Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. - 296 с. :ил.
|
|
|
|
МЫШЕЧНЫЕ ТКАНИ
Мышечные ткани представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака - выраженной сократительной способности, благодаря которой они могут выполнять свою основную функцию - перемещать тело или его части в пространстве.
Важнейшие свойства мышечных тканей. Структурные элементы мышечных тканей (клетки, волокна) обладают удлиненной формой и способны к сокращению благодаря мощному развитию сократительного аппарата. Для последнего характерно высокоупорядоченное расположение актиновых и миозиновых миофиламентов, создающее оптимальные условия для их взаимодействия. Это достигается связью сократимых структур с особыми элементами цитоскелета и плазмолеммой (сарколеммой), выполняющими опорную функцию. В части мышечных тканей миофиламенты образуют органеллы специального значения - миофибриллы. Для мышечного сокращения требуется значительное количество энергии, поэтому в структурных элементах мышечных тканей имеется большое количество митохондрий и трофических включений (липидных капель, гранул гликогена), содержащих субстраты - источники энергии. Поскольку мышечное сокращение протекает с участием ионов кальция, в мышечных клетках и волокнах хорошо развиты структуры, осуществляющие его накопление и выделение - агранулярная эндоплазматическая сеть (саркоплазматическая сеть), кавеолы.
Классификация мышечных тканей основана на признаках их (а) строения и функции (морфофункциональная классификация) и (б) происхождения (гистогенетическая классификация).
Морфофункциональная классификация мышечных тканей выделяет поперечнополосатые (исчерченные) мышечные ткани и гладкую мышечную ткань. Поперечнополосатые мышечные ткани образованы структурными элементами (клетками, волокнами), которые обладают поперечной исчерченностью вследствие особого упорядоченного взаиморасположения в них актиновых и миозиновых миофиламентов. К поперечнополосатым мышечным тканям относят скелетную и сердечную мышечную ткани. Гладкая мышечная ткань состоит из клеток, не обладающих поперечной исчерченностью. Наиболее распространенным видом этой ткани является гладкая мышечная ткань, входящая в состав стенки различных органов (бронхов, желудка, кишки, матки, маточной трубы, мочеточника, мочевого пузыря и сосудов).
Гистогенетическая классификация мышечных тканей выделяет три основных типа мышечных тканей: соматический (скелетная мышечная ткань), целомический (сердечная мышечная ткань) и мезенхимный (гладкая мышечная ткань внутренних органов), а также два дополнительных: миоэпителиальные клетки (видоизмененные эпителиальные сократимые клетки в концевых отделах и мелких выводных протоках некоторых желез) и мионейральные элементы (сократимые клетки нейрального происхождения в радужке глаза).
Скелетная поперечнополосатая (исчерченная) мышечная ткань по своей массе превышает любую другую ткань организма и является самой распространенной мышечной тканью тела человека. Она обеспечивает перемещение тела и его частей в пространстве и поддержание позы (входит в состав локомоторного аппарата), образует глазодвигательные мышцы, мышцы стенки полости рта, языка, глотки, гортани. Аналогичное строение имеет нескелетная висцеральная исчерченная мышечная ткань, которая обнаруживается в верхней трети пищевода, входит в состав наружных анального и уретрального сфинктеров.
Скелетная поперечнополосатая мышечная ткань развивается в эмбриональном периоде из миотомов сомитов, дающих начало активно делящимся миобластам - клеткам, которые располагаются цепочками и сливаются друг с другом в области концов с образованием мышечных трубочек (миотубул), превращающихся в мышечные волокна. Такие структуры, образованные единой гигантской цитоплазмой и многочисленными ядрами, в отечественной литературе традиционно именуют симпластами (в данном случае - миосимпластами), однако этот термин отсутствует в принятой международной терминологии. Некоторые миобласты не сливаются с другими, располагаясь на поверхности волокон и давая начало миосателлитоцитам - мелким клеткам, которые являются камбиальными элементами скелетной мышечной ткани. Скелетная мышечная ткань образована собранными в пучки поперечнополосатыми мышечными волокнами (рис. 87), являющимися ее структурно-функциональными единицами.
Мышечные волокна скелетной мышечной ткани представляют собой цилиндрические образования вариабельной длины (от миллиметров до 10-30 см). Их диаметр также широко варьирует в зависимости от принадлежности к определенной мышце и типу, функционального состояния, степени функциональной нагрузки, состояния питания
и других факторов. В мышцах мышечные волокна образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму, что особенно хорошо видно на поперечных срезах (см. рис. 87). Между мышечными волокнами располагаются тонкие прослойки рыхлой волокнистой соединительной ткани, несущие сосуды и нервы - эндомизий. Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных анизотропных дисков (полос А) и светлых изотропных дисков (полос I). Каждый изотропный диск рассекается надвое тонкой темной линией Z - телофрагмой (рис. 88). Ядра мышечного волокна - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные - лежат на его периферии под сарколеммой и располагаются вдоль волокна. Снаружи сарколемма покрыта толстой базальной мембраной, в которую вплетаются ретикулярные волокна.
Миосателлитоциты (клетки-миосателлиты) - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы мышечного волокна и покрытые общей базальной мембраной (см. рис. 88). Ядро миосателлитоцита - плотное, относительно крупное, органеллы мелкие и немногочисленные. Эти клетки активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Сливаясь с остальной частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии.
Миофибриллы образуют сократительный аппарат мышечного волокна, располагаются в саркоплазме по ее длине, занимая центральную часть, и отчетливо выявляются на поперечных срезах волокон в виде мелких точек (см. рис. 87 и 88).
Миофибриллы обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядоченно, что изотропные и анизотропные диски разных миофибрилл совпадают между собой, обусловливая поперечную исчерченность всего волокна. Каждая миофибрилла образована тысячами повторяющихся последовательно связанных между собой структур - саркомеров.
Саркомер (миомер) является структурно-функциональной единицей миофибриллы и представляет собой ее участок, расположенный между двумя телофрагмами (линиями Z). Он включает анизотропный диск и две половины изотропных дисков - по одной половине с каждой стороны (рис. 89). Саркомер образован упорядоченной системой толстых (миозиновых) и тонких (актиновьх) миофиламентов. Толстые миофиламенты связаны с мезофрагмой (линией М) и сосредоточены в анизотропном диске,
а тонкие миофиламенты прикреплены к телофрагмам (линиям Z), образуют изотропные диски и частично проникают в анизотропный диск между толстыми нитями вплоть до светлой полосы Н в центре анизотропного диска.
Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что в результате взаимодействия актина и миозина в присутствии кальция и АТФ тонкие нити вдвигаются в промежутки между толстыми без изменения их длины. При этом ширина анизотропных дисков не меняется, а ширина изотропных дисков и полос Н - уменьшается. Строгая пространственная упорядоченность взаимодействия множества толстых и тонких миофиламентов в саркомере определяется наличием сложно организованного поддерживающего аппарата, к которому, в частности, относятся телофрагма и мезофрагма. Кальций выделяется из саркоплазматической сети, элементы которой оплетают каждую миофибриллу, после поступления сигнала с сарколеммы по Т-трубочкам (совокупность этих элементов описывается как саркотубулярная система).
Скелетная мышца как орган состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов (рис. 90). Снаружи мышцу покрывает эпимизий - тонкий, прочный и гладкий чехол из плотной волокнистой соединительной ткани, отдающий вглубь органа более тонкие соединительнотканные перегородки - перимизий, который окружает пучки мышечных волокон. От перимизия внутрь пучков мышечных волокон отходят тончайшие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное волокно - эндомизий.
Типы мышечных волокон в скелетной мышце - разновидности мышечных волокон с определенными структурными, биохимическими и функцио нальными различиями. Типирование мышечных волокон производится на препаратах при постановке гистохимических реакций выявления ферментов - например, АТФазы, лактатдегидрогеназы (ЛДГ), сукцинатдегидрогеназы (СДГ) (рис. 91) и др. В обобщенном виде можно условно выделить три основных типа мышечных волокон, между которыми существуют переходные варианты.
Тип I (красные) - медленные, тонические, устойчивые к утомлению, с небольшой силой сокращения, окислительные. Характеризуются малым диаметром, относительно тонкими миофибриллами,
высокой активностью окислительных ферментов (например, СДГ), низкой активностью гликолитических ферментов и миозиновой АТФазы, преобладанием аэробных процессов, высоким содержанием пигмента миоглобина (определяющим их красный цвет), крупных митохондрий и липидных включений, богатым кровоснабжением. Численно преобладают в мышцах, выполняющих длительные тонические нагрузки.
Тип IIВ (белые) - быстрые, тетанические, легко утомляющиеся, с большой силой сокращения, гликолитические. Характеризуются большим диаметром, крупными и сильными миофибриллами, высокой активностью гликолитических ферментов (например, ЛДГ) и АТФазы, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, относительно низким содержанием мелких митохондрий, липидов и миоглобина (определяющим их светлый цвет), значительным количеством гликогена, сравнительно слабым кровоснабжением. Преобладают в мышцах, выполняющих быстрые движения, например, мышцах конечностей.
Тип IIА (промежуточные) - быстрые, устойчивые к утомлению, с большой силой, оксилительно-гликолитические. На препаратах напоминают волокна типа I. В равной степени способны использовать энергию, получаемую путем окислительных и гликолитических реакций. По своим морфологическим и функциональным характеристикам занимают положение, промежуточное между волокнами типа I и IIB.
Скелетные мышцы человека являются смешанными, т. е. содержат волокна различных типов, которые распределены в них мозаично (см. рис. 91).
Сердечная поперечнополосатая (исчерченная) мышечная ткань встречается в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Основным функциональным свойством сердечной мышечной ткани служит способность к спонтанным ритмическим сокращениям, на активность которых влияют гормоны и нервная система. Эта ткань обеспечивает сокращения сердца, которые поддерживают циркуляцию крови в организме. Источником развития сердечной мышечной ткани служит миоэпикардиальная пластинка висцерального листка спланхнотома (целомическая выстилка в шейной части эмбриона). Клетки этой пластинки (миобласты) активно размножаются и постепенно превращаются в сердечные мышечные клетки - кардиомиоциты (сердечные миоциты). Выстраиваясь в цепочки, кардиомиоциты формируют сложные межклеточные соединения - вставочные диски, связывающие их в сердечные мышечные волокна.
Зрелая сердечная мышечная ткань образована клетками - кардиомиоцитами, связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих сердечных мышечных волокон (рис. 92).
Кардиомиоциты (сердечные миоциты) - цилиндрические или ветвящиеся клетки, более крупные в желудочках. В предсердиях они обычно имеют неправильную форму и меньшие размеры. Эти клетки содержат одно или два ядра и саркоплазму, покрыты сарколеммой, которая снаружи окружена базальной мембраной. Их ядра - светлые, с преобладанием эухроматина, хорошо заметными ядрышками - занимают в клетке центральное положение. У взрослого человека значительная часть кардиомиоцитов - полиплоидные, более половины - двуядерные. Саркоплазма кардиомиоцитов содержит многочисленные органеллы и включения, в частности, мощный сократительный аппарат, который сильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых). Сократительный аппарат представлен сердечными исчерченными миофибриллами, по строению сходными с миофибриллами волокон скелетной мышечной ткани (см. рис. 94); в совокупности они обусловливают поперечную исчерченность кардиомиоцитов.
Между миофибриллами у полюсов ядра и под сарколеммой располагаются очень многочисленные и крупные митохондрии (см. рис. 93 и 94). Миофибриллы окружены элементами саркоплазматической сети, связанными с Т-трубочками (см. рис. 94). Цитоплазма кардиомиоцитов содержит кислородсвязывающий пигмент миоглобина и скопления энергетических субстратов в виде липидных капель и гранул гликогена (см. рис. 94).
Типы кардиомиоцитов в сердечной мышечной ткани различаются структурными и функциональными признаками, биологической ролью и топографией. Выделяют три основных типа кардиомиоцитов (см. рис. 93):
1) сократительные (рабочие) кардиомиоциты образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим бjльшую часть их саркоплазмы;
2) проводящие кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они образуют узлы, пучки и волокна проводящей системы сердца и разделяются на несколько подтипов. Характеризуются слабым развитием сократительного аппарата, светлой саркоплазмой и крупными ядрами. В проводящих сердечных волокнах (Пуркинье) эти клетки имеют крупные размеры (см. рис. 93).
3) секреторные (эндокринные) кардиомиоциты располагаются в предсердиях (в особенности, пра-
вом) и характеризуются отростчатой формой и слабым развитием сократительного аппарата. В их саркоплазме вблизи полюсов ядра находятся окруженные мембраной плотные гранулы, содержащие предсердный натриуретический пептид (гормон, вызывающий потерю натрия и воды с мочой, расширение сосудов, снижение артериального давления).
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих сердечное мышечное волокно (см. рис. 92). Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов (см. рис. 94). В области поперечных (ориентированных перпендикулярно расположению миофибрилл) участков вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом и адгезивных фасций. Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска на уровне линии Z. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения (нексусы), обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения.
Гладкая мышечная ткань входит в состав стенки полых (трубчатых) внутренних органов - бронхов, желудка, кишки, матки, маточных труб, мочеточников, мочевого пузыря (висцеральная гладкая мышечная ткань), а также сосудов (васкулярная гладкая мышечная ткань). Гладкая мышечная ткань встречается также в коже, где она образует мышцы, поднимающие волос, в капсулах и трабекулах некоторых органов (селезенка, яичко). Благодаря сократительной активности этой ткани обеспечивается деятельность органов пищеварительного тракта, регуляция дыхания, крово- и лимфотока, выделение мочи, транспорт половых клеток и др. Источником развития гладкой мышечной ткани у эмбриона является мезенхима. Свойствами гладких миоцитов обладают также некоторые клетки, имеющие другое происхождение - миоэпителиальные клетки (видоизмененные сократительные эпителиальные клетки в некоторых железах) и мионейральные клетки радужки глаза (развиваются из нейрального зачатка). Структурно-функциональной единицей гладкой мышечной ткани служит гладкий миоцит (гладкая мышечная клетка).
Гладкие миоциты (гладкие мышечные клетки) - вытянутые клетки преимущественно вере-
теновидной формы, не обладающие поперечной исчерченностью и образующие многочисленные соединения друг с другом (рис. 95-97). Сарколемма каждого гладкого миоцита окружена базальной мембраной, в которую вплетаются тонкие ретикулярные, коллагеновые и эластические волокна. Гладкие миоциты содержат одно удлиненное диплоидное ядро с преобладанием эухроматина и 1-2 ядрышками, расположенное в центральной утолщенной части клетки. В саркоплазме гладких миоцитов умеренно развитые органеллы общего значения располагаются вместе с включениями в конусовидных участках у полюсов ядра. Периферическая ее часть занята сократительным аппаратом - актиновыми и миозиновыми миофиламентами, которые в гладких миоцитах не формируют миофибрилл. Актиновые миофиламенты прикрепляются в саркоплазме к овальным или веретеновидным плотным тельцам (см. рис. 97) - структурам, гомологичным линиям Z в поперечнополосатых тканях; сходные образования, связанные с внутренней поверхностью сарколеммы, называют плотными пластинками.
Сокращение гладких миоцитов обеспечивается взаимодействием миофиламентов и развивается в соответствии с моделью скользящих нитей. Как и в поперечнополосатых мышечных тканях, сокращение гладких миоцитов индуцируется притоком Са2+ в саркоплазму, который в этих клетках выделяется саркоплазматической сетью и кавеолами - многочисленными колбовидными впячиваниями поверхности сарколеммы. Благодаря выраженной синтетической активности гладкие миоциты продуцируют и выделяют (подобно фибробластам) коллагены, эластин и компоненты аморфного вещества. Они способны также синтезировать и секретировать ряд факторов роста и цитокинов.
Гладкая мышечная ткань в органах обычно представлена пластами, пучками и слоями гладких миоцитов (см. рис. 95), внутри которых клетки связаны интердигитациями, адгезивными и щелевыми соединениями. Расположение гладких миоцитов в пластах таково, что узкая часть одной клетки прилежит к широкой части другой. Это способствует наиболее компактной укладке миоцитов, обеспечению максимальной площади их взаимных контактов и высокой прочности ткани. В связи с описанным расположением гладких мышечных клеток в пласте на поперечных срезах соседствуют сечения миоцитов, разрезанных в широкой части и в области узкого края (см. рис. 95).
МЫШЕЧНЫЕ ТКАНИ
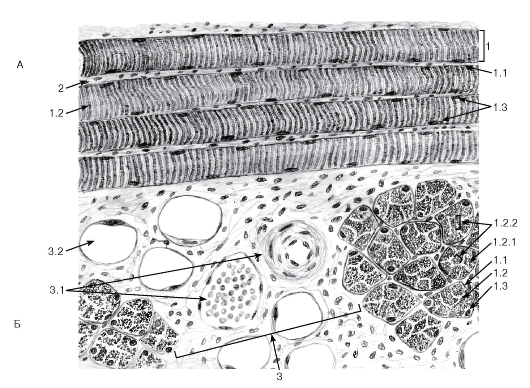
Рис. 87. Скелетная поперечнополосатая мышечная ткань
Окраска: железный гематоксилин
А - продольный срез; Б - поперечный срез:
1 - мышечное волокно: 1.1 - сарколемма, покрытая базальной мембраной, 1.2 - саркоплазма, 1.2.1 - миофибриллы, 1.2.2 - поля миофибрилл (Конгейма); 1.3 - ядра мышечного волокна; 2 - эндомизий; 3 - прослойки рыхлой волокнистой соединительной ткани между пучками мышечных волокон: 3.1 - кровеносные сосуды, 3.2 - жировые клетки
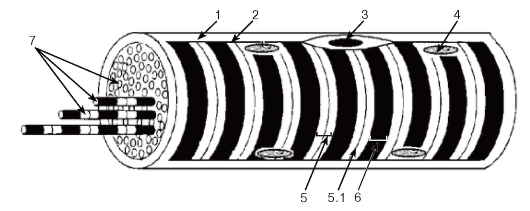
Рис. 88. Скелетное мышечное волокно (схема):
1 - базальная мембрана; 2 - сарколемма; 3 - миосателлитоцит; 4 - ядро миосимпласта; 5 - изотропный диск: 5.1 - телофрагма; 6 - анизотропный диск; 7 - миофибриллы

Рис. 89. Участок миофибриллы волокна скелетной мышечной ткани (саркомер)
Рисунок с ЭМФ
1 - изотропный диск: 1.1 - тонкие (актиновые) миофиламенты, 1.2 - телофрагма; 2 - анизотропный диск: 2.1 - толстые (миозиновые) миофиламенты, 2.2 - мезофрагма, 2.3 - полоса Н; 3 - саркомер
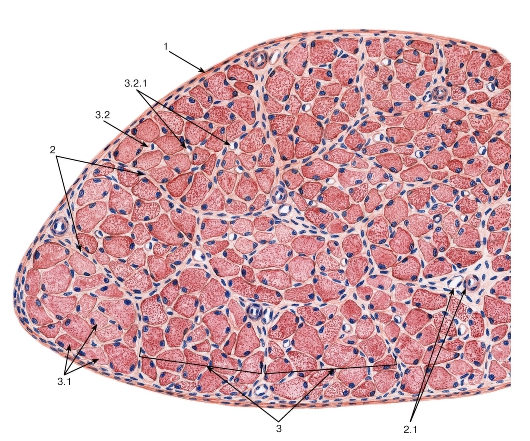
Рис. 90. Скелетная мышца (поперечный срез)
Окраска: гематоксилин-эозин
1 - эпимизий; 2 - перимизий: 2.1 - кровеносные сосуды; 3 - пучки мышечных волокон: 3.1 - мышечные волокна, 3.2 - эндомизий: 3.2.1 - кровеносные сосуды
Рис. 91. Типы мышечных волокон (поперечный срез скелетной мышцы)
Гистохимическая реакция выявления сукцинатдегидрогеназы (СДГ)
1 - волокна I типа (красные волокна) - с высокой активностью СДГ (медленные, окислительные, устойчивые к утомлению); 2 - волокна IIВ типа (белые волокна) - с низкой активностью СДГ (быстрые, гликолитические, утомляемые); 3 - волокна IIА типа (промежуточные волокна) - с умеренной активностью СДГ (быстрые, окислительно-гликолитические, устойчивые к утомлению)
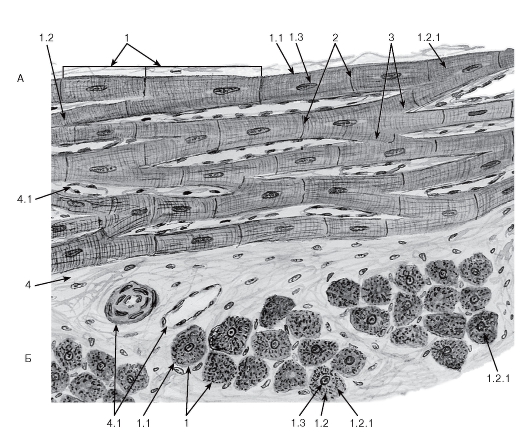
Рис. 92. Сердечная поперечнополосатая мышечная ткань
Окраска: железный гематоксилин
А - продольный срез; Б - поперечный срез:
1 - кардиомиоциты (образуют сердечные мышечные волокна): 1.1 - сарколемма, 1.2 - саркоплазма, 1.2.1 - миофибриллы, 1.3 - ядро; 2 - вставочные диски; 3 - анастомозы между волокнами; 4 - рыхлая волокнистая соединительная ткань: 4.1 - кровеносные сосуды
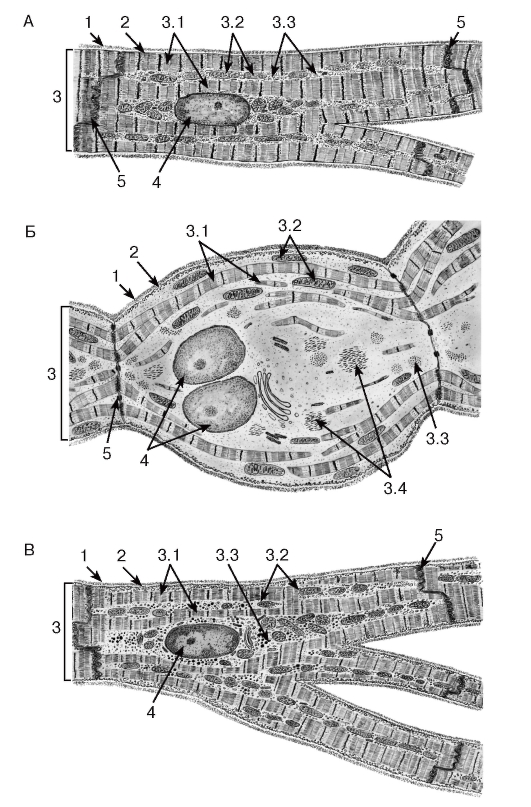
Рис. 93. Ультраструктурная организация кардиомиоцитов различных типов
Рисунки с ЭМФ
A - сократительный (рабочий) кардиомиоцит желудочка сердца:
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии, 3.3 - липидные капли; 4 - ядро; 5 - вставочный диск.
Б - кардиомиоцит проводящей системы сердца (из субэндокардиальной сети волокон Пуркинье):
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии; 3.3 - гранулы гликогена, 3.4 - промежуточные филаменты; 4 - ядра; 5 - вставочный диск.
В - эндокринный кардиомиоцит из предсердия:
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии, 3.3 - секреторные гранулы; 4 - ядро; 5 - вставочный диск
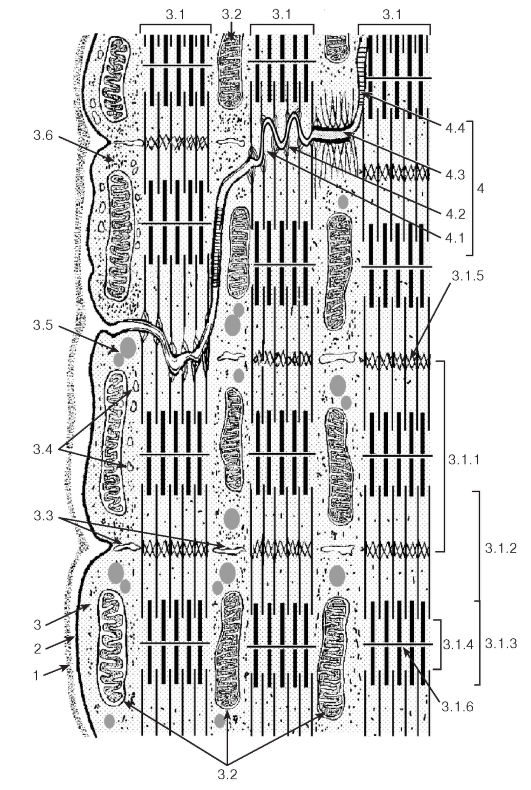
Рис. 94. Ультраструктурная организация области вставочного диска между соседними кардиомиоцитами
Рисунок с ЭМФ
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.1.1 - саркомер, 3.1.2 - изотропный диск, 3.1.3 - анизотропный диск, 3.1.4 - светлая полоса Н, 3.1.5 - телофрагма, 3.1.6 - мезофрагма, 3.2 - митохондрии, 3.3 - Т-трубочки, 3.4 - элементы саркоплазматической сети, 3.5 - липидные капли, 3.6 - гранулы гликогена; 4 - вставочный диск: 4.1 - интердигитации, 4.2 - адгезивная фасция, 4.3 - десмосома, 4.4 - щелевое соединение (нексус)
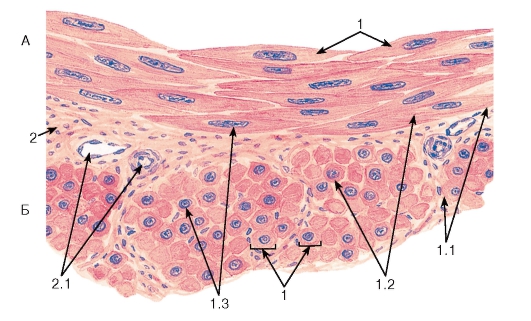
Рис. 95. Гладкая мышечная ткань
Окраска: гематоксилин-эозин
А - продольный срез; Б - поперечный срез:
1 - гладкие миоциты: 1.1 - сарколемма, 1.2 - саркоплазма, 1.3 - ядро; 2 - прослойки рыхлой волокнистой соединительной ткани между пучками гладких миоцитов: 2.1 - кровеносные сосуды
Рис. 96. Изолированные гладкие мышечные клетки
Окраска: гематоксилин
1 - ядро; 2 - саркоплазма; 3 - сарколемма

Рис. 97. Ультраструктурная организация гладкого миоцита (участок клетки)
Рисунок с ЭМФ
1 - сарколемма; 2 - саркоплазма: 2.1 - митохондрии, 2.2 - плотные тельца; 3 - ядро; 4 - базальная мембрана