Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 15. ЭНДОКРИННАЯ СИСТЕМА
Эндокринная система - совокупность структур: органов, частей органов, отдельных клеток, секретирующих в кровь и лимфу гормоны.
Гормоны (от греч. hormau - возбуждаю) - это высокоактивные регуля-торные факторы, оказывающие стимулирующее или угнетающее влияние преимущественно на основные функции организма: обмен веществ, соматический рост, репродуктивные функции.
Эндокринная система совместно с нервной системой осуществляет регуляцию и координацию функций организма. В состав эндокринной системы входят специализированные эндокринные железы, или железы внутренней секреции. Последние - это органы, которые образуют и секретируют в кровь, лимфу и межклеточную среду гормоны, обладающие высокой биологической активностью, оказывающие контактное и дистантное воздействие на процессы жизнедеятельности других клеток и тканей организма. Помимо эндокринных желез, в организме человека существует огромное количество одиночных эндокриноцитов, расположенных в составе эпителиальных тканей кожи, дыхательной, пищеварительной и выделительной систем, составляющие так называемую дисперсную эндокринную систему.
По химическому строению гормоны подразделяются на производные аминокислот (адреналин и норадреналин и др.), пептиды (наиболее многочисленный класс, насчитывающий более 50 гормонов, среди которых - инсулин, глюкагон, ингибин, гастрин и др.), стероидные (половые, коры надпочечников и др.), ненасыщенные жирные кислоты (простагландины). По физиологическому действию различают пусковые гормоны и гормоны-исполнители. Пусковыми гормонами являются нейрогормоны гипоталамуса и гормоны гипофиза, они стимулируют или тормозят синтез и секрецию гормонов в других железах внутренней секреции. Гормоны-исполнители действуют непосредственно на обменные процессы в клетках и тканях-мишенях. Последние бывают гормонозависимыми (функционируют толь-
ко в присутствии конкретного гормона) и гормоночувствительными (могут функционировать без гормональной стимуляции, но их деятельность все же контролируется гормоном).
Клетки-мишени активно захватывают и аккумулируют гормон с помощью специфического для данного гормона белка-рецептора (избирательность связывания гормона). Рецепторы могут быть внутриклеточные (для гормонов, проникающих в цитозоль) или располагаться в виде интегральных белков плазмолеммы (для гормонов, не проникающих в клетку). В последнем случае необходимы дополнительные механизмы передачи гормонального сигнала к внутриклеточным элементам. Передача стимула внутрь клетки осуществляется вторичными медиаторами, или посредниками (цАМФ, цГМФ, ионы кальция и др.). При соединении гормона с рецептором происходит активация гормонзависимого фермента плазмолеммы - аденилил-циклазы. Последняя активирует в цитоплазме образование из АТФ внутриклеточного посредника - циклического аденозинмонофосфата (цАМФ). Далее следует взаимодействие посредника с внутриклеточным рецептором и перемещение комплекса цАМФ-рецептор в ядро и возникновение новых синтезов. При этом ускоряется течение обменных реакций в клетке.
Стероидные гормоны способны проходить через плазмолемму и взаимодействовать с внутриклеточными рецепторами. Они могут действовать и на генетический аппарат клеток-мишеней.
Гормоны обладают высокой биологической активностью, хотя и продуцируются в очень малых количествах. При введении извне эффективными являются чрезвычайно малые концентрации гормонов.
15.1. ВЗАИМОСВЯЗЬ НЕРВНОЙ И ЭНДОКРИННОЙ СИСТЕМ
Общим для нервных и эндокринных клеток является выработка гуморальных регулирующих факторов. Эндокринные клетки синтезируют гормоны и выделяют их в кровь, а нейроны синтезируют нейротрансмиттеры, или переключатели (большинство из которых является нейроаминами): норадреналин, серотонин и другие, выделяющиеся в синаптические щели. В гипоталамусе находятся секреторные нейроны, совмещающие свойства нервных и эндокринных клеток. Они обладают способностью образовывать как нейроамины, так и олигопептидные гормоны (рис. 15.1). Нейроэндокринные клетки объединяют нервную и эндокринную системы в единую нейроэндокринную систему.
В результате новых открытий было показано большое сходство в организации и функционировании структурных элементов нервной и эндокринной систем с таковыми иммунной системы. Так, клетки иммунной системы способны экспрессировать рецепторы для сигнальных молекул, опосредующих воздействия нейроэндокринной системы, а клетки последней могут экспрессировать рецепторы для медиатора иммунной системы. Таким об-
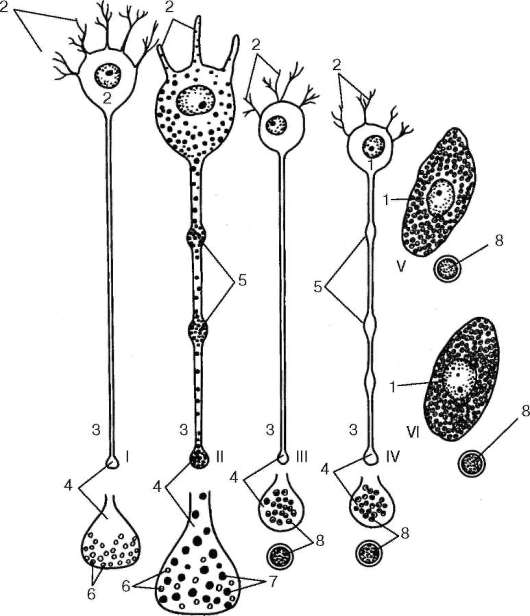
Рис. 15.1. Строение нервных, нейросекреторных и эндокринных клеток (по Б. В. Алешину):
I - холинергический нейрон с ацетилхолиновыми пузырьками в терминалях;
II - гомориположительная нейросекреторная клетка переднего гипоталамуса (пеп-тидохолинергический нейрон), продуцирующая белковые гранулы; III - адренер-гический нейрон с гранулами в терминали, содержащими белковую сердцевину, на которой аккумулируются катехоламины; IV - нейросекреторная пептидадре-нергическая клетка медиобазального гипоталамуса; V - эндокринная клетка (хро-маффинная клетка мозговой части надпочечников) с секреторными гранулами, как в адренергических нейронах (III); VI - эндокринная клетка, вырабатывающая белковые гормоны (парафолликулярные клетки щитовидной железы, энтероциты слизистой оболочки пищеварительного тракта и панкреатические островки), содержит секреторные гранулы с белковой сердцевиной. 1 - перикарион; 2 - дендриты; 3 - аксон; 4 - терминаль аксона; 5 - зоны накопления нейросекрета; 6 - синапти-ческие пузырьки; 7 - гранулы нейрогормона; 8 - структура секреторных гранул
разом, наблюдается трансформация традиционной нейроэндокринологии в нейроиммуноэндокринологию - перспективную область науки в исследовании физиологических основ деятельности мозга и понимания механизмов, лежащих в основе различных патологических процессов.
Внутри эндокринной системы существуют сложные взаимодействия между центральными и периферическими органами этой системы.
Классификация. По происхождению, гистогенезу и гистологическим признакам эндокринные органы классифицируют на три группы: бранхиогенная группа (от греч. branchia - жабры) - железы, происшедшие из глоточных карманов - аналогов жаберных щелей (щитовидная железа, околощитовидные железы); группа надпочечников (корковое и мозговое вещество надпочечников, параганглии); группа мозговых придатков (гипоталамус, гипофиз и эпифиз). Поскольку эндокринные железы составляют единую в функциональном отношении регулирующую систему, существует классификация, в которой учтены межорганные связи и иерархическая зависимость эндокринных органов.
I. Центральные звенья эндокринного комплекса желез (регулируют деятельность большинства периферических эндокринных желез):
1) гипоталамус (нейросекреторные ядра);
2) гипофиз (аденогипофиз и нейрогипофиз);
3) эпифиз.
IIa. Периферические аденогипофиззависимые эндокринные железы и эндо-криноциты:
1) щитовидная железа (тироциты);
2) надпочечники (корковое вещество);
3) гонады (яички, яичники).
IIб. Периферические аденогипофизнезависимые эндокринные железы и эндокриноциты:
1) кальцитониноциты щитовидной железы;
2) околощитовидные железы;
3) мозговое вещество надпочечников и параганглии;
4) эндокринные клетки островков поджелудочной железы (Лангерганса);
5) нейроэндокриноциты в составе неэндокринных органов, эндокрино-циты дисперсной эндокринной системы (APUD-серия клеток).
Среди органов и образований эндокринной системы с учетом их функциональных особенностей различают четыре основные группы.
I. Нейроэндокринные трансдукторы (переключатели), выделяющие нейро-трансмиттеры (посредники) - либерины (стимуляторы) и статины (тормозящие факторы).
Нейрогемальные образования (медиальное возвышение гипоталамуса), задняя доля гипофиза, которые не вырабатывают собственных гормонов, но накапливают гормоны, продуцируемые в нейросекреторных ядрах гипоталамуса.
III. Центральный орган регуляции эндокринных желез и неэндокринных функций - аденогипофиз, осуществляющий регуляцию с помощью вырабатываемых в нем специфических тропных гормонов.
IV. Периферические эндокринные железы и структуры (аденогипофиззависимые и аденогипофизнезависимые).
Как в любой системе, центральные и периферические ее звенья имеют прямые и обратные связи. Гормоны, вырабатываемые в периферических эндокринных образованиях, могут оказывать регулирующее влияние на деятельность центральных звеньев.
Одной из особенностей строения эндокринных органов является обилие в них сосудов, особенно гемокапилляров синусоидного типа и лимфатических капилляров, в которые поступают секретируемые гормоны.
15.2. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ЭНДОКРИННОЙ СИСТЕМЫ
15.2.1. Гипоталамус
Гипоталамус (hypothalamus) является высшим нервным центром регуляции эндокринных функций. Он контролирует и интегрирует все висцеральные функции организма и объединяет эндокринные механизмы регуляции с нервными, будучи мозговым центром симпатического и парасимпатического отделов автономной нервной системы. Субстратом объединения нервной и эндокринной систем являются нейросекреторные клетки, которые у высших позвоночных и человека располагаются в нейросекреторных ядрах гипоталамуса.
Нейроэндокринные трансдукторы (переключатели) и нейрогемальные образования. Медиальное возвышение (eminentia medialis) является нейрогемальным органом гипоталамо-аденогипофизарной системы. Оно образовано эпендимой, отдельные глиальные клетки которой дифференцируются в танициты (tanicyti), отличающиеся разветвленными отростками, контактирующими с клубочками первичной капиллярной сети портальной системы гипоталамо-гипофизарного кровообращения. В гипоталамо-аденогипофизарной системе аккумулируются аденогипофизотропные нейрогормоны - нейротранс-миттеры (либерины и статины), вырабатываемые в мелкоклеточных ядрах среднего и заднего отделов гипоталамуса, которые затем поступают в портальную систему гипофиза. В гипоталамо-нейрогипофизарной системе аналогичным нейрогемальным органом оказывается нейрогипофиз (задняя доля гипофиза), где аккумулируются нонапептидные нейрогормоны (вазопрес-син - антидиуретический гормон и окситоцин), вырабатываемые в крупноклеточных ядрах переднего отдела гипоталамуса, в дальнейшем выделяемые в кровь.
Секреторные нейроны (neuronum secretorium) расположены в ядрах серого вещества гипоталамуса. Строение секреторных нейронов описано в главе 10. Нервные ядра (свыше 30 пар) группируются в его переднем, среднем (медиобазальном и туберальном) и заднем отделах.
В переднем гипоталамусе располагаются парные супраоптические (nuclei supraoptici) и паравентрикулярные (nuclei paraventriculares) ядра. Супраоптические
ядра образованы крупными холинергическими (пептидохолинергическими) нейросекреторными клетками, содержащими как в перикарионах, так и в отростках секреторные гранулы. Аксоны этих клеток проходят через медиальную эминенцию и гипофизарную ножку в заднюю долю гипофиза, где заканчиваются на кровеносных капиллярах утолщенными терминалями. Паравентрикулярные ядра построены более сложно.
Их центральная крупноклеточная часть образована такими же крупными холинергическими нейросекреторными клетками, как в супраоптическом ядре, и их аксонами, идущими в заднюю долю гипофиза. В обоих указанных ядрах крупные нейросекреторные клетки продуцируют белковые (нонапептидные) нейрогормоны - вазопрессин, или антидиуретический гормон (АДГ), и окситоцин. У человека выработка антидиуретического гормона совершается преимущественно в супраоптическом ядре, тогда как продукция окситоцина преобладает в крупноклеточной части паравентри-кулярных ядер.
Периферическая же часть паравентрикулярного ядра состоит из мелких адренергических нейросекреторных клеток. Аксоны этих клеток направляются в медиальную эминенцию.
В мелкоклеточных ядрах среднего (медиобазального и туберального) гипоталамуса их мелкие адренергические (пептидоадренергические) нейро-секреторные клетки вырабатывают аденогипофизотропные нейрогормоны - рилизинг-гормоны, или факторы (от англ. release - освобождать), с помощью которых гипоталамус контролирует деятельность аденогипофиза. Эти нейро-гормоны по своей природе являются низкомолекулярными олигопептидами и разделяются на либерины, стимулирующие выделение и, вероятно, продукцию гормонов передней и средней долей гипофиза, и статины, угнетающие функции аденогипофиза. Среди либеринов различают: соматотропин-рилизинг-фактор - соматолиберин; тиреотропин-рилизинг-фактор - тиролиберин; АКТГ-рилизинг-фактор - кортиколиберин; рилизинг-фактор фолликулости-
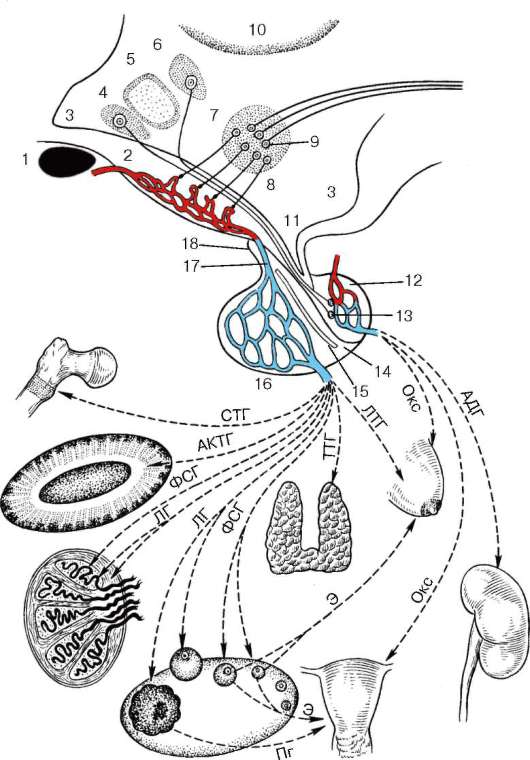
Рис. 15.2. Гипоталамо-гипофизарная система и действие тропных гормонов на органы-мишени (по Б. В. Алешину):
1 - зрительная хиазма; 2 - медиальная эминенция с первичной капиллярной сетью; 3 - полость III желудочка, проекция некоторых гипоталамических ядер на стенку III желудочка; 4 - супраоптическое ядро; 5 - переднее гипоталамическое ядро (преоптическая зона гипоталамуса); 6 - паравентрикулярное ядро; 7 - аркуатно-вентромедиальный комплекс медиобазального гипоталамуса; 8 - таламус; 9 - ней-росекреторные пептидно-адренергические клетки медиобазального гипоталамуса, секретирующие аденогипофизарные гормоны в первичную капиллярную сеть медиальной эминенции (2); 10 - адренергические нейроны медиобазального гипоталамуса, дающие начало нисходящим эфферентным нервным путям (парагипофизарная передача гипоталамических импульсов регулируемым эффекторам); 11 - воронка III желудочка и гипофизарная ножка; 12 - задняя доля гипофиза; 13 - нейросекре-торное тельце Херринга (окончание аксонов нейросекреторных клеток переднего гипоталамуса; 14 - средняя доля гипофиза; 15 - гипофизарная щель; 16 - передняя доля гипофиза с вторичной капиллярной сетью; 17 - портальная (воротная) вена; 18 - туберальная часть аденогипофиза.
Аденогипофизарные гормоны и места их приложения: СТГ - стимулирует рост организма в целом и его отдельных органов (в том числе рост скелета); АКТГ - стимулирует пучковую и сетчатую зоны коры надпочечников; ЛГ - стимулирует овуляцию, образование желтого тела и продукцию последним прогестерона, стимулирует продукцию тестостерона в семеннике; ФСГ - активирует рост фолликулов и выработку ими эстрогенов в яичнике, стимулирует сперматогенез в семеннике; ТТГ - активирует продукцию и секрецию тиреоидного гормона щитовидной железы; ЛТГ - активирует выработку молока в молочных железах. Гормоны, содержащиеся в задней доле гипофиза: окситоцин (Окс) - вызывает сокращение матки и отдачу молока молочными железами; АДГ - стимулирует обратную реабсорбцию воды из первичной мочи в почках (уменьшает диурез) и одновременно повышает артериальное давление; эстрогены яичника (Э) - стимулируют развитие матки и молочных желез
мулирующего гормона - фоллиберин; рилизинг-фактор лютеинизирующего гормона - люлиберин; пролактин-рилизинг-фактор - пролактолиберин; рилизинг-фактор меланоцитостимулирующего гормона - меланолиберин. Среди статинов выделяют: соматотропин-ингибирующий фактор - сомато-статин; пролактинингибирующий фактор - пролактостатин; ингибирующий фактор меланоцитостимулирующего гормона - меланостатин. Важнейшие ядра среднего гипоталамуса локализуются в сером бугре (tuber cinereum) - аркуатное или инфундибулярное (nucl. arcuatus seu infundibularis), дугообразно охватывающее гипофизарную ножку, вентромедиальное (nucl. ventromedialis) и дорсомедиальное (nucl. dorsomedialis). Основным местом выработки гипотала-мических либеринов и статинов оказываются аркуатные и вентромедиальные ядра, а также мелкие пептидоадренергические клетки мелкоклеточной части паравентрикулярного ядра и аналогичные клетки в сером перивентрикуляр-ном веществе, в преоптической зоне гипоталамуса и в супрахиазматическом ядре.
Аксоны мелких пептидоадренергических клеток, равно как и сопровождающих их адренергических нейронов обычного типа, направляются в медиальную эминенцию в составе тубероинфундибулярного дофаминер-гического пучка, берущего начало от дофаминергических нейронов тубе-ральных ядер (рис. 15.2).
Регуляция гипоталамусом периферических эндокринных желез
Влияние гипоталамуса на периферические эндокринные железы осуществляется преимущественно гуморально через секрецию рилизинг-факторов, которые попадают в переднюю долю гипофиза. Такой способ передачи называется трансаденогипофизарным. Кроме того, гипоталамус посылает свои эфферентные импульсы к регулируемым эффекторам прямо по симпатическим или парасимпатическим нервам последних, без опосредования гипофизом, т. е. парагипофизарно.
Парагипофизарная регуляция эндокринных эффекторов может проявляться способностью эндокринных образований непосредственно реагировать (по принципу отрицательной обратной связи) на собственные гормоны, или иммунологические агенты, поступившие в циркуляцию, или на величину эффекта, вызванного ими в организме.
Функции гипоталамуса находятся под контролем головного мозга. В его разных частях находятся нейроэндокринные клетки, которые вырабатывают нейропептиды (более 50). Среди них, например, энкефалин, является интернейрональным медиатором, под воздействием которого по цепочке нейронов происходит выработка нейроаминов - серотонина и норадрена-лина нейронами лимбической системы и норадреналина нейронами ретикулярной формации. Нейроамины влияют на секрецию гипоталамических нейрогормонов. Действие последних стимулирует или тормозит активность аденоцитов гипофиза. Так возникает тесная функциональная связь нервной и эндокринной систем, обеспечивающая контроль, интеграцию и реактивность живых систем.
15.2.2. Гипофиз
Гипофиз, или питуитарная (гороховидная) железа (glandula pituitaria), состоит из аденогипофиза (передняя доля, промежуточная доля, тубераль-ная часть) и нейрогипофиза (задняя доля, стебель, воронка). Аденогипофиз (adenohypophysis) развивается из гипофизарного кармана выстилки верхней части ротовой полости. Гормонпродуцирующие клетки аденогипофиза имеют эпителиальное происхождение (из эпителия ротовой полости). Нейрогипофиз (neurohypophysis) образуется как выпячивание промежуточного пузыря закладки головного мозга (рис. 15.3).
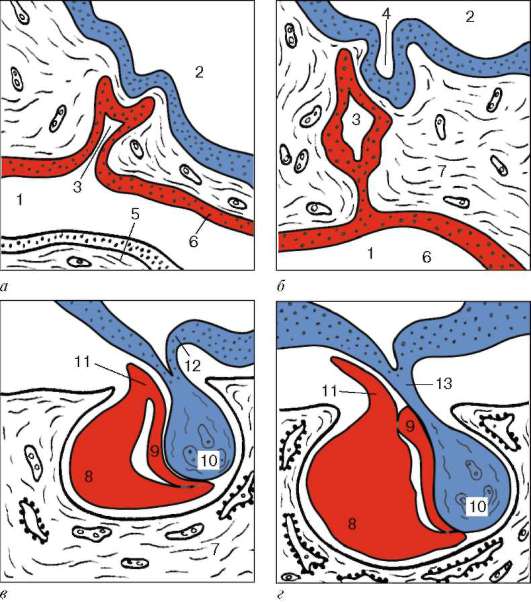
Рис. 15.3. Стадии развития гипофиза (схема):
а - эмбриональные зачатки; б - соединение эктодермального и нейрального эмбриональных зачатков; в, г - формирование основных частей адено- и нейрогипофиза (по О. В. Волковой, с изменениями). 1 - ротовая полость; 2 - полость желудочка мозга; 3 - гипофизарный карман Ратке; 4 - дивертикул промежуточного мозга; 5 - язык; 6 - эктодермальный эпителий ротовой полости; 7 - мезенхима; 8 - передняя стенка кармана Ратке (передняя доля гипофиза); 9 - задняя стенка кармана Ратке (промежуточная доля гипофиза); 10 - задняя доля гипофиза; 11 - туберальная часть; 12 - эпендима; 13 - гипофизарная ножка
Развитие. Закладка гипофиза происходит у зародыша человека на 4- 5-й нед эмбриогенеза как результат взаимодействия двух отдельных зачатков - эпителиального и нейрального. Из эктодермального эпителия, выстилающего ротовую ямку зародыша, выпячивается гипофизарный карман (карман Ратке), направляющийся к основанию формирующегося головного мозга и дающий начало аденогипофизу. Дифференцировка этого эпителиального кармана начинается только после того, как он вступит в соприкосновение с противоположно направленным выпячиванием промежуточного пузыря зачатка головного мозга, которое в дальнейшем окажется воронкой III желудочка. Базальная часть промежуточного пузыря дает начало гипоталамусу.
Дифференцировка эпителиального гипофизарного кармана начинается разрастанием его передней стенки, становящейся передней долей гипофиза и туберальной частью.
Задняя стенка того же кармана становится средней (промежуточной) долей. Между передней и средней долями иногда сохраняется остаток полости гипофизарного кармана в виде гипофизарной щели.
У зародыша человека эта щель облитерирует, и средняя доля гипофиза спаивается с передней. Нейроглия дистального конца воронки, разрастаясь, формирует заднюю долю гипофиза, или нейрогипофиз. Проксимальная же часть воронки, суживаясь, становится гипофизарной ножкой (стебель), связывающей гипоталамус с гипофизом.
В результате дивергентной дифференцировки эпителиоцитов аденогипофи-за возникают клеточные диффероны хромофильных и хромофобных эндокри-ноцитов. Первыми на 9-й нед внутриутробного периода появляются базофиль-ные клетки, а на 4-м мес развития плода - ацидофильные.
К моменту рождения ребенка дифференцировка основных структур гипофиза заканчивается.
Строение. В аденогипофизе различают переднюю долю (lobus anterior), промежуточную часть (pare intermedia) и туберальную часть (pars tuberalis).
Передняя доля образована разветвленными эпителиальными тяжами - трабекулами, формирующими сравнительно густую сеть. Промежутки
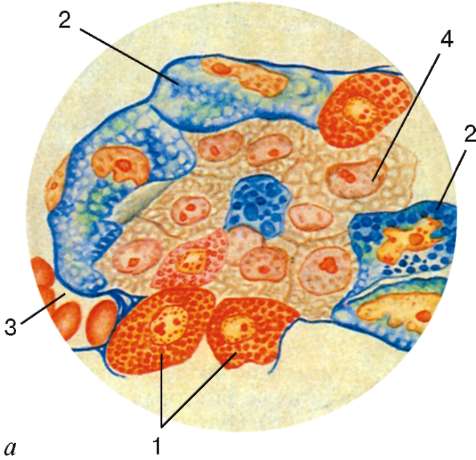
Рис. 15.4. Гипофиз:
а - строение передней доли гипофиза, окраска по Маллори (рисунок Ю. И. Афанасьева): 1 - ацидофильные эндокриноциты; 2 - базофильные эндо-криноциты; 3 - капилляр; 4 - хромофоб-ные эндокриноциты; микрофотографии передней (б) и задней (в) доли гипофиза, окраска азаном: 1 - хромофобные эндо-криноциты; 2 - базофильные эндокри-ноциты; 3 - ацидофильные эндокри-ноциты; 4 - кровеносные капилляры с эритроцитами; 5 - питуициты
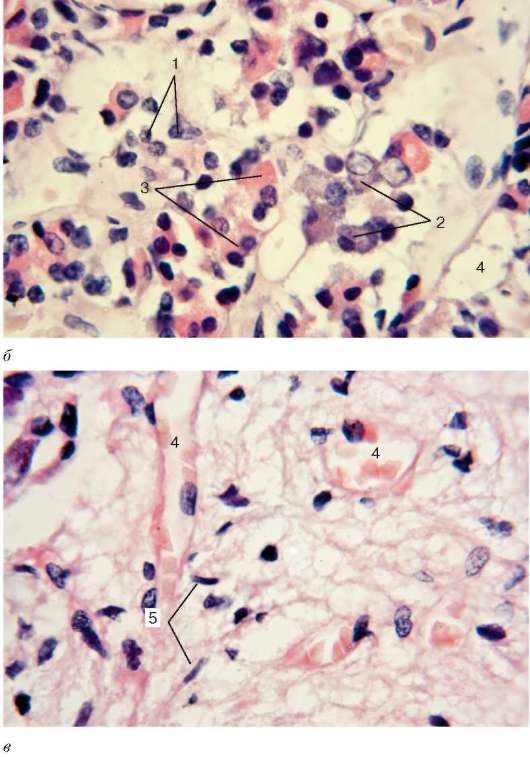
Рис. 15.4. Продолжение
между трабекулами заполнены рыхлой волокнистой соединительной тканью и синусоидными капиллярами, оплетающими трабекулы. Каждая трабекула образована железистыми клетками - эндокриноцитами разных линий дифференцировки (рис. 15.4).
Одни из них, располагающиеся по периферии трабекул, содержат в своей цитоплазме секреторные гранулы, которые интенсивно воспринимают красители. В связи с этим такие клетки именуются хромофильными эндокриноцитами (endocrinocytus chromophilus). Другие же клетки, занимающие середину трабекулы, имеют нечеткие границы, и их цитоплазма окрашивается слабо - хромофобные эндокриноциты (endocrinocytus chromophobus).
Хромофильные эндокриноциты подразделяются на базофильные и ацидофильные по окрашиваемости их секреторных гранул. Базофильные эндо-криноциты (endocrinocytus basophilus) получили такое название потому, что их гранулы окрашиваются основными красителями. Гранулы содержат гликопротеиды, являющиеся материалом для биосинтеза гормонов, продуцируемых этими аденоцитами. Относительное количество этих клеток в норме составляет от 4 до 10 % общего числа эндокриноцитов передней доли. По размерам они сравнительно крупные. Среди них различают две разновидности. Одни из них характеризуются круглой или овальной формой и эксцентрическим положением ядер. Секреторные гранулы диаметром 200-300 нм. Количество таких клеток (базофилов) возрастает во время усиленной продукции гонадотропных гормонов (гонадотропинов), в связи с чем клетки данной разновидности называются гонадотропными эндокри-ноцитами (endocrinocytus gonadotropicus), или гонадотропоцитами (рис. 15.5). Предполагается, что одни из них вырабатывают фолликулостимулирующий гормон (фоллитропин), а другим приписывается образование лютеинизи-рующего гормона (лютропина).
Фоллитропин влияет на формирование половых клеток, лютропин стимулирует образование желтого тела в яичнике и выработку мужского полового гормона интерстициальными клетками яичка.
При недостаточности в организме половых гормонов в передней доле гипофиза по механизму отрицательной обратной связи усиливается выработка гонадотропных гормонов (особенно фоллитропина), в связи с чем в некоторых гипертрофирующихся гонадотропоцитах развивается крупная вакуоль, растягивающая цитоплазму наподобие кольца и оттесняющая уплотняющееся ядро на периферию клетки (см. рис. 15.5, VII). Такие трансформированные гонадотропоциты получили наименование клеток кастрации.
Вторая разновидность базофильных клеток отличается неправильной или угловатой формой (см. рис. 15.5, II). Их секреторные гранулы очень мелкие (диаметром 80-150 нм) и интенсивно окрашиваются альдегидфук-сином. Они содержат меньше гликопротеинов, чем гонадотропоциты. Эти клетки вырабатывают тиротропный гормон - тиротропин, стимулирующий функцию фолликулярных эндокриноцитов щитовидной железы, и называются тиротропными эндокриноцитами (endocrinocytus thyrotropicus), или тиротропоцитами.
Если в организме возникает недостаточность тиреоидных гормонов - йодотиронинов, то происходит перестройка тиротропоцитов. Продукция тиротропина усиливается, вследствие чего некоторые тиротропоциты значительно увеличиваются в объеме, цистерны эндоплазматической сети расширяются, цитоплазма приобретает вид крупноячеистой сети. В этих цистернах обнаруживаются альдегидофуксинофильные гранулы, более крупные, чем в исходных тиротропоцитах.
Такие вакуолизированные тиротропоциты называются клетками тироид-эктомии (см. рис. 15.5, VI).
Для ацидофильных эндокриноцитов (endocrinocytus acidophilus) характерны крупные плотные белковые гранулы, воспринимающие кислые красители
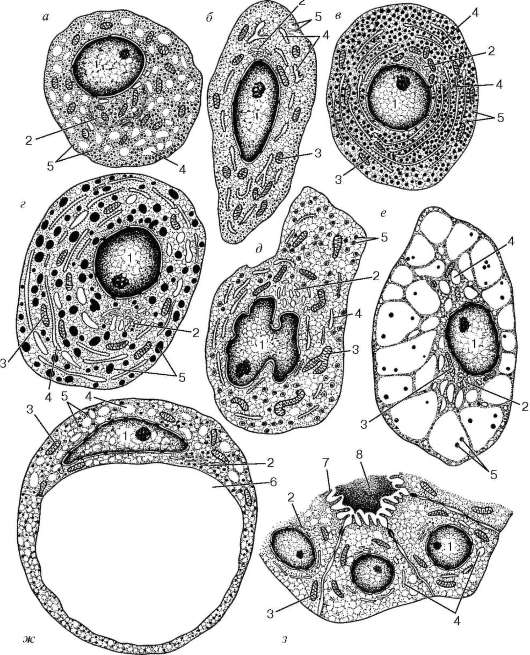
Рис. 15.5. Ультрамикроскопическое строение железистых клеток аденогипофиза (по Б. В. Алешину):
а - гонадотропная фолликулостимулирующая клетка; б - тиротропная клетка; в - соматотропная клетка; г - лактотропная (маммотропная) клетка; д - кор-тикотропная клетка; е - клетка тиреоидэктомии; ж - клетка кастрации; з - фолликулярно-звездчатые клетки псевдофолликула. 1 - ядро; 2 - комплекс Гольджи; 3 - митохондрия; 4 - эндоплазматическая сеть; 5 - секреторные гранулы; 6 - вакуоль; 7 - микроворсинка; 8 - полость псевдофолликула, заполненная кол-лоидоподобной массой
(см. рис. 15.5, III). По размерам эти клетки несколько меньше базофильных, но их количество достигает 30-35 % всех аденоцитов передней доли гипофиза. Форма их округлая или овальная. Ядра располагаются в центре клетки. Сильно развита гранулярная эндоплазматическая сеть.
Ацидофильные эндокриноциты тоже представлены двумя разновидностями. Одни - соматотропные эндокриноциты (endocrinocytus somatotropicus), или соматотропоциты, вырабатывают гормон роста, или соматотропин, регулирующий рост организма; другие - пролактиновые (маммотроп-ные) эндокриноциты, или пролактиноциты (endocrinocytus mammotropicus seu prolactinocyti), продуцируют пролактин (лактотропный гормон). Основное значение пролактина - активирование биосинтеза молока в молочной железе. Продукция этого гормона усиливается у рожениц после родов, во время лактации и вскармливания новорожденного. Кроме того, пролактин удлиняет функционирование желтого тела в яичнике, в связи с чем ранее его иногда называли лютеотропным гормоном.
В соматотропоцитах секреторные гранулы имеют шаровидную форму и достигают в диаметре 350-400 нм. Пролактиноциты отличаются еще более крупными гранулами овальной или удлиненной формы (500-600 нм в длину и 100-120 нм в ширину).
Еще одна группа хромофильных клеток - кортикотропные эндокрино-циты (endocrinocytus corticotropicus), или кортикотропоциты, локализующиеся преимущественно в центральной зоне передней доли гипофиза, продуцируют белковый адренокортикотропный гормон (АКТГ, или кортикотропин), стимулирующий секреторную активность клеток пучковой зоны коркового вещества надпочечников. Их форма неправильная или угловатая, клеточные ядра дольчатые, эндоплазматическая сеть хорошо выражена. Секреторные гранулы имеют строение пузырьков, окруженных мембраной, и содержат внутри плотную белковую сердцевину, причем между мембраной и сердцевиной остается светлое пространство.
Цитоплазма хромофобных эндокриноцитов слабо воспринимает красители и не содержит отчетливых секреторных гранул. Хромофобные клетки составляют около 60 %. Группа хромофобов включает клетки разной степени диф-ференцировки и различного физиологического значения. Среди них можно обнаружить клетки, уже начавшие специализироваться в базофильные или ацидофильные клетки, но еще не успевшие накопить специфические секреторные гранулы. Другие же хромофобные клетки, наоборот, являются вполне специализированными, но лишившимися своих базофильных или ацидофильных секреторных гранул вследствие интенсивной или длительной секреции. К ряду неспециализированных принадлежит лишь небольшая часть хромофобных клеток, которые можно признать резервными. Наконец, среди хромофобов встречаются фолликулозвездчатые (звездчатые) клетки, небольшие по размерам, но обладающие длинными ветвистыми отростками, которыми они соединяются в широкопетлистую сеть. Некоторые из отростков проходят между соседними эндокриноцитами и заканчиваются на синусоидных капиллярах. Иногда звездчатые клетки группируются в небольшие фолликулоподобные образования, в полостях которых накапли-
вается гликопротеиновый секрет. На апикальной поверхности таких звездчатых клеток развиваются микроворсинки, вдающиеся в просвет фолликула (см. рис. 15.5, VIII).
Средняя (промежуточная) часть аденогипофиза представлена узкой полоской эпителия. Эндокриноциты средней доли способны вырабатывать белковый или слизистый секрет, который, накапливаясь между соседними клетками, приводит к формированию в средней доле фолликулоподобных кист. От задней доли эпителий средней доли отделяется тонкой прослойкой рыхлой соединительной ткани.
В средней части аденогипофиза вырабатывается меланоцитостимулиру-ющий гормон (меланоцитотропин), а также липотропин - гормон, усиливающий метаболизм липидов.
Туберальная часть аденогипофиза - отдел, прилежащий к гипофизарной ножке и соприкасающийся с нижней поверхностью медиального возвышения гипоталамуса. Туберальная часть образована эпителиальными тяжами, состоящими из кубических клеток с умеренно базофильной цитоплазмой. От туберальных тяжей, так же как от эпителия средней доли, отходят в переднюю долю трабекулы; в некоторых клетках туберальных тяжей встречаются базофильные гранулы, хотя в небольшом количестве.
Гипоталамо-аденогипофизарное кровоснабжение и его значение в гипо-таламической регуляции аденогипофизарного гормонопоэза (см. рис. 15.2). Система гипоталамо-аденогипофизарного кровоснабжения называется портальной (воротной). Приносящие гипофизарные артерии вступают в медиальное возвышение медиобазального гипоталамуса, где разветвляются в сеть капилляров (первичное капиллярное сплетение портальной системы). Эти капилляры образуют петли и клубочки, с которыми контактируют терминали аксонов нейросекреторных клеток аденогипофизотропной зоны гипоталамуса. Капилляры первичного сплетения собираются в портальные вены, идущие вдоль гипофизарной ножки в переднюю долю, где они распадаются на капилляры синусоидного типа (вторичная капиллярная сеть), разветвляющиеся между трабекулами паренхимы железы. Наконец, синусоиды вторичной капиллярной сети собираются в выносящие вены, по которым кровь, обогатившаяся гормонами передней доли, поступает в общую циркуляцию.
Задняя доля гипофиза, или нейрогипофиз. Задняя доля гипофиза образована в основном клетками эпендимы. Они имеют отростчатую или верете-новидную форму и называются питуицитамы. Их многочисленные тонкие отростки заканчиваются в адвентиции кровеносных сосудов или на базаль-ной мембране капилляров.
В задней доле гипофиза аккумулируются антидиуретический гормон (вазопрессин) и окситоцин, вырабатываемые крупными пептидохо-линергическими нейросекреторными клетками переднего гипоталамуса. Вазопрессин увеличивает реабсорбцию в канальцах почки, окситоцин стимулирует сокращение мускулатуры матки. Аксоны этих нейросекре-торных клеток собираются в гипоталамо-нейрогипофизарные пучки, входят в заднюю долю гипофиза, где заканчиваются крупными терминалями
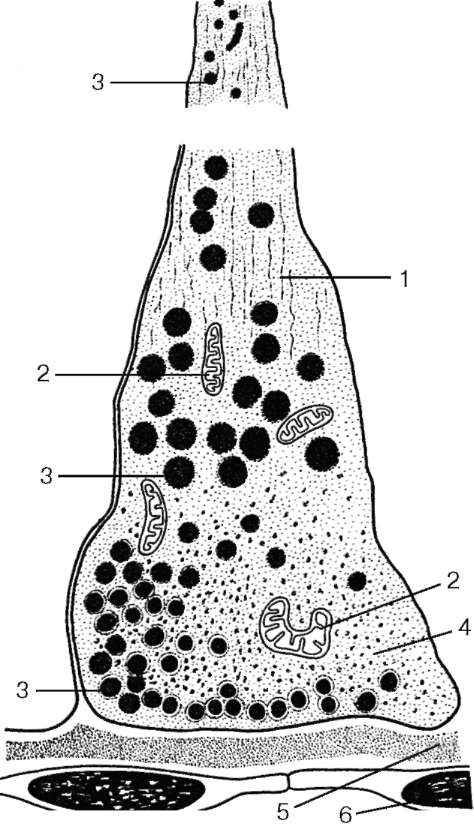
Рис. 15.6. Контакт терминали аксона гипоталамического секреторного нейрона с гемокапилляром задней доли гипофиза (по Гершенфельду): 1 - нейрофиламенты; 2 - митохондрии; 3 - гранулы нейросекрета; 4 - цитоплазма; 5 - базальная мембрана; 6 - эндотелиоциты капилляра
(называемыми тельцами Херринга, или нейросекреторными тельцами) (рис. 15.6). Последние формируют нейрососудистые (нейрогемальные) синапсы, посредством которых ней-росекрет поступает в кровь.
Возрастные изменения. В постна-тальном периоде преимущественно активируются ацидофильные эндокриноциты (очевидно, в связи с обеспечением повышенной продукции соматотропина, стимулирующего быстрый рост тела ребенка), а среди базофилов преобладают тиротропо-циты. В пубертатном периоде, когда наступает половое созревание, увеличивается количество базофиль-ных аденоцитов.
Регенерация. Аденогипофиз обладает ограниченной регенераторной способностью, главным образом за счет дифференциации хромофобных клеток. Задняя доля гипофиза, образованная нейроглией, регенерирует лучше.
15.2.3. Эпифиз
Эпифиз - верхний мозговой придаток (epiphysis cerebri), или шишковидное тело (corpus pineale).
Эпифиз участвует в регуляции процессов, протекающих в организме ритмически или циклически, например овариально-менструального цикла. Ритмические колебания других периодических функций, интенсивность которых закономерно изменяется на протяжении суток, называются циркадными (от лат. circa diem - около дня). Циркадные ритмы явно связаны со сменой дня и ночи (светового и темнового периодов), и их зависимость от эпифиза свидетельствует о том, что гормонообразовательная деятельность последнего определяется его способностью различать смену световых раздражений, получаемых организмом.
Развитие. У зародыша человека эпифиз развивается как выпячивание крыши III желудочка промежуточного мозга на 5-6-й нед развития. В его состав включается субкомиссуральный орган, который развивается из эпендимы III желудочка мозга. У человека и млекопитающих он сильно реду-
цирован (0,2 г). В результате дивергентной дифференцировки нейральных стволовых клеток развиваются два клеточных дифферона - пинеалоцитар-ный и глиоцитарный. Максимального развития эпифиз достигает у детей до 7 лет.
Строение. Снаружи эпифиз окружен тонкой соединительнотканной капсулой, от которой отходят разветвляющиеся перегородки внутрь железы, образующие ее строму и разделяющие ее паренхиму на дольки (рис. 15.7), особенно в пожилом возрасте.
В паренхиме железы различаются секретообразующие пинеалоциты (endocrinocytus pinealis) и опорные глиальные клетки (астроциты). Пинеалоциты располагаются в центральной части долек. Они несколько крупнее опорных клеток, по форме многоугольные, имеют пузыревидные ядра с крупными ядрышками. От тела пинеалоцита отходят длинные отростки, ветвящиеся наподобие дендритов, которые переплетаются с отростками астроцитов. Отростки, булавовидно расширяясь, направляются к капиллярам и контактируют с ними. В цитоплазме этих булавовидных расширений содержатся осмиофильные гранулы, вакуоли и митохондрии.
Среди пинеалоцитов различают светлые пинеалоциты (endocrinocytus lucidus), характеризующиеся светлой гомогенной цитоплазмой, и темные пинеалоциты (endocrinocytus densus) меньшего размера с ацидофильными (а иногда базофильными) включениями в цитоплазме. Обе названные формы представляют собой клетки, находящиеся в различных функциональных состояниях, или клетки, подвергающиеся возрастным изменениям. В цитоплазме пинеалоцитов обнаруживаются многочисленные митохондрии, хорошо развитый комплекс Гольджи, лизосомы, пузырьки агранулярной эндо-плазматической сети, рибосомы и полисомы.
Астроциты преобладают на периферии долек. Их цитоплазма скудна, ядра уплотнены. Длинные отростки направляются к междольковым соединительнотканным перегородкам, образуя своего рода краевую кайму дольки.
Эпифиз вырабатывает антигипоталамические факторы (антигормоны), оказывающие действие на гипофиззависимые эндокринные органы. Действие это обратное (тормозящее, ингибирующее) тропным гормонам аденогипофиза. Большое значение имеет выработка пинеалоцитами анти-гонадотропина, который тормозит секрецию лютропина в аденогипофизе, т. е. играет роль гонадостатина. Антигонадотропин эпифиза и гонадолибе-рин гипоталамуса, действуя как гормоны-антагонисты, совместно осуществляют регуляцию гонадотропной функции гипофиза.
Число регуляторных пептидов, продуцируемых пинеалоцитами, приближается к 40. Из них наиболее важны аргинин-вазотоцин, тиролибе-рин, люлиберин, тиротропин и др. Образование олигопептидных гормонов совместно с нейроаминами (серотонин и мелатонин) демонстрирует принадлежность пинеалоцитов к APUD-серии клеток (см. ниже).
Кровоснабжается эпифиз ветвями средней и задней мозговых артерий.
Иннервируется эпифиз ЦНС и симпатической нервной системой.
Возрастные изменения. У человека эпифиз достигает максимального развития к 5-6 годам жизни, после чего, несмотря на продолжающееся функ-
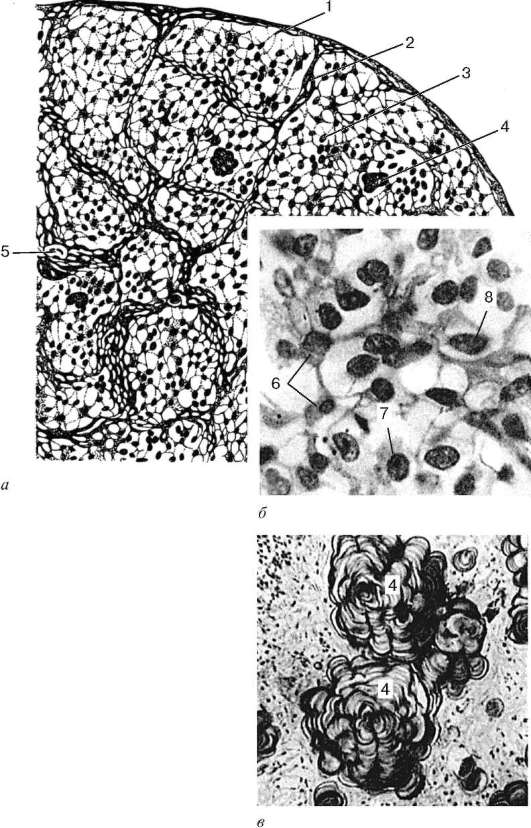
Рис. 15.7. Эпифиз:
а - общий вид эпифиза человека (схема); б - микрофотография паренхимы эпифиза ребенка 1 года 2 мес (по Л. И. Салийчук); в - микрофотография эпифиза человека 63 лет (по Л. И. Салийчук): 1 - соединительнотканная капсула; 2 - соединительнотканная перегородка; 3 - паренхима эпифиза; 4 - мозговой «песок»; 5 - кровеносные сосуды; 6 - глиоциты; 7 - темные пинеалоциты; 8 - светлые пинеалоциты
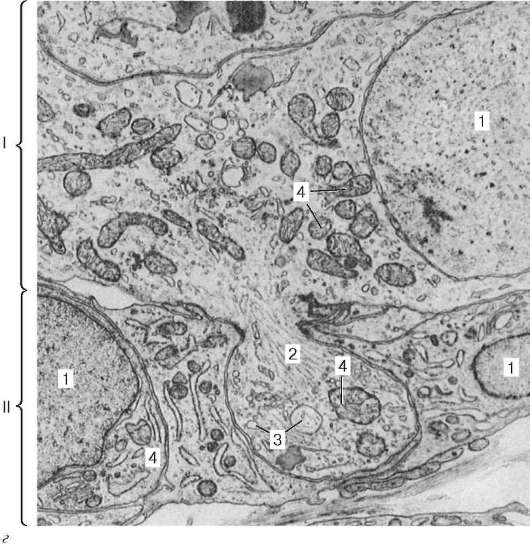
Рис. 15.7. Продолжение
г - эпифиз крысы. Электронная микрофотография, увеличение 12 000 (по Г. А. Косолапову): I - светлый секретирующий пинеалоцит; II - глиоцит; 1 - ядро; 2 - булавовидный отросток; 3 - вакуоли; 4 - митохондрии
ционирование, начинается его возрастная инволюция. Некоторое количество пинеалоцитов претерпевают атрофию, а строма разрастается, и в ней увеличивается отложение фосфатных и карбонатных солей в виде слоистых шариков, называемых мозговым песком (ацервулюс).
15.3. ПЕРИФЕРИЧЕСКИЕ ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Бранхиогенная группа эндокринных желез развивается из зачатков жаберных карманов и включает щитовидную и околощитовидные железы. Из зачатков жаберных карманов развивается также тимус. Щитовидная железа и околощитовидные железы связаны не только общим источником развития, но и функционально, играя главную роль в поддержании метаболического статуса и постоянства состава внутренней среды организма. Гормоны этих желез (тироксин, кальцитонин, паратгормон) регулируют скорость основного обмена и концентрацию кальция в крови.
Щитовидная железа (glandula thyroidea) содержит два клеточных диф-ферона эндокриноцитов, развивающихся из стволовых клеток различного происхождения: Т-тироциты (фолликулярные клетки), наиболее многочисленные, вырабатывающие йодсодержащие гормоны - тироксин (Т4) и трий-одтиронин (Т3), и С-тироциты (парафолликулярные клетки), вырабатывающие гормон тирокальцитонин и ряд других пептидов.
Тироксин и трийодтиронин являются мощными стимуляторами окислительных процессов в клетках, причем трийодтиронин в 5-10 раз активнее тироксина. Эти гормоны усиливают обмен веществ, синтез белков, газообмен, обмен углеводов и жиров. Тиреоидные гормоны оказывают значительное влияние на развитие, рост и дифференцировку клеток и тканей. Они ускоряют развитие костной ткани. Особенно большое влияние гормоны щитовидной железы оказывают на гистогенез нервной ткани. При недостаточности щитовидной железы тормозится дифференци-ровка клеток и тканей головного мозга, нарушается психическое развитие человека. Тиреоидные гормоны оказывают стимулирующее действие на регенерационные процессы в тканях. Для нормальной деятельности щитовидной железы необходимо поступление йода с питьевой водой и пищей в организм. Не содержащий йода третий гормон щитовидной железы - тирокальцитонин - участвует в регуляции кальциевого и фосфорного обмена.
Развитие. Зачаток щитовидной железы возникает у зародыша человека на 3-4-й нед как выпячивание стенки глотки между I и II парами жаберных карманов, которое растет вдоль глоточной кишки в виде эпителиального тяжа. На уровне III-IV пары жаберных карманов этот тяж раздваивается, давая начало формирующимся правой и левой долям щитовидной железы. Начальный эпителиальный тяж (ductus thyreoglossus) атрофируется, и от него сохраняются только перешеек, связывающий обе доли щитовидной железы, и проксимальная часть в виде ямки (foramen coecum) в корне языка. У большинства других млекопитающих атрофируется также дистальный конец эпителиального тяжа, поэтому перешеек не развивается и обе доли щитовидной железы обособляются. Зачатки долей быстро разрастаются, образуя рыхлые сети ветвящихся эпителиальных трабекул, клетки которых дифференцируются в Т-тироциты. Последние объединяются в фолликулы, в промежутки между которыми врастает мезенхима с кровеносными сосудами и нервами. По мере развития в зачаток железы врастают также производные V пары жаберных карманов - клетки так называемых ультимобранхиаль-ных телец. Это С-тироциты, которые имеют нейроэктодермальную природу, и в закладку щитовидной железы они внедряются через ультимобранхиаль-ные зачатки.
Строение. Щитовидная железа окружена соединительнотканной капсулой, прослойки которой направляются вглубь и разделяют орган на дольки, в которых располагаются многочисленные сосуды микроциркуляторного русла и нервы. Основными структурными компонентами паренхимы железы являются фолликулы - замкнутые шаровидные или слегка вытянутые образования варьирующих размеров с полостью внутри, выстланные одним
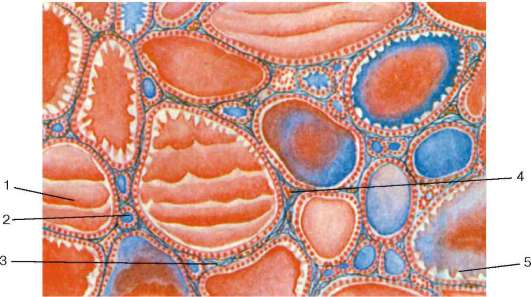
Рис. 15.8. Щитовидная железа (по Б. В. Алешину):
1 - интрафолликулярный коллоид; 2 - микрофолликул; 3 - межфолликулярные соединительнотканные прослойки; 4 - капилляры; 5 - резорбционные вакуоли
слоем эпителиальных клеток, представленных Т-тироцитами (фолликулярными клетками), а также С-тироцитами (парафолликулярными клетками) нейрального происхождения (рис. 15.8).
В дольке железы различают фолликулярные комплексы (микродольки), которые состоят из группы фолликулов, окруженных тонкой соединительнотканной капсулой. В просвете фолликулов накапливается коллоид - секреторный продукт Т-тироцитов, представляющий собой вязкую жидкость, состоящую в основном из тироглобулина. Размер фолликулов и образующих их стенку Т-тироцитов варьирует в нормальных физиологических условиях. Их диаметр от 0,02 до 0,9 мм. В небольших формирующихся фолликулах, еще не заполненных коллоидом, эпителий однослойный призматический. По мере накопления коллоида размеры фолликулов увеличиваются, эпителий становится кубическим, а в сильно растянутых фолликулах, заполненных коллоидом, - плоским. Основная масса фолликулов в норме выстлана Т-тироцитами кубической формы. Увеличение размеров фолликулов обусловлено пролиферацией, ростом и дифферен-цировкой Т-тироцитов, сопровождаемой накоплением коллоида в полости фолликула.
Фолликулы разделяются тонкими прослойками рыхлой волокнистой соединительной ткани с многочисленными кровеносными и лимфатическими капиллярами, оплетающими фолликулы, тучными клетками и лимфоцитами (рис. 15.9).
Т-тироциты, или фолликулярные эндокриноциты (endocrinocytus follicularis), - железистые клетки, составляющие большую часть стенки фолликулов. В фолликулах Т-тироциты образуют выстилку (стенку) и располагаются в один слой на базальной мембране. При умеренной функциональной
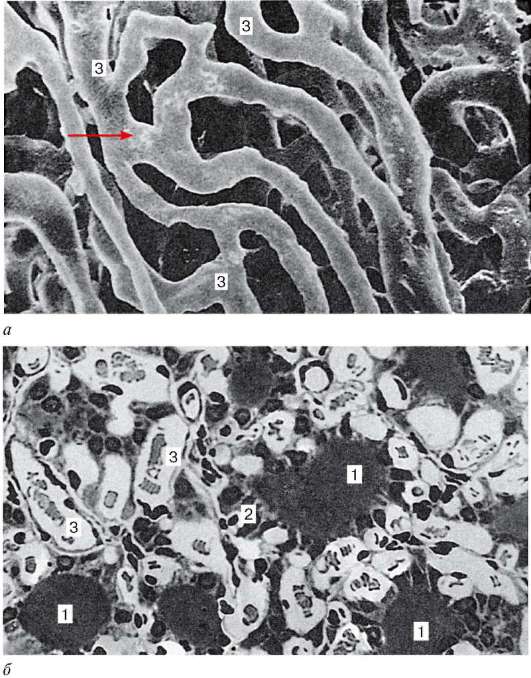
Рис. 15.9. Микроциркуляторное русло щитовидной железы:
а - коррозионный препарат капилляров (стрелка) фолликулов щитовидной железы; б - микрофотография полутонкого среза щитовидной железы (по Н. П. Федченко). 1 - полость фолликула; 2 - Т-тироциты стенки фолликула; 3 - гемокапилляры
активности щитовидной железы (ее нормальной функции) Т-тироциты имеют кубическую форму и шаровидные ядра. Коллоид, секретируемый ими, заполняет в виде гомогенной массы просвет фолликула. На апикальной поверхности Т-тироцитов, обращенной к просвету фолликула, имеются микроворсинки. По мере усиления тиреоидной активности количество и размеры микроворсинок возрастают. Одновременно базальная поверхность Т-тироцитов, почти гладкая в период функционального покоя щитовидной железы, становится складчатой, что увеличивает соприкосновение Т-тироцитов с перифолликулярными пространствами. Соседние клетки в выстилке фолликулов тесно связаны между собой многочисленными дес-мосомами и хорошо развитыми терминальными пластинками. По мере воз-
растания тиреоидной активности на боковых поверхностях Т-тироцитов возникают пальцевидные выступы (интердигитации), входящие в соответствующие вдавления боковой поверхности соседних клеток.
В Т-тироцитах хорошо развиты органеллы, особенно участвующие в белковом синтезе.
Белковые продукты, синтезируемые Т-тироцитами, выделяются в полость фолликула, где завершается образование йодированных тирозинов и тиро-нинов (аминокислот, входящих в состав крупной и сложной молекулы тиро-глобулина). Тиреоидные гормоны могут попасть в циркуляцию лишь после высвобождения из этой молекулы (т. е. после расщепления тироглобулина). Когда потребности организма в тиреоидном гормоне возрастают и функциональная активность щитовидной железы усиливается (гиперфункция щитовидной железы), Т-тироциты фолликулов принимают призматическую форму. Интрафолликулярный коллоид при этом становится более жидким и содержит многочисленные ресорбционные вакуоли. Ослабление функциональной активности (гипофункция щитовидной железы) проявляется, наоборот, уплотнением коллоида, его застоем внутри фолликулов, диаметр и объем которых значительно увеличиваются; высота же Т-тироцитов уменьшается, они принимают уплощенную форму, а их ядра вытягиваются параллельно поверхности фолликула.
Секреторный цикл. В секреторном цикле различают основные фазы: фазу продукции и фазу выведения гормонов (рис. 15.10).
Фаза продукции включает: 1) поступление предшественников тирогло-булина (аминокислот, углеводов, ионов, воды, йодидов), приносимых из кровеносного русла в Т-тироциты; 2) синтез полипептидных цепочек тиро-глобулина в гранулярной эндоплазматической сети и их гликозилирование (соединение с нейтральными сахарами и сиаловой кислотой) с помощью фермента тиропероксидазы в комплексе Гольджи; синтез тиропероксидазы, окисляющей йодиды и обеспечивающей их соединение с тироглобулином на поверхности Т-тироцитов и образование коллоида (йодирование тиро-глобулина). При этом к нейодированному тироглобулину присоединяется сначала один атом йода, а затем и второй, в результате чего образуются моно- и дийодтиронины. Последующая их комплексация дает трийодтиро-нин и тетрайодтиронин (тироксин).
Фаза выведения включает эндоцитоз тиреоидных гормонов, связанных с тироглобулином, которые подвергаются гидролизу с помощью лизосомных протеаз. Тироглобулин расщепляется до аминокислот, а монойодтирозин, дийодтирозин, трийодтиронин (Т3) и тетрайодтиронин (Т4) освобождаются в цитоплазму. Два последних выводятся через базальную мембрану в капилляры и лимфатические капилляры, а монойодтирозин и дийодтиро-зин используются для синтеза новой молекулы тироглобулина.
Тиротропный гормон усиливает функцию щитовидной железы, стимулируя поглощение тироглобулина микроворсинками Т-тироцитов, а также его расщепление в фаголизосомах с высвобождением активных гормонов.
С-тироциты, или парафолликулярные эндокриноциты (endocrinocytus parafollicularis). Во взрослом организме парафолликулярные эндокринные
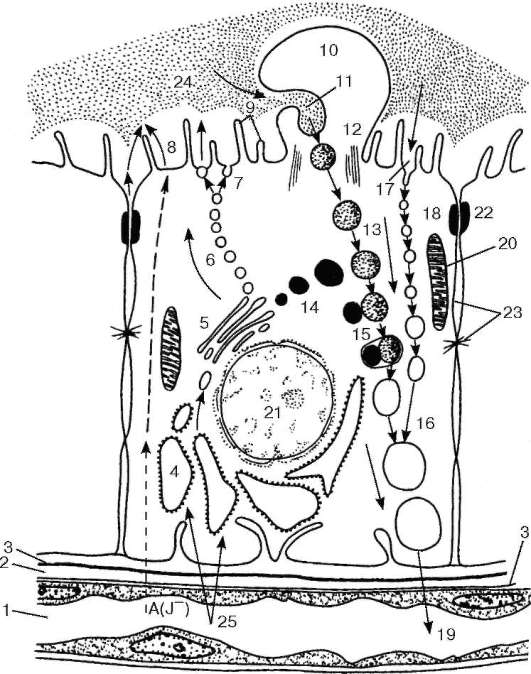
Рис. 15.10. Т-тироцит и процесс секреции (по Б. В. Алешину):
1 - капилляр; 2 - перикапиллярное пространство; 3 - базальные мембраны Т-тироцита и эндотелиоцита (стрелками показано направление транспортировки перерабатываемых веществ); 4 - гранулярная эндоплазматическая сеть; 5 - комплекс Гольджи; 6 - перемещение пузырьков секрета гликопротеида к апикальной плазмолемме; 7 - экзоцитоз в полость фолликула; 8 - поступление атомарного йода в полость фолликула и йодирование гликопротеида; 9 - микроворсинки; 10 - образование псевдоподий в процессе резорбции коллоида; 11 - фагоцитоз интра-фолликулярного коллоида; 12 - микротубулы псевдоподии; 13 - капли фагоцитированного интрафолликулярного коллоида (интрацеллюлярные капли коллоида); 14 - лизосомы; 15 - слияние коллоидной капли с лизосомой; протеолиз тирогло-булина и высвобождение тиреоидного гормона; 16 - вакуоли с продуктами протео-лиза; 17 - протеолиз интрафолликулярного коллоида на апикальной поверхности Т-тироцита и эндоцитоз (пиноцитоз) продуктов этого протеолиза; 18 - транспорт пиноцитозных пузырьков; 19 - отдача тиреоидного гормона в капилляр; 20 - митохондрии; 21 - ядро Т-тироцита; 22 - замыкающая пластинка; 23 - межклеточная щель с десмосомами; 24 - интрафолликулярный коллоид; 25 - поступление веществ из капилляра в Т-тироцит
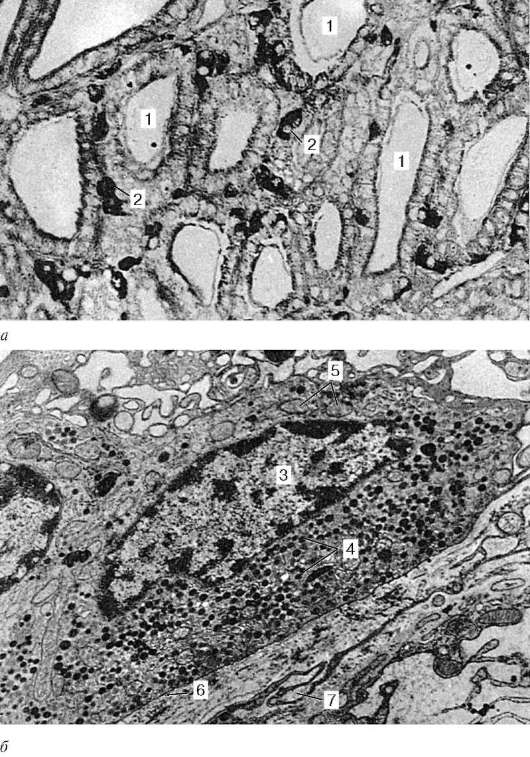
Рис. 15.11. Парафолликулярные эндокриноциты в щитовидной железе крысы: а - микрофотография. Импрегнация нитратом серебра (по Н. П. Федченко); б - электронная микрофотография, увеличение 9000. 1 - полость фолликула щитовидной железы; 2 - парафолликулярные эндокриноциты; 3 - ядро; 4 - секреторные гранулы; 5 - митохондрии; 6 - базальная мембрана; 7 - соединительнотканная оболочка микродольки
клетки локализуются в стенке фолликулов, залегая между основаниями соседних Т-тироцитов, но не достигают своей верхушкой просвета фолликула (интраэпителиальная локализация парафолликулярных клеток). Кроме того, парафолликулярные клетки располагаются также в межфолликулярных прослойках соединительной ткани (рис. 15.11). По размерам
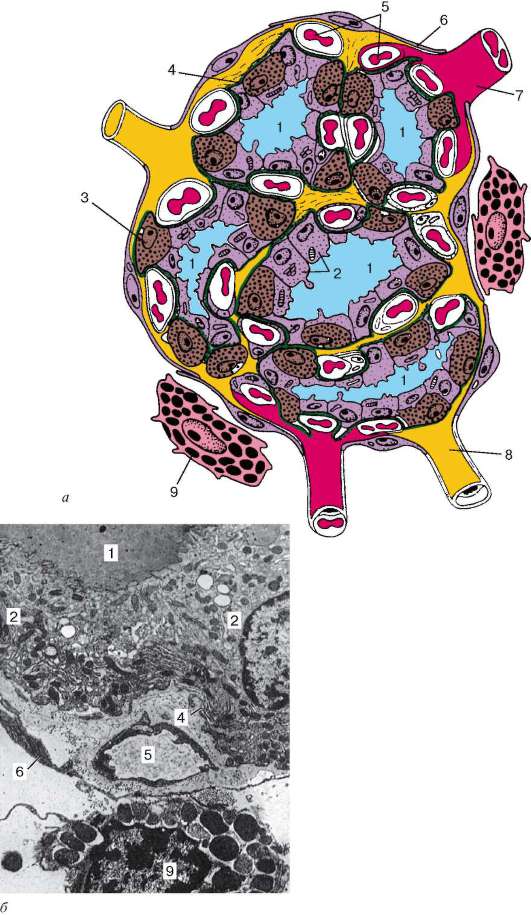
Рис. 15.12. Строение микродольки щитовидной железы (по Н. П. Федченко): а - схема; б - электронная микрофотография, увеличение 10 000. 1 - полость фолликула; 2 - фолликулярные эндокриноциты (Т-тироциты); 3 - пара-фолликулярные эндокриноциты (С-тироциты, кальцитониноциты); 4 - базальная мембрана 5 - гемокапилляр; 6 - соединительнотканная оболочка микродольки; 7 - артериола; 8 - лимфатический капилляр; 9 - тучная клетка
парафолликулярные клетки крупнее Т-тироцитов, имеют округлую, иногда угловатую форму. В отличие от Т-тироцитов парафолликулярные клетки не поглощают йод, но совмещают образование нейроаминов (норадреналина и серотонина) путем декарбоксилирования тирозина и 5-гидрокситриптофана (ароматических аминокислот - предшественников указанных нейроаминов) с биосинтезом белковых (олигопептидных) гормонов - кальцитонина и соматостатина.
Секреторные гранулы, густо заполняющие цитоплазму парафоллику-лярных клеток, обнаруживают сильную осмиофилию и аргирофилию. В цитоплазме парафолликулярных клеток хорошо развиты гранулярная эндоплазматическая сеть и комплекс Гольджи. Секреторные гранулы пара-фолликулярных клеток бывают двух типов. В некоторых парафоллику-лярных клетках преобладают мелкие, но сильно осмиофильные гранулы. Клетки данного типа вырабатывают кальцитонин. В парафолликулярных клетках другого типа содержатся более крупные, но слабо осмиофильные гранулы. Эти клетки продуцируют соматостатин (рис. 15.12).
Васкуляризация. Щитовидная железа обильно снабжается кровью. За единицу времени через щитовидную железу проходит приблизительно столько же крови, сколько через почки, причем интенсивность кровоснабжения значительно увеличивается при усилении функциональной активности органа.
Иннервация. В щитовидной железе много симпатических и парасимпатических нервных волокон. Стимуляция адренергических нервных волокон приводит к небольшому усилению, а парасимпатических - к угнетению функции Т-тироцитов. Основную регулирующую роль играет тиротропный гормон. Парафолликулярные клетки невосприимчивы к гормону гипофиза, но реагируют на симпатические (активирующие) и парасимпатические угнетающие импульсы.
Реактивность и регенерация. Интоксикации, травмы, аутоиммунные процессы, наследственные факторы и другое могут приводить к тиреотоксикозу или к гипотиреозу. Следует иметь в виду, что для нормального функционирования щитовидной железы необходимо достаточное поступление в организм йода. Щитовидная железа обладает высокой способностью к регенерации после травм. Источником роста тиреоидной паренхимы оказывается эпителий фолликулов. Деление Т-тироцитов приводит к увеличению площади сечения фолликула, вследствие чего в нем возникают складки, выступы и сосочки, вдающиеся в полость фолликулов (интрафолликуляр-ная регенерация).
Размножение клеток может привести также к появлению эпителиальных почек, оттесняющих базальную мембрану кнаружи, в межфолликулярное пространство. С течением времени в пролиферирующих Т-тироцитах этих скоплений возобновляется биосинтез тироглобулина, что приводит к образованию микрофолликулов. Микрофолликулы в результате продолжающегося синтеза и накопления коллоида в их полостях увеличиваются в размерах и становятся такими же, как материнские (экстрафолликулярная регенерация). Парафолликулярные клетки в фолликулогенезе участия не принимают.
Примером аутоиммунного заболевания щитовидной железы является болезнь Хашимото. Она возникает вследствие того, что тироглобулин проникает в строму щитовидной железы и, являясь антигеном, вызывает иммунную реакцию организма. При этом сильно разрастается строма железы, инфильтрируется лимфоцитами и плазмоцитами, а фолликулы щитовидной железы содержат мало коллоида и постепенно атрофируются.
15.3.2. Околощитовидные железы
Околощитовидные железы (glandulae parathyroideae) (4-5) расположены на задней поверхности щитовидной железы и отделены от нее капсулой. Масса желез 0,05-0,3 г.
Функциональное значение околощитовидных желез заключается в регуляции метаболизма кальция. Они вырабатывают белковый гормон парати-рин, который стимулирует резорбцию кости остеокластами, повышая содержание кальция в крови, и снижает содержание фосфора в крови, тормозя его резорбцию в почках, уменьшает экскрецию кальция почками, усиливает синтез 1-2,5-дигидроксихолекальциферола (метаболита витамина D), который повышает содержание кальция в сыворотке и его всасывание в пищеварительном тракте.
Паратирин и кальцитонин тесно взаимодействуют в регуляции минерального обмена: кальцитонин снижает уровень кальция в крови; паратирин является антагонистом кальцитонина. Гипокальциемия усиливает секрецию паратирина, а гиперкальциемия, наоборот, подавляет. Кальцитонин и паратирин также действуют на функцию почек и пищеварительный тракт, регулируя экскрецию и поглощение кальция в этих органах.
Развитие. Околощитовидные железы закладываются у зародыша как выступы из эпителия III и IV пары жаберных карманов глоточной кишки. На 5-6-й нед эмбриогенеза образуются четыре зачатка желез в виде эпителиальных почек. На 7-8-й нед эти почки отшнуровываются от стенок жаберных карманов, присоединяясь к задней поверхности щитовидной железы. В процессе гистогенеза эпителия околощитовидных желез составляющие его клетки становятся все более дифференцированными, их размеры увеличиваются, количество гликогена в них уменьшается, цитоплазма приобретает светлую окраску. Они называются главными паратироцитами. У 5-месячного плода главные паратироциты дифференцируются на светлые и темные паратироциты. На 10-м году жизни появляется следующий вид эпителиальных клеток желез - ацидофильные, или оксифильные, пара-тироциты. В виде единичных включений в паренхиме околощитовидных желез могут находиться С-клетки, вырабатывающие кальцитонин.
Строение. Каждая околощитовидная железа окружена тонкой соединительнотканной капсулой. Ее паренхима представлена трабекулами - эпителиальными тяжами либо скоплениями эпителиальных эндокринных клеток - паратироцитов (parathyrocytus endocrinus), разделенными тонкими прослойками рыхлой соединительной ткани с многочисленными капилля-
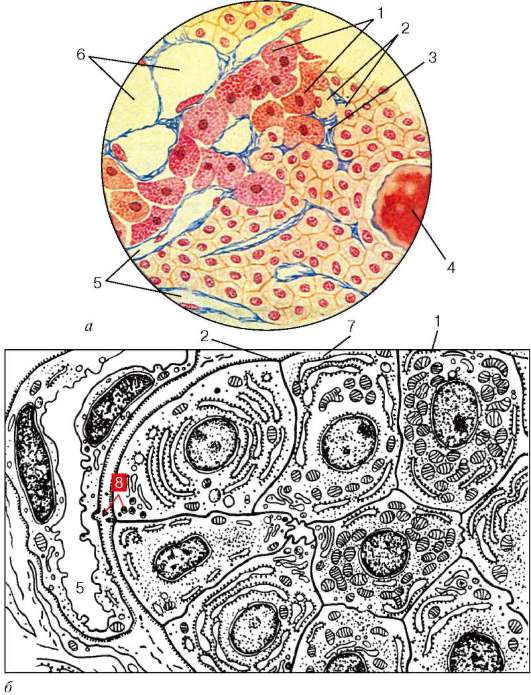
Рис. 15.13. Строение околощитовидной железы человека:
а - окраска по Маллори; б - схема ультрамикроскопического строения (по Ю. И. Афанасьеву). 1 - оксифильные паратироциты; 2 - главные паратироциты; 3 - соединительнотканные прослойки; 4 - фолликул с коллоидоподобным содержимым; 5 - капилляры; 6 - адипоциты; 7 - промежуточные клетки; 8 - пресекре-торные гранулы
рами (рис. 15.13, а, б). Хотя между паратироцитами хорошо развиты межклеточные щели, соседние клетки связаны интердигитациями и десмосома-ми. Различают главные паратироциты (parathyrocytus principalis) и оксифильные паратироциты (parathyrocytus oxyphilicus).
Главные клетки секретируют паратирин, они преобладают в паренхиме железы, имеют небольшие размеры (7-10 мкм), полигональную форму. В периферических зонах цитоплазма базофильна, где рассеяны скопления свободных рибосом (полисом).
Секреторные гранулы имеют диаметр 150-200 нм. При усилении секреторной активности околощитовидных желез главные клетки увеличиваются в объеме. Среди главных паратироцитов различают светлые (parathyrocytus principalis lucidus) неактивные формы и темные (parathyrocytus principalis densus) активные формы. В последних хорошо развиты органеллы синтеза секрета. В цитоплазме светлых клеток встречаются включения гликогена и жира. Обычно на один темный приходится 3-5 светлых паратироцитов.
Оксифильные паратироциты малочисленны, располагаются поодиночке или группами, они значительно крупнее, чем главные паратироциты. В цитоплазме видны оксифильные гранулы, огромное количество митохондрий, слабо развитый комплекс Гольджи. Их рассматривают как стареющие формы главных клеток. Другая точка зрения состоит в том, что это клетки APUD-серии. Выделяют также промежуточный тип клеток.
На секреторную активность околощитовидных желез не оказывают влияния гипофизарные гормоны. Околощитовидная железа по принципу обратной связи быстро реагирует на малейшие колебания содержания кальция в крови. Ее деятельность усиливается при гипокальциемии и ослабляется при гиперкальциемии. Паратироциты обладают рецепторами, способными непосредственно воспринимать прямые влияния на них ионов кальция.
Васкуляризация. Артерии распадаются на многочисленные капилляры, вены не анастомозируют, формируя петлистую сеть. Они лакунообразно расширяются и собираются в субкапсулярные сплетения, сообщающиеся с венами щитовидной железы.
Иннервация. Околощитовидные железы получают обильную симпатическую и парасимпатическую иннервацию. Безмиелиновые волокна заканчиваются терминалями в виде пуговок или колечек между паратироци-тами. Вокруг оксифильных клеток нервные терминали принимают вид корзиночек. Встречаются также инкапсулированные рецепторы. Влияние поступающих нервных импульсов ограничивается сосудодвигательными эффектами.
Возрастные изменения. У новорожденных и детей младшего возраста в паренхиме околощитовидных желез обнаруживаются только главные клетки. Оксифильные клетки появляются не ранее 5-7 лет, к этому времени их количество быстро нарастает. После 20-25 лет постепенно прогрессирует накопление адипоцитов. В пожилом возрасте обнаруживаются фолликулы, содержащие белковую жидкость.
Реактивность и регенерация. Разрастание эпителия околощитовидной железы, приводящее к ее гиперфункции, вызывает нарушение процесса обызвествления костной ткани (остеопороз, остеомаляция) и выведение кальция и фосфора из костей в кровь. При этом происходят резорбция костной ткани, нарастание количества остеокластов, разрастание фиброзной ткани. Кости становятся хрупкими, что приводит к повторным их переломам.
Гипофункция околощитовидной железы (травмы, удаление во время операции, инфекции) вызывает повышение нервно-мышечной возбудимости, ухудшение сократительной способности миокарда, судороги в связи
с недостатком кальция в крови. Физиологическая регенерация протекает медленно за счет митотического деления паратироцитов. При удалении околощитовидной железы возможна подкожная трансплантация фрагментов железы.
15.3.3. Надпочечники
Надпочечники (glandula suprarenalis) - это парные органы, образованные соединением двух отдельных самостоятельных гормонопродуцирую-щих желез, составляющих корковое и мозговое вещество разного происхождения, регуляции и физиологического значения (рис. 15.14). В корковом веществе (коре) надпочечников образуется комплекс стероидных гормонов, которые регулируют обмен углеводов, состав ионов во внутренней среде организма и половые функции - глюкокортикоиды, минералокортикои-ды, половые гормоны. Функция коры, кроме клубочковой зоны, контролируется адренокортикотропным гормоном (АКТГ) гипофиза и гормонами почек - ренин-ангиотензиновой системой.
В мозговом веществе продуцируются катехоламины (эпинефрин и нор-эпинефрин), которые влияют на быстроту сердечных сокращений, сокращение гладких мышц и метаболизм углеводов и липидов.
Развитие. Закладка корковой части обнаруживается у зародыша человека на 5-й нед развития в виде утолщений целомического эпителия по обе стороны корня брыжейки. В дальнейшем эти эпителиальные утолщения, образованные крупными клетками с ацидофильной цитоплазмой, собираются в компактное интерреналовое тело. Ацидофильные клетки становятся зачатком первичной (фетальной) коры будущих надпочечников. На 10-й нед внутриутробного периода первичная кора окружается снаружи мелкими базофильными клетками (происходящими тоже из целомического эпителия), которые дают начало дефинитивной коре надпочечников.
В плодном периоде объем фетальной коры значительно увеличивается, составляя большую часть коркового вещества надпочечника. В течение первого года жизни фетальная кора постепенно исчезает, заменяясь дефинитивной корой. В фетальной коре синтезируются главным образом глюкокортикоиды - кортизол и дегидроэпи-андростерон, которые метаболизируются в печени в 16α-производные, из которых, в свою очередь, образуются в плаценте женские половые гормоны - эстрогены (эстриол, эстрадиол, эстрон).
Из того же целомического эпителия, из которого возникает интерре-наловое тело, закладываются также половые валики - зачатки гонад, что обусловливает их функциональную взаимосвязь и близость химической природы их стероидных гормонов.
Мозговая часть надпочечников закладывается у зародыша человека на 6-7-й нед развития в результате миграции клеток нервного гребня. В процессе миграции происходит дивергентная дифференцировка малодиффе-ренцированных клеток в симпатобласты и хромаффинобласты. Последние
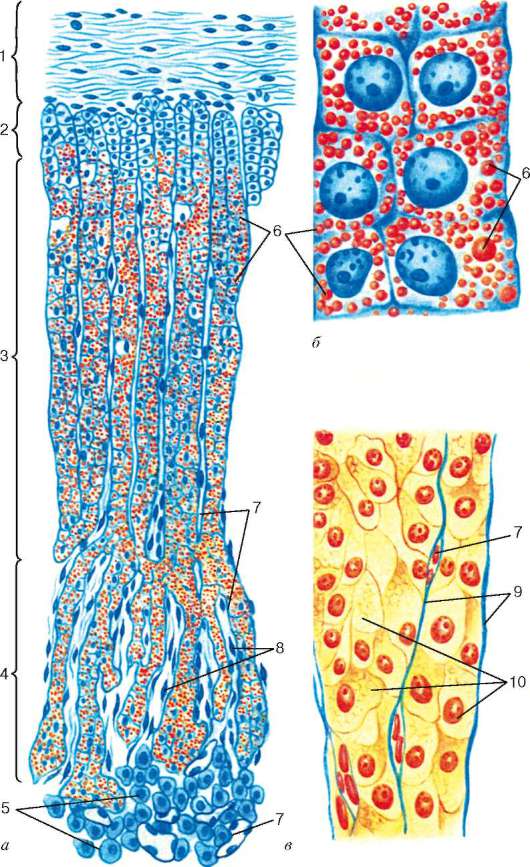
Рис. 15.14. Строение надпочечника (по Б. В. Алешину):
а, б - накопление липидов в кортикостероцитах (окраска суданом III-гематокси-лином); в - кортикостероциты пучковой зоны после растворения липидов (окраска по Маллори). 1 - капсула; 2 - клубочковая зона; 3 - пучковая зона; 4 - сетчатая зона; 5 - хромаффинные клетки мозгового вещества; 6 - капли липидов в кортикостероцитах; 7 - капилляры; 8 - эндотелий; 9 - соединительнотканные прослойки между тяжами кортикостероцитов; 10 - спонгиоциты
внедряются в интерреналовое тело, где размножаются и дифференцируются в хромаффиноциты и дают начало мозговому веществу надпочечников. Следовательно, железистые (хромаффинные) клетки мозгового вещества надпочечников должны рассматриваться как нейроэндокринные. У зародыша хромаффинные клетки вначале содержат только норадреналин (нор-эпинефрин), а на поздних стадиях эмбриогенеза появляются хромаффин-ные клетки, содержащие адреналин (эпинефрин).
Симпатобласты, внедрившиеся в интерреналовое тело, дивергентно дифференцируются в нейроны и глиоциты интрамуральных ганглиев.
Строение. Снаружи надпочечник покрыт соединительнотканной капсулой, в которой различаются два слоя - наружный (плотный) и внутренний (более рыхлый). В толще капсулы нередко обнаруживаются скопления клеток коры в виде узелков различных размеров. Основные структурные компоненты надпочечников - корковое и мозговое вещество.
Корковое вещество надпочечников
Клетки коры надпочечника, или кортикостероциты (corticosterocyti), образуют эпителиальные тяжи, ориентированные перпендикулярно к поверхности надпочечника (см. рис. 15.14).
В коре надпочечника имеется три основные зоны: клубочковая зона (zona glomerulosa), составляющая около 15 % толщины коры, пучковая зона (zona fasciculata) - 75 % и сетчатая зона (zona reticularis) - 10 % толщины коры. Промежутки между эпителиальными тяжами заполнены рыхлой соединительной тканью, по которой проходят кровеносные капилляры и нервные волокна, оплетающие тяжи. Под капсулой имеется тонкий слой мелких малодифференцированных эпителиальных клеток, размножением которых обеспечивается регенерация коры и создается возможность возникновения добавочных интерреналовых телец, иногда обнаруживаемых на поверхности надпочечников и нередко оказывающихся источниками опухолей (в том числе и злокачественных).
Клубочковая зона (zona glomerulosa) образована мелкими кортикостероци-тами (12-15 мкм), которые формируют округлые скопления («клубочки»). В этой зоне клетки содержат мало липидных включений. Их агранулярная эндоплазматическая сеть представлена мелкими пузырьками, между которыми обнаруживаются рибосомы. Многочисленные митохондрии овальной или удлиненной формы отличаются пластинчатыми кристами. Хорошо развит комплекс Гольджи.
В клубочковой зоне вырабатываются минералокортикоиды, главным из которых является альдостерон.
Основная функция минералокортикоидов - поддержание ионного состава электролитов в организме. Минералокортикоиды влияют на реаб-сорбцию и экскрецию ионов в почечных канальцах.
В частности, альдостерон увеличивает реабсорбцию ионов натрия, хлора, бикарбоната и усиливает экскрецию ионов калия и водорода. На синтез и секрецию альдосте-рона влияет ряд факторов. Гормон эпифиза адреногломерулотропин стимулирует образование альдостерона. Стимулирующее влияние на синтез и секрецию альдостерона
оказывают компоненты ренин-ангиотензиновой системы, а тормозящее - натрий-уретические факторы. Простагландины могут оказывать как стимулирующее (Е1 и Е2), так и тормозящее (F1α и F2α) влияние. При гиперсекреции альдостерона происходят задержка натрия в организме, обусловливающая повышение артериального давления, и потеря калия, сопровождающаяся мышечной слабостью.
При пониженной секреции альдостерона отмечаются потеря натрия, сопровождающаяся гипотензией, и задержка калия, ведущая к нарушениям сердечного ритма. Кроме того, минералокортикоиды усиливают воспалительные процессы. Минералокортикоиды жизненно важны. Разрушение или удаление клубочковой зоны приводит к смертельному исходу.
Между клубочковой и пучковой зонами располагается узкая прослойка мелких малодифференцированных клеток. Она называется промежуточной. Предполагается, что размножение клеток данной прослойки обеспечивает регенерацию пучковой и сетчатой зон.
Пучковая зона (zona fasciculata) занимает среднюю часть коры и наиболее выражена. Кортикостероциты этой зоны (рис. 15.15) отличаются крупными размерами (20 мкм), кубической или призматической формой; на поверхности, обращенной к капиллярам, имеются микроворсинки.
Цитоплазма этих клеток изобилует каплями липидов. Митохондрии крупные, округлой или овальной формы, с кристами в виде извитых и ветвящихся трубок (везикулярные кристы). Гладкая эндоплазматическая сеть хорошо выражена. Рибосомы лежат в цитоплазме свободно. В этой зоне наряду со светлыми клетками встречаются в разном количестве темные клетки с уплотненной цитоплазмой, содержащей мало липидных включений, но повышенное количество рибонуклеопротеидов. В темных клетках, помимо агранулярной эндоплазматической сети, имеется гранулярная эндоплазматическая сеть. Светлые и темные клетки представляют разные функциональные состояния одних и тех же кортикостероцитов. Полагают, что в темных клетках осуществляется синтез специфических белков - ферментов, которые в дальнейшем участвуют в образовании кортикостероидов, о чем свидетельствует обильное содержание в цитоплазме темных клеток рибосом. По мере выработки стероидов и их накопления цитоплазма клеток становится светлой, и они вступают в фазу выделения готового секреторного продукта в кровь.
В пучковой зоне вырабатываются глюкокортикоидные гормоны: кор-тикостерон, кортизон и гидрокортизон (кортизол). Они влияют на метаболизм углеводов, белков и липидов и усиливают процессы фосфорили-рования в организме, чем способствуют образованию веществ, богатых энергией, высвобождаемой затем для энергетического обеспечения всех процессов жизнедеятельности, протекающих в каждой клетке организма. Глюкокортикоиды форсируют глюконеогенез (образование глюкозы за счет белков) и отложение гликогена в печени и миокарде, а также мобилизацию тканевых белков. Большие дозы глюкокортикоидов вызывают деструкцию и распад лимфоцитов и эозинофилов крови, приводя к лимфоцитопении и эозинофилопении, а также угнетают воспалительные процессы в организме.
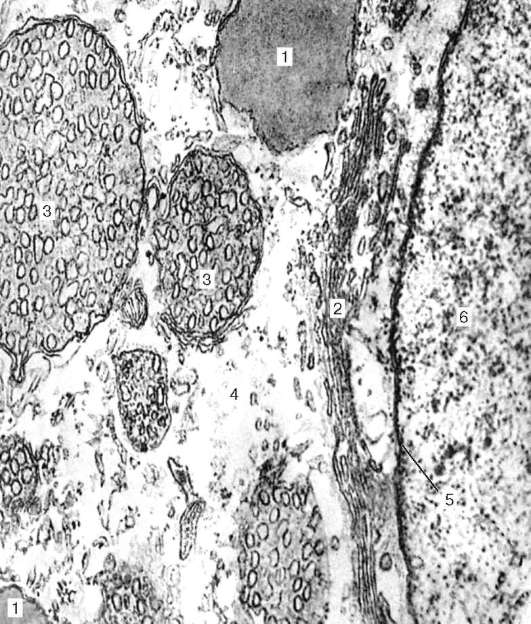
Рис. 15.15. Кортикостероцит пучковой зоны. Электронная микрофотография, увеличение 54 000 (по В. П. Деревянко):
1 - липидные включения; 2 - комплекс Гольджи; 3 - митохондрии; 4 - гиалоплаз-ма; 5 - ядерная оболочка; 6 - ядро
Сетчатая зона (zona reticularis). В ней эпителиальные тяжи разветвляются, формируя рыхлую сеть. Кортикостероциты в сетчатой зоне уменьшаются в размерах и становятся кубическими, округлыми или угловатыми. Содержание в них липидных включений убывает, а число темных клеток возрастает. Кристы митохондрий в клетках трубчатые. Эндоплазматическая сеть преимущественно вакуолярная, в цитоплазме преобладают свободные рибосомы. Комплекс Гольджи хорошо развит.
В сетчатой зоне вырабатывается андрогенстероидный гормон, близкий по химической природе и физиологическим свойствам к тестостерону семенников. Поэтому опухоли коры надпочечников у женщин нередко оказываются причиной вирилизма (развитие вторичных половых признаков мужского пола, в частности усов и бороды). В сетчатой зоне образуются также женские половые гормоны (эстрогены и прогестерон), но в небольших коли-
чествах. Иногда в сетчатой зоне на границе с мозговым веществом сохраняются остатки фетальной коры. Ее клетки отличаются ацидофильной цитоплазмой. Эти остаточные образования иначе называются Х-зоной. Она постоянно обнаруживается в надпочечниках самок некоторых млекопитающих, а у самцов становится заметной после кастрации.
Мозговое вещество надпочечников
Мозговое вещество (medulla) отделено от коркового вещества тонкой, местами прерывающейся прослойкой соединительной ткани. Эта часть надпочечников образована скоплением сравнительно крупных клеток округлой формы - хромаффинных клеток (endocrinocytus medullaris), между которыми находятся кровеносные сосуды (синусоиды). Различают светлые хро-маффинные клетки, или адреноциты, секретирующие адреналин, и темные хромаффинные клетки, или норадреноциты, секретирующие норадреналин. Цитоплазма клеток густо заполнена электронно-плотными секреторными гранулами диаметром 100-500 нм, окаймленными мембраной. Сердцевина гранулы заполнена белком, аккумулирующим секретируемые катехолами-ны - норадреналин и адреналин. После обработки надпочечников раствором бихромата калия в железистых клетках откладывается бурый осадок низших окислов хрома. Подобным же образом эти клетки восстанавливают четырехокись осмия и нитрат серебра, вследствие чего эти клетки получают наименование хромаффинных, или осмиофильных, или аргирофильных.
Электронно-плотные хромаффинные гранулы, помимо биогенных аминов (норадреналин, адреналин, ДОФА), содержат пептиды - энкефалины и хромогранины, что подтверждает их принадлежность к нейроэндокринным клеткам (APUD-серии клеток). Кроме того, в мозговом веществе находятся мультиполярные нейроны автономной нервной системы.
Катехоловые амины (норадреналин и адреналин) оказывают влияние на гладкие мышечные клетки сосудов, пищеварительного тракта, бронхов, на сердечную мышцу, а также на метаболизм углеводов (глюкогенолиз, глюко-неогенез), липидов (липолиз).
Катехоловые амины образуются из аминокислоты тирозина, которая при декар-боксилировании образует ДОФА (дигидрооксифенилаланин), из которого последовательно образуются норадреналин и адреналин. Действие катехоламинов на клетки-мишени реализуется через α- и β-адренергические мембранные рецепторы, связанные с G-белком, активирующим или ингибирующим аденилатциклазу. Образование и выброс в кровь катехоламинов стимулируется при активации симпатической нервной системы.
Регуляция секреторных функций коры надпочечников. Специфическим возбудителем гормонообразовательной деятельности пучковой и сетчатой зон является аденогипофизарный АКТГ.
Регуляция клубочковой зоны более сложна. Поскольку альдостерон образуется из кортикостерона, биосинтез которого стимулируется АКТГ, начальные стадии генеза минералокортикоидов подчиняются влиянию этого аденогипофизарного активатора. Однако переход кортикостерона в
альдостерон определяется дополнительным вмешательством ренина (гормон, вырабатываемый в почке). Кроме того, образование альдостерона стимулирует гормон эпифиза адреногломерулотропин.
Возрастные изменения. Кора надочечников у человека достигает полного развития в возрасте 20-25 лет, когда соотношение толщины ее зон приближается к 1:9:3. После 59 лет толщина коры начинает уменьшаться. В клетках коры постепенно уменьшаются количество и размеры липидных включений, а соединительнотканные прослойки между эпителиальными тяжами утолщаются. Редукция поражает главным образом сетчатую и отчасти клу-бочковую зону. Толщина же пучковой зоны относительно увеличивается, что обеспечивает достаточную интенсивность глюкокортикоидной функции надпочечников вплоть до преклонного возраста. Мозговое вещество надпочечников не претерпевает отчетливо выраженных возрастных изменений. После 40 лет отмечается некоторая гипертрофия хромаффиноцитов, но лишь в преклонном возрасте в них наступают атрофические изменения, ослабевает синтез катехоламинов, а в сосудах и строме обнаруживаются признаки склероза.
Васкуляризация. У мозгового и коркового вещества надпочечника имеется общее кровоснабжение. Артерии, входящие в надпочечник, разветвляются на артериолы, образующие густую подкапсулярную сеть, от которой отходят капилляры, снабжающие кровью кору. Их эндотелий фенестриро-ван, что облегчает поступление кортикальных стероидных гормонов из клеток коры в ток крови. Из сетчатой зоны капилляры вступают в мозговую часть, где принимают вид синусоидов и сливаются в венулы, которые переходят в его венозное сплетение. Наряду с ними в мозговое вещество входят также артерии, берущие начало от подкапсулярной сети. Проходя через кору и обогащаясь продуктами, секретируемыми клетками коры надпочечника, кровь приносит к хромаффиноцитам специальные, вырабатываемые в коре ферменты, которые активируют метилирование норадреналина, т. е. образование адреналина.
В мозговом веществе ветвление кровеносных сосудов таково, что каждый хромаффиноцит одним концом соприкасается с артериальным капилляром, а другим обращен к венозному синусоиду, в который выделяет катехоламины. Венозные синусоиды собираются в центральную вену надпочечника, впадающую в нижнюю полую вену. Таким образом, в кровь поступают одновременно и кортикостероиды, и катехоламины, чем обеспечивается возможность совместного действия обоих регуляторных факторов на эффекторные органы или системы. По другим же венам кровь из коры и мозгового вещества направляется в воротную вену печени, принося в нее адреналин (увеличивающий мобилизацию глюкозы из гликогена) и глюко-кортикоиды, стимулирующие, наоборот, глюконеогенез в печени.
Иннервация. Надпочечники получают эфферентные нервные импульсы главным образом по чревным и блуждающим нервам. В кору входят пост-ганглионарные (безмиелиновые) симпатические волокна, а в мозговую часть (как и в другие симпатические ганглии) преганглионарные (миелиновые) волокна. В капсуле нередко встречаются микроганглии и одиночные ган-
глиозные нервные клетки. В корковой части, деятельность которой активизируется аденогипофизарным АКТГ, роль прямых нервных импульсов ограничивается вазомоторными эффектами. В мозговом веществе, которое не зависит от гипофиза, регуляция его секреторной деятельности осуществляется исключительно нервными импульсами, поступающими главным образом по чревным нервам. Раздражение последних усиливает отдачу адреналина и норадреналина хромаффинными клетками в ток крови.
Реактивность и регенерация. При стрессах, сопровождающихся сильными эмоциональными реакциями страха или ярости, преобладает активность симпатической нервной системы над парасимпатической. При этом повышается не только активность постганглионарных симпатических нейронов, но и секреция клеток мозгового вещества надпочечника. В кровь поступают большие количества норадреналина и адреналина. В результате учащаются и усиливаются сокращения сердца, повышается артериальное давление, увеличивается объем циркулирующей крови в сосудах мышц и центральной нервной системы, в кровь из печени выбрасываются запасы глюкозы. Усиленное выделение адреналина и норадреналина клетками мозгового вещества надпочечника происходит и рефлекторно при резком охлаждении, боли и других видах стресса.
Физиологическая регенерация коры надпочечника осуществляется при участии подкапсулярных клеток и клеток, расположенных между клубочко-вой и пучковой зонами, которые находятся под контролем АКТГ аденоги-пофиза. При удалении одного надпочечника наблюдается компенсаторная гипертрофия и гиперплазия железистых клеток другого надпочечника.
Параганглии, подобно мозговому веществу надпочечников, состоят из хромаффинной ткани, развивающейся из симпатобластов нервного гребня. Различают брюшные, аортальные, каротидные, внутриорганные (в сердце, коже, яичках, матке и т. д.) параганглии. Снаружи они окружены соединительной тканью, прослойки которой проникают между тяжами зернистых эндокриноцитов. Последние диаметром 10-15 мкм имеют овальную или округлую форму и содержат специфические гранулы разного размера, в которых находятся катехоламины. Эндокриноциты окружены поддерживающими глиоцитами параганглия. Капилляр синусоидного типа с фенестрированны-ми эндотелиоцитами прилежит к группе эндокриноцитов в той части, где отсутствуют поддерживающие клетки. Иннервация органа осуществляется симпатической нервной системой.
15.4. ДИСПЕРСНАЯ ЭНДОКРИННАЯ СИСТЕМА (APUD-СЕРИЯ КЛЕТОК)
Совокупность одиночных гормонпродуцирующих клеток называют дисперсной (диффузной) эндокринной системой (ДЭС), а клетки относят к APUD-серии (апудоцитам). Название группы эндокринных клеток, составляющих APUD-серию, происходит от первых букв английских слов amine
precursor uptake and decarboxylation (поглощение и декарбоксилирование аминов и их предшественников). Это название отражает основной признак клеток данной серии - способность накапливать предшественники биогенных аминов, декарбоксилировать их и вырабатывать биогенные амины и полипептидные гормоны. Последние играют важную роль в процессах местной тканевой регуляции. По морфологическим, биохимическим и функциональным признакам выделено более 50 видов клеток APUD-серии. Общей топографической особенностью этих клеток является их расположение около кровеносных сосудов, среди клеток, находящихся в составе эпителия, - полярная дифференцировка (хотя и не всегда отчетливо выраженная), соответствующая выделению секрета (инкрета) в сосуды микро-циркуляторного русла. Эндокринные клетки APUD-серии обнаруживают тесную и прямую зависимость от нервных импульсов, поступающих к ним по симпатическим и парасимпатическим нервам, но не реагируют на троп-ные гормоны передней доли гипофиза; их состояние и активность после гипофизэктомии не нарушаются.
Для клеток APUD-серии характерно присутствие в них нейроаминов (например, серотонина), катехоламинов и других биогенных аминов, которые выявляются благодаря специфической флюоресценции после обработки в парах формальдегида и просмотра препаратов в ультрафиолетовых лучах; поглощение предшественников аминов при введении их в организм (например, 5-окситриптофана, диоксифенилаланина и др.); декарбоксили-рование аминов. Пептидные гормоны выявляются преимущественно имму-ноцитохимическими методами.
В клетках APUD-серии определяется высокое содержание ряда ферментов (эстераз, холинэстераз, альфа-глицерофосфатдегидрогеназ). Иными словами, клетки данной серии сочетают признаки нервных и эндокринных клеток.
Под электронным микроскопом в базальной части этих клеток выявляется большое количество секреторных гранул, белковая сердцевина которых окружена мембраной (рис. 15.16). Клетки APUD-серии дают специфическую реакцию при обработке их антисыворотками к биогенным аминам и полипептидным гормонам.
Гистогенетические и гистофизиологические различия не дают оснований для объединения всех так называемых апудоцитов в единую (генетически) клеточную систему. Несмотря на сходство по некоторым, главным образом гистохимическим, признакам, эндокринные клетки неэндокринных органов являются составными элементами соответствующих тканей, образуя в этих тканях линии дивергентной клеточной дифференцировки, или клеточные диффероны. По этой причине различают клетки APUD-серии: производные нейроэктодермы (нейроэндокриноциты гипоталамуса, эпифиза, пептидергические нейроны ЦНС и ПНС); производные кожной эктодермы (клетки Меркеля, эндокриноциты APUD-серии аденогипофиза); производные кишечной энтодермы (эндокриноциты гастроэнтеропанкреати-ческой системы); производные мезодермы (клетки Лейдига, эндокриноциты теки фолликула яичника) и др.
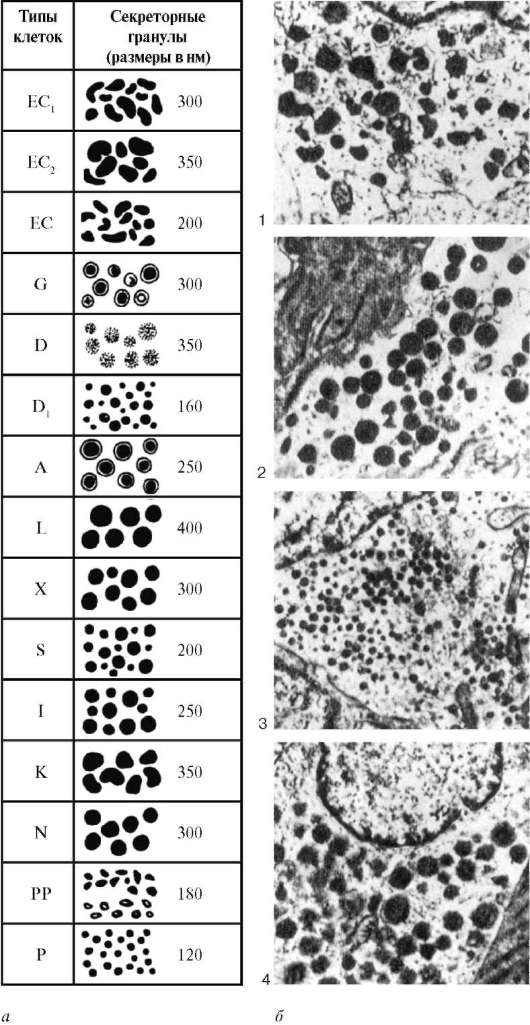
Рис. 15.16. Строение и распределение различных типов эндокриноцитов в пищеварительной системе человека:
а - международная классификация и распределение эндокриноцитов. Ультраструктура их гранул (по Д. Грубе и В. Форссману, с изменениями); б - ультрамикроскопическое строение желудочно-кишечных эндокриноцитов. 1 - ЕС-клетки с гранулами разнообразной формы; 2 - L-клетки с крупными электронно-плотными гранулами; 3 - D1-клетки с многочисленными мелкими гранулами; 4 - D-клетки с крупными умеренно плотными гранулами (по С. М. Говалову, И. М. Коростышевской, М. С. Виноградовой)
В последние годы пептидные гормоны и биогенные амины обнаружены в некоторых неэндокринных клетках: больших гранулярных лимфоцитах (естественных киллерах), тучных клетках, эозинофильных лейкоцитах, некоторых эндотелиальных клетках, тромбоцитах и моноцитах. Предполагается, что эндокринная функция неэндокринных клеток является отражением внутренних ауторегуляторных механизмов выполнения различными клетками присущих им специфических функций.
Таким образом, для эндокриноцитов APUD-серии, несмотря на различные источники их происхождения, характерно наличие в цитоплазме как нейроамина (серотонина), так и пептидного гормона. И тот, и другой секреторный продукт оказывает дистантное или местное (паракринное) воздействие на клетки-мишени, расположенные в данном или другом органе.
Контрольные вопросы
1. Иерархический принцип организации эндокринной системы. Гипоталамус, гипофиз, эпифиз как центральные звенья регуляции функций периферических эндокринных желез.
2. Аденогипофиззависимые эндокринные железы: название, развитие, продуцируемые гормоны, органы и клетки-мишени к их гормонам, значение в регуляции физиологических функций организма.
3. Аденогипофизнезависимые эндокринные железы: название, развитие, продуцируемые гормоны, органы и клетки-мишени к гормонам, значение в регуляции физиологических функций организма.
4. Дисперсная эндокринная система: классификация, топография в организме человека, виды эндокринных клеток, названия и физиологические функции гормонов. Понятие о пара- и аутокринной гормональной регуляции.