Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. - 296 с. :ил.
|
|
|
|
ЭПИТЕЛИАЛЬНЫЕ ТКАНИ
Эпителиальные ткани, или эпителии, - пограничные ткани, которые располагаются на границе с внешней средой, покрывают поверхность тела и слизистых оболочек внутренних органов, выстилают его полости и образуют большинство желез.
Важнейшие свойства эпителиальных тканей: сомкнутое расположение клеток (эпителиоцитов), образующих пласты, наличие хорошо развитых межклеточных соединений, расположение на базальной мембране (особом структурном образовании, которое находится между эпителием и подлежащей рыхлой волокнистой соединительной тканью), минимальное количество межклеточного вещества,
пограничное положение в организме, полярность, высокая способность к регенерации.
Основные функции эпителиальных тканей: барьерная, защитная, секреторная, рецепторная.
Морфологические особенности эпителиоцитов тесно связаны с функцией клеток и их положением в эпителиальном пласте. По форме эпителиоциты разделяют на плоские, кубические и столбчатые (призматические, или цилиндрические). Ядро эпителиоцитов в большинстве клеток сравнительно светлое (преобладает эухроматин) и крупное, по форме соответствует форме клетки. Цитоплазма эпителиоцитов, как правило, содержит хорошо
1 В международной гистологической терминологии отсутствует.
2 В зарубежной литературе термином «синцитий» обычно обозначают и симпластические структуры, а термин «симпласт» практически не используется.
развитые органеллы. В клетках железистого эпителия имеется активный синтетический аппарат. Базальная поверхность эпителиоцитов прилежит к базальной мембране, к которой она прикреплена с помощью полудесмосом - соединений, сходных по строению с половинами десмосом.
Базальная мембрана связывает эпителий и подлежащую соединительную ткань; на светооптическом уровне на препаратах она имеет вид бесструктурной полоски, не окрашивается гематоксилином-эозином, однако выявляется солями серебра и дает интенсивную ШИК-реакцию. На ультраструктурном уровне в ней обнаруживаются два слоя: (1) светлая пластинка (lamina lucida, или lamina rara), прилежащая к плазмолемме базальной поверхности эпителиоцитов, (2) плотная пластинка (lamina densa), обращенная в сторону соединительной ткани. Эти слои различаются содержанием белков, гликопротеинов и протеогликанов. Нередко описывают еще третий слой - ретикулярную пластинку (lamina reticularis), содержащую ретикулярные фибриллы, однако многие авторы рассматривают ее как компонент соединительной ткани, не относя к собственно базальной мембране. Базальная мембрана способствует поддержанию нормальной архитектоники, дифференцировки и поляризации эпителия, обеспечивает его прочную связь с подлежащей соединительной тканью, осуществляет избирательную фильтрацию питательных веществ, поступающих в эпителий.
Межклеточные соединения, или контакты, эпителиоцитов (рис. 30) - специализированные участки на их латеральной поверхности, которые обеспечивают связь клеток друг с другом и способствуют формированию ими пластов, что служит важнейшим отличительным свойством организации эпителиальных тканей.
(1) Плотное (замыкающее) соединение (zonula occludens) представляет собой область частичного слияния наружных листков плазмолемм двух соседних клеток, блокирующую распространение веществ по межклеточному пространству. Оно имеет вид пояска, окружающего клетку по периметру (у ее апикального полюса) и состоящего из анастомозирующих тяжей внутримембранных частиц.
(2) Опоясывающая десмосома, или адгезивный поясок (zonula adherens), локализуется на латеральной поверхности эпителиоцита, охватывая клетку по периметру в виде пояска. К листкам плазмолеммы, утолщенным изнутри в области соединения, прикрепляются элементы цитоскелета - актиновые микрофиламенты. Расширенная межклеточная щель содержит адгезивные белковые молекулы (кадгерины).
(3) Десмосома, или пятно адгезии (macula adherens), состоит из утолщенных дисковидных участков плазмолемм двух соседних клеток (внутриклеточных десмосомных уплотнений, или десмосомных пластинок), которые служат участками прикреп-
ления к плазмолемме промежуточных филаментов (тонофиламентов) и разделены расширенной межклеточной щелью, содержащей адгезивные белковые молекулы (десмоколлины и десмоглеины).
(4) Пальцевидное межклеточное соединение (интердигитация) образовано выпячиваниями цитоплазмы одной клетки, вдающимися в цитоплазму другой, в результате чего увеличивается прочность соединения клеток друг с другом и нарастает площадь поверхности, через которую могут осуществляться межклеточные обменные процессы.
(5) Щелевое соединение, или нексус (nexus), образовано совокупностью трубчатых трансмембранных структур (коннексонов), пронизывающих плазмолеммы соседних клеток и стыкующихся друг с другом в области узкой межклеточной щели. Каждый коннексон состоит из субъединиц, образованных белком коннексином, и пронизан узким каналом, который обусловливает свободный обмен низкомолекулярными соединениями между клетками, обеспечивая их ионное и метаболическое сопряжение. Именно поэтому щелевые соединения относят к коммуникационным соединениям, обеспечивающим химическую (метаболическую, ионную и электрическую) связь между эпителиоцитами, в отличие от плотных и промежуточных соединений, десмосом и интердигитаций, обусловливающих механическую связь эпителиоцитов друг с другом и поэтому именуемых механическими межклеточными соединениями.
Апикальная поверхность эпителиоцитов может быть гладкой, складчатой или содержать реснички, и (или) микроворсинки.
Виды эпителиальных тканей: 1) покровные эпителии (образуют разнообразные выстилки); 2) железистые эпителии (образуют железы); 3) сенсорные эпителии (выполняют рецепторные функции, входят в состав органов чувств).
Классификации эпителиев основаны на двух признаках: (1) строении, которое определяется функцией (морфологическая классификация), и (2) источниках развития в эмбриогенезе (гистогенетическая классификация).
Морфологическая классификация эпителиев разделяет их в зависимости от количества слоев в эпителиальном пласте и формы клеток (рис. 31). По количеству слоев эпителии подразделяют на однослойные (если все клетки расположены на базальной мембране) и многослойные (если на базальной мембране расположен лишь один слой клеток). Если все клетки эпителия связаны с базальной мембраной, но имеют разную форму, а их ядра располагаются в несколько рядов, то такой эпителий именуют многорядным (псевдомногослойным). По форме клеток эпителии подразделяют на плоские, кубические и столбчатые (призматические, цилиндрические). В многослойных эпителиях под их формой подразумевают форму клеток поверхностного слоя. Эта классификация
учитывает также некоторые дополнительные признаки, в частности, наличие специальных органелл (микроворсинчатой, или щеточной, каемки и ресничек) на апикальной поверхности клеток, их способность к ороговению (последний признак относится только к многослойным плоским эпителиям). Особый вид многослойных эпителиев, изменяющих свое строение в зависимости от растяжения, встречается в мочевыводящих путях и называется переходным эпителием (уротелием).
Гистогенетическая классификация эпителиев разработана акад. Н. Г. Хлопиным и выделяет пять основных типов эпителия, развивающихся в эмбриогенезе из различных тканевых зачатков.
1. Эпидермальный тип развивается из эктодермы и прехордальной пластинки.
2. Энтеродермальный тип развивается из кишечной энтодермы.
3. Целонефродермальный тип развивается из целомической выстилки и нефротома.
4. Ангиодермальный тип развивается из ангиобласта (участка мезенхимы, образующего сосудистый эндотелий).
5. Эпендимоглиальный тип развивается из нервной трубки.
Покровные эпителии
Однослойный плоский эпителий образован уплощенными клетками с некоторым утолщением в области расположения дисковидного ядра (рис. 32 и 33). Этим клеткам свойственна диплазматическая дифференцировка цитоплазмы, в которой выделяется расположенная вокруг ядра более плотная часть (эндоплазма), содержащая большую часть органелл, и более светлая наружная часть (эктоплазма) с низким содержанием органелл. Вследствие малой толщины эпителиального пласта через него легко диффундируют газы и быстро транспортируются различные метаболиты. Примерами однослойного плоского эпителия служат выстилка полостей тела - мезотелий (см. рис. 32), сосудов и сердца - эндотелий (рис. 147, 148); он образует стенку некоторых почечных канальцев (см. рис. 33), альвеол легкого (рис. 237, 238). Истонченная цитоплазма клеток этого эпителия на поперечных гистологических срезах обычно прослеживается с трудом, отчетливо выявляются лишь уплощенные ядра; более полное представление о строении эпителиоцитов можно получить на плоскостных (пленочных) препаратах (см. рис. 32 и 147).
Однослойный кубический эпителий образован клетками, содержащими ядро сферической формы и набор органелл, которые развиты лучше, чем в клетках плоского эпителия. Такой эпителий встречается в мелких собирательных протоках мозгового вещества почки (см. рис. 33), почечных ка-
нальцах (рис. 250), в фолликулах щитовидной железы (рис. 171), в мелких протоках поджелудочной железы, желчных протоках печени.
Однослойный столбчатый эпителий (призматический, или цилиндрический) образован клетками с резко выраженной полярностью. Ядро сферической, чаще - эллипсоидной формы обычно смещено к их базальной части, а хорошо развитые органеллы неравномерно распределены по цитоплазме. Такой эпителий образует стенку крупных собирательных протоков почки (см. рис. 33), покрывает поверхность слизистой оболочки желудка
(рис. 204-206), кишки (рис. 34, 209-211, 213-215),
образует выстилку желчного пузыря (рис. 227), крупных желчных протоков и протоков поджелудочной железы, маточной трубы (рис. 271) и матки (рис. 273). Для большинства указанных эпителиев характерны функции секреции и (или) всасывания. Так, в эпителии тонкой кишки (см. рис. 34), встречаются два основных типа дифференцированных клеток - столбчатые каемчатые клетки, или энтероциты (обеспечивают пристеночное пищеварение и всасывание), и бокаловидные клетки, или бокаловидные экзокриноциты (вырабатывают слизь, которая выполняет защитную функцию). Всасывание обеспечивается многочисленными микроворсинками на апикальной поверхности энтероцитов, совокупность которых образует исчерченную (микроворсинчатую) каемку (см. рис. 35). Микроворсинки покрыты плазмолеммой, поверх которой располагается слой гликокаликса, их основу образует пучок актиновых микрофиламентов, вплетающийся в кортикальную сеть микрофиламентов.
Однослойный многорядный столбчатый реснитчатый эпителий наиболее характерен для воздухоносных путей (рис. 36). В нем имеются клетки (эпителиоциты) четырех основных типов: (1) базальные, (2) вставочные, (3) реснитчатые и (4) бокаловидные.
Базальные клетки мелких размеров своим широким основанием прилежат к базальной мембране, а узкой апикальной частью не доходят до просвета. Они являются камбиальными элементами ткани, обеспечивающими ее обновление, и, дифференцируясь, постепенно превращаются во вставочные клетки, которые затем дают начало реснитчатым и бокаловидным клеткам. Последние вырабатывают слизь, которая покрывает поверхность эпителия, перемещаясь по ней благодаря биению ресничек реснитчатых клеток. Реснитчатые и бокаловидные клетки своей узкой базальной частью контактируют с базальной мембраной и прикрепляются к вставочным и базальным клеткам, а апикальной - граничат с просветом органа.
Реснички - органеллы, участвующие в процессах движения, на гистологических препаратах имеют вид тонких прозрачных выростов на апикальной
поверхности цитоплазмы эпителиоцитов (см. рис. 36). При электронной микроскопии обнаруживается, что их основу составляет каркас из микротрубочек (аксонема, или осевая нить), который образован девятью периферическими дублетами (парами) частично слившихся микротрубочек и одной центрально расположенной парой (рис. 37). Аксонема связана с базальным тельцем, которое лежит в основании реснички, по своей структуре идентично центриоли и продолжается в исчерченный корешок. Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дублетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом нексиновыми мостиками и взаимодействуют между собой с помощью динеиновых ручек. При этом соседние дублеты в аксонеме скользят друг относительно друга, обусловливая биение реснички.
Многослойный плоский ороговевающий эпителий состоит из пяти слоев: (1) базального, (2) шиповатого, (3) зернистого, (4) блестящего и (5) рогового (рис. 38).
Базальный слой образован кубическими или столбчатыми клетками с базофильной цитоплазмой, лежащими на базальной мембране. Этот слой содержит камбиальные элементы эпителия и обеспечивает прикрепление эпителия к подлежащей соединительной ткани.
Шиповатый слой образован крупными клетками неправильной формы, связанными друг с другом многочисленными отростками - «шипами». При электронной микроскопии в области шипов выявляются десмосомы и связанные с ними пучки тонофиламентов. По мере приближения к зернистому слою клетки из полигональных постепенно становятся уплощенными.
Зернистый слой - сравнительно тонкий, образован уплощенными (веретеновидными на разрезе) клетками с плоским ядром и цитоплазмой с крупными базофильными кератогиалиновыми гранулами, содержащими один из предшественников рогового вещества - профилаггрин.
Блестящий слой выражен только в эпителии толстой кожи (эпидермисе), покрывающем ладони и подошвы. Он имеет вид узкой оксифильной гомогенной полоски и состоит из уплощенных живых эпителиальных клеток, превращающихся в роговые чешуйки.
Роговой слой (наиболее поверхностный) имеет максимальную толщину в эпителии кожи (эпидермисе) в области ладоней и подошв. Он образован плоскими роговыми чешуйками с резко утолщенной плазмолеммой (оболочкой), не содержащими ядра и органелл, дегидратированными и заполненными роговым веществом. Последнее на ультраструктурном уровне представлено сетью из толстых пучков кератиновых филаментов, погруженных в плотный матрикс. Роговые чешуйки сохраняют связи друг с
другом и удерживаются в составе рогового слоя благодаря частично сохраненным десмосомам; по мере разрушения десмосом в наружных частях слоя чешуйки слущиваются (десквамируют) с поверхности эпителия. Многослойный плоский ороговевающий эпителий образует эпидермис - наружный слой кожи (см. рис. 38, 177), покрывает поверхность некоторых участков слизистой оболочки полости рта (рис. 182).
Многослойный плоский неороговевающий эпителий образован тремя слоями клеток: (1) базальным, (2) промежуточным и (3) поверхностным (рис. 39). Глубокую часть промежуточного слоя иногда выделяют как парабазальный слой.
Базальный слой имеет такое же строение и выполняет те же функции, что и одноименный слой в многослойном плоском ороговевающем эпителии.
Промежуточный слой образован крупными полигональными клетками, которые по мере приближения к поверхностному слою уплощаются.
Поверхностный слой нерезко отделен от промежуточного и образован уплощенными клетками, которые механизмом десквамации постоянно удаляются с поверхностности эпителия. Многослойный плоский неороговевающий эпителий покрывает поверхность роговицы глаза (см. рис. 39, 135), конъюнктивы, слизистых оболочек полости рта- частично (см. рис. 182, 183, 185, 187), глотки, пищевода (рис. 201, 202), влагалища и влагалищной части шейки матки (рис. 274), части мочеиспускательного канала.
Переходный эпителий (уротелий) - особый вид многослойного эпителия, который выстилает большую часть мочевыводящих путей - чашечки, лоханки, мочеточники и мочевой пузырь (рис. 40, 252, 253), часть мочеиспускательного канала. Форма клеток этого эпителия и его толщина зависят от функционального состояния (степени растяжения) органа. Переходный эпителий образован тремя слоями клеток: (1) базальным, (2) промежуточным и (3) поверхностным (см. рис. 40).
Базальный слой представлен мелкими клетками, которые своим широким основанием прилежат к базальной мембране.
Промежуточный слой состоит из удлиненных клеток, более узкой частью направленных к базальному слою и черепицеобразно накладывающихся друг на друга.
Поверхностный слой образован крупными одноядерными полиплоидными или двуядерными поверхностными (зонтичными) клетками, которые в наибольшей степени изменяют свою форму (от округлой до плоской) при растяжении эпителия.
Железистые эпителии
Железистые эпителии образуют большинство желез - структур, которые выполняют секреторную функцию, вырабатывая и выделяя разнообраз-
ные продукты (секреты), обеспечивающие различные функции организма.
Классификация желез основана на учете различных признаков.
По числу клеток железы подразделяют на одноклеточные (например, бокаловидные клетки, клетки диффузной эндокринной системы) и многоклеточные (большинство желез).
По расположению (относительно эпителиального пласта) выделяют эндоэпителиальные (лежащие в пределах эпителиального пласта) и экзоэпителиальные (расположенные за пределами эпителиального пласта) железы. Большинство желез относятся к экзоэпителиальным.
По месту (направлению) выведения секрета железы разделяют на эндокринные (выделяющие секреторные продукты, называемые гормонами, в кровь) и экзокринные (выделяющие секреты на поверхность тела или в просвет внутренних органов).
В экзокринных железах выделяют (1) концевые (секреторные) отделы, которые состоят из железистых клеток, продуцирующих секрет, и (2) выводные протоки, обеспечивающие выделение синтезированных продуктов на поверхность тела или в полость органов.
Морфологическая классификация экзокринных желез основана на структурных признаках их концевых отделов и выводных протоков.
По форме концевых отделов железы подразделяют на трубчатые и альвеолярные (сферической формы). Последние иногда описывают также как ацинусы. При наличии двух типов концевых отделов железы называются трубчатоальвеолярными или трубчато-ацинарными.
По ветвлению концевых отделов выделяют неразветвленные и разветвленные железы, по ветвлению выводных протоков - простые (с неразветвленным протоком) и сложные (с разветвленными протоками).
По химическому составу вырабатываемого секрета железы подразделяют на белковые (серозные), слизистые, смешанные (белково-слизистые), липидные и др.
По механизму (способу) выведения секрета (рис. 41-46) выделяют мерокринные железы (выделение секрета без нарушения структуры клетки), апокринные (с отделением в секрет части апикальной цитоплазмы клеток) и голокринные (с полным разрушением клеток и выделением их фрагментов в секрет).
Мерокринные железы преобладают в организме человека; этот тип секреции хорошо демонстрируется на примере ацинарных клеток поджелудочной железы - панкреатоцитов (см. рис. 41 и 42). Синтез белкового секрета ацинарных клеток происходит
в гранулярной эндоплазматической сети, расположенной в базальной части цитоплазмы (см. рис. 42), отчего эта часть на гистологических препаратах окрашивается базофильно (см. рис. 41). Синтез завершается в комплексе Гольджи, где образуются секреторные гранулы, которые накапливаются в апикальной части клетки (см. рис. 42), обусловливая ее оксифильное окрашивание на гистологических препаратах (см. рис. 41).
Апокринные железы в организме человека немногочисленны; к ним относятся, например, часть потовых желез и молочные железы (см. рис. 43, 44, 279).
В лактирующей молочной железе концевые отделы (альвеолы) образованы железистыми клетками (галактоцитами), в апикальной части которых накапливаются крупные липидные капли, отделяющиеся в просвет вместе с небольшими участками цитоплазмы. Этот процесс отчетливо прослеживается при электронной микроскопии (см. рис. 44), а также на светооптическом уровне при использовании гистохимических методов выявления липидов (см. рис. 43).
Голокринные железы в организме человека представлены единственным видом - сальными железами кожи (см. рис. 45 и 46, а также рис. 181). В концевом отделе такой железы, имеющем вид железистого мешочка, можно проследить деление мелких периферических базальных (камбиальных) клеток, их смещение к центру мешочка с заполнением липидными включениями и превращением в себоциты. Себоциты приобретают вид вакуолизированных дегенерирующих клеток: их ядро сморщивается (подвергается пикнозу), цитоплазма переполняется липидами, а плазмолемма на конечных стадиях разрушается с выделением клеточного содержимого, образующего секрет железы - кожное сало.
Секреторный цикл. Процесс секреции в железистых клетках протекает циклически и включает последовательные фазы, которые могут частично перекрываться. Наиболее типичен секреторный цикл экзокринной железистой клетки, вырабатывающей белковый секрет, который включает (1) фазу поглощения исходных веществ, (2) фазу синтеза секрета, (3) фазу накопления синтезированного продукта и (4) фазу выделения секрета (рис. 47). В эндокринной железистой клетке, синтезирующей и выделяющей стероидные гормоны, секреторный цикл имеет некоторые особенности (рис. 48): после фазы поглощения исходных веществ следует фаза депонирования в цитоплазме липидных капель, содержащих субстрат для синтеза стероидных гормонов, а вслед за фазой синтеза накопления секрета в виде гранул не происходит, синтезированные молекулы сразу же выделяются из клетки механизмами диффузии.
ЭПИТЕЛИАЛЬНЫЕ ТКАНИ
Покровные эпителии
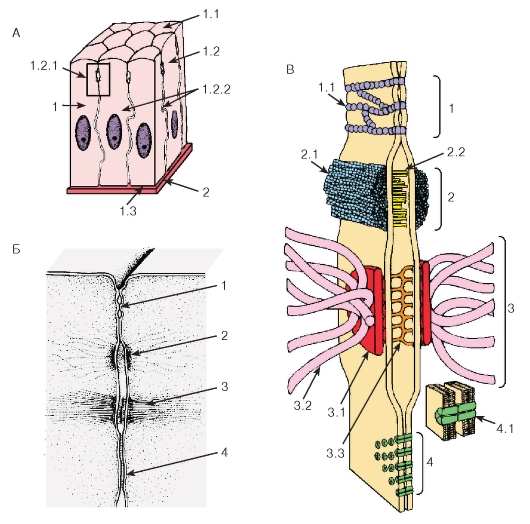
Рис. 30. Схема межклеточных соединений в эпителиях:
А - область расположения комплекса межклеточных соединений (выделена рамкой):
1 - эпителиоцит: 1.1 - апикальная поверхность, 1.2 - латеральная поверхность, 1.2.1 - комплекс межклеточных соединений, 1.2.2 - пальцевидные соединения (интердигитации), 1.3 - базальная поверхность;
2 - базальная мембрана.
Б - вид межклеточных соединений на ультратонких срезах (реконструкция):
1 - плотное (замыкающее) соединение; 2 - опоясывающая десмосома (адгезивный поясок); 3 - десмосома; 4 - щелевое соединение (нексус).
В - трехмерная схема строения межклеточных соединений:
1 - плотное соединение: 1.1 - внутримембранные частицы; 2 - опоясывающая десмосома (адгезивный поясок): 2.1 - микрофиламенты, 2.2 - межклеточные адгезивные белки; 3 - десмосома: 3.1 - десмосомная пластинка (внутриклеточное десмосомное уплотнение), 3.2 - тонофиламенты, 3.3 - межклеточные адгезивные белки; 4 - щелевое соединение (нексус): 4.1 - коннексоны
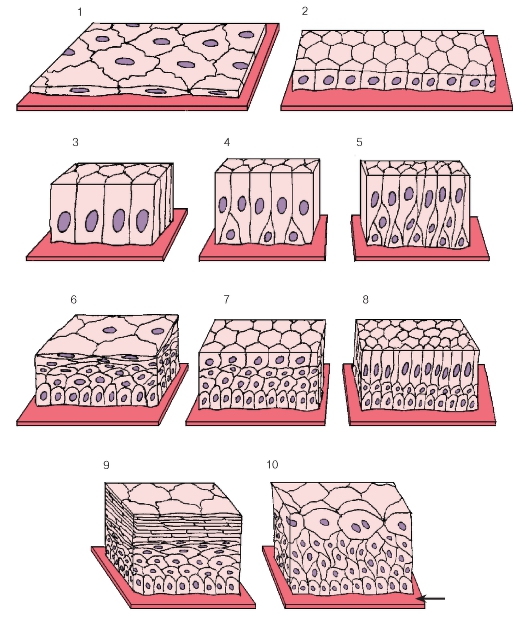
Рис. 31. Морфологическая классификация эпителиев:
1 - однослойный плоский эпителий; 2 - однослойный кубический эпителий; 3 - однослойный (однорядный) столбчатый (призматический) эпителий; 4, 5 - однослойный многорядный (псевдомногослойный) столбчатый эпителий; 6 - многослойный плоский неороговевающий эпителий; 7 - многослойный кубический эпителий; 8 - многослойный столбчатый эпителий; 9 - многослойный плоский ороговевающий эпителий; 10 - переходный эпителий (уротелий)
Стрелкой показана базальная мембрана
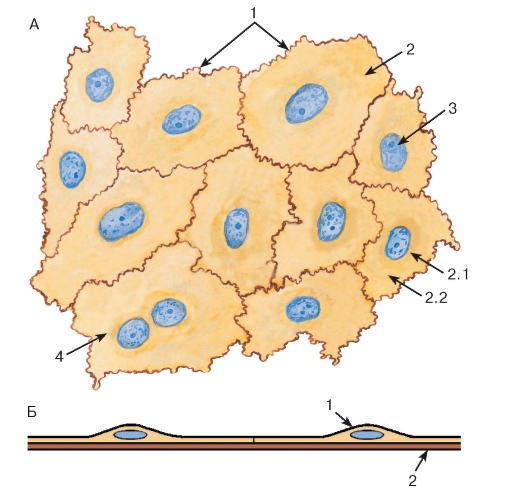
Рис. 32. Однослойный плоский эпителий (мезотелий брюшины):
А - плоскостной препарат
Окраска: азотнокислое серебро-гематоксилин
1 - границы эпителиоцитов; 2 - цитоплазма эпителиоцита: 2.1 - эндоплазма, 2.2 - эктоплазма; 3 - ядро эпителиоцита; 4 - двуядерная клетка
Б - схема строения на срезе:
1 - эпителиоцит; 2 - базальная мембрана
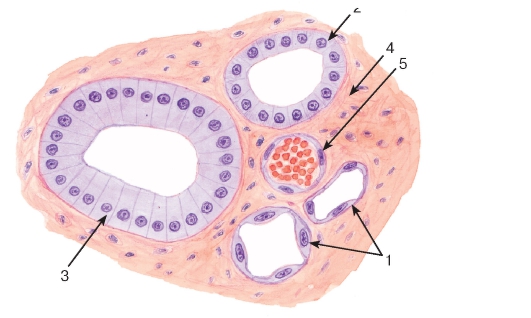
Рис. 33. Однослойные плоский, кубический и столбчатый (призматический) эпителии (мозговое вещество почки)
Окраска: гематоксилин-эозин
1 - однослойный плоский эпителий; 2 - однослойный кубический эпителий; 3 - однослойный столбчатый эпителий; 4 - соединительная ткань; 5 - кровеносный сосуд
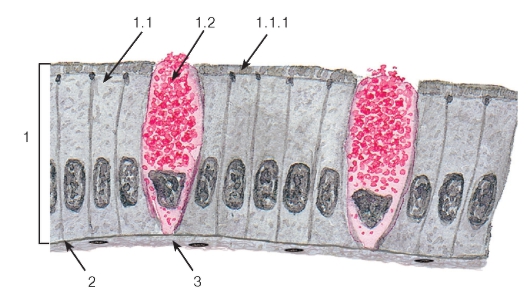
Рис. 34. Однослойный столбчатый каемчатый (микроворсинчатый) эпителий (тонкая кишка)
Окраска: железный гематоксилин-муцикармин
1 - эпителий: 1.1 - столбчатый каемчатый (микроворсинчатый) эпителиоцит (энтероцит), 1.1.1 - исчерченная (микроворсинчатая) каемка, 1.2 - бокаловидный экзокриноцит; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань
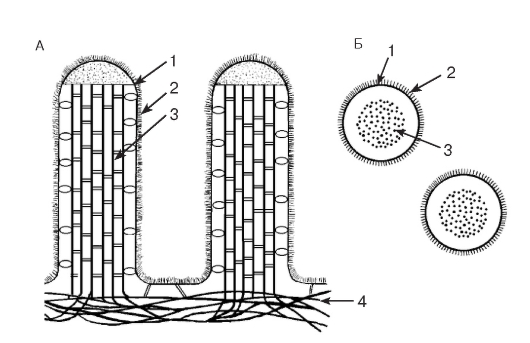
Рис. 35. Микроворсинки клеток кишечного эпителия (схема ультраструктуры):
А - продольные срезы микроворсинок; Б - поперечные срезы микроворсинок:
1 - плазмолемма; 2 - гликокаликс; 3 - пучок актиновых микрофиламентов; 4 - кортикальная сеть микрофиламентов
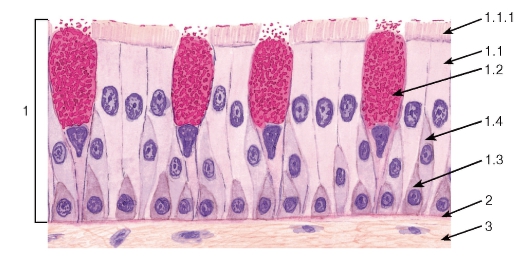
Рис. 36. Однослойный многорядный столбчатый реснитчатый (мерцательный) эпителий (трахея)
Окраска: гематоксилин-эозин-муцикармин
1 - эпителий: 1.1 - реснитчатый эпителиоцит, 1.1.1 - реснички, 1.2 - бокаловидный экзокриноцит, 1.3 - базальный эпителиоцит, 1.4 - вставочный эпителиоцит; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань
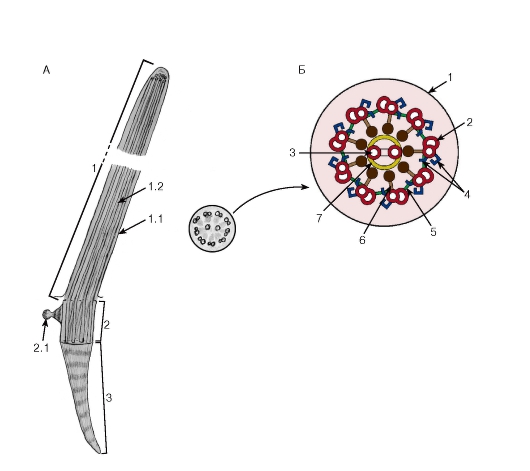
Рис. 37. Ресничка (схема ультраструктуры):
А - продольный срез:
1 - ресничка: 1.1 - плазмолемма, 1.2 - микротрубочки; 2 - базальное тельце: 2.1 - сателлит (центр организации микротрубочек); 3 - базальный корешок
Б - поперечный срез:
1 - плазмолемма; 2 - дуплеты микротрубочек; 3 - центральная пара микротрубочек; 4 - динеиновые ручки; 5 - нексиновые мостики; 6 - радиальные спицы; 7 - центральная оболочка
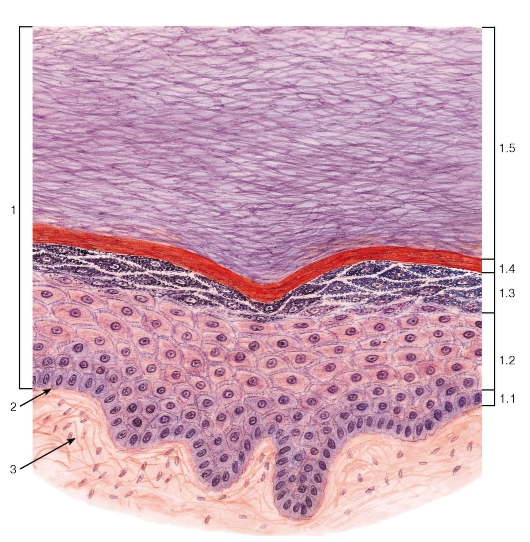
Рис. 38. Многослойный плоский ороговевающий эпителий (эпидермис толстой кожи)
Окраска: гематоксилин-эозин
1 - эпителий: 1.1 - базальный слой, 1.2 - шиповатый слой, 1.3 - зернистый слой, 1.4 - блестящий слой, 1.5 - роговой слой; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань
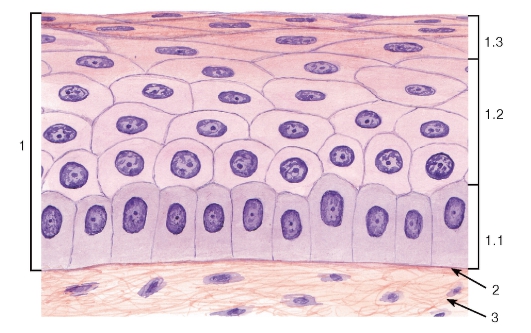
Рис. 39. Многослойный плоский неороговевающий эпителий (роговица)
Окраска: гематоксилин-эозин
1 - эпителий: 1.1 - базальный слой, 1.2 - промежуточный слой, 1.3 - поверхностный слой; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань
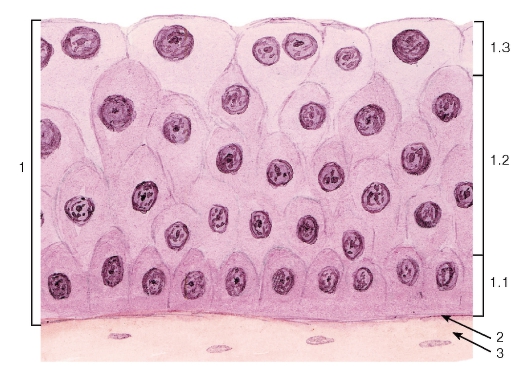
Рис. 40. Переходный эпителий - уротелий (мочевой пузырь, мочеточник)
Окраска: гематоксилин-эозин
1 - эпителий: 1.1 - базальный слой, 1.2 - промежуточный слой, 1.3 - поверхностный слой; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань
Железистые эпителии
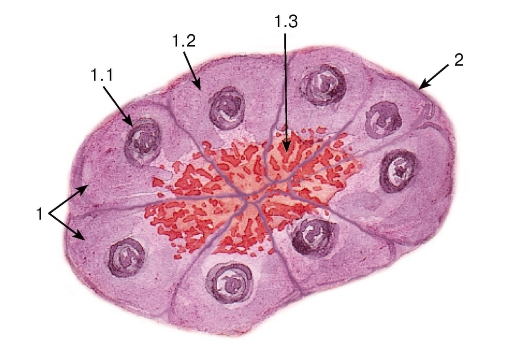
Рис. 41. Мерокринный тип секреции
(концевой отдел поджелудочной железы - ацинус)
Окраска: гематоксилин-эозин
1 - секреторные (ацинарные) клетки - панкреатоциты: 1.1 - ядро, 1.2 - базофильная зона цитоплазмы, 1.3 - оксифильная зона цитоплазмы с гранулами секрета; 2 - базальная мембрана
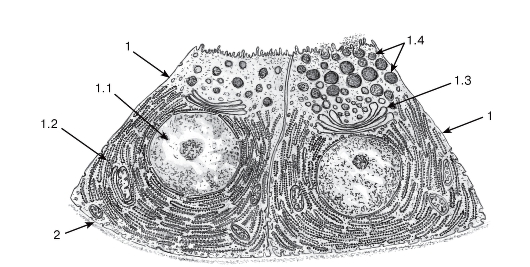
Рис. 42. Ультраструктурная организация железистых клеток при мерокринном типе секреции (участок концевого отдела поджелудочной железы - ацинуса)
Рисунок с ЭМФ
1 - секреторные (ацинарные) клетки - панкреатоциты: 1.1 - ядро, 1.2 - гранулярная эндоплазматическая сеть, 1.3 - комплекс Гольджи, 1.4 - гранулы секрета; 2 - базальная мембрана
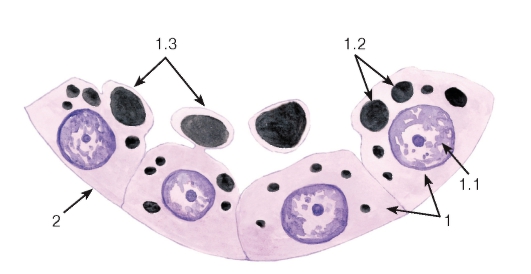
Рис. 43. Апокринный тип секреции (альвеола лактирующей молочной железы)
Окраска: судан черный-гематоксилин
1 - секреторные клетки (галактоциты): 1.1 - ядро, 1.2 - липидные капли; 1.3 - апикальная часть с отделяющимся от нее участком цитоплазмы; 2 - базальная мембрана
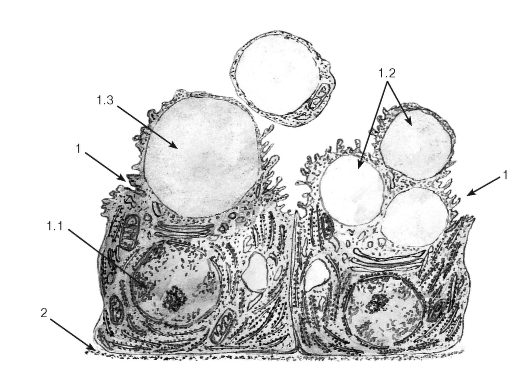
Рис. 44. Ультраструктурная организация железистых клеток при апокринном типе секреции (участок альвеолы лактирующей молочной железы)
Рисунок с ЭМФ
1 - секреторные клетки (галактоциты): 1.1 - ядро; 1.2 - липидные капли; 1.3 - апикальная часть с отделяющимся от нее участком цитоплазмы; 2 - базальная мембрана
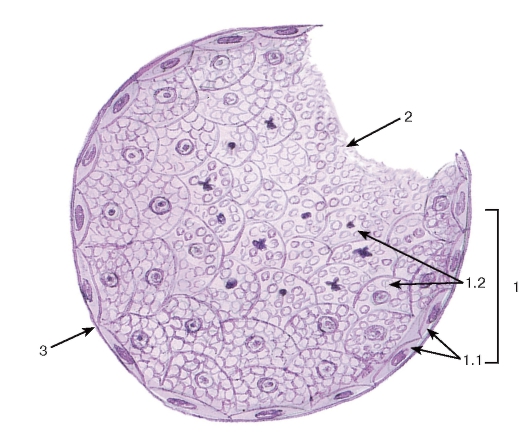
Рис. 45. Голокринный тип секреции (сальная железа кожи)
Окраска: гематоксилин-эозин
1 - клетки железы (себоциты): 1.1 - базальные (камбиальные) клетки, 1.2 - клетки железы на разных стадиях превращения в секрет, 2 - секрет железы; 3 - базальная мембрана
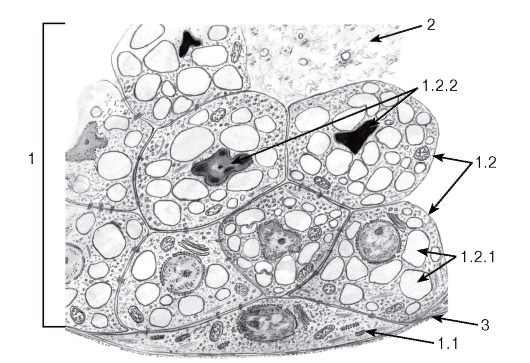
Рис. 46. Ультраструктурная организация железистых клеток при голокринном типе секреции (участок сальной железы кожи)
Рисунок с ЭМФ
1 - клетки железы (себоциты): 1.1 - базальная (камбиальная) клетка, 1.2 - клетки железы на разных стадиях превращения в секрет, 1.2.1 - липидные капли в цитоплазме, 1.2.2 - ядра, претерпевающие пикноз;
2 - секрет железы; 3 - базальная мембрана
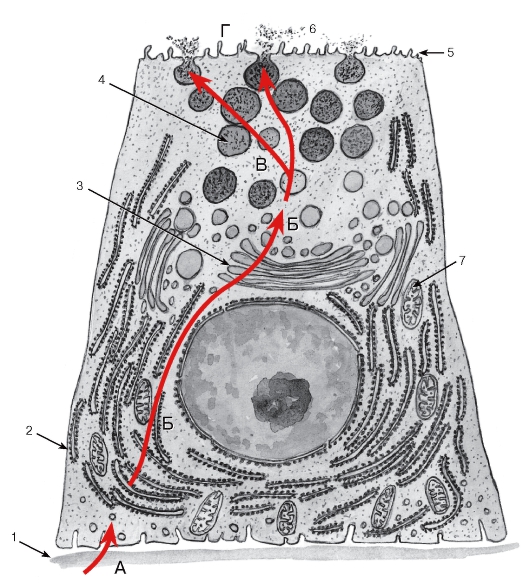
Рис. 47. Структурно-функциональная организация экзокринной железистой клетки в процессе синтеза и выделения белкового секрета
Схема по ЭМФ
А - фаза поглощения клеткой исходных веществ, которые приносятся кровью и транспортируются через базальную мембрану (1); Б - фаза синтеза секрета обеспечивается гранулярной эндоплазматической сетью (2) и комплексом Гольджи (3); В - фаза накопления секрета в виде секреторных гранул (4); Г - фаза выделения секрета через апикальную поверхность клетки (5) в просвет концевого отдела (6). Энергия, необходимая для обеспечения всех указанных процессов, вырабатывается многочисленными митохондриями (7)
Последовательность процессов (фаз) показана красными стрелками
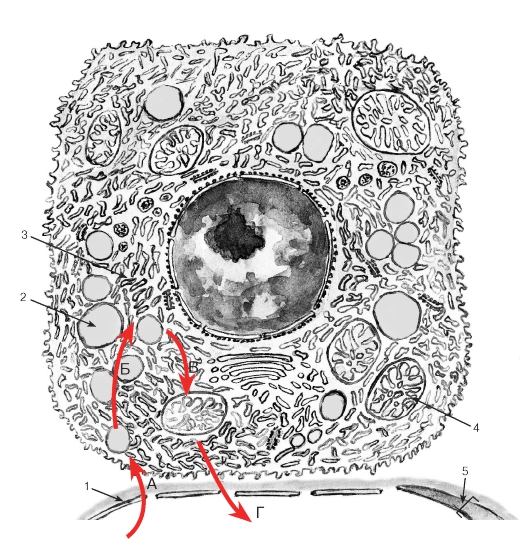
Рис. 48. Структурно-функциональная организация эндокринной железистой клетки в процессе синтеза и выделения стероидных гормонов
Схема по ЭМФ
А - фаза поглощения клеткой исходных веществ, которые приносятся кровью и транспортируются через базальную мембрану (1); Б - фаза депонирования в цитоплазме липидных капель (2), содержащих субстрат (холестерол) для синтеза стероидных гормонов; В - фаза синтеза стероидного гормона обеспечивается гладкой эндоплазматической сетью (3) и митохондриями с тубулярно-везикулярными кристами (4); Г - фаза выделения секрета через базальную поверхность клетки и стенку кровеносного сосуда (5) в кровь. Энергия, необходимая для обеспечения всех указанных процессов, вырабатывается многочисленными митохондриями (4)
Последовательность процессов (фаз) показана красными стрелками