие клетки получили название набухающих. Восстановление паренхимы желез осуществляется главным образом путем внутриклеточной регенерации и редкими делениями камбиальных клеток в составе протоков.
16.2.4. Зубы
Зубы (denti) являются частью жевательного аппарата и состоят главным образом из минерализованных тканей. Они также принимают участие в произношении звуков речи человека, а у животных это еще и орган защиты и нападения. У человека они представлены двумя генерациями: вначале образуются выпадающие, или молочные (20), а затем постоянные (32) зубы. В лунках челюстных костей зубы укрепляются плотной соединительной тканью - периодонтом, который в области шейки зуба образует циркулярную зубную связку. Коллагеновые волокна зубной связки имеют преимущественно радиальное направление. С одной стороны они проникают в цемент корня зуба, а с другой - в альвеолярную кость. Периодонт выполняет не только механическую, но и трофическую функцию, так как в нем проходят кровеносные сосуды, питающие корень зуба.
Развитие. В развитии зубов различают три этапа: 1) образование и обособление зубных зачатков; 2) дифференцировка зубных зачатков; 3) развитие тканей зуба (рис. 16.10).
Первый этап при развитии молочных зубов протекает одновременно с обособлением полости рта и образованием преддверия полости рта. Он начинается в конце 2-го мес внутриутробного развития, когда в эпителии полости рта возникает щечно-губная пластинка, растущая в мезенхиму. Затем в этой пластинке появляется щель, знаменующая обособление полости рта и появление преддверия.
В области закладки однокоренных зубов от дна преддверия растет второе эпителиальное выпячивание в виде валика, превращающегося в зубную пластинку (lamina dentalis). Зубная пластинка в области закладки многокорневых зубов развивается самостоятельно непосредственно из эпителия полости рта. На внутренней поверхности зубной пластинки сначала появляются эпителиальные скопления - зубные зачатки (germen dentis), из которых развиваются эмалевые органы (organum enamelium). Вокруг зубного зачатка клетки мезенхимы уплотняются, формируется зубной мешочек (sacculus dentis). В дальнейшем навстречу к каждому зубному зачатку начинает расти мезенхима в виде зубного сосочка (papilla dentis), вдавливаясь в эмалевый орган (см. рис. 16.10), который становится похожим на двустенный бокал или колпачок.
Второй этап заключается в дифференцировке клеток эмалевого органа, в котором топографически выделяются внутренний и наружный эмалевый эпителий и клетки промежуточного слоя. Внутренний эмалевый эпителий располагается на базальной мембране и становится призматическим (рис. 16.11, а, б). Впоследствии он образует эмаль (enamelum), в связи с чем клетки этого эпителия получили название энамелобластов, или амелобластов (enameloblasti, ameloblasti). Наружный эмалевый эпителий в процессе дальнейшего роста эмалевого органа уплощается, а клетки промежуточного слоя приобретают звездчатую форму вследствие накопления между ними жидкости. Так образуется пульпа эмалевого органа, которая позднее принимает участие в образовании кутикулы эмали (cuticula enameli).
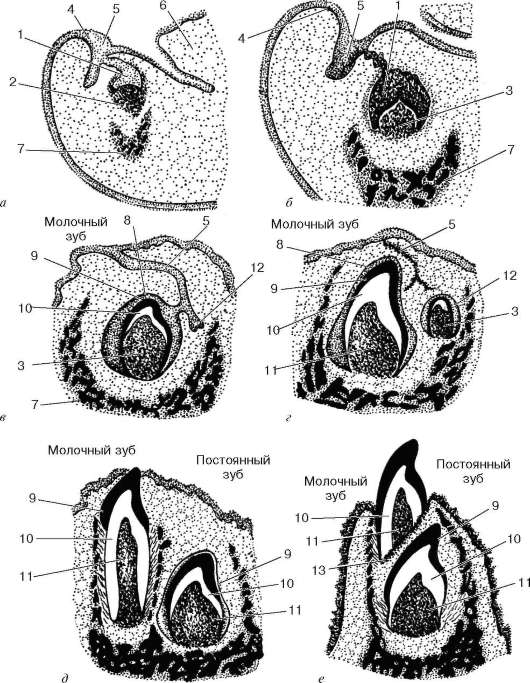
Рис. 16.10. Развитие и прорезывание молочных зубов и закладка постоянного зуба (по А. Хэму и Д. Кормаку):
а-е - последовательные стадии: 1 - закладка молочного зуба; 2 - участок мезенхимы; 3 - зубной сосочек; 4 - многослойный плоский эпителий ротовой полости; 5 - челюстно-губное выпячивание; 6 - язык; 7 - закладка нижней челюсти; 8 - эмалевый орган; 9 - эмаль; 10 - дентин; 11 - пульпа; 12 - закладка постоянного зуба; 13 - остеокласты
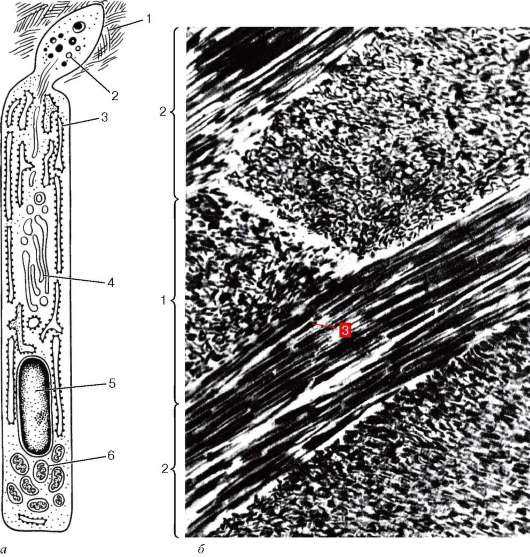
Рис. 16.11. Ультрамикроскопическое строение энамелобласта (амелобласта): а - схема (по Ю. И. Афанасьеву): 1 - эмаль; 2 - гранулы в дистальных отделах энамелобласта; 3 - эндоплазматическая сеть; 4 - комплекс Гольджи; 5 - ядро; 6 - митохондрии; б - микрофотография: 1 - поперечно перерезанные эмалевые призмы; 2 - продольно перерезанные эмалевые призмы; 3 - кристаллы гидроокси-апатитов (по Тревист и Глимчер)
Дифференцировка зубного зачатка начинается в тот период, когда в зубном сосочке разрастаются кровеносные капилляры и появляются первые нервные волокна. В конце 3-го мес эмалевый орган полностью отделяется от зубной пластинки.
Третий этап - развитие тканей зуба - начинается на 4-м мес эмбриогенеза. В периферическом слое пульпы развивающегося зуба мезенхимные клетки дифференцируются сначала в преодонтобласты, а затем одонтобла-
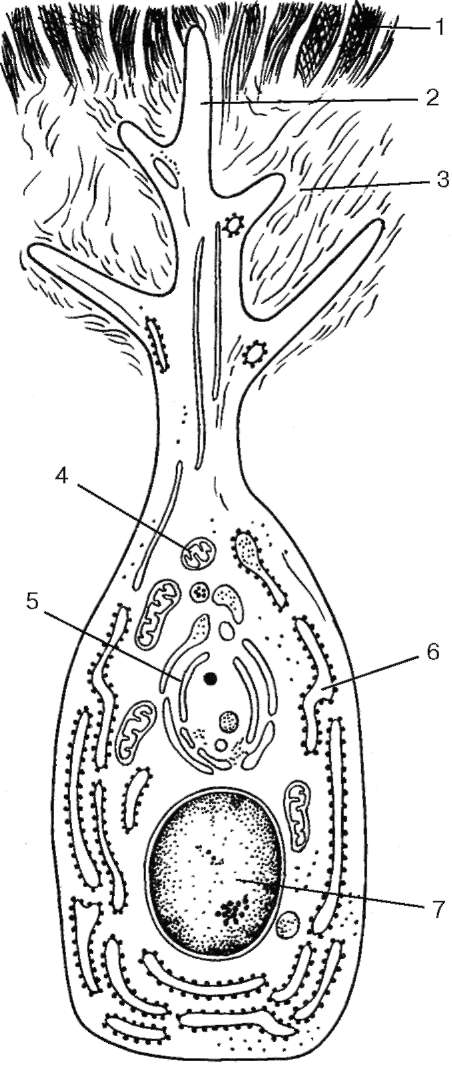
Рис. 16.12. Ультрамикроскопическое строение дентинобласта (одонтобласта) (по Ю. И. Афанасьеву): 1 - дентин; 2 - дистальный отросток дентинобласта; 3 - предентин; 4 - митохондрии; 5 - комплекс Гольджи; 6 - гранулярная эндоплазматическая сеть; 7 - ядро
сты, или дентинобласты (рис. 16.12). Этот процесс начинается раньше и активнее протекает на вершине, а позднее на боковых поверхностях зуба. Он совпадает по времени с подрастанием нервных волокон к ден-тинобластам. Одним из факторов их дифференцировки является базаль-ная мембрана внутреннего эмалевого эпителия. Дентинобласты синтезируют коллаген I типа, гликопротеи-ны, фосфопротеины, протеогликаны и фосфорины, характерные только для дентина. Прежде всего образуется плащевой дентин, расположенный непосредственно под базальной мембраной. Коллагеновые фибриллы в матриксе плащевого дентина располагаются перпендикулярно базальной мембране внутренних клеток эмалевого органа («радиальные волокна Корфа»). Между ради-ально расположенными волокнами залегают отростки дентинобластов.
Минерализация дентина начинается в коронке зуба, а затем в корне, путем отложения кристаллов гидро-оксиапатита на поверхности колла-геновых фибрилл, расположенных вблизи отростков одонтобластов (пе-ритубулярный дентин).
Дентинобласты - клетки мезен-химной природы, высокие призматические, с четко выраженной полярной дифференциацией. Их верхушечная часть имеет отростки, через которые происходит секреция органических веществ, образующих матрицу дентина - предентин. Преколлагеновые и коллагеновые фибриллы матрицы имеют радиальное направление. Это мягкое вещество заполняет промежутки между дентинобластами и клетками внутреннего эмалевого эпителия - энамелобластами. Количество предентина постепенно увеличивается. Позднее, когда происходит кальци-фикация дентина, эта зона входит в состав плащевого дентина. В стадии обызвествления дентина соли кальция, фосфора и других минеральных
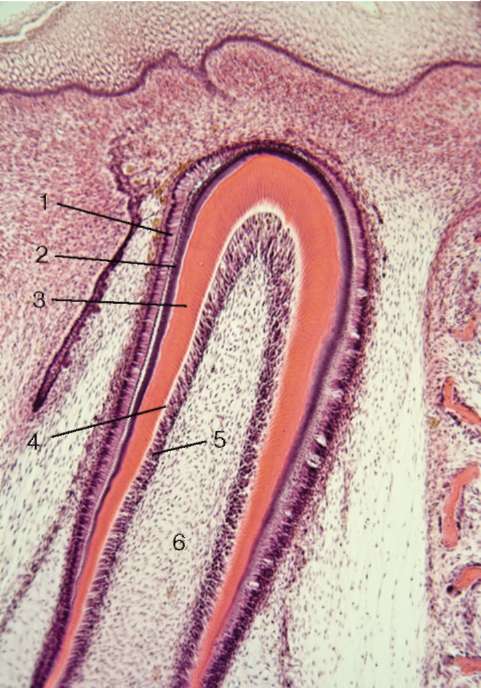
Рис. 16.13. Верхушка зачатка зуба (микрофотография):
1 - энамелобласты; 2 - эмаль; 3 - дентин; 4 - предентин; 5 - слой дентино-бластов; 6 - зубной сосочек
веществ откладываются в виде глы-бок, которые объединяются в глобулы. В дальнейшем развитие дентина замедляется, а около пульпы появляются тангенциальные коллагено-вые волокна околопульпарного дентина (рис. 16. 13).
В конце 5-го мес внутриутробного развития в предентине зачатка зуба начинаются отложение известковых солей и формирование окончательного дентина. Однако процесс обызвествления предентина при этом не захватывает участки, окружающие апикальные отростки ден-тинобластов. Это ведет к возникновению системы радиальных каналов, идущих от внутренней поверхности дентина к наружной поверхности. Кроме того, участки предентина на границе с эмалью также остаются необызвествленными и носят название интерглобулярных пространств.
Параллельно с развитием дентина идет процесс дифференцировки пульпы, в которой фибробласты постепенно синтезируют основное вещество, содержащее преколлагеновые и коллагеновые волокна. Гистохимически в периферической части пульпы, в области расположения дентинобластов и предентина, обнаруживаются ферменты, гидролизующие фосфатные соединения (фосфогидролазы), благодаря которым фосфатные ионы доставляются дентину и эмали.
Отложение первых слоев дентина индуцирует дифференцировку клеток внутреннего эмалевого эпителия (энамелобластов), которые начинают продуцировать эмаль, покрывающую образованный слой дентина.
Энамелобласты - эпителиальные клетки призматической формы с хорошо выраженной полярной дифференциацией. Первые зачатки эмали появляются в виде кутикулярных пластинок на базальной поверхности энаме-лобластов в области коронки зуба. Однако с началом эмалеобразования происходит перемещение, или инверсия, ядра и органелл клетки (центросомы и комплекса Гольджи) в апикальную часть клетки. В результате базаль-ная часть энамелобластов становится апикальной, а апикальная - базаль-ной. После такого изменения полюсов клеток питание клеток начинает осуществляться со стороны пульпы эмалевого органа, а не со стороны дентина. В подъядерной зоне энамелобластов обнаруживаются большое количество рибонуклеиновой кислоты, а также гликоген и высокая активность
щелочной фосфатазы. Этот полюс энамелобласта вытягивается, образуя пальцевидный отросток. При дальнейшем образовании эмали в прилежащих к отросткам участках цитоплазмы энамелобластов появляются гранулы, которые постепенно перемещаются в отростки. Образование эмали происходит путем мерокринной секреции, в результате чего содержимое гранул оказывается в межклеточном пространстве. Начинается их кальцинация и образование предэмалевых призм. Минерализация эмали в отличие от таковой дентина и цемента происходит очень быстро после образования органической матрицы. Этому способствуют амелогенины - белки некол-лагенового типа.
При дальнейшем развитии эмали энамелобласты уменьшаются в размерах и отодвигаются от дентина. К завершению этого процесса, примерно к моменту прорезывания зубов, энамелобласты резко уменьшаются и редуцируются, а эмаль оказывается покрытой лишь тонкой оболочкой - кутикулой, образованной клетками пульпы эмалевого органа. Образование эмали происходит циклически, в результате чего в ее структуре отмечается исчер-ченность. В зрелой эмали содержится более 95 % минеральных веществ.
Наружный эпителий эмалевого органа при прорезывании зуба сливается с эпителием десны и в дальнейшем разрушается. С появлением эмалевых призм поверхность дентина делается неровной. Частичная резорбция дентина, очевидно, способствует укреплению его связи с эмалью и усилению кальцинации эмали высвободившимися солями кальция.
Развитие цемента происходит позднее эмали, незадолго до прорезывания зубов, из окружающей зубной зачаток мезенхимы, образующей зубной мешочек. В последнем различают два слоя: более плотный - наружный и рыхлый - внутренний. В процессе развития цемента во внутреннем слое зубного мешочка в области корня из мезенхимы дифференцируются цемен-тобласты. Цементобласты, подобно остеобластам и дентинобластам, синтезируют коллагеновые белки и другие компоненты межклеточного вещества. По мере развития цементобласты превращаются в отростчатые цементоци-ты, которые погружаются в межклеточное вещество. Цементоциты расположены в полостях и отходящих от них канальцах. Наружный слой зубного мешочка превращается в зубную связку - периодонт. Таким образом, эмалевый орган играет, прежде всего, морфогенетическую роль, определяя форму развивающегося зуба.
Закладка постоянных зубов начинается в конце 4-го - начале 5-го мес внутриутробного развития (первых 10 зубов, сменяющих 10 молочных), а заканчивается в возрасте 2,5-3 лет. Зачаток постоянного зуба находится позади каждого зачатка молочного зуба. Прорезывание молочных зубов у ребенка начинается на 6-7-м мес жизни. К этому времени сформирована только коронка зуба, а формирование корня лишь начинается. Молочные большие коренные зубы (моляры) заменяются постоянными малыми коренными (премолярами).
Закладка постоянных больших коренных зубов происходит на 1- 4-м году жизни. Сначала оба зуба (молочный и постоянный) лежат в общей альвеоле. Затем между ними появляется костная перегородка.
Постоянный зуб развивается очень медленно. Когда наступает время выпадения молочных зубов, т. е. в возрасте 6-7 лет, остеокласты разрушают эту перегородку и корень выпадающего зуба, а постоянный зуб начинает усиленно развиваться. Так же как и молочные, постоянные зубы выталкиваются (прорезываются) под давлением, которое создается в пульпе зуба в связи с образованием основного вещества соединительной ткани. До прорезывания зубов минеральные вещества (кальций, фосфор, фтор и др.) и питательные вещества поступают только из крови. После прорезывания зубов возрастает роль слюны и соответственно ее химического состава.
Строение. Зуб состоит из твердых и мягких частей. В твердой части зуба различают эмаль, дентин и цемент; мягкая часть зуба представлена пульпой.
Эмаль (enamelum) покрывает коронку зуба. Наибольшего развития она достигает у вершины коронки (до 3,5 мм). Эмаль содержит незначительное количество органических веществ (около 3-4 %) и неорганические соли (96-97 %). Среди неорганических веществ подавляющую часть составляют фосфаты и карбонаты кальция и около 4 % - фторид кальция. Эмаль построена из эмалевых призм (prisma enameli) толщиной 3-5 мкм. Каждая призма состоит из тонкой фибриллярной сети, в которой находятся кристаллы гидрооксиапатитов (см. рис. 16.11). Призмы располагаются пучками, имеют извитой ход и залегают почти перпендикулярно к поверхности дентина. На поперечном срезе эмалевые призмы обычно имеют многогранную или вогнуто-выпуклую форму. Между призмами находится менее обыз-вествленное склеивающее вещество. Благодаря S-образно изогнутому ходу призм на продольных шлифах зуба одни из них оказываются рассеченными более продольно, а другие - более поперечно, что обусловливает чередование светлых и темных эмалевых полос. На продольных шлифах можно видеть еще более тонкие параллельные линии. Их появление связывают с периодичностью роста и различной зональной обызвествленностью призм, а также с отражением в структуре эмали силовых линий, возникающих в результате действия силового фактора во время жевания.
Снаружи эмаль покрыта тонкой кутикулой (cuticula enameli), которая на жевательной поверхности зуба быстро стирается и остается заметной лишь на его боковых поверхностях. Химический состав эмали меняется в зависимости от обмена веществ в организме, интенсивности растворения кристаллов гидро-оксиапатита и реминерализации органической матрицы. В небольших пределах эмаль проницаема для воды, ионов, витаминов, глюкозы, аминокислот и других веществ, поступающих непосредственно из полости рта. При этом большую роль играет слюна не только как источник поступления различных веществ, но и как фактор, активно влияющий на процесс проникновения их в ткани зуба. Проницаемость повышается под действием кислот, кальцитонина, спирта, дефицита в пище солей кальция, фосфора, фтора и др. Эмаль и дентин соединяются с помощью взаимных интердигитаций.
Дентин (dentinum) образует большую часть коронки, шейки и корня зубов. Он состоит из органических и неорганических веществ: органического вещества 28 % (главным образом коллагена), неорганических веществ 72 % (главным образом фосфат кальция и магния с примесью фторида кальция).
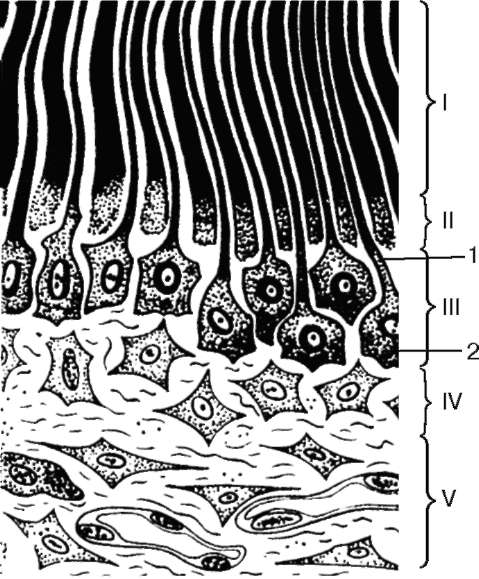
Рис. 16.14. Дентин и пульпа зуба (по Ю. И. Афанасьеву):
I - дентин; II - предентин; III - периферический слой пульпы; IV - промежуточный слой пульпы; V - центральный слой пульпы; 1 - дентинные канальцы с отростками дентинобластов; 2 - тела дентинобластов
Дентин построен из основного вещества, которое пронизано трубочками, или канальцами (tubuli dentinalis) (рис. 16.14). Основное вещество дентина содержит колла-геновые фибриллы и расположенные между ними мукопротеины. Коллагеновые фибриллы в дентине собраны в пучки и имеют преимущественно два направления: радиальное и почти продольное, или тангенциальное. Радиальные волокна преобладают в наружном слое дентина - так называемом плащевом дентине, тангенциальные - во внутреннем, околопульпарном дентине. В периферических участках дентина обнаруживаются так называемые интерглобулярные пространства, которые представляют собой его нео-бызвествленные участки, имеющие вид полостей, с неровными поверхностями. Наиболее крупные интерглобулярные пространства встречаются в коронке зуба, а мелкие, но многочисленные находятся в корне,
где они образуют зернистый слой. Интерглобулярные пространства принимают участие в обмене веществ дентина.
Основное вещество дентина пронизано дентинными канальцами, в которых проходят отростки дентинобластов, расположенных в пульпе зуба, и тканевая жидкость. Канальцы берут начало в пульпе, около внутренней поверхности дентина, и, веерообразно расходясь, заканчиваются на его наружной поверхности. В отростках дентинобластов обнаружена ацетилхолинэстераза, играющая большую роль в передаче нервного импульса. Количество канальцев в дентине, их форма и размеры неодинаковы в различных участках. Более плотно они расположены около пульпы. В дентине корня зуба канальцы ветвятся на всем протяжении, а в коронке они почти не дают боковых ветвей и распадаются на мелкие веточки около эмали. На границе с цементом дентин-ные канальцы также разветвляются, образуя анастомозирующие между собой аркады. Некоторые канальцы проникают в цемент и эмаль, особенно в области жевательных бугорков, и заканчиваются колбовидными вздутиями. Система канальцев обеспечивает питание дентина. Дентин в области, граничащей с эмалью, имеет обычно фестончатый край, что способствует более прочному их соединению. Внутренний слой стенки дентинных канальцев содержит много преколлагеновых аргирофильных волокон, которые сильно минерализованы по сравнению с остальным веществом дентина.

Рис. 16.15. Клеточный цемент корня зуба:
1 - цементоциты; 2 - минерализованное межклеточное вещество
На поперечных шлифах дентина заметны концентрические параллельные линии, появление которых, очевидно, связано с периодичностью роста дентина.
Между дентином и дентинобластами находится полоска предентина, или необызвествленного дентина, состоящего из коллагеновых волокон и аморфного вещества. В опытах с применением радиоактивного фосфора показано, что дентин растет постепенно путем наслоения нерастворимых фосфатов в предентине. Образование дентина не прекращается у взрослого человека. Так, вторичный, или заместительный, дентин, отличающийся нечеткой направленностью дентинных канальцев, наличием многочисленных интерглобулярных пространств, может быть как в предентине, так и пульпе (дентикли). Дентикли образуются при нарушении обмена веществ, при местных воспалительных процессах. Обычно они локализуются около дентинобластов, которые являются источником их развития. Небольшое количество солей может проникать в дентин через периодонт и цемент.
Цемент (cementum) покрывает корень зуба и шейку и в виде тонкого слоя может частично заходить на эмаль. По направлению к верхушке корня цемент утолщается.
По химическому составу цемент приближается к кости. В нем содержится около 30 % органических веществ и 70 % неорганических веществ, среди которых преобладают соли фосфата и карбоната кальция.
По гистологическому строению различают бесклеточный, или первичный, и клеточный, или вторичный, цемент. Бесклеточный цемент располагается преимущественно в верхней части корня, а клеточный - в его нижней части (рис. 16.15). В многокорневых зубах клеточный цемент залегает главным образом у разветвлений корней. Клеточный
цемент содержит клетки - цементоциты - многочисленные коллагено-вые волокна, которые не имеют определенной ориентации; поэтому клеточный цемент по строению и составу сравнивают с грубоволокнистой костной тканью. Цемент не содержит кровеносных сосудов. Клеточный цемент может иметь слоистое строение.
В бесклеточном цементе нет ни клеток, ни их отростков. Он состоит из коллагеновых волокон и аморфного вещества. Коллагеновые волокна проходят в продольном и радиальном направлениях. Радиальные волокна непосредственно продолжаются в периодонт и далее в виде прободающих волокон входят в состав альвеолярной кости. С внутренней стороны они сливаются с коллагеновыми радиальными волокнами дентина.
Питание цемента осуществляется диффузно через кровеносные сосуды периодонта. Циркуляция жидкости в твердых частях зуба происходит за счет ряда факторов: давления крови в сосудах пульпы и периодонта, которое изменяется при перепаде температур в полости рта при дыхании, приеме пищи, жевании и др. Определенный интерес представляют данные о наличии анастомозов дентинных канальцев с отростками клеток цемента. Такая связь канальцев служит дополнительной питательной системой для дентина в случае нарушения кровоснабжения пульпы (воспаление, удаление пульпы, пломбирование канала корня, заращение полости и т. п.).
Пульпа (pulpa dentis), или зубная мякоть, находится в коронковой полости зуба и в корневых каналах. Она состоит из рыхлой соединительной ткани, в которой различают три слоя: периферический, промежуточный и центральный (см. рис. 16.14).
Периферический слой пульпы состоит из нескольких рядов многоотростча-тых клеток грушевидной формы - дентинобластов, отличающихся базофи-лией цитоплазмы. Длина их не превышает 30 мкм, ширина - 6 мкм. Ядро дентинобласта лежит в базальной части клетки. От апикальной поверхности дентинобласта отходит длинный отросток, который проникает в ден-тинный каналец. Полагают, что эти отростки дентинобластов участвуют в снабжении минеральными солями дентина и эмали. Боковые отростки дентинобластов короткие. По своей функции дентинобласты сходны с остеобластами кости. В дентинобластах обнаружена щелочная фосфатаза, играющая активную роль в процессах кальцинирования зубных тканей, а в их отростках, кроме того, выявлены мукопротеиды. В периферическом слое пульпы находятся незрелые коллагеновые волокна. Они проходят между клетками и продолжаются далее в коллагеновые волокна дентина.
В промежуточном слое располагаются незрелые коллагеновые волокна и мелкие клетки, которые, подвергаясь дифференцировке, заменяют отжившие дентинобласты.
Центральный слой состоит из рыхло лежащих клеток, волокон и кровеносных сосудов. Среди клеточных форм этого слоя различают адвентициаль-ные клетки, макрофаги и фибробласты. Между клетками обнаруживаются как аргирофильные, так и коллагеновые волокна. Эластических волокон в пульпе зуба не обнаружено.
Пульпа зуба участвует в питании тканей зуба. Удаление пульпы резко затормаживает обменные процессы, нарушает развитие, рост и регенерацию тканей зуба.
Зубодесневое соединение. Зуб укрепляется в альвеоле челюсти с помощью периодонта и сращения многослойного плоского эпителия с кутикулой шейки зуба.
Периодонт (перицемент) образован плотной соединительной тканью, состоящей из толстых пучков коллагеновых волокон, идущих в основном в горизонтальном и косом направлениях. Периодонт не только удерживает зуб в лунке челюсти, но и амортизирует давление при жевании, а также благодаря большому количеству рецепторных окончаний является рефлексогенной зоной.
Эпителий десны - многослойный плоский ороговевающий, особенно на вестибулярной поверхности. Под эпителием располагается собственная пластинка соединительной ткани, которая плотно срастается с надкостницей альвеолы. Железы в десне отсутствуют. Между поверхностью зуба и десной имеется карман, дно которого расположено на уровне соединения эмали и цемента.
Нарушение целостности зубодесневого соединения может привести к инфицированию и воспалению.
Васкуляризация и иннервация. Сосуды (разветвления верхнечелюстной артерии) вместе с нервами (разветвления тройничного нерва) проникают в полость зуба через основной и дополнительный каналы, расположенные в корне зуба. Артерии входят в корень зуба одним или несколькими стволиками. Разветвляясь в пульпе на множество анастомозирующих капилляров, они собираются далее в вену. В пульпе обнаружено небольшое количество лимфатических капилляров.
Нервы образуют в пульпе зуба два сплетения: глубокое состоит преимущественно из миелиновых волокон, поверхностное - из безмиелиновых. Терминальные разветвления рецепторов пульпы нередко связаны одновременно с соединительной тканью и сосудами пульпы (поливалентные рецепторы). Дентинобласты густо оплетаются тонкими окончаниями тройничного нерва.
Вопрос о природе чувствительности дентина окончательно не решен. Многие исследователи отрицают данные о проникновении нервных окончаний в дентинные канальцы, хотя в начальных отделах этих канальцев окончания иногда обнаруживаются (рис. 16.16).
Возможно, в возникновении боли определенную роль играет гидродинамический механизм раздражения нервных окончаний, расположенных во внутренних отделах дентинных канальцев (передача давления столба жидкости, циркулирующей по дентинным канальцам, на терминали чувствительных нейронов).
Возрастные изменения. На протяжении первых 12-15 лет жизни происходит последовательная смена молочных зубов на постоянные. Первым прорезывается большой коренной зуб (первый моляр), затем центральные и боковые резцы, в 9-14 лет прорезываются премоляры и клыки, и только в 20-25 лет - «зуб мудрости».
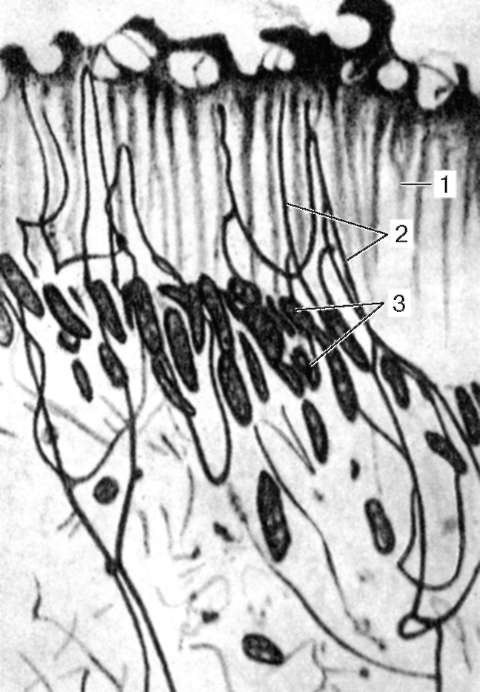
Рис. 16.16. Нервные окончания в зубе (по Бернику):
1 - предентин; 2 - нервные окончания; 3 - дентинобласты
Одновременно с возрастом происходят постепенные изменения в химическом составе и структуре зубов. Эмаль и дентин на их жевательной поверхности стираются. Эмаль тускнеет и может давать трещины, на ней откладывается минерализованный налет. Содержание органических соединений в эмали, дентине и цементе уменьшается, а количество неорганических веществ возрастает. В связи с этим изменяется проницаемость эмали, дентина и цемента для воды, ионов, ферментов, аминокислот и других веществ. С возрастом новообразование дентина почти полностью прекращается, количество же цемента в корне зуба увеличивается. Пульпа зуба с возрастом подвергается атрофии в результате ухудшения питания, вызванного склеротическим изменением ее сосудов. Количество клеточных элементов при этом уменьшается. В дентинобластах отмечается редукция значительной части клеточных
органелл, снижается пиноцитозная активность клетки. Дентинобласты превращаются в дентиноциты. Коллагеновые волокна грубеют. После 40-50 лет в периодонте часто выявляются склеротические изменения сосудов.
Регенерация. Регенерация тканей зуба происходит очень медленно и не полностью. При повреждении дентина или раздражениях его кариозным процессом в зубе со стороны пульпы против очага повреждения образуется небольшое количество заместительного, или вторичного, дентина. Этот процесс сопровождается регенерацией периферического слоя пульпы путем развития малодифференцированных клеток промежуточной зоны в денти-нобласты. Показано также, что в дентинобластическом слое пульпы на всех стадиях развития зуба содержатся клетки, обладающие способностью к пролиферации. Образование дентина происходит примерно через 2 нед после повреждения. Этот процесс начинается с появления предентина. Волокна в основном веществе заместительного дентина в отличие от первичного околопульпарного дентина располагаются без определенного порядка. К концу 4-й нед предентин обызвествляется. Канальцы заместительного дентина имеют неправильную ориентацию и очень слабо ветвятся. Цемент зуба регенерирует плохо. Восстановления эмали после повреждения зуба не происходит вообще. При воздействии на эмаль патогенных факторов эмаль реагирует образованием зон гиперминерализации.
16.2.5. Глотка
В глотке перекрещиваются дыхательный и пищеварительный пути. В ней выделяют три отдела: носовой, ротовой и гортанный. Они различаются по строению слизистой оболочки.
Слизистая оболочка носового отдела глотки покрыта многорядным реснитчатым эпителием, содержит смешанные железы (респираторный тип слизистой оболочки).
Слизистая оболочка ротового и гортанного отделов выстлана многослойным (плоским) эпителием, располагающимся на собственной пластинке слизистой оболочки, в которой имеется хорошо выраженный слой эластических волокон. В подслизистой основе лежат концевые отделы слизистых желез. Выводные протоки их открываются на поверхности эпителия. Слизистая оболочка и подслизистая основа глотки примыкают к мышечной стенке (аналог мышечной оболочки), которая состоит из двух слоев поперечнополосатых мышц - внутреннего продольного и наружного кольцевого. Снаружи глотка окружена адвентициальной оболочкой.
16.2.6. Пищевод
Развитие. Эпителий пищевода образуется из прехордальной пластинки, расположенной в составе выстилки передней кишки, остальные слои - из окружающей мезенхимы. Особый интерес представляет вопрос о развитии эпителия пищевода, который в течение всего эмбрионального периода претерпевает многократные изменения. Сначала эпителиальная выстилка пищевода представлена однослойным призматическим эпителием. У 4-недельного эмбриона он становится двухслойным. После этого наблюдается интенсивное разрастание эпителия, которое приводит к полному закрытию просвета пищевода. Разросшиеся эпителиальные клетки в дальнейшем подвергаются апоптозу, вновь освобождая просвет пищевода. К 3-му мес внутриутробного развития пищевод выстлан многорядным реснитчатым эпителием. С 4-го мес реснитчатые клетки постепенно вытесняются пузырьковидными, содержащими гликоген клетками, которые преобразуются в плоские клетки. С 6-го мес эпителий пищевода становится многослойным плоским. У новорожденных в эпителии могут встречаться островки реснитчатых клеток. У взрослых эти клетки сохраняются изредка только в протоках слизистых желез. Причины трансформации одного вида эпителия в другой неясны. Образование многослойного эпителия в слизистой оболочке пищевода обеспечивает сохранность стенки пищевода при прохождении грубых комков пищи. Железы пищевода появляются в конце 2-го мес, мышечная оболочка пищевода - на 2-м мес, а мышечная пластинка слизистой оболочки - на 4-м мес внутриутробного развития плода.
Строение. Стенка пищевода состоит из слизистой оболочки, подслизистой основы, мышечной и адвентициальной оболочек (рис. 16.17). Слизистая
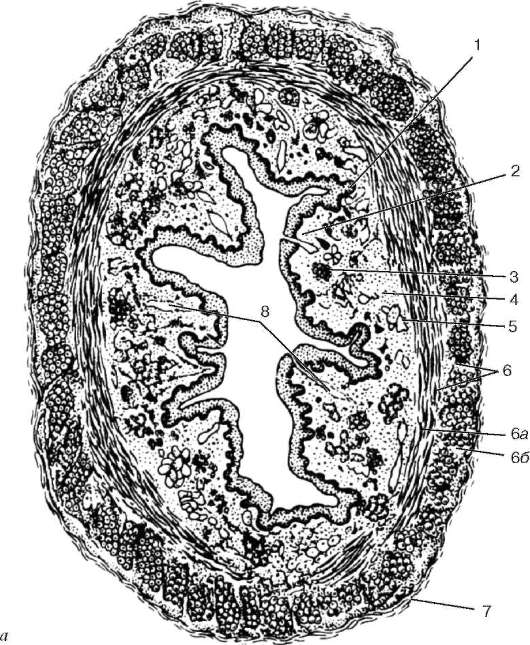
Рис. 16.17. Строение пищевода:
а - поперечный срез в средней трети (схема); б - микрофотография сечения пищевода; в - микрофотография собственных желез пищевода. 1 - многослойный плоский неороговевающий эпителий; 2 - собственная пластинка слизистой оболочки; 3 - мышечная пластинка слизистой оболочки; 4 - подслизистая основа; 5 - собственные железы пищевода; 6 - мышечная оболочка: 6а - циркулярный слой, 6б - продольный слой; 7 - адвентициальная оболочка; 8 - выводные протоки железы
оболочка и подслизистая основа образуют в пищеводе 7-10 продольно расположенных складок, вдающихся в его просвет.
Слизистая оболочка пищевода построена из эпителия, собственной и мышечной пластинок. Эпителий слизистой оболочки - многослойный плоский неороговевающий, но в пожилом возрасте его поверхностные клетки могут подвергаться ороговению. В составе пласта имеется 20-25 клеточных слоев эпителиоцитов, находящихся на последовательных стадиях диф-ференцировки. Кроме того, присутствуют гистологические элементы других клеточных дифферонов (клетки Лангерганса, эндокриноциты, интраэпите-
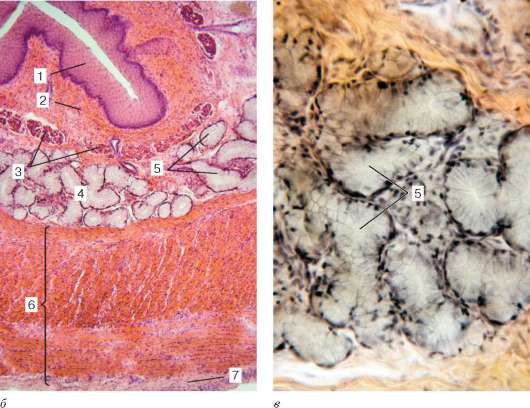
Рис. 16.17. Продолжение (обозначения см. выше)
лиальные лимфоциты). В пищеводе человека плоские клетки поверхностного слоя эпителия содержат небольшое количество зерен кератогиалина.
Собственная пластинка слизистой оболочки образует соединительнотканные сосочки, вдающиеся в эпителий. В ней находятся большие скопления лимфоцитов вокруг протоков слизистых желез, образуя даже отдельные лимфоидные узелки. В собственной пластинке расположены кардиальные железы пищевода (glandulae cardiacae oesophagi). Они представлены двумя группами. Одна группа желез залегает на уровне перстневидного хряща гортани и 5-го кольца трахеи, вторая группа находится в нижней части пищевода, около входа в желудок. По своему строению эти железы напоминают кардиальные железы желудка (отсюда их название). Это простые разветвленные трубчатые железы. Концевые отделы их образованы кубическими и призматическими эпителиоцитами с зернистой цитоплазмой, иногда содержащей муцин. В некоторых концевых отделах кардиальных желез находятся париетальные клетки, вырабатывающие хлориды. Самые мелкие протоки этих желез соединяются в расширенные крупные протоки, которые открываются на вершинах сосочков собственной пластинки слизистой оболочки. Эпителий протоков призматический. Он сходен со слизистым эпителием желудочных ямочек и дает характерную реакцию на муцин. Иногда на месте расположения кардиальных желез слизистая оболочка пищевода приобретает строение, характерное для слизистой оболочки желудка. Кардиальные
железы появляются в пищеводе задолго до того, как его эпителий становится многослойным. Кардиальные железы пищевода содержат большое количество эндокринных клеток (D-, G-, S- и др.), располагающихся главным образом в концевых отделах, а также в выводных протоках. Появление кардиальных желез в пищеводе объясняется меторизисом. Этим термином В. М. Шимкевич (1908) обозначил смещение границ между производными двух соседних эмбриональных зачатков. Полагают, что материал кишечной энтодермы смещается в сторону пищевода и дает начало железам желудочного типа в пределах слизистой оболочки пищевода. Для врача строение и функция кардиальных желез пищевода представляют интерес потому, что именно в местах их расположения часто образуются дивертикулы, кисты, язвы и опухоли пищевода.
Мышечная пластинка слизистой оболочки пищевода состоит из вдоль расположенных пучков гладких мышечных клеток, окруженных сетью эластических волокон. Эта пластинка начинается в виде отдельных небольших пучков на уровне перстневидного хряща гортани; далее по ходу пищевода толщина этого слоя увеличивается и около желудка достигает 200-400 мкм. Мышечная пластинка слизистой оболочки играет большую роль в проведении по пищеводу пищи и в защите внутренней его поверхности от повреждения острыми телами в случае их попадания в пищевод. Раздражение стенки пищевода грубым пищевым комком вызывает снижение тонуса гладких миоцитов мышечной пластинки, а это ведет к расширению соответствующего участка слизистой оболочки. Таким образом, прохождение комка пищи, содержащего грубые частицы, облегчается.
Подслизистая основа пищевода обеспечивает большую подвижность слизистой оболочки по отношению к мышечной оболочке. Вместе со слизистой оболочкой она образует многочисленные продольные складки, которые расправляются во время проглатывания пищи. В подслизистой основе находятся собственные железы пищевода.
Собственные железы пищевода (glandulae oesophagea propriae). Это сложные сильно разветвленные альвеолярно-трубчатые железы (см. рис. 16.17, б, в). Их концевые отделы состоят исключительно из слизистых клеток. Последние окружены миоэпителиоцитами. Секрет изливается в мелкие выводные протоки, которые объединяются в более крупные протоки. Последние проходят через мышечную пластинку слизистой оболочки и в собственной пластинке слизистой оболочки образуют крупные ампулообразные протоки, открывающиеся на поверхности эпителия. Эпителий, выстилающий мелкие протоки, низкий призматический, в более крупных протоках он многослойный плоский, иногда в нем встречаются реснитчатые клетки. Собственные железы пищевода располагаются главным образом на вентральной поверхности его верхней трети. Функция собственных желез пищевода состоит в выделении слизи, постоянно увлажняющей поверхность слизистой оболочки и способствующей прохождению пищевых комков.
Мышечная оболочка состоит из внутреннего циркулярного и наружного продольного слоев, разделенных прослойкой рыхлой волокнистой соединительной ткани. В верхней трети пищевода эти слои представлены висце-
ральной поперечнополосатой мышечной тканью миотомного происхождения, которая является продолжением мышечной ткани глотки. В нижней трети оба слоя образованы только гладкой мышечной тканью мезенхим-ной природы. В средней трети пищевода мышечная оболочка содержит как поперечнополосатую, так и гладкую мышечные ткани. Это обстоятельство может служить ориентиром для определения уровня пищевода на гистологическом срезе. Оба мышечных слоя не всегда лежат точно циркулярно или продольно. Во внутреннем слое встречаются спиральные и косые пучки, в наружном слое отдельные пучки также могут иметь неодинаковое расположение. Утолщение внутреннего циркулярного слоя мышечной оболочки на уровне перстневидного хряща образует верхний сфинктер пищевода, а утолщение этого слоя на уровне перехода пищевода в желудок - нижний сфинктер пищевода. Сокращение мышечной оболочки пищевода способствует проталкиванию пищи в пищеводе по направлению к желудку.
Адвентициальная оболочка с одной стороны связана с прослойками соединительной ткани, расположенными в мышечной оболочке, а с другой - с окружающей пищевод соединительной тканью средостения. В адвенти-циальной оболочке много продольно ориентированных сосудов и нервов. Брюшной отдел пищевода покрыт серозной оболочкой, образованной мезо-телием с подлежащей соединительной тканью.
Васкуляризация. Артерии, входящие в пищевод, образуют сплетения в подслизистой основе (крупнопетлистые и мелкопетлистые), из которых кровь поступает в крупнопетлистое сплетение собственной пластинки слизистой оболочки. Здесь же имеется подэпителиальная сеть кровеносных капилляров. Венозный отток начинается сетью мелких вен в собственной пластинке слизистой оболочки. Эти вены несут кровь в венозное сплетение подслизистой основы, а оттуда - в адвентициаль-ную оболочку. Лимфатическая система пищевода представлена сетями лимфатических капилляров, расположенных в собственной пластинке слизистой оболочки, в подслизистой основе и мышечной оболочке, а также сплетениями лимфатических сосудов, которые лежат в подсли-зистой основе и мышечной оболочке (иногда и в адвентициальной оболочке). Главным коллектором оттока лимфы из пищевода является под-слизистое сплетение.
Иннервация. Интрамуральный нервный аппарат образован связанными между собой четырьмя сплетениями: адвентициальным - наиболее развитым в средней и нижней третях пищевода; субадвентициальным - лежащим на поверхности мышечной оболочки и хорошо выраженным только в верхних частях пищевода; межмышечным - находящимся между циркулярным и продольным мышечным слоями и состоящим из нервных волокон разного калибра, образующих широкопетлистое сплетение, в составе которого располагаются крупные нервные узлы, и подслизистым сплетением, обнаруживающимся на всем протяжении пищевода и состоящим из системы нервных стволов, в местах ветвления которых лежат мелкие нервные узелки. Межмышечное сплетение самое мощное. Чувствительные нервные окончания выявляются в виде инкапсулированных клубочков,
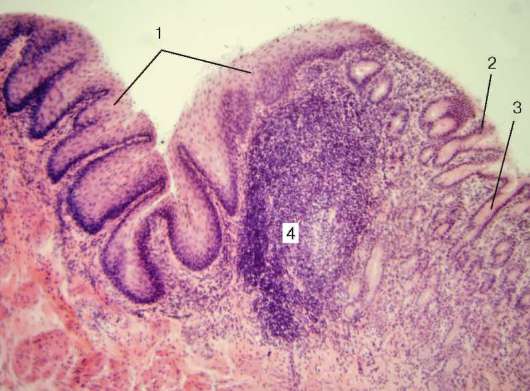
Рис. 16.18. Переход пищевода в желудок (микрофотография):
1 - многослойный плоский неороговевающий эпителий пищевода; 2 - однослойный столбчатый эпителий желудка; 3 - желудочные ямочки; 4 - лимфоидный узелок
кустиков, древовидных разветвлений в мышечных тканях пищевода, в эпителии и соединительной ткани слизистой оболочки и подслизистой основе. В интрамуральных ганглиях обнаруживаются крупные инкапсулированные клубочковые рецепторы, которые, по-видимому, являются баро- и механо-рецепторами.
На гистологических срезах в области границы пищевода и желудка виден стык многослойного плоского эпителия пищевода и однослойного столбчатого эпителия желудка (рис. 16.18).
В собственной пластинке слизистой оболочки пищевода в этой области располагаются кардиальные железы, а в собственной пластинке слизистой оболочки желудка наряду с желудочными железами могут находиться слизистые железы пищевода. Кольцевой слой мышечной оболочки пищевода образует здесь утолщение, формируя сфинктер.
16.3. СРЕДНИЙ И ЗАДНИЙ ОТДЕЛЫ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
В среднем отделе пищеварительного тракта происходит главным образом химическая обработка пищи под воздействием ферментов, вырабатываемых железами, всасывание продуктов переваривания пищи, формирование каловых масс.
16.3.1. Желудок
Желудок выполняет в организме ряд важнейших функций. Главной из них является секреторная. Она заключается в выработке железами желудочного сока. В его состав входят ферменты пепсин, ренин, липаза, а также соляная кислота и слизь.
Пепсин - основной фермент желудочного сока, с помощью которого в желудке начинается процесс переваривания белков. Пепсин вырабатывается в неактивной форме в виде пепсиногена, который в содержимом желудка в присутствии соляной кислоты превращается в активную форму - пепсин.
Из пепсиногена у человека образуется несколько близких по строению пепсинов, а также пепсиноподобный фермент гастриксин. Эти ферменты наиболее активны в кислой среде (для пепсина оптимум рН 1,5-2,5, для гастриксина рН 3,0). Кроме того, в желудочном соке детей грудного возраста открыт фермент ренин, который близок по свойствам пепсину.
Пепсин гидролизует большинство белков, поступивших с пищей, до полипептидов меньших размеров (альбумозы и пептоны), которые далее поступают в кишечник и подвергаются ферментативному распаду до конечных продуктов - свободных аминокислот. Однако некоторые белки (кератины, гистоны, протамины, мукопро-теиды) пепсином не расщепляются.
Ренин у грудных детей превращает растворимый казеиноген молока в нерастворимый казеин (створаживание молока). У взрослых людей эту функцию выполняет пепсин.
Липаза содержится в желудочном соке в небольшом количестве, у взрослых малоактивна, у детей расщепляет жиры молока.
Слизь, покрывая поверхность слизистой оболочки желудка, и бикарбонат в составе слизи (ионы НСО3-) предохраняют ее от действия хлористоводородной кислоты и от повреждения грубыми комками пищи.
Осуществляя химическую переработку пищи, желудок вместе с тем выполняет еще некоторые важные для организма функции. Механическая функция желудка состоит в перемешивании пищи с желудочным соком и перемещении переработанной пищи в двенадцатиперстную кишку. В осуществлении функции принимает участие мускулатура желудка. В стенке желудка образуется антианемический фактор, который способствует поглощению витамина В12, поступающего с пищей. При отсутствии этого фактора у человека развивается злокачественное малокровие. Через стенку желудка происходит всасывание таких веществ, как вода, спирт, соли, сахар и др. Вместе с тем желудок выполняет и экскреторную функцию. Особенно наглядно эта функция проявляется при заболевании почек, когда через стенку желудка выделяется ряд конечных продуктов обмена белков (аммиак, мочевина и др.). Эндокринная функция желудка заключается в выработке ряда биологически активных веществ - гастрина, гистамина, серотонина, мотилина, энтероглюкагона и др. Эти вещества оказывают стимулирующее или тормозящее действие на моторику и секреторную активность железистых клеток желудка и других отделов пищеварительного тракта.
Развитие. Желудок появляется на 4-й нед внутриутробного развития, а в течение 2-го мес формируются все основные его отделы. Однослойный столбчатый эпителий желудка развивается из энтодермы кишечной трубки. Желудочные ямочки образуются в течение 6-10-й нед развития плода, железы закладываются в виде почек на дне желудочных ямочек и, разрастаясь, в дальнейшем располагаются в собственной пластинке слизистой оболочки. Вначале в составе желез появляются париетальные клетки, затем главные и слизистые клетки. В это же время (6-7-я нед) из мезенхимы формируются сначала кольцевой слой мышечной оболочки, затем - мышечная пластинка слизистой оболочки. На 13-14-й нед образуется наружный продольный и несколько позднее - внутренний косой слой мышечной оболочки. Мезотелий возникает из висцерального листка спланхнотома.
Строение. В желудке различают кардиальный, фундальный отделы, тело желудка и пилорический отделы. Стенка желудка состоит из слизистой оболочки, подслизистой основы, мышечной и серозной оболочек.
Для рельефа внутренней поверхности желудка характерно наличие трех видов образований - продольных желудочных складок, желудочных полей и желудочных ямочек.
Желудочные складки (plicae gastricae) образованы слизистой оболочкой и подслизистой основой. Желудочные поля (акае gastricae) представляют собой отграниченные друг от друга бороздками участки слизистой оболочки. Они имеют многоугольную форму и размер поперечника от 1 до 16 мм. Наличие полей объясняется тем, что железы желудка располагаются группами, отделенными друг от друга прослойками соединительной ткани. Поверхностно лежащие вены в этих прослойках просвечивают в виде красноватых линий, выделяя границы между полями. Желудочные ямочки (foveolae gastricae) - углубления эпителия в собственной пластинке слизистой оболочки. Они встречаются по всей поверхности желудка. Число ямочек в желудке достигает почти 3 млн. Желудочные ямочки имеют микроскопические размеры, но величина их неодинакова в различных отделах желудка. В кардиальном отделе и теле желудка их глубина составляет всего 1/4 толщины слизистой оболочки. В пилорической части желудка ямочки более глубокие. Они занимают около половины толщины всей слизистой оболочки. На дне желудочных ямочек открываются протоки желез. Слизистая оболочка наиболее тонкая в кардиальном отделе.
Слизистая оболочка желудка состоит из эпителия, собственной пластинки (l. propria mucosae) и мышечной пластинки (l. muscularis mucosae).
Эпителий, выстилающий поверхность слизистой оболочки желудка и ямочек, однослойный столбчатый железистый (см. рис. 16.19). Все поверхностные слизистые эпителиоциты желудка (epitheliocyti seu mucocyti superficiales gastrici) постоянно выделяют мукоидный (слизеподобный) секрет. Каждая железистая клетка четко подразделяется на две части - базальную и апикальную (рис. 16.20). В базальной части, прилежащей к базальной мембране, лежит овальной формы ядро, над которым располагается комплекс Гольджи. Апикальная часть клетки заполнена зернами или каплями муко-идного секрета. Специфичность секрета поверхностных эпителиоцитов
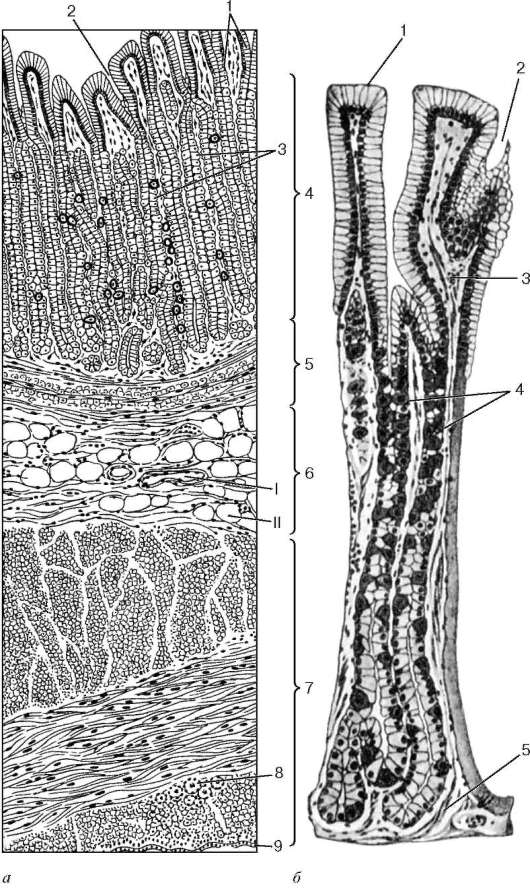
Рис. 16.19. Строение стенки дна желудка:
а, б - схемы; в - микрофотография. 1 - однослойный столбчатый железистый эпителий; 2 - желудочная ямочка; 3 - собственная пластинка слизистой оболочки; 4 - собственные железы желудка; 5 - мышечная пластинка слизистой оболочки; 6 - подслизистая основа: I - кровеносный сосуд, II - жировая клетка; 7 - мышечная оболочка; 8 - межмышечное нервное сплетение; 9 - серозная оболочка
Рис. 16.19. Продолжение (обозначения см. выше)
у человека и животных определяется составом углеводного компонента, в то время как белковая часть характеризуется общностью гистохимических свойств. Углеводный компонент играет определяющую роль в защитной реакции слизистой оболочки желудка на повреждающее действие желудочного сока. Роль поверхностных эпителиоцитов желудка заключается в выработке слизи, которая служит защитой как от механического воздействия грубых частиц пищи, так и от химического действия желудочного сока. Количество слизи в желудке сильно увеличивается при попадании в него раздражающих веществ (алкоголь, кислота, горчица и др.).
В собственной пластинке слизистой оболочки расположены железы желудка, между которыми лежат тонкие прослойки рыхлой волокнистой соединительной ткани. В ней в большем или меньшем количестве
всегда имеются скопления лимфоидных элементов либо в виде диффузных инфильтратов, либо в виде солитарных (одиночных) лимфоидных узелков, которые чаще всего располагаются в области перехода желудка в двенадцатиперстную кишку.
Мышечная пластинка слизистой оболочки состоит из трех слоев, образованных гладкой мышечной тканью: внутреннего и наружного циркулярных и среднего - продольного (см. рис. 16.19). От мышечной пластинки отдельные мышечные клетки отходят в соединительную ткань собственной пластинки слизистой оболочки. Сокращение мышечных элементов слизистой оболочки обеспечивает ее подвижность, а также способствует выведению секрета из желез желудка.
Железы желудка (gll. gastricae) в различных его отделах имеют неодинаковое строение. Различают три вида желудочных желез: собственные железы желудка, пилорические и кардиальные. Количественно преобладают собственные железы желудка. Они залегают в области тела и дна желудка (фундальные). Кардиальные и пилорические железы располагаются в одноименных частях желудка.
Собственные железы желудка (gll. gastricae propriae) - наиболее многочисленные. У человека их насчитывается около 35 млн. Площадь каждой железы составляет приблизительно 100 мм2. Общая секреторная поверхность фундальных желез достигает огромных размеров - около 3-4 м2. По
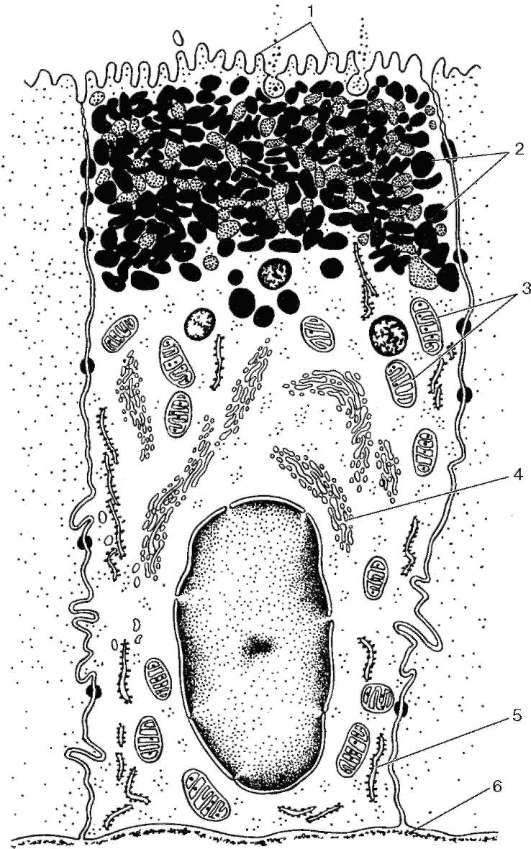
Рис. 16.20. Ультрамикроскопическое строение поверхностных эпителиоцитов желудка (по С. Ито):
1 - микроворсинки; 2 - гранулы слизистого секрета; 3 - митохондрии; 4 - комплекс Гольджи; 5 - гранулярная эндоплазматическая сеть; 6 - базальная мембрана
строению эти железы представляют собой простые неразветвленные, иногда очень слабо разветвленные трубчатые железы (см. рис. 16.19, б, в). Длина одной железы около 0,65 мм, диаметр ее колеблется от 30 до 50 мкм. Железы группами открываются в желудочные ямочки. В каждой железе различают перешеек (isthmus), шейку (cervix) и главную часть (pars principalis), представленную телом (corpus) и дном (fundus). Тело и дно составляют секреторный
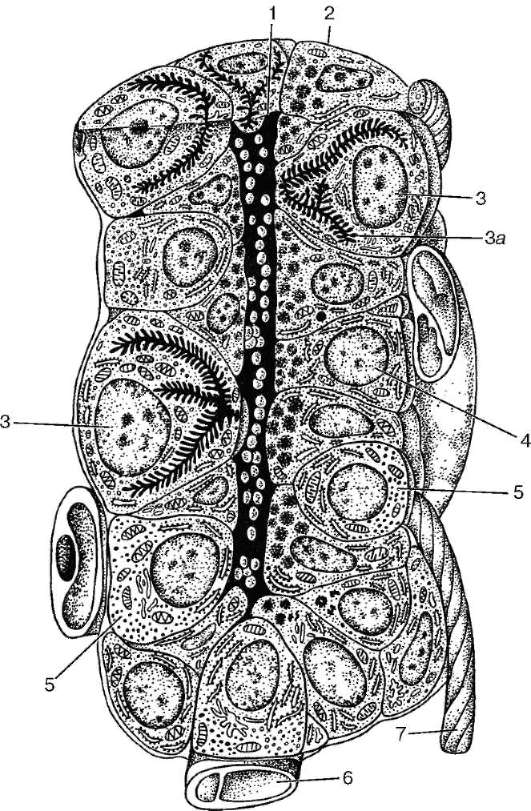
Рис. 16.21. Ультрамикроскопическое строение собственной железы желудка (по Р. В. Крстичу, с изменениями):
1 - просвет железы; 2 - шеечный мукоцит; 3 - париетальные экзокриноциты; 3а - внутриклеточные канальцы; 4 - главные экзокриноциты; 5 - эндокриноциты; 6 - капилляр; 7 - нервное волокно
отдел железы, а шейка и перешеек - ее выводной проток. Просвет в железах очень узкий и почти не виден в препаратах желудка, окрашенных общегистологическими красителями.
Собственные железы желудка содержат пять основных видов железистых клеток: главные экзокриноциты, париетальные экзокриноциты, шеечные слизистые клетки (мукоциты), эндокринные (аргирофильные), малодиффе-ренцированные (камбиальные) эпителиоциты.
Главные экзокриноциты (exocrinocyti principales) располагаются преимущественно в области дна и тела железы (рис. 16.21). Ядра этих клеток имеют округлую форму, лежат в центре клетки. В клетке выделяют базальную
и апикальную части. Базальная часть клетки окрашивается базофиль-но. Здесь находится хорошо развитый синтетический аппарат клетки. На апикальной плазмолемме обнаруживаются короткие микроворсинки. Под плазмолеммой в апикальной части клетки располагаются гранулы белкового секрета диаметром 0,9-1 мкм. Главные экзокриноциты секретируют пепсиноген - профермент (зимоген), который в присутствии соляной кислоты превращается в активную форму - пепсин. Предполагают, что ренин, расщепляющий белки молока, также вырабатывается главными клетками. При изучении различных фаз секреции главных экзокриноцитов выявлено, что в активной фазе выработки и накопления секрета эти клетки крупные, в них хорошо различимы гранулы зимогена. После выделения секрета величина клеток и количество гранул в их цитоплазме заметно уменьшаются. Экспериментально доказано, что при раздражении блуждающего нерва клетки быстро освобождаются от гранул пепсиногена.
Париетальные (обкладочные) экзокриноциты (exocrinocyti parietales) располагаются снаружи от главных и слизистых клеток, плотно прилегая к их базальным частям (см. рис. 16.21). Они больше главных клеток, неправильной формы. Париетальные клетки лежат поодиночке и сосредоточены главным образом в области тела и шейки железы. Цитоплазма этих клеток резко оксифильна. В каждой клетке содержится одно или два ядра округлой формы, лежащие в центре. Внутри клеток располагаются внутриклеточные секреторные канальцы (canaliculis intracellulares) с многочисленными микроворсинками и мелкими везикулами и трубочками, формирующими тубуловезикулярную систему, играющую важную роль в транспорте С1-. Внутриклеточные канальцы переходят в межклеточные канальцы, находящиеся между главными и слизистыми клетками и открывающиеся в просвет железы (см. рис. 16.21). От апикальной поверхности клеток отходят микроворсинки. Для париетальных клеток характерно наличие многочисленных митохондрий. Роль париетальных клеток собственных желез желудка заключается в выработке Н+-ионов и хлоридов, из которых образуется соляная кислота. Париетальные клетки также секретируют внутренний фактор Кастла, необходимый для всасывания витамина В12 в тонкой кишке. В базальной части клеток синтезируется и секретируется бикарбонат. Последний по кровеносным сосудам поступает к поверхностным муко-цитам, поглощается ими и выделяется на апикальной поверхности клетки. Здесь происходит нейтрализация соляной кислоты. Разрушение бикарбо-натного слоя (алкоголем, некоторыми лекарственными препаратами) ведет к очаговой гибели эпителиоцитов желудка.
Шеечные слизистые клетки располагаются только в шейке собственных желез. Ядра у них уплощенные, иногда неправильной треугольной формы, лежат обычно у основания клеток. В апикальной части этих клеток находятся секреторные гранулы. Слизь, выделяемая шеечными клетками, слабо окрашивается основными красителями, но хорошо выявляется муцикарми-ном. По сравнению с поверхностными клетками желудка шеечные клетки меньших размеров и всегда содержат значительно меньшее количество капель слизи. Их секрет по составу отличается от мукоидного секрета,
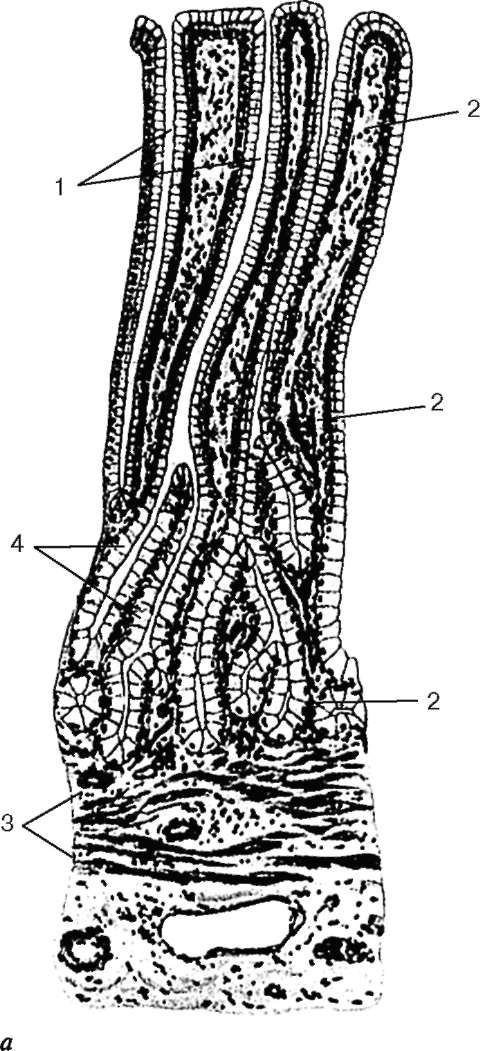
Рис. 16.22. Пилорический отдел желудка: а - схема; б - микрофотография. 1 - желудочные ямочки; 2 - собственная пластинка слизистой оболочки; 3 - мышечная пластинка слизистой оболочки; 4 - пилорические железы
выделяемого железистым эпителием желудка. В шеечных клетках в противоположность другим клеткам фундальных желез часто обнаруживаются фигуры митоза. Полагают, что эти клетки являются источником регенерации как секреторного эпителия желез, так и эпителия желудочных ямок.
В теле собственных желез обнаруживаются слизистые клетки, которые в своей базальной части содержат темное ядро (см. рис. 16.21). В апикальной части этих клеток находятся множество круглых или овальных гранул, небольшое количество митохондрий и комплекс Гольджи. Эти клетки рассматриваются как переходные формы с последующим развитием в главные экзокриноциты.
Согласно унитарной теории, источником развития эпителиоцитов является стволовая клетка в составе шейки железы, которая путем дивергентной дифференцировки дает начало клеточным дифферонам поверхностных, ямочных, слизистых желез, париетальных и главных эк-зокриноцитов, эндокриноцитов. Эндокринные клетки описаны далее.
Пилорические железы (gll. pyloricae) расположены в зоне перехода желудка в двенадцатиперстную кишку (рис. 16.22). Их число составляет около 3,5 млн. Пилорические железы отличаются от собственных желез несколькими признаками: они расположены редко, разветвлены, имеют широкие просветы; большинство
пилорических желез лишено париетальных клеток.
Концевые отделы пилорических желез построены в основном из клеток, напоминающих по своему строению слизистые клетки собственных желез желудка. Ядра их сплющены и лежат у основания клеток. В цитоплазме при использовании специальных методов окраски выявляется слизь. Клетки пилорических желез богаты дипептидазами. Секрет, вырабатываемый пило-рическими железами, имеет щелочную реакцию.
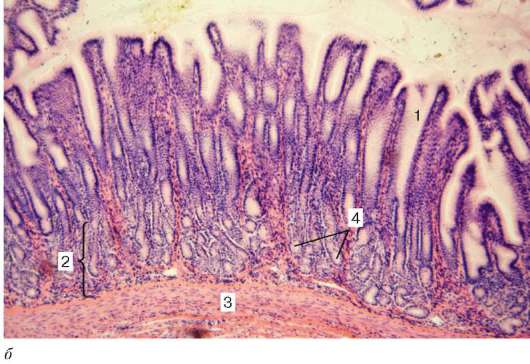
Рис. 16.22. Продолжение (обозначения см. выше)
Строение слизистой оболочки в пилорическом отделе имеет некоторые особенности: желудочные ямочки здесь более глубокие, чем в теле желудка, и занимают около половины всей толщины слизистой оболочки (см. рис. 16.22). Около выхода из желудка эта оболочка имеет хорошо выраженную кольцевую складку, что отражает наличие пилорического сфинктера. Последний образован циркулярно расположенными пучками гладких миоцитов мышечной оболочки желудка. Сфинктер регулирует поступление пищи из желудка в кишечник.
Кардиальные железы (gll. cardiacae) - простые трубчатые железы с сильно разветвленными концевыми отделами. Выводные протоки (шейки) этих желез короткие, выстланы призматическими клетками. Ядра клеток сплющенной формы, лежат у основания клеток. Цитоплазма их светлая. При специальной окраске муцикармином в ней выявляется слизь. По-видимому, секреторные клетки этих желез идентичны клеткам, выстилающим пило-рические железы желудка и кардиальные железы пищевода. В них также обнаружены дипептидазы. Иногда в кардиальных железах встречаются в небольшом количестве главные и париетальные клетки.
Желудочно-кишечные эндокриноциты (endocrinocyti gastrointestinales). В желудке по морфологическим, биохимическим и функциональным признакам выделено несколько видов эндокринных клеток.
ЕС-клетки - самые многочисленные, располагаются в области тела и дна желез между главными клетками. Эти клетки секретируют серотонин и мелатонин. Серотонин стимулирует секрецию пищеварительных ферментов, выделение слизи, двигательную активность. Мелатонин регулирует фотопериодичность функциональной активности (т. е. зависит от действия светового цикла). G-клетки (гастрин-продуцирующие) также многочисленны и находятся главным образом в пилориче-ских железах, а также в кардиальных, располагаясь в области их тела и дна, иногда
шейки. Выделяемый ими гастрин стимулирует секрецию пепсиногена главными клетками, соляной кислоты - париетальными, а также моторику желудка. При гиперсекреции желудочного сока у человека отмечается увеличение числа G-клеток. Кроме гастрина, эти клетки выделяют энкефалин, являющийся одним из эндогенных морфинов. Ему приписывают роль медиации боли. Менее многочисленными являются Р-, ECL-, D-, D1-, А-клетки и др.
Р-клетки секретируют бомбезин, стимулирующий выделение соляной кислоты и панкреатического сока, богатого ферментами, а также усиливают сокращение гладкой мускулатуры желчного пузыря. ECL-клетки (энтерохромаффиноподоб-ные) характеризуются разнообразием формы и располагаются главным образом в теле и дне фундальных желез. Эти клетки вырабатывают гистамин, который регулирует секреторную активность париетальных клеток, выделяющих хлориды. D- и D1-клетки выявляются главным образом в пилорических железах. Они являются продуцентами активных полипептидов. D-клетки выделяют соматостатин, ингиби-рующий синтез белка. D1-клетки секретируют вазоинтестинальный пептид (ВИП), который расширяет кровеносные сосуды и снижает артериальное давление, а также стимулирует выделение гормонов поджелудочной железы. А-клетки синтезируют глюкагон, т. е. имеют сходную функцию с эндокринными А-клетками островков поджелудочной железы.
Подслизистая основа желудка состоит из рыхлой соединительной ткани, содержащей большое количество эластических волокон (см. рис. 16.19, а). В ней расположены артериальное и венозное сплетения, сеть лимфатических сосудов и подслизистое нервное сплетение.
Мышечная оболочка желудка относительно слабо развита в области его дна, хорошо выражена в теле и наибольшего развития достигает в пилори-ческом отделе. В мышечной оболочке различают три слоя, образованных гладкими мышечными клетками (см. рис. 16.19, а). Наружный продольный слой является продолжением продольного мышечного слоя пищевода. Средний - циркулярный, также представляющий собой продолжение циркулярного слоя мышечной оболочки пищевода, наибольшего развития достигает в пилорическом отделе, где образует пилорический сфинктер толщиной около 3-5 см. Внутренний слой представлен пучками гладких мышечных клеток, имеющих косое направление. Между слоями мышечной оболочки располагаются межмышечное нервное сплетение и сплетения лимфатических сосудов.
Серозная оболочка желудка образует наружную часть его стенки, состоит из рыхлой соединительной ткани, которая с поверхности покрыта мезотелием.
Васкуляризация. Артерии, питающие стенку желудка, проходят через серозную и мышечную оболочки, отдавая им соответствующие ветви, а далее переходят в мощное сплетение в подслизистой основе. Веточки от него проникают через мышечную пластинку слизистой оболочки в ее собственную пластинку и образуют там второе сплетение. От этого сплетения отходят мелкие артерии, продолжающиеся в кровеносные капилляры, оплетающие железы и обеспечивающие питание эпителия желудка. Из кровеносных капилляров, находящихся в слизистой оболочке, кровь собирается в мелкие вены. Непосредственно под эпителием проходят отно-
сительно крупные посткапиллярные вены звездчатой формы (vv. stellatae). Повреждение эпителия желудка обычно сопровождается разрывом этих вен и значительным кровотечением. Вены слизистой оболочки, собираясь вместе, формируют сплетение, расположенное в собственной пластинке около артериального сплетения. Второе венозное сплетение располагается в подслизистой основе. Все вены желудка, начиная с вен, лежащих в слизистой оболочке, снабжены клапанами. Лимфатическая сеть желудка берет начало от лимфатических капилляров, слепые концы которых находятся непосредственно под эпителием желудочных ямочек и желез в собственной пластинке слизистой оболочки. Эта сеть сообщается с широкопетлистой сетью лимфатических сосудов, расположенной в подслизистой основе. От лимфатической сети отходят отдельные сосуды, пронизывающие мышечную оболочку. В них вливаются лимфатические сосуды из лежащих между мышечными слоями сплетений.
Иннервация. Желудок имеет два источника эфферентной иннервации: парасимпатический (от блуждающего нерва) и симпатический (из пограничного симпатического ствола). В стенке желудка располагаются три нервных сплетения: межмышечное, подслизистое и субсерозное. Нервные ганглии малочисленны в кардиальной области, увеличиваются в числе и размерах в пилорическом отделе.
Ганглии самого мощного - межмышечного - сплетения построены преимущественно из клеток I типа и незначительного количества клеток II типа. Наибольшее количество клеток II типа наблюдается в пилорическом отделе желудка. Подслизистое сплетение развито слабо. Возбуждение блуждающего нерва ведет к ускорению сокращения желудка и усилению выделения железами желудочного сока. Возбуждение симпатических нервов, наоборот, вызывает замедление сократительной деятельности желудка и ослабление желудочной секреции.
Афферентные волокна образуют чувствительное сплетение, расположенное в мышечной оболочке, волокна которого осуществляют рецептор-ную иннервацию нервных узлов, гладких мышц, соединительной ткани. В желудке обнаружены поливалентные рецепторы.
16.3.2. Тонкая кишка
Тонкая кишка подразделяется на три отдела: двенадцатиперстную, тощую и подвздошную. В тонкой кишке подвергаются химической обработке все виды питательных веществ - белки, жиры и углеводы. В переваривании белков участвуют ферменты панкреатического сока (трипсин, химотрипсин, коллагеназа, эластаза, карбоксилаза) и кишечного сока (ами-нопептидаза, лейцинаминопептидаза, аланинаминопептидаза, трипептида-зы, дипептидазы, энтерокиназа).
Энтерокиназа вырабатывается клетками слизистой оболочки кишки в неактивной форме (киназоген), обеспечивает превращение неактивного фермента трип-синогена в активный трипсин. Пептидазы обеспечивают дальнейший последова-
тельный гидролиз пептидов, начавшийся в желудке, до свободных аминокислот, которые всасываются эпителиоцитами кишечника и поступают в кровь.
В переваривании углеводов также участвуют ферменты поджелудочной железы и кишечного сока: бета-амилаза, амило-1,6-глюкозидаза, олиго-1,6-глюкозидаза, мальтаза (альфа-глюкозидаза), лактаза, которые расщепляют полисахариды и дисахариды до простых сахаров (моносахаридов) - глюкозы, фруктозы, галактозы, всасываемых эпителиоцитами кишки и поступающих в кровь.
Переваривание жиров осуществляют панкреатические липазы, расщепляющие триглицериды, и кишечная липаза, обеспечивающая гидролитическое расщепление моноглицеридов. Продуктами расщепления жиров в кишечнике являются жирные кислоты, глицерин, моноглицериды.
В тонкой кишке происходит также процесс всасывания продуктов расщепления белков, жиров и углеводов в кровеносные и лимфатические сосуды. Кроме того, кишечник выполняет механическую функцию: проталкивает химус в каудальном направлении. Эта функция осуществляется благодаря перистальтическим сокращениям мышечной оболочки кишки. Эндокринная функция, выполняемая специальными секреторными клетками, заключается в выработке биологически активных веществ - серотони-на, гистамина, мотилина, секретина, энтероглюкагона, холецистокинина, панкреозимина, гастрина и ингибитора гастрина. Клетки, синтезирующие эти гормоны, относятся к дисперсной эндокринной системе (APUD-серии клеток) (см. главу 15).
Развитие. Тонкая кишка начинает развиваться на 5-й нед эмбриогенеза. Эпителий ворсинок, крипт и дуоденальные железы тонкой кишки образуются из кишечной энтодермы. На первых этапах дифференциров-ки эпителий однорядный кубический, затем он становится двухрядным, и, наконец, на 7-8-й нед возникает однослойный столбчатый эпителий. На 8-10-й нед развития появляются ворсинки и крипты. В течение 20- 24-й нед формируются циркулярные складки и появляются дуоденальные железы. Клетки кишечного эпителия у 4-недельного эмбриона не дифференцированы и характеризуются высокой пролиферативной активностью. Дифференцировка эпителиоцитов начинается на 6-12-й нед развития. Дивергентная дифференцировка стволовой клетки лежит в основе развития всех разновидностей эпителиоцитов и эндокриноцитов слизистой оболочки кишки. Появляются столбчатые (каемчатые) эпи-телиоциты, для которых характерно интенсивное развитие микроворсинок, увеличивающих резорбционную поверхность. Гликокаликс начинает формироваться к концу эмбрионального - началу плодного периода. В это время в эпителиоцитах отмечаются ультраструктурные признаки резорбции - большое число везикул, лизосом, мультивезикулярных и мекониальных телец.
Бокаловидные экзокриноциты дифференцируются на 5-й нед, эндо-криноциты - на 6-й нед развития. В это время среди эндокриноцитов преобладают переходные клетки с недифференцированными гранулами,
выявляются ЕС-клетки, G-клетки и S-клетки. В плодном периоде преобладают ЕС-клетки, большинство из которых не сообщается с просветом крипт («закрытый» тип); позднее появляется «открытый» тип клеток. Экзокриноциты с ацидофильными гранулами (клетки Панета) малодиффе-ренцированы у эмбрионов и плодов человека. Собственная пластинка слизистой оболочки и подслизистая основа тонкой кишки образуются из мезенхимы на 7-8-й нед эмбриогенеза. Гладкая мышечная ткань в стенке тонкой кишки развивается из мезенхимы неодновременно в различных участках кишечной стенки: на 7-8-й нед появляется внутренний циркулярный слой мышечной оболочки, затем на 8-9-й нед - наружный продольный слой, и, наконец, на 24-28-й нед развития плода возникает мышечная пластинка слизистой оболочки. Серозная оболочка тонкого кишечника закладывается на 5-й нед эмбриогенеза из мезенхимы (ее соединительнотканная часть) и висцерального листка спланхнотома мезодермы (ее мезотелий).
Строение. Стенка тонкой кишки построена из слизистой оболочки, подслизистой основы, мышечной и серозной оболочек (рис. 16.23).
Внутренняя поверхность тонкой кишки имеет характерный рельеф благодаря наличию ряда образований - циркулярных складок, ворсинок и крипт (кишечные железы). Эти структуры увеличивают общую поверхность тонкой кишки, что способствует выполнению основных функций пищеварения. Кишечные ворсинки и крипты являются основными структурно-функциональными единицами слизистой оболочки тонкой кишки.
Циркулярные складки (plicae circulares) образованы слизистой оболочкой и подслизистой основой.
Кишечные ворсинки (villi intestinales) представляют собой выпячивания слизистой оболочки пальцевидной или листовидной формы, свободно вдающиеся в просвет тонкой кишки.
У новорожденных и в раннем постнатальном периоде форма ворсинок пальцевидная, а у взрослых уплощенная - листовидная (или языковидная). Ворсинки уплощенной формы имеют две поверхности - краниальную и каудальную и два края (гребни).
Число ворсинок в тонкой кишке очень велико. Больше всего их в двенадцатиперстной и тощей кишке (22-40 ворсинок на 1 мм2), несколько меньше - в подвздошной кишке (18-31 ворсинка на 1 мм2). В двенадцатиперстной кишке ворсинки широкие и короткие (высота их 0,2-0,5 мм), в тощей и подвздошной кишке они несколько тоньше, но выше (до 0,5-1,5 мм). В образовании каждой ворсинки участвуют структурные элементы всех слоев слизистой оболочки.
Кишечные крипты (железы) (cryptae seu glandulae intestinales) представляют собой углубления эпителия в виде многочисленных трубочек, лежащих в собственной пластинке слизистой оболочки (см. рис. 16.23, а, в). Их устья открываются в просвет между ворсинками. На 1 мм2 поверхности кишки приходится до 100 крипт, а всего в тонкой кишке более 150 млн крипт. Каждая крипта имеет длину около 0,25-0,5 мм, диаметр до 0,07 мм. Общая площадь крипт в тонкой кишке составляет около 14 м2.
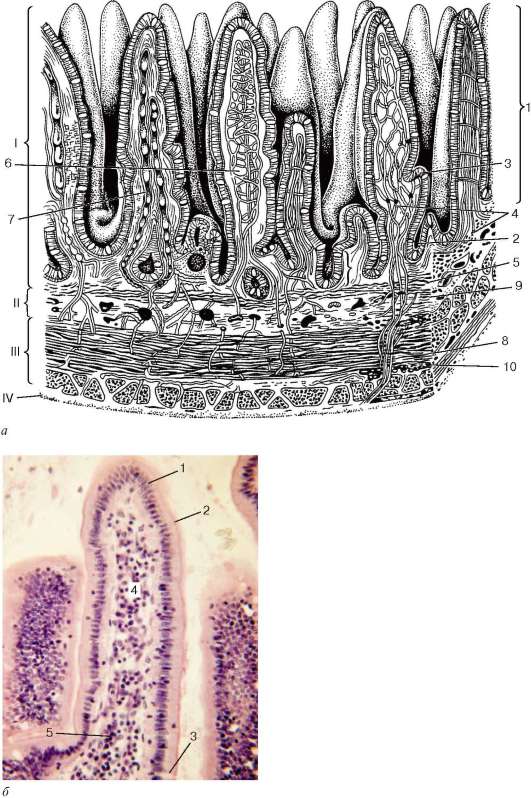
Рис. 16.23. Строение кишки: а - общий план строения (по Крелингу и Крау): I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная оболочка; 1 - кишечные ворсинки; 2 - кишечные железы (крипты); 3 - эпителий; 4 - собственная пластинка слизистой оболочки; 5 - мышечная пластинка слизистой оболочки; 6 - сосудистая сеть; 7 - лимфатическая сеть; 8 - нервное волокно;
9 - подслизистое нервное сплетение;
10 - мышечно-кишечное нервное сплетение; б - продольный срез ворсинки (микрофотография): 1 - однослойный столбчатый эпителий; 2 - микроворсинчатая (щеточная) каемка; 3 - бокаловидный экзокриноцит; 4 - собственная пластинка слизистой оболочки; 5 - лимфоциты
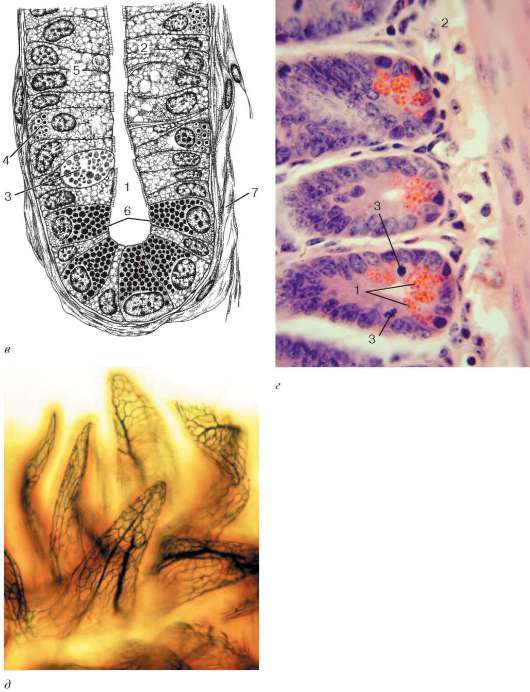
Рис. 16.23. Продолжение в - строение железы (крипты) тонкой кишки (по Н. А. Юриной, Л. С. Румянцевой); 1 - просвет кишки; 2 - столбчатые эпителиоциты; 3 - бокаловидный экзокриноцит; 4 - аргирофильные клетки (эндокриноциты); 5 - мало-дифференцированные эпителиоциты; 6 - экзокриноциты с ацидофильными гранулами; 7 - соединительная ткань собственной пластинки слизистой оболочки; г - крипта подвздошной кишки (микрофотография); 1 - экзокриноциты с ацидофильными гранулами; 2 - собственная пластинка слизистой оболочки; 3 - митотические фигуры в эпителиоци-тах; д - сосуды ворсинки тонкой кишки
Слизистая оболочка тонкой кишки состоит из однослойного столбчатого каемчатого эпителия (epithelium simplex columnarum limbatum), собственной пластинки слизистой оболочки (lamina propria mucosae) и мышечной пластинки слизистой оболочки (lamina muscularis mucosae).
В составе эпителиального пласта тонкой кишки выделяют клеточные диффероны столбчатых эпителиоцитов (epitheliocyti columnares), бокаловидных
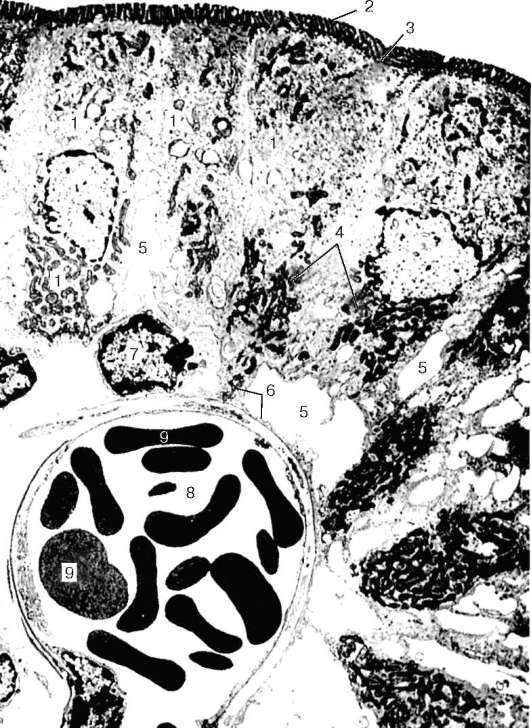
Рис. 16.24. Эпителиальная выстилка тонкой кишки. Электронная микрофотография, увеличение 4000 (по М. Россу, Л. Ромреллу, Г. Кайе):
1 - энтероциты; 2 - микроворсинки исчерченной каемки; 3 - соединяющий комплекс; 4 - митохондрии; 5 - просветы межклеточного пространства между латеральными поверхностями энтероцитов; 6 - базальные отростки энтероцитов; 7 - лимфоцит; 8 - капилляр; 9 - эритроциты
экзокриноцитов (exocrinocyti calciformes), клеток Панета, или экзокриноцитов с ацидофильными гранулами (exocrinocyti cum gzanulis acidophilis), эндокриноцитов (endocrinocyti), а также микроскладчатых эпителиоцитов (М-клеток). Источником развития перечисленных клеток являются стволовые клетки, находящиеся на дне крипт, которые путем асимметричного митоза и дивергентной дифференцировки через стадии клеток-предшественников
развиваются в конкретный вид эпителиоцитов. Клетки-предшественники также находятся в криптах, а в процессе дифференцировки перемещаются в направлении вершины ворсинки, где располагаются дифференцированные клетки, не способные к делению, которые здесь заканчивают жизненный цикл путем апоптоза или слущиваются из-за механического воздействия в процессе пищеварения. Весь цикл обновления (физиологическая регенерация) эпителиоцитов у человека составляет 5-6 сут.
В основе репаративной регенерации лежит аналогичный механизм, и дефект эпителия ликвидируется размножением клеток.
Кроме дифферонов эпителиоцитов, в эпителиальном пласте находятся производные гематогенного дифферона - гранулярные интраэпите-лиальные лимфоциты, располагающиеся в межклеточных пространствах и мигрирующие в собственную пластинку слизистой оболочки и далее в лимфокапилляры. Лимфоциты стимулируются антигенами, попадающими в просвет кишки, и играют важную роль в иммунной защите.
Строение кишечной ворсинки. С поверхности каждая кишечная ворсинка выстлана однослойным столбчатым эпителием (см. рис. 16.23, б). В эпителии различают три основных вида клеток: столбчатые эпителиоциты (и их разновидность - М-клетки), бокаловидные экзокриноциты, эндокриноциты.
Столбчатые эпителиоциты ворсинки (epitheliocyti columnares villi), или энте-роциты, составляют основную массу эпителиального пласта, покрывающего ворсинку. Клетки имеют высоту около 22-26 мкм и ширину около 8 мкм. Они характеризуются выраженной полярностью строения, что отражает их функциональную специализацию - обеспечение всасывания и транспорта веществ, поступающих с пищей.
На апикальной поверхности клеток имеется исчерченная каемка (limbus striatus), образованная множеством микроворсинок (рис. 16.24). Количество микроворсинок на 1 мкм2 поверхности клетки составляет от 60 до 90. Высота каждой микроворсинки у человека около 0,9-1,25 мкм, диаметр - 0,08-0,11 мкм, промежутки между микроворсинками равны 0,01-0,02 мкм. Благодаря огромному числу микроворсинок поверхность всасывания кишки увеличивается в 30-40 раз. На поверхности микроворсинок расположен гликокаликс, представленный липопротеидами и гликопротеинами
(рис. 16.25).
В плазмолемме и гликокаликсе микроворсинок исчерченной каемки обнаружено высокое содержание ферментов, участвующих в расщеплении и транспорте всасывающихся веществ: фосфатазы, нуклеозиддифосфатазы, L-, D-гликозидазы, аминопептидазы и др. Содержание фосфатаз в эпителии тонкой кишки превышает их уровень в печени почти в 700 раз, причем 3/4 их количества находится в каемке. Расщепление пищевых веществ и всасывание их наиболее интенсивно происходят в области исчерченной каемки. Эти процессы получили название пристеночного и мембранного пищеварения в отличие от полостного, совершающегося в просвете кишечной трубки, и внутриклеточного (см. ниже).
В апикальной части энтероцита находится терминальная сеть актиновых и миозиновых микрофиламентов, связанная с межклеточными контакта-
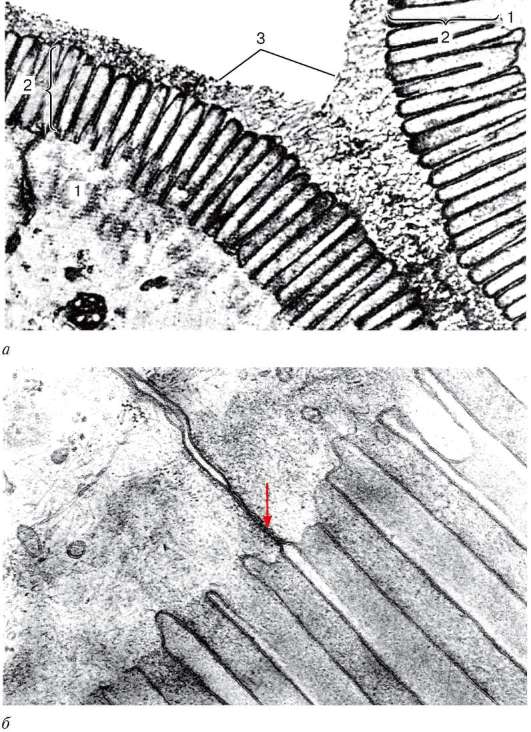
Рис. 16.25. Гликокаликс и микроворсинки исчерченной каемки эпителиоцитов слизистой оболочки тонкой кишки:
а - кишечные ворсинки кошки, электронная микрофотография, увеличение 24 000 (по С. Ито): 1 - апикальные части двух столбчатых эпителиоцитов; 2 - микроворсинки исчерченной каемки; 3 - гликокаликс; б - электронная микрофотография плотного контакта (стрелка) между двумя энтероцитами эпителия тощей кишки кролика, увеличение 50 000 (по В. А. Шахламову)
ми на боковых поверхностях апикальных частей энтероцитов. Различают два типа межклеточных контактов (соединений) - плотные изолирующие контакты (zonula occludens) и адгезивные пояски (zonula adherens), соединяющие соседние клетки и закрывающие сообщение между просветом кишки и межклеточным пространством.
При участии микрофиламентов терминальной сети обеспечивается закрытие межклеточных щелей между энтероцитами, что предотвращает поступление в щели различных веществ в процессе пищеварения. Под терминальной сетью в апикальной части энтероцита расположены трубочки и цистерны гладкой эндоплазматической сети, участвующие в процессах всасывания жиров, а также митохондрии, обеспечивающие энергией процессы всасывания и транспорта метаболитов.
В базальной части столбчатого эпителиоцита находятся ядро овальной формы, синтетический аппарат - рибосомы и гранулярная эндоплазма-тическая сеть. Комплекс Гольджи расположен над ядром, при этом его цистерны лежат вертикально по отношению к поверхности энтероцита. Формирующиеся в области комплекса Гольджи лизосомы и секреторные везикулы перемещаются в апикальную часть клетки и локализуются непосредственно под терминальной сетью и вдоль латеральной плазмолеммы.
Характерно наличие между базальными частями энтероцитов широких межклеточных пространств, ограниченных их латеральными плазмолемма-ми. На латеральных плазмолеммах имеются складки и отростки, которые соединяются с отростками соседних клеток. При активном всасывании жидкости складки расправляются и объем межклеточного пространства увеличивается. В базальных частях энтероцитов имеются тонкие латеральные базальные отростки, контактирующие с подобными отростками соседних клеток и лежащие на базальной мембране. Базальные отростки соединены простыми контактами и обеспечивают закрытие межклеточного пространства между энтероцитами. Наличие межклеточных пространств такого типа характерно для эпителиев, участвующих в транспорте жидкости; при этом эпителий функционирует как селективный барьер.
В латеральной плазмолемме энтероцита локализуются энзимы транспорта ионов, играющие важную роль в переносе метаболитов от апикальной плазмолеммы к латеральной и в межклеточное пространство, а далее через базальную мембрану в l. propria и капилляры.
Энтероциты выполняют также секреторную функцию, продуцируя метаболиты и ферменты, необходимые для терминального пищеварения (пристеночного и мембранного). Синтез секреторных продуктов происходит в гранулярной эндоплазматической сети, а образование секреторных гранул - в комплексе Гольджи, откуда секреторные везикулы, содержащие гликопротеины, транспортируются к поверхности клетки и локализуются под терминальной сетью микрофиламентов и вдоль латеральной плазмо-леммы.
М-клетки (клетки с микроскладками) являются разновидностью энтеро-цитов. Они располагаются на прерывистой базальной мембране на поверхности групповых лимфоидных узелков (пейеровых бляшек) и одиночных лимфоидных узелков. Клетки имеют уплощенную форму, малое число микроворсинок и получили свое название в связи с наличием на их апикальной поверхности микроскладок. С помощью микроскладок они способны захватывать макромолекулы из просвета кишки и формировать эндоци-
тозные везикулы, которые транспортируются к антигенпредставляющим клеткам и лимфоцитам. Последние мигрируют в лимфоидные органы, что стимулирует развитие иммунного ответа на присутствие антигенов.
Бокаловидные экзокриноциты (exocrinocyti caliciformes) в ворсинках расположены поодиночке среди столбчатых клеток. Число их увеличивается по направлению от двенадцатиперстной кишки к подвздошной. По своему строению это типичные слизистые клетки. В них наблюдаются циклические изменения, связанные с накоплением и последующим выделением слизи. В фазе накопления секрета ядра этих клеток оказываются прижатыми к их основанию, в цитоплазме же клеток над ядром видны капли слизи. Комплекс Гольджи и митохондрии располагаются около ядра. Формирование секрета происходит в области комплекса Гольджи. В стадии накопления слизи в клетке обнаруживается большое число измененных митохондрий. Они крупные, светлые, с короткими кристами. После выделения секрета бокаловидная клетка становится узкой, ядро ее уменьшается, цитоплазма освобождается от гранул секрета. Слизь, выделяемая бокаловидными экзокриноцитами, служит для увлажнения поверхности слизистой оболочки кишечника и этим способствует продвижению пищевых частиц, а также участвует в процессах пристеночного пищеварения. Под эпителием ворсинки находится базальная мембрана, за которой следует рыхлая соединительная ткань собственной пластинки слизистой оболочки. В ней проходят кровеносные и лимфатические сосуды и нервы, ориентированные вдоль ворсинки. В строме ворсинки всегда присутствуют отдельные гладкие мышечные клетки - производные мышечной пластинки слизистой оболочки. Пучки гладких миоцитов обвиты сетью ретикулярных волокон, которые связывают их со стромой ворсинки и базальной мембраной. Сокращение миоцитов способствует проталкиванию всасывающихся продуктов гидролиза пищи в кровь и лимфу ворсин кишечника. Другие пучки гладких мышечных клеток, проникающие в подслизистую основу, образуют циркулярные слои вокруг проходящих там сосудов. Сокращение этих мышечных групп регулирует кровоснабжение органа.
Строение кишечной крипты. Эпителиальная выстилка кишечных крипт содержит стволовые клетки, клетки-предшественники столбчатых эпителио-цитов, бокаловидных экзокриноцитов, эндокриноцитов и клеток Панета на всех стадиях развития.
Столбчатые эпителиоциты составляют основную массу эпителиальной выстилки крипт. По сравнению с аналогичными клетками ворсинок они ниже, имеют более тонкую исчерченную каемку и базофильную цитоплазму. В эпителиоцитах нижней половины крипт часто видны фигуры митоза. Эти элементы служат источником регенерации как для эпителиальных клеток ворсинок, так и для клеток крипт. Бокаловидные экзокри-ноциты постоянно находятся в криптах. По строению они не отличаются от таковых в составе ворсинок. Экзокриноциты с ацидофильными гранулами (exocrinocyti cum granulis acidophilis), или клетки Панета, располагаются группами или поодиночке на дне крипт (см. рис. 16.23, в, г). В их апикальной части видны плотные сильно преломляющие свет гранулы. Эти гранулы
резко ацидофильны, окрашиваются эозином в ярко-красный цвет, растворяются в кислотах, но устойчивы к щелочам. Цитохимически в гранулах обнаружены белково-полисахаридный комплекс, ферменты, лизоцим. Цитоплазма базальной части клетки окрашивается базофильно. Вокруг большого округлого ядра располагается немного митохондрий, над ядром находится комплекс Гольджи. Ацидофилия гранул обусловлена наличием богатого аргинином белка. В клетках Панета выявлено большое количество цинка, а также ферментов - кислой фосфатазы, дегидрогеназ и дипептидаз. Наличие в этих клетках ряда ферментов указывает на участие их секрета в процессах пищеварения - расщеплении дипептидов до аминокислот. Не менее важной является антибактериальная функция секрета, связанная с выработкой лизоцима, который разрушает клеточные стенки бактерий и простейших. Таким образом, клетки Панета играют важную роль в регуляции бактериальной флоры тонкой кишки.
Эндокриноцитов в крипте значительно больше, чем в составе эпителия ворсинки. Наиболее многочисленными являются ЕС-клетки, секретирую-щие серотонин, мотилин и вещество Р. А-клетки, продуцирующие энтеро-глюкагон, малочисленны. S-клетки, вырабатывающие секретин, распределены в разных отделах кишечника нерегулярно. Кроме того, в кишечнике найдены I-клетки, секретирующие холецистокинин и панкреозимин - биологически активные вещества, оказывающие стимулирующее действие на функции поджелудочной железы и печени. Обнаружены также G-клетки, вырабатывающие гастрин, D- и D1-клетки, продуцирующие активные пептиды (соматостатин и вазоактивный интестинальный пептид - ВИП) (см. главу 15 и табл. 16.1).
Для собственной пластинки слизистой оболочки (см. рис. 16. 23, а, б) характерно содержание большого количества ретикулярных волокон. Они образуют густую сеть и, подходя к эпителию, участвуют в образовании базальной мембраны. С ретикулярными волокнами тесно связаны отрост-чатые клетки, сходные по строению с ретикулярными клетками кроветворных органов. В собственной пластинке, так же как и в подслизистой основе, постоянно встречаются макрофаги, лимфоциты, а также плазматические клетки, которые относятся к кишечно-ассоциированной лимфоид-ной ткани. В ней расположены сосудистые и нервное сплетения.
Мышечная пластинка слизистой оболочки состоит из двух слоев: внутреннего циркулярного и наружного (более рыхлого) - продольного (см. рис. 16.23, а). Толщина обоих слоев около 40 мкм. В них имеются и косо идущие пучки мышечных клеток. От внутреннего циркулярного мышечного слоя отдельные мышечные клетки отходят в собственную пластинку слизистой оболочки.
Подслизистая основа нередко содержит дольки жировой ткани. В ней располагаются сосуды и подслизистое нервное сплетение.
Мышечная оболочка тонкой кишки состоит из двух слоев: внутреннего - циркулярного (более мощного) и наружного - продольного (см. рис. 16.23, а). Направление хода пучков мышечных клеток в обоих слоях не строго циркулярное и продольное, а спиральное. В наружном слое завитки спи-
рали более растянуты по сравнению с внутренним слоем. Между обоими мышечными слоями располагается прослойка рыхлой волокнистой соединительной ткани, в которой находятся узлы мышечно-кишечного нервного сплетения и сосуды.
Функция мышечной оболочки заключается в перемешивании и проталкивании химуса по ходу кишечника. В тонкой кишке различают сокращения двух видов. Сокращения местного характера обусловлены главным образом сокращениями внутреннего слоя мышечной оболочки. Они совершаются ритмически - 12-13 раз в минуту. Другие сокращения - перистальтические - вызываются действием мышечных элементов обоих слоев и распространяются последовательно по всей длине кишки. Перистальтические сокращения прекращаются после разрушения мышечно-кишечного нервного сплетения. Усиление перистальтики тонкой кишки происходит при возбуждении симпатических нервов, ослабление - при возбуждении блуждающего нерва.
Серозная оболочка покрывает снаружи тонкую кишку со всех сторон, за исключением двенадцатиперстной кишки, которая покрыта брюшиной только спереди, а в остальных частях имеет соединительнотканную оболочку.
Двенадцатиперстная кишка
Особенности строения двенадцатиперстной кишки определяются главным образом наличием дуоденальных желез (glandulae submucosa duodenalis) в подслизистой основе (рис. 16.26). В этот отдел тонкой кишки открываются протоки двух крупных желез - печени и поджелудочной железы. Химус из желудка поступает в двенадцатиперстную кишку и подвергается дальнейшей обработке ферментами кишечного и панкреатического соков и желчных кислот. Здесь же начинаются активные процессы всасывания.
Подслизистые дуоденальные железы (бруннеровы железы). В филогенезе железы появляются у млекопитающих животных, что обусловлено интенсификацией процессов пищеварения в связи с увеличением энергозатрат организма. В эмбриогенезе у млекопитающих и человека дуоденальные железы закладываются и дифференцируются позже других желез - поджелудочной железы, печени, желез желудка. Различия в строении и функции желез связаны с характером питания животных (растительноядные, плотоядные, всеядные). У человека дуоденальные железы закладываются на 20-22-й нед внутриутробного развития. Они расположены в подслизистой основе по всей длине двенадцатиперстной кишки (см. рис. 16.26, а, б).
Дуоденальные железы у всех млекопитающих и человека альвеолярно-трубчатые, разветвленные. Их выводные протоки открываются в крипты либо у основания ворсинок непосредственно в полость кишки. Ведущий клеточный дифферон желез - слизистые клетки (гландулоциты) с характерными гранулами секрета (см. рис. 16.26, б). Кроме них обнаруживаются клетки Панета, бокаловидные клетки, эндокриноциты разных видов (Ес, G, S, D). Камбиальные элементы расположены в устье протоков, поэтому обновление клеток желез идет от протоков в направлении концевых отделов.
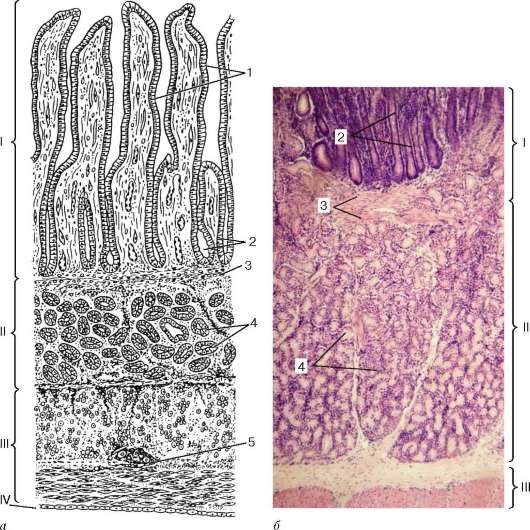
Рис. 16.26. Строение двенадцатиперстной кишки:
а - схема (продольный разрез); б - микрофотография. I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная оболочка. 1 - кишечные ворсинки; 2 - кишечные железы (крипты); 3 - мышечная пластинка слизистой оболочки; 4 - подслизистые (дуоденальные) железы; 5 - мышечно-кишечное нервное сплетение
Секрет гландулоцитов имеет щелочную реакцию, богат нейтральными гликопротеидами с присутствующими в них терминальными дисахарида-ми, в которых галактоза связана с остатками галактозамина или гликоза-мина. В гландулоцитах постоянно отмечаются одновременно синтез, накопление гранул и выделение секрета.
В фазе покоя (вне приема пищи) в гландулоцитах имеют место незначительно выраженные процессы синтеза и экзоцитоза секреторных гранул. При приеме пищи отмечаются усиление секреции путем экзоцитоза гранул, апокринии и даже выделение секрета путем диффузии. Асинхронность
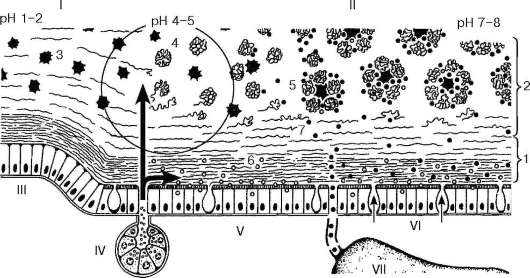
Рис. 16.27. Участие дуоденальных желез в процессах пищеварения в двенадцатиперстной кишке (по А. Н. Яцковскому):
I - желудок; II - двенадцатиперстная кишка; III - однослойный столбчатый эпителий пилорического отдела желудка; IV - дуоденальная железа; V - однослойный столбчатый эпителий ворсинок; VI - бокаловидные экзокриноциты; VII - поджелудочная железа. 1 - пристеночный слой слизи; 2 - содержимое полости кишки (смесь кишечного сока и секрета дуоденальных желез); 3 - химус, поступающий из желудка; 4 - образующиеся флоккулы; 5 - взаимодействие флоккул с частицами химуса; 6 - эндогидролазы; 7 - экзогидролазы
работы отдельных гландулоцитов и различных концевых отделов обеспечивает непрерывность функционирования дуоденальных желез. Их участие в пищеварении отражено на рис. 16.27.
Секрет дуоденальных желез выполняет две основные функции: 1) пищеварительную - участвует в пространственной и структурной организации процессов гидролиза и всасывания; 2) защитную - предохраняет стенку кишечника от механических и химических повреждений.
Секрет дуоденальных желез, соединяясь с пристеночным слоем слизи, придает ему большую вязкость и устойчивость к разрушению. Смешиваясь с дуоденальным кишечным соком, секрет этих желез способствует образованию частиц геля - флоккул, формирующихся при снижении рН в двенадцатиперстной кишке в связи с поступлением закисленного химуса из желудка. Эти флоккулы значительно увеличивают адсорбционные свойства кишечного сока для ферментов, что повышает активность последних. Например, адсорбция и активность фермента трипсина в структурах плотной фазы кишечного сока (после добавления к нему секрета дуоденальных желез) увеличиваются более чем в 2 раза.
Таким образом, секрет дуоденальных желез обладает максимальной способностью к флоккулообразованию (при определенных значениях рН), стимулирует структурирование дуоденального сока и повышает его сорбци-онные свойства. Отсутствие секрета дуоденальных желез в составе химуса
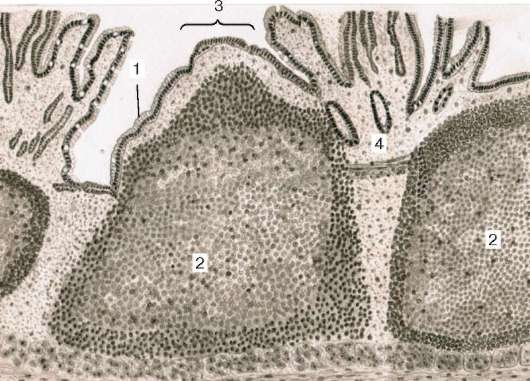
Рис. 16.28. Строение слизистой оболочки подвздошной кишки (схема):
1 - однослойный столбчатый эпителий; 2 - лимфоидные узелки; 3 - выпячивание
слизистой оболочки (купол); 4 - собственная пластинка слизистой оболочки
и пристеночной слизи меняет их физико-химические свойства, в результате чего снижаются сорбционная емкость для эндо- и экзогидролаз и их активность.
Кишечно-ассоциированная лимфоидная ткань. Лимфоидная ткань широко распространена в тонкой кишке в виде лимфоидных узелков и диффузных скоплений в собственной пластинке слизисой оболочки и выполняет защитную функцию.
Одиночные (солитарные) лимфоидные узелки (noduli lymphatici solitarii) встречаются на всем протяжении тонкой кишки. Диаметр их около 0,5-3 мм. Более крупные узелки, лежащие в дистальных отделах тонкой кишки, проникают в мышечную пластинку ее слизистой оболочки и располагаются частично в подслизистой основе (рис. 16.28). Количество одиночных лимфоидных узелков в стенке тонкой кишки детей от 3 до 13 лет составляет около 15 000. По мере старения организма количество их уменьшается.
Сгруппированные лимфоидные узелки (noduli lymphatici aggregati), или пейеро-вы бляшки, как правило, располагаются в подвздошной кишке, но иногда встречаются в тощей и двенадцатиперстной кишке. Число узелков варьирует в зависимости от возраста: в тонкой кишке у детей около 100, у взрослых - около 30-40, а в старческом возрасте их количество значительно уменьшается.
Длина одного сгруппированного лимфоидного узелка может составлять от 2 до 12 см, а ширина - около 1 см. Наиболее крупные из них проникают в подслизистую основу. Ворсинки в слизистой оболочке в местах расположения сгруппированных лимфоидных узелков, как правило, отсутствуют.
Рис. 16.29. Механизм транспорта IgA через эпителиоциты кишечника: I - просвет кишки; II - столбчатые эпителиоциты; III - собственная пластинка слизистой оболочки. 1 - плазмоциты; 2 - димеры IgA; 3 - секреторный компонент, связанный с базальной плазмолеммой (трансмембранный гликопротеин); 4 - комплекс IgA с секреторным компонентом; 5 - эндоцитозная вакуоль; 6 - трансци-тозная вакуоль; 7 - экзоцитоз; 8 - секреторный компонент; 9 - «хвост» молекулы трансмембранного гликопротеина
Для эпителиальной выстилки, расположенной над узелками, характерно, как уже указывалось, наличие М-клеток, через которые транспортируются антигены, стимулирующие лимфоциты. Образующиеся плазмоциты секре-тируют иммуноглобулины (IgA, IgG, IgM), главным из которых является
IgA. На один плазмоцит, секретирующий IgG, приходится 20-30 плазмо-цитов, продуцирующих IgA, и 5 - продуцирующих IgM. IgA в отличие от других иммуноглобулинов более активен, так как не разрушается протео-литическими ферментами кишечника. Резистентность к кишечным про-теазам обусловлена соединением IgA с секреторным компонентом, образуемым эпителиоцитами. В эпителиоцитах синтезируется гликопротеин, который включается в их базальную плазмолемму (трансмембранный гли-копротеин) и служит Fc-рецептором для IgA (рис. 16.29). При соединении IgA с Fc-рецептором образуется комплекс, который с помощью эндоцитоза поступает в эпителиоцит и в составе трансцитозной везикулы переносится в апикальную часть клетки и выделяется в просвет кишки путем экзо-цитоза через апикальную плазмолемму. При выделении указанного комплекса в просвет кишки от него отщепляется только часть гликопротеина, непосредственно связанная с IgA и называемая секреторным компонентом. Остальная его часть («хвост» молекулы) остается в составе плазмолеммы. В просвете кишки IgA осуществляет защитную функцию, нейтрализуя антигены, токсины, микроорганизмы.
Васкуляризация. Артерии, входя в стенку тонкой кишки, образуют три сплетения: межмышечное - между внутренним и наружным слоями мышечной оболочки; широкопетлистое - в подслизистой основе и узкопетлистое - в слизистой оболочке. Из последнего выходят артериолы, образующие кровеносные капилляры вокруг кишечных крипт, и по одной-две артериолы, входящие в каждую ворсинку и распадающиеся там на капиллярные сети. Из кровеносных капилляров ворсинки кровь собирается в венулу, проходящую вдоль ее оси (см. рис. 16.23, а). Вены тонкой кишки образуют два сплетения - сплетение в слизистой оболочке и сплетение в подслизистой основе. Имеются многочисленные артериоловенулярные анастомозы типа замыкающих артерий, регулирующие приток крови к кишечным ворсинкам. Во время акта пищеварения анастомозы между артериями и венами закрыты, и вся масса крови устремляется в слизистую оболочку, к ее ворсинкам. В период голодания анастомозы открыты и основная масса крови проходит, минуя слизистую оболочку. Запирающие вены регулируют объем венозного оттока от тонкой кишки. В случае резкого переполнения эти вены могут депонировать значительные количества крови.
Лимфатические сосуды тонкой кишки представлены очень широко разветвленной сетью. В каждой кишечной ворсинке есть центрально расположенный, слепо оканчивающийся на ее вершине лимфатический капилляр. Просвет его шире, чем в кровеносных капиллярах. Из лимфатических капилляров ворсинок лимфа оттекает в лимфатическое сплетение слизистой оболочки, а из него в соответствующее сплетение подслизистой основы, образованное более крупными лимфатическими сосудами. В это сплетение вливается также густая сеть капилляров, оплетающих одиночные и групповые лимфоидные узелки. Из подслизистого сплетения отходят лимфатические сосуды, находящиеся между слоями мышечной оболочки.
Иннервация. Афферентная иннервация осуществляется мышечно-кишечным чувствительным сплетением (plexus myentericus sensibilis), образо-
Рис. 16.30. Типы пищеварения (по И. А. Морозову)
ванным чувствительными нервными волокнами спинальных ганглиев и их рецепторными окончаниями. Ветвистые и кустиковые нервные окончания часто встречаются в подслизистой основе и собственной пластинке слизистой оболочки. Их терминальные веточки достигают сосудов, дуоденальных желез, эпителия кишечных крипт и ворсинок. Обильные ветвления чувствительных волокон наблюдаются в подвздошной кишке и илеоцекальной области, где преобладают кустиковидные формы рецепторов. Отдельные рецепторы имеются в самих нервных ганглиях.
Эфферентная иннервация осуществляется симпатическими и парасимпатическими нервами. В толще стенки кишки хорошо развиты парасимпатические мышечно-кишечное и подслизистое нервные сплетения.
Мышечно-кишечное сплетение (plexus myentericus) наиболее развито в двенадцатиперстной кишке, где наблюдаются многочисленные, плотно расположенные крупные ганглии. Количество и размеры ганглиев в тонкой кишке уменьшаются в каудальном направлении. В ганглиях различают клетки I и II типа, причем клеток I типа значительно больше. Для тонкой кишки по сравнению с другими отделами пищеварительной трубки характерно наличие большого количества клеток II типа. Их особенно много в двенадцатиперстной кишке, в начальном отделе подвздошной кишки и в илеоцекальной области.
Гистофизиология процессов пищеварения и всасывания в тонкой кишке. Пищеварение в тонкой кишке включает два основных процесса: 1) дальнейшую ферментативную обработку веществ, содержащихся в химусе, до конечных продуктов и подготовку их к всасыванию; 2) всасывание - прохождение продуктов конечного расщепления пищи (мономеров) через эпителий, базальную мембрану, сосудистую стенку и поступление в кровь и лимфу.
Процессы пищеварения происходят в различных зонах кишки, в связи с чем различают внеклеточное пищеварение: полостное (в полости кишки), пристеночное (около стенки кишки), мембранное (на апикальных частях плазмолеммы энтероцитов и их гликокаликсе) - и внутриклеточное пищеварение (в цитоплазме энтероцитов) (рис. 16.30).
Внеклеточное пищеварение в полости кишки осуществляется за счет трех компонентов - ферментов пищеварительных желез (слюнных, поджелудочной), ферментов кишечной флоры и ферментов пищевых продуктов. Пристеночное пищеварение происходит в слизистых отложениях тонкой кишки, которые адсорбируют различные ферменты полостного пищеварения, а также ферменты, выделяемые энтероцитами. Мембранное пищеварение происходит на границе внеклеточной и внутриклеточной среды. На плазмолемме и гликокаликсе энтероцитов пищеварение осуществляется двумя группами ферментов. Первая группа ферментов образуется в поджелудочной железе (альфа-амилаза, липаза, трипсин, химотрипсин, кар-боксипептидаза). Они адсорбируются гликокаликсом и микроворсинками. Вторая группа - ферменты, имеющие кишечное происхождение, они связаны с плазмо-леммой энтероцитов.
Гликокаликс, помимо адсорбции ферментов, участвующих в пищеварении, играет роль фильтра, избирательно пропускающего лишь те вещества, для которых имеются адекватные ферменты. Кроме того, гликокаликс выполняет защитную функцию, обеспечивая изоляцию энтероцитов от бактерий и образованных ими токсических веществ. В гликокаликсе находятся рецепторы гормонов, антигенов, токсинов. Перенос веществ в цитоплазму энтероцита осуществляется путем диффузии, микропиноцитоза и с помощью мембранного транспорта белков (см. главу 4). Внутриклеточное пищеварение происходит внутри столбчатых эпите-лиоцитов с участием комплекса Гольджи и лизосом и включает депонирование, селекцию, транспортировку в лизосому, ресинтез (жиров, специфичных для данного вида), экзоцитоз. В результате последнего мономеры появляются в межклеточном пространстве и перемещаются в кровеносные и лимфатические капилляры (рис. 16.31).
Рис. 16.31. Процесс всасывания жира через эпителий тонкой кишки: а - столбчатый эпителиоцит после голодания животного; б - столбчатый эпите-лиоцит после приема жирной пищи (схемы по Карделю и другим, с изменениями); в - хиломикроны в лимфокапиллярах подслизистой основы двенадцатиперстной кишки крысы через 6 ч после приема пищи (по А. Н. Яцковскому). 1 - микроворсинки исчерченной каемки; 2 - гликокаликс; 3 - капли липидов; 4 - эндоплаз-матическая сеть; 5 - митохондрии; 6 - хиломикроны; 7 - комплекс Гольджи; 8 - межклеточное пространство; 9 - хиломикроны, поступающие через межклеточное пространство в лимфатический капилляр (10); 11 - хиломикроны в соединительной ткани; 12 - хиломикроны в лимфокапилляре
16.3.3. Толстая кишка
Толстая кишка выполняет важные функции - интенсивное всасывание воды из химуса и формирование каловых масс. Способность к всасыванию жидкостей используют во врачебной практике для введения больным питательных веществ с помощью клизм. В толстой кишке выделяется значительное количество слизи, которая облегчает продвижение содержимого по кишке и способствует склеиванию непереваренных частиц пищи. Одной из функций толстой кишки является выделительная. Через ее слизистую оболочку выделяется ряд веществ, например кальций, магний, фосфаты, соли тяжелых металлов и т. п. В толстой кишке вырабатываются витамин К и витамин В. Этот процесс осуществляется с участием бактериальной флоры, постоянно присутствующей в кишечнике. С помощью бактерий в толстой кишке происходит переваривание клетчатки.
Развитие. Эпителий ободочной кишки и тазовой части прямой кишки развивается из энтодермы. В кожной и промежуточной зонах анальной части прямой кишки эпителий имеет эктодермальное происхождение. Выражена граница между эпителием кишечного и кожного типа, расположенная между столбчатой и промежуточной зонами прямой кишки. Эпителий кишечной трубки сильно разрастается на 6-7-й нед внутриутробной жизни плода. Ворсинки и крипты в слизистой оболочке кишки закладываются почти одновременно. Позднее сюда врастает мезенхима, что ведет к сильному выпячиванию ворсинок в просвет кишки. На 4-м мес эмбрионального развития закладка толстой кишки содержит большое количество ворсинок. В дальнейшем усиленный рост поверхности слизистой оболочки приводит к растягиванию и сглаживанию этих ворсинок. К рождению ворсинки в толстой кишке исчезают.
Мышечная оболочка толстой кишки развивается на 3-м мес, а мышечная пластинка слизистой оболочки - на 4-м мес развития плода.
Ободочная кишка
Стенка ободочной кишки образована слизистой оболочкой, подслизи-стой основой, мышечной и серозной оболочками.
Для рельефа внутренней поверхности ободочной кишки характерно наличие большого количества циркулярных складок и кишечных крипт
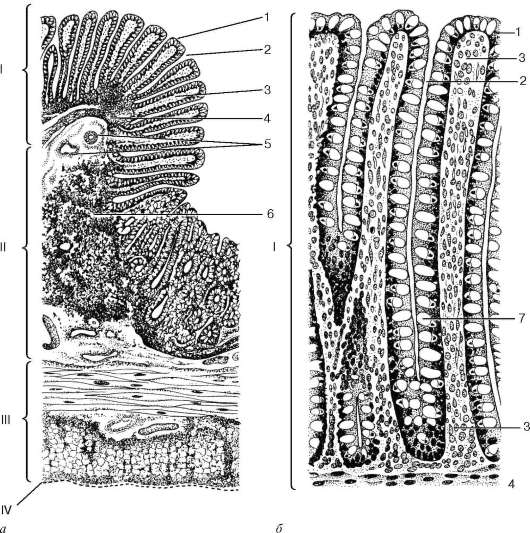
Рис. 16.32. Строение толстой кишки:
а - схема строения стенки толстой кишки; б - строение крипт. I - слизистая оболочка; II - подслизистая основа; III - мышечная оболочка; IV - серозная оболочка. 1 - энтероциты (колоноциты); 2 - кишечные железы (крипты); 3 - собственная пластинка слизистой оболочки; 4 - мышечная пластинка слизистой оболочки; 5 - кровеносные сосуды; 6 - одиночный лимфоидный узелок; 7 - бокаловидный экзокриноцит
(желез), значительно увеличивающих ее площадь. В отличие от тонкой кишки здесь отсутствуют ворсинки (рис. 16.32).
Циркулярные складки образуются на внутренней поверхности кишки из слизистой оболочки и подслизистой основы. Они располагаются поперек и имеют полулунную форму (отсюда название «полулунные складки»). Кишечные железы (крипты) в ободочной кишке развиты лучше, чем в тонкой, расположены чаще, размеры их больше (0,4-0,7 мм), они шире, содержат очень много бокаловидных экзокриноцитов.
Слизистая оболочка ободочной кишки, как и тонкой, включает эпителий, собственную пластинку (l. propria) и мышечную пластинку (l. muscularis mucosae).
Эпителий слизистой оболочки однослойный столбчатый. Он состоит из трех основных видов клеток: столбчатых эпителиоцитов, бокаловидных экзокриноцитов и желудочно-кишечных эндокриноцитов. Кроме того, имеются малодифференцированные эпителиоциты. Столбчатые эпителиоциты располагаются на поверхности слизистой оболочки и в ее криптах. По своему строению они сходны с аналогичными клетками тонкой кишки, но у них более тонкая исчерченная каемка (рис. 16.33).
Бокаловидные клетки, выделяющие слизь, имеются в большом количестве в криптах. Строение их было описано ранее. У основания кишечных крипт лежат малодифференцированные (камбиальные) эпителиоциты. В них часто видны фигуры митотического деления. За счет этих клеток происходит физиологическая регенерация эпителия толстой кишки. Изредка встречаются энтероэндокринные клетки (EC- и ECL-клетки) и клетки с ацидофильными гранулами (клетки Панета).
Собственная пластинка слизистой оболочки образует тонкие соединительнотканные прослойки между кишечными криптами. В этой пластинке часто встречаются одиночные лимфоидные узелки, из которых лимфоциты мигрируют в окружающую соединительную ткань и проникают в эпителий.
Мышечная пластинка слизистой оболочки более выражена, чем в тонкой кишке, и состоит из двух слоев. Внутренний слой более плотный, образован преимущественно циркулярно расположенными пучками гладких миоцитов. Наружный слой представлен пучками гладких миоцитов, ориентированных частично продольно, частично косо по отношению к оси кишки. Мышечные клетки в этом слое расположены более рыхло, чем во внутреннем.
Подслизистая основа содержит много жировых клеток. Здесь располагаются сосудистые, а также нервное подслизистое сплетения. В подслизистой основе ободочной кишки всегда очень много лимфоидных узелков; они распространяются сюда из собственной пластинки слизистой оболочки.
Мышечная оболочка представлена двумя слоями гладких мышц: внутренним - циркулярным и наружным - продольным. Наружный слой мышечной оболочки в ободочной кишке имеет особое строение. Этот слой не сплошной, и пучки гладких миоцитов в нем собраны в три ленты, тянущиеся вдоль всей ободочной кишки. В участках кишки, лежащих между лентами, обнаруживается лишь тонкий слой, состоящий из незначительного количества продольно расположенных пучков гладких миоцитов. Данные участки образуют вздутия, выбухающие наружу. Между двумя слоями мышечной оболочки есть прослойка рыхлой соединительной ткани, в которой проходят сосуды и находится мышечно-кишечное нервное сплетение.
Серозная оболочка покрывает ободочную кишку снаружи. Иногда она имеет пальцеобразные выросты. Эти выросты представляют собой скопления жировой ткани, покрытые брюшиной.
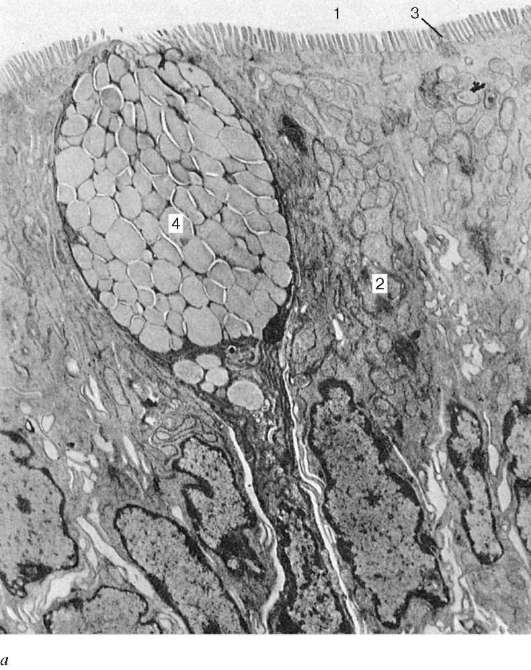
Рис. 16.33. Верхняя часть железы (крипты) толстой кишки собаки. Электронные микрофотографии:
а - по В. А. Шахламову; б - по К. А. Зуфарову. 1 - просвет кишечной железы (крипты); 2 - энтероциты (колоноциты); 3 - микроворсинки; 4 - бокаловидные экзокри-ноциты; 5 - лимфоцит
Червеобразный отросток
Для этого органа характерны большие скопления лимфоидной ткани. Червеобразный отросток имеет просвет (рис. 16.34) треугольной формы у детей и круглой - у взрослых. С годами этот просвет может исчезать, зарастая соединительной тканью.
Развитие. В развитии червеобразного отростка (аппендикса) плода человека можно выделить два основных периода. Первый период (8-12 нед) характеризуется отсутствием лимфоидных узелков, формированием однослойного столбчатого эпителия на поверхности и в криптах, появлением
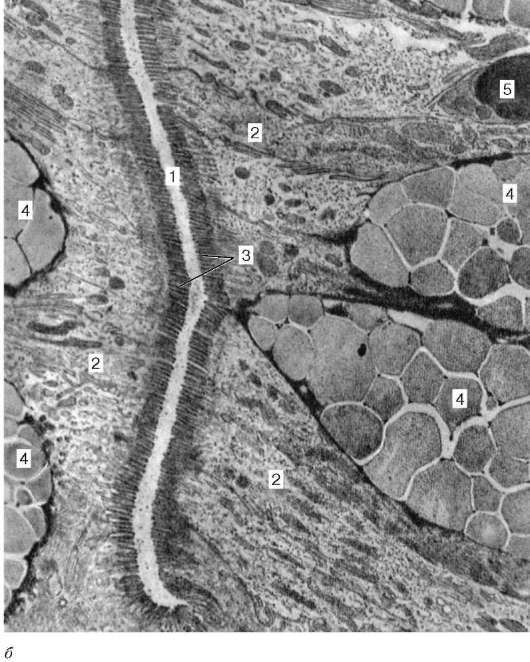
Рис. 16.33. Продолжение (обозначения см. выше)
эндокриноцитов и началом заселения лимфоцитами собственной пластинки слизистой оболочки. Для второго периода (17-31-я нед развития) характерны интенсивное развитие лимфоидной ткани и лимфоидных узелков без светлых центров, образование куполов над узелками. Эпителий, покрывающий купол, однослойный кубический, иногда плоский, инфильтрирован лимфоцитами. Вокруг зоны купола расположены высокие складки слизистой оболочки. На дне крипт дифференцируются экзокриноциты с ацидофильными гранулами. В процессе развития аппендикс заселяется как Т-лимфоцитами, так и В-лимфоцитами. Завершение основных морфогене-тических процессов отмечается к 40-й нед внутриутробного развития, когда число лимфоидных узелков в органе достигает 70, количество эндокриноцитов максимально (среди них преобладают ЕС- и S-клетки).
Слизистая оболочка червеобразного отростка имеет кишечные железы (крипты), покрытые однослойным столбчатым эпителием, в составе кото-
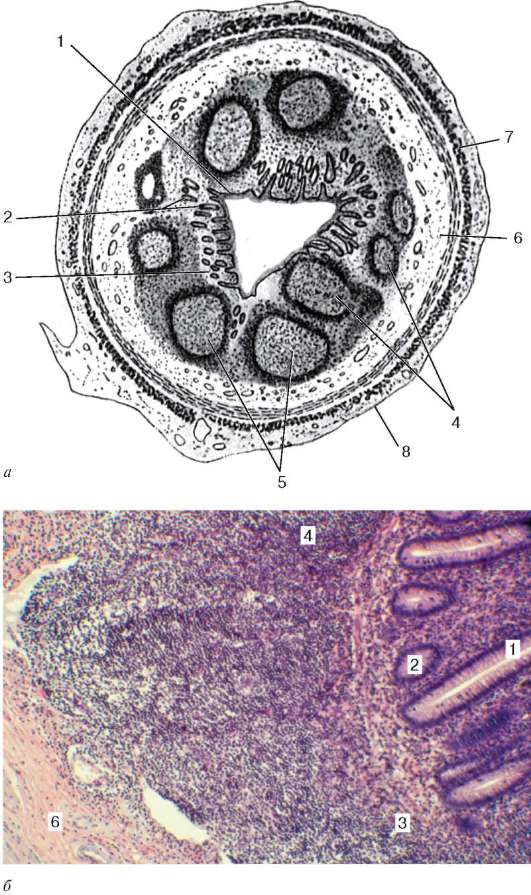
Рис. 16.34. Червеобразный отросток:
а - схема строения; б - микрофотография. 1 - эпителий; 2 - кишечные железы (крипты); 3 - собственная пластинка слизистой оболочки; 4 - лимфоидные узелки; 5 - герминативные центры; 6 - подслизистая основа; 7 - мышечная оболочка; 8 - серозная оболочка
рого встречаются бокаловидные клетки, М-клетки и энтероэндокриноциты (см. рис. 16.34). На дне кишечных крипт чаше, чем в других отделах толстой кишки, встречаются экзокриноциты с ацидофильными гранулами (клетки Панета). Здесь же располагаются недифференцированные (камбиальные) эпителиоциты и эндокринные клетки, причем их здесь больше, чем в криптах тонкой кишки (в среднем в каждой около пяти клеток). Собственная пластинка слизистой оболочки без резкой границы (вследствие слабого развития мышечной пластинки слизистой оболочки) переходит в подслизистую основу. В собственной пластинке и в подслизистой основе располагаются многочисленные крупные местами сливающиеся скопления лимфоидной ткани. При попадании инфекции в просвет отростка наступают изменения строения его стенки. В лимфоидных узелках возникают крупные светлые центры, лимфоциты инфильтрируют соединительную ткань собственной пластинки, и часть их проходит через эпителий в просвет червеобразного отростка. В этих случаях в просвете отростка можно видеть слущенные эпителиоциты и скопления погибших лимфоцитов. В подслизистой основе располагаются кровеносные сосуды и нервное подслизистое сплетение.
Мышечная оболочка имеет два слоя: внутренний - циркулярный и наружный - продольный. Продольный мышечный слой отростка сплошной в отличие от соответствующего слоя ободочной кишки. Снаружи отросток обычно покрыт серозной оболочкой, которая образует собственную брыжейку отростка.
Червеобразный отросток осуществляет защитную функцию, скопления лимфоидной ткани в нем являются составной частью кишечно-ассоциированной лимфоидной ткани.
Прямая кишка
Стенка прямой кишки состоит из тех же оболочек, что и ободочной кишки. В тазовой части прямой кишки ее слизистая оболочка имеет три поперечные складки, в образовании которых участвуют подслизистая основа и кольцевой слой мышечной оболочки. Ниже этих складок располагаются 8-10 продольных складок, между которыми видны углубления.
В анальной части кишки различают три зоны: столбчатую, промежуточную и кожную. В столбчатой зоне продольные складки образуют заднепроходные столбы. В промежуточной зоне эти образования соединяются, образуя зону слизистой оболочки с гладкой поверхностью в виде кольца шириной около 1 см (zona haemorrhoidalis).
Слизистая оболочка прямой кишки состоит из эпителия, собственной и мышечной пластинок. Эпителий в верхнем отделе прямой кишки однослойный столбчатый, в столбчатой зоне нижнего отдела - многослойный кубический, в промежуточной - многослойный плоский неороговевающий, в кожной - многослойный плоский ороговевающий. Переход от многослойного кубического эпителия к многослойному плоскому совершается резко в виде зигзагообразной - аноректальной линии (linea anorectalis).
Переход в эпителий кожного типа постепенный. В эпителии прямой кишки встречаются столбчатые эпителиоциты с микроворсинчатой
каемкой, бокаловидные экзокриноциты и эндокринные (ECL) клетки. Последних особенно много в столбчатой зоне. Эпителий в верхней части прямой кишки образует кишечные крипты. Они несколько длиннее, чем в ободочной кишке, но менее многочисленны. В нижних отделах кишки крипты постепенно исчезают.
Собственная пластинка слизистой оболочки принимает участие в формировании складок прямой кишки. Здесь располагаются одиночные лимфоид-ные узелки и сосуды. В области столбчатой зоны в этой пластинке залегает сеть тонкостенных кровеносных лакун, кровь из которых оттекает в геморроидальные вены. В собственной пластинке этой области проходят протоки желез, расположенных в подслизистой основе.
В промежуточной зоне собственная пластинка содержит большое количество эластических волокон, а также лимфоцитов и тучных клеток. Здесь находятся и единичные сальные железы.
В кожной зоне, окружающей анальное отверстие, к сальным железам присоединяются волосы. Потовые железы в собственной пластинке слизистой оболочки появляются на расстоянии 1-1,5 см от ануса, представляют собой трубчатые железы, концевые отделы которых свернуты кольцом (gll. circumanales). Это железы апокринного типа секреции.
Мышечная пластинка слизистой оболочки, как и в других отделах толстой кишки, состоит из двух слоев. Пучки гладких миоцитов ее постепенно переходят в узкие продольные пучки, тянущиеся до столбчатой зоны.
В подслизистой основе располагаются сосудистые и нервные сплетения. Здесь же встречаются пластинчатые нервные тельца. В подслизистой основе лежит сплетение геморроидальных вен (рис. 16.35). При нарушении тонуса стенок этих сосудов появляются варикозные расширения. При патологических изменениях эти образования могут быть причиной кровотечений. В подслизистой основе столбчатой зоны прямой кишки имеется 6-8 разветвленных трубчатых образований, тянущихся до циркулярного слоя мышечной оболочки, прободающих его и слепо заканчивающихся в межмышечной соединительной ткани. На концах их образуются ампулярные расширения, которые выстланы одним или двумя слоями кубических клеток. Эпителий главных протоков этих рудиментарных анальных желез (gll. anales) состоит из нескольких слоев полигональных клеток. Устье протока выстлано многослойным плоским эпителием. Эти эпителиальные трубочки расценивают как гомологи анальных желез животных. У человека при патологических условиях они могут служить местом образования фистул.
Мышечная оболочка состоит из двух слоев: внутреннего - циркулярного и наружного - продольного. Циркулярный слой на разных уровнях прямой кишки образует два утолщения, которые выделяются как отдельные анатомические образования - сфинктеры внутренний и наружный (m. sphincter ani internus et externus). Мышца наружного сфинктера образована поперечнополосатой мышечной тканью. Наружный продольный слой мышечной оболочки прямой кишки в противоположность другим отделам толстой кишки сплошной. Между обоими мышечными слоями имеется прослойка рыхлой соединительной ткани, в которой лежат мышечно-кишечные сплетения и сосуды.
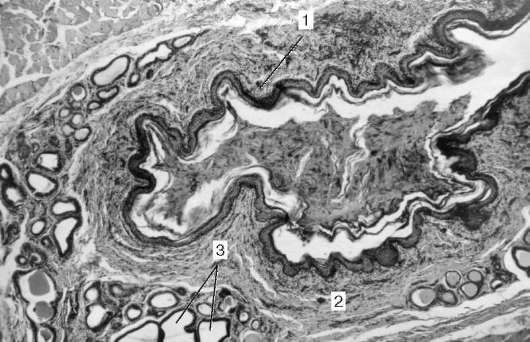
Рис. 16.35. Строение стенки прямой кишки человека (микропрепарат П. И. Гала-ховой):
1 - многослойный плоский ороговевающий эпителий; 2 - собственная пластинка слизистой оболочки; 3 - геморроидальные вены
Серозная оболочка покрывает прямую кишку в верхней ее части; в нижних отделах прямая кишка имеет соединительнотканную оболочку.
Иннервация. В парасимпатическом мышечно-кишечном нервном сплетении толстой кишки, начиная с проксимальных отделов, моторные нейроны I типа постепенно заменяются чувствительными нейронами II типа, которые в прямой кишке становятся преобладающими.
Афферентная иннервация в прямой кишке хорошо развита. В толстой кишке афферентные волокна образуют в мышечной оболочке чувствительное сплетение. Чувствительные окончания имеют вид кустиков и термина-лей, оканчивающихся в гладких мышцах.
16.4. ПЕЧЕНЬ
Печень (hepar) - самая крупная железа пищеварительного тракта. Функции печени чрезвычайно разнообразны. В ней обезвреживаются многие продукты обмена веществ, инактивируются гормоны, биогенные амины, а также ряд лекарственных препаратов. Печень участвует в защитных реакциях организма против микробов и чужеродных веществ в случае проникновения их извне. В ней образуется гликоген - главный источник поддержания постоянной концентрации глюкозы в крови. В печени синтезируются важнейшие белки плазмы крови: фибриноген, альбумины, протромбин и др. Здесь метаболизируется железо и образуется желчь, необходимая для всасывания жиров в кишечнике. Большую роль она играет в обмене холестерина, который является важным компонентом клеточных мембран. В печени накапливаются необходимые
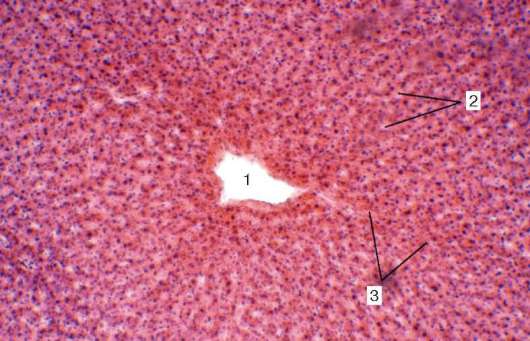
Рис. 16.36. Печень человека:
1 - центральная вена; 2 - синусоидальные капилляры; 3 - печеночные балки
для организма жирорастворимые витамины - A, D, Е, К и др. Кроме того, в эмбриональном периоде печень является органом кроветворения. Столь многочисленные и важные функции печени определяют ее значение для организма как жизненно необходимого органа.
Развитие. Зачаток печени образуется из энтодермы в конце 3-й нед эмбриогенеза и имеет вид мешковидного выпячивания вентральной стенки туловищной кишки (печеночная бухта). В процессе роста печеночная бухта подразделяется на верхний (краниальный) и нижний (каудальный) отделы. Краниальный отдел служит источником развития печени и печеночного протока, каудальный - желчного пузыря и желчного протока. Устье печеночной бухты, в которое впадают краниальный и каудальный отделы, образует общий желчный проток. В гистогенезе происходит дивергентная дифференцировка стволовых клеток в составе краниального отдела печеночной бухты, в результате которой возникают диффероны эпителиоцитов печени (гепатоцитов) и эпителиоцитов желчных протоков (холангиоцитов). Эпителиальные клетки краниального отдела печеночной бухты быстро разрастаются в мезенхиме брыжейки, формируя многочисленные тяжи. Между эпителиальными тяжами располагается сеть широких кровеносных капилляров, происходящих из желточной вены, которая в процессе развития дает начало воротной вене.
Сложившаяся таким путем железистая паренхима печени своим строением напоминает губку. Дальнейшая дифференцировка печени происходит во второй половине внутриутробного периода развития и в первые годы после рождения. При этом по ходу ветвей воротной вены внутрь печени врастает соединительная ткань, разделяя ее на печеночные дольки.
Строение. Поверхность печени покрыта соединительнотканной капсулой, которая плотно срастается с висцеральным листком брюшины. Паренхима
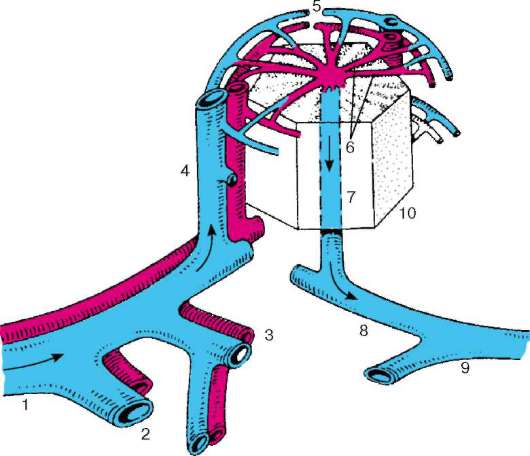
Рис. 16.37. Кровеносная система печени (по Е. Ф. Котовскому):
1 - воротная вена и печеночная артерия; 2 - долевая вена и артерия; 3 - сегментарная вена и артерия; 4 - междольковая артерия и вена; 5 - вокругдольковая вена и артерия; 6 - внутридольковые гемокапилляры; 7 - центральная вена; 8 - поддоль-ковая вена; 9 - печеночные вены; 10 - печеночная долька
печени образована печеночными дольками (lobuli hepaticus). Печеночные дольки - структурно-функциональные единицы печени (рис. 16.36).
Существует несколько представлений об их строении. Согласно классическому представлению, печеночные дольки имеют форму шестигранных призм с плоским основанием и слегка выпуклой вершиной. Их ширина не превышает 1,5 мм, тогда как высота, несмотря на значительные колебания, несколько больше. Иногда простые дольки сливаются (по 2 и более) своими основаниями и формируют более крупные сложные печеночные дольки. Количество долек в печени человека достигает 500 тыс. Междольковая соединительная ткань образует строму органа. В ней проходят кровеносные сосуды и желчные протоки, структурно и функционально связанные с печеночными дольками. У человека междольковая соединительная ткань развита слабо, и вследствие этого печеночные дольки плохо отграничены друг от друга. Такое строение характерно для здоровой печени. Наоборот, интенсивное развитие соединительной ткани, сопровождающееся атрофией (уменьшением) печеночных долек, является признаком тяжелого заболевания печени, известного под названием «цирроз».
Кровеносная система. Исходя из классического представления о строении печеночных долек, кровеносную систему печени условно можно разделить на три части: систему притока крови к долькам, систему циркуляции крови внутри них и систему оттока крови от долек (рис. 16.37).
Система притока представлена воротной веной и печеночной артерией. Воротная вена, собирая кровь от всех непарных органов брюшной полости, богатую веществами, всосавшимися в кишечнике, доставляет ее в печень. Печеночная артерия приносит кровь от аорты, насыщенную кислородом. В печени эти сосуды многократно разделяются на всё более мелкие сосуды: долевые, сегментарные, междольковые вены и артерии (vv. и аа. interlobulares), вокругдольковые вены и артерии (vv. и аа. perilobulares). На всем протяжении эти сосуды сопровождаются аналогичными по названию желчными протоками (ductuli biliferi).
Вместе ветви воротной вены, печеночной артерии и желчные протоки составляют так называемые печеночные триады. Рядом с ними лежат лимфатические сосуды.
Междольковые вены и артерии, подразделяющиеся по размеру на 8 порядков, идут вдоль боковых граней печеночных долек. Отходящие от них вокругдольковые вены и артерии опоясывают дольки на разных уровнях.
Междольковые и вокругдольковые вены являются сосудами со слаборазвитой мышечной оболочкой. Однако в местах разветвления в их стенках наблюдаются скопления мышечных элементов, образующих сфинктеры. Соответствующие междольковые и вокругдольковые артерии относятся к сосудам мышечного типа. При этом артерии обычно в несколько раз меньше по диаметру, чем рядом лежащие вены.
От вокругдольковых вен и артерий начинаются кровеносные капилляры. Они входят в печеночные дольки и сливаются, образуя внутридолько-вые синусоидные сосуды, которые составляют систему циркуляции крови в печеночных дольках. По ним течет смешанная кровь в направлении от периферии к центру долек. Соотношение между венозной и артериальной кровью во внутридольковых синусоидных сосудах определяется состоянием сфинктеров междольковых вен. Внутридольковые капилляры относятся к синусоидному (до 30 мкм в диаметре) типу капилляров с прерывистой базальной мембраной. Они идут между тяжами печеночных клеток - печеночными балками, радиально сходясь к центральным венам (vv. centrales), которые лежат в центре печеночных долек.
Центральными венами начинается система оттока крови от долек. По выходе из долек эти вены впадают в поддольковые вены (vv. sublobulares), проходящие в междольковых перегородках. Поддольковые вены не сопровождаются артериями и желчными протоками, т. е. не входят в состав триад. По этому признаку их легко отличить от сосудов системы воротной вены - междольковых и вокругдольковых вен, приносящих кровь к долькам.
Центральные и поддольковые вены - сосуды безмышечного типа. Они сливаются и образуют ветви печеночных вен, которые в количестве 3- 4 выходят из печени и впадают в нижнюю полую вену. Ветви печеночных вен имеют хорошо развитые мышечные сфинктеры. С их помощью регулируется отток крови от долек и всей печени в соответствии с ее химическим составом и массой.
Таким образом, печень снабжается кровью из двух мощных источников - воротной вены и печеночной артерии. Благодаря этому через печень

Рис. 16.38. Ультрамикроскопическое строение печени (по Е. Ф. Котовскому): 1 - внутридольковый синусоидный сосуд; 2 - эндотелиальная клетка; 3 - ситовидные участки; 4 - звездчатые макрофаги; 5 - перисинусоидальное пространство; 6 - ретикулярные волокна; 7 - микроворсинки гепатоцитов; 8 - гепатоциты; 9 - желчный капилляр; 10 - перисинусоидальные жиронакапливающие клетки; 11 - жировые включения в цитоплазме жиронакапливающей клетки; 12 - эритроциты в капилляре
проходит за непродолжительное время вся кровь организма, обогащаясь белками, освобождаясь от продуктов азотистого обмена и других вредных веществ. Паренхима печени имеет огромное число кровеносных капилляров, и вследствие этого кровоток в печеночных дольках осуществляется медленно, что способствует обмену между кровью и клетками печени, выполняющими защитную, обезвреживающую, синтетическую и другие важные для организма функции. При необходимости в сосудах печени может депонироваться большая масса крови.
Классическая печеночная долька (lobulus hepaticus classicus seu poligonalis). Согласно классическому представлению, печеночные дольки образованы печеночными балками и внутридольковыми синусоидными кровеносными капиллярами. Печеночные балки, построенные из гепатоцитов - печеночных эпителиоци-тов, расположены в радиальном направлении. Между ними в том же направлении от периферии к центру долек проходят кровеносные капилляры.
Внутридольковые кровеносные капилляры выстланы плоскими эндотелио-цитами. В области соединения эндотелиальных клеток друг с другом имеются мелкие поры. Эти участки эндотелия называются ситовидными (рис. 16.38).
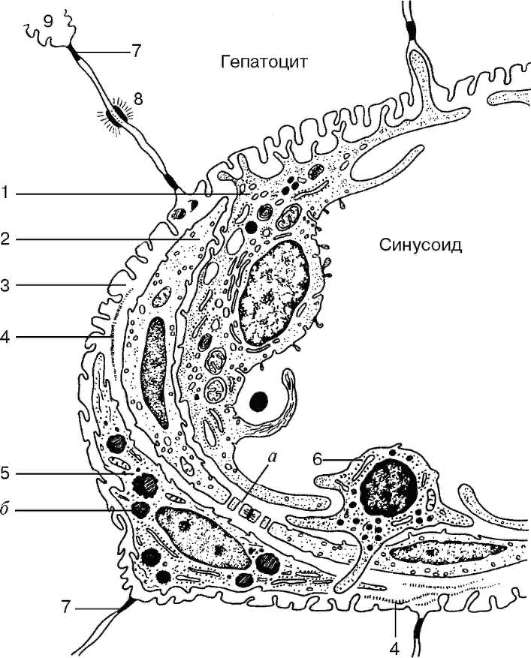
Рис. 16.39. Строение синусоида печени:
1 - звездчатый макрофаг (клетка Купфера); 2 - эндотелиоцит: а - поры (сетевидная зона); 3 - перисинусоидальное пространство (пространство Диссе); 4 - ретикулярные волокна; 5 - жиронакапливающая клетка с каплями липида (б); 6 - ямочная клетка (печеночная НК-клетка, гранулированный лимфоцит); 7 - плотные контакты гепатоцитов; 8 - десмосома гепатоцитов; 9 - желчный капилляр (по Е. Ф. Котовскому)
Между эндотелиоцитами рассеяны многочисленные звездчатые макрофаги (клетки Купфера), не образующие сплошного пласта. В отличие от эндо-телиоцитов они имеют моноцитарное происхождение и являются макрофагами печени (macrophagocytus stellatus), с которыми связаны ее защитные реакции (фагоцитоз эритроцитов, участие в иммунных процессах, разрушение бактерий). Звездчатые макрофаги имеют отростчатую форму и строение, типичное для фагоцитов. К звездчатым макрофагам и эндотелиальным клеткам со стороны просвета синусоидов прикрепляются с помощью псевдоподии ямочные клетки (pit-клетки, печеночные НК-клетки). В их цитоплазме, кроме органелл, присутствуют секреторные гранулы (рис. 16.39). Эти клетки относятся к большим гранулярным лимфоцитам, которые обладают естественной киллерной активностью и одновременно эндокринной
функцией. Благодаря этому печеночные НК-клетки в зависимости от условий могут осуществлять противоположные эффекты: например, при заболеваниях печени они, как киллеры, уничтожают поврежденные гепатоциты, а в период выздоровления, подобно эндокриноцитам (апудоцитам), стимулируют пролиферацию печеночных клеток. Основная часть НК-клеток находится в зонах, окружающих сосуды портального тракта (триады).
Базальная мембрана на большом протяжении у внутридольковых капилляров отсутствует, за исключением их периферических и центральных отделов. Капилляры окружены узким (0,2-1 мкм) перисинусоидальным пространством (Диссе). Через поры в эндотелии капилляров составные части плазмы крови могут попадать в это пространство, а в условиях патологии сюда проникают и форменные элементы. В нем, кроме жидкости, богатой белками, находятся микроворсинки гепатоцитов, иногда отростки звездчатых макрофагов, аргирофильные волокна, оплетающие печеночные балки, а также отростки клеток, известных под названием жиронакапливающие клетки. Эти небольшие (5-10 мкм) клетки располагаются между соседними гепатоцитами. Они постоянно содержат не сливающиеся друг с другом мелкие капли жира, много рибосом и единичные митохондрии. Количество жиронакапливающих клеток может резко возрастать при ряде хронических заболеваний печени. Полагают, что эти клетки, подобно фибробластам, способны к волокнообразованию, а также к депонированию жирорастворимых витаминов. Кроме того, клетки участвуют в регуляции просвета синусоидов и секретируют факторы роста.
Печеночные балки состоят из гепатоцитов, связанных друг с другом дес-мосомами и по типу «замка». Балки анастомозируют между собой, и поэтому их радиальное направление в дольках не всегда четко заметно. В печеночных балках и анастомозах между ними гепатоциты располагаются двумя рядами, тесно прилегающими друг к другу. В связи с этим на поперечном срезе каждая балка представляется состоящей из двух клеток. По аналогии с другими железами печеночные балки можно считать концевыми отделами печени, так как образующие их гепатоциты секретируют глюкозу, белки крови и ряд других веществ.
Между рядами гепатоцитов, составляющих балку, располагаются желчные капилляры, или канальцы, диаметром от 0,5 до 1 мкм. Эти капилляры не имеют собственной стенки, так как образованы соприкасающимися били-арными поверхностями гепатоцитов, на которых имеются небольшие углубления, совпадающие друг с другом и вместе образующие просвет желчного капилляра (рис. 16.40, а, б). Просвет желчного капилляра не сообщается с межклеточной щелью благодаря тому, что мембраны соседних гепатоцитов в этом месте плотно прилегают друг к другу, образуя замыкательные пластинки. Поверхности гепатоцитов, ограничивающие желчные капилляры, имеют микроворсинки, которые вдаются в их просвет.
Полагают, что циркуляция желчи по этим капиллярам (канальцам) регулируется с помощью микрофиламент, располагающихся в цитоплазме гепа-тоцитов вокруг просвета канальцев. При угнетении их сократительной способности в печени может наступить холестаз, т. е. застой желчи в канальцах и протоках. На обычных гистологических препаратах желчные капилляры
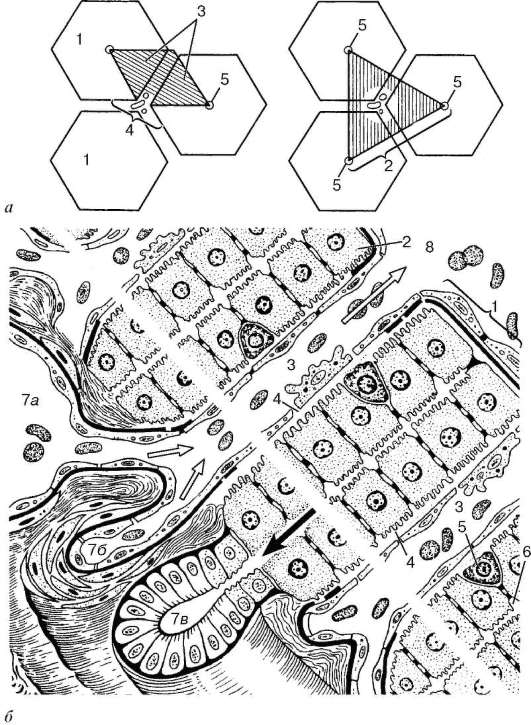
Рис. 16.40. Строение долек (а) и балок (б) печени (по Е. Ф. Котовскому): а - схема строения портальной дольки и ацинуса печени: 1 - классическая печеночная долька; 2 - портальная долька; 3 - печеночный ацинус; 4 - триада; 5 - центральные вены; б - схема строения печеночной балки: 1 - печеночная балка (пластинка); 2 - гепатоцит; 3 - кровеносные капилляры; 4 - перисинусоидальное пространство; 5 - жиронакапливающая клетка; 6 - желчный каналец; 7а - вокруг-дольковая вена; 7б - вокругдольковая артерия; 7 в - вокругдольковый желчный проток; 8 - центральная вена
остаются незаметными и выявляются только при специальных методах обработки (импрегнация серебром или инъекции капилляров окрашенной массой через желчный проток). На таких препаратах видно, что желчные капилляры слепо начинаются на центральном конце печеночной балки, идут вдоль
нее, слегка изгибаясь и отдавая в стороны короткие слепые выросты. Ближе к периферии дольки формируются желчные проточки (холангиолы, канальцы Геринга), стенка которых представлена как гепатоцитами, так и эпителио-цитами (холангиоцитами). По мере увеличения калибра проточка стенка его становится сплошной, выстланной однослойным эпителием. В его составе располагаются малодифференцированные (камбиальные) холангиоциты. Холангиолы впадают в междольковые желчные протоки (ductuli interlobulares).
Таким образом, желчные капилляры располагаются внутри печеночных балок, тогда как между балками проходят кровеносные капилляры. Поэтому каждый гепатоцит в печеночной балке имеет две стороны. Одна сторона - билиарная - обращена к просвету желчного капилляра, куда клетки секретируют желчь (экзокринный тип секреции), другая - васкулярная - направлена к кровеносному внутридольковому капилляру, в который клетки выделяют глюкозу, мочевину, белки и другие вещества (эндокринный тип секреции). Между кровеносными и желчными капиллярами нет непосредственной связи, так как их отделяют друг от друга печеночные и эндотелиальные клетки. Только при заболеваниях (паренхиматозная желтуха и др.), связанных с повреждением и гибелью части печеночных клеток, желчь может поступать в кровеносные капилляры. В этих случаях желчь разносится кровью по всему организму и окрашивает его ткани в желтый цвет (желтуха).
Согласно другой точки зрения о строении печеночных долек, они состоят из широких пластинок (laminae hepaticae), анастомозирующих между собой. Между пластинами располагаются кровяные лакуны (vas sinusoidem), по которым медленно циркулирует кровь. Стенка лакун образована эндо-телиоцитами и звездчатыми макрофагоцитами. От пластин они отделены перилакунарным пространством.
Существуют представления о гистофункциональных единицах печени, отличных от классических печеночных долек. В качестве таковых рассматриваются так называемые портальные печеночные дольки и печеночные ацинусы. Портальная долька (lobulus portalis) включает сегменты трех соседних классических печеночных долек, окружающих триаду. Поэтому она имеет треугольную форму, в ее центре лежит триада, а на периферии, т. е. по углам, - вены (центральные). В связи с этим в портальной дольке кровоток по кровеносным капиллярам направлен от центра к периферии (см. рис. 16.40, а). Печеночный ацинус (acinus hepaticus) образован сегментами двух рядом расположенных классических долек, благодаря чему имеет форму ромба. У острых его углов проходят вены (центральные), а у тупого угла - триада, от которой внутрь ацинуса идут ее ветви (вокругдольковые). От этих ветвей к венам (центральным) направляются гемокапилляры (см. рис. 16.40, а). Таким образом, в ацинусе, как и в портальной дольке, кровоснабжение осуществляется от его центральных участков к периферическим.
Печеночные клетки, или гепатоциты, составляют 60 % всех клеточных элементов печени. Они выполняют большую часть функций, присущих печени. Гепатоциты имеют неправильную многоугольную форму. Диаметр их достигает 20-25 мкм. Многие из них (до 20 % в печени человека) содержат два ядра и больше. Количество таких клеток зависит от функционального
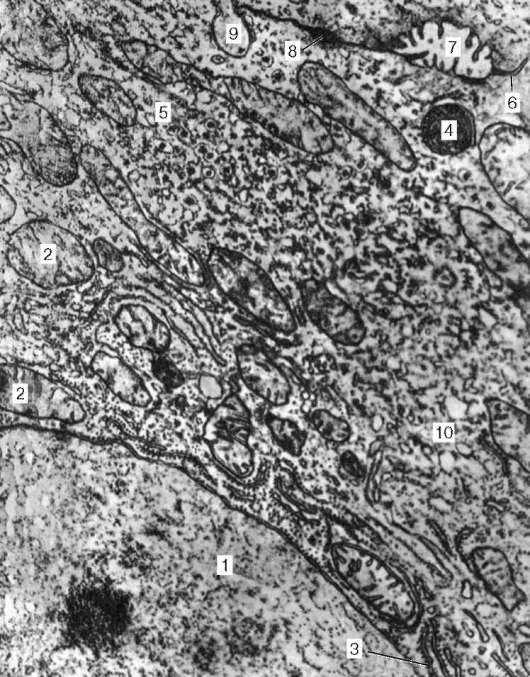
Рис. 16.41. Гепатоцит. Электронная микрофотография, увеличение 8000 (препарат Е. Ф. Котовского):
1 - ядро; 2 - митохондрии; 3 - гранулярная эндоплазматическая сеть; 4 - лизосо-ма; 5 - гликоген; 6 - граница между гепатоцитами; 7 - желчный капилляр; 8 - десмо-сома; 9 - соединение по типу «замка»; 10 - агранулярная эндоплазматическая сеть
состояния организма: например, беременность, лактация, голодание заметно отражаются на их содержании в печени (рис. 16.41).
Ядра гепатоцитов круглой формы, их диаметр колеблется от 7 до 16 мкм. Это объясняется наличием в печеночных клетках наряду с обычными ядрами (диплоидными) более крупных - полиплоидных. Число этих ядер с возрастом постепенно увеличивается и к старости достигает 80 %.
Цитоплазма печеночных клеток окрашивается не только кислыми, но и основными красителями, так как отличается большим содержанием РНП. В ней присутствуют все виды общих органелл. Гранулярная эндоплазма-тическая сеть имеет вид узких канальцев с прикрепленными рибосомами. В центролобулярных клетках она расположена параллельными рядами, а
в периферических - в разных направлениях. Агранулярная эндоплазматическая сеть в виде трубочек и пузырьков встречается либо в небольших участках цитоплазмы, либо рассеяна по всей цитоплазме. Гранулярный вид сети участвует в синтезе белков крови, а агранулярный - в метаболизме углеводов. Кроме того, эндоплазматическая сеть за счет образующихся в ней ферментов осуществляет дезинтоксикацию вредных веществ (а также инактивацию ряда гормонов и лекарств). Около канальцев гранулярной эндо-плазматической сети располагаются пероксисомы, с которыми связан обмен жирных кислот. Большинство митохондрий имеет округлую или овальную форму и размер 0,8-2 мкм. Реже наблюдаются митохондрии нитчатой формы, длина которых достигает 7 мкм и более. Митохондрии отличаются сравнительно небольшим числом крист и умеренно плотным матриксом. Они равномерно распределены в цитоплазме. Количество их в одной клетке может меняться. Комплекс Гольджи в период интенсивного желчеотделения перемещается в сторону просвета желчного капилляра. Вокруг него встречаются отдельные или небольшими группами лизосомы. На васкулярной и билиарной поверхностях клеток имеются микроворсинки.
Гепатоциты содержат различного рода включения: гликоген, липиды, пигменты и другие, образующиеся из продуктов, приносимых кровью. Количество их меняется в различные фазы деятельности печени. Наиболее легко эти изменения обнаруживаются в связи с процессами пищеварения. Уже через 3-5 ч после приема пищи количество гликогена в гепатоцитах возрастает, достигая максимума через 10-12 ч. Через 24-48 ч после еды гликоген, постепенно превращаясь в глюкозу, исчезает из цитоплазмы клеток. В тех случаях, когда пища богата жирами, в цитоплазме клеток появляются капли жира, причем раньше всего - в клетках, расположенных на периферии печеночных долек. При некоторых заболеваниях накопление жира в клетках может переходить в их патологическое состояние - ожирение. Процессы ожирения гепатоцитов резко проявляются при алкоголизме, травмах мозга, лучевой болезни и др. В печени наблюдается суточный ритм секреторных процессов: днем преобладает выделение желчи, а ночью - синтез гликогена. По-видимому, этот ритм регулируется при участии гипоталамуса и гипофиза. Желчь и гликоген образуются в разных зонах печеночной дольки: желчь обычно вырабатывается в периферической зоне, и только затем этот процесс постепенно распространяется на центральную зону, а отложение гликогена осуществляется в обратном направлении - от центра к периферии дольки. Гепатоциты непрерывно выделяют в кровь глюкозу, мочевину, белки, жиры, а в желчные капилляры - желчь.
Желчевыводящие пути. К ним относятся внутрипеченочные и внепече-ночные желчные протоки. К внутрипеченочным принадлежат междольковые желчные протоки, а к внепеченочным - правый и левый печеночные протоки, общий печеночный, пузырный и общий желчный протоки. Междольковые желчные протоки вместе с разветвлениями воротной вены и печеночной артерии образуют в печени триады. Стенка междольковых протоков состоит из однослойного кубического, а в более крупных протоках - из цилиндрического эпителия, снабженного каемкой, и тонкого слоя рыхлой соединительной ткани. В апикальных отделах эпителиальных клеток протоков нередко встре-
чаются в виде зерен или капель составные части желчи. На этом основании предполагают, что междольковые желчные протоки выполняют секреторную функцию. Печеночные, пузырный и общий желчный протоки имеют примерно одинаковое строение. Это сравнительно тонкие трубки диаметром около 3,5-5 мм, стенка которых образована тремя оболочками. Слизистая оболочка состоит из однослойного высокого призматического эпителия и хорошо развитого слоя соединительной ткани (собственная пластинка). Для эпителия этих протоков характерно наличие в его клетках лизосом и включений желчных пигментов, что свидетельствует о резорбтивной, т. е. всасывательной, функции эпителия протоков. В эпителии нередко встречаются эндокринные и бокаловидные клетки. Количество последних резко увеличивается при заболеваниях желчных путей. Собственная пластинка слизистой оболочки желчных протоков отличается богатством эластических волокон, расположенных продольно и циркулярно. В небольшом количестве в ней имеются слизистые железы. Мышечная оболочка тонкая, состоит из спирально расположенных пучков гладких миоцитов, между которыми много соединительной ткани. Мышечная оболочка хорошо выражена лишь в определенных участках протоков - в стенке пузырного протока при переходе его в желчный пузырь и в стенке общего желчного протока при впадении его в двенадцатиперстную кишку. В этих местах пучки гладких миоцитов располагаются главным образом циркулярно. Они образуют сфинктеры, которые регулируют поступление желчи в кишечник. Адвентициальная оболочка состоит из рыхлой соединительной ткани.
16.5. ЖЕЛЧНЫЙ ПУЗЫРЬ
Желчный пузырь (vesica biliaris seu fellae) представляет собой тонкостенный орган (толщина стенки 1,5-2 мм), вмещающий 40-70 мл желчи. Стенка желчного пузыря состоит из трех оболочек: слизистой, мышечной и адвен-тициальной (рис. 16.42).
Желчный пузырь со стороны брюшной полости покрыт серозной оболочкой. Слизистая оболочка образует многочисленные складки. Она выстлана однослойным столбчатым эпителием. В составе эпителия различают поверхностные эпителиоциты, бокаловидные клетки и базальные (камбиальные) клетки. Поверхностные столбчатые эпителиоциты (холецистоциты) имеют микроворсинчатую каемку. Под эпителием располагается собственная пластинка слизистой оболочки, содержащая большое количество эластических волокон. Там же встречаются жировые, тучные и плазматические клетки. В области шейки пузыря в ней находятся альвеолярно-трубчатые железы, выделяющие слизь. Эпителий слизистой оболочки обладает способностью всасывать воду и некоторые другие вещества из желчи, заполняющей полость пузыря. Поэтому пузырная желчь всегда более густой консистенции и более темного цвета, чем желчь, изливающаяся непосредственно из печени. Мышечная оболочка желчного пузыря состоит из пучков гладких миоцитов, расположенных в виде сети, в которой миоциты располагают-
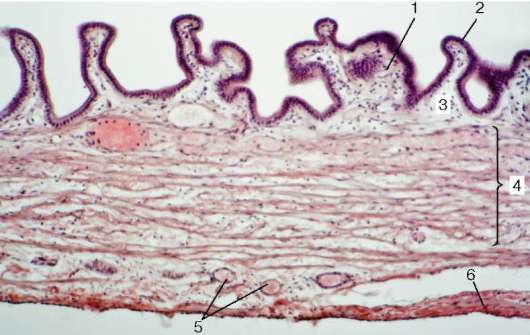
Рис. 16.42. Строение желчного пузыря:
1 - складки слизистой оболочки; 2 - эпителий слизистой оболочки; 3 - собственная пластинка слизистой оболочки; 4 - мышечная оболочка; 5 - кровеносные сосуды; 6 - наружная оболочка
ся преимущественно циркулярно. В области шейки пузыря циркулярные пучки мышечных клеток особенно сильно развиты. Вместе с мышечным слоем пузырного протока они образуют сфинктер. Между пучками мышечных клеток всегда имеются хорошо выраженные прослойки рыхлой соединительной ткани. Адвентициальная оболочка желчного пузыря состоит из плотной соединительной ткани, в которой содержится много толстых эластических волокон, образующих сети. Со стороны брюшной полости стенку желчного пузыря покрывает серозная оболочка.
Иннервация. В капсуле печени находится автономное нервное сплетение, ветви которого, сопровождая кровеносные сосуды, продолжаются в меж-дольковую соединительную ткань. Они иннервируют кровеносные сосуды, желчные протоки и гепатоциты в печеночных балках, формируя окончания в виде небольших утолщений. По ходу нервов располагаются отдельные нервные клетки.
Возрастные изменения. С возрастом в гепатоцитах повышается количество липофусцина, который окрашивает клетки в бурый цвет. Число делящихся клеток резко уменьшается. Ядра гепатоцитов увеличиваются в размерах - гипертрофируются, в них повышается содержание ДНК и наблюдается гиперхроматоз. В старческом возрасте отмечается разрастание соединительной ткани между печеночными дольками.
Регенерация. Печень обладает высокой способностью к физиологической и репаративной регенерации. У животных при удалении от 50 до 70 % ткани печени исходная ее масса восстанавливается на 10-14-е сут. Процессы регенерации происходят путем компенсаторного увеличения размеров клеток (гипертрофии) и размножения гепатоцитов. Стимулирует регенерацию печени пища, богатая углеводами и белками. Однако хорошая регенераци-
онная способность печени не характерна для человека. В целом гепатоциты и холангиоциты относят к растущей клеточной популяции.
16.6. ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
Поджелудочная железа (pancreas) является смешанной железой, включающей экзокринную и эндокринную части. В экзокринной части вырабатывается панкреатический сок (около 1500-3000 мл в сутки), богатый пищеварительными ферментами - трипсиногенами, липазами, амилазой и другими, поступающими по выводному протоку в просвет двенадцатиперстной кишки, где его проферменты активируются и участвуют в расщеплении белков, жиров и углеводов до конечных продуктов. В эндокринной части синтезируется ряд гормонов: инсулин, глюкагон, соматостатин, ВИП, панкреатический полипептид, - принимающих участие в регуляции углеводного, белкового и жирового обмена в тканях и др.
Развитие. Поджелудочная железа развивается из энтодермы и мезенхимы. Ее зачаток появляется в конце 3-й нед эмбриогенеза в виде дорсального и вентральных выпячиваний стенки туловищного отдела эмбриональной кишки, врастающих в брыжейку. Из них формируются головка, тело и хвост железы. На 3-м мес плодного периода энтодермальные зачатки начинают дифференцироваться на экзокринные и эндокринные отделы железы. Из эпителиальных разрастаний зачатка железы вначале образуются выводные протоки, а затем концевые отделы - панкреатические аци-нусы. Последними из камбиальных клеток терминальных участков выводных протоков развиваются эндокринные клетки островков поджелудочной железы. Из мезенхимы развиваются соединительнотканные элементы стро-мы, а также сосуды. К моменту рождения обе части железы приобретают дифференцированное состояние, однако в постнатальном онтогенезе продолжается структурно-функциональная перестройка органа.
Строение. Поджелудочная железа с поверхности покрыта тонкой соединительнотканной капсулой, срастающейся с висцеральным листком брюшины. Ее паренхима разделена на дольки, между которыми проходят соединительнотканные тяжи. В них расположены кровеносные сосуды, нервы, интрамураль-ные нервные ганглии, пластинчатые тельца и выводные протоки (рис. 16.43). Дольки включают экзокринные и эндокринные части железы. На долю первой части приходится около 97 %, а второй - до 3 % массы железы.
16.6.1. Экзокринная часть
Эта часть поджелудочной железы в дольках представлена панкреатическими ацинусами, вставочными и внутридольковыми протоками, а также междольковыми протоками и общим панкреатическим протоком, открывающимся в двенадцатиперстную кишку.
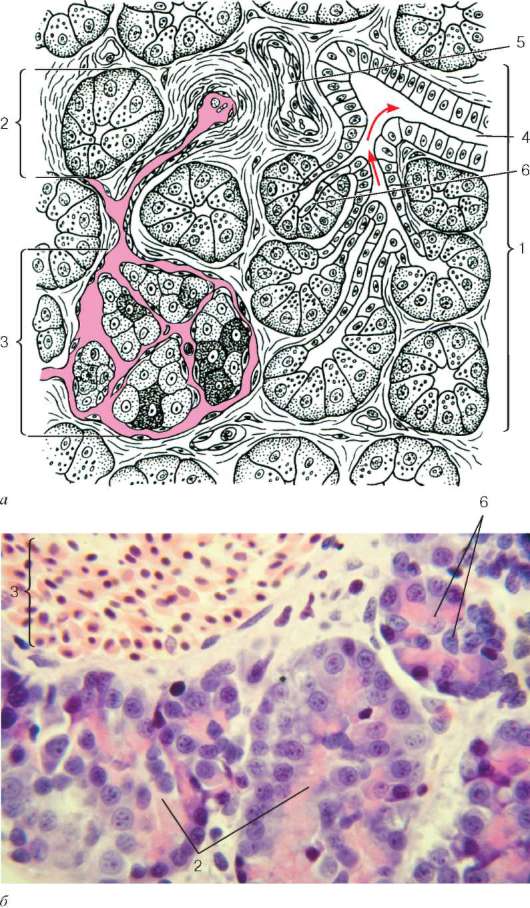
Рис. 16.43. Строение поджелудочной железы:
а - схема (по Ю. И. Афанасьеву и В. В. Яглову); б - микрофотография. 1 - долька; 2 - панкреатические ацинусы; 3 - панкреатический островок; 4 - внутридолько-вый проток; 5 - кровеносные сосуды; 6 - центроацинозные клетки
Структурно-функциональной единицей экзокринной части поджелудочной железы является панкреатический ацинус (acinus pancreaticus). Он включает секреторный отдел и вставочный проток, которым начинается вся про-токовая система железы. Внешне ацинус напоминает мешочек размером 100-150 мкм. Между ацинусами располагаются ретикулярные волокна, кровеносные капилляры, а также нервные волокна и нервные ганглии вегетативной нервной системы. Ацинусы состоят из 8-12 крупных экзокринных панкреатоцитов ( pancreatocyti exocrini), расположенных на базальной мембране, и нескольких мелких центроацинарных клеток (см. рис. 16.43).
Экзокринные панкреатоциты выполняют секреторную функцию, синтезируя пищеварительные ферменты панкреатического сока. Они имеют форму конуса с суженной верхушкой и широким основанием, лежащим на базальной мембране ацинуса. Плазмолемма на базальной поверхности клеток образует внутренние складки, а на апикальной поверхности - микроворсинки. Между боковыми поверхностями клеток имеются контакты типа замыкательных пластинок и десмосом. Верхушечные (апикальные) части клеток называют зимо-генными зонами, а противоположные базальные части клеток - гомогенными зонами. Зимогенные зоны клеток окрашиваются кислыми красителями, т. е. являются оксифильными. Зимогенная зона клеток в основном занята крупными секреторными гранулами (диаметром до 80 нм). Среди них выделяют гранулы различной степени зрелости (плотности). Зимогенные гранулы содержат синтезируемые в клетках ферменты в неактивной форме, т. е. в виде зимогена. В гомогенной зоне преобладает гранулярная эндоплазматическая сеть, состоящая из массы плоских мешочков, мембраны которых усеяны рибосомами. В них осуществляется синтез ферментов панкреатического сока. Обилие рибосом обусловливает базофилию этой зоны. Ядра экзокринных панкреатоцитов, содержащие 1-2 ядрышка, преимущественно располагаются в их базальной части. В надъядерной части клеток расположен хорошо развитый комплекс Гольджи. Митохондрии рассеяны по всей цитоплазме, но большинство их находится под плазмолеммой и рядом с комплексом Гольджи. Они отличаются разнообразной формой.
Секреторную деятельность экзокринные панкреатоциты осуществляют циклически. Их секреторный цикл, включающий фазы поглощения исходных веществ, синтеза секрета, накопления его и затем выделения по мерокринному типу, занимает в среднем 1,5-2 ч. Однако в зависимости от физиологических потребностей организма в пищеварительных ферментах этот цикл может сократиться или, наоборот, увеличиться.
Выделившийся из экзокринных панкреатоцитов секрет попадает во вставочный проток (ductus intercalates), стенка которого состоит из мелких клеток. В одних случаях они примыкают к секреторным клеткам сбоку, имея общую с ними базальную мембрану, в других - проникают в центр ацинуса, располагаясь на апикальной поверхности экзокринных панкреа-тоцитов. При такой локализации они называются центроацинарными клетками (cellulae centroacinosi).
Центроацинарные клетки имеют неправильную, уплощенную форму, их овальное ядро окружено узким слоем светлой цитоплазмы, бедной органел-
лами. На свободной поверхности, обращенной в просвет ацинуса, имеются единичные микроворсинки.
Вставочные протоки переходят в межацинозные протоки (ductus interacinosi). Стенки этих протоков выстланы однослойным кубическим эпителием. Их плазмолемма образует внутренние складки и микроворсинки. Клетки соединяются друг с другом с помощью десмосом. В цитоплазме клеток много митохондрий и хорошо развит комплекс Гольджи. Существует мнение, что эпителиальные клетки протоков продуцируют жидкий компонент панкреатического сока.
Межацинозные протоки впадают в более крупные внутридольковые протоки (ductus intralobulares), стенки которых покрыты однослойным кубическим эпителием. Ядра эпителиальных клеток крупные, в цитоплазме находятся немногочисленные митохондрии, слабовыраженный комплекс Гольджи, свободные рибосомы, гладкая эндоплазматическая сеть. Вокруг протоков расположена рыхлая соединительная ткань, в которой проходят кровеносные капилляры и нервные волокна.
Внутридольковые протоки далее продолжаются в междольковые протоки (ductus interlobulares), которые лежат в соединительнотканных перегородках (септах) между дольками. Они впадают в общий проток поджелудочной железы, проходящий в ее толще от хвоста к головке, где он открывается (вместе с общим желчным протоком) в полость двенадцатиперстной кишки. Все эти протоки выстланы слизистой оболочкой, состоящей из однослойного столбчатого эпителия и собственной пластинки слизистой оболочки. В устье общего протока, кроме того, имеются циркулярно расположенные гладкие миоциты, образующие его сфинктер.
В эпителии протоков имеются бокаловидные клетки, а также эндокрино-циты, вырабатывающие гормоны панкреозимин и холецистокинин. Под их влиянием стимулируются секреторная активность экзокринных панкреа-тоцитов и выделение желчи из печени. В собственной пластинке слизистой оболочки протоков находятся мелкие слизистые железы.
16.6.2. Эндокринная часть
Эта часть поджелудочной железы представлена панкреатическими островками (insulae pancreaticae), или островками Лангерганса, лежащими между панкреатическими ацинусами (см. рис. 16.43; рис. 16.44). Они обычно имеют округлую или овальную форму, но наряду с этим могут встречаться островки лентовидной и звездчатой формы. В среднем их диаметр колеблется от 100 до 300 мкм. Наибольшее количество островков располагается в хвостовой части железы. Общее их число колеблется от 1 до 2 млн и более.
Островки состоят из эндокринных клеток, между которыми находятся кровеносные капилляры фенестрированного типа, окруженные перикапил-лярным пространством. Именно сюда прежде всего поступают инсулярные гормоны.
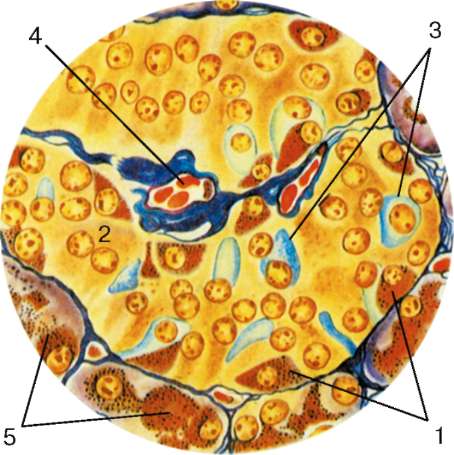
Рис. 16.44. Панкреатический островок: 1 - А-клетки (ацидофильные, глюкаго-ноциты); 2 - В-клетки (базофильные, инсулиноциты); 3 - D-клетки (сомато-статиноциты); 4 - кровеносные капилляры; 5 - панкреатические ацинусы
Эндокринные клетки, в отличие от экзокринных панкреатоцитов, имеют меньшие размеры. В их цитоплазме умеренно развита гранулярная эндо-плазматическая сеть, но хорошо представлены комплекс Гольджи, митохондрии (мелкие) и секреторные гранулы (рис. 16.45). Эти гранулы по своим физико-химическим и морфологическим свойствам неодинаковы в различных клетках островков. На этом основании среди эндокринных клеток различают следующие основные виды: В-клетки (базофильные, инсулоциты), А-клетки (ацидофильные, глюкагоноциты), D-клетки (дендритические, соматостатиноциты), РР-клетки (клетки, продуцирующие панкреатический полипептид).
В-клетки составляют основную массу клеток островков (около 70- 75 %). Большая часть их лежит в центре островков. Секреторные В-гранулы клеток не растворяются в воде, но полностью растворяются в спирте. Они проявляют базофильные свойства, окрашиваясь альдегидфуксином, ген-циановым фиолетовым в синий цвет. Гранулы имеют размер около 275 нм. Между их содержимым и покрывающей мембраной имеется широкий светлый ободок. В некоторых гранулах содержится плотная кристаллическая структура, в которой обнаруживается цинк. Гранулы В-клеток состоят из гормона инсулина. Одним из наиболее ярких эффектов инсулина является его гипогликемическое действие, так как он способствует усвоению глюкозы крови клетками тканей. Поэтому при недостатке инсулина количество глюкозы в тканях снижается, а содержание ее в крови резко возрастает, что приводит к сахарному мочеизнурению (сахарный диабет).
А-клетки составляют примерно 20-25 % всей совокупности эндокрино-цитов. В островках они занимают преимущественно периферическое положение. А-гранулы клеток устойчивы к спирту, но растворяются в воде. Они обладают оксифильными свойствами, в связи с чем окрашиваются кислым фуксином в ярко-красный цвет. Размеры гранул около 230 нм. Их плотное содержимое отделено от окружающей мембраны узким светлым ободком. В гранулах А-клеток обнаружен гормон глюкагон. По своему действию он является антагонистом инсулина. Под его влиянием в тканях происходит усиленное расщепление гликогена до глюкозы. В связи с этим в случаях его недостатка количество глюкозы в крови может снижаться.
Следовательно, инсулин и глюкагон строго поддерживают физиологическую концентрацию сахара в крови и определяют содержание гликогена в тканях (прежде всего в печени).
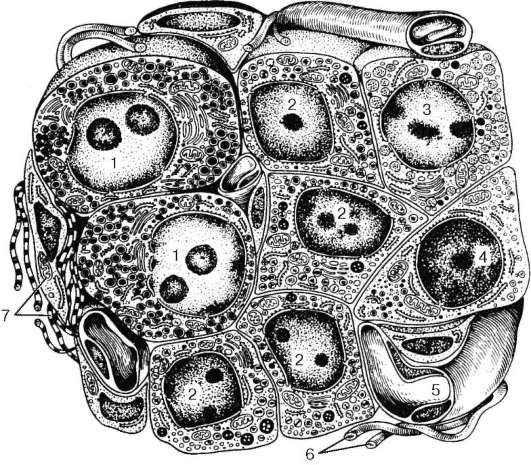
Рис. 16.45. Ультрамикроскопическое строение клеток панкреатического островка (по Ю. И. Афанасьеву):
1 - эндокриноцит А; 2 - эндокриноцит В; 3 - эндокриноцит D; 4 - эндокриноцит РР; 5 - капилляр; 6 - нервные волокна; 7 - рыхлая соединительная ткань
D-клетки, число которых в островках невелико (5-10 %), располагаются в основном на их периферии, имеют грушевидную и реже звездчатую форму. D-гранулы среднего размера (325 нм), умеренной плотности и лишены светлого ободка. D-клетки секретируют гормон соматостатин. Этот гормон задерживает выделение инсулина и глюкагона А- и В-клетками, а также подавляет синтез ферментов экзокринными панкреатоцитами.
РР-клетки (2-5 %) вырабатывают панкреатический полипептид, стимулирующий выделение желудочного и панкреатического сока. Это полигональные клетки с очень мелкими зернами в цитоплазме (размер гранул не более 140 нм). РР-клетки обычно локализуются по периферии островков в области головки железы, а также встречаются вне островков в экзокринной части железы и в составе эпителия протоков.
Помимо экзокринных панкреатоцитов и эндокринных клеток, в дольках поджелудочной железы описан еще один вид секреторных клеток - промежуточные, или ациноостровковые, клетки, которые, однако, встречаются редко (0,08 %) (рис. 16.46). Они располагаются группами вокруг островков среди панкреатических ацинусов. Характерной особенностью промежуточных клеток является наличие в них гранул двух типов - крупных зимо-генных, присущих экзокринным панкреатоцитам, и мелких, типичных для эндокринных клеток. Их митохондрии делятся по величине на крупные и
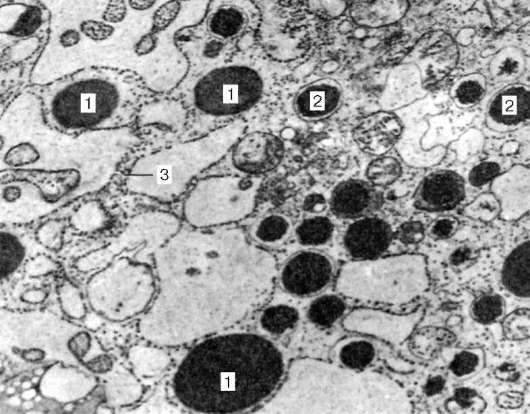
Рис. 16.46. Ациноостровковая клетка. Электронная микрофотография, увеличение 10 000 (препарат В. В. Яглова):
1 - зимогенные гранулы; 2 - эндокринные гранулы; 3 - гранулярная эндоплазма-тическая сеть
мелкие, а гранулярная эндоплазматическая сеть по степени развития занимает промежуточное положение. Гранулы ациноостровковых клеток напоминают гранулы одного из видов клеток островков - А, В или D. В связи с этим предложено классифицировать ациноостровковые клетки по их гормональному профилю на три типа: А, В и D. Большая часть ациноостров-ковых клеток выделяет в кровь как эндокринные, так и зимогенные гранулы. Реже встречаются клетки, из которых те и другие гранулы поступают в выводные протоки железы.
Васкуляризация. Поджелудочная железа снабжается кровью, приносимой по ветвям чревной и верхней брыжеечной артерий. Разветвления этих артерий в междольковой соединительной ткани и внутри долек образуют густые капиллярные сети, оплетающие ацинусы и проникающие в островки. Существует мнение, что эти капиллярные сети между собой не сообщаются. Согласно другому предположению, в дольках железы существует портальная система сосудов, когда приносящая артериола распадается на капилляры островков, а затем они собираются в выносящие артериолы, от которых начинается новая сеть капилляров, оплетающих ацинусы экзо-кринной части железы. Оттекающая из поджелудочной железы венозная кровь поступает в воротную вену.
Лимфатическая система начинается щелевидными капиллярами вокруг ацинусов и островков. Лимфатические капилляры вливаются в лимфатические сосуды, которые проходят вблизи кровеносных.
Иннервация. Эфферентная иннервация поджелудочной железы осуществляется блуждающим и симпатическими нервами. Симпатические волокна сопровождают кровеносные сосуды, являясь по своему значению сосудо-двигательными. В поджелудочной железе имеются интрамуральные автономные ганглии. Основную массу их нервных клеток составляют холинер-гические нейроны. Вместе с тем в ганглиях содержатся и пептидергические нейроны, секретирующие полипептидные гормоны. Нервные волокна холи-нергических и пептидергических нейронов заканчиваются на клетках панкреатических ацинусов и вдоль капилляров проходят в островки, регулируя секреторную функцию железы.
Чувствительные нервные волокна образуют в междольковой соединительной ткани разнообразные рецепторы, в том числе пластинчатые тельца.
Возрастные изменения в поджелудочной железе проявляются, прежде всего, в изменении соотношения между ее экзокринной и эндокринной частями. Островки наиболее сильно развиты в железе в первые годы жизни. С возрастом их число постепенно уменьшается.
Регенерация. Пролиферативная (митотическая) активность клеток поджелудочной железы крайне низкая, поэтому в физиологических условиях в ней происходит обновление клеток путем внутриклеточной регенерации. При удалении части органа регистрируется митотическая активность в зоне регенерации. Последующая дифференцировка эпителия регенерата завершается образованием новых ацинусов. Наиболее ярко проявляется регенерационная гипертрофия эпителиоцитов в неповрежденных участках экзокринной паренхимы. При повреждении панкреатических островков восстановление эндокриноцитов осуществляется путем кратковременной митотической активности эндокриноцитов, а также дифференцировки промежуточных (ациноостровковых) клеток в эндокриноциты. Не исключается участие в регенерации эпителиоцитов протоков поджелудочной железы, которые дифференцируются в эндокриноциты.
Контрольные вопросы
1. Сравнительная морфофизиологическая характеристика разных отделов пищеварительной трубки в аспекте последовательности химического расщепления пищи.
2. Развитие, строение, смена молочных и постоянных зубов.
3. Крупные слюнные железы: развитие, сравнительная структурно-функциональная характеристика.
4. Развитие, строение, функции и регенерация печени.
5. Развитие, строение, функции и особенности регенерации поджелудочной железы.