Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. - 296 с. :ил.
|
|
|
|
ОРГАНЫ ЖЕНСКОЙ ПОЛОВОЙ СИСТЕМЫ
Органы женской половой системы включают: 1) внутренние (расположенные в тазу) - женские гонады - яичники, маточные трубы, матку, влагалище; 2) наружные - лобок, малые и большие половые губы и клитор. Они достигают полного развития с наступлением полового созревания, когда устанавливается их циклическая деятельность (овариально-менструальный цикл), продолжающаяся в течение репродуктивного периода женщины и прекращающаяся с его завершением, после чего органы половой системы утрачивают функцию и атрофируются.
Яичник
Яичник выполняет две функции - генеративную (образование женских половых клеток - овогенез) и эндокринную (синтез женских половых гормонов). Снаружи он одет кубическим поверхностным эпителием (видоизмененным мезотелием) и состоит из коркового и мозгового вещества (рис. 264).
Корковое вещество яичника - широкое, нерезко отделено от мозгового. Основную его массу составляют фолликулы яичника, образованные половыми клетками (овоцитами), которые окружены фолликулярными эпителиоцитами.
Мозговое вещество яичника - небольшое, содержит крупные извитые кровеносные сосуды и особые хилусные клетки.
Строма яичника представлена плотной соединительнотканной белочной оболочкой, лежащей под поверхностным эпителием, и своеобразной веретеновидноклеточной соединительной тканью, в которой в виде завихрений плотно располагаются веретеновидные фибробласты и фиброциты.
Овогенез (за исключением завершающего этапа) протекает в корковом веществе яичника и включает 3 фазы: 1) размножения, 2) роста и 3) созревания.
Фаза размножения овогоний происходит внутриутробно и завершается до рождения; большая часть образовавшихся клеток гибнет, меньшая вступает в фазу роста, превращаясь в первичные овоциты, развитие которых блокируется в профазе I деления мейоза, во время которой (как и при сперматогенезе) происходит обмен сегментов хромосом, обеспечивающий генетическое разнообразие гамет.
Фаза роста овоцита складывается из двух периодов: малого и большого. Первый отмечается до полового созревания в отсутствие гормональной сти-
муляции; второй происходит только после него под действием фолликулостимулирующего гормона (ФСГ) гипофиза и характеризуется периодическим вовлечением фолликулов в циклическое развитие, завершающееся их созреванием.
Фаза созревания начинается с возобновления деления первичных овоцитов в зрелых фолликулах непосредственно перед наступлением овуляции. При завершении I деления созревания образуются вторичный овоцит и мелкая, почти лишенная цитоплазмы клетка - первое полярное тельце. Вторичный овоцит сразу же вступает во II деление созревания, которое, однако, останавливается в метафазе. При овуляции вторичный овоцит выделяется из яичника и поступает в маточную трубу, где в случае оплодотворения спермием завершает фазу созревания с образованием гаплоидной зрелой женской половой клетки (яйцеклетки) и второго полярного тельца. Полярные тельца в дальнейшем разрушаются. В отсутствие оплодотворения половая клетка подвергается дегенерации на стадии вторичного овоцита.
Овогенез протекает при постоянном взаимодействии развивающихся половых клеток с эпителиальными в составе фолликулов, изменения которых известны как фолликулогенез.
Фолликулы яичника погружены в строму и состоят из первичного овоцита, окруженного фолликулярными клетками. В них создается микроокружение, необходимое для поддержания жизнеспособности и роста овоцита. Фолликулы обладают и эндокринной функцией. Размеры и строение фолликула зависят от стадии его развития. Различают: примордиальные, первичные, вторичные и третичные фолликулы (см. рис. 264-266).
Примордиальные фолликулы - самые мелкие и многочисленные, располагаются в виде скоплений под белочной оболочкой и состоят из мелкого первичного овоцита, окруженного однослойным плоским эпителием (фолликулярными эпителиоцитами).
Первичные фолликулы состоят из более крупного первичного овоцита, окруженного одним слоем кубических или столбчатых фолликулярных клеток. Между овоцитом и фолликулярными клетками впервые становится заметной прозрачная оболочка, имеющая вид бесструктурного оксифильного слоя. Она состоит из гликопротеинов, вырабатывается овоцитом и способствует увеличению площади поверхности взаимного обмена веществ между ним и фолликулярными клетками. По мере дальнейшего
роста фолликулов толщина прозрачной оболочки нарастает.
Вторичные фолликулы содержат продолжающий расти первичный овоцит, окруженный оболочкой из многослойного кубического эпителия, клетки которого делятся под влиянием ФСГ. В цитоплазме овоцита накапливается значительное количество органелл и включений, в ее периферической зоне образуются кортикальные гранулы, которые в дальнейшем участвуют в образовании оболочки оплодотворения. В фолликулярных клетках также нарастает содержание органелл, образующих их секреторный аппарат. Прозрачная оболочка утолщается; в нее проникают микроворсинки овоцита, контактирующие с отростками фолликулярных клеток (см. рис. 25). Утолщается базальная мембрана фолликула между этими клетками и окружающей стромой; последняя образует соединительнотканную оболочку (теку) фолликула (см. рис. 266).
Третичные (везикулярные, антральные) фолликулы формируются из вторичных вследствие секреции фолликулярными клетками фолликулярной жидкости, которая сначала накапливается в мелких полостях фолликулярной оболочки, в дальнейшем сливающихся в единую полость фолликула (антрум). Овоцит находится внутри яйценосного бугорка - скопления фолликулярных клеток, выступающего в просвет фолликула (см. рис. 266). Остальные фолликулярные клетки носят название гранулезы и вырабатывают женские половые гормоны эстрогены, уровни которых в крови нарастают по мере роста фолликулов. Тека фолликула разделяется на два слоя: наружный слой теки содержит фибробласты теки, во внутреннем слой теки образуются стероидпродуцирующие эндокриноциты теки.
Зрелые (предовуляторные) фолликулы (граафовы фолликулы) - крупные (18-25 мм), выступают над поверхностью яичника.
Овуляция - разрыв зрелого фолликула с выбросом из него овоцита, как правило, происходит на 14-е сутки 28-дневного цикла под влиянием выброса ЛГ. За несколько часов до овуляции овоцит, окруженный клетками яйценосного бугорка, отделяется от стенки фолликула и свободно плавает в его полости. При этом фолликулярные клетки, связанные с прозрачной оболочкой, удлиняются, образуя так называемый лучистый венец. В первичном овоците происходит возобновление мейоза (заблокированного в профазе I деления) с образованием вторичного овоцита и первого полярного тельца. Вторичный овоцит далее вступает во II деление созревания, которое блокируется в метафазе. Разрыв стенки фолликула и покрываю-
щих ее тканей яичника происходит в небольшом истонченном и разрыхленном выпячивающемся участке - стигме. При этом из фолликула выделяются овоцит, окруженный клетками лучистого венца, и фолликулярная жидкость.
Желтое тело образуется вследствие дифференцировки клеток гранулезы и теки овулировавшего фолликула, стенки которого спадаются, образуя складки, а в просвете находится сгусток крови, замещающийся в дальнейшем соединительной тканью (см. рис. 265).
Развитие желтого тела (лютеогенез) включает 4 стадии: 1) пролиферации и васкуляризации; 2) железистого метаморфоза; 3) расцвета и 4) обратного развития.
Стадия пролиферации и васкуляризации характеризуется активным размножением клеток гранулезы и теки. В гранулезу из внутреннего слоя теки врастают капилляры, а разделяющая их базальная мембрана разрушается.
Стадия железистого метаморфоза: клетки гранулезы и теки превращаются в полигональные светлоокрашенные клетки - лютеоциты (гранулезные и теки), в которых формируется мощный синтетический аппарат. Основную массу желтого тела составляют крупные светлые гранулезные лютеоциты, по его периферии лежат мелкие и темные лютеоциты теки (рис. 267).
Стадия расцвета характеризуется активной функцией лютеоцитов, вырабатывающих прогестерон - женский половой гормон, способствующий возникновению и протеканию беременности. Эти клетки содержат крупные липидные капли, контактируют с обширной капиллярной сетью
(рис. 268).
Стадия обратного развития включает последовательность дегенеративных изменений лютеоцитов с их разрушением (лютеолитическое тело) и замещением плотным соединительнотканным рубцом - беловатым телом (см. рис. 265).
Атрезия фолликулов - процесс, включающий остановку роста и разрушение фолликулов, который, затрагивая мелкие фолликулы (примордиальные, первичные), приводит к их полному разрушению и бесследному замещению соединительной тканью, а при развитии в крупных фолликулах (вторичных и третичных) вызывает их преобразования с формированием атретических фолликулов. При атрезии гибнут овоцит (сохраняется лишь его прозрачная оболочка) и гранулезные клетки, а клетки внутренней теки, напротив, разрастаются (рис. 269). Некоторое время атретический фолликул активно синтезирует стероидные гормоны,
в дальнейшем разрушается, замещаясь соединительной тканью - беловатым телом (см. рис. 265).
Все описанные последовательные изменения фолликулов и желтого тела, протекающие циклически в течение репродуктивного периода жизни женщины и сопровождающиеся соответствующими колебаниями уровней половых гормонов, получили название овариального цикла.
Хилусные клетки образуют скопления вокруг капилляров и нервных волокон в области ворот яичника (см. рис. 264). Они сходны с интерстициальными эндокриноцитами (клетками Лейдига) яичка, содержат липидные капли, хорошо развитую агранулярную эндоплазматическую сеть, иногда - мелкие кристаллы; вырабатывают андрогены.
Маточная труба
Маточные трубы представляет собой мышечные трубчатые органы, протягивающиеся вдоль широкой связки матки от яичника до матки.
Функции маточных труб: (1) захват овоцита, выделяющегося из яичника при овуляции, и его перенос в направлении матки; (2) создание условий для транспорта спермиев из матки; (3) обеспечение среды, необходимой для оплодотворения и начального развития эмбриона; (5) перенос эмбриона в матку.
Анатомически маточная труба подразделяется на 4 отдела: воронку с бахромкой, открывающуюся в области яичника, расширенную часть - ампулу, узкую часть - перешеек и короткий интрамуральный (интерстициальный) сегмент, расположенный в стенке матки. Стенка маточной трубы состоит из трех оболочек: слизистой, мышечной и серозной (рис. 270 и 271).
Слизистая оболочка образует многочисленные ветвящиеся складки, сильно развитые в воронке и ампуле, где они почти целиком заполняют просвет органа. В перешейке эти складки укорачиваются, а в интерстициальном сегменте превращаются в короткие гребешки (см. рис. 270).
Эпителий слизистой оболочки - однослойный столбчатый, образован клетками двух типов - реснитчатыми и секреторными. В нем постоянно присутствуют лимфоциты.
Собственная пластинка слизистой оболочки - тонкая, образована рыхлой волокнистой соединительной тканью; в бахромке содержит крупные вены.
Мышечная оболочка утолщается от ампулы к интрамуральному сегменту; состоит из нерезко разграниченных толстого внутреннего циркулярного
и тонкого наружного продольного слоев (см. рис. 270 и 271). Ее сократительная активность усиливается эстрогенами и угнетается прогестероном.
Серозная оболочка характеризуется наличием под мезотелием толстого слоя соединительной ткани, содержащего сосуды и нервы (подсерозная основа), а в ампулярном отделе - пучков гладкой мышечной ткани.
Матка
Матка представляет собой полый орган с толстой мышечной стенкой, в котором происходит развитие эмбриона и плода. В ее расширенную верхнюю часть (тело) открываются маточные трубы, суженная нижняя (шейка матки) выступает во влагалище, сообщаясь с ним шеечным каналом. В состав стенки тела матки входят три оболочки (рис. 272): 1) слизистая оболочки (эндометрий), 2) мышечная оболочка (миометрий) и 3) серозная оболочка (периметрий).
Эндометрий в течение репродуктивного периода претерпевает циклическую перестройку (менструальный цикл) в ответ на ритмические изменения секреции гормонов яичником (овариальный цикл). Каждый цикл завершается разрушением и удалением части эндометрия, которые сопровождаются выделением крови (менструальным кровотечением).
Эндометрий состоит из покровного однослойного столбчатого эпителия, который образован секреторными и реснитчатыми эпителиоцитами, и собственной пластинки - стромы эндометрия. Последняя содержит простые трубчатые маточные железы, которые открываются на поверхность эндометрия (рис. 272). Железы образованы столбчатым эпителием (сходным с покровным): их функциональная активность и морфологические особенности существенно меняются в ходе менструального цикла. Строма эндометрия содержит отростчатые фибробластоподобные клетки (способные к ряду превращений), лимфоциты, гистиоциты и тучные клетки. Между клетками располагается сеть коллагеновых и ретикулярных волокон; эластические волокна обнаруживаются лишь в стенке артерий. В эндометрии выделяют два слоя, которые различаются по строению и функции: 1) базальный и 2) функ циональный (см. рис. 272 и 273).
Базальный слой эндометрия прикрепляется к миометрию, содержит донышки маточных желез, окруженные стромой с плотным расположением клеточных элементов. Он мало чувствителен к гормонам, имеет стабильное строение и служит источником восстановления функционального слоя.
Получает питание из прямых артерий, отходящих от радиальных артерий, которые проникают в эндометрий из миометрия. Он содержит проксимальные отделы спиральных артерий, служащих продолжением радиальных в функциональный слой.
Функциональный слой (при его полном развитии) много толще базального; содержит многочисленные железы и сосуды. Он высоко чувствителен к гормонам, под влиянием которых изменяются его строение и функция; в конце каждого менструального цикла (см. ниже) этот слой разрушается, вновь восстанавливаясь в следующем. Снабжается кровью за счет спиральных артерий, которые разделяются на ряд артериол, связанных с капиллярными сетями.
Миометрий - самая толстая оболочка стенки матки - включает три нерезко разграниченных мышечных слоя: 1) подслизистый - внутренний, с косым расположением пучков гладких мышечных клеток; 2) сосудистый - средний, наиболее широкий, с циркулярным или спиральным ходом пучков гладких мышечных клеток, содержащий крупные сосуды; 3) надсосудистый - наружный, с косым или продольным расположением пучков гладких мышечных клеток (см. рис. 272). Между пучками гладких миоцитов располагаются прослойки соединительной ткани. Структура и функция миометрия зависят от женских половых гормонов эстрогенов, усиливающих его рост и сократительную активность, которая угнетается прогестероном. В родах сократительная деятельность миометрия стимулируется гипоталамическим нейрогормоном окситоцином.
Периметрий имеет типичное строение серозной оболочки (мезотелий с подлежащей соединительной тканью); он покрывает матку неполностью - в тех участках, где он отсутствует, имеется адвентициальная оболочка. В периметрии находятся симпатические нервные ганглии и сплетения.
Менструальный цикл - закономерные изменения эндометрия, которые повторяются в среднем каждые 28 суток и условно разделяются на три фазы: (1) менструальную (кровотечения), (2) пролиферации, (3) секреции (см. рис. 272 и 273).
Менструальная фаза (1-4-й дни) в первые два дня характеризуется удалением разрушенного функционального слоя (образовавшегося в предыдущем цикле) вместе с небольшим количеством крови, после чего от всего эндометрия остается лишь базальный слой. Поверхность эндометрия, не покрытая эпителием, в последующие два дня подвергается эпителизации вследствие миграции эпителия из донышек желез на поверхность стромы.
Фаза пролиферации (5-14-й дни цикла) характеризуется усиленным ростом эндометрия (под влиянием эстрогенов, выделяемых растущим фолликулом) с образованием структурно оформленных, но функционально неактивных узких маточных желез, к концу фазы приобретающих штопорообразный ход. Отмечается активное митотическое деление клеток желез и стромы эндометрия. Происходит формирование и рост спиральных артерий, мало извитых в этой фазе.
Фаза секреции (15-28-й дни цикла) и характеризуется активной деятельностью маточных желез, а также изменениями стромальных элементов и сосудов под влиянием прогестерона, выделяемого желтым телом. В середине фазы эндометрий достигает максимального развития, его состояние оптимально для имплантации эмбриона; в конце фазы функциональный слой подвергается некрозу вследствие спазма сосудов. Выработка и выделение секрета маточными железами начинается с 19-го дня и усиливается к 20-22-му. Железы имеют извитой вид, их просвет часто мешковидно растянут и заполнен секретом, содержащим гликоген и гликозаминогликаны. Строма отекает, в ней формируются островки крупных полигональных предецидуальных клеток. Вследствие интенсивного роста спиральные артерии становятся резко извитыми, закручиваясь в виде клубков. В отсутствие наступления беременности вследствие регрессии желтого тела и снижения уровней прогестерона на 23-24-й дни секреция желез эндометрия завершается, ухудшается его трофика и начинаются дегенеративные изменения. Отек стромы уменьшается, маточные железы становятся складчатыми, пилообразными, многие их клетки гибнут. Спиральные артерии спазмируются на 27-й день, прекращая кровоснабжение функционального слоя и вызывая его гибель. Некротизированный и пропитанный кровью эндометрий отторгается, чему способствуют периодические сокращения матки.
Шейка матки имеет строение толстостенной трубки; она пронизана шеечным каналом, который начинается в полости матки внутренним зевом и заканчивается во влагалищной части шейки наружным зевом.
Слизистая оболочка шейки матки образована эпителием и собственной пластинкой и отличается по строению от аналогичной оболочки тела матки. Канал шейки характеризуется многочисленными продольными и поперечными ветвящимися пальмовидными складками слизистой оболочки. Он выстлан однослойным столбчатым эпителием, который вдается в собственную пластинку, образуя
около 100 разветвленных шеечных желез (рис. 274).
Эпителий канала и желез включает два типа клеток: численно преобладающие железистые слизистые клетки (мукоциты) и реснитчатые эпителиоциты. Изменения слизистой оболочки шейки матки в течение менструального цикла проявляются колебаниями секреторной активности шеечных мукоцитов, которая в середине цикла увеличивается примерно в 10 раз. Шеечный канал в норме заполнен слизью (шеечная пробка).
Эпителий влагалищной части шейки матки,
как и во влагалище, - многослойный плоский неороговевающий, содержащий три слоя: базальный, промежуточный и поверхностный. Граница этого эпителия с эпителием шеечного канала - резкая, проходит преимущественно выше наружного зева (см. рис. 274), однако ее расположение непостоянно и зависит от эндокринных влияний.
Собственная пластинка слизистой оболочки шейки матки образована рыхлой волокнистой соединительной тканью с высоким содержанием плазматических клеток, вырабатывающих секреторные IgA, которые переносятся в слизь эпителиальными клетками и обеспечивают поддержание местного иммунитета в женской половой системе.
Миометрий состоит преимущественно из циркулярных пучков гладких мышечных клеток; содержание соединительной ткани в нем значительно выше (особенно во влагалищной части), чем в миометрии тела, сеть эластических волокон развита сильнее.
Плацента
Плацента - временный орган, образующийся в матке во время беременности и обеспечивающий связь между организмами матери и плода, благодаря которой осуществляется рост и развитие последнего.
Функции плаценты: (1) трофическая - обеспечение питания плода; (2) дыхательная - обеспечение газообмена плода; (3) выделительная (экскреторная) - удаление продуктов обмена плода; (4) барьерная - защита организма плода от воздействия токсических факторов, предотвращение попадания в организм плода микроорганизмов; (5) эндокринная - синтез гормонов, обеспечивающих течение беременности, подготовку материнского организма к родам; (6) иммунная - обеспечение иммунной совместимости матери и плода. Принято различать материнскую и плодную части плаценты.
Хориальная пластинка располагается под амниотической оболочкой; она образована во-
локнистой соединительной тканью, которая содержит хориальные сосуды - ветви пупочных артерий и пупочной вены (рис. 275). Хориальная пластинка покрыта слоем фибриноида - однородного бесструктурного оксифильного вещества гликопротеиновой природы, которое образуется тканями материнского и плодного организма и покрывает различные участки плаценты.
Ворсины хориона отходят от хориальной пластинки. Крупные ворсины сильно ветвятся, образуя ворсинчатое дерево, которое погружено в межворсинчатые пространства (лакуны), заполненные материнской кровью. Среди ветвей ворсинчатого дерева в зависимости от калибра, положения в этом дереве и функции выделяют ворсины нескольких типов (крупные, промежуточные и терминальные). Крупные, в частности, стволовые (якорные) ворсины выполняют опорную функцию, содержат крупные ветви пупочных сосудов и регулируют поступление крови плода в капилляры мелких ворсин. Якорные ворсины соединены с децидуальной оболочкой (базальной пластинкой) клеточными колонками, образованными вневорсинчатым цитотрофобластом. Терминальные ворсины отходят от промежуточных и являются областью активного обмена между кровью матери и плода. Образующие их компоненты остаются неизменными, однако соотношения между ними претерпевает существенные изменения в различные сроки беременности (рис. 276).
Строма ворсин образована рыхлой волокнистой соединительной тканью, содержащей фибробласты, тучные и плазматические клетки, а также особые макрофаги (клетки Гофбауэра) и кровеносные капилляры плода.
Трофобласт покрывает ворсины снаружи и представлен двумя слоями - внешним слоем синцитиотрофобластом и внутренним - цитотрофобластом.
Цитотрофобласт - слой одноядерных кубических клеток (клеток Лангханса) - с крупными эухроматическими ядрами и слабо или умеренно базофильной цитоплазмой. Они сохраняют свою высокую пролиферативную активность в течение всей беременности.
Синцитиотрофобласт образуется в результате слияния клеток цитотрофобласта, поэтому он представлен обширной цитоплазмой вариабельной толщины с хорошо развитыми органеллами и многочисленными микроворсинками на апикальной поверхности, а также многочисленными ядрами, которые мельче, чем в цитотрофобласте.
Ворсины в ранние сроки беременности покрыты непрерывным слоем цитотрофобласта и широким слоем синцитиотрофобласта с равномерно распределенными ядрами. Их объемная рыхлая строма незрелого типа содержит отдельные макрофаги и небольшое количество слабо развитых капилляров, находящихся преимущественно в центре ворсинок (см. рис. 276).
Ворсины в зрелой плаценте характеризуются изменениями стромы, сосудов и трофобласта. Строма становится более рыхлой, макрофаги в ней единичны, капилляры имеют резко извитой ход, располагаются ближе к периферии ворсинки; в конце беременности появляются так называемые синусоиды - резко расширенные сегменты капилляров (в отличие от синусоидов печени и костного мозга покрыты непрерывной эндотелиальной выстилкой). Относительное содержание клеток цитотрофобласта в ворсинах уменьшается во второй половине беременности, а их слой утрачивает свою непрерывность, и к родам в нем остаются лишь отдельные клетки. Синцитиотрофобласт истончается, местами образует истонченные участки, приближенные к эндотелию капилляров. Его ядра уменьшаются, часто гиперхромны, образуют компактные скопления (узлы), подвергаются апоптозу и вместе с фрагментами цитоплазмы отделяются в материнский кровоток. Слой трофобласта покрывается снаружи и замещается фибриноидом (см. рис. 276).
Плацентарный барьер - совокупность тканей, разделяющих материнский и плодный кровоток, сквозь которые осуществляется двусторонний обмен веществ между матерью и плодом. В ранние сроки беременности толщина плацентарного барьера максимальна и представлена следующими слоями: фибриноид, синцитиотрофобласт, цитотрофобласт, базальная мембрана цитотрофобласта, соединительная ткань стромы ворсины, базальная мембрана капилляра ворсины, его эндотелий. Толщина барьера существенно снижается к концу беременности в связи с отмеченными выше тканевыми перестройками (см. рис. 276).
Материнская часть плаценты образована базальной пластинкой эндометрия (базальной децидуальной оболочкой), от которой в межворсинчатые пространства отходят соединительнотканные перегородки (септы), не доходящие до хориальной пластинки и не разграничивающие целиком это пространство на отдельные камеры. В децидуальной оболочке содержатся особые децидуальные клетки, которые образуются при беременности из предецидуальных клеток, появляющихся в строме
эндометрия в секреторной фазе каждого менструального цикла. Децидуальные клетки - крупные, овальной или многоугольной формы, с округлым, эксцентрически расположенным светлым ядром и ацидофильной вакуолизированной цитоплазмой, содержащей развитый синтетический аппарат. Эти клетки секретируют ряд цитокинов, факторов роста и гормонов (пролактин, эстрадиол, кортиколиберин, релаксин), которые, с одной стороны, в совокупности ограничивают глубину инвазии трофобласта в стенку матки, с другой - обеспечивают локальную толерантность иммунной системы матери по отношению к аллогенному плоду, что обусловливает успешное течение беременности.
Влагалище
Влагалище - толстостенный растяжимый трубчатый орган, соединяющий преддверие влагалища с шейкой матки. Стенка влагалища состоит из трех оболочек: слизистой, мышечной и адвентициальной.
Слизистая оболочка выстлана толстым многослойным плоским неороговевающим эпителием, лежащим на собственной пластинке (см. рис. 274). Эпителий включает базальный, промежуточный и поверхностный слои. В нем постоянно обнаруживаются лимфоциты, антиген-представляющие клетки (Лангерганса). Собственная пластинка состоит из волокнистой соединительной ткани с большим количеством коллагеновых и эластических волокон и обширным венозным сплетением.
Мышечная оболочка состоит из пучков гладких мышечных клеток, образующих два нечетко разграниченных слоя: внутренний циркулярный и наружный продольный, которые продолжаются в аналогичные слои миометрия.
Адвентициальная оболочка образована соединительной тканью, которая сливается с адвентицией прямой кишки и мочевого пузыря. Содержит крупное венозное сплетение и нервы.
Молочная железа
Молочная железа является частью репродуктивной системы; ее структура существенно варьирует в разные периоды жизни, что обусловлено различиями гормонального фона. У взрослой женщины молочная железа состоит из 15-20 долей - трубчато-альвеолярных желез, которые разграничены тяжами плотной соединительной ткани и, расходясь радиально от соска, далее разделяются на множественные дольки. Между дольками много жировой
ткани. На соске доли открываются млечными протоками, расширенные участки которых (млечные синусы) расположены под ареолой (пигментированным околососковым кружком). Млечные синусы выстланы многослойным плоским эпителием, остальные протоки - однослойным кубическим или столбчатым эпителием и миоэпителиальными клетками. Сосок и ареола содержат большое количество сальных желез, а также пучки радиальных (продольных) гладких мышечных клеток.
Функционально неактивная молочная железа
содержит слабо развитый железистый компонент, который состоит преимущественно из протоков. Концевые отделы (альвеолы) не сформированы и имеют вид терминальных почек. Большая часть органа занята стромой, представленной волокнистой соединительной и жировой тканями (рис. 277). При беременности под влиянием высоких концентраций гормонов (эстрогенов и прогестерона в сочетании с пролактином и плацентарным лактогеном) происходит структурно-функциональная перестройка железы. Она включает резкое разрастание эпителиальной ткани с удлинением и ветвлением протоков, формированием альвеол при уменьшении объема жировой и волокнистой соединительной тканей.
Функционально активная (лактирующая) молочная железа образована дольками, состоящими из концевых отделов (альвеол), заполненных моло-
ком, и внутридольковых протоков; между дольками в прослойках соединительной ткани (междольковых перегородках) располагаются междольковые протоки (рис. 278). Секреторные клетки (галактоциты) содержат развитую гранулярную эндоплазматическую сеть, умеренное число митохондрий, лизосом, крупный комплекс Гольджи (см. рис. 44). Они вырабатывают продукты, которые секретируются различными механизмами. Белок (казеин), а также молочный сахар (лактоза) выделяются мерокринным механизмом путем слияния мембраны секреторных белковых гранул с плазмолеммой. Мелкие липидные капельки сливаются с образованием более крупных липидных капель, которые направляются в апикальную часть клетки и выделяются в просвет концевого отдела вместе с окружающими их участками цитоплазмы (апокринная секреция) - см. рис. 43 и 279.
Выработка молока регулируется эстрогенами, прогестероном и пролактином в сочетании с инсулином, кортикостероидами, гормоном роста и тиреоидными гормонами. Выделение молока обеспечивается миоэпителиальными клетками, которые своими отростками охватывают галактоциты и сокращаются под влиянием окситоцина. В лактирующей молочной железе соединительная ткань имеет вид тонких перегородок, инфильтрированных лимфоцитами, макрофагами, плазматическими клетками. Последние вырабатывают иммуноглобулины класса А, которые транспортируются в секрет.
ОРГАНЫ ЖЕНСКОЙ ПОЛОВОЙ СИСТЕМЫ
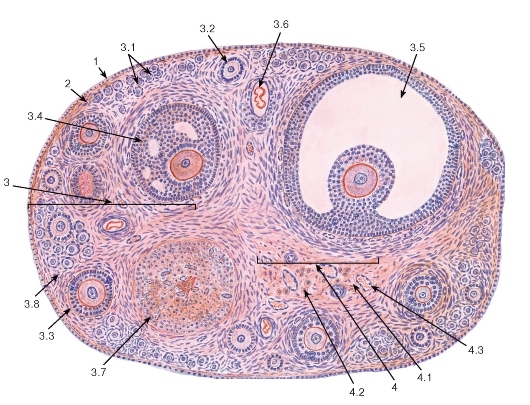
Рис. 264. Яичник (общий вид)
Окраска: гематоксилин-эозин
1 - поверхностный эпителий (мезотелий); 2 - белочная оболочка; 3 - корковое вещество: 3.1 - примордиальные фолликулы, 3.2 - первичный фолликул, 3.3 - вторичный фолликул, 3.4 - третичный фолликул (ранний антральный), 3.5 - третичный (зрелый предовуляторный) фолликул - граафов пузырек, 3.6 - атретический фолликул, 3.7 - желтое тело, 3.8 - строма коркового вещества; 4 - мозговое вещество: 4.1 - рыхлая волокнистая соединительная ткань, 4.2 - хилусные клетки, 4.3 - кровеносные сосуды
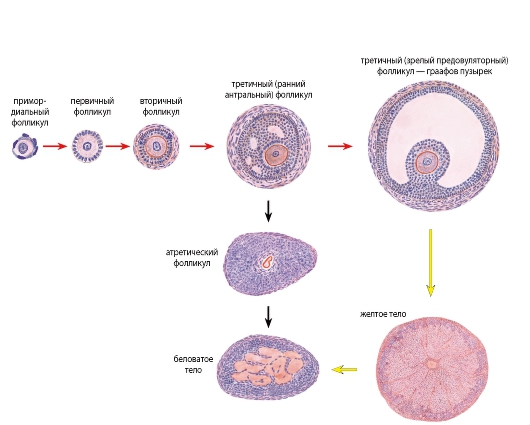
Рис. 265. Яичник. Динамика преобразования структурных компонентов - овариальный цикл (схема)
На схеме показан ход преобразований в процессах овогенеза и фолликулогенеза (красные стрелки), образования и развития желтого тела (желтые стрелки) и атрезии фолликулов (черные стрелки). Конечной стадией преобразования желтого тела и атретического фолликула является беловатое тело (образовано рубцовой соединительной тканью)
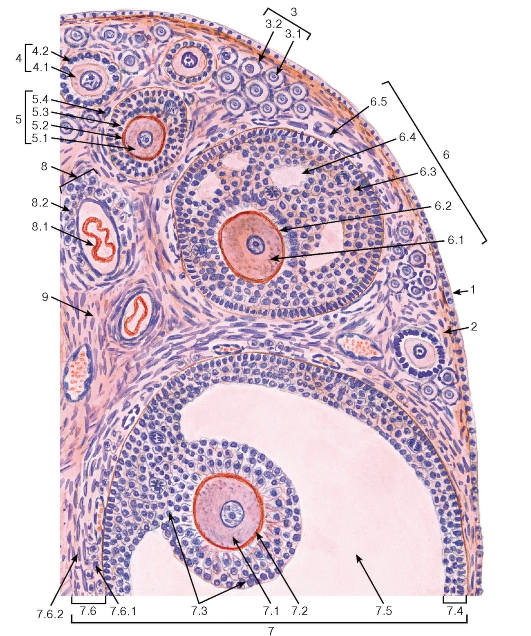
Рис. 266. Яичник. Участок коркового вещества
Окраска: гематоксилин-эозин
1 - поверхностный эпителий (мезотелий); 2 - белочная оболочка; 3 - примордиальные фолликулы:
3.1 - первичный овоцит, 3.2 - фолликулярные клетки (плоские); 4 - первичный фолликул: 4.1 - первичный овоцит, 4.2 - фолликулярные клетки (кубические, столбчатые); 5 - вторичный фолликул: 5.1 - первичный ооцит, 5.2 - прозрачная оболочка, 5.3 - фолликулярные клетки (многослойная оболочка) - гранулеза; 6 - третичный фолликул (ранний антральный): 6.1 - первичный овоцит, 6.2 - прозрачная оболочка, 6.3 - фолликулярные клетки - гранулеза, 6.4 - полости, содержащие фолликулярную жидкость, 6.5 - тека фолликула; 7 - зрелый третичный (предовуляторный) фолликул - граафов пузырек: 7.1 - первичный овоцит,
7.2 - прозрачная оболочка, 7.3 - яйценосный бугорок, 7.4 - фолликулярные клетки стенки фолликула - гранулеза, 7.5 - полость, содержащая фолликулярную жидкость, 7.6 - тека фолликула, 7.6.1 - внутренний слой теки, 7.6.2 - наружный слой теки; 8 - атретический фолликул: 8.1 - остатки овоцита и прозрачной оболочки, 8.2 - клетки атретического фолликула; 9 - рыхлая волокнистая соединительная ткань (строма яичника)
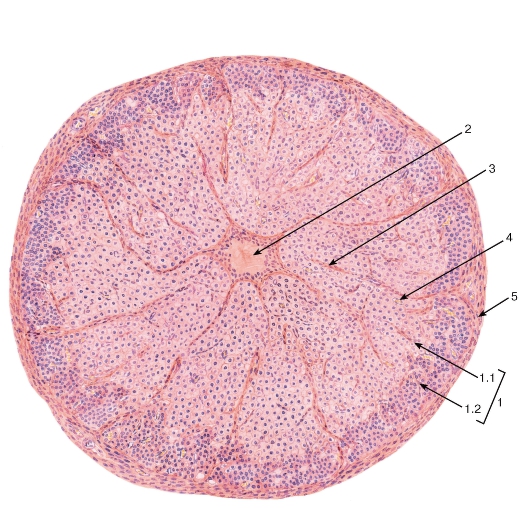
Рис. 267. Яичник. Желтое тело в фазе расцвета
Окраска: гематоксилин-эозин
1 - лютеоциты: 1.1 - гранулезные лютеоциты, 1.2 - лютеоциты теки; 2 - зона кровоизлияния; 3 - прослойки рыхлой волокнистой соединительной ткани; 4 - кровеносные капилляры; 5 - соединительнотканная капсула (уплотнение стромы яичника)
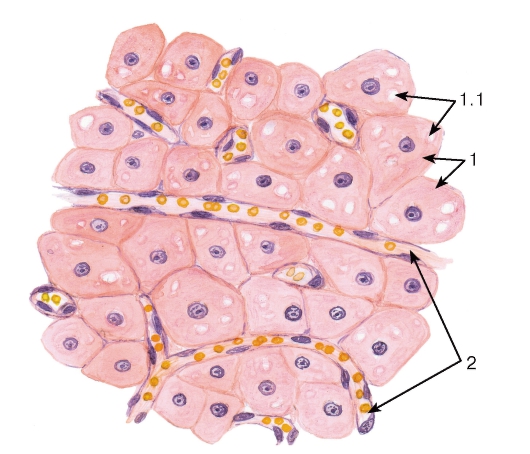
Рис. 268. Яичник. Участок желтого тела
Окраска: гематоксилин-эозин
1 - гранулезные лютеоциты: 1.1 - липидные включения в цитоплазме; 2 - кровеносные капилляры
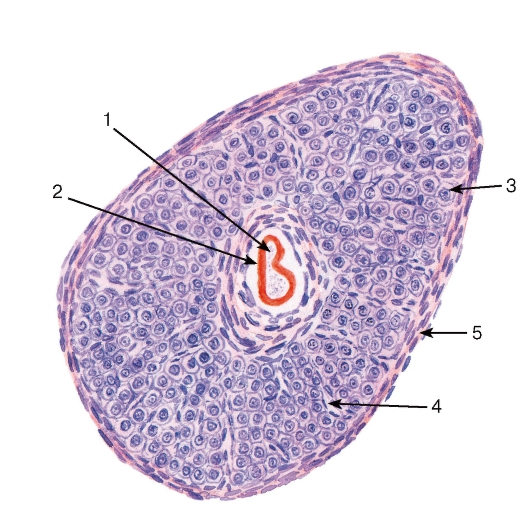
Рис. 269. Яичник. Атретический фолликул
Окраска: гематоксилин-эозин
1 - остатки разрушевшегося овоцита; 2 - остатки прозрачной оболочки; 3 - железистые клетки; 4 - кровеносный капилляр; 5 - соединительнотканнная капсула (уплотнение стромы яичника)
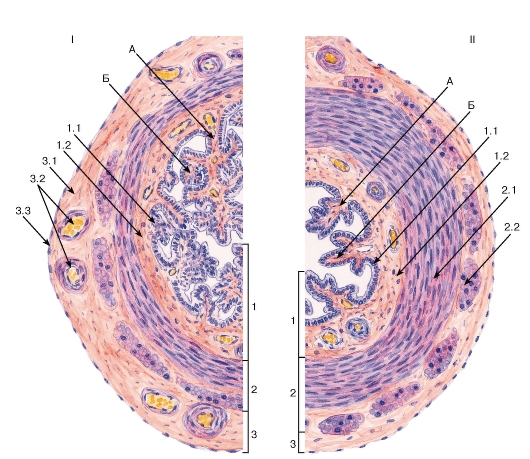
Рис. 270. Маточная труба (общий вид)
I - ампулярная часть; II - перешеек Окраска: гематоксилин-эозин
А - первичные складки слизистой оболочки; Б - вторичные складки слизистой оболочки
1 - слизистая оболочка: 1.1 - однослойный столбчатый реснитчатый эпителий, 1.2 - собственная пластинка; 2 - мышечная оболочка: 2.1 - внутренний циркулярный слой, 2.2 - наружный продольный слой; 3 - серозная оболочка: 3.1 - рыхлая волокнистая соединительная ткань, 3.2 - кровеносные сосуды, 3.3 - мезотелий
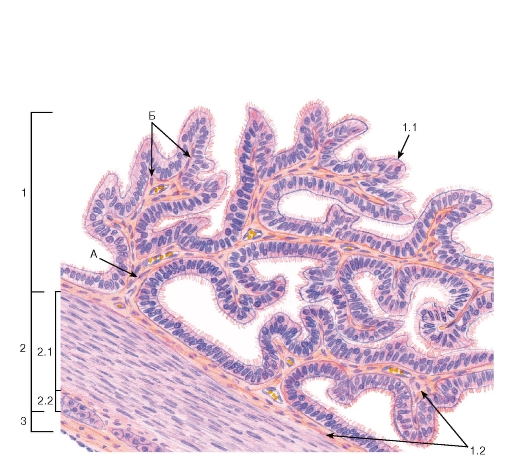
Рис. 271. Маточная труба (участок стенки)
Окраска: гематоксилин-эозин
А - первичные складки слизистой оболочки; Б - вторичные складки слизистой оболочки
1 - слизистая оболочка: 1.1 - однослойный столбчатый реснитчатый эпителий, 1.2 - собственная пластинка; 2 - мышечная оболочка: 2.1 - внутренний циркулярный слой, 2.2 - наружный продольный слой; 3 - серозная оболочка
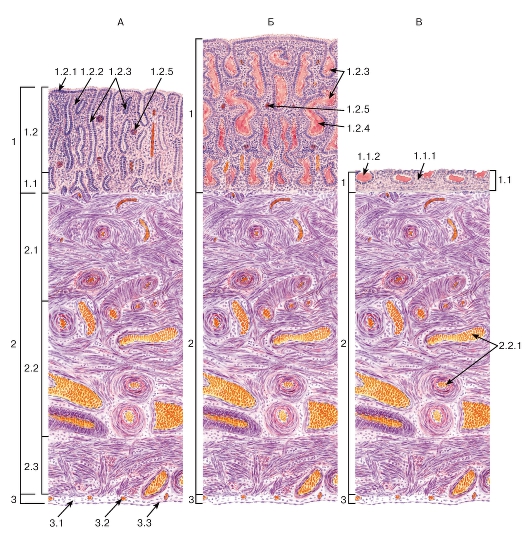
Рис. 272. Матка в различных фазах менструального цикла
Окраска: ШИК-реакция и гематоксилин
А - фаза пролиферации; Б - фаза секреции; В - менструальная фаза
1 - слизистая оболочка (эндометрий): 1.1 - базальный слой, 1.1.1 - собственная пластинка слизистой оболочки (строма эндометрия), 1.1.2 - донышки маточных желез, 1.2 - функциональный слой, 1.2.1 - однослойный столбчатый покровный эпителий, 1.2.2 - собственная пластинка (строма эндометрия), 1.2.3 - маточные железы, 1.2.4 - секрет маточных желез, 1.2.5 - спиральная артерия; 2 - мышечная оболочка (миометрий): 2.1 - подслизистый мышечный слой, 2.2 - сосудистый мышечный слой, 2.2.1 - кровеносные сосуды (артерии и вены), 2.3 - надсосудистый мышечный слой; 3 - серозная оболочка (периметрий): 3.1 - рыхлая волокнистая соединительная ткань, 3.2 - кровеносные сосуды, 3.3 - мезотелий
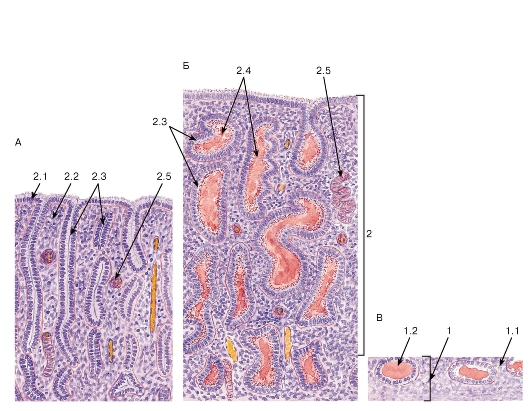
Рис. 273. Эндометрий в различных фазах менструального цикла
Окраска: ШИК-реакция и гематоксилин
А - фаза пролиферации; Б - фаза секреции; В - менструальная фаза
1 - базальный слой эндометрия: 1.1 - собственная пластинка слизистой оболочки (строма эндометрия), 1.2 - донышки маточных желез, 2 - функциональный слой эндометрия, 2.1 - однослойный столбчатый покровный эпителий, 2.2 - собственная пластинка (строма эндометрия), 2.3 - маточные железы, 2.4 - секрет маточных желез, 2.5 - спиральная артерия
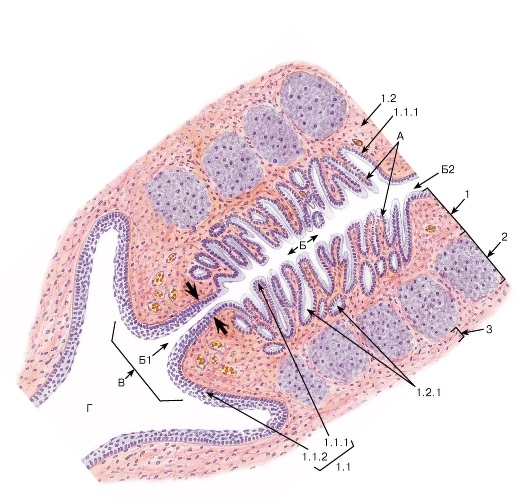
Рис. 274. Шейка матки
Окраска: ШИК-реакция и гематоксилин
А - пальмовидные складки; Б - шеечный канал: Б1 - наружный зев, Б2 - внутренний зев; В - влагалищная часть шейки матки; Г - влагалище
1 - слизистая оболочка: 1.1 - эпителий, 1.1.1 - однослойный столбчатый железистый эпителий шеечного канала, 1.1.2 - многослойный плоский неороговевающий эпителий влагалищной части шейки матки, 1.2 - собственная пластинка слизистой оболочки, 1.2.1 - шеечные железы; 2 - мышечная оболочка; 3 - адвентициальная оболочка
Область «стыка» многослойного плоского неороговевающего и однослойного столбчатого железистого эпителиев показана жирными стрелками
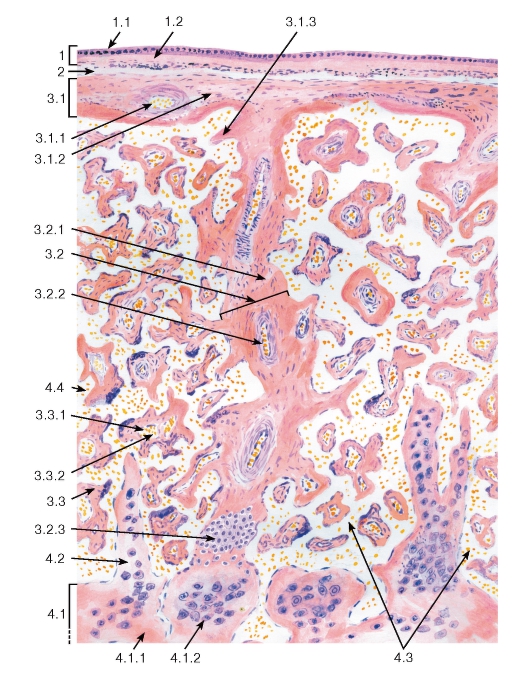
Рис. 275. Плацента (общий вид)
Окраска: гематоксилин-эозин Комбинированный рисунок
1 - амниотическая оболочка: 1.1 - эпителий амниона, 1.2 - соединительная ткань амниона; 2 - амниохориальное пространство; 3 - плодная часть: 3.1 - хориальная пластинка, 3.1.1 - кровеносные сосуды, 3.1.2 - соединительная ткань, 3.1.3 - фибриноид, 3.2 - стволовая («якорная») ворсина хориона,
3.2.1 - соединительная ткань (строма ворсины), 3.2.2 - кровеносные сосуды, 3.2.3 - колонки цитотрофобласта (периферический цитотрофобласт), 3.3 - терминальная ворсина, 3.3.1 - кровеносный капилляр,
3.3.2 - кровь плода; 4 - материнская часть: 4.1 - децидуальная оболочка, 4.1.1 - рыхлая волокнистая соединительная ткань, 4.1.2 - децидуальные клетки, 4.2 - соединительнотканная септа, 4.3 - межворсинчатые пространства (лакуны), 4.4 - материнская кровь
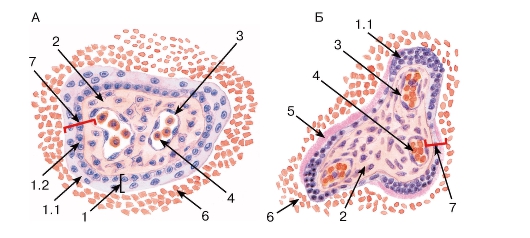
Рис. 276. Терминальные ворсины плаценты
А - ранней плаценты; Б - поздней (зрелой) плаценты Окраска: гематоксилин-эозин
1 - трофобласт: 1.1 - синцитиотрофобласт, 1.2 - цитотрофобласт; 2 - эмбриональная соединительная ткань ворсины; 3 - кровеносный капилляр; 4 - кровь плода; 5 - фибриноид; 6 - кровь матери; 7 - плацентарный барьер
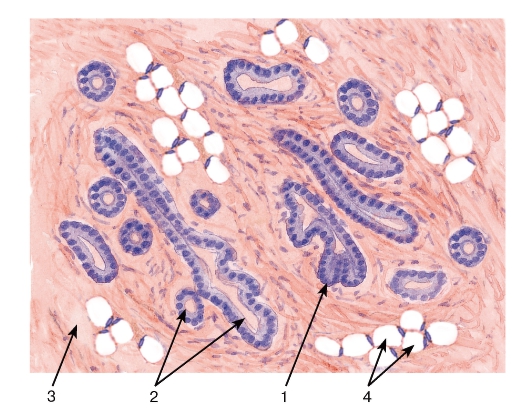
Рис. 277. Молочная железа (нелактирующая)
Окраска: гематоксилин-эозин
1 - терминальные почки (несформированные концевые отделы); 2 - выводные протоки; 3 - соединительнотканная строма; 4 - жировая ткань
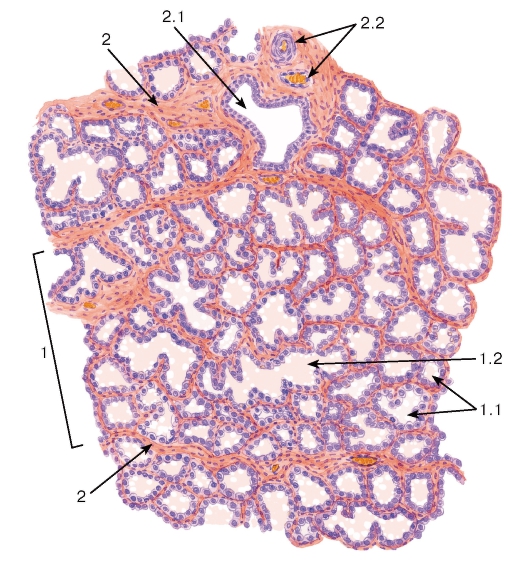
Рис. 278. Молочная железа (лактирующая)
Окраска: гематоксилин-эозин
1 - долька железы, 1.1 - концевые отделы (альвеолы), 1.2 - внутридольковый проток; 2 - междольковые соединительнотканные прослойки: 2.1 - междольковый выводной проток, 2.2 - кровеносные сосуды
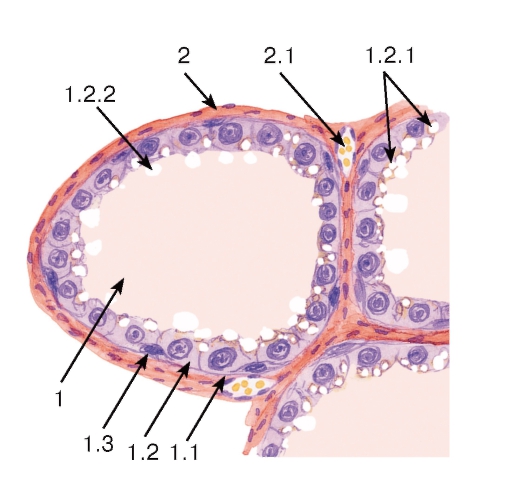
Рис. 279. Молочная железа (лактирующая). Участок дольки
Окраска: гематоксилин-эозин
1 - концевой отдел (альвеола): 1.1 - базальная мембрана, 1.2 - секреторные клетки (галактоциты), 1.2.1 - липидные капли в цитоплазме, 1.2.2 - выделение липидов механизмом апокринной секреции, 1.3 - миоэпителиоциты; 2 - прослойки рыхлой волокнистой соединительной ткани: 2.1 - кровеносный сосуд