Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. - 296 с. :ил.
|
|
|
|
ЦИТОЛОГИЯ
Цитология - наука о закономерностях строения, развития и жизнедеятельности клетки. В последние годы широкое распространение получил близкий по смыслу термин «биология клетки», который особенно часто используют в тех случаях, когда речь идет об изучении фундаментальных закономерностей жизнедеятельности клетки. Термин «цитология» стал использоваться более ограниченно для обозначения прикладных, в частности, диагностических исследований клеточного материала. В отечественной научной и учебной литературе термин «цитология» часто используется в обоих указанных значениях. Цитологию подразделяют на общую и частную. Общая цитология изучает наиболее общие структурнофункциональные свойства, присущие всем клеткам организма. Как правило, ее изучение предшествует освоению курса гистологии. Частная цитология рассматривает специфические характеристики клеток конкретных тканей и органов, обусловленные особенностями их развития, жизнедеятельности и выполняемых функций. Обычно изучение вопросов частной цитологии включено в материал соответствующих разделов частной гистологии.
Клетка - элементарная структурная, функциональная и генетическая единица в составе всех растительных и животных организмов. Организм взрослого человека состоит примерно из 1013 клеток, которые подразделяют более чем на 200 типов, существенно различающихся своими структурными и функциональными особенностями. Вместе с тем, клетки всех типов характеризуются сходством общей организации и строения важнейших компонентов.
На светооптическом уровне клетки обычно изучают после их фиксации и окрашивания - в исследуемом цитологическом материале (мазках, отпечатках) или на гистологических срезах тканей и органов (рис. 1 и 2). Фиксация обеспечивает сохранность различных структур клетки, окрашивание способствует их выявлению благодаря неодинаковому
сродству данных структур к гистологическим красителям. Наиболее распространенная общеобзорная окраска сочетает основной краситель гематоксилин с кислым красителем эозином (см. рис. 1 и 2). Гематоксилин, как и другие основные красители, связывается со структурами, содержащими кислоты, которые именуются базофильными. К ним относятся ядро (вследствие высокого содержания ДНК и РНК), а также цитоплазма - при высоком содержании в ней рибосом или гранулярной эндоплазматической сети. Эозин, подобно другим кислым красителям, окрашивает различные структуры, содержащие основные вещества (оксифильные, или ацидофильные) - цитоплазму клеток (в особенности, при высоком содержании в ней митохондрий и некоторых белковых секреторных гранул), а также отдельные компоненты межклеточного вещества (например, коллагеновые волокна). Оценить способность тех или иных компонентов клетки связываться с определенными красителями можно, сопоставляя клетки на срезах, окрашенных различными способами (см. рис. 2).
Компоненты клетки. Каждая клетка состоит из двух основных компонентов - ядра и цитоплазмы (см. рис. 1). Ядро отделено от цитоплазмы ядерной оболочкой; при световой микроскопии в неделящейся (интерфазной) клетке в нем выявляются темно окрашенный (гетеро-)хроматин, ядрышко в виде темной крупной гранулы и бесструктурная кариоплазма (см. рис. 1). Более мелкие структурные детали ядра не определяются.
Цитоплазма отделена от внешней (для данной клетки) среды плазмолеммой (клеточной мембраной) и содержит органеллы и включения (рис. 10), погруженные в неструктурированную прозрачную гиалоплазму. Органеллы - постоянно присутствующие в цитоплазме структуры, выполняющие определенные функции в клетке. Включения - временные компоненты цитоплазмы, образующиеся в результате накопления продуктов метаболизма клеток.
СТРУКТУРЫ ЦИТОПЛАЗМЫ, ВЫЯВЛЯЕМЫЕ ПРИ СВЕТОВОЙ МИКРОСКОПИИ
Структуры цитоплазмы, выявляемые при световой микроскопии, сравнительно немногочисленны. С помощью специальных методов окраски в ней можно выявить лишь отдельные виды
органелл, в частности, митохондрии (рис. 3) и комплекс Гольджи (рис. 4 и 101). Большинство же органелл выявляются лишь при использовании электронной микроскопии (рис. 10-16).
С помощью гистохимических методов на уровне светового микроскопа в цитоплазме клеток можно выявить разнообразные по своему химическому составу в ключения, в частности, гранулы гликогена (рис. 5) и липидные капли (рис. 6 и 7). Секреторные включения (секреторные гранулы) отчетливо выявляются в апикальной части клеток концевых отделов поджелудочной железы
(панкреатоцитов) и с помощью стандартных методов окраски препаратов (рис. 8). Пигментные включения цитоплазмы определяются благодаря их собственной окраске (рис. 9); ядро на этом препарате выделено с помощью дополнительной окраски (как и на препаратах, представленных на рис. 4-6) для оценки его расположения, формы и размеров.
СТРУКТУРЫ ЦИТОПЛАЗМЫ, ВЫЯВЛЯЕМЫЕ ПРИ ЭЛЕКТРОННОЙ МИКРОСКОПИИ
Структуры цитоплазмы, выявляемые при электронной микроскопии, в обобщенном виде представлены на объемной схеме ультраструктурной организации клетки, которая демонстрирует также и ультраструктуру клеточного ядра (см. рис. 10). На этой схеме показана плазмолемма (клеточная мембрана), которая занимает в клетке пограничное положение и обеспечивает многообразные процессы взаимодействия клетки с окружающей ее средой (другими клетками, межклеточным веществом). Поверхность клетки, покрытая плазмолеммой, имеет различный рельеф: на одних участках она может быть сравнительно гладкой, на других - содержит зоны специализированных межклеточных соединений (на латеральных участ ках), на третьих (на апикальном полюсе) - покрывает микроворсинки и реснички (на рис. 10 последние не показаны) - специализированные выпячивания цитоплазмы, основу которых образуют высоко организованные элементы цитоскелета. На рис. 10 показано также участие плазмолеммы в процессах эндоцитоза и экзоцитоза. Отдельные органеллы и их группы далее более детально изображены на рисунках, сделанных с электронных микрофотографий (рис. 11-16).
Митохондрии - мембранные органеллы длиной 2-10 мкм и диаметром 0,2-2 мкм, обеспечивающие клетку энергией, которая генерируется благодаря процессам окисления и аккумулируется в виде фосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот. Наиболее типичное строение имеют митохондрии с ламеллярными кристами - пластинчатыми складками внутренней митохондриальной мембраны. Кристы обращены в митохондриальный матрикс - зернистое вещество умеренной плотности, заполняющее полость митохондрии (см. рис. 11) и содержащее множество ферментов, крупные митохондриальные гранулы (с ионами Са2+ и Mg2+), а также (не видны на рисунке) митохондриальные рибосомы и митохондриальную
ДНК. Реже встречаются митохондрии с тубулярно-везикулярными кристами, которые совместно с гладкой эндоплазматической сетью принимают участие в синтезе стероидов (см. рис. 12). Митохондрии располагаются в тех участках цитоплазмы, где происходит активное потребление энергии, например, в области органелл, участвующих в синтезе белков (см. рис. 11).
Рибосомы - мелкие (диаметр - 15-30 нм) плотные немембранные органеллы, состоящие из двух асимметричных субъединиц и обеспечивающие синтез белка из аминокислот (в особенности, молекул, которые после синтеза остаются в гиалоплазме). Они образуют цепочки (полирибосомы, или полисомы), свободно располагающиеся в гиалоплазме или связанные с поверхностью мембран эндоплазматической сети.
Эндоплазматическая сеть - органелла, обеспечивающая синтез углеводов, липидов и белков, а также начальные посттрансляционные изменения последних. Она имеет мембранное строение и состоит из системы уплощенных, удлиненных, трубчатых и везикулярных образований.
Гранулярная эндоплазматическая сеть обеспечивает биосинтез и начальное гликозилирование мембранных белков и белков, предназначенных для экспорта из клетки. Она образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы, придающие мембранам зернистый (гранулярный) вид (см. рис. 11).
Агранулярная эндоплазматическая сеть представляет собой трехмерную систему мембранных анастомозирующих трубочек, канальцев, цистерн и пузырьков, на поверхности которых рибосомы отсутствуют (см. рис. 12). Агранулярная эндоплазматическая сеть участвует в синтезе липидов (в частности, стероидов - совместно с митохондриями с тубулярно-везикулярными кристами, с которыми она обычно соседствует), гликогена, обеспечивает детоксикацию различных веществ и накопление ионов Са2+.
Комплекс Гольджи совместно с рибосомами и эндоплазматической сетью образует синтетический аппарат клетки. Это - сложно организованная поляризованная мембранная органелла, которая представлена тремя основными элементами: (1) стопкой изогнутых уплощенных, расширяющихся по краям мешочков (цистерн), (2) пузырьками и (3) крупными вакуолями, или секреторными пузырьками (см. рис. 13). В комплексе Гольджи выделяют две поверхности, обладающие структурными и функциональными различиями: (а) цис-поверхность (незрелую, формирующуюся) - выпуклой формы, обращенную к эндоплазматической сети; (б) транс-поверхность (зрелую) - вогнутой формы, обращенную к плазмолемме и связанную с отделяющимися от цистерн вакуолями. Функции комплекса Гольджи: синтез полисахаридов и гликопротеинов (гликокаликса, слизи); химические изменения (процессинг) молекул, транспортируемых из гранулярной эндоплазматической сети, конденсация секреторного продукта и образование секреторных гранул; обеспечение новообразованных гранул мембраной и упаковка в нее секреторных продуктов; сортировка белков на транс-поверхности перед их окончательным транспортом.
Эндосомы и лизосомы образуют аппарат внутриклеточного переваривания, функция которого состоит в регулируемом внутриклеточном расщеплении макромолекул внеклеточного и внутриклеточного происхождения.
Эндосомы - мембранные пузырьки с постепенно закисляющимся содержимым (pH 6,0-5,5), которые обеспечивают перенос макромолекул с поверхности клетки в лизосомы и их частичный или полный гидролиз на стадиях, предшествующих лизосомальному уровню деградации.
Лизосомы - мембранные органеллы диаметром 0,1-2 мкм, активно участвующие в завершающих этапах процесса полного внутриклеточного переваривания захваченных клеткой макромолекул посредством широкого спектра литических ферментов при низких значениях рН (5,0 и ниже). Лизосомы, не способные полностью переварить находящиеся в них молекулы, преобразуются в остаточные тельца, которые могут долго находиться в цитоплазме или выделять свое содержимое за пределы клетки. Распространенным типом остаточных телец в организме человека являются липофусциновые гранулы - мембранные пузырьки, содержащие труднорастворимый коричневый эндогенный пигмент липофусцин (см. рис. 12), который рассматривают как «пигмент старения» или «изнашивания».
Пероксисомы (микротельца) - мембранные пузырьки диаметром 0,05-1,5 мкм с умеренно плотным однородным или мелкозернистым матриксом,
содержащим многочисленные ферменты, катализирующие окислительно-восстановительные реакции, и каталазу. В матриксе иногда выявляется более плотная кристаллоидная сердцевина (нуклеоид), (см. рис. 14) - область конденсации ферментов. Пероксисомы участвуют в расщеплении жирных кислот и других метаболических реакциях (обмен аминокислот, оксалата и полиаминов, синтез некоторых фосфолипидов). Они защищают клетку от действия перекиси водорода, оказывающей сильный повреждающий эффект, а также разрушают ряд токсических веществ.
Цитоскелет представляет собой сложную динамичную систему микротрубочек, микрофиламентов и промежуточных филаментов. Эти компоненты цитоскелета являются немембранными органеллами, образующими в клетке трехмерные сети. Они входят также в состав ряда других более сложно организованных органелл (ресничек, жгутиков, микроворсинок, клеточного центра) и клеточных соединений (десмосом, полудесмосом, опоясывающих десмосом).
Микротрубочки - наиболее крупные компоненты цитоскелета (диаметр около 24-25 нм) - полые цилиндрические образования длиной до нескольких микрометров (см. рис. 15).
Их стенка состоит из димерами из белковых молекул α- и β-тубулина. Важнейшие функции микротрубочек: поддержание формы и полярности клетки, обеспечение внутриклеточного транспорта, движения ресничек и хромосом в митозе (формируют ахроматиновое веретено, необходимое для клеточного деления). Микротрубочки в цитоплазме образуют сети, либо располагаются в виде пучков, например, в отростках нейронов, в составе митотического веретена (см. рис. 15).
Микротрубочки образуют основу других органелл (центриолей, ресничек, жгутиков), частично сливаясь друг с другом с формированием пар, или дублетов (в аксонеме ресничек и жгутиков - см. рис. 37), и триплетов (в базальном тельце и центриоли).
Центриоль (см. рис. 16) - цилиндрическая структура длиной 0,3-0,5 мкм и диаметром 0,15- 0,2 мкм из девяти триплетов частично слившихся микротрубочек, объединенных поперечными белковыми мостиками («ручками»). С каждым триплетом посредством ножек связаны сателлиты - глобулярные белковые тельца, которые являются центрами образования микротрубочек. В клеточном центре содержатся две центриоли, которые располагаются во взаимно перпендикулярных плоскостях.
Микрофиламенты - тонкие нити диаметром 5-7 нм, образованные преимущественно белком актином, лежат в цитоплазме поодиночке, в виде
сетей или пучками. В большинстве клеток концентрируются в области кортикальной (терминальной) сети под плазмолеммой. Взаимодействуя с другими белками, обладают сократимостью, в частности, они обеспечивают сократимость мышечных клеток, процессы экзо- и эндоцитоза, образование псевдоподий и миграцию клеток. Микрофиламенты участвуют в организации структуры некоторых межклеточных соединений (опоясывающих десмосом), образуют основу («каркас») некоторых органелл, например, микроворсинок (рис. 35), стереоцилий.
Промежуточные филаменты - белковые нити толщиной около 10 нм, располагающиеся в цитоплазме в виде трехмерной сети, окружающей ядро. Они обеспечивают распределение органелл по определенным участкам цитоплазмы клетки, участвуют в образовании рогового вещества в эпителии, входят в состав десмосом и полудесмосом.
В клетках различных тканей промежуточные филаменты различаются по своей химической природе и молекулярной массе.
ЯДРО, ДЕЛЕНИЕ И ГИБЕЛЬ КЛЕТКИ
Ядро клетки. При исследовании клеток под электронным микроскопом в ядре (рис. 17) обнаруживаются многие структурные детали, которые не выявляются под световым микроскопом (ср. с рис. 1).
Ядерная оболочка, которая практически не определяется под световым микроскопом, имеет сложную ультраструктурную организацию: она состоит из двух ядерных мембран (наружной и внутренней), разделенных узким перинуклеарным пространством и содержит ядерные поры, через которые осуществляется обмен молекулами между ядром и цитоплазмой. На поверхности наружной ядерной мембраны имеются рибосомы, к внутренней ядерной мембране прилегает часть скоплений хроматина.
Хроматин состоит из комплекса ДНК и белка и соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими нитями и неразличимы как индивидуальные структуры. Выделяют эу- и гетерохроматин.
Эухроматин - деспирализованные и практически не окрашенные участки хромосом, которые активно участвуют в транскрипции. Гетерохроматин соответствует конденсированным интенсивно окрашенным участкам хромосом, не участвующим в транскрипции. Скопления гетерохроматина располагаются под ядерной оболочкой и вокруг ядрышка, более мелкие глыбки разбросаны по всему ядру.
Ядрышко образовано специализированными участками хромосом - ядрышковыми организаторами. Функции ядрышка заключаются в синтезе рибосомальной РНК и ее сборке в предшественники субъединиц рибосом. В интерфазном ядре на светооптическом уровне оно выявляется как мелкая плотная базофильная гранула (см. рис. 1). Под электронным микроскопом обнаруживается сложная структурная организация ядрышка, в частности, выявляется образующая его плотная
широкопетлистая сеть - нуклеолонема (ядрышковая нить), которая состоит из волокнистой части (молекул рибосомальной РНК) и гранулярной части (субъединиц рибосом).
Нуклеоплазма (кариоплазма) - жидкий, не окрашивающийся компонент ядра, в котором располагаются хроматин и ядрышко. Она содержит воду и ряд растворенных и взвешенных в ней молекул.
Клеточный цикл - совокупность явлений между двумя последовательными делениями клетки или между ее образованием и гибелью. Он обеспечивает функцию воспроизведения и передачи генетической информации. Клеточный цикл включает собственно митоз, или фазу М (митотическое деление), и интерфазу - промежуток между делениями.
Интерфаза значительно более длительна, чем митоз (обычно занимает не менее 90% всей продолжительности клеточного цикла) и подразделяется на три части (рис. 18): постмитотический интервал (G1), фазу синтеза ДНК (S) и премитотический интервал (G2).
Переход от одного периода интерфазы к другому (в особенности, от G1 к S), а также вступление клетки в митоз регулируются сложными молекулярными механизмами и опосредуются специальными факторами.
Митотическое деление клеток (митоз, или непрямое деление) является универсальным механизмом деления клеток. Оно следует за G2-интервалом и завершает клеточный цикл, обеспечивая равномерное распределение генетического материала в дочерние клетки. Митоз включает 4 основные фазы: профазу, метафазу, анафазу и телофазу
(рис. 19).
Профаза начинается с конденсации хромосом, которые становятся видимыми как нитевидные структуры. Ядрышко и ядерная оболочка к концу профазы исчезают, кариоплазма смешивается с цитоплазмой. Центриоли мигрируют к противопо-
ложным полюсам клетки и дают начало волокнам митотического (ахроматинового) веретена, часть которых прикрепляются к хромосомам.
Метафаза соответствует максимальному уровню конденсации хромосом, которые выстраиваются в области экватора митотического веретена, образуя картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со стороны полюсов).
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды и движения дочерних хромосом к противоположным полюсам клетки. Она завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картины звезд (стадия дочерних звезд).
Телофаза - конечная стадия митоза, в течение которой реконструируются ядра дочерних клеток и завершается их разделение. Вокруг конденсированных хромосом дочерних клеток восстанавливается кариолемма, вновь появляются ядрышки. Хромосомы прогрессивно деспирализуются и исчезают, замещаясь картиной хроматина интерфазного ядра. Одновременно происходит перешнуровка цитоплазмы с формированием двух дочерних клеток, между которыми осуществляется равномерное распределение органелл.
Гибель клеток, наряду с их размножением и дифференцировкой, является одним из ключевых процессов и факторов в обеспечении нормальной жизнедеятельности различных тканей. При гибели клеток могут наблюдаться два главных принципиально различных вида морфологических изменений, которые соответствуют различным механизмам ее развития - некроз и апоптоз.
Некроз возникает под действием резко выраженных повреждающих факторов (перегревания, переохлаждения, недостатка кислорода, нарушения кровоснабжения, химических препаратов, механической травмы и др.). Он охватывает различные по численности группы клеток, сопровождается набуханием цитоплазмы и отдельных органелл, обширными повреждениями мембран, уплотнением,
сморщиванием, распадом и лизисом ядра. Выделяющиеся при разрыве плазмолеммы продукты распада клетки привлекают лейкоциты и макрофаги, способствуя воспалительной реакции.
Апоптоз - физиологическая (запрограммированная) гибель клеток - активный, регулируемый генетической программой процесс, который запускается внешними факторами. Он обычно протекает асинхронно в отдельных клетках или мелких клеточных группах, разделенных численно превосходящими жизнеспособными клетками. Апоптоз наблюдается в различных тканях человека и животных в норме, патологии, эмбриональном развитии и у взрослого, являясь одним из фундаментальных и универсальных биологических механизмов тканевого гомеостаза. Апоптоз развивается под влиянием физиологических активаторов (индукторов апоптоза), разнообразных повреждающих физических и химических факторов, нарушения баланса регуляторных воздействий, некоторых микробов (в особенности, вирусов).
Морфологические проявления апоптоза обладают характерными особенностями (рис. 20). На ранних стадиях процесса происходит утрата клетками специализированных структур на их поверхности (например, межклеточных соединений), их отделение от соседних. Происходят уплотнение ядра (с периферическим отложением гетерохроматина в виде крупных полулуний) и конденсация цитоплазмы, которая приводит ко все более компактному расположению органелл, сохраняющих свою целостность. Нарастающая конденсация цитоплазмы сочетается с изменением формы клетки - она образует многочисленные крупные вздутия и выпячивания; ядро окончательно уплотняется, сморщивается (кариопикноз) и распадается на фрагменты, окруженные мембраной. Выпячивания клетки, содержащие органеллы и фрагменты ядра, отшнуровываются, формируя крупные окруженные мембраной фрагменты - апоптотические тельца. Последние быстро захватываются и перевариваются соседними клетками или местными фагоцитами; воспалительная реакция при этом не развивается.
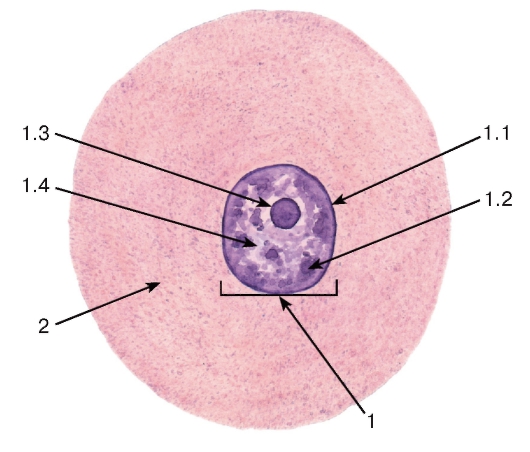
Рис. 1. Строение клетки по данным световой микроскопии
(на примере нервной клетки чувствительного узла спинномозгового нерва)
Окраска: гематоксилин-эозин
1 - ядро: 1.1 - ядерная оболочка (кариолемма), 1.2 - хроматин, 1.3 - ядрышко, 1.4 - кариоплазма;
2 - цитоплазма
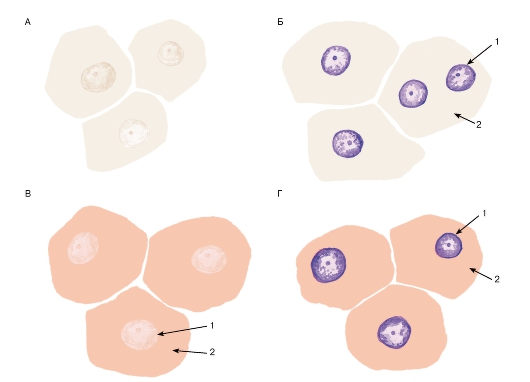
Рис. 2. Клетки на гистологических срезах, окрашенных различными способами:
А - неокрашенный срез; Б - срез, окрашенный гематоксилином; В - срез, окрашенный эозином; Г - срез, окрашенный гематоксилином и эозином: 1 - ядро; 2 - цитоплазма
СТРУКТУРЫ ЦИТОПЛАЗМЫ (выявляемые при световой микроскопии)
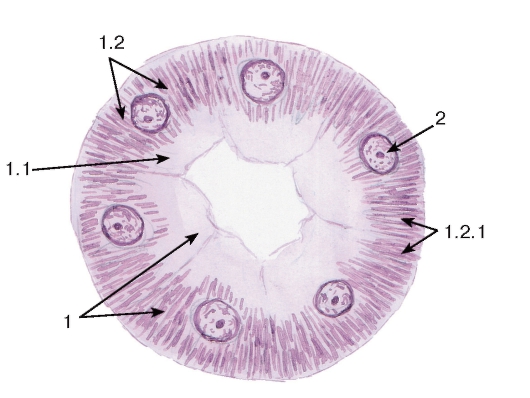
Рис. 3. Митохондрии
(в эпителиальных клетках почечного канальца)
Окраска: железный гематоксилин
1 - цитоплазма клетки почечного канальца: 1.1 - апикальная часть, 1.2 - базальная часть, 1.2.1 - митохондрии; 2 - ядро
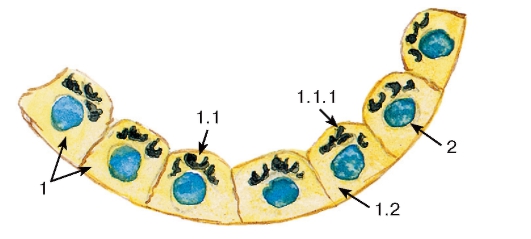
Рис. 4. Комплекс Гольджи
(в фолликулярных клетках щитовидной
железы - тироцитах)
Окраска: азотнокислое серебро-гематоксилин
1 - цитоплазма тироцита: 1.1 - апикальная часть, 1.1.1 - комплекс Гольджи, 1.2 - базальная часть;
2 - ядро
Примечание. Здесь и далее использованный вариант окраски (импрегнации) азотнокислым серебром не указывается

Рис. 5. Включения гликогена
(в клетках печени - гепатоцитах)
Окраска: ШИК-реакция и гематоксилин
1 - цитоплазма гепатоцита: 1.1 - гранулы гликогена;
2 - ядро
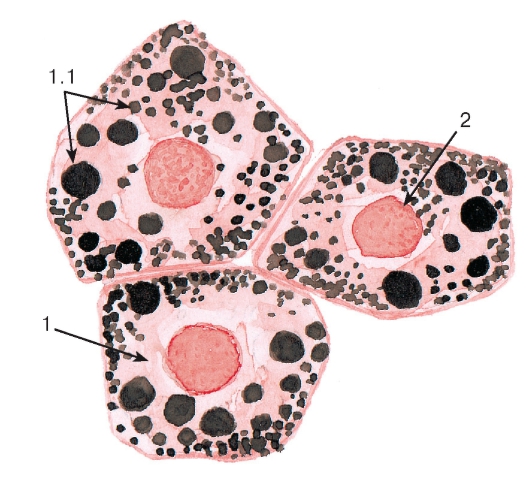
Рис. 6. Липидные включения
(в клетках печени - гепатоцитах)
Окраска: судан черный-сафранин
1 - цитоплазма гепатоцита: 1.1 - липидные капли;
2 - ядро
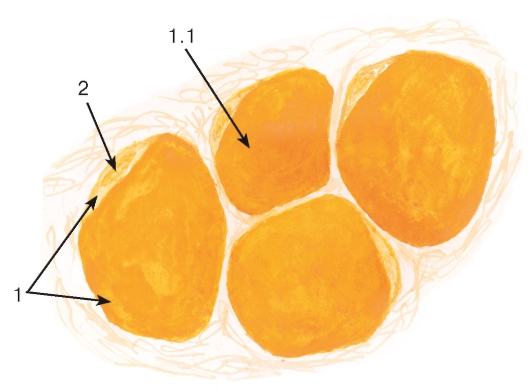
Рис. 7. Липидные включения
(в жировых клетках - адипоцитах)
Окраска: судан III
1 - цитоплазма адипоцита: 1.1 - липидная капля, занимающая большую часть цитоплазмы; 2 - область расположения ядра
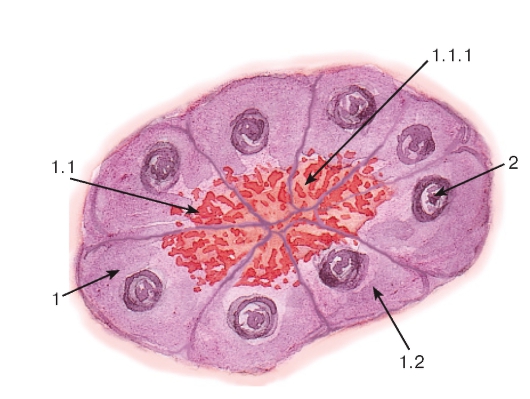
Рис. 8. Секреторные включения
(в клетках концевых отделов поджелудочной
железы - панкреатоцитах)
Окраска: гематоксилин-эозин
1 - цитоплазма панкреатоцита: 1.1 - апикальная часть, 1.1.1 - гранулы секрета, 1.2 - базальная часть; 2 - ядро
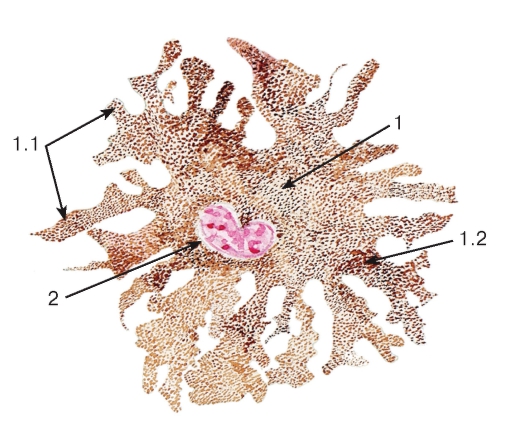
Рис. 9. Пигментные включения
(в пигментных клетках кожи аксолотля):
плоскостной (пленочный) препарат
Окраска ядра: кармин
1 - цитоплазма пигментной клетки: 1.1 - отростки, 1.2 - гранулы пигмента (меланина); 2 - ядро
СТРУКТУРЫ ЦИТОПЛАЗМЫ (выявляемые при электронной микроскопии)
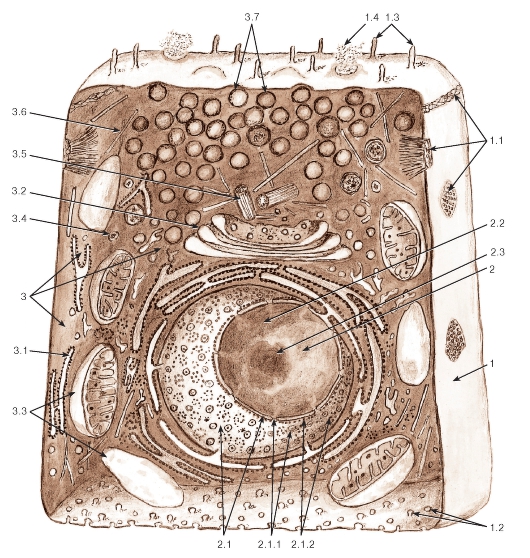
Рис. 10. Объемная схема ультраструктурной организации клетки:
1 - плазмолемма: 1.1 - участки межклеточных соединений на латеральной поверхности, 1.2 - эндоцитозные пузырьки на базальной поверхности, 1.3 - микроворсинки на апикальной поверхности, 1.4 - участок выделения секрета; 2 - ядро: 2.1 - ядерная оболочка, 2.1.1. - ядерные поры, 2.1.2 - рибосомы на поверхности ядерной оболочки, 2.2 - хроматин, 2.3 - ядрышко; 3 - цитоплазма: 3.1 - гранулярная эндоплазматическая сеть, 3.2 - комплекс Гольджи, 3.3 - митохондрии, 3.4 - лизосомы, 3.5 - клеточный центр, 3.6 - микротрубочки, 3.7 - секреторные гранулы
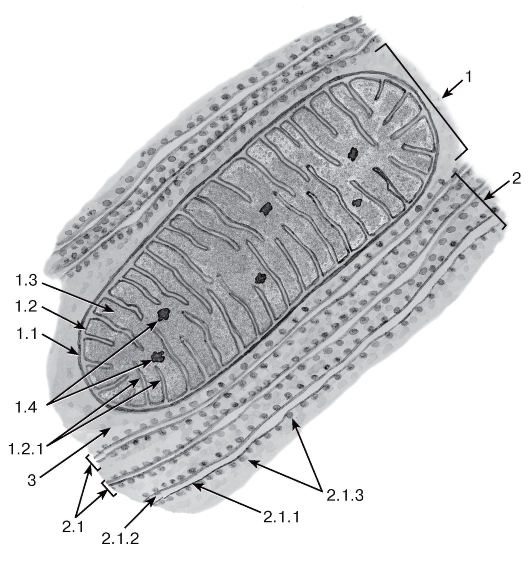
Рис. 11. Митохондрия с ламеллярными кристами и гранулярная эндоплазматическая сеть
Рисунок с электронной микрофотографии (ЭМФ)
1 - митохондрия: 1.1 - наружная митохондриальная мембрана, 1.2 - внутренняя митохондриальная мембрана, 1.2.1 - кристы, 1.3 - митохондриальный матрикс, 1.4 - митохондриальные гранулы; 2 - гранулярная эндоплазматическая сеть: 2.1 - цистерны, 2.1.1 - мембрана, 2.1.2- просвет цистерны, 2.1.3 - рибосомы; 3 - гиалоплазма
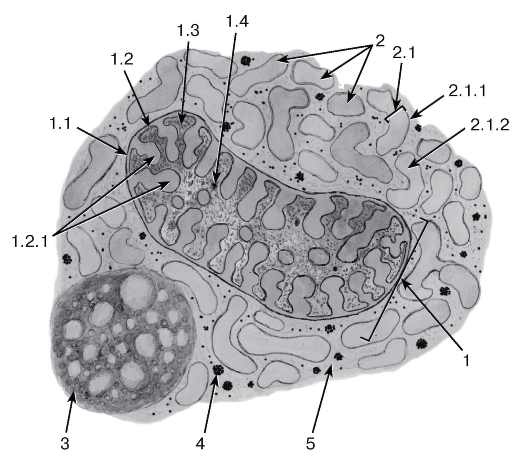
Рис. 12. Митохондрия с тубулярно-везикулярными кристами, агранулярная эндоплазматическая сеть, липофусциновая гранула и гранулы гликогена
Рисунок с ЭМФ
1 - митохондрия: 1.1 - наружная митохондриальная мембрана, 1.2 - внутренняя митохондриальная мембрана, 1.2.1 - кристы, 1.3 - митохондриальный матрикс, 1.4 - митохондриальные гранулы; 2 - агранулярная эндоплазматическая сеть: 2.1 - цистерна, 2.1.1 - мембрана, 2.1.2 - просвет цистерны; 3 - липофусциновая гранула; 4 - гранулы гликогена; 5 - гиалоплазма
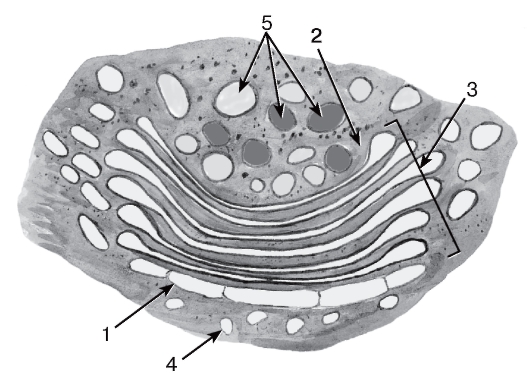
Рис. 13. Комплекс Гольджи
Рисунок с ЭМФ
1 - цис-поверхность; 2 - транс-поверхность; 3 - цистерны (мешочки); 4 - пузырьки; 5 - вакуоли
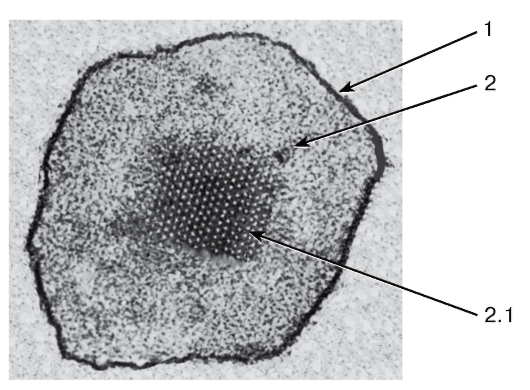
Рис. 14. Пероксисома
Рисунок с ЭМФ
1 - мембрана; 2 - матрикс: 2.1 - кристаллоидная сердцевина (нуклеоид)
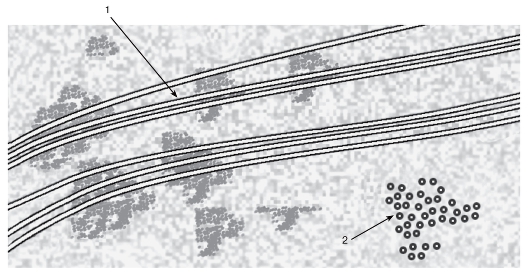
Рис. 15. Микротрубочки митотического веретена
Рисунок с ЭМФ
1 - продольный срез; 2 - поперечный срез
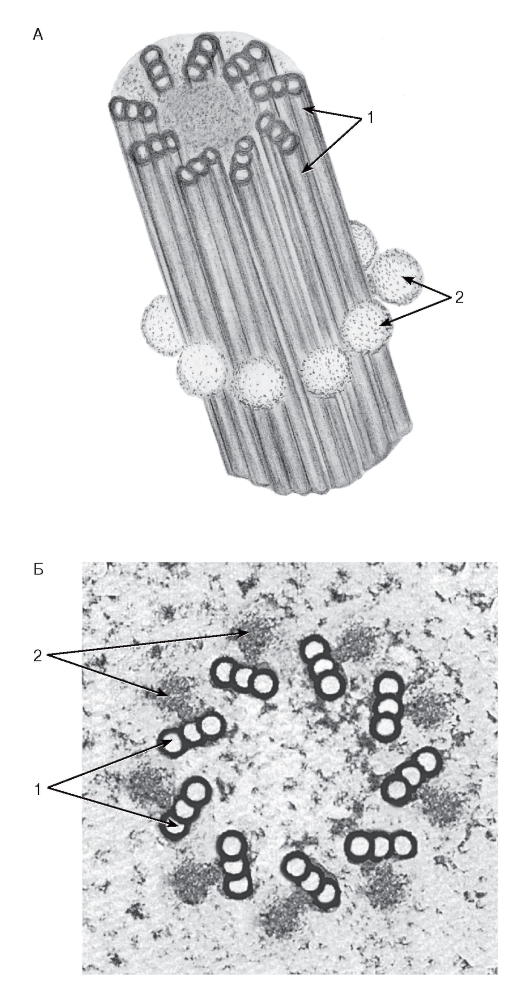
Рис. 16. Центриоль:
А - общий вид (объемная реконструкция); Б - поперечный срез (рисунок с ЭМФ)
1 - триплеты микротрубочек; 2 - центриолярные сателлиты (центры организации микротрубочек)
ЯДРО, ДЕЛЕНИЕ И ГИБЕЛЬ КЛЕТКИ
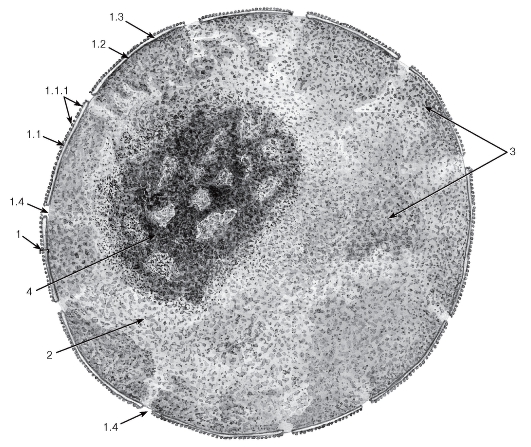
Рис. 17. Ультраструктурная организация ядра клетки
Рисунок с ЭМФ
1 - ядерная оболочка: 1.1 - наружная ядерная мембрана, 1.1.1 - рибосомы, 1.2 - внутренняя ядерная мембрана, 1.3 - перинуклеарное пространство, 1.4 - ядерные поры; 2 - нуклеоплазма (кариоплазма); 3 - хроматин; 4 - ядрышко
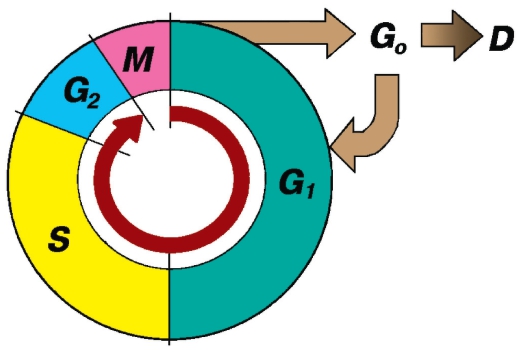
Рис. 18. Клеточный цикл (схема):
М - митоз (фаза М) Периоды (части) интерфазы:
G1-период, или фаза G1 (постмитотический интервал), наступает сразу после митотического деления клетки, характеризуется активным ее ростом, восстановлением необходимого набора органелл и синтезом «запускающих» белков-активаторов S-периода.
Gg-период(фаза G0) - репродуктивный покой. Часто это период гетеросинтеза, когда клетка дифференцируется и выполняет свои специфические функции вплоть до своей гибели (D). При стимуляции некоторые клетки способны возвращаться в G1-период.
S-период, или фаза синтеза ДНК, характеризуется репликацией (удвоением) содержания ДНК и синтезом белков, обеспечивающих упаковку вновь синтезируемой ДНК. Удваивается число центриолей. G2-период, или фаза G2 (премитотический интервал), продолжается до начала митоза. В течение этого времени в клетке происходит подготовка к делению: созревание центриолей, накопление энергии, синтез РНК и белков, необходимых для процесса деления
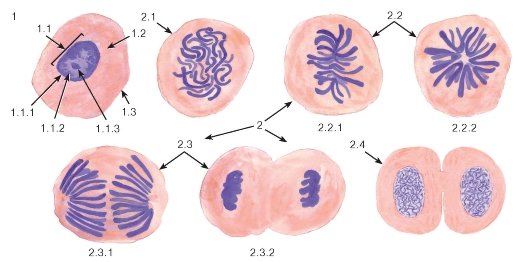
Рис. 19. Митотическое деление животных клеток
Окраска: железный гематоксилин
1 - клетка в интерфазе: 1.1 - ядро, 1.1.1 - ядерная оболочка, 1.1.2 - хроматин, 1.1.3 - ядрышко, 1.2 - цитоплазма, 1.3 - плазмолемма; 2 - митотически делящиеся клетки: 2.1 - профаза, 2.2 - метафаза: 2.2.1 - метафазная (экваториальная) пластинка, 2.2.2 - «материнская звезда», 2.3 - анафаза: 2.3.1 - ранняя, 2.3.2 - поздняя, 2.4 - телофаза
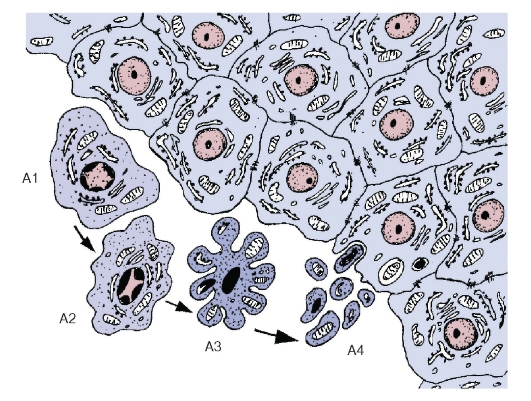
Рис. 20. Апоптоз. Морфологические изменения клеток при апоптозе (схема):
А1 - начало апоптоза: утрата клеткой соединений с соседними интактными клетками и ее отделение от них. А2 - сжатие и уплотнение цитоплазмы и ядра, изменение формы клетки, распределение гетерохроматина в виде полулуний под кариолеммой. A3 - нарастающее сжатие и уплотнение клетки, образование вздутий и выростов на ее поверхности, кариопикноз. А4 - распад клетки на фрагменты, окруженные плазмолеммой (апоптотические тельца), и их фагоцитоз соседними интактными клетками