Гистология, эмбриология, цитология : учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
|
|
|
|
Глава 12. ОРГАНЫ ЧУВСТВ
12.1. ОБЩАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА И КЛАССИФИКАЦИЯ
Органы чувств обеспечивают восприятие различных раздражителей, действующих на организм; преобразование и кодирование внешней энергии в нервный импульс, передачу по нервным путям в подкорковые и корковые центры, где происходят анализ поступившей информации и формирование субъективных ощущений. Органы чувств - это анализаторы внешней и внутренней среды, которые обеспечивают адаптацию организма к конкретным условиям.
Соответственно в каждом анализаторе различают три части: периферическую (рецепторную), промежуточную и центральную.
Периферическая часть представлена органами, в которых находятся специализированные рецепторные клетки. По специфичности восприятия стимулов различают механорецепторы (рецепторы органа слуха, равновесия, тактильные рецепторы кожи, рецепторы аппарата движения, барорецепторы), хеморецепторы (органов вкуса, обоняния, сосудистые интерорецепторы), фоторецепторы (сетчатки глаза), терморецепторы (кожи, внутренних органов), болевые рецепторы.
Промежуточная (проводниковая) часть анализатора представляет собой цепь вставочных нейронов, по которым нервный импульс от рецепторных клеток передается к корковым центрам. На этом пути могут быть промежуточные, подкорковые, центры, где происходят обработка афферентной информации и переключение ее на эфферентные центры.
Центральная часть анализатора представлена участками коры больших полушарий. В центре осуществляются анализ поступившей информации, формирование субъективных ощущений. Здесь информация может быть заложена в долговременную память или переключена на эфферентные пути.
Классификация органов чувств . В зависимости от строения и функции рецепторной части органы чувств делятся на три типа.
К первому типу относятся органы чувств, у которых рецепторами являются специализированные нейросенсорные клетки (орган зрения, орган обоняния), преобразующие внешнюю энергию в нервный импульс.
Ко второму типу относятся органы чувств, у которых рецепторами являются не нервные, а эпителиальные клетки (сенсоэпителиальные). От них
преобразованное раздражение передается дендритам чувствительных нейронов, которые воспринимают возбуждение сенсоэпителиальных клеток и порождают нервный импульс (органы слуха, равновесия, вкуса).
К третьему типу относятся проприоцептивная (скелетно-мышечная) кожная и висцеральная сенсорные системы. Периферические отделы в них представлены различными инкапсулированными и неинкапсулированны-ми рецепторами (см. главу 10).
12.2. ОРГАН ЗРЕНИЯ
Глаз (ophthalmos oculus) - орган зрения, представляющий собой периферическую часть зрительного анализатора, в котором рецепторную функцию выполняют нейросенсорные клетки сетчатой оболочки.
12.2.1. Развитие глаза
Глаз развивается из различных эмбриональных зачатков (рис. 12.1). Сетчатка и зрительный нерв формируются из нервной трубки путем образования сначала так называемых глазных пузырьков, сохраняющих связь с эмбриональным мозгом при помощи полых глазных стебельков. Передняя часть глазного пузырька впячивается внутрь его полости, благодаря чему он приобретает форму двустенного глазного бокала. Часть эктодермы, расположенная напротив отверстия глазного бокала, утолщается, инвагинирует и отшнуровывается, давая начало зачатку хрусталика. Эктодерма претерпевает эти изменения под влиянием индукторов дифференцировок, образующихся в глазном пузырьке. Первоначально хрусталик имеет вид полого эпителиального пузырька. Затем клетки эпителия его задней стенки удлиняются и превращаются в так называемые хрусталиковые волокна, заполняющие полость пузырька. В процессе развития внутренняя стенка глазного бокала преобразуется в сетчатку, а наружная - в пигментный слой сетчатки. На 4-й нед эмбриогенеза зачаток сетчатки состоит из однородных малодиффе-ренцированных клеток. На 5-й нед появляется разделение сетчатки на два слоя: наружный (от центра глаза) - ядерный, и внутренний слой, не содержащий ядер. Наружный ядерный слой играет роль матричной зоны, где наблюдаются многочисленные фигуры митоза. В результате последующей дивергентной дифференцировки стволовых (матричных) клеток развиваются клеточные диффероны различных слоев сетчатки. Так, в начале 6-й нед из матричной зоны начинают выселяться нейробласты, образующие внутренний слой. В конце 3-го мес определяется слой крупных ганглиозных нейронов. В последнюю очередь в сетчатке появляется наружный ядерный слой, состоящий из нейросенсорных клеток - палочковых и колбочковых нейронов. Происходит это незадолго до рождения. Помимо нейробластов в матричном слое сетчатки образуются глиобласты - источники развития клеток глии.
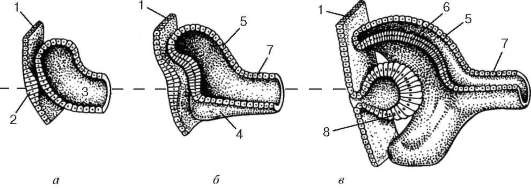
Рис. 12.1. Развитие глаза:
а-в - сагиттальные разрезы глаз эмбрионов на различных стадиях развития. 1 - эктодерма; 2 - хрусталиковая плакода - будущий хрусталик; 3 - глазной пузырек; 4 - сосудистая выемка; 5 - наружная стенка глазного бокала - будущий пигментный слой сетчатки; 6 - внутренняя стенка глазного бокала; 7 - стебелек - будущий зрительный нерв; 8 - хрусталиковый пузырек
Высокодифференцированными среди них становятся радиальные глиоциты (мюллеровы волокна), пронизывающие всю толщу сетчатки.
Стебелек глазного бокала пронизывается аксонами, образующимися в сетчатке ганглиозных мультиполярных нейронов. Эти аксоны и формируют зрительный нерв , направляющийся в мозг. Из окружающей глазной бокал мезенхимы формируются сосудистая оболочка и склера. В передней части глаза склера переходит в покрытую многослойным плоским эпителием (эктодермальным) прозрачную роговицу. Изнутри роговица выстлана однослойным эпителием нейроглиального происхождения. Сосуды и мезенхима, проникающие на ранних стадиях развития внутрь глазного бокала, совместно с эмбриональной сетчаткой принимают участие в образовании стекловидного тела и радужки. Мышца радужки, суживающая зрачок, развивается из краевого утолщения наружного и внутреннего листков глазного бокала, а мышца, расширяющая зрачок, - из наружного листка. Таким образом, обе мышцы радужки по своему происхождению являются нейральными.
12.2.2. Строение глаза
Глазное яблоко (bulbus oculi) состоит из трех оболочек. Наружная (фиброзная) оболочка глазного яблока (tunica fibrosa bulbi), к которой прикрепляются наружные мышцы глаза, обеспечивает защитную функцию. В ней различают передний прозрачный отдел - роговицу и задний непрозрачный отдел - склеру. Средняя (сосудистая) оболочка (tunica vasculosa bulbi) играет основную роль в обменных процессах. Она имеет три части: часть радужки, часть цилиарного тела и собственно сосудистую - хороидею (choroidea).
Внутренняя оболочка глаза - сетчатка (tunica interna bulbi, retina) - сенсорная, рецепторная часть зрительного анализатора, в которой происходят
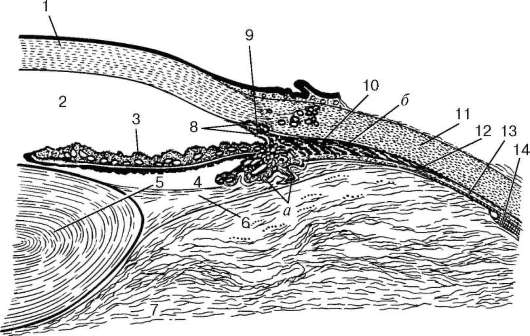
Рис. 12.2. Строение переднего отдела глазного яблока (схема):
1 - роговица; 2 - передняя камера глаза; 3 - радужка; 4 - задняя камера глаза; 5 - хрусталик; 6 - ресничный поясок (циннова связка); 7 - стекловидное тело; 8 - гребенчатая связка; 9 - венозный синус склеры; 10 - ресничное (цилиарное) тело: а - отростки ресничного тела; б - ресничная мышца; 11 - склера; 12 - сосудистая оболочка; 13 - зубчатая линия; 14 - сетчатка
под воздействием света фотохимические превращения зрительных пигментов, фототрансдукция, изменение биоэлектрической активности нейронов и передача информации о внешнем мире в подкорковые и корковые зрительные центры.
Оболочки глаза и их производные формируют три функциональных аппарата: светопреломляющий, или диоптрический (роговица, жидкость передней и задней камер глаза, хрусталик и стекловидное тело); аккомодационный (радужка, ресничное тело с ресничными отростками); рецепторный аппарат (сетчатка).
Наружная фиброзная оболочка - склера (sclera), образована плотной оформленной волокнистой соединительной тканью, содержащей пучки коллагеновых волокон, между которыми находятся уплощенной формы фибробласты и отдельные эластические волокна (рис. 12.2). Пучки кол-лагеновых волокон, истончаясь, переходят в собственное вещество роговицы.
Толщина склеры в заднем отделе вокруг зрительного нерва наибольшая - 1,2- 1,5 мм, кпереди склера истончается до 0,6 мм у экватора и до 0,3-0,4 мм позади места прикрепления прямых мышц. В области диска зрительного нерва большая часть (2/3) истонченной фиброзной оболочки сливается с оболочкой зрительного нерва, а истонченные внутренние слои образуют решетчатую пластинку (lamina cribrosa). При повышении внутриглазного давления фиброзная оболочка истончается, что является причиной некоторых патологических изменений.
Светопреломляющий аппарат глаза
Светопреломляющий (диоптрический) аппарат глаза включает роговицу, хрусталик, стекловидное тело, жидкость (водянистую влагу) передней и задней камер глаза.
Роговица (cornea) занимает 1/16 площади фиброзной оболочки глаза и, выполняя защитную функцию, отличается высокой оптической гомогенностью, пропускает и преломляет световые лучи и является составной частью светопреломляющего аппарата глаза.
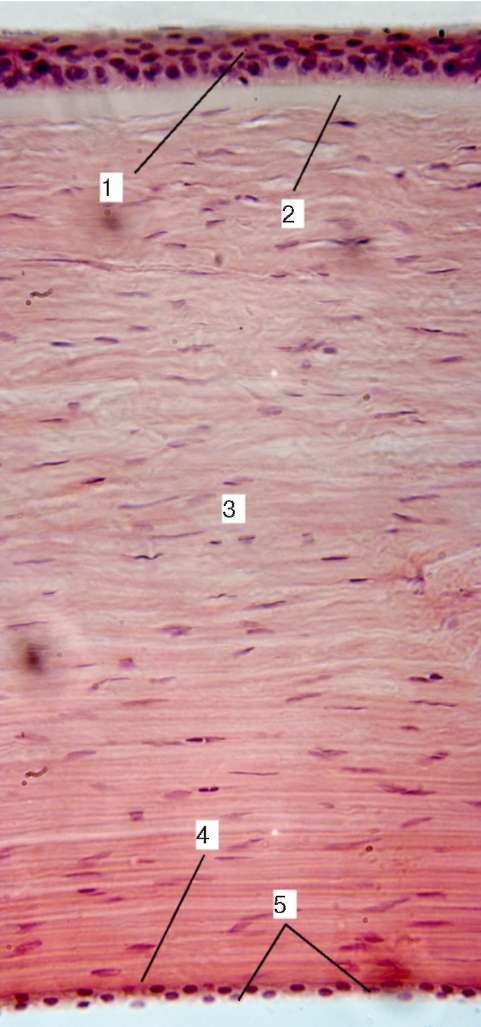
Рис. 12.3. Роговица глаза: 1 - многослойный плоский неорогове-вающий эпителий; 2 - передняя пограничная пластинка; 3 - собственное вещество; 4 - задняя пограничная пластинка; 5 - задний эпителий роговицы
Толщина роговицы 0,8-0,9 мкм в центре и 1,1 мкм на периферии, радиус кривизны 7,8 мкм, показатель преломления - 1,37, сила преломления 40 диоптрий.
В роговице микроскопически выделяют пять слоев: 1) передний многослойный плоский неорого-вевающий эпителий; 2) переднюю пограничную пластинку (боумено-ву мембрану); 3) собственное вещество; 4) заднюю пограничную пластинку (десцеметову мембрану); 5) задний эпителий (эндотелий передней камеры) (рис. 12.3).
Клетки переднего эпителия роговицы (кератоциты) плотно прилегают друг к другу, располагаются в пять слоев, соединены десмосомами (см. рис. 12.3). Базальный слой расположен на передней пограничной пластинке. В патологических условиях (при недостаточно прочной связи базального слоя и передней пограничной пластинки) происходит отслойка базального слоя от пограничной пластинки. Клетки базаль-ного слоя эпителия (камбиальные) имеют призматическую форму и овальное ядро, расположенное близко к вершине клетки. К базально-му слою примыкают 2-3 слоя многогранных клеток. Их вытянутые в стороны отростки внедряются между соседними клетками эпителия, подобно крыльям (крылатые, или шиповатые, клетки). Ядра кры-
латых клеток округлые. Два поверхностных эпителиальных слоя состоят из резко уплощенных клеток и не имеют признаков ороговения. Удлиненные узкие ядра клеток наружных слоев эпителия располагаются параллельно поверхности роговицы. В эпителии имеются многочисленные свободные нервные окончания, обусловливающие высокую тактильную чувствительность роговицы. Поверхность роговицы увлажнена секретом слезных и конъюнктивальных желез, который защищает глаз от вредных физико-химических воздействий внешнего мира, бактерий. Эпителий роговицы отличается высокой регенерационной способностью. Под эпителием роговицы расположена бесструктурная передняя пограничная пластинка (lamina limitans anterior) - боуменова мембрана - толщиной 6-9 мкм. Это гомогенный слой беспорядочно расположенных коллагеновых фибрилл - продукт жизнедеятельности эпителиоцитов. Граница между боуменовой мембраной и эпителием хорошо выражена, слияние боуменовой мембраны со стромой происходит незаметно.
Собственное вещество роговицы (substantia propria cornea) - строма - состоит из гомогенных тонких соединительнотканных пластинок, взаимопересе-кающихся под углом, но правильно чередующихся и расположенных параллельно поверхности роговицы. В пластинках и между ними располагаются отростчатые плоские клетки, являющиеся разновидностями фибробластов. Пластинки состоят из параллельно расположенных пучков коллагеновых фибрилл диаметром 0,3-0,6 мкм (по 1000 в каждой пластинке). Клетки и фибриллы погружены в основное вещество, богатое гликозаминогликана-ми (в основном кератинсульфатами), которое обеспечивает прозрачность собственного вещества роговицы. Оптимальная концентрация воды в стро-ме (75-80 %) поддерживается механизмом транспорта ионов натрия через задний эпителий. Переход прозрачной роговицы в непрозрачную склеру происходит в области лимба роговицы (limbus corneae). Собственное вещество роговицы не имеет кровеносных сосудов.
Задняя пограничная пластинка (lamina limitans posterior) - десцеметова мембрана - толщиной 5-10 мкм, представлена коллагеновыми волокнами диаметром 10 нм, погруженными в аморфное вещество. Это стекловидная, сильно преломляющая свет структура. Она состоит из двух слоев: наружного - эластического, внутреннего - кутикулярного и является производным клеток заднего эпителия. Характерными особенностями задней пограничной пластинки являются прочность, резистентность к химическим агентам и расплавляющему действию гнойного экссудата при язвах роговицы.
При гибели передних слоев десцеметова мембрана выпячивается в виде прозрачного пузырька (десцеметоцеле). На периферии она утолщается, и у людей пожилого возраста на этом месте могут формироваться округлые бородавчатые образования - тельца Гассаля-Генле.
У лимба десцеметова мембрана, истончаясь и разволокняясь, переходит в трабекулярный аппарат склеры (см. ниже).
Задний эпителий (epithelium posterius), или эндотелий передней камеры, состоит из одного слоя гексагональных клеток. Ядра клеток круглые или слегка овальные, их ось располагается параллельно поверхности роговицы. Клетки нередко содержат вакуоли. На периферии роговицы задний эпителий переходит непосредственно на волокна трабекулярной сети, образуя наружный покров каждого трабекулярного волокна, вытягиваясь в длину. Задний эпителий защищает роговицу от воздействия влаги передней камеры.
Процессы обмена в роговице обеспечиваются диффузией питательных веществ из передней камеры глаза за счет краевой петлистой сети роговицы, многочисленными концевыми капиллярными ветвями, образующими густое перилимбальное сплетение.
Лимфатическая система роговицы формируется из узких лимфатических щелей, сообщающихся с ресничным венозным сплетением.
Роговица отличается высокой чувствительностью, что объясняется наличием в ней нервных окончаний. Длинные цилиарные нервы, представляя ветви назоцили-арного нерва, отходящего от первой ветви тройничного нерва, на периферии роговицы проникают в ее толщу, теряют миелин на некотором расстоянии от лимба, делясь дихотомически. Нервные ветви образуют следующие сплетения: в собственном веществе роговицы, претерминальное и под передней пограничной пластинкой - терминальное, суббазальное (сплетение Райзера).
При воспалительных процессах кровеносные капилляры и клетки (лейкоциты, макрофаги и др.) проникают из области лимба в собственное вещество роговицы, что приводит к ее помутнению и ороговению, образованию бельма.
Передняя камера образована роговицей (наружная стенка) и радужкой (задняя стенка), в области зрачка - передней капсулой хрусталика. На крайней ее периферии в углу передней камеры имеется радужно-роговичный (камерный) угол (spatia anguli iridocornealis) с небольшим участком ресничного (цилиарного) тела. Камерный (так называемый фильтрационный) угол граничит с дренажным аппаратом - шлеммовым каналом. Состояние камерного угла играет большую роль в обмене водянистой влаги и в изменении внутриглазного давления. Соответственно вершине угла в склере проходит кольцевидно располагающийся желобок (sulcus sclerae internus). Задний край желобка несколько утолщен и образует склеральный валик, сформированный за счет круговых волокон склеры (заднее пограничное кольцо Швальбе). Склеральный валик служит местом прикрепления поддерживающей связки цилиарного тела и радужки - трабекулярного аппарата, заполняющего переднюю часть склерального желобка. В задней части он прикрывает шлеммов канал.
Трабекулярный аппарат, ранее ошибочно называвшийся гребенчатой связкой, состоит из двух частей: склерокорнеальной (lig. sclerocorneale), занимающей большую часть трабекулярного аппарата, и второй, более нежной, - увеаль-ной части, которая расположена с внутренней стороны и является собственно гребенчатой связкой (lig. pectinatum). Склерокорнеальный отдел трабекуляр-ного аппарата прикрепляется к склеральной шпоре, частично сливается с цилиарной мышцей (мышца Брюкке). Склерокорнеальная часть трабеку-лярного аппарата состоит из сети трабекул, имеющих сложную структуру.
В центре каждой трабекулы, представляющей плоский тонкий тяж, проходит коллагеновое волокно, обвитое, укрепленное эластическими волокнами и покрытое снаружи футляром из гомогенной стекловидной оболочки, являющейся продолжением задней пограничной пластинки. Между сложным переплетом корнеосклеральных волокон остаются многочисленные свободные щелевидные отверстия - фонтановы пространства, выстланные эндотелием передней камеры, переходящим с задней поверхности роговицы. Фонтановы пространства направлены к стенке венозного синуса склеры (sinus venosus sclerae) - шлеммова канала, расположенного в нижнем отделе склерального желобка шириной 0,25 см. В некоторых местах он разделяется на ряд канальцев, далее сливающихся в один ствол. Внутри шлеммов канал выстлан эндотелием. С его наружной стороны отходят широкие, иногда варикозно-расширенные сосуды, образующие сложную сеть анастомозов, от которых берут начало вены, отводящие водянистую влагу из передней и задней камер в глубокое склеральное венозное сплетение.
Хрусталик (lens). Это прозрачное двояковыпуклое тело, форма которого меняется во время аккомодации глаза к видению близких и отдаленных объектов. Вместе с роговицей и стекловидным телом хрусталик составляет основную светопреломляющую среду. Радиус кривизны хрусталика варьирует от 6 до 10 мм, показатель преломления составляет 1,42. Хрусталик покрыт прозрачной капсулой толщиной 11-18 мкм. Это базальная мембрана эпителия, которая содержит коллаген, сульфатированный гликозоами-ногликан и др. Передняя стенка хрусталика состоит из однослойного плоского эпителия (epithelium lentis). По направлению к экватору эпителиоциты становятся выше и образуют ростковую зону хрусталика. Эта зона является камбиальной для клеток передней и задней поверхности хрусталика. Новые эпителиоциты преобразуются в хрусталиковые волокна (fibrae lentis). Каждое волокно представляет собой прозрачную шестиугольную призму. В цитоплазме хрусталиковых волокон находится прозрачный белок - кристаллин. Волокна склеиваются друг с другом особым веществом, которое имеет такой же, как и они, коэффициент преломления. Центрально расположенные волокна теряют свои ядра, укорачиваются и, накладываясь друг на друга, образуют ядро хрусталика.
Хрусталик поддерживается в глазу с помощью волокон ресничного пояска (zonula ciliaris), образованного радиально расположенными пучками нерастяжимых волокон, прикрепленных с одной стороны к ресничному (цили-арному) телу, а с другой - к капсуле хрусталика, благодаря чему сокращение мышц ресничного тела передается хрусталику. Знание закономерностей строения и гистофизиологии хрусталика позволило разработать методы создания искусственных хрусталиков и широко внедрить в клиническую практику их пересадку, что сделало возможным лечение больных с помутнением хрусталика (катаракта).
Стекловидное тело (corpus vitreum). Это прозрачная масса желеобразного вещества, заполняющего полость между хрусталиком и сетчаткой, в составе которого 99 % воды. На фиксированных препаратах стекловидное тело имеет сетчатое строение. На периферии оно более плотное, чем в центре.
Через стекловидное тело проходит канал - остаток эмбриональной сосудистой системы глаза - от сосочка сетчатки до задней поверхности хрусталика. Стекловидное тело содержит белок витреин и гиалуроновую кислоту, из клеток в нем обнаружены гиалоциты, макрофаги и лимфоциты. Показатель преломления стекловидного тела равен 1,33.
Аккомодационный аппарат глаза
Аккомодационный аппарат глаза (радужка, ресничное тело с ресничным пояском) обеспечивает изменение формы и преломляющей силы хрусталика, фокусировку изображения на сетчатке, а также приспособление глаза к интенсивности освещения.
Радужка (iris). Представляет собой дисковидное образование с отверстием изменчивой величины (зрачок) в центре. Она является производным сосудистой (в основном) и сетчатой оболочек. Сзади радужка покрыта пигментным эпителием сетчатой оболочки. Расположена между роговицей и хрусталиком на границе между передней и задней камерами глаза (рис. 12.4). Край радужки, соединяющий ее с ресничным телом, называется ресничным (цилиар-ным) краем. Строма радужки состоит из рыхлой волокнистой соединительной ткани, богатой пигментными клетками. Здесь располагаются мионейральные клетки. Радужка осуществляет свою функцию в качестве диафрагмы глаза с помощью двух мышц: суживающей (musculus sphincter pupillae) и расширяющей (musculus dilatator pupillae) зрачок.
В радужке различают пять слоев: передний (наружный) эпителий, покрывающий переднюю поверхность радужки, передний пограничный (наружный бессосудистый) слой, сосудистый слой, задний (внутренний) пограничный слой и задний (пигментный) эпителий.
Передний эпителий (epithelium anterius iridis) представлен нейроглиальными плоскими полигональными клетками. Он является продолжением эпителия, покрывающего заднюю поверхность роговицы.
Передний пограничный слой (stratum limitans anterius) состоит из основного вещества, в котором располагаются значительное количество фибробластов и пигментных клеток. Различное положение и количество меланинсодержащих клеток обусловливают цвет глаз. У альбиносов пигмент отсутствует и радужка имеет красный цвет в связи с тем, что через ее толщу просвечивают кровеносные сосуды. В пожилом возрасте наблюдается депигментация радужки, и она делается более светлой.
Сосудистый слой (stratum vasculosum) состоит из многочисленных сосудов, пространство между которыми заполнено рыхлой волокнистой соединительной тканью с пигментными клетками.
Задний пограничный слой (stratum limitans posterius) не отличается по строению от переднего слоя.
Задний пигментный эпителий (epithelium posterius pigmentosum) является продолжением двухслойного эпителия сетчатки, покрывающего ресничное тело и отростки. Он включает диффероны видоизмененных глиоцитов и пигментоцитов.
Ресничное, или цилиарное, тело (corpus ciliare). Ресничное тело является производным сосудистой и сетчатой оболочек. Выполняет функцию фиксации хрусталика и изменения его кривизны, тем самым участвуя в акте
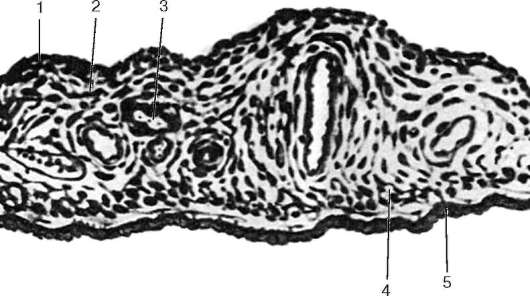
Рис. 12.4. Радужка:
1 - однослойный плоский эпителий; 2 - передний пограничный слой; 3 - сосудистый слой; 4 - задний пограничный слой; 5 - задний пигментный эпителий
аккомодации. На меридиональных срезах через глаз цилиарное тело имеет вид треугольника, который своим основанием обращен в переднюю камеру глаза. Ресничное тело подразделяется на две части: внутреннюю - ресничный венец (corona ciliaris) и наружную - ресничное кольцо (orbiculus ciliaris). От поверхности ресничного венца отходят по направлению к хрусталику ресничные отростки (processus ciliares), к которым прикрепляются волокна ресничного пояска (см. рис. 12.2). Основная часть ресничного тела, за исключением отростков, образована ресничной, или цилиарной, мышцей (m. cilia-ris), играющей важную роль в аккомодации глаза. Она состоит из пучков гладких мышечных клеток нейроглиальной природы, располагающихся в трех различных направлениях.
Различают наружные меридиональные мышечные пучки, лежащие непосредственно под склерой, средние радиальные и циркулярные мышечные пучки, образующие кольцевой мышечный слой. Между мышечными пучками расположена рыхлая волокнистая соединительная ткань с пигментными клетками. Сокращение цилиарной мышцы приводит к расслаблению волокон круговой связки - ресничного пояска хрусталика, вследствие чего хрусталик становится выпуклым и его преломляющая сила увеличивается.
Ресничное тело и ресничные отростки покрыты глиальным эпителием. Последний представлен двумя слоями: внутренний - непигментированные цилиндрические клетки - аналог мюллеровых волокон, наружный - продолжением пигментного слоя сетчатки. Эпителиальные клетки, покрывающие ресничное тело и отростки, принимают участие в образовании водянистой влаги, заполняющей обе камеры глаза.
Сосудистая оболочка (choroidea) осуществляет питание пигментного эпителия и нейронов, регулирует давление и температуру глазного яблока. В ней различают надсосудистую, сосудистую, сосудисто-капиллярную пластинки и базальный комплекс.
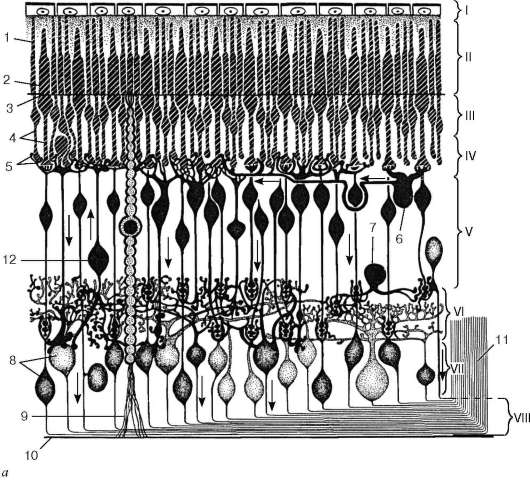
Рис. 12.5. Сетчатка:
а - схема нейронного состава сетчатки: 1 - палочки; 2 - колбочки; 3 - наружный пограничный слой; 4 - центральные отростки нейросенсорных клеток (аксоны);
5 - синапсы аксонов нейросенсорных клеток с дендритами биполярных нейронов;
6 - горизонтальный нейрон; 7 - амакринный нейрон; 8 - ганглиозные нейроны; 9 - радиальный глиоцит; 10 - внутренний пограничный слой; 11 - волокна зрительного нерва; 12 - центрифугальный нейрон
Надсосудистая пластинка (lamina suprachoroidea) толщиной 30 мкм представляет самый наружный слой сосудистой оболочки, прилежащий к склере. Она образована рыхлой волокнистой соединительной тканью, содержит большое количество пигментных клеток (меланоцитов), коллагеновых фибрилл, фибробластов, нервных сплетений и сосудов. Тонкие (диаметром 2-3 мкм) коллагеновые волокна этой ткани направлены от склеры к хороидее, параллельно склере, имеют косое направление в передней части, переходят в ресничную мышцу.
Сосудистая пластинка (lamina vasculosa) состоит из переплетающихся артерий и вен, между которыми располагаются рыхлая волокнистая соединительная ткань, пигментные клетки, отдельные пучки гладких миоцитов. Сосуды хороидеи являются ветвями задних коротких цилиарных артерий (орбитальные ветви глазной
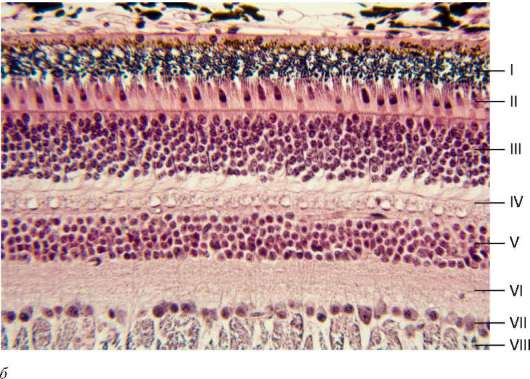
Рис. 12.5. Продолжение
б - микрофотография: I - пигментный эпителий сетчатки; II - палочки и колбочки нейросенсорных клеток; III - наружный ядерный слой; IV - наружный сетчатый слой; V - внутренний ядерный слой; VI - внутренний сетчатый слой; VII - слой ганглиозных нейронов; VIII - слой нервных волокон
артерии), которые проникают на уровне диска зрительного нерва в глазное яблоко, а также ветвями длинных цилиарных артерий.
Сосудисто-капиллярная пластинка (lamina choroicapillaris) содержит гемокапил-ляры висцерального или синусоидного типа, отличающиеся неравномерным калибром. Между капиллярами располагаются уплощенные фибробласты.
Базальный комплекс (complexus basalis) - мембрана Бруха (lamina vitrea, lamina elastica, membrana Brucha) - очень тонкая пластинка (1-4 мкм), располагающаяся между сосудистой оболочкой и пигментным слоем (эпителием) сетчатки. В ней различают наружный коллагеновый слой с зоной тонких эластических волокон, являющихся продолжением волокон сосудисто-капиллярной пластинки; внутренний коллагеновый слой, волокнистый (фиброзный), более толстый слой; третий слой представлен базальной мембраной пигментного эпителия. Через базальный комплекс в сетчатку поступают вещества, необходимые для нейросенсорных клеток.
Рецепторный аппарат глаза
Рецепторный аппарат глаза представлен зрительной частью сетчатой оболочки (сетчатки).
Внутренняя чувствительная оболочка глазного яблока, сетчатка (tunica interna sensoria bulbi, retina) состоит из наружного пигментного слоя (stratum pigmentosum) и внутреннего слоя нейросенсорных клеток (stratum nervosum) (рис. 12.5, а, б). Функционально выделяют заднюю большую зрительную часть сетчатки (pars
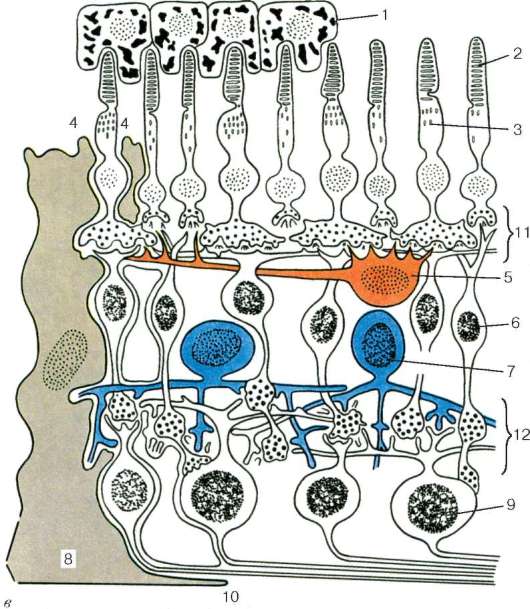
Рис. 12.5. Продолжение
в - синаптические связи в сетчатке (схема по Е. Бойкоту, Дж. Даулингу): 1 - пигментный слой; 2 - палочки; 3 - колбочки; 4 - зона расположения наружного пограничного слоя; 5 - горизонтальные нейроны; 6 - биполярные нейроны; 7 - ама-кринные нейроны; 8 - радиальные глиоциты; 9 - ганглиозные нейроны; 10 - зона расположения внутреннего пограничного слоя; 11 - синапсы между нейросенсор-ными клетками, биполярными и горизонтальными нейронами в наружном сетчатом слое; 12 - синапсы между биполярными, амакринными и ганглиозными нейронами во внутреннем сетчатом слое
optica retinae), меньшие части - ресничную, покрывающую ресничное тело (pars ciliares retinae), и радужковую, покрывающую заднюю поверхность радужки (pars iridica retina). В заднем полюсе глаза находится желтоватого цвета пятно (macula lutea) с небольшим углублением - центральной ямкой (fovea centralis).
Свет входит в глаз через роговицу, водянистую влагу передней камеры, хрусталик, жидкость задней камеры, стекловидное тело и, пройдя через толщу всех слоев сетчатки, попадает на отростки нейросенсорных клеток, в
наружных сегментах которых начинаются физиологические процессы возбуждения, фототрансдукции. Таким образом, сетчатка глаза человека относится к типу так называемых инвертированных органов, т. е. таких, в которых фоторецепторы направлены от света и образуют самые глубокие слои сетчатки, обращенные к слою пигментного эпителия.
Сетчатка состоит из трех типов радиально расположенных нейронов и двух слоев синапсов. Первый тип нейронов, расположенных наружно, - это палочковые и колбочко-вые нейроны, второй тип - биполярные нейроны, осуществляющие контакты между первым и третьим типом, третий тип - ганглиозные нейроны. Кроме того, имеются нейроны, осуществляющие горизонтальные связи, - горизонтальные и амакринные.
Наружный ядерный слой содержит тела палочковых и колбочковых нейронов, внутренний ядерный слой - тела биполярных, горизонтальных и ама-кринных нейронов, а слой ганглиоз-ных клеток - тела ганглиозных и перемещенных амакринных нейронов (см. рис. 12.5).
В наружном сетчатом слое контакты между колбочковыми нейронами и палочковыми нейронами осуществляются с вертикально ориентированными биполярными и горизонтально ориентированными горизонтальными нейронами. Во внутреннем сетчатом слое осуществляется переключение информации с вертикально ориентированных биполярных нейронов на ганглиоз-ные клетки, а также на различные виды вертикально и горизонтально направленные амакринные нейроны. В этом слое происходят кульми-
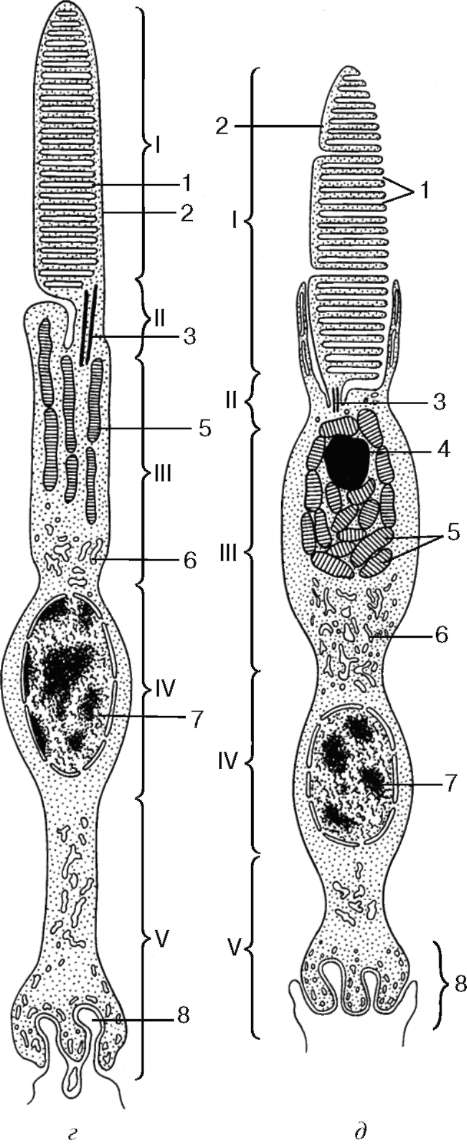
Рис. 12.5. Продолжение г, д - ультрамикроскопическое строение палочковой и колбочковой нейросенсорных клеток (схема по Ю. И. Афанасьеву):
I - наружный сегмент; II - связующий отдел; III - внутренний сегмент; IV - перикарион; V - аксон. 1 - диски (в палочках) и полудиски (в колбочках);
2 - плазмолемма; 3 - базальные тельца ресничек; 4 - липидное тело; 5 - митохондрии; 6 - эндоплазматическая сеть; 7 - ядро; 8 - синапс
нация всех интегральных процессов, связанных со зрительным образом, и передача информации через зрительный нерв в мозг. Через все слои сетчатки проходят радиальные глиальные клетки (клетки Мюллера).
В сетчатке выделяют также наружный пограничный слой, который состоит из множества описанных выше синаптических комплексов, расположенных между клетками Мюллера и нейросенсорными клетками; слой нервных волокон, который состоит из аксонов ганглиозных клеток. Последние, достигнув внутренней части сетчатки, поворачивают под прямым углом и затем идут параллельно внутренней поверхности сетчатки к месту выхода зрительного нерва. Они не содержат миелина и не имеют шванновских оболочек, что обеспечивает их прозрачность. Внутренний пограничный слой представлен окончаниями отростков клеток Мюллера и их базальными мембранами.
Нейросенсорные клетки делятся на два типа: палочковые и колбочковые (см. рис. 12.5). Палочковые нейроны являются рецепторами сумеречного (ночного зрения), колбочковые нейроны - рецепторами дневного зрения. Морфологически нейросенсорные клетки представляют собой длинные цилиндрической формы клетки, которые имеют несколько отделов. Дистальная часть рецепторов - это видоизмененная ресничка. Наружный сегмент (палочка или колбочка) - содержит фоторецепторные мембраны, где и происходит поглощение света и начинается зрительное возбуждение. Наружный сегмент связан с внутренним сегментом соединительной ножкой - ресничкой (цилией). Во внутреннем сегменте находятся множество митохондрий и полирибосом, цистерны комплекса Гольджи и небольшое количество элементов гранулярной и гладкой эндоплазматической сети. В этом сегменте происходит синтез белка. Далее сужающаяся часть клетки заполнена микротрубочками (миоид), затем идет расширенная часть с ядром. Тело клетки, расположенное проксимальнее внутреннего сегмента, переходит в аксональный отросток, который формирует синапс с дендри-тами биполярных и горизонтальных нейронов. Однако палочковые клетки отличаются от колбочковых клеток (см. рис. 12.5, г, д). У палочковых нейронов наружный сегмент цилиндрической формы, а диаметр внутреннего сегмента равен диаметру наружного. Наружные сегменты колбочковых клеток обычно конические, а внутренний сегмент по диаметру значительно превосходит наружный.
Наружный сегмент представляет собой стопку плоских мембранных мешочков - дисков, число которых доходит до 1000. В процессе эмбрионального развития диски палочек и колбочек образуются как складки - впячи-вания плазматической мембраны реснички.
В палочках новообразование складок продолжается у основания наружного сегмента в течение всей жизни. Вновь появившиеся складки оттесняют старые в дистальном направлении. При этом диски отрываются от плазмо-леммы и превращаются в замкнутые структуры, полностью отделенные от плазмолеммы наружного сегмента. Отработанные диски фагоцитируются клетками пигментного эпителия. Дистальные диски колбочек так же, как у палочек, фагоцитируются пигментными клетками.
Таким образом, фоторецепторный диск в наружном сегменте палочковых нейронов полностью отделен от плазматической мембраны. Он образован двумя фоторецепторными мембранами, соединенными по краям и внутри диска, на всем его протяжении имеется узкая щель. У края диска щель расширяется, образуется петля, внутренний диаметр которой составляет несколько десятков нанометров. Параметры диска: толщина - 15 нм, ширина внутридискового пространства - 1 нм, расстояние между дисками - междискового цитоплазматического пространства - 15 нм.
У колбочек в наружном сегменте диски не замкнуты и внутридисковое пространство сообщается с внеклеточной средой (см. рис. 12.5, д). У них более крупное округлое и светлое ядро, чем у палочек. Во внутреннем сегменте колбочек имеется участок, называемый эллипсоидом, состоящий из липидной капли и скопления плотно прилегающих друг к другу митохондрий. От ядросодержащей части нейросенсорных клеток отходят центральные отростки - аксоны, которые образуют синапсы с дендритами биполярных и горизонтальных нейронов, а также с карликовыми и плоскими биполярными нейронами. Длина колбочек в центре желтого пятна около 75 мкм, толщина - 1-1,5 мкм.
Толщина фоторецепторной мембраны наружного сегмента палочковых нейронов составляет около 7 нм. Основным белком фоторецепторной мембраны (до 95-98 % интегральных белков) является зрительный пигмент родопсин, который обеспечивает поглощение света и запускает фоторецеп-торный процесс.
Зрительный пигмент представляет собой хромогликопротеид. Эта сложная молекула содержит одну хромофорную группу, две олигосахаридные цепочки и водонерастворимый мембранный белок опсин. Хромофорной группой зрительных пигментов служит ретиналь-1 (альдегид витамина А) или ретиналь-2 (альдегид витамина А2). Все зрительные пигменты, содержащие ретиналь-1, относятся к родопсинам, а содержащие ретиналь-2 - к порфиропсинам. Светочувствительная молекула зрительного пигмента при поглощении одного кванта света претерпевает ряд последовательных превращений, в результате которых обесцвечивается. Фотолиз родопсина запускает каскад реакций, в результате происходит гиперполяризация нейрона и уменьшение выделения медиатора.
Среди колбочковых нейронов выделяют три типа, различающиеся зрительными пигментами с максимальной чувствительностью в длинноволновой (558 нм), средневолновой (531 нм) и коротковолновой (420 нм) части спектра. Один из пигментов - йодопсин - чувствителен к длинноволновой части спектра. Пигмент, чувствительный к коротковолновой части спектра, более сходен с родопсином. У человека гены, кодирующие пигмент коротковолновой части спектра и родопсина, находятся на длинном плече 3-й и 7-й хромосом и имеют сходство по структуре. Различные видимые нами цвета зависят от соотношения трех видов стимулируемых колбочковых нейронов.
Отсутствие длинно- и средневолновых колбочковых нейронов обусловлено соответствующими изменениями гена на Х-хромосоме, которые определяют два
типа дихромазии: протанопию и дейтеранопию. Протанопия - нарушение цветоощущения на красный цвет (ранее ошибочно называлось дальтонизмом). У Джона Дальтона благодаря последним достижениям молекулярной генетики выявлена дей-теранопия (нарушение цветоощущения на зеленый цвет).
Горизонтальные нервные клетки (neuron horisontalis) располагаются в один или два ряда. Они отдают множество дендритов, которые контактируют с аксонами нейросенсорных клеток. Аксоны горизонтальных нейронов, имеющие горизонтальную ориентацию, могут тянуться на довольно значительном расстоянии и вступать в контакт с аксонами как палочковых, так и колбоч-ковых нейронов. Передача возбуждения с горизонтальных клеток на синапсы нейросенсорной клетки и биполярного нейрона вызывает временную блокаду в передаче импульсов от фоторецепторов (эффект латерального торможения), что увеличивает контраст в зрительном восприятии.
Биполярные нервные клетки (neuron bipolaris) соединяют палочковые и кол-бочковые нейроны с ганглиозными нейронами сетчатки. В центральной части сетчатки несколько палочковых нейронов соединяются с одним биполярным нейроном, а колбочковые нейроны контактируют в соотношении 1:1 или 1:2. Такое сочетание обеспечивает более высокую остроту цветового видения по сравнению с черно-белым. Биполярные нейроны имеют радиальную ориентацию. Различают несколько видов биполярных нейронов по строению, содержанию синаптических пузырьков и связям с фоторецепторами (например, биполярные нейроны палочки, биполярные нейроны колбочки). Биполярные клетки играют существенную роль в концентрации импульсов, получаемых от нейросенсорных клеток и затем передаваемых ганглиозным нейронам.
Взаимоотношения биполярных нейронов с палочковыми и колбочко-выми нейронами различаются. Например, несколько палочковых клеток (15-20) в наружном сетчатом слое образуют синаптические связи с одним биполярным нейроном. Аксон последнего в составе внутреннего сетчатого слоя взаимодействует с различными типами амакринных нейронов, которые, в свою очередь, формируют синапсы с ганглиозным нейроном. Физиологический эффект заключается в ослаблении или усилении сигнала палочкового нейрона, что обусловливает чувствительность зрительной системы к единичному кванту света.
Амакринные клетки относятся к интернейронам, которые осуществляют связь на втором синаптическом уровне вертикального пути: нейросенсорная клетка → биполярный нейрон → ганглиозный нейрон. Их синаптическая активность во внутреннем сетчатом слое проявляется в интеграции, модуляции, включении сигналов, идущих к ганглиозным нейронам.
Эти клетки, как правило, не имеют аксонов, однако некоторые ама-кринные клетки содержат длинные аксоноподобные отростки. Синапсы амакринных клеток бывают химическими и электрическими. Например, дистальные дендриты амакринной клетки А образуют синапсы с аксонами палочковых биполярных нейронов, а проксимальные дендриты - с ган-глиозными нейронами. Более крупные дендриты А формируют электриче-
ские синапсы с аксонами колбочковых биполярных нейронов. В передаче нервного импульса от палочковых нейронов большую роль играют дофа-минергические и ГАМКергические амакринные клетки. Они ремодели-руют нервные импульсы и осуществляют обратную связь с палочковыми нейронами.
Ганглиозные нейроны - наиболее крупные клетки сетчатки, имеющие большой диаметр аксонов, способных проводить электрические сигналы. В их цитоплазме хорошо выражено хроматофильное вещество. Они собирают информацию от всех слоев сетчатки как по вертикальным путям (нейросен-сорные клетки → биполярные нейроны → ганглиозные нейроны), так и по латеральным путям (нейросенсорные клетки → горизонтальные нейроны → биполярные нейроны → амакринные нейроны → ганглиозные нейроны) и передают ее в мозг. Тела ганглиозных нейронов образуют ганглиозный слой (stratum ganglionicum), а их аксоны (более миллиона волокон) формируют внутренний слой нервных волокон (stratum neurofibrarum) и далее зрительный нерв. Ганглиозные нейроны гетероморфны. Они отличаются друг от друга по морфологическим и функциональным свойствам.
Нейроглия. Три дифферона глиальных клеток найдены в сетчатке человека: клетки Мюллера (радиальные глиоциты), протоплазматические астроциты и микроглиоциты. Через все слои сетчатки проходят длинные, узкие радиальные глиальные клетки. Их удлиненное ядро лежит на уровне ядер биполярных нейронов. Базальные отростки клеток участвуют в образовании внутреннего, а апикальные отростки - наружного пограничного слоя. Клетки регулируют ионный состав окружающей нейроны среды, участвуют в процессах регенерации, играют опорную и трофическую роль.
Пигментный слой, эпителий (stratum pigmentosum), наружный слой сетчатки - состоит из призматических полигональных пигментных клеток - пиг-ментоцитов. Своими основаниями клетки располагаются на базальной мембране, которая входит в состав мембраны Бруха сосудистой оболочки. Общее количество пигментных клеток, содержащих коричневые гранулы меланина, варьирует от 4 до 6 млн. В центре желтого пятна пигментоциты более высокие, а на периферии они уплощаются, становятся шире. Апикальные части плазмолеммы пигментных клеток контактируют непосредственно с дистальной частью наружных сегментов нейросенсорных клеток.
Апикальная поверхность пигментоцитов имеет два вида микроворсинок: длинные микроворсинки, которые располагаются между наружными сегментами нейросенсорных клеток, и короткие микроворсинки, которые взаимодействуют с концами наружных сегментов нейросенсорных клеток. Один пигментоцит контактирует с 30-45 наружными сегментами нейросен-сорных клеток, а вокруг одного наружного сегмента палочковых нейронов обнаруживается 3-7 отростков пигментоцитов, содержащих меланосомы, фагосомы и органеллы общего значения. В то же время вокруг наружного сегмента колбочкового нейрона - 30-40 отростков пигментоцитов, которые длиннее и не содержат органелл, за исключением меланосом. Фагосомы образуются в процессе фагоцитоза дисков наружных сегментов нейросен-сорных клеток.
Наличие пигмента в отростках (меланосом) обусловливает поглощение 85-90 % света, попадающего в глаз. Под воздействием света меланосомы перемещаются в апикальные отростки пигментоцитов, а в темноте мелано-сомы возвращаются в перикарион. Это перемещение происходит с помощью микрофиламентов при участии гормона меланотропина. Пигментный эпителий, располагаясь вне сетчатки, выполняет ряд важных функций: оптическую защиту и экранирование от света; транспорт метаболитов, солей, кислорода и т. п. из сосудистой оболочки к нейросенсорным клеткам и обратно, фагоцитоз дисков наружных сегментов нейросенсорных клеток и доставку материала для постоянного обновления плазматической мембраны последних; участие в регуляции ионного состава в субретинальном пространстве.
В пигментном эпителии велика опасность развития темновых и фотоокислительных деструктивных процессов. Все ферментативные и неферментативные звенья антиокислительной защиты присутствуют в клетках пигментного эпителия: пигментоциты участвуют в защитных реакциях, тормозящих перекисное окисление липидов с помощью ферментов микропероксисом и функциональных групп меланосом. Например, в них найдена высокая активность пероксидазы, как селе-нозависимой, так и селенонезависимой, и высокое содержание альфа-токоферола. Меланосомы в клетках пигментного эпителия, обладающие антиоксидантным свойством, служат специфическими участниками системы антиоксидантной защиты. Они эффективно связывают прооксидантные зоны (ионы железа) и не менее эффективно взаимодействуют с активными формами кислорода.
На внутренней поверхности сетчатки у заднего конца оптической оси глаза имеется округлое или овальное желтое пятно диаметром около 2 мм. Слегка углубленный центр этого образования называется центральной ямкой (fovea centralis) (рис. 12.6, а).
Центральная ямка - место наилучшего восприятия зрительных раздражений. В этой области внутренний ядерный и ганглиозный слои резко истончаются, а несколько утолщенный наружный ядерный слой представлен главным образом телами колбочковых нейронов.
Кнутри от центральной ямки (fovea centralis) имеется зона длиной 1,7 мм, в которой отсутствуют нейросенсорные клетки - слепое пятно, а аксоны ганглиозных нейронов формируют зрительный нерв. Последний при выходе из сетчатки через решетчатую пластинку склеры виден как диск зрительного нерва (discus nervi optici) с приподнятыми в виде валика краями и небольшим углублением в центре (excavatio disci).
Зрительный нерв - промежуточная часть зрительного анализатора. По нему информация о внешнем мире передается от сетчатки в центральные отделы зрительной системы. Впереди турецкого седла и воронки гипофиза волокна зрительного нерва образуют перекрест (хиазма), где волокна, идущие от носовой половины сетчатки, перекрещиваются, а идущие от вилочной части сетчатки не перекрещиваются. Далее в составе зрительного тракта перекрещенные и неперекрещенные нервные волокна направляются в латеральное коленчатое тело промежуточного мозга соответствующей гемисферы (подкорковые зрительные центры) и верхние холмики крыши среднего мозга. В латеральном коленчатом теле аксоны третьего
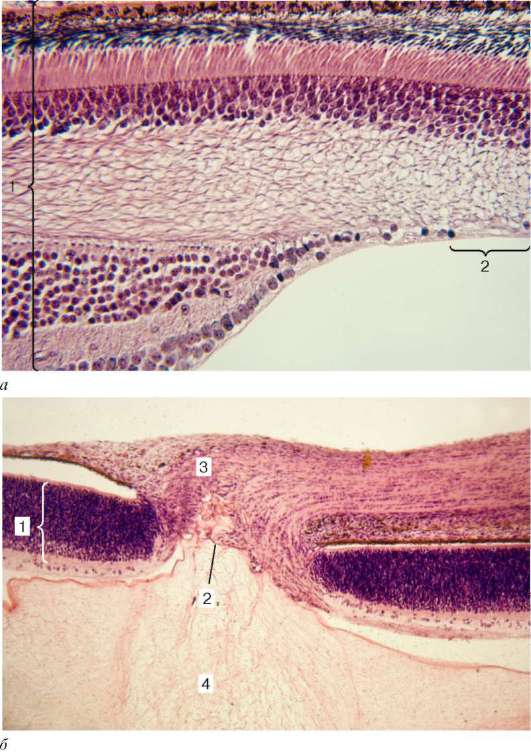
Рис. 12.6. Центральная ямка (а) и диск зрительного нерва (б):
а: 1 - сетчатка; 2 - центральная ямка (желтое пятно); б: 1 - сетчатка; 2 - диск зрительного нерва («слепое пятно»); 3 - зрительный нерв; 4 - стекловидное тело. Микрофотографии
нейрона заканчиваются и контактируют со следующим нейроном, аксоны которого, проходя под чечевицеобразную часть внутренней капсулы, формируют зрительную лучистость (radiatio optica), направляются в затылочную долю, зрительные центры, располагающиеся в области шпорной борозды, и в экстрастриарные зоны.
Регенерация сетчатки. Процессы физиологической регенерации палочковых и колбочковых нейронов происходят в течение всей жизни. Ежесуточно в каждой палочковой клетке ночью или в каждой колбочковой клетке днем
формируется около 80 мембранных дисков. Процесс обновления каждой палочковой клетки длится 9-12 сут.
Один пигментоцит ежесуточно фагоцитирует около 2-4 тыс. дисков, в нем образуется 60-120 фагосом, каждая из которых содержит 30-40 дисков.
Таким образом, пигментоциты обладают исключительно высокой фагоцитарной активностью, которая повышается при напряжении функции глаза в 10-20 раз и более.
Выявлены циркадные ритмы утилизации дисков: отделение и фагоцитоз сегментов палочковых клеток происходят обычно утром, а колбочковых - ночью.
В механизмах отделения отработанных дисков важную роль играет ретинол (витамин А), который в больших концентрациях накапливается в наружных сегментах палочковых клеток на свету и, обладая сильно выраженными мембранолитиче-скими свойствами, стимулирует указанный выше процесс. Циклические нуклеоти-ды (цАМФ) тормозят скорость деструкции дисков и их фагоцитоз. В темноте, когда цАМФ много, скорость фагоцитоза невелика, а на свету, когда содержание цАМФ снижено, она возрастает.
Васкуляризация. Ветви глазничной артерии формируют две группы разветвлений: одна образует ретинальную сосудистую систему сетчатки, васку-ляризующую сетчатку и часть зрительного нерва; вторая образует цилиар-ную систему, снабжающую кровью сосудистую оболочку, ресничное тело, радужку и склеру. Лимфатические капилляры располагаются только в склеральной конъюнктиве, в других участках глаза они не найдены.
Вспомогательный аппарат глаза
К вспомогательному аппарату глаза относятся глазные мышцы, веки и слезный аппарат.
Глазные мышцы. Они представлены поперечнополосатыми (исчерченными) мышечными волокнами миотомного происхождения, которые прикрепляются сухожилиями к склере и обеспечивают движение глазного яблока.
Веки (palpebrae). Веки развиваются из кожных складок, образующихся кверху и книзу от глазного бокала. Они растут по направлению друг к другу и спаиваются своим эпителиальным покровом. К 7-му мес внутриутробного развития спайка исчезает. Передняя поверхность век - кожная, задняя - конъюнктива - продолжается в конъюнктиву глаза (слизистую оболочку) (рис. 12.7). Внутри века, ближе к его задней поверхности, располагается тарзальная пластинка, состоящая из плотной волокнистой соединительной ткани. Ближе к передней поверхности в толще век залегает кольцевая мышца. Между пучками мышечных волокон располагается прослойка рыхлой соединительной ткани. В этой прослойке оканчивается часть сухожильных волокон мышцы, поднимающей верхнее веко.
Другая часть сухожильных волокон этой мышцы прикрепляется прямо к проксимальному краю тарзальной (соединительнотканной) пластинки. Наружная поверхность покрыта тонкой кожей, состоящей из тонкого многослойного плоского ороговевающего эпителия и рыхлой соединительной ткани, в которой залегают волосяные эпителиальные влагалища коротких пушковых волос, а также ресниц (по краям смыкающихся частей век).
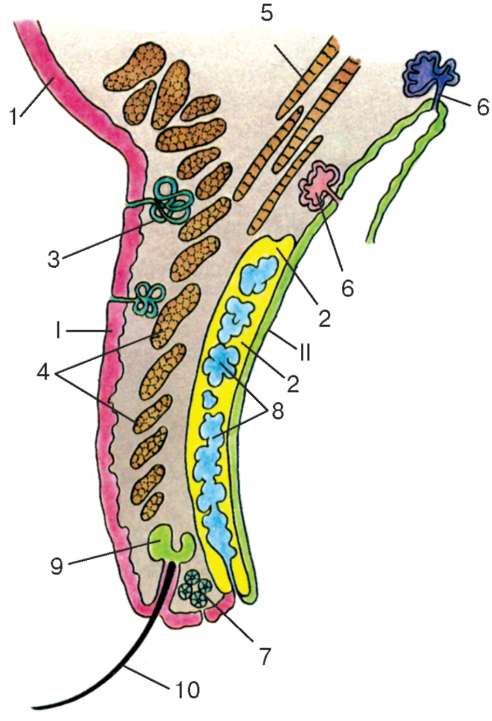
Рис. 12.7. Веко (сагиттальный срез): I - передняя (кожная поверхность); II - внутренняя поверхность (конъюнктива). 1 - многослойный плоский ороговеваю-щий эпителий (эпидермис) и соединительная ткань (дерма); 2 - рудиментарная хрящевая пластинка; 3 - трубчатые мерокринные потовые железы; 4 - круговая мышца века; 5 - мышца, поднимающая веко; 6 - слезные железы; 7 - апо-кринные потовые железы; 8 - простые трубчато-альвеолярные (мейбомиевы) железы, вырабатывающие сальный секрет; 9 - простые разветвленные альвеолярные голокринные (ресничные) железы, выделяющие сальный секрет; 10 - ресница
В соединительной ткани кожи находятся мелкие трубчатые мерокринные потовые железы. Около волосяных фолликулов встречаются апокринные потовые железы. В воронку корня ресницы открываются мелкие простые разветвленные сальные железы. Вдоль внутренней поверхности века, покрытой конъюнктивой, располагаются 20-30 и более особого вида простых разветвленных трубчато-альвеолярных голокринных (мейбомие-вых) желез (в верхнем веке их больше, чем в нижнем), вырабатывающих сальный секрет. Над ними и в области свода ( fornix) лежат мелкие слезные железы. Центральная часть века на всем его протяжении состоит из плотной волокнистой соединительной ткани и пучков волокон поперечнополосатой мышечной ткани, ориентированных по вертикали (m. levator palpebrae superioris), а вокруг глазной щели кольцевая мышца (m. orbicularis oculi). Сокращения этих мышц обеспечивают смыкание век, а также смазывание передней поверхности глазного яблока слезной жидкостью и липидным секретом желез.
Сосуды века образуют две сети - кожную и конъюнктивальную. Лимфатические сосуды формируют третье дополнительное, тарзальное сплетение.
Конъюнктива - тонкая прозрачная слизистая оболочка, которая покрывает заднюю поверхность век
и переднюю часть глазного яблока. В области роговицы конъюнктива срастается с ней. Многослойный не-ороговевающий эпителий располагается на соединительнотканной основе. В составе эпителия находятся бокаловидные клетки, вырабатывающие слизь. Под эпителием в соединительной ткани конъюнктивы в области век имеется хорошо выраженная капиллярная сеть, способствующая всасыванию лекарственных препаратов (капель, мазей), которые наносятся на поверхность конъюнктивы.
Слезный аппарат глаза. Он состоит из слезопродуцирующей слезной железы и слезоотводящих путей - слезное мясцо, слезные канальцы, слезный мешок и слезно-носовой канал.
Слезная железа располагается в слезной ямке глазницы и состоит из нескольких групп сложных альвеолярно-трубчатых серозных желез. Концевые отделы включают диффероны секреторных клеток (лакримоци-тов) и миоэпителиоцитов. Слабощелочной секрет слезных желез содержит около 1,5 % хлорида натрия, незначительное количество альбумина (0,5 %), лизоцим, оказывающий бактерицидное действие, и IgA. Слезная жидкость увлажняет и очищает роговицу глаза. Она непрерывно выделяется в верхний конъюнктивальный свод, а оттуда движением век направляется на роговицу, медиальный угол глазной щели, где образуется слезное озерцо. Сюда открываются устья верхнего и нижнего слезных канальцев, каждый из которых впадает в слезный мешок, а он продолжается в слезно-носовой проток, открывающийся в нижний носовой ход. Стенки слезного мешка и слезно-носового протока выстланы двух- и многорядным эпителием.
Возрастные изменения. С возрастом ослабляется функция всех аппаратов глаза. В связи с изменением общего метаболизма в организме в хрусталике и роговице часто происходят уплотнение межклеточного вещества и помутнение, которое практически необратимо. У пожилых людей откладываются липиды в роговице и склере, что обусловливает их потемнение. Утрачивается эластичность хрусталика, и ограничивается его аккомодационная возможность. Склеротические процессы в сосудистой системе глаза нарушают трофику тканей, особенно сетчатки, что приводит к изменению структуры и функции рецепторного аппарата.
12.3. ОРГАНЫ ОБОНЯНИЯ
Обоняние - самый древний вид сенсорного восприятия. Обонятельный анализатор представлен двумя системами - основной и вомероназальной, каждая из которых имеет три части: периферическую (органы обоняния), промежуточную, состоящую из проводников (аксоны обонятельных нейро-сенсорных эпителиоцитов и нервных клеток обонятельных луковиц), и центральную, локализующуюся в обонятельном центре коры большого мозга.
Основной орган обоняния (organum ofactus), являющийся периферической частью сенсорной системы, представлен ограниченным участком слизистой оболочки носа - обонятельной областью, покрывающей у человека верхнюю и отчасти среднюю раковины носовой полости, а также верхнюю часть носовой перегородки. Внешне обонятельная область отличается от респираторной части слизистой оболочки желтоватым цветом.
Периферической частью вомероназальной, или дополнительной, обонятельной системы является вомероназальный (якобсонов) орган (organum vomeronasale Jacobsoni). Он имеет вид парных эпителиальных трубок, замкнутых с одного конца и открывающихся другим концом в полость носа.
У человека вомероназальный орган расположен в соединительной ткани основания передней трети носовой перегородки по обе ее стороны на границе между хрящом перегородки и сошником. Кроме якобсонова органа, вомероназальная система включает вомероназальный нерв, терминальный нерв и собственное представительство в переднем мозге - добавочную обонятельную луковицу. Этот орган хорошо развит у пресмыкающихся и млекопитающих. Обонятельные нейросенсорные эпителиоциты специализированы на восприятии феромонов (веществ, выделяемых специализированными железами).
Функции вомероназальной системы связаны с функциями половых органов (регуляция полового цикла и сексуального поведения) и эмоциональной сферой.
Развитие. Источником образования всех частей органа обоняния являются обособляющаяся часть нейроэктодермы, симметричные локальные утолщения эктодермы - обонятельные плакоды, расположенные в области передней части головы зародыша, и мезенхима. Материал плакоды впячивается в подлежащую мезенхиму, формируя обонятельные мешки, связанные с внешней средой посредством отверстий (будущие ноздри). В составе стенки обонятельного мешка находятся обонятельные стволовые клетки, которые на 4-м мес внутриутробного развития путем дивергентной дифференцировки развиваются в нейросенсорные (обонятельные) клетки, поддерживающие и базальные эпителиоциты. Часть клеток обонятельного мешка идет на построение обонятельной (боуменовой) железы. В дальнейшем центральные отростки нейросенсорных клеток, объединившись между собой, образуют в совокупности 20-40 нервных пучков (обонятельных путей - fila olfactoria), устремляющихся через отверстия в хрящевой закладке будущей решетчатой кости к обонятельным луковицам головного мозга. Здесь осуществляется синаптический контакт между терминалями аксонов и дендритами митральных нейронов обонятельных луковиц.
Вомероназальный орган формируется в виде парной закладки на 6-й нед развития в нижней части перегородки носа. К 7-й нед развития завершается формирование полости вомероназального органа, а вомероназальный нерв соединяет его с добавочной обонятельной луковицей. В вомероназальном органе плода 21-й нед развития имеются поддерживающие эпителиоциты с ресничками и микроворсинками и обонятельные нейросенсорные эпите-лиоциты с микроворсинками. Структурные особенности вомероназального органа указывают на его функциональную активность уже в перинатальном периоде (рис. 12.8, 12.9).
Строение. Основной орган обоняния - периферическая часть обонятельного анализатора - состоит из пласта многорядного цилиндрического эпителия высотой 60-90 мкм, в котором различают обонятельные нейросен-сорные клетки, поддерживающие и базальные эпителиоциты (рис. 12.10, А, Б). От подлежащей соединительной ткани они отделены хорошо выраженной базальной мембраной. Обращенная в носовую полость поверхность обонятельной выстилки покрыта слоем слизи.
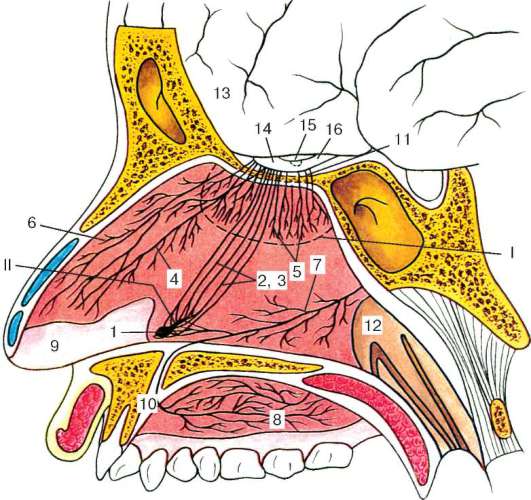
Рис. 12.8. Топография рецепторных полей и проводящих путей обонятельных анализаторов. Сагиттальное сечение головы человека на уровне носовой перегородки (по В. И. Гулимовой):
I - рецепторное поле основного органа обоняния (обозначено пунктирной линией);
II - рецепторное поле вомероназального органа. 1 - вомероназальный орган; 2 - вомероназальный нерв; 3 - терминальный нерв; 4 - передняя ветвь терминального нерва; 5 - волокна обонятельного нерва; 6 - внутренние носовые ветви решетчатого нерва; 7 - носонёбный нерв; 8 - нёбные нервы; 9 - слизистая оболочка носовой перегородки; 10 - носонёбный канал; 11 - отверстия решетчатой пластинки; 12 - хоана; 13 - передний мозг; 14 - основная обонятельная луковица; 15 - дополнительная обонятельная луковица; 16 - обонятельный тракт
Нейросенсорные, или рецепторные, обонятельные эпителиоциты (epithe-liocyti neurosensoriae olfactoriae) располагаются между поддерживающими эпителиоцитами и имеют короткий периферический отросток - дендрит и длинный - центральный - аксон. Их ядросодержащие части занимают, как правило, срединное положение в толще обонятельной выстилки.
У собак, которые отличаются хорошо развитым органом обоняния, насчитывается около 225 млн обонятельных клеток, у человека их число значительно меньше, но все же достигает 6 млн (30 тыс. на 1 мм2). Различают два вида обонятельных клеток. У одних клеток дистальные части периферических отростков заканчиваются характерными утолщениями - обонятельными булавами, или луковицами дендритов (clava olfactoria). Меньшая часть клеток обонятельного эпителия имеет обонятельные микроворсинки (микровиллы).
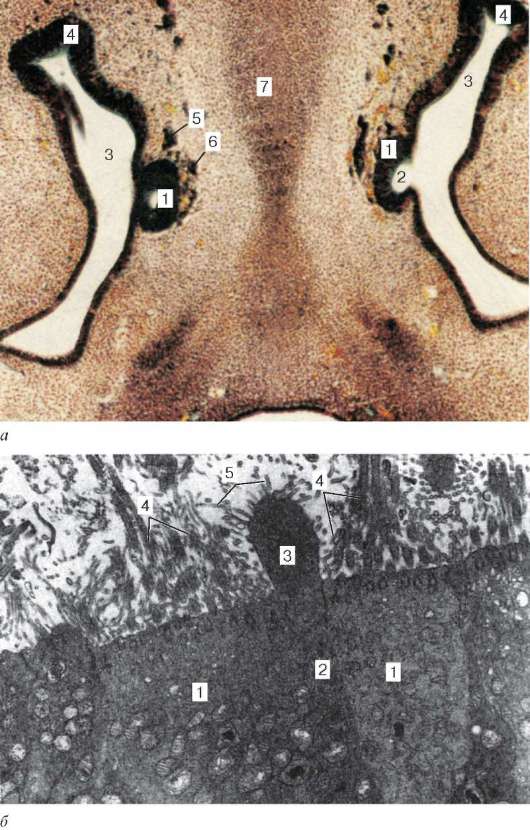
Рис. 12.9. Развитие вомероназального органа у эмбриона человека (по В. И. Гу-лимовой):
а - микрофотография поперечного среза головы эмбриона 7 нед развития, окраска по Маллори: 1 - вомероназальный орган; 2 - полость вомероназального органа; 3 - полость носа; 4 - слизистая оболочка стенки носовой полости; 5 - вомероназальный нерв; 6 - терминальный нерв; 7 - закладка носовой перегородки; б - электронная микрофотография вомероназального эпителия плода человека 21 нед развития (увеличение 12 000): 1 - опорные клетки; 2 - нейросенсорный эпителиоцит; 3 - булава нейросенсорного эпителиоцита; 4 - реснички; 5 - микроворсинки
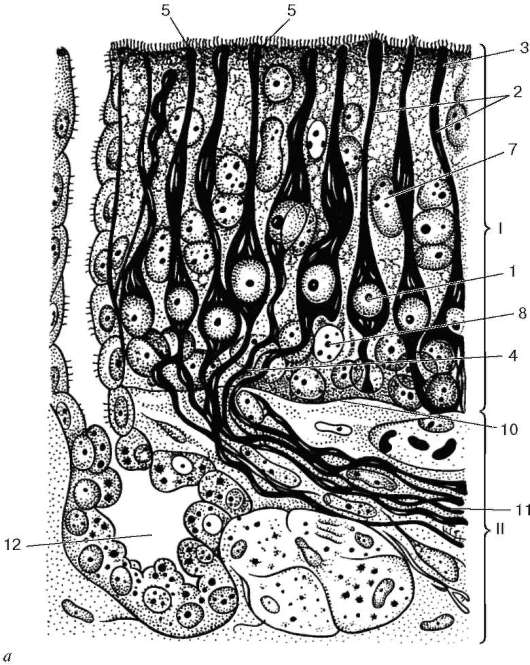
Рис. 12.10. Строение обонятельного эпителия (схема):
а - микроскопическое строение (по Я. А. Винникову и Л. К. Титовой); б - ультрамикроскопическое строение (по А. А. Бронштейну, с изменениями); в - регенерация обонятельных нейросенсорных эпителиоцитов (по Л. Арденсу): А, Б, В - дифференцирующаяся нейросенсорная клетка; Г, Д - разрушающаяся клетка. I - обонятельный эпителий; II - собственная пластинка слизистой оболочки. 1 - нейросенсор-ные клетки; 2 - периферические отростки (дендриты); 3 - обонятельные луковицы дендритов; 4 - центральные отростки (аксоны); 5 - обонятельные реснички; 6 - микроворсинки; 7 - поддерживающие эпителиоциты; 8 - базальные эпителиоциты; 9 - малодифференцированные нейроны; 10 - базальная мембрана; 11 - нервные стволики - аксоны нейросенсорных клеток; 12 - обонятельная железа
Обонятельные булавы нейросенсорных клеток на своей округлой вершине несут до 10-12 подвижных обонятельных ресничек (см. рис. 12.10, Б, В). Реснички содержат продольно ориентированные фибриллы: 9 пар периферических и 2 - центральных, отходящих от базальных телец. Обонятельные реснички подвижны и являются своеобразными антеннами для молекул
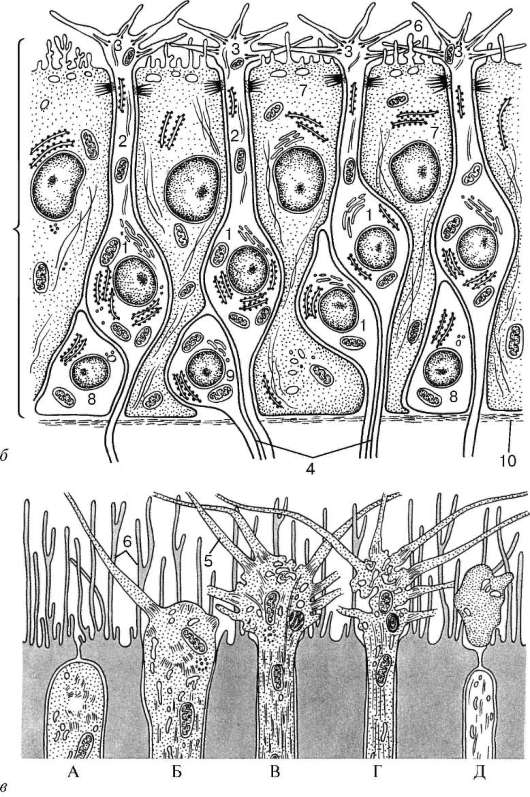
Рис. 12.10. Продолжение
пахучих веществ. Периферические отростки обонятельных клеток могут сокращаться под действием пахучих веществ. Ядра обонятельных нейросенсорных клеток светлые, с одним или двумя крупными ядрышками. Около ядра отчетливо выявляется гранулярная эндоплазматическая сеть. Базальная часть клетки продолжается в тонкий, слегка извивающийся аксон, который проходит между поддерживающими эпителиоцитами.
Обонятельные клетки с микроворсинками сходны по строению с описанными выше нейросенсорными клетками с булавой. Микроворсинки слу-
жат для увеличения мембранной поверхности клетки, воспринимающей запахи. В соединительнотканном слое центральные отростки нейросенсор-ных клеток формируют пучки безмиелинового обонятельного нерва.
Поддерживающие эпителиоциты (epitheliocytus sustentans) - глиальные по происхождению, формируют эпителиальный пласт, в котором и располагаются нейросенсорные эпителиоциты. На апикальной поверхности поддерживающих эпителиоцитов имеются многочисленные микроворсинки длиной до 2 мкм. Поддерживающие эпителиоциты проявляют признаки апокринной секреции и обладают высоким уровнем метаболизма. В цитоплазме обнаруживается гранулярная эндоплазматическая сеть. Митохондрии большей частью скапливаются в апикальной части, где находится также большое число гранул и вакуолей. Комплекс Гольджи располагается над овальным ядром. Подъядерная часть клетки сужается, достигая базальной мембраны в промежутках между базальными эпителиоцитами. В цитоплазме поддерживающих клеток содержится коричнево-желтый пигмент.
Базальные эпителиоциты (epitheliocytus basales) кубической формы находятся на базальной мембране и снабжены цитоплазматическими выростами, окружающими пучки центральных отростков обонятельных клеток. Цитоплазма их заполнена рибосомами и не содержит тонофибрилл. Базальные эпителиоциты относятся к камбию обонятельного эпителия и служат источником регенерации его клеток.
Эпителий вомероназального органа состоит из рецепторной и респираторной частей. Рецепторная часть по строению сходна с обонятельным эпителием основного органа обоняния. Главное отличие состоит в том, что обонятельные булавы нейросенсорных эпителиоцитов вомероназального органа несут на своей поверхности неподвижные микроворсинки.
Промежуточная, или проводниковая, часть основной обонятельной сенсорной системы начинается обонятельными безмиелиновыми нервными волокнами, которые объединяются в 20-40 нитевидных стволиков (fila olfactoria) и через отверстия решетчатой кости направляются в обонятельные луковицы (см. рис. 12.10). Каждая обонятельная нить представляет собой безмиелиновое волокно, содержащее от 20 до 100 и более осевых цилиндров аксонов нейросенсорных эпителиоцитов, погруженных в цитоплазму лем-моцитов. В обонятельных луковицах расположены вторые нейроны обонятельного анализатора. Это крупные нервные клетки, называемые митральными, имеют синаптические контакты с несколькими тысячами аксонов нейросенсорных клеток одноименной, а частично и противоположной стороны. Обонятельные луковицы построены по типу коры полушарий большого мозга, имеют концентрически расположенные шесть слоев: 1 - слой обонятельных клубочков; 2 - наружный зернистый слой; 3 - молекулярный слой; 4 - слой тел митральных нейронов; 5 - внутренний зернистый слой; 6 - слой центробежных волокон.
Контакт аксонов нейросенсорных эпителиоцитов с дендритами митральных нейронов происходит в клубочковом слое, где суммируются возбуждения рецеп-торных клеток. Здесь же осуществляется взаимодействие рецепторных клеток между собой и с мелкими ассоциативными клетками. В обонятельных клубочках
реализуются и центробежные эфферентные влияния, исходящие из вышележащих эфферентных центров (переднее обонятельное ядро, обонятельный бугорок, ядра миндалевидного тела, препириформная кора). Наружный зернистый слой образован телами пучковых нейронов и многочисленными синапсами с дополнительными дендритами митральных нейронов, аксонами межклубочковых клеток и дендро-дендритическими синапсами митральных нейронов. В 4-м слое лежат тела митральных нейронов. Их аксоны проходят через 4-5-й слои луковиц, а на выходе из них образуют обонятельные контакты вместе с аксонами пучковых клеток. В области 6-го слоя от аксонов митральных нейронов отходят возвратные коллатерали, распределяющиеся в разных слоях. Внутренний зернистый слой образован скоплением нейронов, которые по своей функции являются тормозными. Их дендриты образуют синапсы с возвратными коллатералями аксонов митральных нейронов.
Промежуточная, или проводниковая, часть вомероназальной системы представлена безмиелиновыми волокнами вомероназального нерва, которые, подобно основным обонятельным волокнам, объединяются в нервные стволики, проходят через отверстия решетчатой кости и соединяются с добавочной обонятельной луковицей, которая расположена в дорсомедиаль-ной части основной обонятельной луковицы и имеет сходное строение.
Центральный отдел обонятельной сенсорной системы локализуется в древней коре - в гиппокампе и в новой - гиппокамповой извилине, куда направляются аксоны митральных нейронов (обонятельный тракт). Здесь происходит окончательный анализ обонятельной информации (расшифровка кода запаха).
Сенсорная обонятельная система через ретикулярную формацию связана с автономной нервной системой, иннервирующей органы пищеварительной и дыхательной систем, что объясняет рефлекторные реакции со стороны последних на запахи.
Обонятельные железы. В подлежащей рыхлой волокнистой ткани обонятельной области располагаются концевые отделы трубчато-альвеолярных обонятельных (боуменовых) желез (см. рис. 12.10), выделяющие секрет, который содержит большое количество белков, олигонуклеотидов, гликозаминоглика-нов и др. В слизи обнаружены одорантсвязывающие белки - неспецифические переносчики пахучих молекул. В концевых отделах желез снаружи лежат уплощенные клетки - миоэпителиальные, внутри - клетки, секретирующие по мерокринному типу. Прозрачный, водянистый секрет желез вместе с секретом поддерживающих эпителиоцитов увлажняет поверхность обонятельной слизистой оболочки, что является необходимым условием для функционирования нейросенсорных эпителиоцитов. В этом секрете, омывающем обонятельные реснички нейросенсорной клетки, растворяются пахучие вещества, присутствие которых только в этом случае и воспринимается рецепторными белками, вмонтированными в плазмолемму ресничек. Каждый запах вызывает электрический ответ многих нейросенсорных эпителиоцитов обонятельной выстилки, в которой возникает мозаика электрических сигналов. Эта мозаика индивидуальна для каждого запаха и является кодом запаха.
Васкуляризация. Слизистая оболочка полости носа обильно снабжена кровеносными и лимфатическими сосудами. Сосуды микроциркуляторного
типа напоминают кавернозные тела. Кровеносные капилляры синусоидно-го типа образуют сплетения, которые способны депонировать кровь. При действии резких температурных раздражителей и молекул пахучих веществ слизистая оболочка носа может сильно набухать и покрываться значительным слоем слизи, что затрудняет рецепцию.
Возрастные изменения. Чаще всего они обусловлены перенесенными в течение жизни воспалительными процессами (риниты), которые приводят к атрофии рецепторных клеток и разрастанию респираторного эпителия.
Регенерация. У млекопитающих в постнатальном периоде онтогенеза обновление рецепторных обонятельных клеток происходит в течение 30 сут. В конце жизненного цикла нейросенсорные эпителиоциты подвергаются деструкции, фагоцитируются поддерживающими эпителиоцитами. Малодифференцированные нейроны базального слоя способны к мито-тическому делению, лишены отростков. В процессе их дифференцировки увеличивается объем клеток, появляются специализированный дендрит, растущий к поверхности, и аксон, растущий в сторону базальной мембраны, который в дальнейшем устанавливает контакт с митральным нейроном обонятельной луковицы. Клетки постепенно перемещаются к поверхности, замещая погибшие нейросенсорные эпителиоциты. На дендрите формируются специализированные структуры (микроворсинки и реснички). При некоторых вирусных поражениях обонятельных клеток не происходит их восстановление и обонятельная область замещается респираторным эпителием.
12.4. ОРГАН ВКУСА
Орган вкуса (organum gustus) - периферическая часть вкусового анализатора представлена рецепторными эпителиальными клетками во вкусовых почках (caliculi gustatoriae). Они воспринимают вкусовые (пищевые и непищевые) раздражения, генерируют и передают рецепторный потенциал афферентным нервным окончаниям, в которых появляются нервные импульсы. Информация поступает в подкорковые и корковые центры. При участии сенсорной системы обеспечиваются такие реакции, как отделение секрета слюнных желез, выделение желудочного сока и другое, поведенческие реакции на поиск пищи и т. п. Вкусовые почки располагаются в многослойном плоском эпителии боковых стенок желобоватых, листовидных и грибовидных сосочков языка человека (рис. 12.11). У детей, а иногда и у взрослых вкусовые почки могут находиться на губах, задней стенке глотки, нёбных дужках, наружной и внутренней поверхностях надгортанника. Количество вкусовых почек у человека достигает 2000.
Развитие органа вкуса. Вкусовые почки начинают развиваться на 6- 7-й нед эмбриогенеза человека. Они формируются как выпячивания слизистой оболочки языка на его дорсальной поверхности. Источником развития сенсоэпителиальных клеток вкусовых почек является многослойный
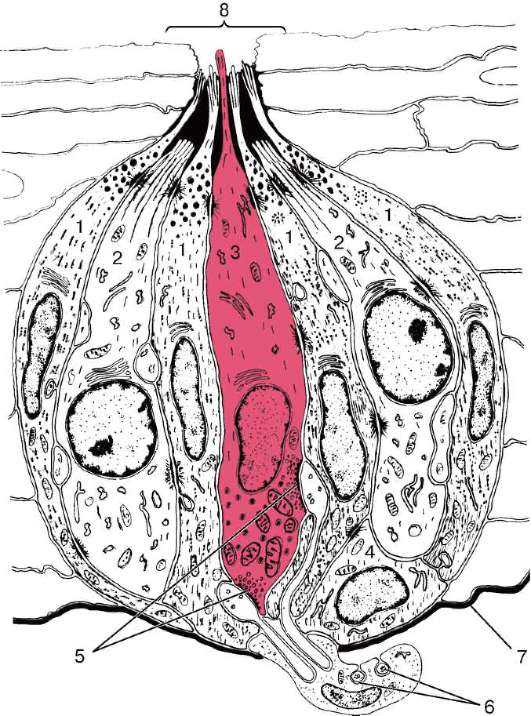
Рис. 12.11. Вкусовая почка:
1 - вкусовой эпителиоцит I типа; 2 - вкусовой эпителиоцит II типа; 3 - вкусовой эпителиоцит III типа; 4 - вкусовой эпителиоцит IV типа; 5 - синаптические контакты с клеткой III типа; 6 - нервные волокна, окруженные леммоцитом; 7 - базальная мембрана; 8 - вкусовая пора
эпителий сосочков языка. Он подвергается дифференцировке под индуцирующим воздействием окончаний нервных волокон язычного, языкоглоточ-ного и блуждающего нервов. В результате дивергентной дифференцировки малодифференцированных предшественников возникают различные типы вкусовых эпителиоцитов. Таким образом, иннервация вкусовых почек появляется одновременно с возникновением их зачатков.
Строение. Каждая вкусовая почка имеет эллипсоидную форму размером 27-115 мкм в высоту и 16-70 мкм в ширину и занимает всю толщу многослойного эпителиального пласта сосочка языка. Она состоит из плотно прилежащих друг к другу 40-60 гетероморфных эпителиоцитов различного типа. От подлежащей соединительной ткани вкусовая почка отделяется базальной мембраной. Вершина почки сообщается с поверхностью языка при помощи вкусовой поры (porus gustatorius). Вкусовая пора ведет в неболь-
шое углубление между поверхностными эпителиальными клетками сосочков - вкусовую ямку (см. рис. 12.11).
Среди вкусовых клеток различают несколько морфофункциональных типов. Вкусовые эпителиоциты I типа на своей апикальной поверхности имеют до 40 микроворсинок, являющихся адсорбентами вкусовых раздражителей. В цитоплазме обнаруживаются многочисленные электронно-плотные гранулы, гранулярная эндоплазматическая сеть, митохондрии, пучки микрофиламентов и микротрубочек цитоскелета. Все это придает цитоплазме темный вид.
Вкусовые эпителиоциты II типа имеют светлую цитоплазму, в которой обнаруживаются цистерны гладкой эндоплазматической сети, лизосомы и мелкие вакуоли. Апикальная поверхность содержит мало микроворсинок. Вышеперечисленные клетки не формируют синаптических контактов с нервными волокнами и относятся к поддерживающим.
Вкусовые эпителиоциты III типа, относительная доля которых во вкусовой почке равна 5-7 %, характеризуются наличием в цитоплазме пузырьков диаметром 100-200 нм с электронно-плотной сердцевиной. На апикальной поверхности клетки находится крупный отросток с микроворсинками, проходящими через вкусовую пору. Эти клетки формируют синапсы с афферентными волокнами и являются сенсоэпителиальными.
Вкусовые эпителиоциты IV типа (базальные) располагаются в базальной части вкусовой почки. Эти малодифференцированные клетки характеризуются небольшим объемом цитоплазмы вокруг ядра и слабым развитием органелл. В клетках выявляются фигуры митоза. Базальные клетки в отличие от сенсоэпителиальных и поддерживающих клеток никогда не достигают поверхности эпителиального слоя. Базальные клетки относятся к камбиальным.
Периферические (перигеммальные) клетки имеют серповидную форму, содержат мало органелл, но в них много микротрубочек и они связаны с нервными окончаниями.
Во вкусовой ямке между микроворсинками находится электронно-плотное вещество с высокой активностью фосфатаз и значительным содержанием рецепторного белка и гликопротеидов, которое играет роль адсорбента для вкусовых веществ, попадающих на поверхность языка. Энергия внешнего воздействия трансформируется в рецепторный потенциал. Под его влиянием из сенсоэпителиальной клетки (эпителиоцита III типа) выделяется медиатор (серотонин или норэпинефрин), который, действуя на нервное окончание сенсорного нейрона, вызывает в нем генерацию нервного импульса. Нервный импульс передается далее в промежуточную часть анализатора.
Во вкусовых почках передней части языка обнаружен сладкочувствитель-ный рецепторный белок, а в задней части - горькочувствительный. Вкусовые вещества адсорбируются на примембранном слое плазмолеммы микроворсинок, в которую вмонтированы специфические рецепторные белки. Одна и та же вкусовая клетка способна воспринимать несколько вкусовых раздражений. При адсорбции воздействующих молекул происходят конформа-ционные изменения рецепторных белковых молекул, которые приводят к
локальному изменению проницаемости мембран сенсоэпителиальной клетки и деполяризации или гиперполяризации плазмолеммы.
В каждую вкусовую почку входит и разветвляется около 50 афферентных нервных волокон, формирующих синапсы с базальными отделами сен-соэпителиальных клеток. На одной сенсоэпителиальной клетке могут быть окончания нескольких нервных волокон, а одно волокно кабельного типа может иннервировать несколько вкусовых почек. В формировании вкусовых ощущений принимают участие неспецифические афферентные окончания (тактильные, болевые, температурные), имеющиеся в слизистой оболочке полости рта, глотке, возбуждение которых добавляет окраску вкусовых ощущений («острый вкус перца» и др.).
Промежуточная часть вкусового анализатора. Центральные отростки ганглиев лицевого, языкоглоточного и блуждающего нервов вступают в ствол головного мозга к ядру одиночного пути, где находится второй нейрон вкусового пути. Здесь может происходить переключение импульсов на эфферентные пути к мимической мускулатуре, слюнным железам, к мышцам языка. Большая часть аксонов ядра одиночного пути достигает таламуса, где находится 3-й нейрон вкусового пути, аксоны которого заканчиваются на 4-м нейроне в коре большого мозга нижней части постцентральной извилины (центральная часть вкусового анализатора). Здесь формируются вкусовые ощущения.
Регенерация. Сенсорные и поддерживающие эпителиоциты вкусовой почки непрерывно обновляются. Продолжительность их жизни примерно 10 сут. При разрушении вкусовых эпителиоцитов нейроэпителиальные синапсы прерываются и вновь образуются на новых сенсоэпителиальных клетках.
12.5. ОРГАН СЛУХА И РАВНОВЕСИЯ
Орган слуха и равновесия, или преддверно-улитковый орган (organum vestibulo-cochleare), - наружное, среднее и внутреннее ухо, осуществляющее восприятие звуковых, гравитационных и вибрационных стимулов, линейных и угловых ускорений.
12.5.1. Наружное ухо
Наружное ухо (auris externa) включает ушную раковину, наружный слуховой проход и барабанную перепонку.
Ушная раковина (auricular) состоит из тонкой пластинки эластического хряща, покрытой кожей с немногочисленными тонкими волосами и сальными железами. Потовых желез в ее составе мало.
Наружный слуховой проход образован хрящом, являющимся продолжением эластического хряща раковины, и костной частью. Поверхность прохода покрыта тонкой кожей, содержащей волосы и связанные с ними сальные желе-
зы. Глубже сальных желез расположены трубчатые церуминозные (сальные) железы (glandula ceruminosa), выделяющие ушную серу, обладающую бактерицидными свойствами. Их протоки открываются самостоятельно на поверхности слухового прохода или в выводные протоки сальных желез. Количество желез уменьшается по мере приближения к барабанной перепонке.
Барабанная перепонка (membrana tympanica) овальной, слегка вогнутой формы, толщиной 0,1 мм. Одна из слуховых косточек среднего уха - молоточек - сращена с помощью своей ручки с внутренней поверхностью барабанной перепонки. От молоточка к барабанной перепонке проходят кровеносные сосуды и нервы. Барабанная перепонка в средней части состоит из двух слоев, образованных пучками коллагеновых и эластических волокон и залегающими между ними фибробластами. Волокна наружного слоя расположены радиально, а внутреннего - циркулярно. В верхней части барабанной перепонки количество коллагеновых волокон уменьшается (перепонка Шрапнеля). На наружной ее поверхности располагается очень тонким слоем (50-60 мкм) многослойный плоский эпителий, на внутренней поверхности, обращенной в среднее ухо, - слизистая оболочка толщиной около 20-40 мкм, покрытая однослойным плоским эпителием.
12.5.2. Среднее ухо
Среднее ухо (auris media) состоит из барабанной полости, слуховых косточек и слуховой (евстахиевой) трубы.
Барабанная полость - уплощенное пространство объемом около 2 см3, выстланное слизистой оболочкой. Эпителий - однослойный плоский, местами переходящий в кубический или цилиндрический. В слизистой оболочке и костных стенках среднего уха проходят ветки лицевого, языко-глоточного, блуждающего нервов. На медиальной стенке барабанной полости имеются два отверстия, или «окна». Первое - овальное окно. В нем располагается основание стремечка, которое удерживается с помощью тонкой связки по окружности окна. Овальное окно отделяет барабанную полость от вестибулярной лестницы улитки. Второе окно круглое, находится несколько позади овального. Оно закрыто волокнистой мембраной. Круглое окно отделяет барабанную полость от барабанной лестницы улитки.
Слуховые косточки - молоточек, наковальня, стремечко как система рычагов передают колебания барабанной перепонки наружного уха к овальному окну, от которого начинается вестибулярная лестница внутреннего уха.
Слуховая труба, соединяющая барабанную полость с носовой частью глотки, имеет хорошо выраженный просвет диаметром 1-2 мм. В области, прилежащей к барабанной полости, слуховая труба окружена костной стенкой, а ближе к глотке содержит островки гиалинового хряща. Просвет трубы выстлан многорядным призматическим реснитчатым эпителием. В нем имеются бокаловидные железистые клетки. На поверхности эпителия открываются протоки слизистых желез. Через слуховую трубу регулируется давление воздуха в барабанной полости среднего уха.
12.5.3. Внутреннее ухо
Внутреннее ухо (auris interna) состоит из костного лабиринта и расположенного в нем перепончатого лабиринта, в котором находятся рецептор-ные клетки - волосковые клетки органа слуха и равновесия. Рецепторные клетки (сенсоэпителиальные по происхождению) представлены в органе слуха - в спиральном органе улитки, а в органе равновесия - в пятнах маточки и мешочка (эллиптического и сферического мешочков) и в трех ампулярных гребешках полукружных каналов.
Развитие внутреннего уха. У 3-недельного эмбриона человека на уровне ромбовидного мозга (см. главу 11) обнаруживаются парные утолщения ней-роэктодермы - слуховые плакоды. Материал слуховых плакод впячивается в подлежащую мезенхиму, возникают слуховые ямки. Последние полностью погружаются во внутреннюю среду и отшнуровываются от эктодермы - образуются слуховые пузырьки. Развитие их контролируется мезенхимой, ромбовидным мозгом и мезодермой (рис. 12.12). Слуховой пузырек располагается вблизи первой жаберной щели.
Стенка слухового пузырька состоит из многорядного нейроэпите-лия, который секретирует эндолимфу, заполняющую просвет пузырька. Одновременно слуховой пузырек контактирует с эмбриональным слуховым нервным ганглием, который вскоре делится на две части - ганглий преддверия и ганглий улитки. В процессе дальнейшего развития пузырек меняет свою форму, перетягиваясь на две части: первая - вестибулярная - превращается в эллиптический пузырек - маточку (utriculus) с полукружными каналами и их ампулами, вторая - образует сферический пузырек - мешочек (sacculus) и закладку улиткового канала. Улитковый канал постепенно растет, завитки его увеличиваются, и он отделяется от эллиптического пузырька. На месте прилегания слухового ганглия к слуховому пузырьку стенка последнего утолщается. Клетки слухового пузырька с 7-й нед раз-
Рис. 12.12. Развитие слухового пузырька у эмбриона человека (по Арею, с изменениями):
а - стадия 9 сомитов; б - стадия 16 сомитов; в - стадия 30 сомитов. 1 - эктодерма; 2 - слуховая плакода; 3 - мезодерма; 4 - глотка; 5 - слуховая ямка; 6 - мозговой пузырь; 7 - слуховой пузырек
вития путем дивергентной дифференцировки дают начало клеточным диф-феронам улитки, полукружных каналов, маточки и мешочка. Дифферон рецепторных (сенсоэпителиальных) клеток возникает лишь при контакте малодифференцированных клеток с отростками нейронов слухового нервного ганглия.
Рецепторные и поддерживающие эпителиоциты органа слуха и равновесия обнаруживаются у эмбрионов длиной 15-18,5 мм. Улитковый канал вместе со спиральным органом развивается в виде трубки, которая впячивается в завитки костной улитки. Одновременно развиваются и пери-лимфатические пространства. В улитке у эмбриона длиной 43 мм имеется перилимфатическое пространство барабанной лестницы, а у эмбрионов длиной 50 мм - и перилимфатическое пространство вестибулярной лестницы. Несколько позднее происходят процессы окостенения и формирования костного лабиринта улитки и полукружных каналов.
Улитковый канал
Восприятие звуков осуществляется в спиральном органе, расположенном по всей длине улиткового канала перепончатого лабиринта. Улитковый канал представляет собой спиральный слепо заканчивающийся мешок длиной 3,5 см, заполненный эндолимфой и окруженный снаружи перилимфой. Улитковый канал и окружающие его заполненные перилимфой пространства барабанной и вестибулярной лестницы в свою очередь заключены в костную улитку, образующую у человека 2,5 завитка вокруг центрального костного стержня (модиолуса).
Улитковый канал на поперечном разрезе имеет форму треугольника, стороны которого образованы вестибулярной (преддверной) мембраной (мембрана Рейсснера), сосудистой полоской и базилярной пластинкой. Вестибулярная мембрана (membrana vestibularis) образует верхнемедиальную стенку канала. Она представляет собой тонкофибриллярную соединительнотканную пластинку, покрытую однослойным плоским эпителием, обращенным к эндолимфе, и слоем плоских фиброцитоподобных клеток, обращенным к перилимфе (рис. 12.13).
Наружная стенка образована сосудистой полоской (stria vascularis), расположенной на спиральной связке (ligamentum spirale). В составе сосудистой полоски различают многочисленные краевые клетки с большим количеством митохондрий в цитоплазме. Апикальная поверхность этих клеток
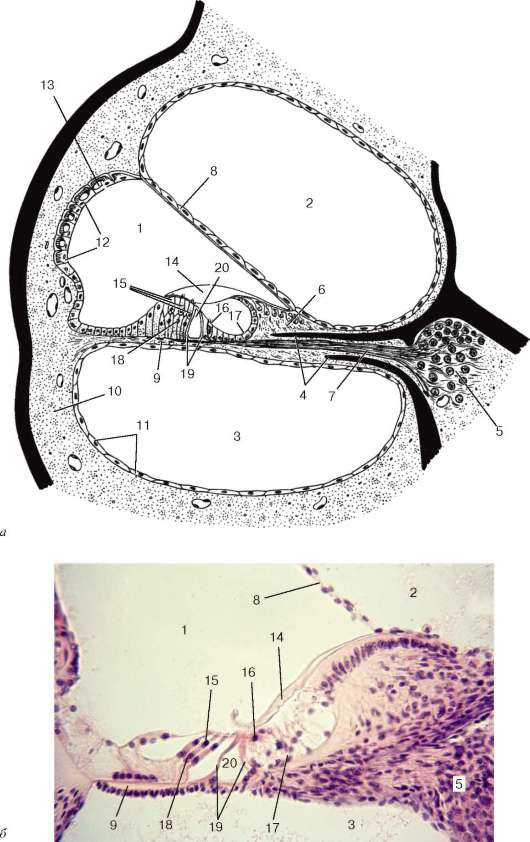
Рис. 12.13. Строение перепончатого канала улитки и спирального органа: а - схема; б - спиральный орган (микрофотография). 1 - перепончатый канал улитки; 2 - вестибулярная лестница; 3 - барабанная лестница; 4 - спиральная костная пластинка; 5 - спиральный узел; 6 - спиральный гребень; 7 - дендриты нервных клеток; 8 - вестибулярная мембрана; 9 - базилярная пластинка; 10 - спиральная связка; 11 - эпителий, выстилающий барабанную лестницу; 12 - сосудистая полоска; 13 - кровеносные сосуды; 14 - покровная мембрана; 15 - наружные волосковые (сен-соэпителиальные) клетки; 16 - внутренние волосковые (сенсоэпителиальные) клетки; 17 - внутренние поддерживающие эпителиоциты; 18 - наружные поддерживающие эпителиоциты; 19 - наружные и внутренние столбчатые эпителиоциты; 20 - туннель

Рис. 12.14. Ультрамикроскопическое строение сосудистой полоски (а) (по Ю. И. Афанасьеву):
б - микрофотография сосудистой полоски. 1 - светлые базальные клетки; 2 - темные призматические клетки; 3 - митохондрии; 4 - кровеносные капилляры; 5 - базальная мембрана
омывается эндолимфой. Клетки осуществляют транспорт ионов натрия и калия, обеспечивают высокую концентрацию ионов калия в эндолим-фе. Промежуточные (звездчатой формы) и базальные (плоские) клетки не имеют контакта с эндолимфой. Базальные клетки относят к камбию сосудистой полоски. Здесь же обнаруживаются нейроэндокриноциты, вырабатывающие пептидные гормоны - серотонин, мелатонин, адреналин и другие, которые участвуют в регуляции объема эндолимфы. Между клетками проходят гемокапилляры. Предполагают, что клетки сосудистой полоски продуцируют эндолимфу, которая играет значительную роль в трофике спирального органа (рис. 12.14).
Нижняя (базилярная) пластинка (lamina basilaris), на которой располагается спиральный орган, построена наиболее сложно. С внутренней стороны она прикрепляется к спиральной костной пластинке в том месте, где ее надкостница - спиральный край (лимб) делится на две части: верхнюю - вестибулярную губу и нижнюю - барабанную губу. Последняя переходит в базилярную пластинку, которая на противоположной стороне прикрепляется к спиральной связке.
Базилярная пластинка представляет собой соединительнотканную пластинку, которая в виде спирали тянется вдоль всего улиткового канала. На стороне, обращенной к спиральному органу, она покрыта базальной мембраной эпителия этого органа. В основе базилярной пластинки лежат тонкие коллагеновые волокна, которые тянутся в виде непрерывного радиального пучка от спиральной костной пластинки до спиральной связки, выступающих в полость костного канала улитки. Характерно, что длина волокон неодинакова по всей длине улиткового канала. Более длинные (около 505 мкм) волокна находятся на вершине улитки, короткие (около 105 мкм) - в ее основании. Располагаются волокна в гомогенном основном веществе. Волокна состоят из тонких фибрилл диаметром около 30 нм, ана-стомозирующих между собой с помощью еще более тонких пучков. Со стороны барабанной лестницы базилярная пластинка покрыта слоем плоских фиброцитоподобных клеток мезенхимной природы.
Поверхность спирального края покрыта плоским эпителием. Его клетки обладают способностью к секреции. Выстилка спиральной бороздки (sulcus spiralis) представлена несколькими рядами крупных плоских полигональных клеток, которые непосредственно переходят в поддерживающие эпителиоциты, примыкающие к внутренним волосковым клеткам спирального органа.
Покровная мембрана (membrana tectoria) имеет связь с эпителием вестибулярной губы. Она представляет собой лентовидную пластинку желеобразной консистенции, которая тянется в виде спирали по всей длине спирального органа, располагаясь над вершинами его сенсоэпителиальных волосковых клеток. Эта пластинка состоит из тонких радиально направленных коллагеновых волокон. Между волокнами находится прозрачное склеивающее вещество, содержащее гликозаминогликаны.
Спиральный орган
Спиральный, или кортиев, орган расположен на базилярной мембране перепончатого лабиринта улитки. Это эпителиальное образование повторяет ход улитки. Его площадь расширяется от базального завитка улитки к апикальному. Состоит из двух групп клеток - волосковых (сенсоэпите-лиальных, кохлеоцитов) и поддерживающих. Каждая из этих групп клеток подразделяется на внутренние и наружные (см. рис. 12.13). Эти две группы разделяет туннель.
Внутренние волосковые клетки (cochleocyti internae) имеют кувшинообраз-ную форму (рис. 12.15) с расширенной базальной и искривленной апикальной частями, лежат в один ряд на поддерживающих внутренних фаланговых эпителиоцитах (epitheliocyti phalangeae internae). Их общее количество у человека достигает 3500. На апикальной поверхности имеется ретикулярная пластинка, на которой расположены от 30 до 60 коротких микроворсинок - стереоцилии (длина их в базальном завитке улитки примерно 2 мкм, а в верхушечном - больше в 2-2,5 раза). В базальной и апикальной частях клеток имеются скопления митохондрий, элементы гладкой и гранулярной эндо-плазматической сети, актиновые и миозиновые миофиламенты. Наруж-
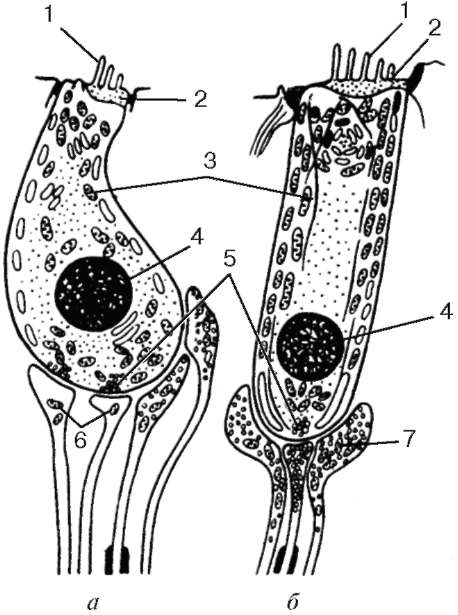
Рис. 12.15. Ультраструктурная организация внутренних (а) и наружных (б) во-лосковых клеток (схема). 1 - волоски; 2 - кутикула; 3 - митохондрии; 4 - ядра; 5 - синаптические пузырьки в цитоплазме сенсоэпители-альных клеток; 6 - светлые нервные окончания; 7 - темные нервные окончания
ная поверхность базальной половины клетки покрыта сетью преимущественно афферентных нервных окончаний.
Наружные волосковые клетки (coch-leocyti externae) имеют цилиндрическую форму, лежат в 3-5 рядов во вдавлениях поддерживающих наружных фаланговых эпителиоцитов (epitheliocyti phalangeae externae). Общее количество наружных эпителиальных клеток у человека может достигать 12 000-20 000. Они, как и внутренние волосковые клетки, имеют на своей апикальной поверхности кутикулярную пластинку со стерео-цилиями, которые образуют щеточку из нескольких рядов в виде буквы V (рис. 12.16). Стереоцилии числом 100- 300 своими вершинами прикасаются к внутренней поверхности покровной мембраны. Они содержат многочисленные плотно расположенные фибриллы, имеющие в своем составе сократительные белки (актин и миозин), благодаря чему после наклона они вновь принимают исходное вер-
тикальное положение.
Цитоплазма клеток содержит агранулярную эндоплазматическую сеть, элементы цитоскелета, богата окислительными ферментами, имеет большой запас гликогена. Все это позволяет клетке сокращаться. Клетки иннер-вируются преимущественно эфферентными волокнами.
Наружные волосковые клетки значительно чувствительнее к звукам большей интенсивности, чем внутренние. Высокие звуки раздражают только волосковые клетки, расположенные в нижних завитках улитки, а низкие звуки - волосковые клетки вершины улитки.
Во время звукового воздействия на барабанную перепонку ее колебания передаются на молоточек, наковальню и стремечко, а далее через овальное окно на перилимфу, базилярную пластинку и покровную мембрану. В ответ на звук возникают колебания, которые воспринимаются волоско-выми клетками, так как происходит радиальное смещение покровной мембраны, в которую погружены кончики стереоцилий. Отклонение стерео-цилий волосковых клеток изменяет проницаемость механочувствительных ионных каналов и возникает деполяризация плазмолеммы. Нейромедиатор (глутамат) выделяется из синаптических пузырьков и воздействует на рецепторы афферентных терминалей нейронов слухового ганглия. Афферентная
информация по слуховому нерву передается в центральные части слухового анализатора.
Поддерживающие эпителиоциты спирального органа в отличие от волосковых своими основаниями непосредственно располагаются на базальной мембране. В их цитоплазме обнаруживаются тонофибриллы. Внутренние фаланговые эпителиоциты, лежащие под внутренними волосковыми клетками, связаны между собой плотными и щелевыми контактами. На апикальной поверхности имеются тонкие пальцевидные отростки (фаланги). Этими отростками вершины волосковых клеток отделены друг от друга.
На базилярной мембране располагаются также наружные фалан-говые клетки. Они залегают в 3- 4 ряда в непосредственной близости от наружных столбчатых эпителио-цитов. Эти клетки имеют призматическую форму. В их базальной части располагается ядро, окруженное пучками тонофибрилл. В верхней трети, на месте соприкосновения с наружными волосковыми клетками, в наружных фаланговых эпителио-цитах есть чашевидное вдавление, в которое входит основание наружных волосковых клеток. Только один узкий отросток наружных поддерживающих эпителиоцитов доходит своей тонкой вершиной - фалангой - до верхней поверхности спирального органа.
В спиральном органе расположены также так называемые внутренние и наружные столбчатые эпителиоциты (epitheliocyti columnaris internae et externae). На месте своего соприкосновения они сходятся под острым углом друг к другу и образуют правильный треугольный канал - туннель, заполненный эндолимфой. Туннель тянется по спирали вдоль всего спирального органа. Основания столбчатых эпителиоцитов прилежат друг к другу и располагаются на базальной мембране. Через туннель проходят нервные волокна.
Вестибулярная часть перепончатого лабиринта (labyrinthus vestibularis) - место расположения рецепторов органа равновесия. Она состоит из двух пузырьков - эллиптического, или маточки (utriculus), и сферического, или круглого, мешочка (sacculus), сообщающихся при помощи узкого канала и связанных с тремя полукружными каналами, локализующимися в костных

Рис. 12.16. Наружная поверхность клеток спирального органа. Сканирующая электронная микрофотография, увеличение 2500 (препарат К. Койчева): 1 - наружные волосковые клетки; 2 - внутренние волосковые клетки; 3 - границы поддерживающих эпителиоцитов
каналах, расположенных в трех взаимно перпендикулярных направлениях. Эти каналы на месте соединения их с маточкой имеют расширения - ампулы. В стенке перепончатого лабиринта в области маточки и мешочка и ампул есть участки, содержащие чувствительные клетки - вестибулоциты. Эти участки называются пятнами, или макулами, соответственно: пятно маточки (macula utriculi) располагается в горизонтальной плоскости, а пятно круглого мешочка (macula sacculi) - в вертикальной плоскости. В ампулах эти участки называются гребешками, или кристами (crista ampullaris). Стенка вестибулярной части перепончатого лабиринта состоит из однослойного плоского эпителия, за исключением области крист полукружных каналов и макул, где он превращается в кубический и призматический.
Пятна мешочков (макулы). Эти пятна выстланы эпителием, расположенным на базальной мембране и состоящим из чувствительных и опорных клеток (рис. 12.17). Поверхность эпителия покрыта особой студенистой ото-литовой мембраной (membrana statoconiorum), в которую включены состоящие из карбоната кальция кристаллы - отолиты, или статоконии (statoconia). Макула маточки - место восприятия линейных ускорений и земного притяжения (рецептор гравитации, связанный с изменением тонуса мышц, определяющих установку тела). Макула мешочка, являясь также рецептором гравитации, одновременно воспринимает и вибрационные колебания.
Вестибулярные волосковые клетки (cellulae sensoriae pilosae) непосредственно обращены своими вершинами, усеянными волосками, в полость лабиринта. По строению волосковые клетки подразделяются на два вида (см. рис. 12.17, б). Грушевидные вестибулоциты отличаются округлым широким основанием, к которому примыкает нервное окончание, образующее вокруг него футляр в виде чаши. Столбчатые вестибулоциты формируют точечные контакты с афферентными и эфферентными нервными волокнами. На наружной поверхности этих клеток имеется кутикула, от которой отходят 60-80 неподвижных волосков - стереоцилии длиной около 40 мкм и одна подвижная ресничка - киноцилия, имеющая строение сократительной реснички.
Пятно мешочка содержит около 18 000 рецепторных клеток, а пятно маточки - около 33 000. Киноцилия всегда полярно располагается по отношению к пучку стереоцилий. При смещении стереоцилий в сторону кино-цилии клетка возбуждается, а если движение направлено в противоположную сторону, происходит торможение клетки. В эпителии макул различно поляризованные клетки собираются в четыре группы, благодаря чему во время скольжения отолитовой мембраны стимулируется только определен-
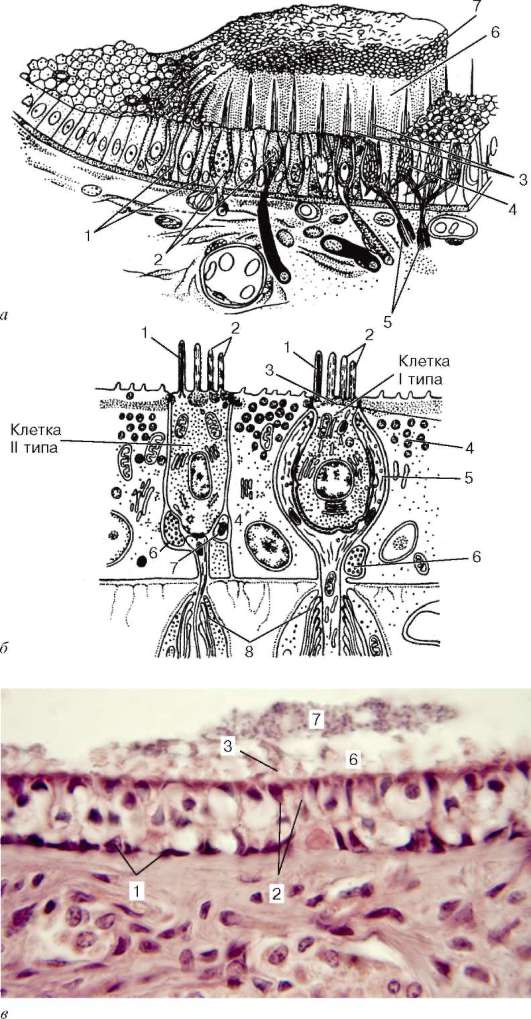
Рис. 12.17. Макула:
а - строение на светооптическом уровне (схема по Кольмеру):
1 - поддерживающие эпителиоциты; 2 - волосковые (сенсоэпителиальные) клетки; 3 - волоски; 4 - нервные окончания; 5 - миелиновые нервные волокна; 6 - студенистая отолитовая мембрана; 7 - отолиты; б - строение на ультрамикроскопическом уровне (схема): 1 - киноцилия; 2 - стереоцилия; 3 - кутикула; 4 - поддерживающий эпителиоцит; 5 - чашевидное нервное окончание; 6 - эфферентное нервное окончание; 7 - афферентное нервное окончание; 8 - миелиновое нервное волокно (дендрит); в - микрофотография (обозначения см. «а»)
ная группа клеток, регулирующая тонус определенных мышц туловища; другая группа клеток в это время тормозится. Полученный через афферентные синапсы импульс передается через вестибулярный нерв в соответствующие части вестибулярного анализатора.
Поддерживающие эпителиоциты (epitheliocyti sustentans), располагаясь между волосковыми, отличаются темными овальными ядрами. Они имеют большое количество митохондрий. На их вершинах обнаруживается множество микроворсинок.
Ампулярные гребешки (кристы). Они в виде поперечных складок находятся в каждом ампулярном расширении полукружного канала. Ампулярный гребешок выстлан вестибулярными волосковыми и поддерживающими эпителиоцитами. Апикальная часть этих клеток окружена желатинообраз-ным прозрачным куполом (cupula gelatinosa), который имеет форму колокола, лишенного полости. Его длина достигает 1 мм. Тонкое строение волоско-вых клеток и их иннервация сходны с таковыми волосковых клеток макул маточки и мешочка (рис. 12.18). В функциональном отношении жела-тинозный купол - рецептор угловых ускорений. При движении головы или ускоренном вращении всего тела купол легко меняет свое положение. Отклонение купола под влиянием движения эндолимфы в полукружных каналах стимулирует волосковые клетки. Их возбуждение вызывает рефлекторный ответ той части скелетной мускулатуры, которая корригирует положение тела и движение глазных мышц.
Иннервация. На волосковых эпителиоцитах спирального и вестибулярного органов расположены афферентные нервные окончания биполярных нейронов, тела которых располагаются в основании спиральной костной пластинки, образуя спиральный ганглий. Основная часть нейронов (первый тип) относится к большим биполярным клеткам, которые содержат крупное ядро с ядрышком и мелкодиспергированным хроматином. В цитоплазме имеются многочисленные рибосомы, редко встречающиеся нейрофила-менты. Ко второму типу нейронов относятся мелкие псевдоуниполярные нейроны, отличающиеся ацентричным расположением ядра с плотным хроматином, малым количеством рибосом и большой концентрацией нейрофи-ламентов в цитоплазме, слабой миелинизацией нервных волокон.
Нейроны первого типа получают афферентную информацию исключительно от внутренних волосковых клеток, а нейроны второго типа - от наружных волосковых клеток. Иннервация внутренних и наружных воло-сковых клеток кортиева органа осуществляется двумя типами волокон. Внутренние волосковые клетки снабжены преимущественно афферентными волокнами, которые составляют около 95 % всех волокон слухового нерва, а наружные волосковые клетки получают преимущественно эфферентную иннервацию (составляет 80 % всех эфферентных волокон улитки).
Эфферентные волокна происходят из перекрещенного и неперекрещен-ного оливо-кохлеарных пучков. Число волокон, пересекающих туннель, может быть около 8000.
На базальной поверхности одной внутренней волосковой клетки бывает до 20 синапсов, образуемых афферентными волокнами слухового нерва.
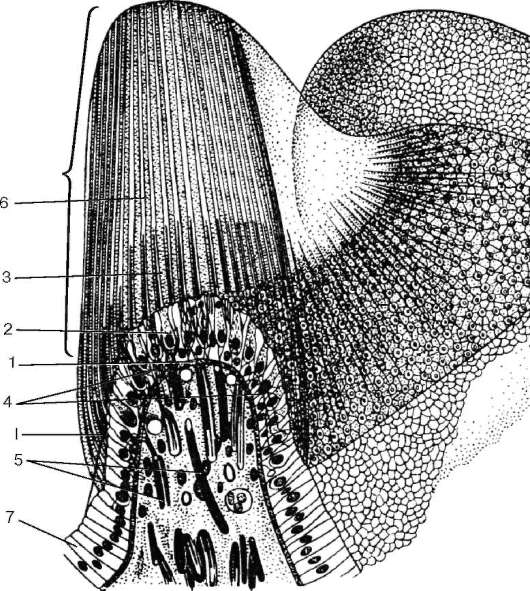
Рис. 12.18. Строение ампулярного гребешка (схема по Кольмеру, с изменениями): I - гребешок; II - желатинозный купол. 1 - поддерживающие эпителиоциты; 2 - во-лосковые (сенсоэпителиальные) клетки; 3 - волоски; 4 - нервные окончания; 5 - миелиновые нервные волокна; 6 - желатинозное вещество пограничного купола; 7 - эпителий, выстилающий стенку перепончатого канала
Эфферентные терминали составляют не более одной на каждой внутренней волосковой клетке, в них находятся круглые прозрачные пузырьки диаметром до 35 нм. Под внутренними волосковыми клетками видны многочисленные аксодендритические синапсы, образованные эфферентными волокнами на афферентных волокнах, которые содержат не только светлые, но и более крупные гранулированные пузырьки диаметром 100 нм и более
(рис. 12.19).
На базальной поверхности наружных волосковых клеток афферентные синапсы немногочисленны (разветвления одного волокна иннервируют до 10 клеток). В этих синапсах видны немногочисленные круглые светлые пузырьки диаметром 35 нм и более мелкие (6-13 нм). Эфферентные синапсы более многочисленны - до 13 на 1 клетку. В эфферентных терминалях находятся круглые светлые пузырьки диаметром около 35 нм и гранулированные - диаметром 100-300 нм. Кроме того, на боковых поверхностях
Рис. 12.19. Иннервация и медиаторное обеспечение спирального органа (схема): 1 - внутренняя волосковая (сенсоэпителиальная) клетка; 2 - наружные волосковые (сенсоэпителиальные) клетки; 3 - рецепторы на волосковых клетках; 4 - эфферентное окончание на дендрите рецепторного нейрона; 5 - эфферентные окончания на наружных волосковых клетках; 6 - биполярные нейроны спирального узла; 7 - покровная мембрана
наружных сенсоэпителиальных клеток имеются терминали в виде тонких веточек с синаптическими пузырьками диаметром до 35 нм. Под наружными волосковыми клетками имеются контакты эфферентных волокон на афферентных волокнах.
Медиаторы синапсов. Тормозящие медиаторы. Ацетилхолин - основной медиатор в эфферентных терминалях на наружных и внутренних волосковых клетках. Его роль заключается в подавлении ответов волокон слухового нерва на акустическую стимуляцию. Опиоиды (энкефалины) обнаружены в эфферентных терминалях под внутренними и наружными волосковыми клетками в виде крупных (больше 100 нм) гранулированных пузырьков. Их роль - модуляция активности других медиаторов: ацетилхолина, норадреналина, гамма-аминомасляной кислоты (ГАМК) - путем непосредственного взаимодействия с рецепторами или изменения проницаемости мембраны для ионов и медиаторов.
Возбуждающие медиаторы (аминокислоты). Глутамат обнаружен в области основания внутренних волосковых клеток и в больших нейронах спирального ганглия. Аспартат найден вокруг наружных волосковых клеток в афферентных терминалях, содержащих ГАМК, и в малых нейронах спирального ганглия. Их роль - регуляция деятельности каналов К+ и Na+.
Нейроны коркового центра слуховой сенсорной системы расположены в верхней височной извилине, где происходит интеграция качеств звука (интенсивность, тембр, ритм, тон) на клетках 3-й и 4-й пластинок коры. Корковый центр слуховой сенсорной системы имеет многочисленные ассоциативные связи с корковыми центрами других сенсорных систем, а также с моторной зоной коры.
Васкуляризация. Артерия перепончатого лабиринта берет свое начало от верхней мозговой артерии. Она делится на две ветви: вестибулярную и общую кохлеарную. Вестибулярная артерия снабжает кровью нижние и боковые части маточки и мешочка, а также верхние боковые части полукружных каналов, образуя капиллярные сплетения в области слуховых пятен. Кохлеарная артерия снабжает кровью спиральный ганглий и через надкостницу вестибулярной лестницы и спиральной костной пластинки доходит до внутренних частей базальной мембраны спирального органа. Венозная система лабиринта складывается из трех независимых друг от друга венозных сплетений, находящихся в улитке, преддверии и полукружных каналах. Лимфатические сосуды в лабиринте не обнаружены. Спиральный орган сосудов не имеет.
Возрастные изменения. С возрастом у человека могут возникать нарушения органа слуха. При этом изменяются отдельно или совместно звукопроводящая и звуковоспринимающая системы. Это связано с тем, что в области овального окна костного лабиринта появляются очаги оссификации, распространяющиеся на подкожную пластинку стремечка. Стремечко теряет подвижность в овальном окне, что резко снижает порог слышимости. С возрастом чаще поражаются нейроны сенсорного аппарата, которые гибнут и не восстанавливаются.
Контрольные вопросы
1. Принципы классификации органов чувств.
2. Развитие, строение органа зрения, основы физиологии зрения.
3. Орган слуха и равновесия: развитие, строение, функции.
4. Органы вкуса и обоняния. Особенности развития и строения их рецеп-торных клеток.