Клиническая генетика. Геномика и протеомика наследственной патологии : учеб. пособие. - 3-е изд., перераб. и доп. - Мутовин Г.Р. 2010. - 832 с. : ил
|
|
|
|
ГЛАВА 2 СТРУКТУРНАЯ ГЕНОМИКА
ОРГАНИЗАЦИЯ НАСЛЕДСТВЕННОГО МАТЕРИАЛА
Наследственность (или наследственный материал) - это генетическая информация, заключенная в молекулах ДНК и РНК. Основная часть ДНК (3,1467 млрд н.п.) находится в ядре, минимальная часть ДНК (16,6 тыс. н.п.) - в цитоплазме (митохондриях) клетки.
Выделяют следующие уровни организации наследственного материала:
• молекулярно-генетический (генный или геномный) уровень - нуклеотидные последовательности молекул ДНК и РНК одной клетки;
• цитогенетический (хромосомный или гетерохроматиновый) уровень - нуклеотидные и аминокислотные последовательности хромосом одной клетки;
• популяционный генетический и фенотипический уровень - генотипы (геномы) и фенотипы (феномы) всех организмов одного и того же биологического вида или геномно-протеомный организменный уровень.
В зависимости от перечисленных уровней в первой половине XX в. в генетике выделились научные направления: молекулярная генетика, цитогенетика и популяционная генетика. В дальнейшем первые два направления трансформировались в структурную геномику, продолжением (следствием) которой стала функциональная геномика (протеомика).
Рассмотрение уровней организации наследственного материала у человека целесообразно начать с наиболее знакомого популяционного генетического и фенотипического уровня.
Как сказано в главе 1, термин «популяция» впервые применен в 1903 г. В. Иогансеном. В современной редакции под ним подразумевается совокупность особей (организмов) одного биологического вида, населяющих определенную географическую территорию.
Биологический вид Homo sapiens (Человек разумный) на Земле включает почти 6,5 млрд людей, принадлежащих к различным расам,
нациям, народам и народностям, составляющим большие и малые популяции или популяционные группы.
К наиболее известным из них относятся представители 400 основных популяционных групп (около 8% общей популяции человека).
Существует еще огромное число этнических групп (этносов), различающихся по языковым диалектам и наречиям. Жители Земли общаются между собой на 2100 различных языках, объединяющих около 5 тыс. диалектов и наречий.
В России традиционно проживали две основные расы (европейская и азиатская), представленные многочисленными национальностями, народами и народностями. В конце XX в. у нас насчитывалось 164 популяционные группы. В XXI в. их стало еще больше из-за огромного миграционного притока населения в Россию из бывших союзных республик СССР.
Во всех без исключения популяционных группах всегда можно отличить разных людей друг от друга по генотипическим и фенотипическим признакам как в норме, так и при патологии.
Под генотипическими различиями подразумеваются структурные (физические, химические, генетические, биохимические) и функциональные особенности наследственного материала на геномном (молекулы ДНК и РНК) и хромосомном (геномно-протеомном) уровнях, под фенотипическими различиями - внешние и/или внутренние особенности на уровне фенотипа отдельной клетки и целого организма (соответствующие понятия будут рассмотрены ниже).
В случае патологии практически в каждой популяции выделяют характерные для нее болезни (например, еврейские болезни - Вильсона- Коновалова, Леша-Найяна, Тея-Сакса, меледская болезнь или ладонноподошвенный гиперкератоз у населения, проживающего вблизи Средиземного моря, финские болезни - аспарагилглюкозоаминурия, шведский и японский варианты наследственного нефрита и т.д.).
Теперь рассмотрим молекулярно-генетический и хромосомный уровни организации наследственного материала.
СТРОЕНИЕ МОЛЕКУЛЫ ДНК
Химическое строение молекулы ДНК в виде двойной спирали предугадано Джеймсом Уотсоном, Френсисом Криком и Моррисом Уилкинсом и впервые продемонстрировано на рентгенограмме Розалиндой Франклин в 1953 г. (рис. 1).
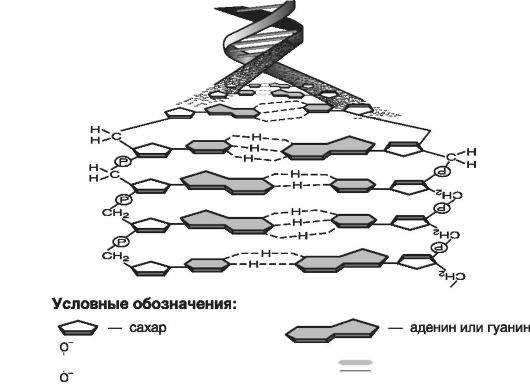
Рис. 1. Схема структуры молекулы ДНК
Как видно из приведенной на рис. 1 схемы, молекула ДНК представляет собой линейный полимер, сформированный из двух полинуклеотидных цепей (нитей) или ДНК-полинуклеотидов, закрученных между собой в правую спираль.
Каждая линейная цепь состоит из нуклеотидов или мономеров, которые можно представить как фосфат (P) - сахар - основание, т.е. данная последовательность нуклеотидов - первичная структура
ДНК.
В ДНК четыре базовых азотистых основания: аденин - А, гуанин - Г, тимин - Т и цитозин - Ц. Из них первые два - пурины, последние два - пиримидины.
Связь нуклеотидов внутри каждой цепи ДНК обеспечивается чередованием фосфатных групп и пятиуглеродного сахара - дезоксирибозы. Между ними существует прочная фосфодиэфирная связь.
Обе линейные цепи закручены в правую спираль таким образом, что внутри спирали расположены основания, а снаружи - сахарофосфатные комплексы. Спираль - вторичная структура ДНК.
Каждая линейная цепь имеет два конца: начало- 5'-конец и З'-конец.
Связь двух цепей в молекуле ДНК обеспечивается комплементарностью пар оснований. Это означает, что, когда основания А и Т противоположных цепей молекулы расположены друг против друга, между ними спонтанно образуются две слабые водородные связи, тем самым создается АТ-пара (аденин и тимин). В свою очередь, ГЦ-пара (гуанин и цитозин) также представляет собой комплементарную пару, но между гуанином и цитозином возникают три слабые водородные связи.
Спонтанный процесс образования пар оснований (т.е. нуклеотидных пар - н.п.) называется гибридизацией ДНК. Другие комбинации пар оснований не комплементарны.
Цепи ДНК между собой антипараллельны (имеют противоположную направленность), т.е. определяющее направление в одной цепи от (5'-конца до З'-конца) будет противоположно таковому в другой цепи (от З'-конца до 5'-конца).
Простую полинуклеотидную структуру принято обозначать как последовательность первых букв названий азотистых оснований. Фрагмент двухцепочечной молекулы ДНК с противоположной направленностью цепей изображается так:
5' Ц А Г T 3' 3' Г T Ц A 5'
Молекула ДНК - своего рода «винтовая лестница», условные «перила» которой (слева и справа) - сахарофосфатные комплексы, а условные «ступени» - АТ- и ГЦ-пары оснований. Каждая «ступень» состоит как бы из двух половинок: слева - пурин, справа - пиримидин.
Процесс разделения цепей ДНК (или их фрагментов) под действием температуры (около 95 °С) приводит к разрыву водородных связей между комплементарными парами оснований и называется плавлением ДНК.
При последующем охлаждении водородные связи между комплементарными парами восстанавливаются. Такая техника гибридизации называется отжигом ДНК. Она послужила основой для изучения молекулярной структуры гена.
СТРОЕНИЕ И ТИПЫ МОЛЕКУЛЫ РНК
Молекула РНК образуется на молекуле ДНК в ходе транскрипции (считывание наследственной информации). По структуре, хотя и напоминающей структуру молекулы ДНК, молекула РНК принципи-
ально иная: она представляет собой не две, а одну цепочку, в которой вместо дезоксирибозы содержится рибоза, а вместо тимина - урацил (пиримидиновое основание).
В эукариотических клетках существуют разные типы молекул
РНК.
• Рибосомные РНК (рРНК). Существуют в виде 5 субъединиц, образующих рибосомы. Они занимают 80% общего объема разных типов РНК.
• Транспортные РНК (тРНК). Их 20 типов, соответствующих 20 основным аминокислотам. Они участвуют в параллельно протекающих процессах трансляции и биосинтеза и занимают 15% объема.
• Малые ядерные РНК (мяРНК). Это небольшие гетерогенные РНК, участвующие в сплайсинге (см. ниже). Их объем менее 1%.
• Информационные РНК (иРНК), или мессенджер РНК (мРНК). «Мессенджер» означает «посланец» или «гонец» (из ядра в цитоплазму).
Иногда используется еще одно название: РНК - транскрипт. Их 2 типа: пре-РНК (копия первичной структуры гена) и мРНК (собственно ген) - это гетерогенные РНК, образующиеся в ходе транскрипции ДНК и занимающие около 5% объема.
МОЛЕКУЛА ДНК - ГЛАВНАЯ МОЛЕКУЛА ЖИЗНИ
Молекулу ДНК характеризуют следующие уникальные свойства.
• Бессмертие и стабильность функционирования. Только молекула ДНК передается из поколения в поколение в стабильном виде. Она обеспечивает непрерывность жизни биологического вида (но не индивида) благодаря сохранению и воспроизводству закодированной в ее структуре наследственной информации о конкретном биологическом виде, обновляющейся в каждом поколении.
• Обеспечение эволюционной преемственности и уникальности наследственной информации в ряду поколений с помощью механизмов взаимодействия между материнским и отцовским (по происхождению) геномами.
• Механизм экспрессии (работы) генов как источник белковых продуктов онтогенеза.
• Наличие восстановительных механизмов (систем репарации) на случай повреждения собственной структуры молекулы.
ГЕНЕТИЧЕСКИЙ КОД
Генетический код - условная запись наследственной информации в молекулах ДНК и РНК. В состав белковых молекул (полипептидных цепочек) входят 20 основных аминокислот. Включение в полипептидную цепь одной аминокислоты определяется последовательностью из трех азотистых оснований - триплетом (ДНКили РНК-кодон), кодирующей единицей. Последовательность оснований в одном кодоне имеет принципиальное значение для синтеза аминокислот. Так, если три основания расположены в определенном порядке (тимин, цитозин, аденин), то производится серин, если в другом (аденин, цитозин, тимин), то треонин.
Соответствие ДНКили РНК-кодона той или иной аминокислоте составляет сущность канонического (классического) генетического кода (табл. 1).
Таблица 1. Соответствие ДНКили РНК-кодонов основным аминокислотам
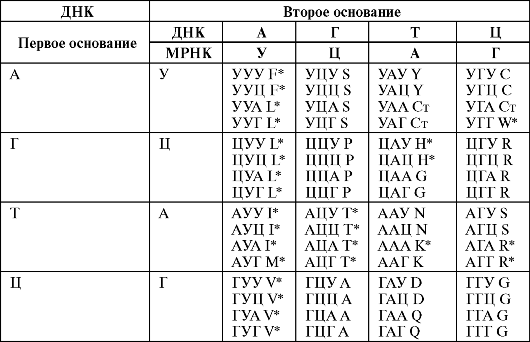 Обозначения: Азотистые
основания: А - аденин, Г - гуанин, Ц - цитозин, Т - тимин, У - урацил;
основные аминокислоты: А - аланин, R - аргинин, N - аспарагин, D -
аспарагиновая кислота, V - валин, H - гистидин, G - глицин, Q -
глутамин, E - глутаминовая кислота, I - изолейцин, L - лейцин, K -
лизин, М - метионин, P - пролин, S - серин, Y - тирозин, Т - треонин, W -
триптофан, F - фенилаланин, C - цистеин, Ст - стоп-кодон; * -
незаменимые аминокислоты, АУГ М - инициаторный кодон.
Обозначения: Азотистые
основания: А - аденин, Г - гуанин, Ц - цитозин, Т - тимин, У - урацил;
основные аминокислоты: А - аланин, R - аргинин, N - аспарагин, D -
аспарагиновая кислота, V - валин, H - гистидин, G - глицин, Q -
глутамин, E - глутаминовая кислота, I - изолейцин, L - лейцин, K -
лизин, М - метионин, P - пролин, S - серин, Y - тирозин, Т - треонин, W -
триптофан, F - фенилаланин, C - цистеин, Ст - стоп-кодон; * -
незаменимые аминокислоты, АУГ М - инициаторный кодон.
Как следует из табл. 1, из четырех базовых оснований, входящих в молекулу ДНК или молекулу РНК, можно образовать 64 триплета (ДНК- и РНК-кодоны). Из них три не кодируют основные аминокислоты: УАА, УАГ и УГА - это «стоп-кодоны», прекращающие биосинтез белков в рибосомах. Еще один кодон - ЛУГ - выполняет две функции: кодирует аминокислоту метионин и является инициаторным (индикаторным) кодоном; при контакте с ним РНК-полимераза начинает биосинтез белка в рибосомах.
Оставшиеся триплеты (всего 61) кодируют основные аминокислоты. Причем одну аминокислоту может кодировать как один, так и сразу несколько триплетов - это вырожденный генетический код . Например, только двум аминокислотам соответствуют по одному кодону (метионин - АУГ и триптофан - УГГ); остальным аминокислотам соответствуют 2 кодона и более. Так, у лейцина их 6: УУА, УУГ, ЦУУ, ЦУЦ, ЦУА и ЦУГ.
Свойства генетического кода
Свойства генетического кода:
• триплетность;
• линейность (кодоны в мРНК считываются в направлении 5' -> 3');
• неперекрываемость (каждый нуклеотид в мРНК входит в состав только одного кодона);
• вырожденность (одна аминокислота может кодироваться однимшестью кодонами, различающимися, как правило, по третьему нуклеотиду в триплете);
• универсальность (генетический код универсален для всех видов живых организмов).
Условия функционирования и фундаментальные генетические процессы в клетке
В определенных физиологических условиях (рН среды 7,35; t среды 37 °С) ДНК представляет собой «живой» кодирующий материал, функционирующий только в присутствии специфических ферментов.
Если необходимые параметры отсутствуют, то клеточная ДНК - «мертвый» кодирующий материал, который, однако, можно «оживить», поместив в указанные физиологические условия.
Помимо уже отмеченных выше ДНК- и РНК-полимераз, следует выделить играющие особую роль другие филогенетически устойчи-
вые ферменты клетки - ДНК-хеликазы, способные к разделению двух цепей молекулы ДНК и переводящие ее из нативного состояния в одноцепочечное. Эти ферменты участвуют только в тех генетических процессах, которые непосредственно связаны с разделением родительских цепей молекулы ДНК (репликация, транскрипция, рекомбинация и репарация) или на них основаны. Они называются фундаментальными генетическими процессами клетки. К ним относится также биосинтез белка.
Отдельно рассмотрим каждый из этих процессов.
Репликация ДНК
Репликация ДНК обеспечивает воспроизводство самой молекулы (удвоение) и, следовательно, передачу закодированной в ней информации из поколения в поколение, в том числе измененной информации, если таковая появилась.
В упрощенном виде поясним: в ходе репликации под действием ДНК-полимеразы в молекуле ДНК разрываются слабые водородные связи между АТ- и ГЦ-парами оснований, двойная спираль расходится, и на каждой из разошедшихся цепей (как на матрице) строится комплементарная ей дочерняя цепь. Между матричной и дочерней цепями восстанавливаются водородные связи и формируются АТ- и ГЦ-пары оснований.
Таким образом, в клетке осуществляется комплементарный синтез ДНК, получивший название полуконсервативного способа репликации; он характерен для всех эукариот.
Репликация ДНК начинается в точке инициации, где происходит разъединение двойной спирали и образование одноцепочечных участков ДНК, служащих матрицей для синтеза дочерних цепей. Этот процесс называется вилкой репликации (рис. 2).
Как показано на рис. 2, новая цепь ДНК растет в направлении от 5'-конца к З'-концу матричной цепи, имеющей противоположное направление (от 3'-конца к 5'-концу).
Первый дезоксирибонуклеотид присоединяется к 3'-концу РНК-праймера (короткий двухцепочечный участок со свободным 3 '-концом), выстраивая дочернюю цепочку ДНК.
Поскольку все полимеразы строят цепи только в одном направлении, то синтез ДНК на 3' - 5' родительской цепи будет идти непрерывно - это лидирующая цепь, растущая в направлении движения вилки репликации, и для ее продолжения необходим один акт инициации.
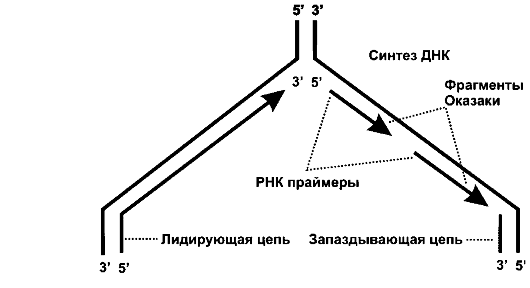 Рис. 2. Схема вилки репликации (по: Гриффите А. и соавт., 2000)
Рис. 2. Схема вилки репликации (по: Гриффите А. и соавт., 2000)
На другой матричной цепи ДНК синтез дочерней цепи осуществляется короткими фрагментами Оказаки - это запаздывающая (отстающая) цепь; она растет в направлении, противоположном движению репликативной вилки.
Синтез ДНК прекращается, когда встречаются вилки соседних репликонов. Скорость репликации зависит от количества вилок.
Точность репликации чрезвычайно высока: ошибки происходят с частотой, не превышающей 1:10-10, и корректируются ДНКполимеразами.
Благодаря полуконсервативному способу репликации обеспечивается удвоение наследственного материала в исходной материнской клетке, после чего она считается подготовленной к последующему делению на две дочерние клетки, в которые поровну распределяется удвоенная молекула ДНК т.е. в каждой из дочерних клеток количество ДНК, соответствует таковому в материнской клетке.
Теоретически возможны еще два механизма репликации: а) консервативный механизм, когда новая молекула состоит из двух новых цепей, а старая молекула - из двух старых цепей ДНК (напоминает механизм однородительской изодисомии - см. главу 28);
б) дисперсный механизм, когда каждая из двух вновь образованных молекул содержит в обеих цепях как новые, так и старые участки молекулы ДНК.
Распределение ДНК по дочерним клеткам обеспечивается в ходе митоза - деление соматической клетки, а также мейоза - деления половой клетки (см. главу 9).
Генетический контроль репликации и контроль клеточного цикла у человека осуществляет генная сеть, включающая более 400 генов, локализованных на хромосомах: 1-4, 6-9, 11, 12, 14, 16, 17, 19, 22 и Х-хромосоме.
В некоторых случаях происходит многократная репликация отдельных групп генов, являющаяся одним из важных механизмов регуляции генной активности, позволяющим синтезировать (амплифицировать) огромное число копий генов, которые затем транскрибируются, создавая необходимое количество требуемого генного продукта.
Транскрипция ДНК, процессинг и сплайсинг мРНК
Транскрипция ДНК заключается в считывании или переписывании наследственной информации, закодированной в конкретном участке матричной цепи ДНК, и образовании или синтезе мРНК.
Ключевой фермент РНК-полимераза состоит из трех типов:
- I тип находится в ядрышке и отвечает за транскрипцию рРНК;
- II тип находится в нуклеоплазме и обеспечивает 20-40% клеточной активности, отвечая за синтез гетерогенной ядерной РНК (гяРНК) - это предшественник мРНК;
- III тип находится в нуклеоплазме и отвечает за синтез малых ядерных РНК (мяРНК), тРНК и 5БРНК.
Вместе с тем, многие мяРНК транскрибируются РНК-полимеразой II типа.
В ходе транскрипции РНК-полимераза II «узнает» в молекуле ДНК промотор (место посадки РНК-полимеразы) и присоединяется к нему, а затем, перемещаясь вдоль ДНК, последовательно расплетает двойную спираль.
Начиная с промотора, на матричной цепи ДНК копируется ее зеркальное отражение - дочерняя цепь (или мРНК), по мере продвижения РНК-полимеразы постепенно отходящая от матрицы; двойная спираль ДНК позади фермента восстанавливается.
Когда РНК-полимераза II достигает конца копируемого участка, мРНК полностью отделяется от матрицы. Транскрипция завершается в области терминатора, расположенного в 3'-некодирующей части гена (см. ниже).
Транскрибируемые участки молекулы ДНК способны считываться с образованием активного функционального продукта в виде пре-
РНК (будущая мРНК).
Затем образовавшаяся пре-РНК подвергается процессингу (созреванию), в ходе которого из нее вырезаются интроны (некодирующая часть).
Процессинг мРНК включает следующее:
• 1. Происходит метилирование мРНК. На 5'-конце имеется КЭПмодифицированный в 7-м положении метилированный остаток гуанозин-5-трифосфата, соединенный с концевым нуклеозидом способом: 5'-5'. КЭП (определение см. ниже) участвует в регуляции, трансляции и стабилизации мРНК, защищая ее от действия 5'-эндонуклеаз.
• 2. Происходит кэпирование с участием гуанилтрансферазы и метилтрансферазы.
• 3. Происходит полиаденилирование или присоединение, последовательности полиадениловой кислоты к 3'-концу РНК после окончания ее синтеза с участием фермента поли(А)-полимеразы.
• 4. После вырезания интронов оставшиеся в мРНК экзоны (кодирующая часть) соединяются между собой в единую цепочку - это сплайсинг мРНК, или завершающая стадия ее созревания, т.е. заключительное событие процессинга мРНК.
В последние годы установлено: у эукариот за раскручивание цепи ДНК на участке транскрипции и за ее спирализацию после окончания синтеза мРНК, а также отсоединение транскрипта от нити ДНК, отвечают ДИК-топоизомеразы, а в процессе сплайсинга мРНК участвуют рибозимы РИК, обладающие ферментной активностью.
Эукариотические мРНК стабильны в течение нескольких часов и даже суток.
Таким образом, транскрипция, процессинг и сплайсинг происходят в ядре клетки. Затем мРНК готова к выходу из ядра в цитоплазму и трансляции в рибосомах.
Трансляция информации и биосинтез белка в рибосомах
Молекулы зрелой мРНК в виде гранул выходят из ядра в цитоплазму, где соединяются с рибосомами. Именно в них осуществляется трансляция или передача информации, записанной как последовательность нуклеотидов в мРНК, в последовательность аминокислот в молекулах белков (полипептидных цепочек).
Трансляция информации протекает параллельно с биосинтезом белков. В ее осуществлении принимает участие макромолекулярный комплекс, включающий: мРНК, рРНК, тРНК, амино-тРНК-
синтетазы, белковые факторы инициации, элонгации (удлинение или наращивание полипептида) и терминации трансляции.
Рибосомы - своеобразные «молекулярные машины», представляющие собой цитоплазматические структуры клетки, состоящие из двух неравных субъединиц с разными константами седиментации (60S и 40S) и содержащие все взаимодействующие в ходе трансляции молекулы, что обеспечивает биосинтез белков под контролем мРНК (рис. 3).
На этапе инициации трансляции малая субъединица рибосомы, инициаторная тРНК и фактор инициации «узнают» кодон-инициатор (ЛУГ) у 5'-конца молекулы мРНК.
Потом «включается» большая субъединица рибосомы, и в ней начинается биосинтез полипептидной цепочки, протекающий в три этапа: присоединение тРНК, образование пептидной связи и продвижение на три нуклеотида. Затем цикл повторяется.
При узнавании кодонов, являющихся стоп-сигналами, белковые факторы терминации транскрипции освобождают полипептидную цепь от рибосомы. Иными словами, биосинтез белка - это управляемая мРНК саморегулирующаяся система клетки.
В биосинтезе белка одновременно участвуют до 100 рибосом. Такой комплекс получил название полирибосомы.
Рекомбинация ДНК
Рекомбинация ДНК - это перегруппировка (перемешивание) генов, поступивших в молекулу ДНК по материнской (50% генов) и отцовской (50% генов) линии. Ее результатом, как правило, становится взаимный (реципрокный) или (редко) односторонний (нереципрокный) перенос участков ДНК с одной молекулы ДНК на другую молекулу ДНК (с несестринской хроматиды одной хромосомы на несестринскую хроматиду другой хромосомы).
Такой механизм обеспечивает эволюционную преемственность и уникальность наследственной информации в ряду поколений (за счет создаваемой гетерогенности), а также играет роль в рекомбинационной репарации ДНК (см. главы 5 и 10).
Репарация ДНК
Репарация ДНК - восстановление структуры молекулы ДНК в случае ее повреждения.
Абсолютной стабильности структуры ДНК не бывает. Это относится как к генам (участкам молекулы ДНК), так и к белкам, в том числе к белкам хромосом.
В соответствии с особенностями гомеостаза (см. ниже), контролируемого генотипом организма, в случае повреждений структуры гена (например, ошибок репликации ДНК) или производимого геном белка (например, ошибок транскрипции мРНК) вслед за дестабилизацией молекулярной структуры должна следовать ее стабилизация. Именно сохранение стабильной молекулярной структуры ДНК обеспечивают восстановительные (репарационные) механизмы, реализуемые с помощью разных ферментных систем клетки (см. главу 10).
Теперь рассмотрим хромосомный уровень организации наследственного материала.
Структурная организация хромосомы
Хромосома есть наследственный материал, организованный в виде хроматинового (гетерохроматинового) или нуклеопротеидного комплекса. Это молекула ДНК, связанная с множеством белковых молекул.
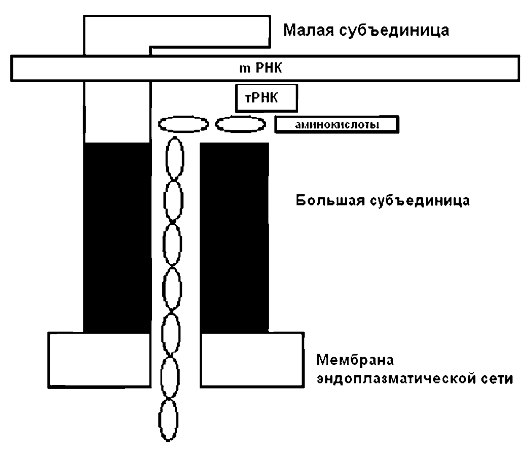 Рис. 3. Схема строения и функционирования рибосомы
Рис. 3. Схема строения и функционирования рибосомы
Хромосомы, видимые в световом микроскопе в метафазе митоза, - это метафазные хромосомы, или компактные хроматиновые структуры.
Прежде чем охарактеризовать морфологию хромосомы, рассмотрим особенности ДНК, которая в метафазной хромосоме находится в наиболее конденсированном состоянии: здесь она плотно упакована для доставки в дочерние клетки и поэтому функционально инертна (неактивна).
Степень конденсации ДНК в хромосоме очень важна, ибо в упакованном состоянии молекула недоступна для действия ферментов и сигнальных молекул.
Если оценить ДНК хромосомы по степени конденсации, то наиболее упакованными будут области центромера, состоящие из повторяющихся последовательностей ДНК (рис. 4).
Как показано на рис. 4, области центромера удерживают две сестринские хроматиды. На внешней поверхности располагаются комплексы белков кинетохора (область вокруг центромера), к волокнам которого прикрепляются микротрубочки митотического веретена.
После деления клетки ее хромосомы становятся менее упакованными и называются интерфазными хромосомами.
В отличие от ДНК метафазных хромосом, ДНК интерфазных хромосом функционально активна. Вместе с тем, она обладает еще большой степенью конденсации по сравнению с нитью метафазной
ДНК.
На рис. 5 приведено схематическое изображение гаплоидного (одинарного) набора хромосом человека (идиограмма) при их дифференциальном окрашивании. При таком окрашивании участки хромосом различаются по содержанию АТ- и ГЦ-пар оснований. В частности, Q-сегменты или совпадающие с ними G-сегменты соответствуют участкам, богатым АТ-парами, и содержат тканеспецифические гены, реплицирующиеся во второй половине фазы синтеза ДНК (S-период). R-сегменты соответствуют участкам, богатым ГЦ-парами, и содержат общеклеточные гены, или гены «домашнего хозяйства», реплицирующиеся в первой половине S-периода (см. главу 9).
На рис. 6 представлены варианты схематического изображения Х-хромосомы человека при дифференциальном окрашивании.
Ядерная ДНК
Согласно результатам исследований, структура ДНК включает следующее.
1. Уникальные последовательности (или экзоны) - кодирующую часть молекулы. Они, как правило, - единичные копии ДНК. Имеют размеры от нескольких сот до двух-трех тыс. н.п. Тысяча пар - одна килобаза (кб), миллион пар - одна мегабаза (мгб).
Экзонами представлено большинство структурных генов. Они локализованы по всему геному (в основном между ALU-повторами) и занимают около 3-5% общего объема.
Не вся кодирующая ДНК представлена одними экзонами.
Во-первьх, в диплоидном ядерном геноме каждая такая последовательность уже не уникальна, потому что представлена двумя аллельными копиями, локализованными в идентичных локусах на гомологичных хромосомах отцовского и материнского происхождения. Во-вторых, существует множество рассеянных по всему геному неаллельных кодирующих (высокогомологичных) последовательностей ДНК, повторяю-
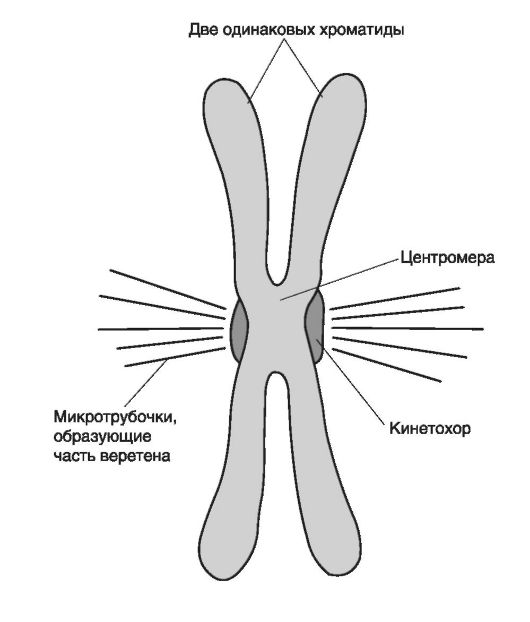
Рис. 4. Морфология хромосомы (по: В. Эллиот, Д. Эллиот, 2002)
щихся многократно. Именно они формируют многочисленные семейства генов, объединенных в кластеры (пучки), включающие: гены, гомологичные по коротким мотивам; гены, кодирующие протяженные высококонсервативные домены; псевдогены (поврежденные копии или фрагменты генов); непроцессированные и процессированные гены (как экспрессирующиеся, так и неэкспрессирующиеся).
2. Повторяющиеся последовательности (или интроны) - некодирующая часть молекулы. Они могут располагаться как в самих генах, так и между ними. Во втором случае их называют спейсерами или межгенными промежутками.
 Рис. 5. Схематическое изображение хромосом человека (идиограмма гаплоидного набора):
Рис. 5. Схематическое изображение хромосом человека (идиограмма гаплоидного набора):

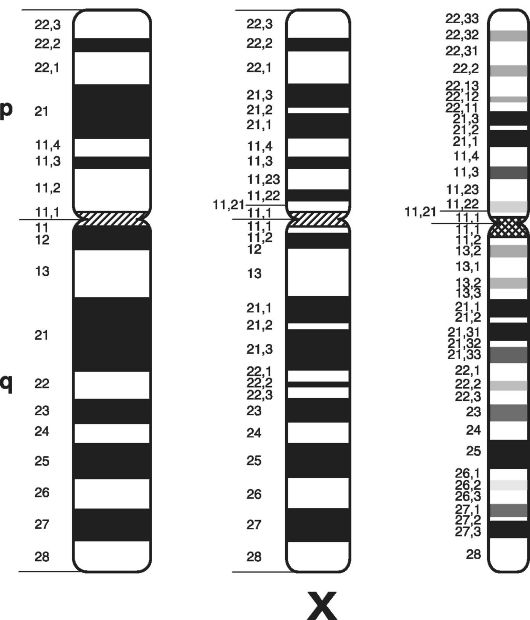 Рис. 6. Варианты
схематического изображения Х-хромосомы человека: левая часть рисунка -
указаны локусы и районы; центральная часть - локусы, районы и сегменты;
правая часть - локусы, районы, сегменты и субсегменты;
Рис. 6. Варианты
схематического изображения Х-хромосомы человека: левая часть рисунка -
указаны локусы и районы; центральная часть - локусы, районы и сегменты;
правая часть - локусы, районы, сегменты и субсегменты;
р - короткое плечо; q - длинное плечо хромосомы
На долю повторяющихся последовательностей ДНК, находящихся внутри генов, приходится 35% общего объема генома. Повторы внутри генов и повторы между генами разделены на длинные (LINE-повторы или L1-элементы), занимающие 20% объема, средние (SINE-повторы или элементы) - 10%, короткие (ALU-повторы или элементы) - 10%, сателлитную ДНК - 15% и другие повторы - 5%.
Размеры (длина) и количество повторов: L1-элементы - свыше 6 кб, их до 100 тыс. копий; SINE-элементы - 0,34-6 кб, их свыше 100 тыс. копий; ALU-элементы - до 0,3 кб, их до 900 тыс. копий.
L1- и ALU-элементы семейства локализованы по всему геному. Например, если хромосому 21 разрезать с помощью рестриктазы на 50 фрагментов (всего в этой хромосоме около 51 млн н.п.), то в большинстве их (но не во всех) обнаружатся как L1-элементы, так и ALU-элементы.
Сателлитная ДНК - это семейство высокоповторяющихся последовательностей (от 5-10 до 170-340 н.п.), локализованных в основном в гетерохроматиновых районах.
Функции повторяющихся последовательностей ДНК остаются пока плохо изученными, хотя уже получен ряд интересных данных (см. главы 1 и 27).
Следует учитывать: структурные и функциональные особенности хромосомной ДНК крайне важны при рассмотрении вопроса об ее преобразованиях в клетке и организме.
Чуть более 50% хромосомной ДНК (22 аутосомы + 1 Х-хромосома + 1 «митохондриальная» хромосома) поступает в будущий организм по материнской линии (вместе с яйцеклеткой), а чуть менее 50% (22 аутосомы + 1 Y- или Х-хромосома) - по отцовской линии (вместе со сперматозоидом, см. главу 5). В результате оплодотворения в зиготе формируется единый комплекс хромосом (кариотип) и единый комплекс расположенных в них генов (генотип).
Говоря об организации ДНК на хромосомном уровне, следует особо выделить ее уникальность, основанную на гетерогенности (разнообразии) молекулярной структуры. Этот термин был впервые применен С.Н. Давиденковым в конце 30-х годов XX в. для обозначения разнообразия генетических причин наследственных болезней нервной системы, а в дальнейшем стал широко использоваться для других целей, например для обозначения гетерогенности белков (по первичной структуре, аминоконцевой последовательности, белковому пику и другим особенностям).
В случае гетерогенности ДНК имеются в виду уникальные различия между двумя случайно выбранными гомологичными хромосомами любой одной пары или различия по одному нуклеотиду в среднем на каждые 300-500 нуклеотидов (всего в геноме человека 3,165 млрд п.н.). Именно эта особенность лежит в основе метода ДНК-диагностики при идентификации личности человека, а также тестировании генов наследственных болезней. По мнению американских ученых, разрешающая способность метода ДНК-диагностики
абсолютна: 99,999999%.
Белки хромосомы
ДНК в составе хромосомы соединена с белками - это нативный хроматин, представленный гистонами (основные или щелочные белки), богатыми аргинином и лизином; гистоны, благодаря своему положительному заряду, образуют ионные связи с отрицательно заряженными фосфатными группами, расположенными на внешней стороне двойной спирали ДНК.
Среди гистонов выделены 5 классов: Н1, Н2А, Н2В, Н3 и Н4. Гистон Н1 вдвое длиннее других и отвечает за целостность нуклеосомной структуры хромосомы: на одну молекулу гистона Н1 приходится одна нуклеосома.
Стабилизация нативного хроматина обеспечивается взаимодействием в основном двух гистонов: Н1 и Н3. Причем, не исключено участие в ней еще и негистоновых (кислых) белков; их количество невелико и варьирует.
Негистоновые белки участвуют в образовании длинных петель ДНК, прикрепленных к осевым белковым структурам.
Природа негистоновых белков пока плохо изучена. В их состав входят сложные белки, ферменты, а также регуляторные белки.
Дискретной единицей хромосомы служит нуклеосома - частица размерами около 200 н.п. с диаметром около 10 нм и белковым остовом (гистоновый октамер), включающим по две молекулы каждого из гистонов (кроме Н1) - это хроматин, напоминающий «бусины на нити».
Вокруг белкового остова по его длине накручена линкерная ДНК (30-40 н.п.); ее диаметр - 2 нм (рис. 7).
Укладка ДНК на нуклеосомном уровне - I уровень структурной организации хромосомной нити.
На II супернуклеосомном уровне организации диаметр хромосомной нити (хроматиновой фибриллы) равен 30 нм - это, по-видимому, уровень метафазной хромосомы (модель соленоида). На данном уров-
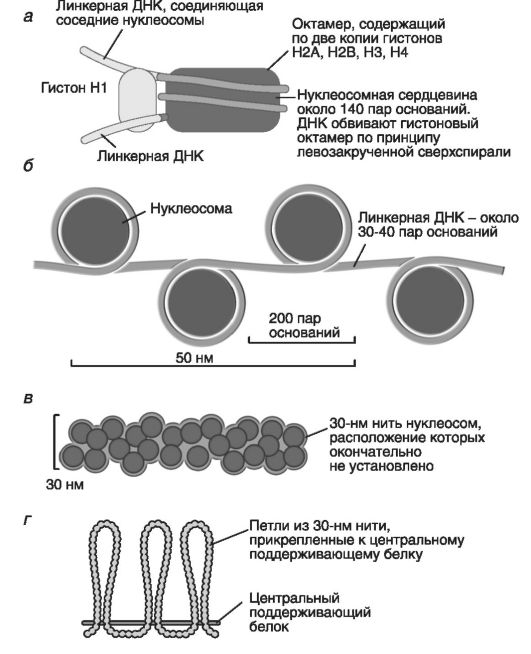 Рис. 7. Компактизация хроматина на нуклеосомном уровне (по Албертс Б. и соавт., 1994):
Рис. 7. Компактизация хроматина на нуклеосомном уровне (по Албертс Б. и соавт., 1994):
а - нуклеосома; б - нуклеосомы с линкерными участками ДНК; в - нить хроматина 30 нм толщиной; г - петли хроматиновой нити, прикрепленные к центральному поддерживающему белку
не хроматиновая фибрилла состоит из упакованных нуклеосом и ее петли прикреплены к центральному поддерживающему белку.
Предполагается существование еще III уровня структурной организации хромосомной нити - вероятно, он должен соответствовать структурам интерфазного ядра, наблюдаемым под световым микроскопом (рис. 8).
Интроны как участки хромосом могут входить или не входить в структуру генов. Как правило, интроны не содержат информацию, необходимую для синтеза белка. В таких участках хромосомная ДНК остается в конденсированном, генетически инертном (неактивном) состоянии - это гетерохроматин. В то же время функционирующие области хромосом (или эухроматин) менее конденсированы и более активны.
Морфологическая неоднородность хромосом определяется изменчивостью конденсации ДНК. В частности, проходя к началу митоза полный цикл конденсации, одни районы интерфазных хромосом деконденсируются в эухроматин, а другие районы остаются в конденсированном состоянии (гетерохроматиновом) в течение всего митотического цикла.
В метафазной хромосоме такое подразделение на эухроматин и гетерохроматин сохраняется, и гетерохроматин наиболее интенсивно
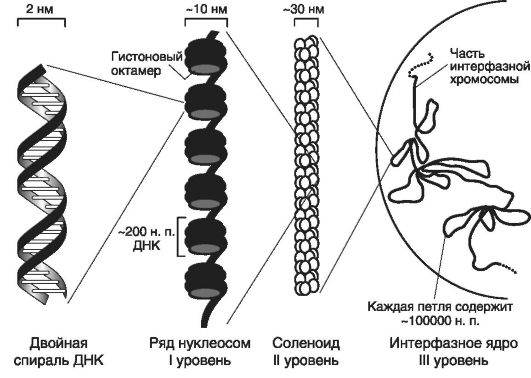 Рис. 8. Уровни структурной организации хромосомной нити (по Томпсон M. и соавт., 1991)
Рис. 8. Уровни структурной организации хромосомной нити (по Томпсон M. и соавт., 1991)
выявляется в виде не полностью конденсированных участков (или вторичных перетяжек).
Например, вторичные перетяжки длинных плеч хромосом 1, 9 и 16, коротких плеч акроцентрических хромосом и длинного плеча Y-хромосомы бедны структурными генами или не содержат их вообще.
Наоборот, эухроматин реализует информацию посредством мРНК.
Хромосома - структура с большим преобладанием линейного размера над поперечным. В метафазе митоза общая длина гаплоидного набора хромосом при средней степени их конденсации составляет около 1500 мкм с поперечником хроматиды около 0,5 мкм. Сопоставление этих величин говорит об исключительных масштабах физического преобразования хромосомной нити при ее переходе из интерфазного в метафазное состояние, включающем два взаимосвязанных процесса: многократное уменьшение длины и многократное увеличение поперечника хромосомы.
Митохондриальная ДНК
Митохондриальная ДНК (мтДНК) представляет собой полуавтономную генетическую систему, работающую под контролем ядерной ДНК. Это единственная кольцевая ДНК, имеющая размер в
16 569 н.п. (рис. 9).
В настоящее время описаны особенности строения и функционирования мтДНК (их не менее 10), отличающиеся от ядерной ДНК.
Во-первых, мтДНК полуавтономна (существует возможность ее переноса между культивируемыми клетками человека в процессе слияния и разъединения органелл).
Во-вторых, мтДНК поступает в клетку исключительно по материнской линии (см. главу 26).
В-третьих, мтДНК подвергается репликативному отбору как при митозе, так и при мейозе. Поэтому в одной клетке обнаруживается смесь мутантной и нормальной мтДНК (гетероплазмия).
В ходе митотического и мейотического цитокинеза доля и той, и другой мтДНК значительно варьирует в дочерних клетках вплоть до образования в них гомоплазмии.
Названные механизмы имеют большое значение в развитии митохондриальных болезней, при которых расстройства энергетического метаболизма в клетках прямо пропорциональны доле мутантных копий мтДНК в митохондриях; она составляет от 0 до 100%.
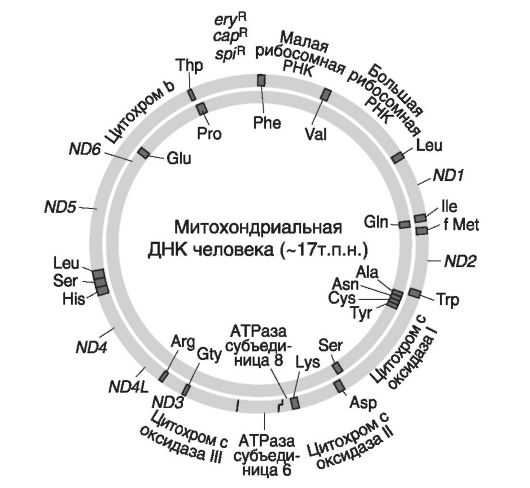 Рис. 9. Карта мтДНК у человека (по: Гриффитс А. и соавт, 2000)
Рис. 9. Карта мтДНК у человека (по: Гриффитс А. и соавт, 2000)
В-четвертых, для мтДНК характерен феномен пороговой экспрессии (доля мутантных копий) в той или иной ткани. Пороговая экспрессия наиболее высока в нервной ткани, поперечно-полосатых мышцах, сетчатке глаз, островках Лангерганса и других тканях.
В-пятых, эволюционная изменчивость мтДНК в 10-20 раз выше, чем ядерной ДНК (в первом случае мутации возникают чаще, чем во втором).
Хотя только 20% участников (компонентов) процессов окислительного фосфорилирования кодируются генами митохондриального генома, а остальные 80% - генами ядерной ДНК, нарушения тканевого дыхания возникают гораздо чаще из-за мутаций в мтДНК.
В-шестых, в мтДНК нет интронов.
В-седьмых, в большей части мРНК, содержащейся в мтДНК, отсутствуют 5'- и З'-нетранслируемые последовательности.
В-восьмых, двойное кольцо мтДНК имеет внутреннюю тяжелую цепь (Н-цепь) и внешнюю легкую цепь (L-цепь). У них разная плотность генов: в первой цепи больше пурина, во второй - пиримидина.
В митохондриальной ДНК есть в легкой цепи некодирующий район (или D-петля), служащий регуляторной областью и местом локализации участка начала репликации Н-цепи и транскрипции.
Синтез Н-цепи идет по часовой стрелке вокруг мтДНК; после прохождения 2/3 цепи начинается репликация L-цепи против часовой стрелки, т.е. репликация мтДНК - двухступенчатый асинхронный процесс.
В-девятых, выявлены отличия генетического кода мтДНК от кода ядерной ДНК. Например, метионин в мтДНК кодирует кодон АУА, а в ядерной ДНК - кодон АУГ; стоп-кодоны мтДНК - триплеты АГА и АГГ, тогда как в ядерной ДНК они кодируют аргинин.
В-десятых, наличествуют и другие особенности, связанные с транскрипцией, локализацией ферментов и т.д.
ОСНОВНЫЕ ПОНЯТИЯ СТРУКТУРНОЙ ГЕНОМИКИ
Ген и модель его общей структуры
Под понятием «ген» подразумевается структурно-функциональная единица наследственного материала (генома).
Ген - это участок молекулы ДНК, кодирующий молекулу полипептида (белка), молекулы мРНК, рРНК, тРНК.
В последние годы произошло очередное уточнение понятия. Теперь считается, что ген - ассоциированный с регуляторными последовательностями фрагмент молекулы ДНК, соответствующий единице транскрипции на хромосоме.
Согласно модели общей структуры гена, он - «конкретная единица молекулы ДНК (ее кодирующая и некодирующая части) с расположенными по флангам регуляторными последовательностями».
Ген имеет (рис. 10) слева 5'-конец (начало гена), а справа З'-конец (окончание гена); между ними находятся экзоны, интроны и другие структуры.
Исключением из этой модели являются гены альфа- и бетаинтерферонов, гистонов и гены мтДНК, у которых нет интронов.
Место расположения гена в хромосоме (его позиция в ней) называется генным (генетическим) локусом.
Состояние локуса играет принципиальную роль: в компактном виде он интактен, и РНК-полимераза лишена доступа к промотору. В результате декомпактизации локус приобретает форму петли, связанной с ядерным матриксом (внутренний белковый скелет ядра).
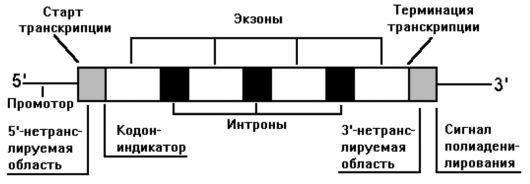 Рис. 10. Модель общей структуры гена человека
Рис. 10. Модель общей структуры гена человека
На рис. 11 приведена структура бета-глобинового гена человека. Это самый короткий ген у человека: в нем 1100 н.п., образующих 3 экзона (90, 222 и 126 н.п.) и 2 интрона (116 и 646 н.п.).
Как следует из рис. 10 и 11, в ген входят следующие тонкие структурные элементы.
• Промотор, или место присоединения РНК-полимеразы. Он включает ТАТА-бокс (остатки тимина и аденина), ГЦ-бокс (остатки гуанина и цитозина) и ЦААТ-бокс (остатки цитозина, аденина и тимина). Боксы расположены перед точкой инициации транскрипции (кэп-сайт). Следует отметить: ни один из элементов промотора не является обязательным для транскрипции, поэтому в разных генах разные комбинации этих элементов. Например, в 20% всех генов человека (включая гены «домашнего хозяйства») нет ТАТА-боксов, а некоторые гены имеют множественные копии одного и того же элемента или лишены их полностью (ЦААТбоксы, ГЦ-боксы и др.).
• Кэп-сайт - основной инициаторный комплекс (ОИК). Он необходим для генов, транскрибируемых РНК-полимеразой II (гены II типа). Для присоединения РНК-полимеразы II к ОИК требуется ряд белков - общих факторов транскрипции (они есть во всех клетках).
• Экзоны и разделяющие их интроны.
• СрО - островки (динуклеотиды) или участки, разделяющие экзоны и интроны (на рис. 11 это: аа31, аа104, аа105, аа146); они расположены не внутри, а в начале гена (около 5'-конца), что позволяет предположить их участие в регуляции.
• 5'- и З'-нетранслируемые области.
• Небольшие регуляторные сайты, участвующие в инициации (кодон АТТ) или терминации трансляции (кодон ТАА).
• Небольшие регуляторные участки, необходимые для инициации, усиления (энхансеры), ослабления (сайленсеры) и терминации транскрипции.
• Участок сигнала полиаденилирования.
Почти все мРНК имеют «поли-А-хвост«, присоединяющийся к З'-концу перед выходом молекулы мРНК из ядра. Считается, что «поли-А-хвост» защищает мРНК от быстрой деструкции, ибо деаденилирование часто ей предшествует.
Предполагается также, что такой «хвост» влияет на транспорт, трансляцию и распад мРНК, значительно сокращая период ее полужизни.
В последние годы обнаружен белок, связывающийся с «хвостом», но механизм его действия пока неясен.
Размеры других генов человека весьма вариабельны. Как отмечалось, самый короткий из них - бета-глобиновый ген, ну а самый протяженный - ген дистрофина (2,2 млн н.п. и более 2000 экзонов).
Зная размеры гена, можно выяснить размер кодируемого им белка (и наоборот), поскольку одна аминокислота закодирована тремя нуклеотидами. Например, белок, кодируемый геном с размером 3000 н.п., содержит 1000 аминокислотных остатков.
Свойства гена как единицы функционирования
Главное свойство гена - его экспрессия или работа, в ходе которой последовательность нуклеотидов в молекуле ДНК переводит-
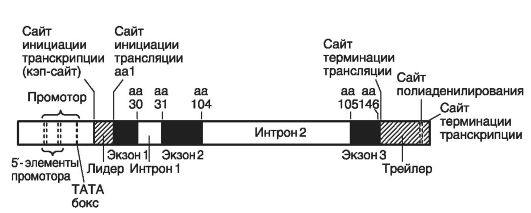 Рис. 11. Структура бета-глобинового гена человека (по: Гилберт С., 1995)
Рис. 11. Структура бета-глобинового гена человека (по: Гилберт С., 1995)
ся в последовательность аминокислот в молекуле полипептида (см. главу 3).
Следующее свойство гена - аллельное состояние. Организационная структура гена - диаллельная модель (рис. 12), т.е. большинство генов существуют в виде двух альтернативных вариантов - аллелей, локализованных в одинаковых (идентичных) локусах на гомологичных хромосомах.
В диплоидной клетке каждая пара гомологичных хромосом содержит одну хромосому отцовского происхождения (имеет отцовские аллели) и другую хромосому материнского происхождения (материнские аллели).
Гены, расположенные в одинаковых локусах на гомологичных хромосомах, называются аллельными генами, т.е. у каждой пары родительских генов одна материнская и одна отцовская копии (аллели) одного и того же гена.
Если гены одной родительской пары находятся в разных (неидентичных) локусах как гомологичных, так и негомологичных родительских пар хромосом, то это неаллельные гены.
В классической генетике выделен ряд понятий, связанных с аллельными генами.
Во-первых, «гомозиготность» и «гетерозиготность». Термины введены в 1903 г. одним из основоположников генетики - англичанином Вильямом Бэтсоном.
В случае гомозиготности каждый из двух аллелей одного и того же гена обладает одинаковой по содержанию информацией. В случае гетерозиготности каждый из двух аллелей одного гена обладает разной по
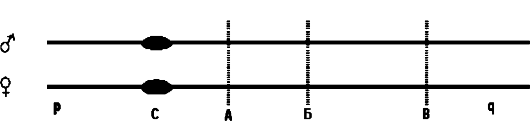 Рис. 12. Диаллельная
модель организационной структуры гена Гомологичные хромосомы: отцовская
(верхняя) и материнская (нижняя); р - короткое плечо; q - длинное
плечо; С - центромера; А, Б и В - идентичные генные локусы длинных плеч
двух хромосом и содержащиеся в них аллельные копии одного гена
(соответствуют местам пересечения пунктирных линий с хромосомами)
Рис. 12. Диаллельная
модель организационной структуры гена Гомологичные хромосомы: отцовская
(верхняя) и материнская (нижняя); р - короткое плечо; q - длинное
плечо; С - центромера; А, Б и В - идентичные генные локусы длинных плеч
двух хромосом и содержащиеся в них аллельные копии одного гена
(соответствуют местам пересечения пунктирных линий с хромосомами)
содержанию информацией. Организмы с такими аллельными парами называются гомозиготными и гетерозиготными соответственно.
Во-вторых, аллельные и неаллельные гены образуют в одной и той же клетке две группы родительских генов (отцовскую и материнскую по происхождению).
В каждой группе любой ген отличается от альтернативного ему гена по содержанию информации, закодированной в его нуклеотидной последовательности.
В-третьих, если речь идет о совокупности (сцеплении) аллелей одного и того же гена (например, аллелей полиморфного маркера), то такое состояние называется гаплотипом (см. ниже), а локусы, где расположены подобные аллели, называются полиморфными локусами (см. ниже). Их в геноме у человека выделено около 6 тыс.
Обнаружение в одной популяции неравновесия в сцеплении полиморфных аллелей одного локуса позволяет определить точку повреждения гена (точковую мутацию) и время (возраст) ее появления.
Другие свойства гена.
• Дискретность. У биологического вида развитие разных признаков контролируется разными генами, локализованными в разных локусах на разных хромосомах. Например, ген хореи Гентингтона расположен на коротком плече хромосомы 4, сегмент 1, подсегмент 6 (записывается как 4р16), ген атаксии Фридрейха - на длинном плече хромосомы 9, сегменты 1 и 2, подсегменты 3 и 1.1 (записывается как 9q13-q21.1).
• Дозированность. Ген обусловливает развитие признака до определенного количественного предела (дозы); параллельно дозе гена усиливается выраженность (экспрессивность) признака. Превышение дозы в рамках одного биологического вида не наблюдается. Например, у человека размеры ушной раковины или длина спинки носа имеют определенные пределы, характерные для Homo sapiens, и никогда не достигнут размеров ушной раковины или длины хобота у слона.
• Специфичность. Ген обеспечивает развитие только определенного признака или их группы. Например, ген, контролирующий окраску глаз, не способен сформировать такой признак, как курчавость волос.
В случае развития группы признаков под влиянием одного гена речь идет о плейотропном действии или эффекте (см. главу 5). Например, ген атаксии-телеангиэктазии (11q22.3) ответственен за
следующие признаки: нарушение координации и походки, появление сосудистых звездочек (телеангиэктазий) на склерах, коже, слизистых оболочках органов дыхания и пищеварения, а также иммунодефицит и др.
• Стабильность. При отсутствии мутаций (нарушений структуры и функции) ген передается в ряду поколений в стабильном виде (см. главу 5).
Гаплотип
Гаплотип - совокупность или сочетание (сцепление) аллелей одного и того же гена.
Аллели, входящие в гаплотип, происходят из одного базового аллеля, т.е. для них характерен «эффект основателя».
Новые аллели мутантного гена, сформированные на отцовской или материнской хромосоме, в последующих поколениях постепенно «размываются» за счет рекомбинационной изменчивости и меняют содержание информации о признаке.
Аллели одного гаплотипа, как и множественные аллели одного гена (см. ниже), кодируют количественные признаки. Однако для вторых аллелей не характерен «эффект основателя».
Классификация генов
Гены человека делятся на ряд классов: белок-кодирующие; гены, кодирующие РНК; гены РНК-полимераз; митохондриальные гены.
Класс белок-кодирующих генов: это общеклеточные гены (гены «домашнего хозяйства»); тканеспецифические гены (гены терминальной дифференцировки или специфически функционирующие гены); гены, кодирующие транскрипционные факторы: PAX I-9, HOX I-9, SOX I-10 (см. главу 8); ген SRY (ген, определяющий мужской пол). В последние годы в этот класс включены гены, контролирующие:
• структуру белков, участвующих в процессах синтеза аминокислот, азотистых оснований и сахаров;
• синтез белков, обеспечивающих матричные процессы (репликацию, транскрипцию и трансляцию).
Следует выделить уникальные особенности строения некоторых белок-кодирующих генов - они включают две области: структурную часть и фланкирующие регуляторные участки с 5'- и 3'-сторон. Например, глобиновые гены человека образуют два мультигенных семейства (кластера), структурно напоминающих оперон у прока-
риот, но без общих регуляторных участков (промотора, оператора и терминатора), ибо каждый кластер имеет свои регуляторные участки. В частности, первый кластер альфа-глобиновых генов локализован в коротком плече хромосомы 16 в определенной последовательности: сначала идут последовательности генов, кодирующих цепи эмбриональных гемоглобинов, затем - цепи трех псевдогенов, а ближе к 3 '-концу - цепи генов альфа 1 и 2.
Второй кластер бета-глобиновых генов расположен в коротком плече хромосомы 11 в следующей последовательности: сначала ген эмбриональной цепи, затем 2 гена фетальных цепей HbF, которые отличаются по одному кодону, потом один псевдоген бета-цепи, сигма-ген, кодирующий сигма-цепь гемоглобина взрослого человека HbA2, и бета-ген.
Эти гены функционируют на разных этапах онтогенеза в той же последовательности.
Класс РНК-кодирующих генов: гены, отвечающие за синтез мРНК,
рРНК, тРНК и мяРНК.
Гены тРНК повторяются умеренно, числом достигая 1300. Особенность их строения: расчлененный и находящийся внутри гена промотор, один блок которого расположен на расстоянии 8-30 н.п. слева от инициаторного кодона, а второй блок - справа, на расстоянии 51 н.п.
Сам промотор содержит А- и В-боксы с каноническими последовательностями. Если расстояние между боксами уменьшается, то уровень транскрипции снижается либо траскрипция полностью прекращается. В свою очередь, замена одного нуклеотида в В-боксе искажает структуру тРНК.
Крайне интересны данные о ранее неизвестных молекулах мяРНК. По-видимому, они участвуют в образовании гибридных молекул сложных рибопротеиновых частиц, называемых сплайсосомами. Процесс формирования сплайсосом идет путем спаривания с основаниями консенсусных последовательностей ДНК или меток сочленений экзонов с интронами: GU-начало метки и AG-конец метки. Такое спаривание происходит в местах сплайсинга на первичном РНК-транскрипте, что определяет интроны, подлежащие удалению.
Мутации в мяРНК могут привести к неправильному сплайсингу, влекущему за собой невозможность удаления интронов из РНКтранскрипта. Например, при бета-талассемии не происходит образо-
вания нормальных количеств бета-гемоглобина из-за замены гуанина на аденин в 5'-конце, что вызывает блок процессинга мРНК.
Иными словами, молекулы мяРНК катализируют химические процессы.
В последние годы предложена новая классификация, где в зависимости от типа транскрибируемых РНК-полимераз выделяют 3 класса генов:
• гены рРНК, транскрибируемые РНК-полимеразой I (это гены 18S; 5,8S и 28S);
• гены, транскрибируемые РНК-полимеразой II (это гены, кодирующие белки);
• гены, транскрибируемые РНК-полимеразой III (это гены 5SpРНК,
тРНК и мяРНК).
Класс митохондриальных генов: 37 генов, не содержащих интронов: гены рибосомных ^S-рРНК и ^S-рРНК; 22 гена тРНК; гены трех субъединиц цитохром-с-оксидазы; гены шестой и восьмой субъединиц АТФ-азы; ген цитохрома b; гены семи субъединиц NADH- дегидрогеназы.
Генетический полиморфизм
Генетический (геномный) полиморфизм (ГП) - вариации наследственного материала в пределах одного биологического вида (или основа разнообразия людей).
В классической генетике понятие ГП соответствует нескольким наследственным вариантам, из которых наиболее редкий вариант встречается с частотой, превышающей частоту обратного мутирования (см. главу 5).
Если говорить о ГП как о генетических маркерах признаков организма, то их можно считать полиморфными только тогда, когда наиболее редкий их вариант встречается чаще, чем в 1% случаев.
Речь идет о частоте встречаемости четвертого варианта диаллельной модели, когда изменены обе аллельные копии одного и того же гена.
Как известно, при первом варианте и материнская, и отцовская копии сохраняют стабильную наследственную информацию о признаке; второй вариант - материнская копия несколько изменена, а отцовская - нет; третий вариант - отцовская копия несколько изменена, а материнская - нет; четвертый вариант - обе копии несколько изменены.
Наличие разных вариантов существования диаллельной модели традиционно (хотя и не совсем верно) считается множественным аллелизмом (см. выше). Но так как аллелей всего только два, а вариантов не менее четырех (точнее, их всегда гораздо больше), то правильнее следовало бы назвать такое состояние многовариантностью существования двух аллелей одного и того же гена.
Вместе с тем, известны серии множественных аллелей. Мутации в них по отношению друг к другу могут быть доминантными, рецессивными, а также вероятны неполное доминирование, сверхдоминирование, кодоминирование, условное доминирование, плейотропный эффект (см. главу 4).
Локусы, в которых расположены такие аллели, называются полиморфными (см. выше). Например, у человека в полиморфных локусах находятся полиморфные семейства молекул HLA-антигенов !иП классов (см. главу 15).
Другой пример полиморфности локусов - разная частота распространенности IV группы крови (по классификации АВО): среди населения ряда регионов мира она встречается у 3-4%, тогда как в других регионах - у 10-12%.
В молекулярном плане ГП - это наличие небольших (совместимых с жизнью) нарушений структуры нуклеотидов в молекуле ДНК (нарушений порядка последовательности) и, следовательно, в молекуле белка (см. главу 3).
В структурной геномике главной формой молекулярного полиморфизма является однонуклеотидный полиморфизм (ОНП) или варианты последовательностей ДНК у разных лиц с вовлечением одной пары нуклеотидов. ОНП характерен для 93% генов человека.
Выделяют качественный и количественный ГП. Первый представляет собой замену одного нуклеотида (или SNP), его частота 1:300 н.п. Второй - повторы нуклеотидов с различной степенью выраженности.
ГП - это также вариабельность числа тандемных повторов (одинаковые повторы следуют друг за другом) или STR в виде одного, двух, трех повторов и более.
Если протяженность тандемных повторов еще больше (от 10 до 100 нуклеотидов), то имеет место генетический полиморфизм по размеру повторов, или VNTR.
Кроме того, в геноме человека выделены области протяженной гомологии, возникшей в результате дупликации относительно круп-
ных фрагментов ДНК. Их эволюционное значение трудно переоценить. Например, рассмотрим дупликацию фрагментов ДНК, включающих ген CYP21 - ген цитохрома Р450 - и гены комплемента. В ходе эволюции одна из копий гена - ген CYP2^ - порой «накапливает» мутации и лишается активности, превращаясь в псевдоген. В результате между псевдогеном CYP2^ и сохранившим свою активность геном CYP2? может произойти генная конверсия (в гомологичных хромосомах) с перемещением участка гена CYP2? на ген CYP2^; или возникнет делеция гена CYP2?, обусловив развитие адреногенитального синдрома с гиперплазией коры надпочечников, вызванной недостаточностью 21-гидроксилазы.
Другой пример - дупликация гена миелинового белка РМР22; она вероятна в результате гомологичной рекомбинации между крупными повторами (REP), фланкирующими содержащую ген область ДНК (1,5 млн н.п.). Такая дупликация - наиболее частая причина невральной амиотрофии Шарко-Мари-Тус.
Вместе с тем делеция гена, кодирующего этот белок, ведет к менее тяжелой по проявлениям болезни - наследственной нейропатии.
Еще одна важная особенность ГП - различия между аллелями по числу копий повтора.
Исключение из этого правила - тринуклеотидный повтор цитозин-аденин-гуанин - ЦАГ (кодон глутаминовой кислоты), способный включаться в структурные гены в случае болезней экспансии числа нуклеотидных повторов (см. главу 28).
В целом установлено: разнообразные аллельные формы ГП могут образовываться за счет точковых мутаций, рекомбинаций и неравного кроссинговера.
В настоящее время создаются карты SNP, насыщенные SNP- маркерами. С их помощью проводится идентификация генов, ответственных за широко распространенные мультифакториальные и онкологические заболевания, болезни с отсроченной манифестацией признаков.
Особая роль принадлежит полиморфизму митохондриальной ДНК
(мтДНК).
В зиготе и во всех произошедших из нее соматических клетках при формировании многоклеточного организма транскрибируются и затем транслируются обе цепи мтДНК материнского происхождения, образующие единый транскрипт, расщепляющийся по сайтам, расположенным по обе стороны от каждого гена тРНК. Большая
часть генов тРНК (14 из 22) транскрибируется по часовой стрелке. После вырезания тРНК из мтДНК освобождаются молекулы рРНК
и мРНК.
В случае возникновения мутаций в мтДНК в клетке наблюдается гетероплазмия или присутствие разных типов мтДНК (нормальная и мутантная); при гомоплазмии все типы мтДНК одинаковые.
Мутации в мтДНК делятся на точковые и структурные (делеции). Они ведут к изменению белков, входящих в комплексы дыхательной цепи митохондрий, и, следовательно, обусловливают снижение энергетического обеспечения клеток или дефицит синтеза АТР, что ведет к болезням тех органов и тканей, в которых происходят энергоемкие процессы.
ГЕНОТИП И ФЕНОТИПИЧЕСКОЕ ПРОЯВЛЕНИЕ
Генотип человека
Генотип - совокупность всех генов организма (или полигенный комплекс). Регистрируемый результат действия генотипа - фенотипическое проявление (см. главу 3).
Уточнение о фенотипическом проявлении необходимо потому, что в геноме человека наряду с кодирующими последовательностями ДНК имеются некодирующие, неэкспрессирующиеся (нетранскрибирующиеся) последовательности без фенотипического проявления - псевдогены (или ложные гены).
В результате выполнения международной программы «Геном человека» установлено: генотип человека включает 35-40 тыс. генов. Такие данные соответствуют ранее известным - о выделении 35 тыс. типов мРНК в культуре злокачественных клеток человека, названной клетками HeLa (инициалы одной онкологической больной). Каждый тип мРНК есть результат транскрипции индивидуального структурного гена (для сравнения: генотип кишечной палочки - 4 тыс., дрожжей - 7 тыс. генов).
Генотип человека с одной стороны - основа его гомеостаза (см. ниже), а с другой стороны - основа программы его онтогенеза.
В частности, индивиды с генотипами, не обеспечивающими подавление отрицательного действия патологических генов и факторов среды, оставляют меньше потомков, чем те, у которых нежелательные эффекты подавляются.
Подавление генотипом отрицательных эффектов зависит от активности имеющихся в любой клетке генов-модификаторов. Они модифицируют действие нормальных аллелей в аллельных и неаллельных парах таким образом, что те начинают оказывать доминантное влияние на патологические гены.
В целом же генотип - основа эволюционной стабильности биологического вида, обеспечиваемой уникальными свойствами молекулы ДНК. В свою очередь, стабильность генотипа обусловливает надежность гомеостаза как отдельных клеток, так и организма в целом.
Гомеостаз и системный характер организации молекулярной жизни
Гомеостаз - фенотипическое проявление генотипа, свойство организма сохранять постоянство (целостность) внутренней среды при совместимых с жизнью условиях внешней среды.
Гомеостаз базируется на стабильности разнообразных и бесконечных в течение жизни генетически детерминированных преобразований или межмолекулярных реакций, сложившихся в ходе эволюции. Среди них реакции: ДНК-ДНК, ДНК-РНК, РНК-РНК, белок-ДНК, белок-РНК, белок-белок, белок-субстрат.
Выделяют разные виды гомеостаза:
• структурный гомеостаз, целостность морфологической организации на разных уровнях дискретности организма;
• химический гомеостаз, целостность состава внеклеточной (жидкой) части внутренней среды (кровь, лимфа, цереброспинальная жидкость);
• гомеостаз метаболических процессов в печени, почках, мышцах и других органах;
• гомеостаз газообмена в легких;
• гомеостаз кислотно-щелочного состояния крови;
• гомеостаз буферных свойств гемоглобина и т.д.
Многие из указанных видов гомеостаза имеют общие механизмы и тесно связаны друг с другом.
Именно объединение отдельных видов гомеостаза в целостную приспособительную систему организма в ответ на действие факторов внешней среды отражает системный характер организации различных форм молекулярной жизни (см. главы 7 и 8).
Такое объединение достигается за счет совместного функционирования основных регуляторных (и одновременно защитных)
систем организма: нервной, эндокринной, иммунной, ферментной (см. главы 13-15). Эти системы различаются по скорости и времени сохранения ответа на действие факторов внешней среды (раздражителей). Относительно быстрые реакции организма обеспечиваются нервной системой.
Гормональные и иммунные воздействия распространяются на клетки, ткани и органы гораздо медленнее, чем нервные импульсы, но зато и сохраняются более длительное время.
Наглядным примером генерализованного ответа на необычное по силе и продолжительности воздействие раздражителей служит стресс-реакция, обусловливающая быстрое привлечение энергетических ресурсов для адаптации организма к новым условиям.
При действии раздражителей сигналы из анализаторных отделов коры головного мозга поступают в гипоталамус, передающий их гипофизу, где резко возрастает синтез гормонов, и они выбрасываются в кровь. Включаются три основные «эндокринные оси»: адренокортикотропная, соматотропная и тиреоидная - они связаны с активностью надпочечников и щитовидной железы. Затем происходят соответствующие изменения в состоянии других систем. Общий итог - повышение уровня сопротивляемости организма.
Известны два способа генетического контроля гомеостаза.
Первый способ - контроль за элементарными реакциями непосредственно в клетке (например, пассивный транспорт веществ, синтез ферментов).
Второй способ - контроль за более сложными реакциями, протекающими на уровне органов и систем организма (например, регуляция функций на основе нормы реакции, регуляция иммунитета, мышечной и нервной активности, умственной деятельности и др.).
ГЕНОМ ЧЕЛОВЕКА И ЕГО РАЗМЕРЫ
Геном человека - полная генетическая система, ответственная за происхождение, развитие и наследование всех структурных и функциональных особенностей организма.
С молекулярных позиций геном человека - одинарное количество нуклеотидных последовательностей ядерной ДНК и мтДНК, распределенных по 25 хромосомам (22 аутосомы, Х-хромосома, Y-хромосома и хромосома, условно названная митохондриальной). Относительно всех 25 хромосом - это их полный гаплоидный набор.
Митохондриальная хромосома (в отличие от ДНК аутосом и гоносом) и ее мтДНК не связаны с белками, т.е. она существует в «голом виде» (см. выше).
В гаплоидном наборе хромосом молекулярная масса ДНК равна 3,6 χ 10-12 г. В диплоидном наборе она в 2 раза больше: 7,3 χ 10-12 г.
В настоящее время выделяют три параметра, характеризующие размеры генома при диплоидном наборе хромосом: физический, генетический и цитогенетический. У человека имеют место следующие параметры.
• Физический размер - 3,1467 χ 109, или 3,1467 млрд н.п. ДНК. В соматической клетке количество ДНК в 2 раза больше: 6,3 χ 109 н.п.
• Генетический размер - 3,0-3,5 тыс. сантиморганид [См] (общее количество генетических обменов между гомологичными хромосомами в ходе одного мейоза).
• Цитогенетический размер - 2,5-3,0 тыс. полос (бэндов), выявляемых на всех хромосомах при дифференциальном (прометафазном) окрашивании.
О размерах генома и генотипа человека судят исходя из следующего примера: если деспирализовать ДНК всех 25 хромосом и вытянуть каждую из них в отдельную ниточку, а затем состыковать все ниточки конец в конец, то общая длина хромосомной ДНК составит 120-130 см (суммарная длина всех нуклеотидных последовательностей ДНК). Но общая длина только экзонной части ДНК - всего 8-10 см (суммарная длина нуклеотидных последовательностей, входящих в экзонную часть ДНК).
Следует указать на отсутствие корреляции между размерами биологического вида и размерами его генома (для сравнения: у саламандры и лилии размер генома почти в 30 раз больше, чем у человека).
Кариотип, кариотип человека
Кариотип - полный набор хромосом соматической клетки конкретного биологического вида с учетом количества, формы, размера, структуры (при дифференциальном окрашивании) и других особенностей.
Кариотип человека - совокупность морфологических особенностей полного хромосомного набора ядра соматической клетки, свойственная виду Homo sapiens (для женского пола - 46,ХХ; для мужского пола - 46,XY, т.е. двойной, диплоидный набор хромосом, включающий 44 аутосомы и 2 половые хромосомы - гоносомы).
Популяция, популяция человека
Популяция - совокупность особей одного биологического вида (группы видов), постоянно находящихся (проживающих) на ограниченной географической территории.
Как сказано в главе 1, термин впервые ввел В. Иогансен в 1903 г. для обозначения неоднородной группы особей одного биологического вида. В 1926 г. С.С. Четвериков, изучавший разнообразие популяций, стал основателем нового научного направления - популяционной генетики; в ней наиболее важными факторами считаются естественный отбор и изоляция.
Популяция человека - совокупность всех людей Земли (примерно 6,5 млрд человек).
Генофонд человека и его свойства
Генофонд человека - совокупность всех генов (генотипов) в общей популяции человека как биологического вида (гены всех живущих на Земле людей).
Понятие «генофонд« впервые ввел А.С. Серебровский в 1928 г.
Основные свойства генофонда: генетическая целостность; глубокая дифференцированность (или неоднородность) генотипов при сохраняющейся общей совокупности генов и целостности генофонда; зависимость генофонда современного человека от генофонда его предков; наличие генетического груза (см. ниже).
Первые три свойства (особенно первое) обеспечивают сохранение (бессмертие) человека как вида Homo sapiens на основе непрерывного воспроизводства (репродуктивного процесса).
Репродуктивный процесс у человека
С генетической точки зрения, репродуктивный процесс у человека
есть воспроизводство новых генотипов в каждом последующем поколении на основе слияния двух родительских генотипов.
В ходе такого воспроизводства осуществляется постоянный обмен наследственным материалом между организмами женского и мужского пола, формируются новые генотипы их потомков, и каждое новое поколение потомков представлено генотипами, содержащими около 50% генов каждого родителя.
Репродуктивный процесс, осуществляемый в каждом поколении человека, вносит свой вклад (большой или малый) в генофонд чело-
века, зависящий от жизнеспособности и репродуктивной активности каждого потомка в окружающей его среде обитания.
Условия окружающей среды по-разному влияют на жизнеспособность и репродуктивную активность разных людей (разных генотипов). Поэтому генофонд человека (человечества) постепенно изменяется (совершенствуется) в результате естественного отбора, и более приспособленные люди (генотипы) чаще выживают и оставляют потомство, сохраняя, однако, генетическую целостность генофонда.
Таким образом, биологический смысл репродуктивного процесса у человека заключается в передаче из поколения в поколение и сохранении целостности генофонда.
Генетический груз у человека
Генетический груз как одно из свойств генофонда человека прямо связан с геномным, протеомным и репродуктивным здоровьем человека (см. главу 1). Еще в 1932 г. Г. Мёллер дал определение генетического груза как меньшей части общей популяции с измененной наследственностью. Сюда относят людей с наследственной патологией, развившейся в результате отрицательного действия факторов окружающей среды или переданной от предыдущих поколений.
Такие люди менее приспособлены к выживанию, часто подолгу болеют и избирательно гибнут в процессе естественного отбора. Продолжительность их жизни почти в 2 раза меньше, чем у здоровых людей.