Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. - 296 с. :ил.
|
|
|
|
ЧАСТНАЯ ГИСТОЛОГИЯ. ОРГАНЫ НЕРВНОЙ СИСТЕМЫ
Нервная система осуществляет объединение частей организма в единое целое (интеграцию), обеспечивает регуляцию разнообразных процессов, координацию функции различных органов и тканей и взаимодействие организма с внешней средой. Она воспринимает многообразную информацию, поступающую из внешней среды и из внутренних органов, перерабатывает ее и генерирует сигналы, обеспечивающие ответные реакции, адекватные действующим раздражителям. В основе деятельности нервной системы лежат рефлекторные дуги - цепочки нейронов, которые обеспечивают реакции рабочих органов (органов-мишеней) в ответ на раздражение рецепторов. В рефлекторных дугах нейроны, связанные друг с другом синапсами, образуют три звена: рецепторное (афферентное), эффекторное и расположенное между ними ассоциативное (вставочное).
Отделы нервной системы
Анатомическое подразделение отделов нервной системы:
(1) центральная нервная система (ЦНС) -
включает головной и спинной мозг;
(2) периферическая нервная система - включает периферические нервные ганглии (узлы), нервы и нервные окончания (описаны в разделе «Нервная ткань»).
Физиологическое подразделение отделов нервной системы (в зависимости от характера иннервации органов и тканей):
(1) соматическая (анимальная) нервная система - контролирует преимущественно функции произвольного движения;
(2) автономная (вегетативная) нервная система - регулирует деятельность внутренних органов, сосудов и желез.
Автономная нервная система подразделяется на взаимодействующие друг с другом симпатический и парасимпатический отделы, которые различаются локализацией периферических узлов и центров в мозгу, а также характером влияния на внутренние органы.
В соматическую и автономную нервную систему входят звенья, расположенные в ЦНС и периферической нервной системе. Функционально ведущей тканью органов нервной системы является нервная ткань, включающая нейроны и глию. Скопления нейронов в ЦНС обычно называют ядрами, а в периферической нервной системе - ганглиями (узлами). Пучки нервных волокон в центральной нервной системе носят названия трактов, в периферической - нервов.
Органы периферической нервной системы
Нервы (нервные стволы) связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами. Они образованы пучками миелиновых и безмиелиновых нервных волокон, которые объединены соединительнотканными компонентами (оболочками): эндоневрием, периневрием и эпиневрием (рис. 114-118). Большинство нервов являются смешанными, т. е. включают афферентные и эфферентные нервные волокна.
Эндоневрий - тонкие прослойки рыхлой волокнистой соединительной ткани с мелкими кровеносными сосудами, окружающие отдельные нервные волокна и связывающие их в единый пучок.
Периневрий - оболочка, покрывающая каждый пучок нервных волокон снаружи и отдающая перегородки вглубь пучка. Он имеет пластинчатое строение и образован концентрическими пластами уплощенных фибробластоподобных клеток, связанных плотными и щелевыми соединениями. Между слоями клеток в пространствах, заполненных жидкостью, располагаются компоненты базальной мембраны и продольно ориентированные коллагеновые волокна.
Эпиневрий - наружная оболочка нерва, связывающая воедино пучки нервных волокон. Он состоит из плотной волокнистой соединительной ткани, содержащей жировые клетки, кровеносные и лимфатические сосуды (см. рис. 114).
Структуры нерва, выявляемые с помощью различных методов окраски. Различные гистологические методы окраски позволяют более детально и избирательно изучить отдельные компоненты
нерва. Так, осмирование дает контрастное окрашивание миелиновых оболочек нервных волокон (позволяя оценить их толщину и дифференцировать миелиновые и безмиелиновые волокна), однако отростки нейронов и соединительнотканные компоненты нерва остаются очень слабо окрашенными или неокрашенными (см. рис. 114 и 115). При окраске гематоксилином-эозином миелиновые оболочки не окрашиваются, отростки нейронов имеют слабо базофильное окрашивание, однако хорошо выявляются ядра нейролеммоцитов в нервных волокнах и все соединительнотканные компоненты нерва (см. рис. 116 и 117). При окраске азотнокислым серебром ярко окрашиваются отростки нейронов; миелиновые оболочки остаются неокрашенными, соединительнотканные компоненты нерва выявляются слабо, их структура не прослеживается (см. рис. 118).
Нервные ганглии (узлы) - структуры, образованные скоплениями нейронов вне ЦНС, - разделяются на чувствительные и автономные (вегетативные). Чувствительные ганглии содержат псевдоуниполярные или биполярные (в спиральном и вестибулярном ганглиях) афферентные нейроны и располагаются преимущественно по ходу задних корешков спинного мозга (чувствительные узлы спинномозговых нервов) и некоторых черепно-мозговых нервов.
Чувствительные ганглии (узлы) спинномозговых нервов имеют веретеновидную форму и покрыты капсулой из плотной волокнистой соединительной ткани. По периферии ганглия находятся плотные скопления тел псевдоуниполярных нейронов, а центральная часть занята их отростками и расположенными между ними тонкими прослойками эндоневрия, несущими сосуды (рис. 121).
Псевдоуниполярные чувствительные нейроны характеризуются сферическим телом и светлым ядром с хорошо заметным ядрышком (рис. 122). Цитоплазма нейронов содержит многочисленные митохондрии, цистерны гранулярной эндоплазматической сети, элементы комплекса Гольджи (см. рис. 101), лизосомы. Каждый нейрон окружен слоем прилежащих к нему уплощенных клеток олигодендроглии (сателлитными глиальными клетками, или мантийными глиоцитами) с мелкими округлыми ядрами; снаружи глиальной оболочки имеется тонкая соединительнотканная капсула (см. рис. 122). От тела псевдоуниполярного нейрона отходит отросток, разделяющийся Т-образно на периферическую (афферентную, дендритную) и центральную (эфферентную, аксональную) ветви, которые покрываются миелиновыми оболочками. Периферический отросток (афферентная ветвь) заканчивается рецепторами,
центральный отросток (эфферентная ветвь) в составе заднего корешка вступает в спинной мозг (см. рис. 119).
Автономные нервные ганглии образованы скоплениями мультиполярных нейронов, на которых многочисленные синапсы образуют преганглионарные волокна - отростки нейронов, чьи тела лежат в ЦНС (см. рис. 120).
Классификация автономных ганглиев. По локализации: ганглии могут располагаться вдоль позвоночника (паравертебральные ганглии) или впереди него (превертебральные ганглии), а также в стенке органов - сердца, бронхов, пищеварительного тракта, мочевого пузыря и др. (интрамуральные ганглии - см., например, рис. 203, 209, 213, 215) или вблизи их поверхности.
По функциональному признаку автономные нервные ганглии разделяются на симпатические и парасимпатические. Эти ганглии различаются своей локализацией (симпатические лежат пара- и превертебрально, парасимпатические - интрамурально или вблизи органов), а также локализацией нейронов, дающих преганглионарные волокна, характером нейромедиаторов и направленностью реакций, опосредуемых их клетками. Большинство внутренних органов имеют двойную автономную иннервацию. Общий план строения симпатических и парасимпатических нерв ных ганглиев сходен.
Строение автономных ганглиев. Автономный ганглий снаружи покрыт соединительнотканной капсулой и содержит диффузно или группами расположенные тела мультиполярных нейронов, их отростки в виде безмиелиновых или (реже) миелиновых волокон и эндоневрий (рис. 123). Тела нейронов - базофильные, неправильной формы, содержат эксцентрично расположенное ядро; встречаются многоядерные и полиплоидные клетки. Нейроны окружены (обычно не полностью) оболочками из глиальных клеток (сателлитными глиальными клетками, или мантийными глиоцитами). Снаружи от глиальной оболочки располагается тонкая соединительнотканная оболочка (рис. 124).
Интрамуральные ганглии и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена некоторыми авторами выделяются в самостоятельный метасимпатический отдел автономной нервной системы. В интрамуральных ганглиях описаны нейроны трех типов (см. рис. 120):
1) Длинноаксонные эфферентные нейроны (клетки I типа Догеля) с короткими дендритами и длинным аксоном, идущим за пределы узла
к клеткам рабочего органа, на которых он образует двигательные или секреторные окончания.
2) Равноотростчатые афферентные нейроны (клетки II типа Догеля) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в центральную нервную систему.
3) Ассоциативные клетки (клетки III типа Догеля) - местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типов. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
Рефлекторные дуги в соматическом (анимальном) и автономном (вегетативном) отделах нервной системы обладают рядом особенностей (см. рис. 119 и 120). Основные различия заключаются в ассоциативном и эффекторном звеньях, поскольку рецепторное звено сходно: оно образовано афферентными псевдоуниполярными нейронами, тела которых располагаются в чувствительных ганглиях. Периферические отростки этих клеток образуют чувствительные нервные окончания, а центральные вступают в спинной мозг в составе задних корешков.
Ассоциативное звено в соматической дуге представлено вставочными нейронами, дендриты и тела которых расположены в задних рогах спинного мозга, а аксоны направляются в передние рога, передавая импульсы на тела и дендриты эфферентных нейронов. В автономной дуге дендриты и тела вставочных нейронов расположены в боковых рогах спинного мозга, а аксоны (преганглионарные волокна) покидают спинной мозг в составе передних корешков, направляясь в один из автономных ганглиев, где и оканчиваются на дендритах и телах эфферентных нейронов.
Эффекторное звено в соматической дуге образовано мультиполярными мотонейронами, тела и дендриты которых лежат в передних рогах спинного мозга, а аксоны выходят из спинного мозга в составе передних корешков, направляются к чувствительному ганглию и далее в составе смешанного нерва - к скелетной мышце, на волокнах которой их веточки образуют нейро-мышечные синапсы. В автономной дуге эффекторное звено образовано мультиполярными нейронами, тела которых лежат в составе автономных ганглиев, а аксоны (постганглионарные волокна) в составе нервных стволов и их ветвей направляются к клеткам рабочих органов - гладких мышц, желез, сердца.
Органы центральной нервной системы Спинной мозг
Спинной мозг имеет вид округлого тяжа, расширенного в шейном и пояснично-крестцовом отделах и пронизанного центральным каналом. Он состоит из двух симметричных половин, разделенных спереди передней срединной щелью, сзади - задней срединной бороздой, и характеризуется сегментарным строением; с каждым сегментом связана пара передних (двигательных, вентральных) и пара задних (чувствительных, дорсальных) корешков. В спинном мозгу различают серое вещество, расположенное в его центральной части, и белое вещество, лежащее по периферии (рис. 125).
Серое вещество на поперечном разрезе имеет вид бабочки (см. рис. 125) и включает парные передние (вентральные), задние (дорсальные) и боковые (латеральные) рога. Рога серого вещества обеих симметричных частей спинного мозга связаны друг с другом в области передней и задней серой спайки. В сером веществе находятся тела, дендриты и (частично) аксоны нейронов, а также глиальные клетки. Между телами нейронов находится нейропиль - сеть, образованная нервными волокнами и отростками глиальных клеток. Нейроны располагаются в сером веществе в виде не всегда резко разграниченных скоплений (ядер).
Задние рога содержат несколько ядер, образованных мультиполярными вставочными нейронами, на которых оканчиваются аксоны псевдоуниполярных клеток чувствительных ганглиев (см. рис. 119), а также волокна нисходящих путей из лежащих выше (супраспинальных) центров. Аксоны вставочных нейронов а) оканчиваются в сером веществе спинного мозга на мотонейронах, лежащих в передних рогах (см. рис. 119); б) образуют межсегментарные связи в пределах серого вещества спинного мозга; в) выходят в белое вещество спинного мозга, где образуют восходящие и нисходящие проводящие пути (тракты).
Боковые рога, хорошо выраженные на уровне грудных и крестцовых сегментов спинного мозга, содержат ядра, образованные телами мультиполярных вставочных нейронов, которые относятся к симпатическому и парасимпатическому отделам автономной нервной системы (см. рис. 120). На дендритах и телах этих клеток оканчиваются аксоны: а) псевдоуниполярных нейронов, несущих импульсы от рецепторов, расположенных во внутренних органах, б) нейронов центров регуляции вегетативных функций, тела которых находятся в продолговатом мозгу. Аксоны автономных нейронов, выходя из спинного мозга в составе передних корешков, образуют преган-
глионарные волокна, направляющиеся к симпатическим и парасимпатическим узлам.
Передние рога содержат мультиполярные двигательные нейроны (мотонейроны), объединенные в ядра, каждое из которых обычно тянется на несколько сегментов. Различают крупные α-мотонейроны и рассеянные среди них более мелкие γ-мотонейроны. На отростках и телах мотонейронов имеются многочисленные синапсы, оказывающие на них возбуждающие и тормозные воздействия. На мотонейронах оканчиваются: коллатерали центральных отростков псевдоуниполярных клеток чувствительных узлов; вставочных нейронов, тела которых лежат в задних рогах спинного мозга; аксоны местных мелких вставочных нейронов (клеток Реншоу), связанных с коллатералями аксонов мотонейронов; волокна нисходящих путей пирамидной и экстрапирамидной систем, несущие импульсы из коры большого мозга и ядер ствола мозга. Тела мотонейронов содержат крупные глыбки хроматофильного вещества (см. рис. 100) и окружены глиоцитами (рис. 126). Аксоны мотонейронов покидают спинной мозг в составе передних корешков, направляются к чувствительному ганглию и далее в составе смешанного нерва - к скелетной мышце, на волокнах которой они образуют нейро-мышечные синапсы (см. рис. 119).
Центральный канал (см. рис. 128) проходит в центре серого вещества и окружен передней и задней серыми спайками (см. рис. 125). Он заполнен спинномозговой жидкостью и выстлан одним слоем кубических или столбчатых клеток эпендимы, апикальная поверхность которых покрыта микроворсинками и (частично) ресничками, а латеральные связаны комплексами межклеточных соединений.
Белое вещество спинного мозга окружает серое (см. рис. 125) и разделяется передними и задними корешками на симметричные задние, боковые и передние канатики. Оно состоит из продольно идущих нервных волокон (преимущественно миелиновых), образующих нисходящие и восходящие проводящие пути (тракты). Последние отделены друг от друга тонкими прослойками соединительной ткани и астроцитов, которые встречаются и внутри трактов (рис. 127). Проводящие пути включают две группы: проприоспинальные (осуществляют связь между различными отделами спинного мозга) и супраспинальные пути (обеспечивают связь спинного мозга со структурами головного мозга - восходящие и нисходящие тракты).
Мозжечок
Мозжечок является частью головного мозга и представляет собой центр равновесия, поддер-
жания мышечного тонуса и координации движений. Он образован двумя полушариями с большим числом бороздок и извилин на поверхности и узкой средней частью (червем). Серое вещество образует кору мозжечка и ядра; последние залегают в глубине его белого вещества.
Кора мозжечка характеризуется высокой упорядоченностью расположения нейронов, нервных волокон и глиальных клеток всех типов. Она отличается богатством межнейронных связей, которая обеспечивают переработку поступающей в нее разнообразной сенсорной информации. В коре мозжечка различают три слоя (снаружи внутрь): 1) молекулярный слой; 2) слой клеток Пуркинье (слой грушевидных нейронов); 3) зернистый слой (рис. 129 и 130).
Молекулярный слой содержит сравнительно небольшое количество мелких клеток, в нем находятся тела корзинчатых и звездчатых нейронов. Корзинчатые нейроны располагаются во внутренней части молекулярного слоя. Их короткие дендриты образуют связи с параллельными волокнами в наружной части молекулярного слоя, а длинный аксон идет поперек извилины, отдавая через определенные интервалы коллатерали, которые спускаются к телам клеток Пуркинье и, разветвляясь, охватывают их наподобие корзинок, образуя тормозные аксо-соматические синапсы (см. рис. 130). Звездчатые нейроны - мелкие клетки, тела которых лежат выше тел корзинчатых нейронов. Их дендриты образуют связи с параллельными волокнами, а разветвления аксона формируют тормозные синапсы на дендритах клеток Пуркинье и могут участвовать в образовании корзинки вокруг их тел.
Слой клеток Пуркинье (слой грушевидных нейронов) содержит лежащие в один ряд тела клеток Пуркинье, оплетенные коллатералями аксонов корзинчатых клеток («корзинками»).
Клетки Пуркинье (грушевидные нейроны) - крупные клетки с телом грушевидной формы, содержащим хорошо развитые органеллы. От него в молекулярный слой отходят 2-3 первичных (стволовых) дендрита, интенсивно ветвящихся с образованием конечных (терминальных) дендритов, достигающих поверхности молекулярного слоя (см. рис. 130). На дендритах находятся многочисленные шипики - контактные зоны возбуждающих синапсов, образуемых параллельными волокнами (аксонами зернистых нейронов), и тормозных синапсов, образуемых лазящими волокнами. Аксон клетки Пуркинье отходит от основания ее тела, покрывается миелиновой оболочкой, пронизывает зернистый слой и проникает в белое вещество, являясь единственным эфферентным путем его коры.
Зернистый слой содержит близко расположенные тела зернистых нейронов, больших зведчатых нейронов (клеток Гольджи), а также клубочки мозжечка - особые округлые сложные синаптические контактные зоны между моховидными волокнами, дендритами зернистых нейронов и аксонами больших зведчатых нейронов.
Зернистые нейроны - наиболее многочисленные нейроны коры мозжечка - мелкие клетки с короткими дендритами, имеющими вид «птичьей лапки», на которых в клубочках мозжечка розетки моховидных волокон образуют многочисленные синаптические контакты. Аксоны зернистых нейронов направляются в молекулярный слой, где Т-образно делятся на две ветви, идущие параллельно длине извилины (параллельные волокна) и образующие возбуждающие синапсы на дендритах клеток Пуркинье, корзинчатых и звездчатых нейронов, а также больших звездчатых нейронов.
Большие звездчатые нейроны (клетки Гольджи) крупнее зернистых нейронов. Их аксоны в пределах клубочков мозжечка образуют тормозные синапсы на дендритах зернистых нейронов, а длинные дендриты поднимаются в молекулярный слой, где ветвятся и образуют связи с параллельными волокнами.
Афферентные волокна коры мозжечка включают моховидные и лазящие волокна (см. рис. 130), которые проникают в кору мозжечка из спинного мозга, продолговатого мозга и моста.
Моховидные волокна мозжечка заканчиваются расширениями (розетками) - клубочках мозжечка, образуя синаптические контакты с дендритами зернистых нейронов, на которых оканчиваются также и аксоны больших звездчатых нейронов. Клубочки мозжечка снаружи не полностью окружены плоскими отростками астроцитов.
Лазящие волокна мозжечка проникают в кору из белого вещества, проходя через зернистый слой до слоя клеток Пуркинье и стелясь по телам и дендритам этих клеток, на которых они оканчиваются возбуждающими синапсами. Коллатеральные ветви лазящих волокон образуют синапсы на других нейронах всех типов.
Эфферентные волокна коры мозжечка представлены аксонами клеток Пуркинье, которые в виде миелиновых волокон направляются в белое вещество и достигают глубоких ядер мозжечка и вестибулярного ядра, на нейронах которых они образуют тормозные синапсы (клетки Пуркинье являются тормозными нейронами).
Кора полушарий большого мозга
Кора полушарий большого мозга представляет собой высший и наиболее сложно организован-
ный нервный центр, деятельность которого обеспечивает регуляцию разнообразных функций организма и сложные формы поведения. Кора образована слоем серого вещества, покрывающего белое вещество, на поверхности извилин и в глубине борозд. Серое вещество содержит нейроны, нервные волокна и клетки нейроглии всех видов. На основании различий плотности расположения и строения клеток (цитоархитектоники), хода волокон (миелоархитектоники) и функциональных особенностей различных участков коры в ней выделяют 52 нерезко разграниченные поля.
Нейроны коры - мультиполярные, различных размеров и форм, включают более 60 видов, среди которых выделены два основных типа - пирамидные и непирамидные.
Пирамидные клетки - специфический для коры полушарий тип нейронов; по разным оценкам, составляют 50-90 % всех нейронов коры. От апикального полюса их конусовидного (на срезах - треугольного) тела к поверхности коры отходит длинный (апикальный) покрытый шипиками дендрит (рис. 133), направляющийся в молекулярную пластинку коры, где он ветвится. От базальной и латеральных частей тела вглубь коры и в стороны от тела нейрона расходятся несколько более коротких боковых (латеральных) дендритов, которые, ветвясь, распространяются в пределах того же слоя, где находится тело клетки. От середины базальной поверхности тела отходит длинный и тонкий аксон, идущий в белое вещество и дающий коллатерали. Различают гигантские, большие, промежуточные и малые пирамидные клетки. Основная функция пирамидных клеток - обеспечение связей внутри коры (промежуточные и малые клетки) и образование эфферентных путей (гигантские и большие клетки).
Непирамидные клетки располагаются практически во всех слоях коры, воспринимая поступающие афферентные сигналы, а их аксоны распространяются в пределах самой коры, передавая импульсы на пирамидные нейроны. Эти клетки весьма разнообразны и преимущественно являются разновидностями звездчатых клеток. Основная функция непирамидных клеток - интеграция нейронных цепей внутри коры.
Цитоархитектоника коры полушарий большого мозга. Нейроны коры располагаются нерезко разграниченными слоями (пластинками), которые обозначаются римскими цифрами и нумеруются снаружи внутрь. На срезах, окрашенных гематоксилином-эозином, связи между нейронами не прослеживаются, поскольку выявляются лишь
тела нейронов и начальные участки их отростков
(рис. 131).
I - молекулярная пластинка располагается под мягкой мозговой оболочкой; содержит сравнительно небольшое число мелких горизонтальных нейронов с длинными ветвящимися дендритами, отходящими в горизонтальной плоскости от веретеновидного тела. Их аксоны участвуют в образовании тангенциального сплетения волокон этого слоя. В молекулярном слое имеются многочисленные дендриты и аксоны клеток более глубоко расположенных слоев, образующих межнейронные связи.
II - наружная зернистая пластинка образована многочисленными мелкими пирамидными и звездчатыми клетками, дендриты которых ветвятся и поднимаются в молекулярную пластинку, а аксоны либо уходят в белое вещество, либо образуют дуги и также направляются в молекулярную пластинку.
III - наружная пирамидная пластинка характеризуется преобладанием пирамидных нейронов, размеры которых увеличиваются вглубь слоя от малых до больших. Апикальные дендриты пирамидных клеток направляются в молекулярную пластинку, а латеральные образуют синапсы с клетками данной пластинки. Аксоны этих клеток оканчиваются в пределах серого вещества или направляются в белое. Помимо пирамидных клеток, пластинка содержит разнообразные непирамидные нейроны. Пластинка выполняет преимущественно ассоциативные функции, связывая клетки как в пределах данного полушария, так и с противоположным полушарием.
IV - внутренняя зернистая пластинка содержит малые пирамидные и звездчатые клетки. В этой пластинке оканчивается основная часть таламических афферентных волокон. Аксоны клеток этой пластинки образуют связи с клетками выше- и нижележащих пластинок коры.
V - внутренняя пирамидная пластинка образована большими пирамидными нейронами, а в области моторной коры (прецентральной извилины) - гигантскими пирамидными нейронами (клетки Беца). Апикальные дендриты пирамидных нейронов достигают молекулярной пластинки, латеральные дендриты распространяются в пределах той же пластинки. Аксоны гигантских и больших пирамидных нейронов проецируются на ядра головного и спинного мозга, наиболее длинные из них в составе пирамидных путей достигают каудальных сегментов спинного мозга.
VI - мультиформная пластинка образована разнообразными по форме нейронами, причем ее
наружные участки содержат более крупные клетки, а внутренние - более мелкие и редко расположенные. Аксоны этих нейронов уходят в белое вещество в составе эфферентных путей, а дендриты проникают до молекулярной пластики.
Миелоархитектоника коры полушарий большого мозга. Нервные волокна коры полушарий большого мозга включают три группы: 1) афферентные; 2) ассоциативные и комиссуральные; 3) эфферентные.
Афферентные волокна приходят в кору из ниже расположенных отделов головного мозга в виде пучков в составе вертикальных полосок - радиальных лучей (см. рис. 132).
Ассоциативные и комиссуральные волокна - внутрикорковые волокна, которые соединяют между собой различные области коры внутри одного или в разных полушариях соответственно. Эти волокна образуют пучки (полоски), которые проходят параллельно поверхности коры в пластинке I (тангенциальная пластинка), в пластинке II (дисфиброзная пластинка, или полоска Бехтерева), в пластинке IV (полоска наружной зернистой пластинки, или наружная полоска Байярже) и в пластинке V (полоска внутренней зернистой пластинки, или внутренняя полоска Байярже) - см. рис. 132. Последние две системы являются сплетениями, образованными конечными отделами афферентных волокон.
Эфферентные волокна связывают кору с подкорковыми образованиями. Эти волокна идут в нисходящем направлении в составе радиальных лучей.
Типы строения коры полушарий большого мозга.
В отдельных участках коры, связанных с выполнением разных функций, преобладает развитие тех или иных ее слоев, на основании чего различают агранулярный и гранулярный типы коры.
Агранулярный тип коры характерен для ее моторных центров и отличается наибольшим развитием пластинок III, V и VI коры при слабом развитии пластинок II и IV (зернистых). Такие участки коры служат источниками нисходящих проводящих путей.
Гранулярный тип коры характерен для областей расположения чувствительных корковых центров. Он отличается слабым развитием слоев, содержащих пирамидные клетки, при значительной выраженности зернистых (II и IV) пластинок.
Белое вещество головного мозга представлено пучками нервных волокон, которые поднимаются к серому веществу коры из ствола мозга и спускаются к стволу мозга от корковых центров серого вещества.
ОРГАНЫ НЕРВНОЙ СИСТЕМЫ
Органы периферической нервной системы
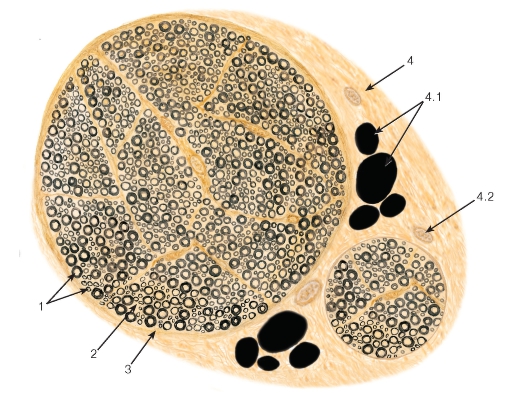
Рис. 114. Нерв (нервный ствол). Поперечный срез
Окраска: осмирование
1 - нервные волокна; 2 - эндоневрий; 3 - периневрий; 4 - эпиневрий: 4.1 - жировая ткань, 4.2 - кровеносный сосуд
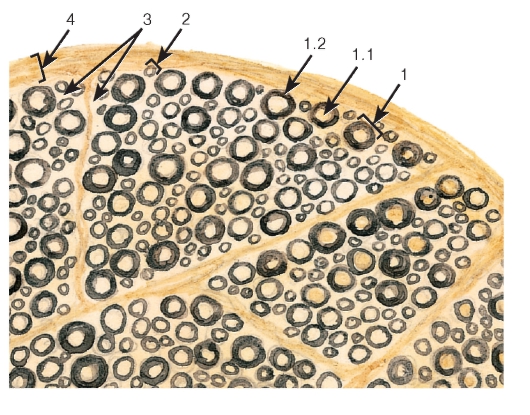
Рис. 115. Участок нерва (нервного ствола)
Окраска: осмирование
1 - миелиновое волокно: 1.1 - отросток нейрона, 1.2 - миелиновая оболочка;
2 - безмиелиновое волокно; 3 - эндоневрий; 4 - периневрий
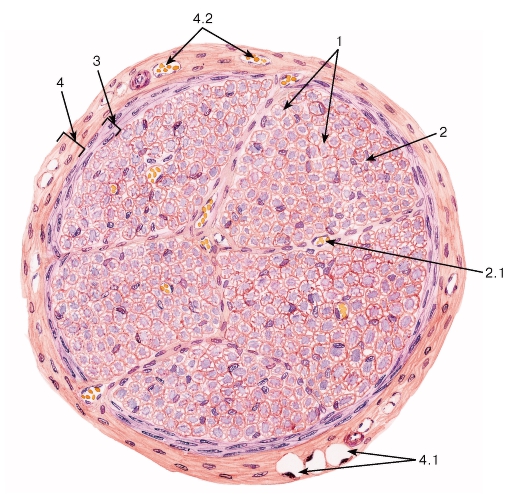
Рис. 116. Нервный ствол (нерв). Поперечный срез
Окраска: гематоксилин-эозин
1 - нервные волокна; 2 - эндоневрий: 2.1 - кровеносный сосуд; 3 - периневрий; 4 - эпиневрий: 4.1 - жировые клетки, 4.2 - кровеносные сосуды
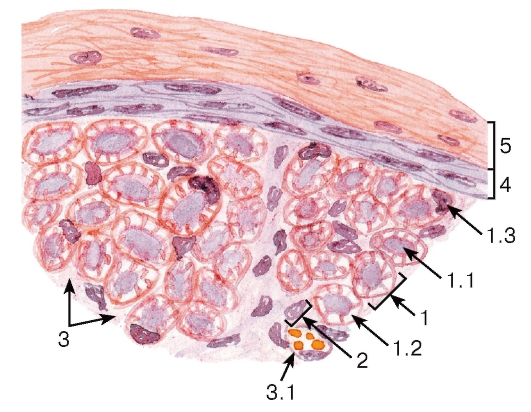
Рис. 117. Участок нервного ствола (нерва)
Окраска: гематоксилин-эозин
1 - миелиновое волокно: 1.1 - отросток нейрона, 1.2 - миелиновая оболочка, 1.3 - ядро нейролеммоцита; 2 - безмиелиновое волокно; 3 - эндоневрий: 3.1 - кровеносный сосуд; 4 - периневрий; 5 - эпиневрий
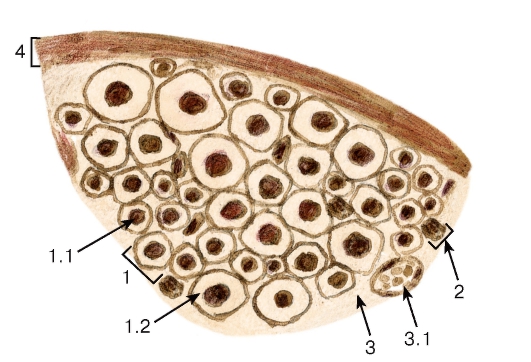
Рис. 118. Участок нервного ствола (нерва)
Окраска: азотнокислое серебро
1 - миелиновое волокно: 1.1 - отросток нейрона, 1.2 - миелиновая оболочка; 2 - безмиелиновое волокно; 3 - эндоневрий: 3.1 - кровеносный сосуд; 4 - периневрий
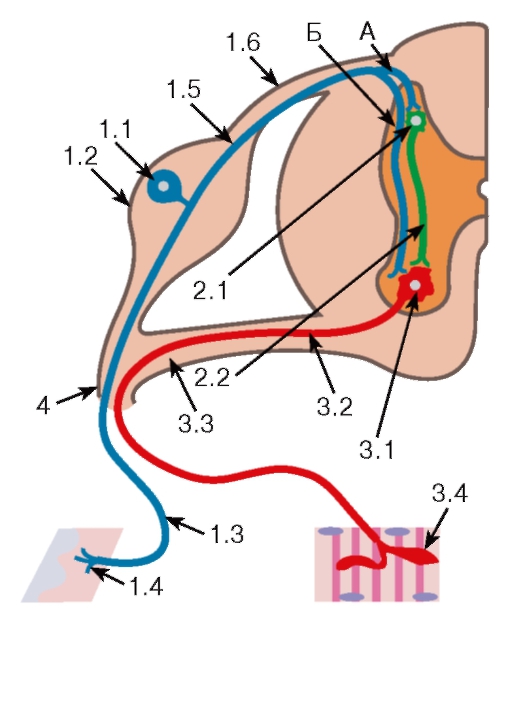
Рис. 119. Соматическая рефлекторная дуга
1. Рецепторное звено образовано афферентными (чувствительными) псевдоуниполярными нейронами, тела которых (1.1) располагаются в чувствительных узлах спинномозгового нерва (1.2). Периферические отростки (1.3) этих клеток образуют чувствительные нервные окончания (1.4) в коже или скелетной мышце. Центральные отростки (1.5) вступают в спинной мозг в составе задних корешков (1.6) и направляются в задние рога серого вещества, образуя синапсы на телах и дендритах вставочных нейронов (трехнейронные рефлекторные дуги, А), или проходят в передние рога к мотонейронам (двухнейронные рефлекторные дуги, Б).
2. Ассоциативное звено представлено мультиполярными вставочными нейронами (2.1), дендриты и тела которых лежат в задних рогах. Их аксоны (2.2) направляются в передние рога, передавая нервные импульсы на тела и дендриты эффекторных нейронов.
3. Эфферентное звено образовано мультиполярными мотонейронами (3.1). Тела и дендриты этих нейронов лежат в передних рогах, формируя двигательные ядра. Аксоны (3.2) мотонейронов выходят из спинного мозга в составе передних корешков (3.3) и далее в составе смешанного нерва (4) направляются к скелетной мышце, где веточки аксона образуют нейро-мышечные синапсы (3.4)
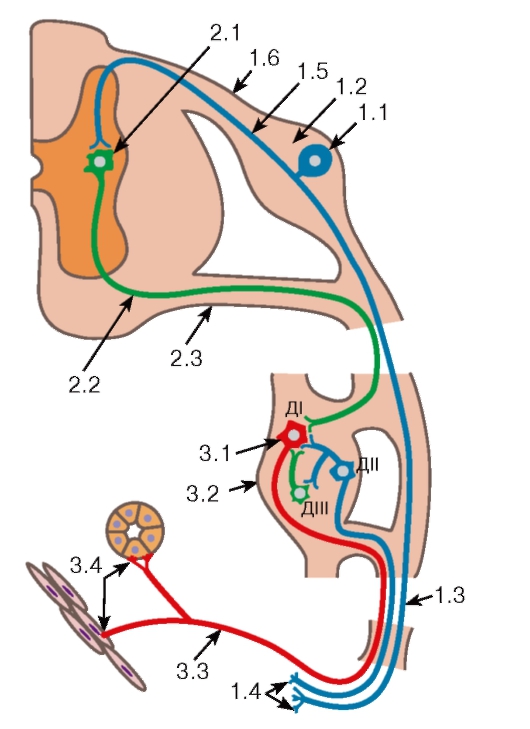
Рис. 120. Автономная (вегетативная) рефлекторная дуга
1. Рецепторное звено образовано афферентными (чувствительными) псевдоуниполярными нейронами, тела которых (1.1) лежат в чувствительных узлах спинномозгового нерва (1.2). Периферические отростки (1.3) этих клеток образуют чувствительные нервные окончания (1.4) в тканях внутренних органов. Центральные отростки (1.5) вступают в спинной мозг в составе зад них корешков (1.6) и направляются в боковые рога серого вещества, образуя синапсы на телах и дендритах вставочных нейронов.
2. Ассоциативное звено представлено мультиполярными вставочными нейронами (2.1), дендриты и тела которых расположены в боковых рогах спинного мозга. Аксоны этих нейронов являются преганглионарными волокнами (2.2). Они покидают спинной мозг в составе передних корешков (2.3), направляясь в один из вегетативных ганглиев, где и заканчиваются на телах и дендритах их нейронов.
3. Эфферентное звено образовано мультиполярными или биполярными нейронами, тела которых (3.1) лежат в автономных ганглиях (3.2). Аксоны этих клеток являются постганглионарными волокнами (3.3). В составе нервных стволов и их ветвей они направляются к клеткам рабочих органов - гладким мышцам, железам, сердцу, образуя на них окончания (3.4). В вегетативных ганглиях помимо «длинноаксонных» эфферентных нейронов - клеток I типа Догеля (ДI), имеются «равноотростчатые» афферентные нейроны - клетки II типа Догеля (ДII), которые входят в качестве рецепторного звена в состав местных рефлекторных дуг, и ассоциативные клетки III типа Догеля (ДIII) - мелкие вставочные нейроны
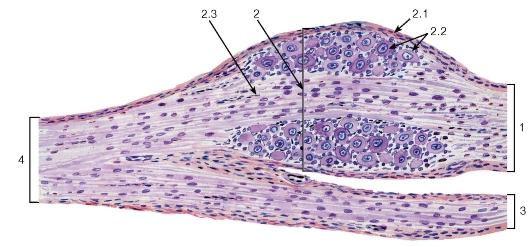
Рис. 121. Чувствительный ганглий спинномозгового нерва
Окраска: гематоксилин-эозин
1 - задний корешок; 2 - чувствительный ганглий спинномозгового нерва: 2.1 - соединительнотканная капсула, 2.2 - тела псевдоуниполярных чувствительных нейронов, 2.3 - нервные волокна; 3 - передний корешок; 4 - спинномозговой нерв
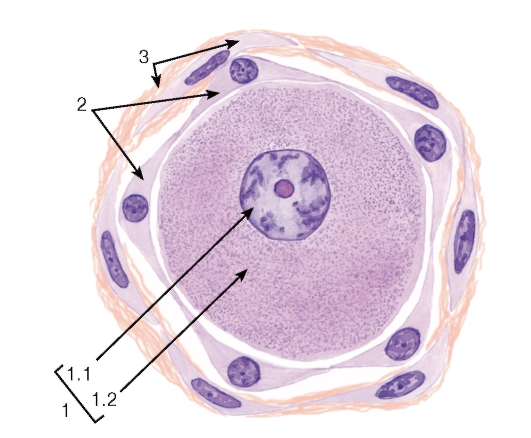
Рис. 122. Псевдоуниполярный нейрон чувствительного ганглия спинномозгового нерва и его тканевое микроокружение
Окраска: гематоксилин-эозин
1 - тело псевдоуниполярного чувствительного нейрона: 1.1 - ядро, 1.2 - цитоплазма; 2 - сателлитные глиальные клетки; 3 - соединительнотканная капсула вокруг тела нейрона
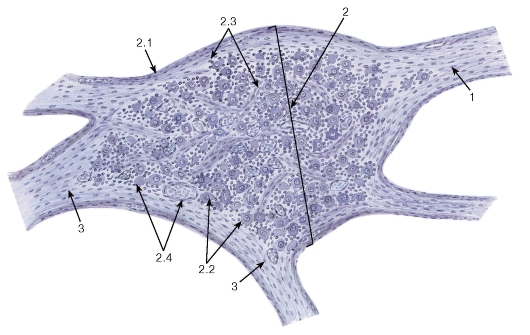
Рис. 123. Автономный (вегетативный) ганглий из солнечного сплетения
Окраска: железный гематоксилин
1 - преганглионарные нервные волокна; 2 - автономный ганглий: 2.1 - соединительнотканная капсула, 2.2 - тела мультиполярных вегетативных нейронов, 2.3 - нервные волокна, 2.4 - кровеносные сосуды; 3 - постганглионарные волокна
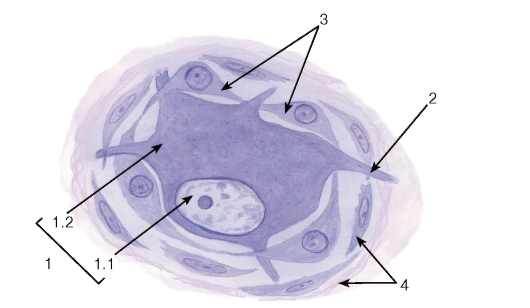
Рис. 124. Мультиполярный нейрон автономного ганглия и его тканевое микроокружение
Окраска: железный гематоксилин
1 - тело мультиполярного нейрона: 1.1 - ядро, 1.2 - цитоплазма; 2 - начало отростков; 3 - глиоциты; 4 - соединительнотканная оболочка
Органы центральной нервной системы
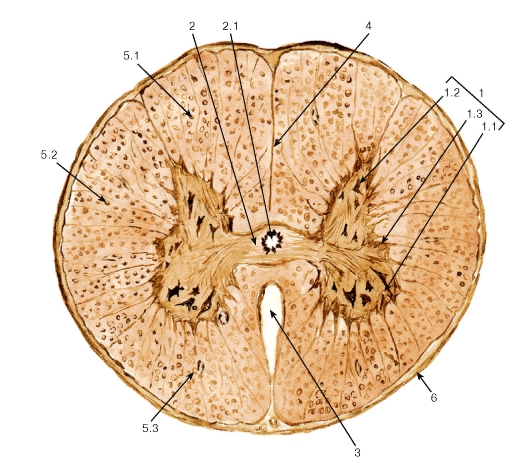
Рис. 125. Спинной мозг (поперечный срез)
Окраска: азотнокислое серебро
1 - серое вещество: 1.1 - передний (вентральный) рог, 1.2 - задний (дорсальный) рог, 1.3 - боковой (латеральный) рог; 2 - передняя и задняя серые спайки: 2.1 - центральный канал; 3 - передняя срединная щель; 4 - задняя срединная борозда; 5 - белое вещество (тракты): 5.1 - дорсальный канатик, 5.2 - латеральный канатик, 5.3 - вентральный канатик; 6 - мягкая оболочка спинного мозга
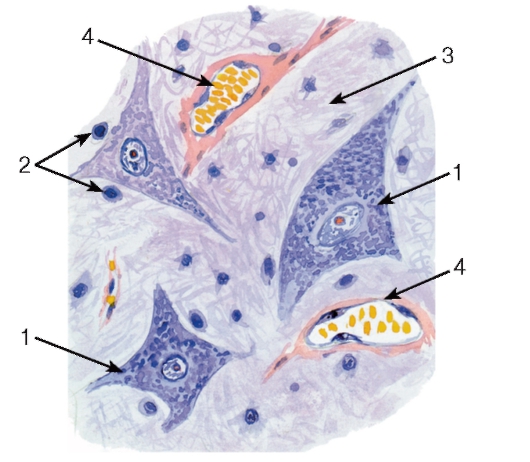
Рис. 126. Спинной мозг.
Участок серого вещества (передние рога)
Окраска: гематоксилин-эозин
1 - тела мультиполярных двигательных нейронов;
2 - глиоциты; 3 - нейропиль; 4 - кровеносные сосуды
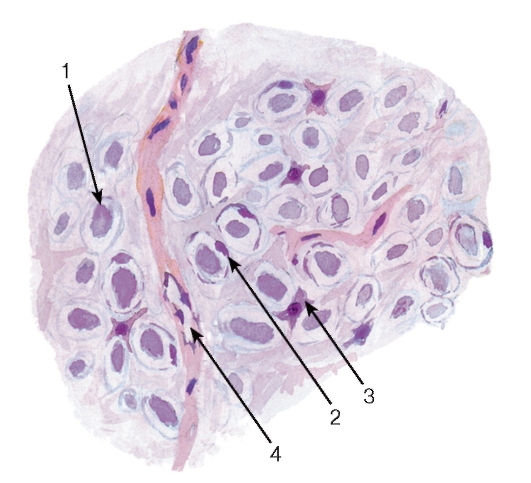
Рис. 127. Спинной мозг. Участок белого вещества
Окраска: гематоксилин-эозин
1 - миелиновые нервные волокна; 2 - ядра олигодендроцитов; 3 - астроциты; 4 - кровеносный сосуд
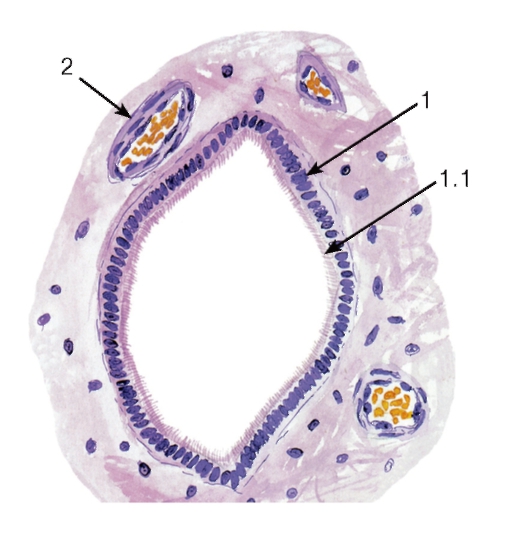
Рис. 128. Спинной мозг. Центральный канал
Окраска: гематоксилин-эозин
1 - эпендимоциты: 1.1 - реснички; 2 - кровеносный сосуд
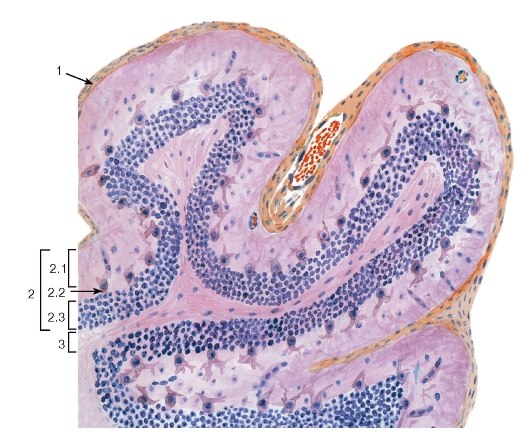
Рис. 129. Мозжечок. Кора
(срез, перпендикулярный ходу извилин)
Окраска: гематоксилин-эозин
1 - мягкая оболочка головного мозга; 2 - серое вещество (кора): 2.1 - молекулярный слой, 2.2 - слой клеток Пуркинье (грушевидных нейронов), 2.3 - зернистый слой; 3 - белое вещество
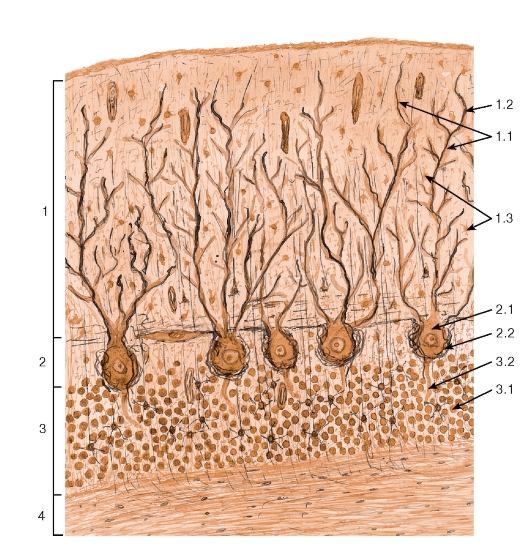
Рис. 130. Мозжечок. Участок коры
Окраска: азотнокислое серебро
1 - молекулярнай слой: 1.1 - дендриты клеток Пуркинье, 1.2 - афферентные (лазящие) волокна, 1.3 - нейроны молекулярного слоя; 2 - слой клеток Пуркинье (грушевидных нейронов): 2.1 - тела грушевидных нейронов (клеток Пуркинье), 2.2 - «корзинки», образованные коллатералями аксонов корзинчатых нейронов; 3 - зернистый слой: 3.1 - тела зернистых нейронов, 3.2 - аксоны клеток Пуркинье; 4 - белое вещество
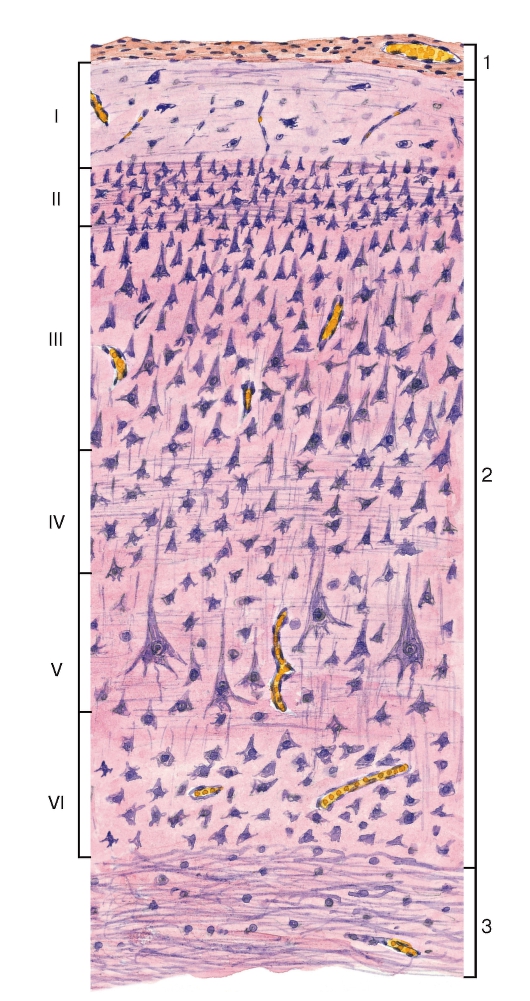
Рис. 131. Полушарие большого мозга. Кора. Цитоархитектоника
Окраска: гематоксилин-эозин
1 - мягкая оболочка головного мозга; 2 - серое вещество: пластинки (слои) коры обозначены римскими цифрами: I - молекулярная пластинка, II - наружная зернистая пластинка, III - наружная пирамидная пластинка, IV - внутренняя зернистая пластинка, V - внутренняя пирамидная пластинка, VI - мультиформная пластинка; 3 - белое вещество

Рис. 132. Полушарие большого мозга. Кора.
Миелоархитектоника
(схема)
1 - тангенциальная пластинка; 2 - дисфиброзная пластинка (полоска Бехтерева); 3 - радиальные лучи; 4 - полоска наружной зернистой пластинки (наружная полоска Байярже); 5 - полоска внутренней зернистой пластинки (внутренняя полоска Байярже)
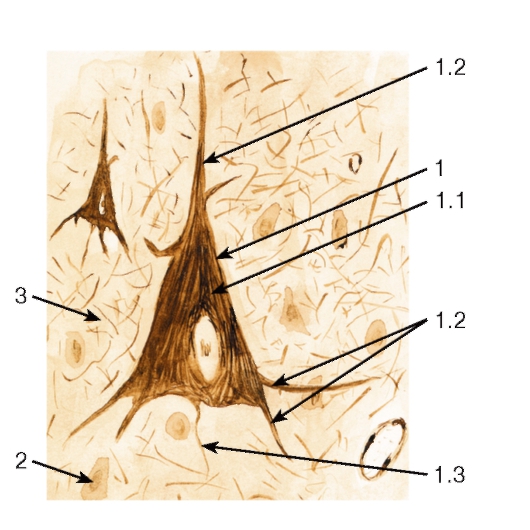
Рис. 133. Большой пирамидный нейрон полушария большого мозга
Окраска: азотнокислое серебро
1 - большой пирамидный нейрон: 1.1 - тело нейрона (перикарион), 1.2 - дендриты, 1.3 - аксон;
2 - глиоциты; 3 - нейропиль