Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. - 296 с. :ил.
|
|
|
|
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
(ткани внутренней среды)
Соединительные ткани, или ткани внутренней среды, представляют собой группу тканей с разнообразными морфофункциональными характеристиками, которые образуют внутреннюю среду организма и поддерживают ее постоянство. Эти ткани никогда непосредственно не граничат с внешней средой и полостями тела.
Общие признаки соединительных тканей: 1) развитие в эмбриональном периоде из общего источника - мезенхимы, которая является полипотентным (образующим ряд тканей) и гетерогенным (состоящим из разных по происхождению клеток) зачатком (см. рис. 49), 2) высокое содержание межклеточного вещества.
Функции соединительных тканей разнообразны. Наиболее общая функция всех соединительных тканей - поддержание постоянства внутренней среды организма (гомеостатическая). Она включает ряд частных функций, к которым относятся: трофическая (обеспечение других тканей питательными веществами); дыхательная (обеспечение газообмена в других тканях);
регуляторная (влияние на деятельность других тканей посредством биологически активных веществ и контактных взаимодействий); защитная (обеспечение разнообразных защитных реакций); транспортная (обусловливает все предыдущие, так как обеспечивает перенос питательных веществ, газов, регуляторных веществ, защитных факторов и клеток); опорная, механическая - формирование стромы (поддерживающих и опорных элементов для других тканей) и капсул различных органов, а также образование (в качестве функционально ведущих тканей) органов, выполняющих роль опорных и защитных элементов в организме (сухожилий, связок, хрящей, костей).
Классификация соединительных тканей выделяет пять подгрупп:
(1) Кровь, лимфа - своеобразные соединительные ткани с жидким межклеточным веществом (плазмой), в котором находятся клетки (лейкоциты) и постклеточные структуры (эритроциты, тромбоциты). Эти ткани выполняют ряд функций, связанных с транспортом веществ, дыханием и защитными реакциями.
КЛАССИФИКАЦИЯ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ

(2) Кроветворные ткани (лимфоидная, миелоидная) обеспечивают процессы гемоцитопоэза - постоянного образования форменных элементов крови, возмещающего их естественную убыль.
(3) Волокнистые соединительные ткани (собственно соединительные ткани) - наиболее типичные представители данной группы тканей, в межклеточном веществе которых ярко выражен волокнистый компонент. Подразделяются на несколько видов в зависимости от относительного объема, занимаемого в ткани волокнами, и их ориентации.
(4) Соединительные ткани со специальными свойствами (жировая, ретикулярная, пигментная, слизистая) - выполняют разнообразные специализированные функции в организме. Частично сходны по строению с волокнистыми соединительными тканями, однако характеризуются резким преобладанием специфических клеток (например, жировая и пигментная ткани) или неволокнистых компонентов межклеточного вещества (слизистая ткань).
(5) Скелетные соединительные ткани (хрящевые и костные) - характеризуются плотным и прочным межклеточным веществом (обызвествленным в костных тканях), обеспечивающим их высокие механические свойства, благодаря которым они выполняют опорную функцию по отношению к организму в целом (в составе скелета) или некоторым органам (входя в их состав).
Кровь и кроветворные ткани
Кровь - своеобразная жидкая ткань, относящаяся к группе тканей внутренней среды, которая циркулирует в сосудах благодаря ритмическим сокращениям сердца. В состав крови входят (1) форменные элементы (эритроциты, лейкоциты и тромбоциты) и (2) плазма крови - жидкое межклеточное вещество, содержащее ряд неорганических ионов и органических веществ (белков, углеводов, липидов). Из форменных элементов только лейкоциты являются истинными клетками; эритроциты и тромбоциты человека относятся к постклеточным структурам.
Функции крови. Наиболее общая функция - транспортная (перенос разнообразных веществ) - включает ряд частных функций: дыхательную (перенос газов), трофическую (перенос питательных веществ), экскреторную (удаление из тканей продуктов метаболизма), регуляторную (перенос гормонов, факторов роста и других биологически активных регуляторных веществ), терморегуляторную (распределение тепла между органами и его выделение во внешнюю среду). Гомеостатическая функ-
ция крови обеспечивает поддержание постоянства внутренней среды организма. Защитная функция направлена на нейтрализацию чужеродных антигенов, обезвреживание микроорганизмов неспецифическими и специфическими (иммунными) механизмами.
Количественные показатели содержания форменных элементов, регистрируемые при анализах крови, включают концентрации форменных элементов, которые учитывают в гемограмме, а также лейкоцитарную формулу.
Концентрации форменных элементов определяют при анализе крови в расчете на 1 мкл (1 мм3) или 1 л крови с помощью специальных счетных камер или автоматических анализаторов. Результаты анализа записывают в виде гемограммы (см. с. 62).
Лейкоцитарная формула определяется на мазках крови путем дифференциального подсчета количества различных лейкоцитов. Результаты регистрируются в виде таблицы, в которой содержание клеток каждого вида представлено в процентах по отношению к общему количеству лейкоцитов, принятому за 100 % (см. с. 62).
Морфологические особенности форменных элементов хорошо выявляются на мазках (рис. 50), в которых они распластываются по поверхности стекла и обычно имеют несколько большие размеры, чем на срезах. Мазки окрашивают специальными смесями красителей (метиленового синего, азура и эозина). В нашей стране наибольшее распространение получил вариант такой окраски по Романовскому-Гимзе.
Эритроциты - наиболее многочисленные форменные элементы крови - образуются в миелоидной ткани в красном костном мозгу (рис. 56, 57, 68); у человека и млекопитающих они относятся к постклеточным структурам, поскольку в ходе развития утрачивают ядро и органеллы.
Морфологическая характеристика эритроцитов. Эритроциты окрашиваются оксифильно и имеют вид двояковогнутого диска диаметром 7,2-7,5 мкм, что определяет более светлую окраску их центральной части по сравнению с периферической (см. рис. 50). Благодаря такой форме они имеют большую поверхность, активно насыщаются кислородом, способны к обратимой деформации. Форма эритроцитов поддерживается деятельностью ионных насосов в их плазмолемме, а также особыми элементами цитоскелета. Изменения формы эритроцитов возникают при их старении и в патологических условиях. При электронной микроскопии определяется высокая плотность цитоплазмы эритроцитов, содержащей кислород-связывающий пигмент гемоглобин в виде мелких гранул. Помимо зрелых эритроцитов,
в кровотоке в небольшом количестве обнаруживаются ретикулоциты - молодые формы эритроцитов, частично сохранившие органеллы, которые выявляются в виде базофильной сеточки (см. рис. 57).
Функции эритроцитов осуществляются только внутри сосудистого русла и включают дыхательную (благодаря высокому содержанию в них гемоглобина) и регуляторную (обусловлена способностью переносить на своей поверхности биологически активные вещества).
Тромбоциты (кровяные пластинки) образуются в миелоидной ткани в красном костном мозгу в результате фрагментации периферических участков цитоплазмы мегакариоцитов (рис. 58) и относятся к постклеточным структурам.
Морфологическая характеристика тромбоцитов. Тромбоциты - мелкие дисковидные двояковыпуклые безъядерные структуры диаметром 2-4 мкм, циркулирующие в крови. На мазках крови тромбоциты часто обнаруживаются в виде скоплений; в них выявляется светлая прозрачная наружная часть - гиаломер и центральная окрашенная часть, содержащая азурофильные гранулы, - грануломер (см. рис. 50). При электронной микроскопии в тромбоцитах обнаруживаются гранулы нескольких типов, митохондрии, мощно развитый цитоскелет и системы мембранных трубочек и канальцев. Гранулы тромбоцитов содержат факторы свертывания, факторы роста, АДФ, АТФ, ионы, гистамин.
Функции тромбоцитов осуществляются как внутри сосудистого русла, так и вне его: повреждение стенки сосуда вызывает активацию тромбоцитов с изменением их формы, адгезией в области повреждения, агрегацией (слипанием друг с другом) и секреторной реакцией, в результате чего развиваются реакции гемостаза (остановки кровотечения) и гемокоагуляции (свертывания крови). Выделившиеся факторы роста способствуют регенерации сосудистой стенки, по завершении которой тромб лизируется.
Лейкоциты (белые кровяные клетки) представляют собой группу морфологически и функционально разнообразных подвижных форменных элементов, которые циркулируют в крови, а в дальнейшем мигрируют через стенку мелких сосудов в соединительную ткань, где они участвуют в различных защитных реакциях. Таким образом, лейкоциты осуществляют свои функции за пределами сосудистого русла.
Классификация лейкоцитов основана на присутствии в их цитоплазме специфических гранул. По этому признаку все лейкоциты разделяют на гранулоциты и агранулоциты. В зависимости от окраски специфических гранул гранулоциты подразделя-
ют на базофильные, эозинофильные (ацидофильные) и нейтрофильные. Помимо специфических гранул в цитоплазме гранулоцитов содержатся неспецифические, или азурофильные, гранулы, которые представляют собой лизосомы. Ядро гранулоцитов обычно дольчатое (сегментированное), менее зрелые их формы имеют палочковидное ядро. Агранулоциты содержат в цитоплазме лишь неспецифические (азурофильные) гранулы. Их ядро обычно имеет округлую или бобовидную форму. К агранулоцитам относятся моноциты и лимфоциты.
Нейтрофильные гранулоциты (нейтрофилы) - наиболее распространенный вид лейкоцитов и, в частности, гранулоцитов. Они попадают в кровь из красного костного мозга, где в миелоидной ткани происходит их образование (рис. 59).
Морфологическая характеристика нейтрофильных гранулоцитов. Размеры нейтрофильных гранулоцитов на мазках варьируют в пределах 10- 15 мкм и примерно в 1,5 раза превышают размеры эритроцитов (см. рис. 50). Ядро нейтрофильных гранулоцитов имеет неодинаковое строение в клетках разной степени зрелости. В сегментоядерных нейтрофильных гранулоцитах (наиболее зрелых и численно преобладающих) ядро интенсивно окрашено и обычно содержит 3-4 сегмента, соединенных узкими перетяжками (см. рис. 50 и 51). В менее зрелых палочкоядерных нейтрофильных гранулоцитах оно не сегментировано или содержит лишь намечающиеся перетяжки, в наименее зрелых и самых малочисленных юных нейтрофильных гранулоцитах (метамиелоцитах) ядро бобовидной формы. Цитоплазма нейтрофильных гранулоцитов на светооптическом уровне слабооксифильна. Она содержит сравнительно многочисленные (50- 200 в каждой клетке) цитоплазматические гранулы двух основных типов. Первичные (азурофильные, или неспецифические) гранулы - относительно крупные и соответствуют выявляемой на светооптическом уровне зернистости, которая окрашивается азуром в розово-фиолетовый цвет. Вторичные (специфические нейтрофильные) гранулы, численно преобладающие в зрелых клетках - мелкие, плохо выявляются под световым микроскопом. Гранулы обоих типов содержат широкий спектр антимикробных веществ, активных в нейтральной и кислой среде.
Функции нейтрофильных гранулоцитов: уничтожение микроорганизмов после их захвата (фагоцитоза) или внеклеточно нефагоцитарным механизмом - путем выделения антимикробных веществ в межклеточное пространство; разрушение и переваривание поврежденных клеток и тканей; участие в регуляции деятельности других клеток (благодаря выработке ряда цитокинов).
Базофильные гранулоциты (базофилы) - самая малочисленная группа лейкоцитов и, в частности, гранулоцитов. Они образуются в миелоидной ткани в красном костном мозгу (рис. 61), откуда попадают в кровь. По морфологическим и функциональным свойствам они близки, но не идентичны тучным клеткам соединительной ткани.
Морфологическая характеристика базофильных гранулоцитов. Размеры базофильных гранулоцитов на мазках составляют 9-12 мкм, т. е. примерно соответствуют размерам нейтрофилов или несколько меньше их. Их ядра - дольчатые или S-образные, нередко маскируются ярко окрашенными цитоплазматическими гранулами (см. рис. 50), которые подразделяются на два типа - специфические и азурофильные (рис. 52). Специфические базофильные гранулы - крупные, хорошо видны в световой микроскоп, окрашиваются метахроматически - с изменением оттенка основного красителя вследствие высокого содержания сульфатированных гликозаминогликанов. На электронно-микроскопическом уровне их матрикс варьирует по плотности (см. рис. 52). Эти гранулы содержат гепарин, гистамин, ферменты, хемотаксические факторы.
Функции базофильных гранулоцитов связаны с выделением биологически активных веществ, содержащихся в их гранулах или вновь синтезируемых при активации. При выделении небольших количеств этих веществ базофилы выполняют регуляторную (гомеостатическую) функцию; при массивной (анафилактической) дегрануляции развиваются аллергические реакции, протекающие с сокращением гладких мышечных клеток, расширением сосудов, повышением их проницаемости и повреждением тканей.
Эозинофильные гранулоциты (эозинофилы) образуются в миелоидной ткани в красном костном мозгу (рис. 60), откуда попадают в кровь.
Морфологическая характеристика эозинофильных гранулоцитов. Размеры эозинофильных гранулоцитов на мазках больше, чем нейтрофильных (см. рис. 50) и составляют 12-17 мкм. Их ядро состоит из двух, реже - трех сегментов. Они легко узнаются на мазках благодаря многочисленным крупным специфическим эозинофильным гранулам, заполняющим их цитоплазму вместе с азурофильными гранулами. Под электронным микроскопом эозинофильные гранулы обычно имеют овальную форму и содержат плотное кристаллоидное тело (рис. 53). В специфических гранулах накапливаются белки, обладающие антимикробным и антипаразитарным действием, а также инактивирующие некоторые биологически активные соединения (гистамин, гепарин, простагландины).
Функции эозинофильных гранулоцитов: защитная - внутриклеточное и внеклеточное уничтожение микроорганизов, паразитов (гельминтов и простейших); иммунорегуляторная - ограничение области аллергической реакции путем инактивации ее медиаторов, а также выработка ряда медиаторов воспаления и цитокинов.
Моноциты относятся к агранулоцитам. Они образуются в миелоидной ткани в красном костном мозгу (рис. 62), из которого попадают в кровь; после циркуляции в кровеносном русле перемещаются из него в ткани, где превращаются в различные виды макрофагов.
Морфологическая характеристика моноцитов.
На мазках крови моноциты являются самыми крупными клетками среди лейкоцитов (см. рис. 50) и имеют округлую форму; под электронным микроскопом обнаруживаются различные цитоплазматические выпячивания. Ядро моноцитов - крупное (занимает до половины площади клетки на мазке), эксцентрично расположенное, бобовидной или подковообразной формы, светлое, с одним или несколькими мелкими ядрышками. Цитоплазма - слабобазофильная, содержит развитые органеллы (рис. 55) и азурофильные гранулы.
Функции моноцитов связаны с их превращением в макрофаги после миграции из сосудов в ткани. К ним относятся: обеспечение реакций неспецифической защиты организма против микробов, опухолевых и зараженных вирусами клеток; участие в специфиче ских (иммунных) защитных реакциях; захват и внутриклеточное переваривание различных стареющих и погибших клеток, а также их фрагментов и компонентов межклеточного вещества; секреция ферментов, цитокинов, факторов роста.
Лимфоциты относятся к агранулоцитам; источником их развития служат миелоидная ткань (красный костный мозг) и лимфоидная ткань (лимфоидные органы) (рис. 63), из которых они попадают в кровь и лимфу. Большая часть этих клеток после циркуляции в крови проникают из сосудов в различные ткани, впоследствии вновь возвращаясь в кровь (явление рециркуляции).
Морфологическая характеристика лимфоцитов. Лимфоциты по размерам подразделяют на малые (наиболее зрелые и многочисленные по сравнению с другими лимфоцитами крови), средние (менее зрелые) и большие (наименее зрелые).
Малые лимфоциты по размерам сравнимы с эритроцитами (см. рис. 50), их ядро - темное, круглое, овальное или бобовидное, на мазке занимает большую часть клетки (см. рис. 50 и 54). Цитоплазма окружает ядро узким ободком, окрашивается резко базофильно, содержит слабо развитые органеллы и азурофильные гранулы.
Средние лимфоциты крупнее малых и сходны с ними морфологически, однако их ядро светлее, а цитоплазма развита значительнее и занимает больший объем в клетке (см. рис. 50, 63, 64).
Большие лимфоциты со светлым ядром и развитой цитоплазмой обычно отсутствуют в крови (за одним исключением - см. ниже). В значительном количестве большие лимфоциты встречаются лишь в лимфоидной ткани, где они обычно являются активно делящимися (бластными) формами развивающихся клеток лимфоидного ряда - лимфобластами или иммунобластами (см. рис. 56, 63, 64, 67).
Большие гранулярные лимфоциты - особая разновидность больших лимфоцитов, циркулирующих в крови, которые выполняют функцию натуральных киллеров (НК-клеток) - одного из видов эффекторных клеток иммунной системы (см. рис. 56). Они характеризуются бобовидным ядром и светлой цитоплазмой с крупными азурофильными гранулами, содержимое которых обеспечивает цитотоксическую активность этих клеток.
Лимфоциты, сходные морфологически, могут выполнять неодинаковые функции и различаются экспрессией маркеров на своей поверхности, выявляемых специальными иммуноцитохимическими методами. По этим признакам, а также по месту дифференцировки и характеру рецепторов антигенов выделяют Т-лимфоциты и В-лимфоциты (см. ниже).
Функции лимфоцитов. Лимфоциты являются главными клетками иммунной системы и обеспечивают реакции специфического иммунитета (иммунная функция), защищающие организм от чужеродных антигенов (а также измененных собственных). Механизмы взаимодействия лимфоцитов различных видов между собой и с другими клетками в ходе реализации иммунного ответа схематично представлены на рис. 67. Регуляторная функция лимфоцитов отражает их способность регулировать деятельность клеток других типов в иммунных реакциях, процессах роста, дифференцировки и регенерации тканей посредством контактных взаимодействий и секреции цитокинов.
Основные закономерности кроветворения схематично представлены на рис. 56, который отражает общепризнанную ныне унитарную теорию крове творения. На основании способности к самообновлению, клеточному делению и образованию различных форменных элементов клетки, участвующие в процессе кроветворения, можно разделить на семь классов:
I класс - плюрипотентные гемопоэтические стволовые клетки, или стволовые клетки крови, которые могут образовывать любые форменные
элементы и обладают способностью к самообновлению. По строению эти клетки напоминают малые лимфоциты и могут быть идентифицированы иммуноцитохимически по набору антигенов на клеточной поверхности. У взрослого человека они сосредоточены преимущественно в красном костном мозгу, однако обнаруживаются в крови, циркулируя в которой они попадают в другие органы кроветворения.
II класс - частично детерминированные полипотентные родоначальные клетки, которые способны к ограниченному самоподдержанию и дают начало форменным элементам нескольких (но не всех) видов. Родоначальные клетки, относящиеся к данному и следующему (III) классам, называют также колониеобразующими единицами (КОЕ) или клетками, формирующими колонии (КФК). К этому классу относят родоначальные клетки лимфоцитопоэза (КОЕ-лимфоцитов) и миелопоэза (КОЕ-гранулоцитов, эритроцитов, моноцитов и мегакариоцитов).
III класс - унипотентные (коммитированные) родоначальные клетки детерминированы в направлении развития только одного вида форменных элементов (за исключением КОЕ-гранулоцитов и моноцитов). Они обладают низким потенциалом самоподдержания. Эти клетки, как и клетки предшествующих классов, не идентифицируются морфологически и внешне сходны с малыми лимфоцитами.
К ним относят родоначальные клетки эритроцитов, мегакариоцитов, базофилов, эозинофилов, КОЕ-нейтрофилов и моноцитов, дающие родоначальные клетки нейтрофилов и моноцитов, а также коммитированные клетки лимфоцитопоэза - про-В-лимфоциты и протимоциты.
IV класс - морфологически распознаваемые предшественники - бластные формы, которые представляют отдельные линии развития форменных элементов. Пролиферативная активность этих клеток ограничена; способностью к самоподдержанию они не обладают.
V и VI классы - созревающие (дифференцирующиеся) пролиферирующие и неделящиеся клетки. Эти клетки подвергаются структурной и функциональной дифференцировке, образуя соответствующий вид форменных элементов, в ходе которой они (за исключением лимфоцитов и моноцитов) утрачивают способность к делению.
VII класс - зрелые (дифференцированные) форменные элементы, циркулирующие в крови. Они неспособны к делению (за исключением лимфоцитов и моноцитов). Форменные элементы крови в тканях включают лейкоциты, которые покидают сосудистое русло и мигрируют в ткани, где выполняют свои функции.
Структурные преобразования клеток в ходе гемоцитопоэза (начиная со стадии морфологически распознаваемых предшественников) представлены на рис. 57-63.
Эритроцитопоэз (эритропоэз) - процесс образования и созревания эритроцитов - происходит в миелоидной ткани. Он включает (см. рис. 57):
(1) уменьшение размеров клеток по мере их созревания; (2) изменение окраски цитоплазмы от интенсивно базофильной в базофильном эритробласте (в связи с большим числом полирибосом) до оксифильной (обусловленной накоплением синтезированного гемоглобина) в ортохроматофильном (ацидофильном) эритробласте, через стадию полихроматофильного эритробласта; (3) постепенное снижение содержания и в конечном итоге утрату всех органелл (их остатки сохраняются в ретикулоците); (4) снижение, а в дальнейшем - утрату способности к делению (на стадии ортохроматофильного эритробласта; (5) конденсацию ядра и его последующее удаление из клетки (в конце стадии ортохроматофильного эритробласта).
Тромбоцитопоэз - процесс образования и созревания тромбоцитов - происходит в миелоидной ткани. Первой морфологически распознаваемой клеткой тромбоцитопоэза служит мегакариобласт, который, делясь и дифференцируясь, превращается в промегакариоцит - клетку с диплоидным бобовидным ядром (см. рис. 58). В этой клетке по мере ее преобразования в мегакариоцит происходит активная полиплоидизация с последующим эндомитозом и образованием многочисленных связанных перемычками долей ядра. Клетка резко увеличивается в объеме, краевая (периферическая) зона ее цитоплазмы накапливает гранулы и пересекается демаркационными каналами (границами будущих тромбоцитов). Тромбоциты образуются в результате процесса частичной фрагментации этой зоны цитоплазмы, которая может образовывать узкие длинные лентовидные отростки - филоподии (протромбоциты).
Гранулоцитопоэз - образование и дифференцировка гранулоцитов - происходит в миелоидной ткани. Процесс дифференцировки предшественников гранулоцитов в зрелые клетки включает (см. рис. 59-61): (1) уменьшение размеров клетки;
(2) снижение, а в дальнейшем (со стадии метамиелоцита) - утрату способности к делению; (3) изменение формы ядра - от округлой (в промиелоцитах и миелоцитах) до бобовидной (в метамиелоцитах) и палочковидной (в палочкоядерных клетках), его сегментацию (с образованием сегментоядерных гранулоцитов); нарастание конденсации ядерного хроматина; (4) выработку и накопление гранул
в цитоплазме с постепенным увеличением доли специфических гранул. Одновременно изменяются функциональные свойства клеток (нарастает подвижность, экспрессируются разнообразные рецепторы).
Моноцитопоэз - процесс развития моноцитов - происходит в миелоидной ткани. Преобразование монобластов в промоноциты и моноциты включает (см. рис. 62): (1) дальнейшее увеличение размеров клетки преимущественно за счет нарастания объема цитоплазмы, (2) снижение базофилии цитоплазмы, (3) накопление в ней азурофильных гранул, (4) изменение формы ядра, которое из округлого (в монобластах и промоноцитах) становится бобовидным (в моноцитах). Мигрируя в ткани, моноциты превращаются в различные виды макрофагов (вместе с которыми образуют единую моноцитарно-макрофагальную систему). При этом в клетке увеличивается содержание лизосом, митохондрий, пиноцитозных пузырьков и размер комплекса Гольджи, плазмолемма формирует многочисленные складки, микроворсинки, нарастает содержание на ней рецепторов, повышается подвижность клетки, ее метаболическая и фагоцитарная активность.
Лимфоцитопоэз - развитие лимфоцитов - происходит в миелоидной ткани в красном костном мозгу и лимфоидной ткани в различных лимфоидных органах и характеризуется их поэтапной миграцией (см. также раздел «Органы кроветворения и иммуногенеза»). Стадии морфологически распознаваемых предшественников клеток лимфоидного ряда соответствуют Т- и В-лимфобласты, которые образуются из унипотентных (коммитированных) родоначальных клеток - про-В-лимфоцитов и про-Т-лимфоцитов (протимоцитов) соответственно (см. рис. 56). Развитие Т- и В-лимфоцитов из родоначальных клеток связано с их пролиферацией и дифференцировкой и разделяется на две фазы: антиген-независимую и антиген-зависимую (см. рис. 56, 63 и 67).
1. Антиген-независимая фаза развития Т- и В-лимфоцитов (см. рис. 63) включает их пролиферацию, дифференцировку и селекцию и осуществляется в отсутствие антигенов в центральных (первичных) органах кроветворения и иммуногенеза - тимусе и красном костном мозгу соответственно, где эти клетки приобретают специфические (способные распознавать конкретный антиген) рецепторы. Морфологически этот процесс протекает как последовательное преобразование лимфобластов в незрелые (средние) лимфоциты и развитие из них зрелых (малых) лимфоцитов (см. рис. 63).
2. Антиген-зависимая фаза развития лимфоцитов происходит в периферических (вторичных) органах кроветворения и иммуногене-
за (лимфатических узлах, селезенке, миндалинах, пейеровых бляшках, аппендиксе и др.). Она осуществляется в результате взаимодействия антигенов со специфическими рецепторами на зрелых лимфоцитах, которое вызывает активацию лимфоцитов, их бласт-трансформацию (превращение в бластную форму - иммунобласт) и пролиферацию. Завершением этих процессов служит формирование эффекторных и регуляторных Т-лимфоцитов, плазматических клеток, а также Т- и В-клеток памяти.
Кроветворные (гемоцитопоэтические) ткани являются особыми видами соединительных тканей, или тканей внутренней среды, которые обеспечивают физиологическую регенерацию форменных элементов крови (гемоцитопоэз). Они представлены лимфоидной и миелоидной тканями (см. рис. 64 и 68). В состав каждой из этих тканей входят два компонента: (1) форменные элементы крови на различных стадиях развития (описание см. выше); (2) ретикулярная ткань.
Ретикулярная ткань относится к соединительным тканям со специальными свойствами, которая образует структурную основу (строму) кроветворных тканей и обеспечивает развитие форменных элементов крови в органах кроветворения и иммуногенеза (за исключением тимуса, в котором ее роль играет специализированная эпителиальная ткань). Компонентами ретикулярной ткани являются клетки и межклеточное вещество.
Ретикулярные клетки - крупные отростчатые, формирующие трехмерную сеть фибробластоподобные клетки с большим округлым центрально расположенным светлым ядром с крупным ядрышком и слабооксифильной цитоплазмой (см. рис. 64).
Межклеточное вещество ретикулярной ткани представлено ретикулярными волокнами (образованы коллагеном III типа), которые формируют разветвленную трехмерную сеть, оплетающую ретикулярные клетки, и основным аморфным веществом. Ретикулярные волокна не выявляются стандартными методами окраски, обладают аргирофилией и дают ШИК-реакцию.
Функция ретикулярной ткани - обеспечение процессов кроветворения путем создания необходимого микроокружения для развивающихся клеток крови - включает ряд частных функций: опорную, трофическую, секреторную, фагоцитарную.
Лимфоидная ткань (см. рис. 64) состоит из трехмерной сети, образованной ретикулярными клетками и волокнами (в тимусе - отростчатыми эпителиальными клетками), в петлях которой находятся лимфоциты на различных стадиях развития, плазматические клетки и макрофаги, а в периферических лимфоидных органах - также и дендритные
антиген-представляющие клетки. Лимфоидная ткань располагается в лимфоидных органах (органах иммунной системы) - тимусе, селезенке, лимфатических узлах, миндалинах, пейеровых бляшках, червеобразном отростке - и многочисленных лимфоидных образованиях, имеющихся в стенке органов различных систем.
Плазматические клетки (плазмоциты) - конечная стадия развития В-лимфоцитов (см. рис. 56, 65-67). Они обеспечивают синтез и секрецию иммуноглобулинов (антител), встречаются в периферических лимфоидных органах (см. рис. 64), рыхлой волокнистой (рис. 69) и миелоидной (рис. 68) ткани. На светооптическом уровне эти клетки характеризуются овальной или округлой формой, эксцентрическим расположением ядра со своеобразной картиной хроматина в виде «спиц колеса», выраженной базофилией цитоплазмы, кроме светлого околоядерного участка - «дворика» (см. рис. 65). Под электронным микроскопом в этих клетках выявляются многочисленные цистерны гранулярной эндоплазматической сети, которые заполняют большую часть цитоплазмы, за исключением области «дворика», занятой комплексом Гольджи и центриолями (см. рис. 66).
Иммуногенез - развитие и осуществление реакций клеточного и гуморального иммунитета, защищающих организм от чужеродных антигенов (а также измененных собственных), - соответствует антиген-зависимому этапу лимфоцитопоэза. Процессы иммуногенеза протекают преимущественно в лимфоидной ткани периферических органов иммунной системы, структурная организация которых обеспечивает оптимальные условия для эффективного взаимодействия клеток (см. рис. 67). Иммунные реакции влючают (1) контактное воздействие клеток-эффекторов иммунной системы на клетки-мишени, несущие чужеродные или измененные собственные антигены (клеточный иммунитет, обеспечивается Т-лимфоцитами), и (2) воздействие на мишени антител, продуцируемых плазмоцитами и переносимых кровью и тканевыми жидкостями (гуморальный иммунитет, обеспечивается В-лимфоцитами при взаимодействии с Т-лимфоцитами). Индукция иммунных реакций осуществляетсядендритными антиген-представляющими клетками, которые захватывают антигены и представляют их лимфоцитам в переработанном виде в комплексе с молекулами главного комплекса гистосовместимости II типа. С антиген-представляющими клетками взаимодействуют регуляторные Т-лимфоциты (Т-хелперы) и эффекторные Т-лимфоциты (Т-киллеры, или цитотоксические Т-клетки), которые подвергаются бласттрансформации и про-
лиферации с образованием популяций соответствующих регуляторных и эффекторных клеток, а также Т-клеток памяти. Т-хелперы играют основную роль в распознавании антигена, запуске реакций клеточного и гуморального иммунитета, регуляции взаимодействий Т-лимфоцитов друг с другом и с В-лимфоцитами, продукции лимфокинов. Основная их функция - стимулирующее (хелперное) влияние на эффекторные клетки. Они подразделяются на два подкласса: клетки Т-хелперы 1 типа отвечают преимущественно за реакции клеточного иммунитета и воспаления, а Т-хелперы 2 типа - стимулируют реакции гуморального иммунитета. Активированные Т-цитоксические клетки уничтожают зараженные вирусами или опухолевые клетки. Активированные антигенами В-лимфоциты при взаимодействии с Т-хелперами 2 типа подвергаются бласттрансформации и пролиферации с образованием популяций В-клеток памяти и плазматических клеток (см. рис. 67).
Миелоидная ткань образована ретикулярной тканью, в петлях которой располагаются гемопоэтические стволовые клетки и многочисленные развивающиеся форменные элементы крови, относящиеся ко всем ее росткам, поскольку в ней осуществляются процессы эритропоэза, тромбоцитопоэза, гранулоцитопоэза, моноцитопоэза и (частично) лимфоцитопоэза (см. рис. 68, а также 157, 158). Эритроциты развиваются в составе групп - эритробластических островков. Зрелые форменные элементы мигрируют в просвет особых кровеносных сосудов - синусоидов (венулярных синусов), по периферии которых располагаются макрофаги. Характерными компонентами миелоидной ткани являются жировые клетки - адипоциты. Миелоидная ткань входит в состав красного костного мозга.
Волокнистые соединительные ткани
Волокнистые соединительные ткани являются наиболее типичными представителями группы соединительных тканей, отчего их называют также собственно соединительными тканями. Как и другие ткани этой группы, они характеризуются высоким содержанием межклеточного вещества. В последнем значительное место занимают волокна (что отражено в наименовании этих тканей), которые выполняют важную функциональную роль; пространства между волокнами заполнены основным аморфным веществом.
Функции волокнистых соединительных тканей
включают все основные функции, свойственные соединительным тканям, наиболее важными из них являются: (1) трофическая, (2) регуляторная, (3) защитная и (4) опорная (механическая).
Классификация волокнистых соединительных тканей основана на соотношении клеток и межклеточного вещества, а также свойствах и особенностях организации (степени упорядоченности) последнего. В соответствии с классификацией выделяют рыхлую волокнистую соединительную ткань (см. рис. 69 и 71) и плотную волокнистую соединительную ткань (рис. 71-73).
1. Рыхлая волокнистая соединительная ткань характеризуется сравнительно невысоким содержанием волокон в межклеточном веществе, относительно большим объемом основного аморфного вещества, многочисленным и разнообразным клеточным составом.
2. Плотная волокнистая соединительная ткань отличается преобладанием в межклеточном веществе волокон при незначительном объеме, занимаемом основным аморфным веществом, относительно малочисленным и однообразным клеточным составом. Плотную волокнистую соединительную ткань, в свою очередь, подразделяют на:
(а) оформленную (в которой все волокна ориентированы в одном направлении);
(б) неоформленную (с различной ориентацией волокон).
Рыхлая волокнистая соединительная ткань является самым распространенным видом соединительных тканей (см. рис. 69) и выполняет все функции, свойственные соединительным тканям, взаимодействуя с другими тканями, связывая их между собой (что оправдывает общее название этой группы тканей) и способствуя поддержанию гомеостаза в организме. Эта ткань обнаруживается повсеместно, во всех органах - она образует их строму (основу), в частности, междольковые прослойки и прослойки между слоями и оболочками, заполняет пространства между функциональными элементами других тканей, сопровождает нервы и сосуды, входит в состав кожи и слизистых оболочек. Рыхлая волокнистая соединительная ткань содержит разнообразные клетки и межклеточное вещество, включающее волокна различных видов и основное аморфное вещество.
Клетки рыхлой волокнистой соединительной ткани представляют собой сложную гетерогенную популяцию функционально разнообразных и взаимодействующих между собою и с компонентами межклеточного вещества элементов.
Фибробласты - наиболее распространенные и функционально ведущие клетки рыхлой волокнистой соединительной ткани. Они вырабатывают (и частично разрушают) все компоненты межклеточного вещества (волокна и основное аморфное вещество), регулируют деятельность других клеток соединительных тканей. Зрелый
фибробласт - крупная отростчатая клетка с нерезкими границами и светлым ядром, содержащим мелкодисперсный хроматин и 1-2 ядрышка (см. рис. 69). Цитоплазма слабо базофильна и характеризуется диплазматической дифференцировкой - нерезким разделением на эндоплазму (внутреннюю, более плотную часть, окружающую ядро) и эктоплазму (периферическую, сравнительно светлую часть, образующую отростки). Эндоплазма содержит большую часть органелл мощно развитого синтетического аппарата, а также лизосомы, митохондрии; эктоплазма заполнена преимущественно элементами цитоскелета (рис. 70). Предшественниками фибробластов в ткани считают адвентициальные клетки - мелкие малодифференцированные веретеновидные уплощенные клетки, располагающиеся по ходу капилляров (см. рис. 69).
Конечной формой развития фибробласта служит фиброцит - узкая веретенообразная, неспособная к пролиферации клетка с длинными тонкими отростками, плотным ядром и слабо развитым синтетическим аппаратом. Фиброциты преобладают в плотной волокнистой соединительной ткани (см. рис. 71-73).
Макрофаги (гистиоциты) - вторые по численности (после фибробластов) клетки рыхлой волокнистой соединительной ткани - образуются из моноцитов после их миграции в соединительную ткань из просвета кровеносных сосудов (см. рис. 56 и 62). Морфологические признаки гистиоцитов зависят от их функциональной активности. Покоящиеся гистиоциты имеют вид мелких клеток с четкими контурами, небольшим темным ядром и плотной цитоплазмой. Активированные гистиоциты обладают изменчивой формой (см. рис. 69). Их ядро светлее, чем в покоящихся клетках, но темнее, чем в фибробластах. Цитоплазма с неровными краями содержит многочисленные крупные фаголизосомы, которые в виде вакуолей хорошо видны под световым микроскопом, придавая ей вспененный вид. (см. рис. 69). Ультраструктурная организация активированного гистиоцита характеризуется многочисленными выростами цитоплазмы и псевдоподиями, значительным числом лизосом, умеренно развитым комплексом Гольджи (см. рис. 70). Функции гистиоцитов: поглощение и переваривание поврежденных, зараженных, опухолевых и погибших клеток, компонентов межклеточного вещества, а также экзогенных материалов и микроорганизмов; индукция иммунных реакций (в качестве антиген-представляющих клеток); регуляция деятельности клеток других типов благодаря секреции цитокинов, факторов роста, ферментов.
Жировые клетки (адипоциты), согласно принятым представлениям, образуются из общих с фибробластами предшественников путем накопления липидных включений. Адипоциты - большие клетки сферической формы (в скоплениях деформируются, становясь многогранными) с уплощенным и смещенным к периферии ядром и почти целиком заполняющей цитоплазму, одной крупной, жировой каплей (по этой причине адипоциты белой жировой ткани называют однокапельными). Остальная часть цитоплазмы образует тончайший ободок, окружающий жировую каплю и расширяющийся до уплощенного полулуния в участке вокруг ядра (см. рис. 69 и 71). При стандартных методах обработки гистологического материала липиды, находящиеся в жировой капле, растворяются, в результате чего адипоцит приобретает вид пустого пузырька с тончайшим слоем цитоплазмы и уплощенным ядром. Для выявления липидов на гистологических препаратах используют специальные методы фиксации и проводки материала, обеспечивающие их сохранность, а также окраски срезов (наиболее часто - суданом черным или суданом III) - см. рис. 7. Жировые клетки являются нормальным компонентом рыхлой волокнистой соединительной ткани и в небольшом количестве встречаются в ней повсеместно. Ткань, в которой адипоциты являются структурно и функционально ведущими клеточными элементами, называют жировой и относят к одному из видов соединительных тканей со специальными свойствами (см. рис. 71).
Жировые клетки накапливают липиды, которые служат источником энергии в организме (трофическая функция), они также выделяют ряд цитокинов и других биологически активных пептидов - адипокинов, влияющих на другие клетки (регуляторная функция). Жировая ткань обеспечивает ряд дополнительных функций, к которым относятся: опорная, защитная и пластическая - она окружает различные органы и заполняет пространства между ними, защищая их от механических травм, служит опорным и фиксирующим элементом; теплоизолирующая - она препятствует чрезмерной потере тепла организмом; депонирующая - жировая ткань накапливает жирорастворимые витамины и стероидные гормоны (особенно эстрогены); эндокринная - жировая ткань синтезирует эстрогены и гормон, регулирующий потребление пищи - лептин.
Тучные клетки развиваются в тканях из предшественника, имеющего костномозговое происхождение. Это - клетки удлиненной или округлой формы, с овальным или округлым ядром, которое на светооптическом уровне часто прослеживается с
трудом, так как маскируется метахроматическими гранулами, лежащими в цитоплазме (см. рис. 69). При электронной микроскопии выявляются выросты цитоплазмы и микроворсинки, умеренно развитые синтетический аппарат и элементы цитоскелета, липидные капли, а также гранулы с морфологически вариабельным содержимым (см. рис. 70). Гранулы тучных клеток сходны по строению и составу с гранулами базофилов, но не идентичны им; в них содержатся: гепарин, гистамин, дофамин, хемотаксические факторы, гиалуроновая кислота, гликопротеины, фосфолипиды и ферменты. При активации эти клетки вырабатывают также простагландины, тромбоксан, простациклин и лейкотриены. При постепенном выделении небольших доз этих биологически активных веществ тучные клетки (как и базофилы) выполняют регуляторные функции, направленные на поддержание гомеостаза. Регуляторная функция тучных клеток связана также с выработкой ими цитокинов и факторов роста. При быстрой массивной (анафилактической) дегрануляции тучных клеток в ответ на антиген (аллерген) развиваются аллергические реакции, протекающие со спазмом гладких мышечных клеток, расширением сосудов, повышением их проницаемости, повреждением тканей. Клинические проявления массивной дегрануляции тучных клеток зависят от ее распространенности и локализации в организме и имеют различную степень тяжести вплоть до анафилактического шока и смерти. В тканях тучные клетки располагаются преимущественно около мелких сосудов - периваскулярно (см. рис. 69), что, вероятно, связано с их регуляторной функцией и влиянием на проницаемость сосудов.
Плазматические клетки (плазмоциты) и их предшественники - В-лимфоциты - в небольших количествах постоянно содержатся в различных участках рыхлой волокнистой соединительной ткани (см. рис. 69). Они имеют мелкие размеры, располагаются поодиночке или группами, и (как и в лимфоидной ткани) вырабатывают и выделяют антитела (иммуноглобулины), обеспечивая тем самым гуморальный иммунитет. Характерные морфологические и функциональные признаки плазмоцитов описаны ранее и показаны на рис. 65 и 66.
Дендритные антиген-представляющие клетки развиваются из предшественников костномозгового происхождения. Они встречаются в рыхлой волокнистой соединительной ткани, эпителиях, лимфоидной ткани (см. рис. 67), лимфе и крови. Эти клетки обладают высокой активностью захвата, процессинга и представления антигенов лимфоцитам, морфологически характеризуются отростчатой формой.
Лейкоциты (гранулоциты и агранулоциты) являются нормальными клеточными компонентами рыхлой волокнистой соединительной ткани (см. рис. 69), в которую они мигрируют из мелких сосудов, однако их содержание в ней в норме незначительно. Выделяя цитокины, эти клетки влияют друг на друга, остальные клетки соединительной ткани и на клетки соседних тканей. Локальное увеличение числа лейкоцитов в рыхлой волокнистой соединительной ткани выявляется при воспалении.
Пигментные клетки имеют нейральное происхождение и являются потомками клеток, выселившихся в эмбриональном периоде из нервного гребня. Они имеют отростчатую форму; их цитоплазма содержит пигмент меланин. В рыхлой волокнистой соединительной ткани человека и других млекопитающих пигментные клетки встречаются сравнительно редко. Численное преобладание этих клеток над другими клеточными элементами соединительной ткани характерно для радужки и сосудистой оболочки глаза. Такую ткань называют пигментной и относят к одному из видов соединительных тканей со специальными свойствами (см. выше).
Межклеточное вещество рыхлой волокнистой соединительной ткани состоит из волокон трех типов (коллагеновых, ретикулярных и эластических) и основного аморфного вещества.
Коллагеновые волокна образованы коллагеном I типа и состоят из фибрилл, которые выявляются только под электронным микроскопом. На гистологических препаратах коллагеновые волокна имеют вид оксифильных продольно исчерченных извитых тяжей, идущих в различных направлениях поодиночке и часто образующих пучки вариабельной толщины (см. рис. 71). Они хорошо выявляются при окраске железным гематоксилином (см. рис. 69). Коллагеновые волокна обеспечивают высокие механические свойства соединительной ткани, определяют ее архитектонику, связывают клетки с межклеточным веществом и отдельные компоненты последнего между собой; влияют на свойства клеток.
Ретикулярные волокна имеют малый диаметр и, как правило, формируют тонкие растяжимые трехмерные сети. Они образованы коллагеном III типа, не обнаруживаются при стандартных гистологических окрасках и требуют специальных методов окрашивания (солями серебра, ШИК- реакцией). Основная функция ретикулярных волокон - опорная. Они встречаются в рыхлой волокнистой соединительной ткани (особенно во вновь образованной или подвергающейся перестройке), а также во всех других видах соединительной
ткани. Ретикулярные волокна особенно многочисленны в кроветворных (миелоидной и лимфоидной) тканях.
Эластические волокна образованы белками эластином (преобладает и образует основу волокна) и фибриллином (располагается по периферии зрелого волокна). Они обладают способностью к обратимой деформации, придавая эластические свойства ткани. Эластические волокна тоньше коллагеновых, ветвятся и анастомозируют друг с другом, формируя трехмерные сети (см. рис. 69); в отличие от коллагеновых волокон, они обычно не образуют пучки. На светооптическом уровне они не выявляются стандартными методами окраски и обнаруживаются при использовании избирательных методов (чаще всего - орсеина, рис. 154), однако окрашиваются железным гематоксилином (см. рис. 69).
Основное аморфное вещество заполняет промежутки между волокнистыми компонентами межклеточного вещества и окружает клетки. При изучении под светооптическим и электронным микроскопами оно имеет аморфное строение, прозрачно, характеризуется слабой базофилией (см. рис. 69) и низкой электронной плотностью. На молекулярном уровне оно обладает сложной организацией и состоит из макромолекулярных гидратированных комплексов протеогликанов и структурных гликопротеинов.
Плотная волокнистая соединительная ткань характеризуется (1) очень высоким содержанием волокон (преимущественно коллагеновых), формирующих толстые пучки и занимающих основную часть объема ткани, (2) малым количеством основного аморфного вещества в составе межклеточного вещества, (3) сравнительно низким содержанием клеточных элементов и (4) преобладанием одного (главного) типа клеток - фиброцитов - над остальными (особенно в плотной оформленной ткани).
Главное свойство плотной волокнистой соединительной ткани - очень высокая механическая прочность - обусловлено присутствием мощных пучков коллагеновых волокон. Ориентация этих волокон соответствует направлению действия сил, вызывающих деформацию ткани.
Плотная волокнистая неоформленная соединительная ткань характеризуется расположением в трех различных плоскостях пучков коллагеновых волокон, которые переплетаются между собою, формируя трехмерную сеть (см. рис. 71). Содержание основного аморфного вещества невелико, клетки немногочисленны. Такая ткань образует капсулы различных органов и глубокий (сетчатый) слой дермы (см. рис. 71), в котором
эта ткань занимает основной объем (см. также рис. 177). В составе дермы, между слоем плотной волокнистой соединительной ткани и эпидермисом, располагается рыхлая волокнистая соединительная ткань, а глубже плотной волокнистой ткани находится жировая ткань, образующая гиподермис(см. рис. 71 и 177).
Плотная волокнистая оформленная соединительная ткань содержит толстые пучки коллагеновых волокон, располагающиеся параллельно друг другу (в направлении действия нагрузки), и небольшое количество основного аморфного вещества (рис. 72 и 73). Содержание клеток невелико; среди них подавляющее большинство составляют фиброциты. Описанное строение имеет ткань, образующая сухожилия, связки, фасции и апоневрозы.
Сухожилие как орган включает пучки коллагеновых волокон различных порядков с расположенными между ними фиброцитами и окружающие пучки оболочки (прослойки) из рыхлой и плотной неоформленной соединительных тканей. В сухожилии выделяют первичные, вторичные и третичные сухожильные пучки (см. рис. 72 и 73). Первичные сухожильные (коллагеновые) пучки располагаются между рядами фиброцитов. Вторичные сухожильные (коллагеновые) пучки образованы группой первичных пучков, окруженных снаружи оболочкой из рыхлой волокнистой неоформленной соединительной ткани - эндотендинием. Третичные сухожильные (коллагеновые) пучки состоят из нескольких вторичных пучков, которые окружены снаружи оболочкой из плотной волокнистой неоформленной соединительной ткани - перитендинием, отдающего вглубь сухожилия прослойки эндотендиния. Сухожилие в целом может представлять собой третичный пучок, в некоторых случаях оно складывается из нескольких третичных пучков, окруженный общей оболочкой - эпитендинием.
Скелетные соединительные ткани
Скелетные соединительные ткани включают хрящевые и костные ткани, объединенные в единую группу на основании ряда признаков: (1) общей функции - опорной; (2) общего источника развития в эмбриогенезе (мезенхимы); (3) сходства строения - и хрящевые, и костные ткани образованы клетками и преобладающим по объему межклеточным веществом, имеющим значительную механическую прочность, которое является функционально ведущим, так как обеспечивает выполнение этими тканями опорной функции.
Хрящевые ткани входят в состав органов дыхательной системы (носа, гортани, трахеи, брон-
хов), ушной раковины, суставов, межпозвонковых дисков; у плода ими образована значительная часть скелета. Хрящевая ткань играет важную роль и в обеспечении роста костей. Хрящевые ткани состоят из клеток (хондроцитов) и межклеточного вещества (хрящевого матрикса), образованного волокнами и основным аморфным веществом. В состав последнего входят протеогликаны, формирующие крупные агрегаты, и гликопротеины; характерно высокое содержание воды. Хрящевые ткани образуют структуры органного порядка - хрящи (см. ниже).
Классификация хрящевых тканей основана на особенностях строения и биохимического состава их межклеточного вещества и выделяет: (1) гиалиновую хрящевую ткань, (2) эластическую хрящевую ткань и (3) волокнистую (фиброзную) хрящевую ткань.
Гиалиновая хрящевая ткань является наиболее распространенным ее видом в организме человека. Она образует скелет у плода, вентральные концы ребер, хрящи носа, гортани (частично), трахеи и крупных бронхов, покрывает суставные поверхности. Хондроциты имеют овальную или сферическую форму и располагаются в полостях - лакунах поодиночке или (в глубине хряща) в виде изогенных групп (агрегатов хондроцитов), насчитывающих до 8-12 клеток (рис. 74). Межклеточное вещество (хрящевой матрикс) на гистологических препаратах кажется однородным; оно содержит коллаген II типа; протеогликаны, а также гликопротеины. На гистологических препаратах выявляется территориальный матрикс, который непосредственно окружает хрящевые клетки или их изогенные группы в виде округлого базофильного облачка, и интертерриториальный матрикс - наиболее старая часть межклеточного вещества со слабобазофильной или оксифильной окраской (см. рис. 74).
Эластическая хрящевая ткань образует хрящи, которые обладают гибкостью и способностью к обратимой деформации. Из нее состоят хрящи ушной раковины, наружного слухового прохода, евстахиевой трубы, надгортанника, некоторые хрящи гортани, а также хрящевые пластинки и островки средних бронхов. Хондроциты в этой ткани располагаются в лакунах, где они лежат поодиночке или в виде небольших (до четырех клеток) изогенных групп. Матрикс, помимо коллагена II типа, протеогликанов и гликопротеинов, содержит эластические волокна, образующие плотную сеть (рис. 75).
Волокнистая (фиброзная) хрящевая ткань образует хрящи, которые обладают значительной механической прочностью. Она обнаруживается в межпозвонковых дисках, лонном симфизе, участ-
ках прикрепления сухожилий и связок к костям или гиалиновым хрящам. Хондроциты в этой ткани имеют округлую или удлиненную форму и располагаются в лакунах поодиночке или в виде мелких изогенных групп, нередко выстраиваются в колонки вдоль пучков коллагеновых волокон (рис. 76). Матрикс, помимо коллагена II типа, протеогликанов и гликопротеинов, содержит большое количество коллагена I типа в виде коллагеновых волокон, которые часто располагаются параллельными пучками (см. рис. 76).
Хрящ как орган включает функционально ведущую и количественно преобладающую хрящевую ткань, которая образует два нерезко разграниченные слоя (зоны), а также покрывающую его снаружи соединительнотканную оболочку - надхрящницу (см. рис. 74).
Зона молодого хряща - сравнительно тонкая, расположена под надхрящницей и состоит из уплощенных хондроцитов, которые лежат поодиночке параллельно поверхности хряща и окружены гомогенным оксифильным матриксом.
Зона зрелого хряща располагается глубже предыдущей и представлена округлыми хондроцитами, собранными в изогенные группы и окруженными преимущественно базофильным матриксом, который разделяется на территориальный и интертерриториальный (см. рис. 74).
Надхрящница обеспечивает механическую связь хряща с другими структурами (сухожилиями, связками и др.), она содержит кровеносные сосуды (обеспечивает питание хряща), нервы и камбиальные элементы хрящевой ткани. В ее состав входят два слоя: наружный волокнистый (фиброзный) и внутренний хондрогенный (см. рис. 74).
Волокнистый слой - толстый, образован плотной волокнистой неоформленной соединительной тканью. Он обеспечивает механическую прочность надхрящницы, ее связь с другими структурами.
Хондрогенный слой - тонкий, состоит из рыхлой волокнистой соединительной ткани, среди клеток которой присутствуют малодифференцированные камбиальные клетки, способные дифференцироваться в хондробласты.
Костные ткани образуют скелет, который защищает внутренние органы от повреждений, входит в локомоторный аппарат и является важнейшим депо минеральных веществ в организме. Костная ткань образована клетками и обызвествленным межклеточным веществом - костным матриксом (рис. 77). Костные ткани образуются в результате процесса остеогистогенеза, или остео генеза, который начинается и наиболее активно протекает у эмбриона (эмбриональный остеогистогенез), продолжаясь и после рождения (постнатальный
остеогистогенез). Формирование костей (как органов) завершается, в среднем, к 25 годам, однако гистогенез костной ткани при этом не прекращается, поскольку у взрослого в физиологических условиях она подвергается постоянной внутренней перестройке.
Развитие костной ткани (остеогистогенез, или остеогенез) может происходить двумя путями: (1) непосредственно из мезенхимы или эмбриональной соединительной ткани (прямой остеогенез, или интрамембранозное окостенение); (2) на месте ранее образованной хрящевой модели кости (непрямой остеогенез, или хрящевое окостенение).
Прямой остеогенез характерен для развития грубоволокнистой костной ткани, образующей первоначально плоские кости черепа, ключицы, конечных фаланг пальцев. Он включает: 1) образование остеогенных островков - скоплений активно размножающихся клеток мезенхимы; 2) дифференцировку клеток остеогенных островков в остеобласты и образование ими органического матрикса кости (остеоида), главным компонентом которого является коллаген I типа; 3) обызвествление (минерализацию) остеоида остеобластами путем отложения кристаллов гидроксиапатита.
В ходе прямого остеогенеза формируются оксифильные костные трабекулы (балки), содержащие обызвествленный костный матрикс (см. рис. 77). На их поверхности находятся остеобласты, которые дифференцируются из клеток остеогенной (образующейся из мезенхимы) соединительной ткани. Они откладывают костный матрикс на поверхность балок, а затем погружаются в него и превращаются в остеоциты, тела которых располагаются в лакунах, а связывающие их отростки проходят в костных канальцах (часто не видны при стандартных окрасках). Трабекулы частично подвергаются разрушению вследствие деятельности остеокластов, которые формируют на их поверхности углубления - эрозионные, или резорбционные, лакуны (см. рис. 77).
Клетки костной ткани включают остеобласты, остеоциты и остеокласты (см. рис. 77 и 78).
Остеобласты синтезируют и секретируют неминерализованное межклеточное вещество (матрикс) кости (остеоид), участвуют в его обызвествлении, регулируют поток кальция и фосфора в костную ткань и из нее. А ктивные остеобласты - кубические или столбчатые клетки с округлым ядром с крупным ядрышком, базофильной цитоплазмой (см. рис. 77), образующей отростки, посредством которых эти клетки связаны с другими остеобластами и остеоцитами. На ультраструктурном уровне в остеобластах выяв-
ляется мощно развитый синтетический аппарат, большое число митохондрий, пузырьков, а на их поверхности - многочисленные микроворсинки (см. рис. 78). Неактивные (покоящиеся) остеобласты - клетки, выстилающие кость, - образуются из активных остеобластов и в покоящейся кости покрывают большую часть ее поверхности. Они имеют вид уплощенных клеток с веретеновидными (на срезе) ядрами и редуцированными органеллами.
Остеоциты - основной тип клеток зрелой костной ткани, которые поддерживают нормальное состояние костного матрикса. Они образуются из остеобластов, которые окружаются со всех сторон обызвествленным матриксом, уменьшаются в размерах, утрачивают способность к делению и к активной синтетической деятельности, теряют большую часть органелл. Уплощенные тела остеоцитов лишены полярности и находятся в узких костных полостях - лакунах, где они окружены коллагеновыми фибриллами и узкой полоской остеоида (см. рис. 77, 78 и 80). Отростки остеоцитов располагаются в узких костных канальцах и связывают соседние клетки благодаря щелевым соединениям между ними.
Остеокласты - подвижные многоядерные гигантские клетки, образующиеся вследствие слияния моноцитов, осуществляющие разрушение (резорбцию) костной ткани. Они располагаются в образованных ими углублениях на поверхности костной ткани - эрозионных (резорбционных) лакунах (см. рис. 77, 78). Остеокласты достигают крупных размеров и содержат до нескольких десятков ядер (на отдельном срезе обычно видна лишь их часть). Цитоплазма - ацидофильная, пенистая, с высоким содержанием лизосом, митохондрий, пузырьков (см. рис. 77 и 78). Ее участок, прилежащий к кости, образует многочисленные складки клеточной мембраны - микроскладчатую кайму (гофрированный край), в этой области происходит резорбция костной ткани по границе, известной как эрозионный (резорбционный) фронт. Процесс разрушения костного матрикса остеокластом включает закисление содержимого эрозионной лакуны, обусловливающее растворение минерального компонента матрикса, и разрушение его органических компонентов лизосомальными ферментами, выделяемыми в лакуну.
Развитие кости на месте хряща (ранее образованной хрящевой модели), или непрямой остеогенез, характерен для развития подавляющего большинства костей скелета человека. Первоначально формируется хрящевая модель будущей кости, которая служит основой для ее развития, а в дальнейшем она разрушается и замещается
костью. Непрямой остеогенез включает следующие стадии:
1. Образование хрящевой модели кости завершается формированием из мезенхимы покрытого надхрящницей гиалинового хряща, сходного по форме с будущей костью.
2. Образование перихондрального костного кольца (костной манжетки) начинается в середине диафиза хрящевой модели и распространяется к его краям; оно является результатом дифференцировки в надхрящнице остеобластов, которые вырабатывают костный матрикс и образуют вокруг хряща цилиндрическое костное кольцо (манжетку), нарушающее питание хряща и вызывающее в нем дистрофические изменения и кальцификацию (обызвествление).
3. Образование эндохондральной костной ткани происходит в результате проникновения остеогенных клеток внутрь обызвествленной хрящевой ткани диафиза вместе с врастающими в нее из надкостницы кровеносными сосудами. Эти клетки дифференцируются в остеобласты, образующие эндохондральную кость внутри разрушающегося хряща. В центральной части диафиза эндохондральная кость разрушается остеокластами, образуя костномозговую полость, которая заполняется красным костным мозгом. Эндохондральная кость сохраняется только в области зоны оссификации (линии окостенения) - зигзагообразной границы с кальцифицированным и разрушающимся хрящом, остатки которого она окружает. На рис. 79 представлена картина, соответствующая данной стадии непрямого остеогенеза.
Хрящевая ткань, взаимодействующая с надвигающейся на нее эндохондральной костной тканью, подвергается изменениям, разделяясь на четыре зоны. В направлении от эпифиза к диафизу описаны: (1) зона покоя (неизмененного хряща) - наиболее удаленная от эндохондральной кости; (2) зона пролиферации - содержит колонки (столбики) активно делящихся уплощенных хондроцитов; (3) зона гипертрофии - состоит из крупных округлых дегенеративно измененных пузырчатых хондроцитов; (4) зона кальцификации (обызвествленного хряща) - непрерывно разрушается и замещается разрастающейся эндохондральной костью (см. рис. 79).
4. Образование эндохондральной (энхондральной) кости в эпифизах и формирование эпифизарных пластинок роста. Образование эндохондральной кости в эпифизах приводит к тому, что неизмененный гиалиновый хрящ в области, прилежащей к диафизу (метафизе), формирует эпифизарную хрящевую пластинку роста. Рост кости в длину обеспечивается пролиферацией хондроцитов в этой пластинке с их дифференцировкой и образованием
матрикса, который постепенно кальцифицируется, разрушается и замещается со стороны диафиза эндохондральной костной тканью. Снижение, а в дальнейшем - прекращение пролиферации хондроцитов в эпифизарной хрящевой пластинке приводит к ее истончению и полному исчезновению с замещением костной тканью, связывающей диафиз с эпифизом. После этого дальнейший рост кости в длину прекращается.
Классификация костных тканей основана на различиях строения межклеточного вещества, в частности, степени упорядоченности расположения в нем коллагеновых волокон. Выделяют (1) грубо волокнистую костную ткань и (2) пластинчатую костную ткань.
Грубоволокнистая костная ткань (см. рис. 80) характеризуется неупорядоченным расположением коллагеновых волокон в матриксе. Она отличается относительно небольшой механической прочностью и обычно образуется тогда, когда остеобласты формируют остеоид с высокой скоростью (в костной ткани плода, при заживлении переломов). Лакуны остеоцитов, содержащие их тела, не имеют закономерной ориентации. В ходе нормального развития и при регенерации костной ткани грубоволокнистая костная ткань постепенно замещается пластинчатой. У взрослого она сохраняется лишь в заросших швах черепа и участках прикрепления некоторых сухожилий к костям.
Пластинчатая костная ткань у взрослого образует практически весь костный скелет. Ее минерализованный матрикс состоит из костных пластинок, образованных параллельно расположенными коллагеновыми волокнами. Лакуны остеоцитов, содержащие их тела, располагаются между пластинками упорядоченно, а костные канальцы с отростками остеоцитов пронизывают пластинки под прямыми углами.
Кость как орган обладает сложной архитектоникой и тканевым составом (см. рис. 81-83). Функционально ведущей тканью кости служит пластинчатая костная ткань, снаружи и со стороны костномозговой полости она покрыта соединительнотканными оболочками (более толстой надкостницей и тонким эндостом). Кость содержит костный мозг, кровеносные и лимфатические сосуды и нервы. В кости как органе различают компактное вещество и губчатое (трабекулярное) вещество, которые образованы пластинчатой костной тканью и плавно переходят друг в друга.
Компактное вещество (компактная кость) образует диафизы трубчатых костей (см. рис. 81 и 82) и наружный слой костной ткани всех других костей. Костные пластинки компактного вещества образуют следующие системы:
(1) Остеоны - цилиндрические структуры, расположенные вдоль длинной оси кости (см. рис. 81 и 82), являются морфофункциональными единицами компактной кости. Они состоят из концентрических костных пластинок, расположенных вокруг канала остеона (центрального канала), в котором проходят кровеносные сосуды, нервные волокна, окруженные небольшим количеством рыхлой волокнистой соединительной ткани, содержащей остеогенные клетки-предшественники (камбиальные элементы). Между пластинками остеона залегают лакуны остеоцитов; наружной границей остеона является цементирующая (спайная) линия, образованная преимущественно основным веществом и почти не содержащая волокон. Каналы остеонов сообщаются друг с другом, с надкостницей и костномозговой полостью за счет поперечно или косо идущих перфорирующих (фолькмановских) каналов, содержащих сосуды. В отличие от каналов остеона, эти каналы не окружены концентрически расположенными костными пластинками.
(2) Интерстициальные, или промежуточные (вставочные), пластинки заполняют пространства между остеонами и являются остатками ранее существовавших остеонов, разрушенных в процессе перестройки кости.
(3) Наружные и внутренние опоясывающие пластинки образуют самый наружный и самый внутренний слои компактного вещества кости и располагаются параллельно поверхности кости под надкостницей и эндостом соответственно.
Губчатое вещество (трабекулярная кость) состоит из трехмерной сети анастомозирующих костных трабекул, разделенных межтрабекулярными пространствами, содержащими костный мозг (см. рис. 83). Трабекулы губчатого вещества кости образованы параллельно лежащими костными пластинками неправильной формы, объединенными в трабекулярные пакеты (морфофункциональные единицы губчатого вещества).
Надкостница покрывает кость снаружи (см. рис. 81) и прочно прикреплена к ней толстыми пучками перфорирующих коллагеновых пучков (шарпеевских волокон), которые проникают и вплетаются в слой наружных опоясывающих пластинок кости.
В надкостнице имеются два слоя: наружный волокнистый (фиброзный) слой образован плотной волокнистой неоформленной соединительной тканью, внутренний остеогенный слой состоит из рыхлой волокнистой соединительной ткани, в которой располагаются камбиальные остеогенные клетки.
Функции надкостницы: трофическая - благодаря сосудам, которые проникают в кость; регенераторная - обусловлена наличием камбиальных элементов; механическая, опорная - обеспечи-
вает механическую связь кости с другими структурами (сухожилиями, связками, мышцами).
Эндост - тонкая выстилка кости со стороны костного мозга, аналогичная надкостнице, состоящая из непрерывного слоя плоских клеток. Содержит остеогенные клетки и остеокласты.
Соединения костей
Соединения костей разделяются на непрерывные - синартрозы, являющиеся неподвижными или малоподвижными, и прерывные - суставы, или диартрозы, обеспечивающие подвижность костей.
Непрерывные соединения костей (синартрозы)
разделяют в зависимости от характера ткани, осуществляющей связь между костями, на три типа:
1. Синдесмозы - соединения костей посредством плотной волокнистой соединительной ткани. У человека к таким соединениям относят межкостные перепонки, связывающие кости предплечья, голени, швы между костями черепа в период роста.
2. Синхондрозы - соединения костей посредством хрящевой ткани. Примерами таких соединений служат реберно-грудинное сочленение с помощью гиалинового хряща, лонное сращение, образованное в основном волокнистой хрящевой тканью, а также межпозвонковые диски, состоящие из механически прочного фиброзного кольца из волокнистого хряща, которое заключает исполняющее роль амортизатора полужидкое студенистое ядро.
3. Синостозы - соединения костей посредством костной ткани - возникают в качестве завершающей стадии развития скелета путем замещения синхондрозов и синдесмозов (например, соединение костей таза и костей черепа после завершения их роста).
Прерывные соединения костей (диартрозы, синовиальные соединения, или суставы) обеспечивают свободные движения костей, которые удерживаются посредством связок и окружены плотной соединительнотканной суставной капсулой (сумкой), охватывающей их концы в виде муфты. Для достижения минимального трения суставные поверхности костей покрыты гладким суставным хрящом и смачиваются синовиальной жидкостью, заполняющей суставную полость (рис. 84). Суставной хрящ (обычно гиалиновый) прочно прикреплен к кости (рис. 85), имеет гладкую поверхность и обеспечивает не только скольжение, но амортизацию толчков. Питание суставного хряща осуществляется из двух источников: из синовиальной жидкости (основной путь) и со стороны субхондральной костной ткани, контактирующей с кальцифицированным хрящом.
Суставной хрящ по строению отчасти сходен с хрящевой эпифизарной пластинкой роста кости. В нем выделяют: (1) тангенциальную зону (поверхностную); (2) переходную зону (промежуточную) и (3) радиальную зону (базальную), которые связаны с субхондральной кальцифицированной пластинкой (см. рис. 85).
1. Тангенциальная зона состоит из бесклеточной пластинки, обращенной в полость сустава, и тангенциального слоя уплощенных хондроцитов. Коллагеновые волокна в этой зоне располагаются в большинстве почти параллельно (тангенциально) суставной поверхности.
2. Переходная (промежуточная) зона содержит слой округлых хондроцитов и слой изогенных групп хондроцитов.
3. Радиальная (базальная) зона образована колонками хондроцитов, а также слоем гипертрофированных хондроцитов (необызвествленного хряща). Между колонками коллагеновые волокна ориентированы преимущественно под углом к суставной поверхности, приближаясь к ней в виде дуг, в глубокой части зоны они располагаются радиально - перпендикулярно суставной поверхности. Границей между радиальной зоной и субхондральной кальцифицированной пластинкой, прикрепленной к субхондральной костной ткани, служит волнообразная базофильная пограничная линия, соответствующая фронту минерализации.
Суставная капсула (сумка) герметически окружает область сустава, прочно прикрепляясь к надкостнице костей выше и ниже расположения суставных поверхностей и ограничивая суставную полость. Она образована двумя слоями - наружным волокнистым слоем (фиброзная мембрана) и внутренним синовиальным слоем (синовиальная мембрана) (см. рис. 85).
Волокнистый слой образован плотной волокнистой соединительной тканью, которая переходит в надкостницу.
Синовиальный слой выстилает изнутри суставную сумку за исключением суставных поверхнос-
тей, покрытых хрящом; в отдельных участках он образует синовиальные складки и синовиальные ворсинки. Синовиальный слой может либо вплотную прилежать к фиброзной оболочке, либо отделяться от нее слоем рыхлой волокнистой соединительной или жировой ткани. Он имеет сложное строение и состоит из двух слоев (см. рис. 85): субинтимального фиброваскулярного слоя (включая его глубокую и поверхностную части) и синовиальной интимы (синовиальной внутренней оболочки). Субинтимальный фиброваскулярный слой представлен клетками, а также коллагеновыми и эластическими волокнами, имеющими в нем различную ориентацию.
Синовиальная интима обращена в полость сустава и состоит из 1-6 слоев синовиальных клеток (синовиоцитов), расположенных в виде несплошных эпителиоидных пластов, в которых между клетками лежат компоненты межклеточного вещества.
Синовиоциты - специализированные клетки соединительной ткани - разделяются на два основных типа (рис. 86), между которыми имеются промежуточные варианты:
1. Фагоцитирующие синовиальные клетки, или синовиоциты А, - удлиненные макрофагоподобные клетки с овальным ядром, многочисленными митохондриями, умеренно развитым синтетическим аппаратом, высоким содержанием лизосом, фагосом, пиноцитозных пузырьков. На их поверхности имеются многочисленные ветвящиеся микроворсинки. Функция этих клеток связана с поглощением (резорбцией) компонентов синовиальной жидкости.
2. Секреторные синовиальные клетки, или синовиоциты В, - полигональные или отростчатые фибробластоподобные клетки с круглым ядром, многочисленными митохондриями, хорошо развитым синтетическим аппаратом и плотными секреторными гранулами. Эти клетки образуют компоненты матрикса и секретируют ряд веществ (протеогликаны и гиалуроновую кислоту) в синовиальную жидкость, которая смачивает суставные поверхности, исполняя роль смазки, и обеспечивает питание суставных хрящей.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
(ткани внутренней среды)
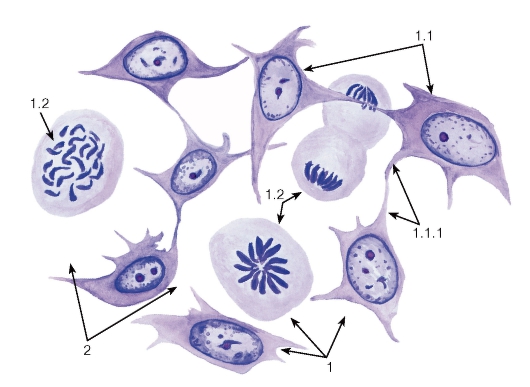
Рис. 49. Мезенхима зародыша - источник развития соединительных тканей
Окраска: гематоксилин-эозин
1 - клетки: 1.1 - клетки в интерфазе, 1.1.1 - отростки клеток, 1.2 - митотически делящиеся клетки;
2 - межклеточные промежутки
Кровь и кроветворные ткани
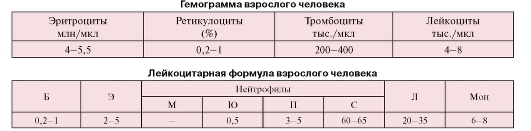
Б - базофилы, Э - эозинофилы, М - миелоциты, Ю - юные (метамиелоциты), П - палочкоядерные, С - сегментоядерные, Л - лимфоциты, Мон - моноциты
Доли различных лейкоцитов приведены в процентах от их общего числа
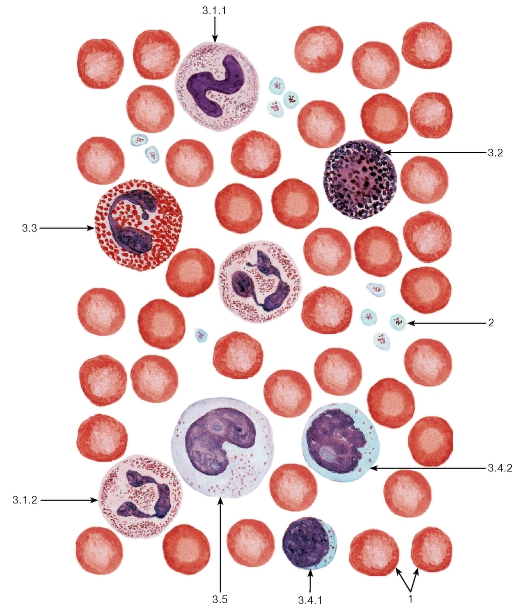
Рис. 50. Кровь человека (мазок)
Окраска: по Романовскому-Гимзе
1 - эритроциты; 2 - тромбоциты; 3 - лейкоциты: 3.1 - нейтрофильные гранулоциты (3.1.1 - палочкоядерный, 3.1.2 - сегментоядерный), 3.2 - базофильный гранулоцит, 3.3 - эозинофильный гранулоцит, 3.4 - лимфоциты (3.4.1 - малый лимфоцит, 3.4.2 - средний лимфоцит), 3.5 - моноцит
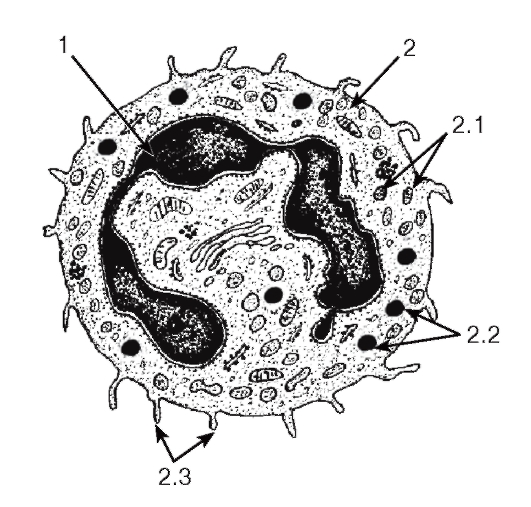
Рис. 51. Ультраструктура сегментоядерного нейтрофильного гранулоцита
Рисунок с ЭМФ
1 - ядро; 2 - цитоплазма: 2.1 - специфическиегранулы, 2.2 - неспецифические гранулы, 2.3 - псевдоподии
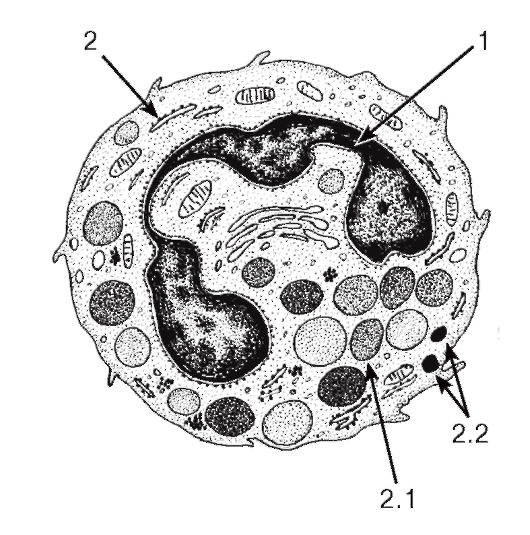
Рис. 52. Ультраструктура базофильного гранулоцита
Рисунок с ЭМФ
1 - ядро; 2 - цитоплазма: 2.1 - специфические гранулы, 2.2 - неспецифические гранулы
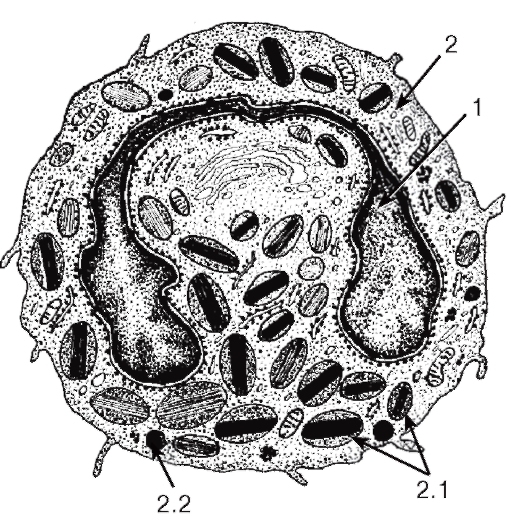
Рис. 53. Ультраструктура эозинофильного гранулоцита
Рисунок с ЭМФ
1 - ядро; 2 - цитоплазма: 2.1 - специфические гранулы с кристаллоидными телами; 2.2 - неспецифические гранулы
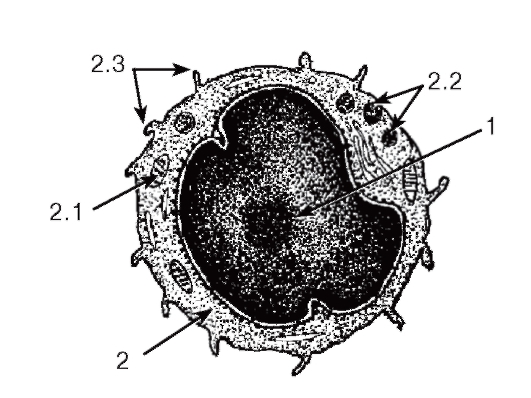
Рис. 54. Ультраструктура лимфоцита
Рисунок с ЭМФ
1 - ядро; 2 - цитоплазма: 2.1 - митохондрия, 2.2 - неспецифические (азурофильные) гранулы, 2.3 - псевдоподии
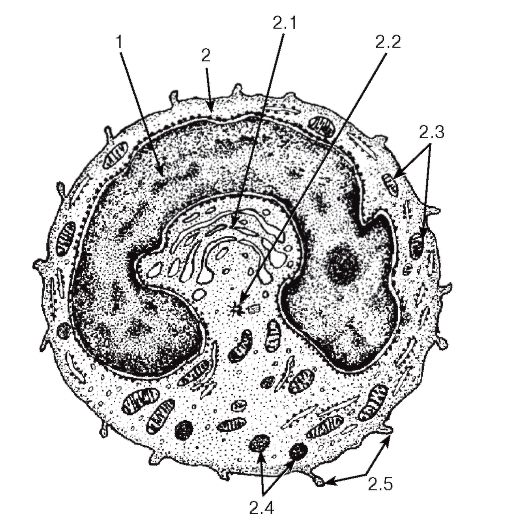
Рис. 55. Ультраструктура моноцита
Рисунок с ЭМФ
1 - ядро; 2 - цитоплазма: 2.1 - комплекс Гольджи, 2.2 - центриоли, 2.3 - митохондрии, 2.4 - неспецифические гранулы, 2.5 - псевдоподии
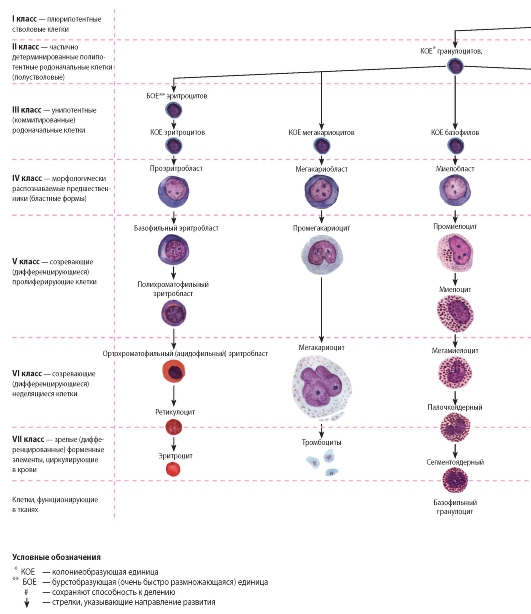
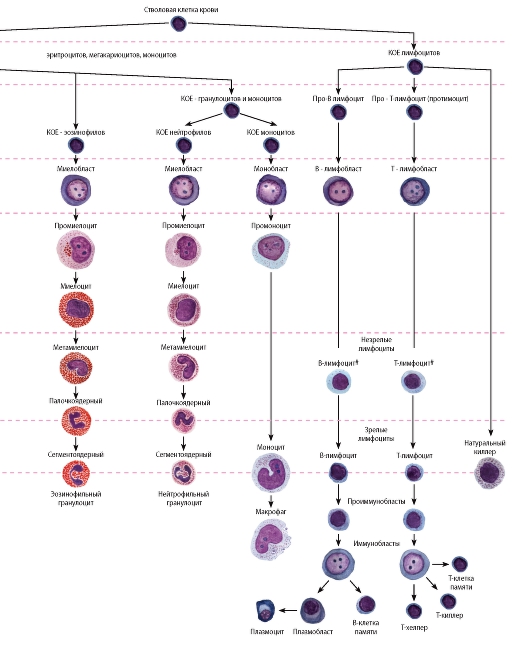
Рис. 56. Схема кроветворения
Структурные преобразования гемопоэтических клеток (начиная со стадии морфологически распознаваемых предшественников)
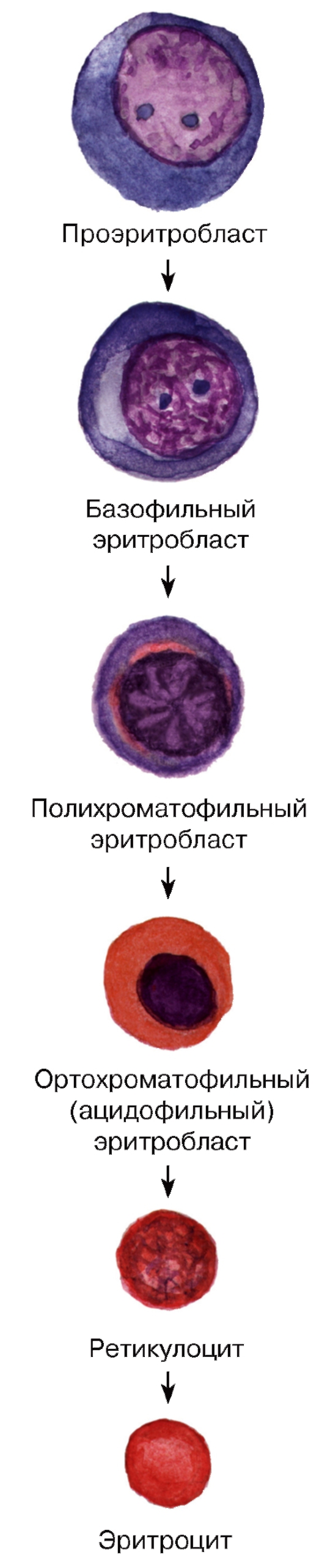
Рис. 57. Эритропоэз
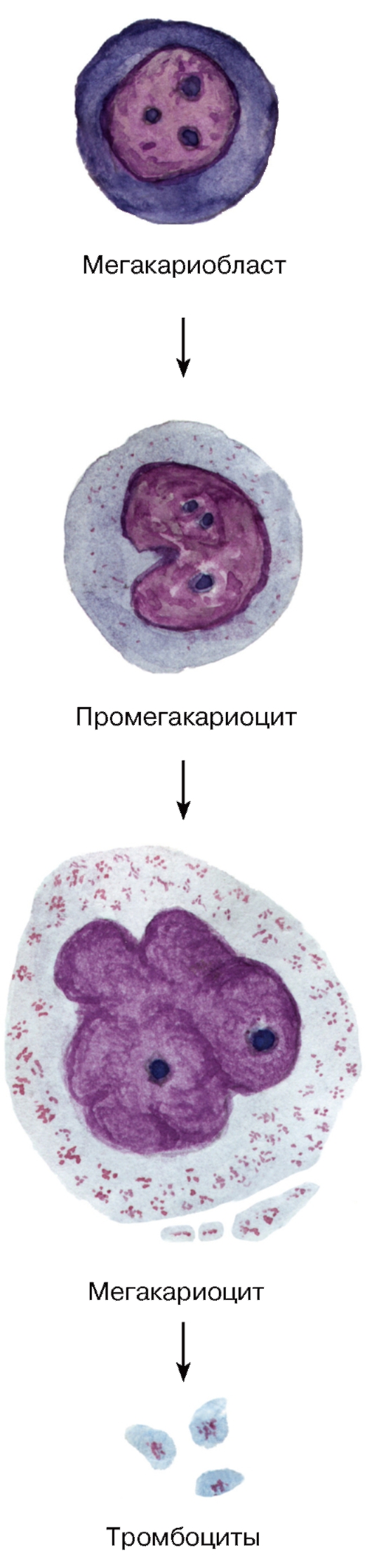
Рис. 58. Тромбоцитопоэз
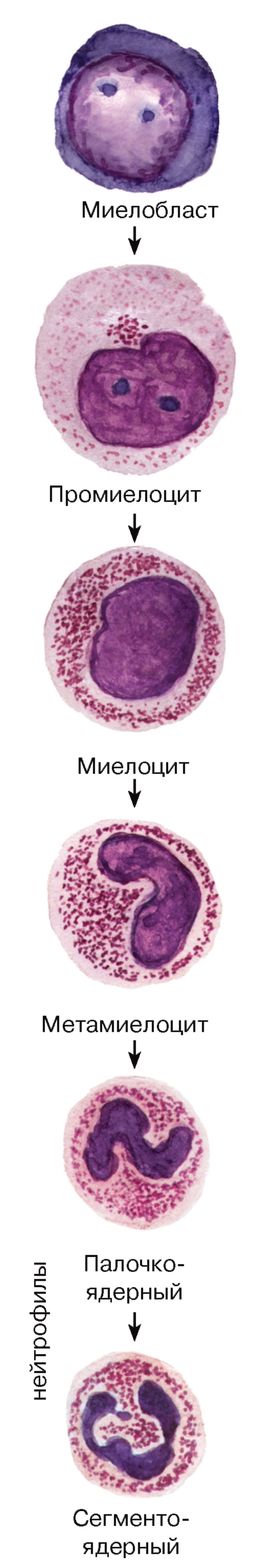
Рис. 59. Гранулоцитопоэз: образование нейтрофильных гранулоцитов
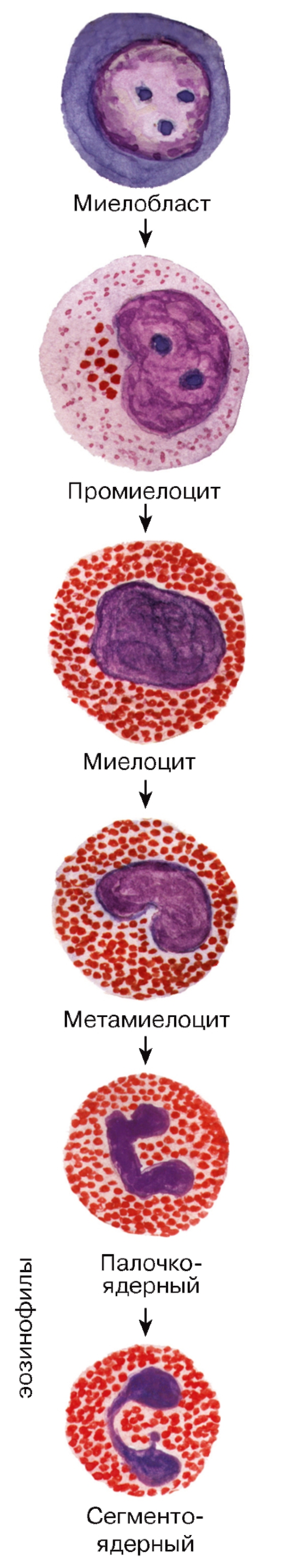
Рис. 60. Гранулоцитопоэз: образование эозинофильных гранулоцитов
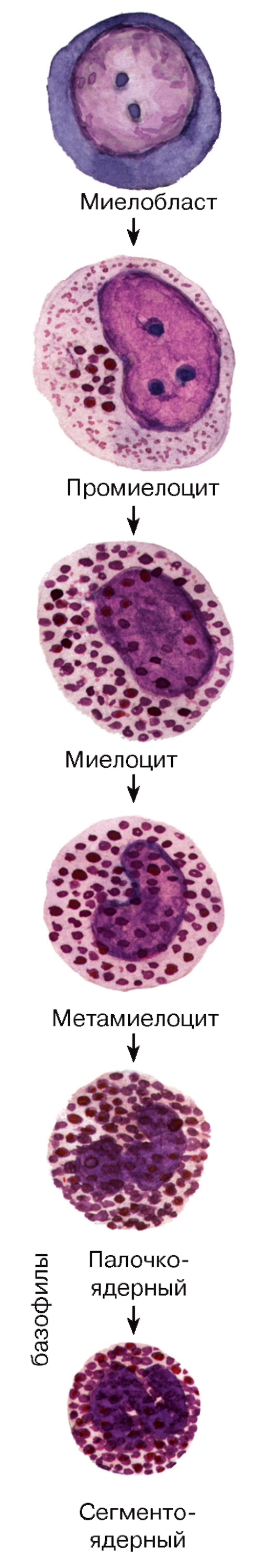
Рис. 61. Гранулоцитопоэз: образование базофильных гранулоцитов
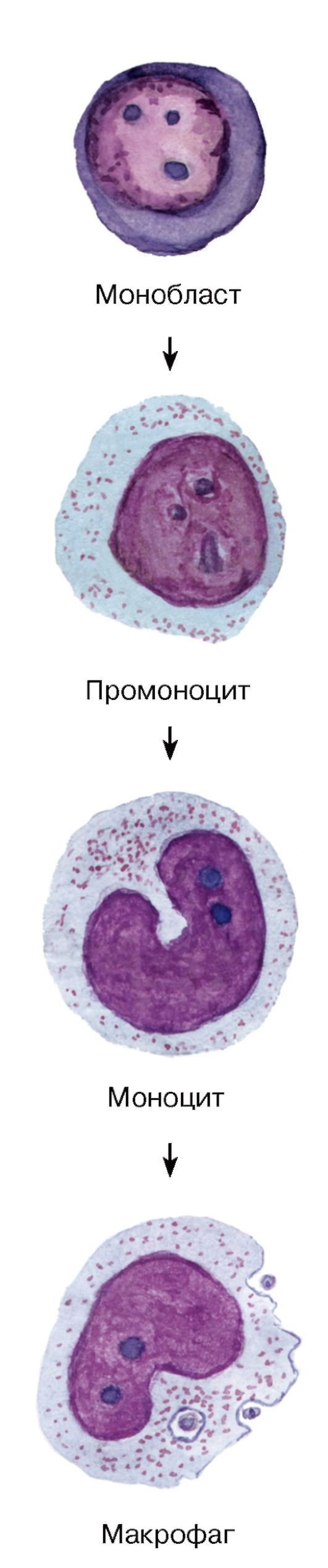
Рис. 62. Моноцитопоэз: образование моноцитов и макрофагов
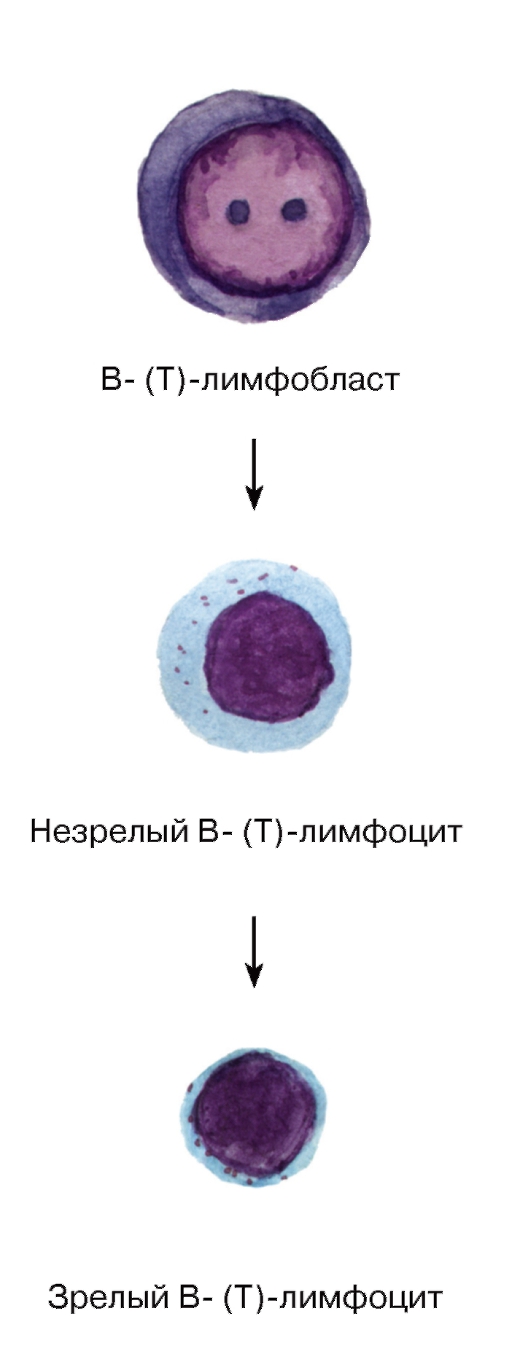
Рис. 63. Лимфоцитопоэз (антиген-независимый этап)
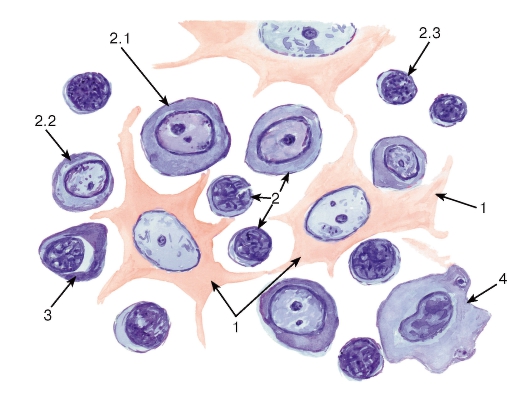
Рис. 64. Лимфоидная ткань (лимфатический узел)
Окраска: гематоксилин-эозин
1 - клетки ретикулярной ткани; 2 - лимфоциты: 2.1 - большой лимфоцит (лимфобласт), 2.2 - средний лимфоцит (незрелый), 2.3 - малый лимфоцит (зрелая форма); 3 - плазмоцит; 4 - макрофаг
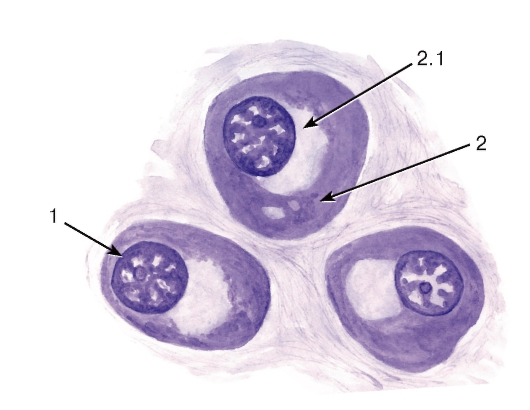
Рис. 65. Плазмоциты в лимфоидной ткани (лимфатический узел в условиях иммунной реакции)
Окраска: галлоцианин
1 - ядро; 2 - цитоплазма: 2.1 - околоядерный «дворик»
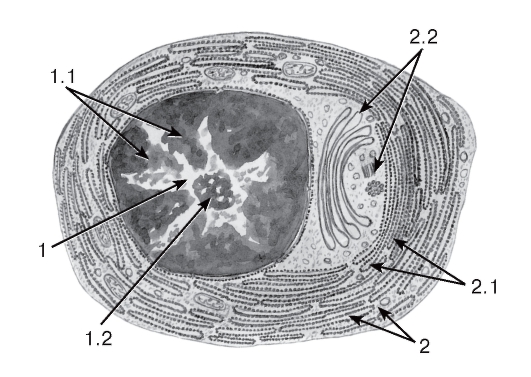
Рис. 66. Ультраструктура плазмоцита
Рисунок с ЭМФ
1 - ядро: 1.1 - гетерохроматин в виде спиц колеса, 1.2 - ядрышко; 2 - цитоплазма: 2.1 - цистерны гранулярной эндоплазматической сети, 2.2 - комплекс Гольджи и центриоли (соответствуют расположению околоядерного «дворика» на гистологических препаратах)
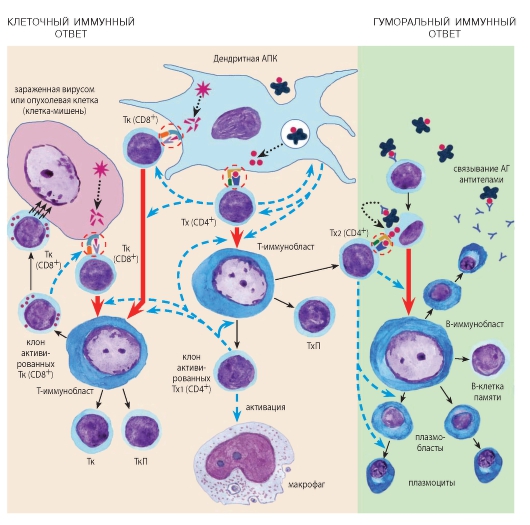
Рис. 67. Схема иммуногенеза:
КЛЕТОЧНЫЙ ИММУНИТЕТ. Образование Тх. Дендритные АПК поглощают экзогенные АГ, подвергают их процессингу и экспрессируют на своей поверхности в виде комплекса ЭАГ/ МНС II. Тх (CD4+) связываются с этим комплексом посредством ТКР и молекулы CD4 (двойное иммунное распознавание). При этом АПК и Тх воздействуют друг на друга цитокинами. Активированные Тх подвергаются БТП, превращаясь в один из двух подклассов (Тх1 или Тх2). Тх1 стимулируют преимущественно реакции клеточного иммунитета: они выделяют цитокины, способствующие образованию Тх, Тк и активирующие макрофаги. Тх2 стимулируют преимущественно реакции гуморального иммунитета (см. ниже). Часть Тх превращаются в ТхП.
Образование Тк. Дендритные АПК и зараженные вирусами или опухолевые клетки-мишени подвергают процессингу эндогенные АГ и экспрессируют их на своей поверхности в виде комплекса ЭАГ/МНС I. Тк (CD8+) связываются с этим комплексом посредством ТКР и молекулы CD8 (двойное иммунное распознавание). Последующие активация, БТП и дифференцировка Тк требуют помощи со стороны Тх1, выделяющих соответствующие цитокины. Активированные Тк выделяют цитокины и уничтожают клетки-мишени, распознавая комплекс АГ/МНС I на их поверхности (не показано), прикрепляясь к ним и выделяя цитотоксические вещества, накопленные в цитоплазматических гранулах. Часть Тк превращаются в ТкП.
ГУМОРАЛЬНЫЙ ИММУНИТЕТ. В-лимфоциты специфически связывают экзогенные АГ с помощью поверхностных иммуноглобулиновых рецепторов, поглощают их, подвергают процессингу и экспрессируют на своей поверхности в виде комплекса ЭАГ/МНС II. Тх2 (CD4+) связываются с этим комплексом посредством ТКР и молекулы CD4 (двойное иммунное распознавание), выделяя цитокины, которые активируют В-лимфоциты, стимулируют их БТП и дифференцировку в плазматические клетки, секретирующие иммуноглобулины (антитела). При воздействии АГ формируются также В-клетки памяти.
Представленная схема соответствует антиген-зависимому этапу лимфоцитопоэза

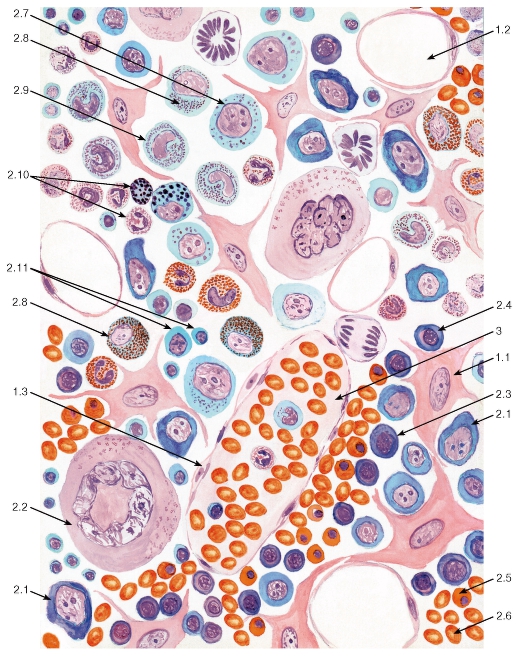
Рис. 68. Миелоидная ткань (красный костный мозг)
Окраска: азур II-эозин
1 - клетки стромы: 1.1 - ретикулярная клетка, 1.2 - жировая клетка (адипоцит), 1.3 - макрофаг; 2 - гемопоэтические клетки: 2.1 - бластные формы, 2.2 - мегакариоцит, 2.3 - эритробласт базофильный, 2.4 - эритробласт полихроматофильный, 2.5 - эритробласт ортохроматофильный, 2.6 - зрелый эритроцит, 2.7 - промиелоцит, 2.8 - миелоцит, 2.9 - метамиелоцит (юный), 2.10 - зрелые гранулоциты, 2.11 - лимфоциты; 3 - синусоид, содержащий зрелые форменные элементы крови
Волокнистые соединительные ткани
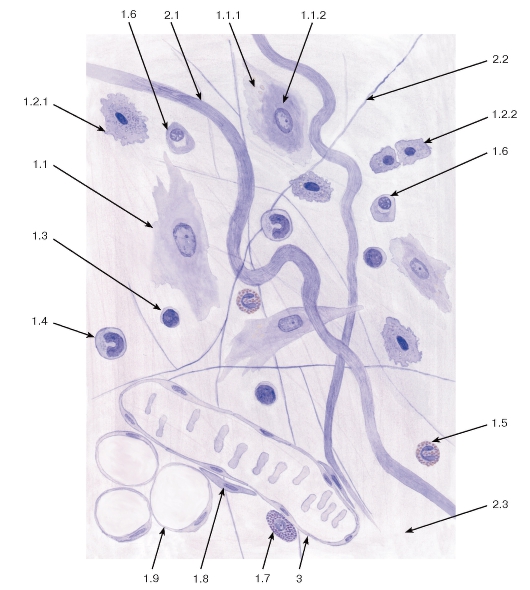
Рис. 69. Рыхлая волокнистая соединительная ткань (пленочный препарат)
Окраска: железный гематоксилин
1 - клетки: 1.1 - фибробласт, 1.1.1 - эктоплазма, 1.1.2 - эндоплазма, 1.2 - гистиоцит (макрофаг), 1.2.1 - активированный гистиоцит, 1.2.2 - неактивные гистиоциты, 1.3 - лимфоцит, 1.4 - моноцит, 1.5 - эозинофил, 1.6 - плазмоцит, 1.7 - тучная клетка, 1.8 - адвентициальная клетка, 1.9 - адипоцит;
2 - межклеточное вещество: 2.1 - коллагеновое волокно, 2.2 - эластическое волокно, 2.3 - основное (аморфное) вещество; 3 - кровеносный сосуд
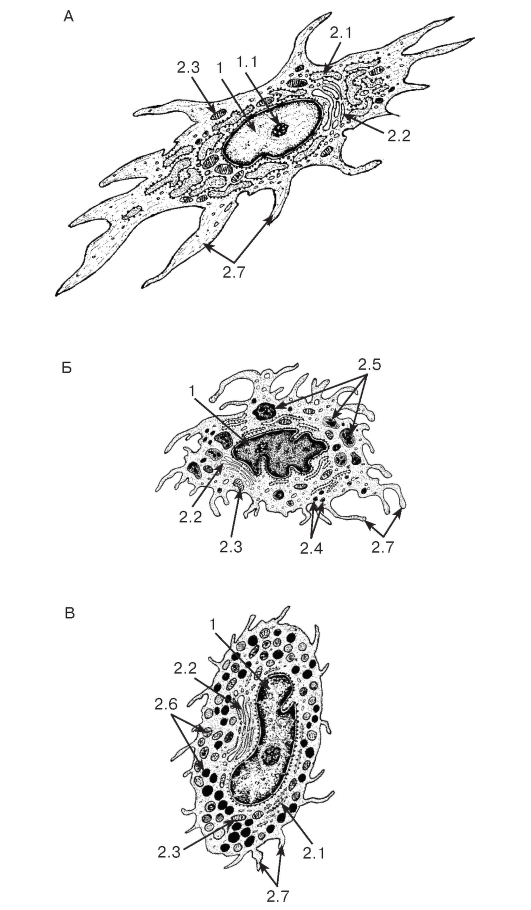
Рис. 70. Ультраструктурная организация клеток соединительной ткани
Рисунки с ЭМФ
А - фибробласт; Б - гистиоцит; В - тучная клетка
1 - ядро: 1.1 - ядрышко; 2 - цитоплазма: 2.1 - цистерна гранулярной эндоплазматической сети, 2.2 - комплекс Гольджи, 2.3 - митохондрии, 2.4 - лизосомы, 2.5 - фаголизосомы, 2.6 - секреторные гранулы, 2.7 - отростки
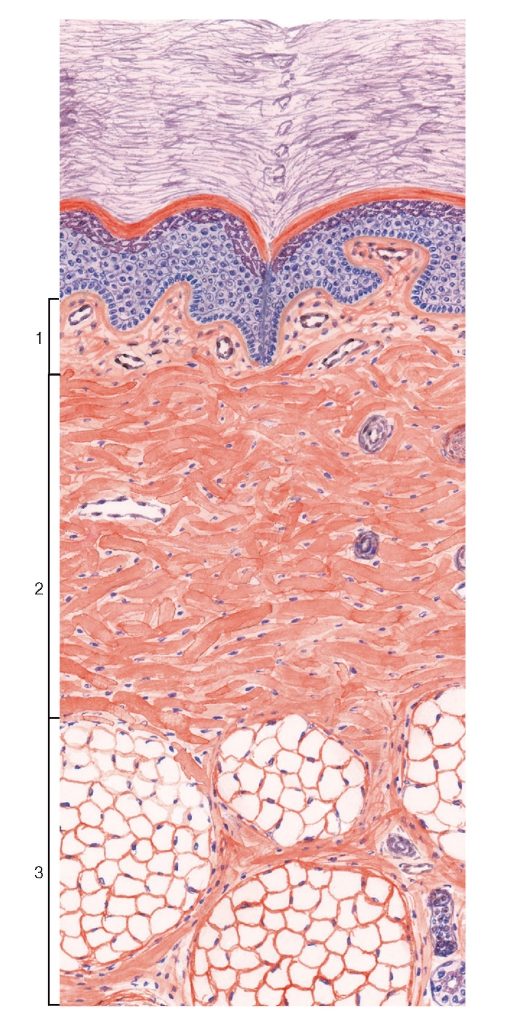
Рис. 71. Различные виды соединительных тканей (кожа пальца)
Окраска: гематоксилин-эозин
1 - рыхлая волокнистая соединительная ткань; 2 - плотная волокнистая неоформленная соединительная ткань; 3 - жировая ткань
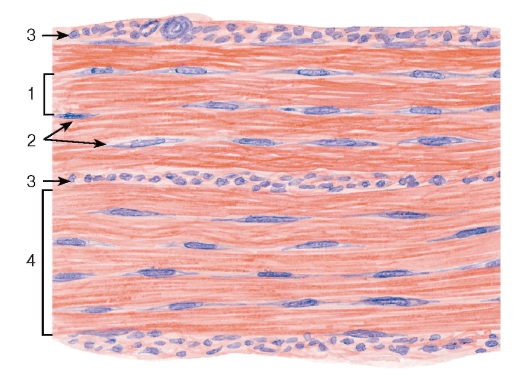
Рис. 72. Плотная волокнистая оформленная соединительная ткань (сухожилие, продольный срез)
Окраска: гематоксилин-эозин
1 - первичный сухожильный пучок; 2 - сухожильные клетки (фиброциты); 3 - эндотендиний; 4 - вторичный сухожильный пучок
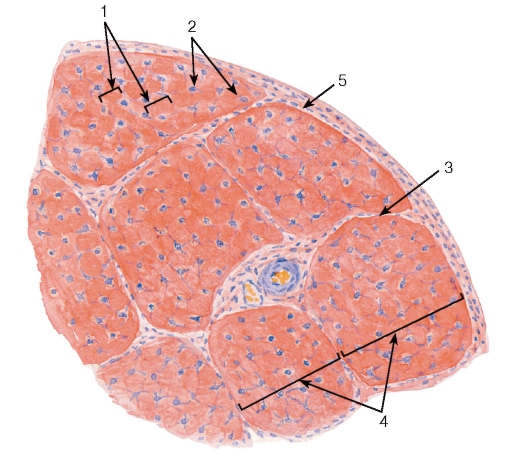
Рис. 73. Плотная волокнистая оформленная соединительная ткань (сухожилие, поперечный срез)
Окраска: гематоксилин-эозин
1 - первичные сухожильные пучки; 2 - сухожильные клетки (фиброциты); 3 - эндотендиний; 4 - вторичные сухожильные пучки; 5 - перитендиний
Скелетные соединительные ткани
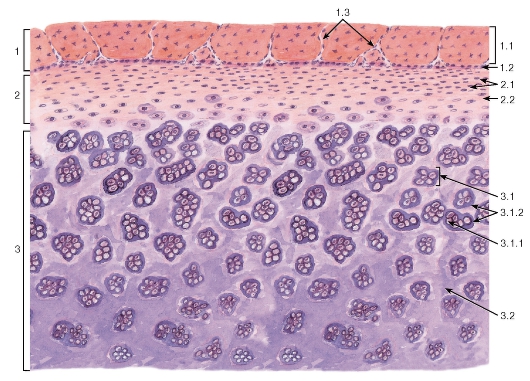
Рис. 74. Гиалиновая хрящевая ткань (участок гиалинового хряща)
Окраска: гематоксилин-эозин
1 - надхрящница: 1.1 - наружный фиброзный слой, 1.2 - внутренний (хондрогенный) клеточный слой, 1.3 - кровеносные сосуды; 2 - зона молодого хряща: 2.1 - хондроциты, 2.2 - межклеточное вещество (хрящевой матрикс); 3 - зона зрелого хряща: 3.1 - клеточная территория, 3.1.1 - изогенная группа хондроцитов, 3.1.2 - территориальный матрикс, 3.2 - интертерриториальный матрикс
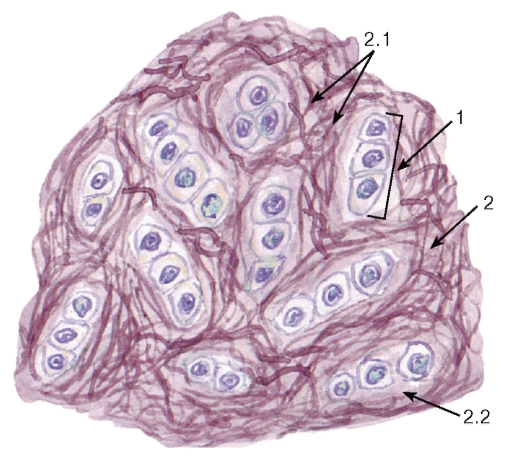
Рис. 75. Эластическая хрящевая ткань (участок эластического хряща)
Окраска: орсеин-гематоксилин
1 - изогенная группа хондроцитов; 2 - межклеточное вещество (хрящевой матрикс): 2.1 - эластические волокна, 2.2 - основное вещество
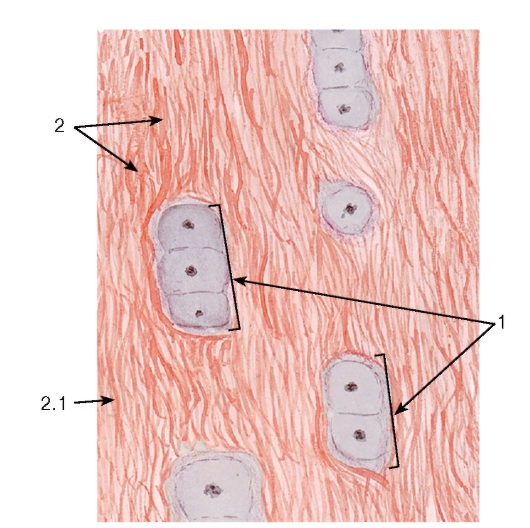
Рис. 76. Волокнистая (фиброзная) хрящевая ткань (участок волокнистого хряща)
Окраска: гематоксилин-эозин
1 - изогенные группы хондроцитов; 2 - межклеточное вещество (хрящевой матрикс): 2.1 - коллагеновые волокна
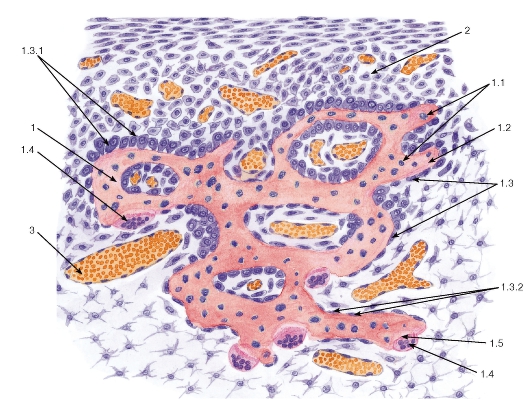
Рис. 77. Развитие костной ткани непосредственно из мезенхимы (прямой остеогенез)
Окраска: гематоксилин-эозин
1 - костная трабекула: 1.1 - лакуны остеоцитов, 1.2 - обызвествленное межклеточное вещество, 1.3 - остеобласты, 1.3.1 - активные остеобласты, 1.3.2 - неактивные остеобласты, 1.4 - остеокласты, 1.5 - эрозионная лакуна; 2 - клетки остеогенной (дифференцирующейся из мезенхимы) соединительной ткани; 3 - кровеносный сосуд
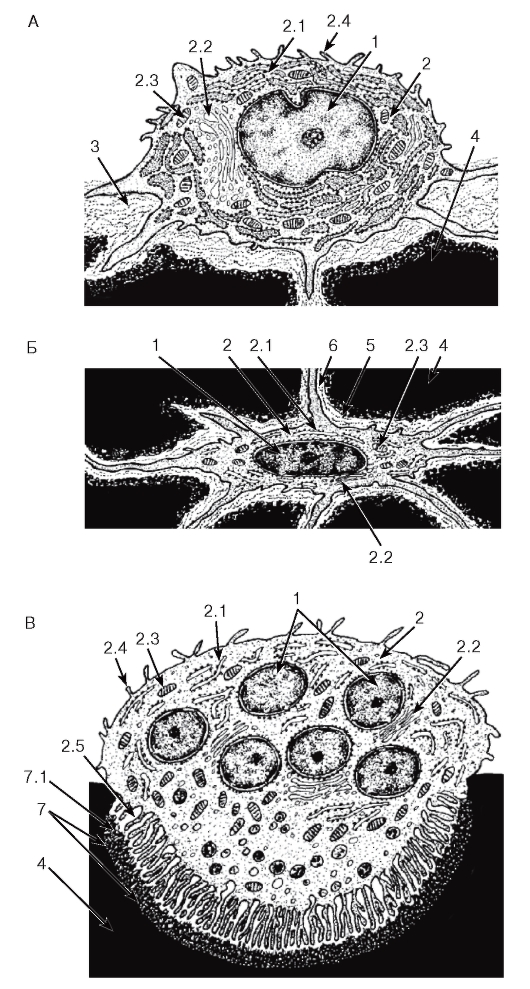
Рис. 78. Ультраструктурная организация клеток костной ткани
Рисунки с ЭМФ
А - остеобласт; Б - остеоцит; В - остеокласт
1 - ядро (ядра); 2 - цитоплазма: 2.1 - цистерны гранулярной эндоплазматической сети, 2.2 - комплекс Гольджи, 2.3 - митохондрии, 2.4 - микроворсинки, 2.5 - микроскладчатая кайма (цитоплазматические отростки); 3 - остеоид; 4 - обызвествленное межклеточное вещество; 5 - лакуна остеоцита (содержит тело клетки); 6 - костные канальцы с отростками остеоцита; 7 - эрозионная лакуна: 7.1 - эрозионный фронт
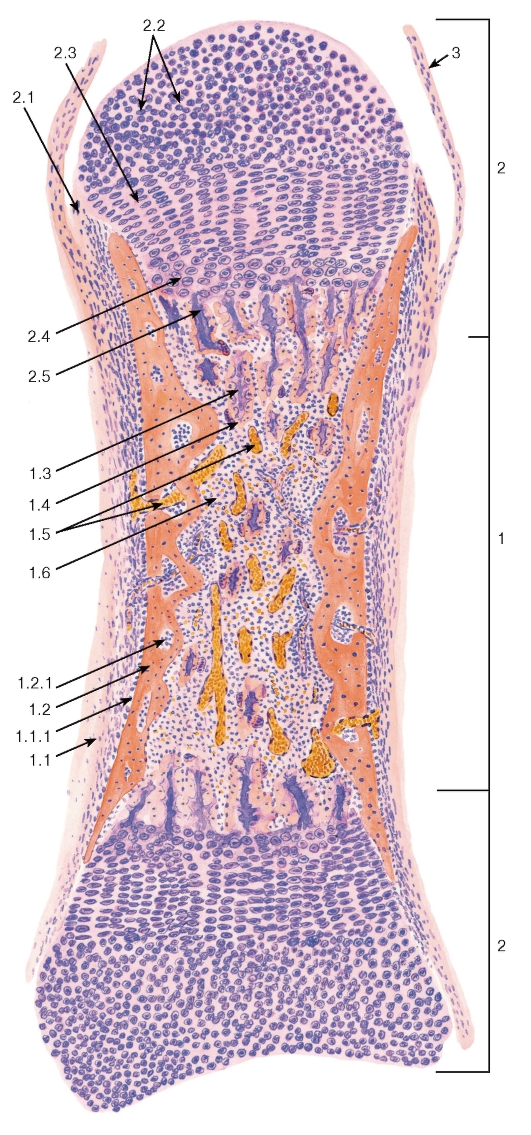
Рис. 79. Развитие кости на месте хряща (непрямой остеогенез)
Окраска: гематоксилин-эозин
1 - диафиз: 1.1 - надкостница, 1.1.1 - осте огенный слой (внутренний слой надкостницы), 1.2 - перихондральное костное кольцо, 1.2.1 - отверстие, 1.3 - остатки обызвествленного хряща, 1.4 - эндохондральная кость, 1.5 - кровеносные сосуды, 1.6 - формирующийся костный мозг; 2 - эпифизы: 2.1 - надхрящница, 2.2 - зона покоя, 2.3 - зона пролиферации (с колонками хондроцитов), 2.4 - зона гипертрофии, 2.5 - зона кальцификации; 3 - суставная сумка
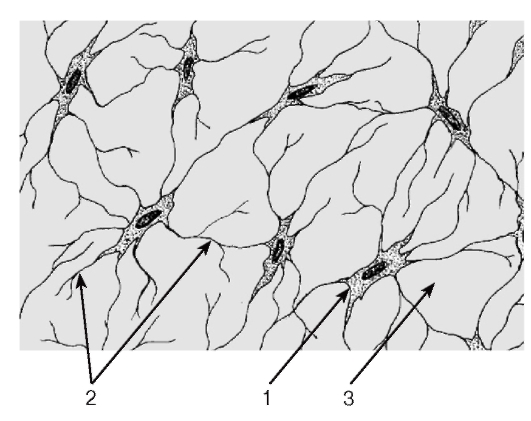
Рис. 80. Грубоволокнистая костная ткань (тотальный плоскостной препарат)
Не окрашен
1 - лакуна остеоцита (место расположения тела клетки); 2 - костные канальцы (содержащие отростки остеоцитов); 3 - межклеточное вещество
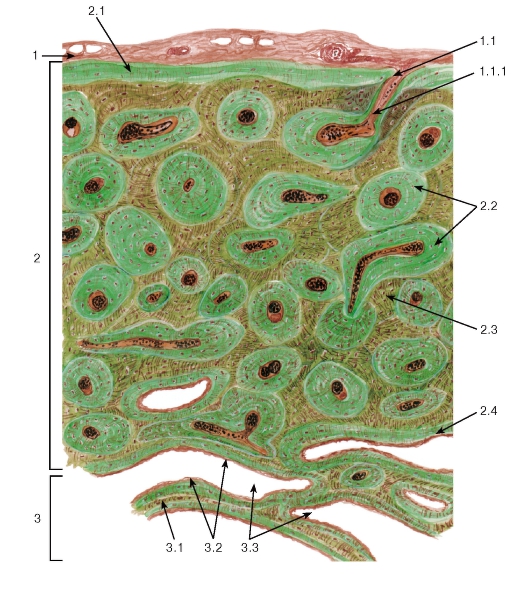
Рис. 81. Пластинчатая костная ткань (поперечный срез диафиза декальцинированной трубчатой кости)
Окраска: тионин-пикриновая кислота
1 - надкостница: 1.1 - перфорирующий (фолькмановский) канал, 1.1.1 - кровеносный сосуд;
2 - компактное вещество кости: 2.1 - наружные опоясывающие пластинки, 2.2 - остеоны, 2.3 - интерстициальные пластинки, 2.4 - внутренние опоясывающие пластинки; 3 - губчатое вещество кости: 3.1 - костные трабекулы, 3.2 - эндост, 3.3 - межтрабекулярные пространства
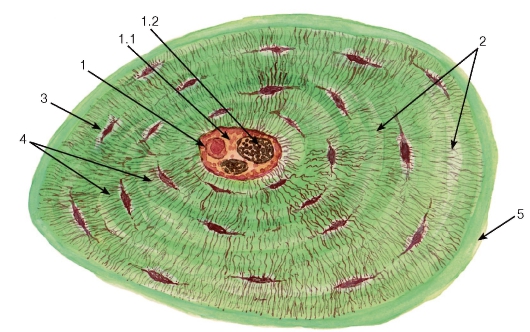
Рис. 82. Поперечный срез остеона
(диафиз декальцинированной трубчатой кости)
Окраска: тионин-пикриновая кислота
1 - канал остеона: 1.1 - соединительная ткань, 1.2 - кровеносные сосуды; 2 - концентрические костные пластинки; 3 - лакуна остеоцита, содержащее его тело; 4 - костные канальцы с отростками остеоцитов; 5 - цементирующая линия
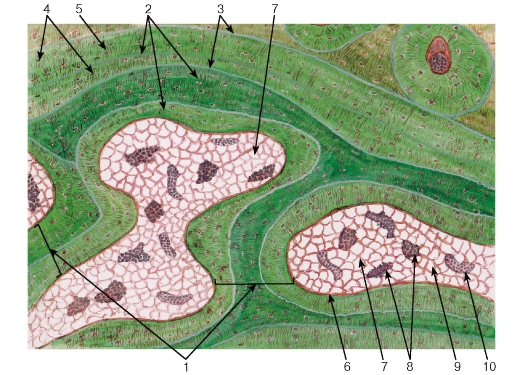
Рис. 83. Пластинчатая костная ткань. Участок губчатого вещества (диафиз декальцинированной трубчатой кости)
Окраска: тионин-пикриновая кислота
1 - костные трабекулы; 2 - пакеты костных пластинок; 3 - цементирующие линии; 4 - лакуны остеоцитов, содержащие их тела; 5 - костные канальцы с отростками остеоцитов; 6 - эндост; 7 - межтрабекулярные пространства; 8 - костный мозг; 9 - жировая ткань; 10 - кровеносный сосуд
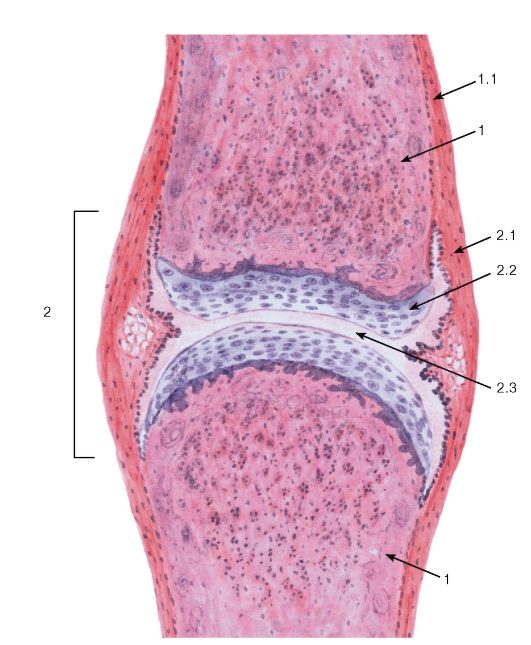
Рис. 84. Синовиальное соединение (сустав). Общий вид
Окраска: гематоксилин-эозин
1 - кость: 1.1 - надкостница; 2 - синовиальное соединение (сустав): 2.1 - суставная капсула (сумка), 2.2 - суставной хрящ (гиалиновый), 2.3 - суставная полость (содержит синовиальную жидкость)
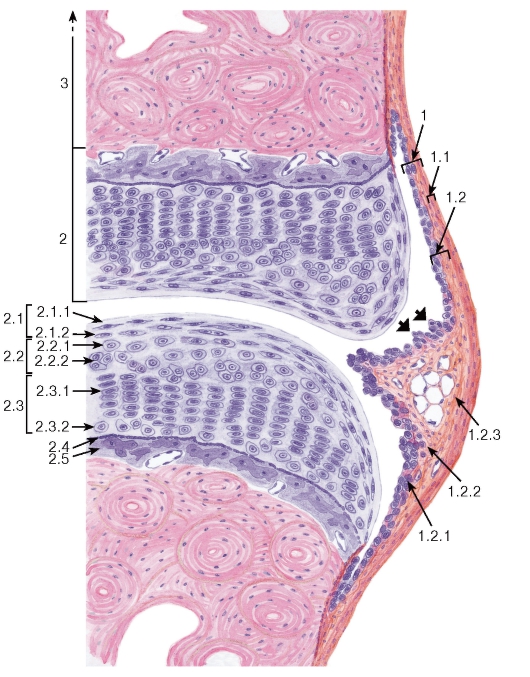
Рис. 85. Участок синовиального соединения (сустава)
Окраска: гематоксилин-эозин
1 - суставная капсула (сумка): 1.1 - волокнистый слой, 1.2 - синовиальный слой, образующий синовиальные ворсинки (показаны жирными стрелками),1.2.1 - синовиальная интима (синовиоциты), 1.2.2 - глубокая часть субинтимального фиброваскулярного слоя, 1.2.3 - поверхностная часть субинтимального фиброваскулярного слоя; 2 - суставной хрящ (гиалиновый): 2.1 - тангенциальная зона, 2.1.1 - бесклеточная пластинка, 2.1.2 - уплощенные хондроциты, 2.2 - промежуточная зона, 2.2.1 - округлые хондроциты, 2.2.2 - изогенные группы хондроцитов, 2.3 - радиальная зона, 2.3.1 - колонки хондроцитов, 2.3.2 - слой гипертрофированных (дистрофически измененных) хондроцитов, 2.4 - пограничная линия (фронт минерализации), 2.5 - кальцифицированный гиалиновыйхрящ; 3 - субхондральная костная ткань
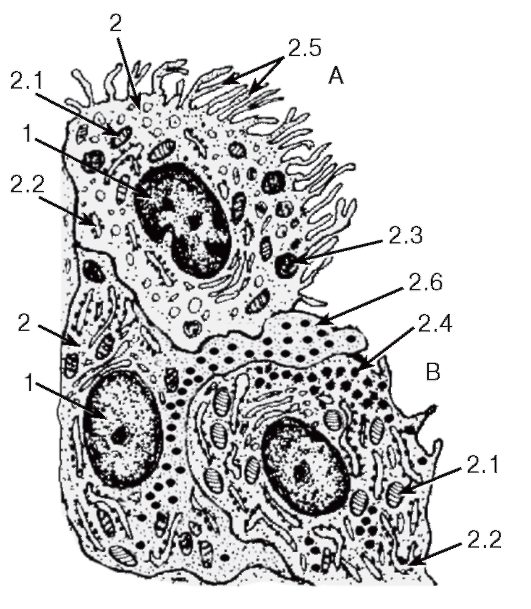
Рис. 86. Ультраструктурная организация синовиальных клеток (синовиоцитов)
Рисунок с ЭМФ
A - синовиоцит A (фагоцитирующая синовиальная клетка);
B - синовиоциты В (секреторные синовиальные клетки):
1 - ядро, 2 - цитоплазма: 2.1 - митохондрии, 2.2 - цистерны гранулярной эндоплазматической сети, 2.3 - лизосомы, 2.4 - секреторные гранулы, 2.5 - микроворсинки, 2.6 - цитоплазматический отросток