Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 5. ФИЗИОЛОГИЯ НЕЙРОНОВ
Нервная система состоит из двух типов клеток - нервных (нейроны) и глиальных (нейроглия). К нейроглии относятся астроциты, микроглия, миелинобразующие клетки (олигодендроциты ЦНС и шванновские клетки периферических нервов). По отношению к нейронам глиоциты выполняют трофическую, опорную и изолирующую (электрический изолятор) функции. Нейрон - основная структурно-функциональная единица нервной системы. Количество нейронов в мозге человека превышает 100х109. Основная функция нейронов - генерация, передача и интеграция нервных импульсов.
Нейроны
В нервной клетке различают тело (перикарион) и отростки - дендриты и аксон (рис. 5-1). Как правило, нейрон имеет несколько дендритов и один аксон. Отростки нейрона и поверхность перикариона участвуют в образовании синапсов. Нейроны - возбудимые клетки - осуществляют передачу электрических сигналов в пределах самой нервной клетки (а между нейронами при помощи нейромедиаторов в синапсах) и этим обеспечивают способность мозга к переработке информации. Сома нейрона синтезирует белки и различные клеточные материалы и распределяет их по дендритам и аксону.
Транспорт веществ в нейроне и его отростках. Транспортные механизмы способны перемещать клеточные компоненты в антероградном направлении в сторону от сомы нейрона или в ретроградном направлении в сторону сомы нейрона. Кинезин и микротубулоассоциированный белок ответственны за антероградный аксональный транспорт органелл и везикул; динеин и другой микротубулоассоциированный белок отвечают за ретроградный транспорт. Антеро-
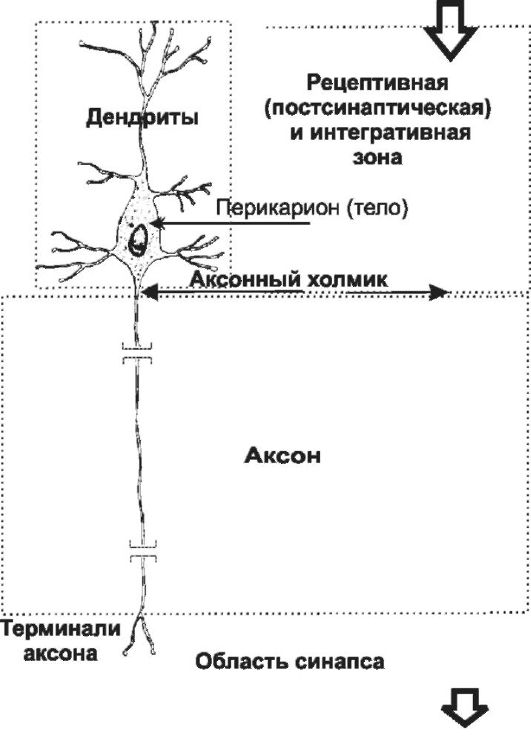
Рис. 5-1. Многоотростчатый нейрон [11]. Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них - аксон, все другие - дендриты. Справа и сверху вниз: функциональные зоны нервной клетки - рецептивная (дендритная), аксон (область проведения ПД), концевых разветвлений аксона (пресинаптическая).
градный транспорт подразделяется на быстрый и медленный. Скорость медленного аксоплазматического транспорта 1-2 мм в день. Скорость быстрого аксоплазматического транспорта около
Классификации нейронов
Существует множество классификаций нервных клеток, так как нейроны различаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в нейронных сетях и по множеству других признаков. В зависимости от класса выполняемой функции выделяют афферентные (чувствительные, сенсорные), эфферентные (двига-
тельные, моторные) и вставочные нервные клетки (ассоциативные нейроны, или интернейроны).
Исходя из характера электрогенеза в различных зонах нервных клеток (рецептивная и интегративная зоны, начальный сегмент ак- сона, аксон, терминали аксона) предложена универсальная классификация (см. рис. 5-1). Эта классификация учитывает направление (вход и выход) сигналов в пределах одной нервной клетки, но находящейся в составе нейронных сетей или цепочек, а также характер мембранного электрогенеза (изменения МП в рецептив- ной и пресинаптической зонах и передача ПД по аксону).
Мембранный потенциал покоя
Мембранный потенциал (МП) покоя - разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя (при отсутствии раздражителей) - рассмотрен в главе 2 (раздел «Мембранный электрогенез»). МП покоя нервной клетки составляет -70 мВ, и его величина, устанавливаемая безворотными ионными каналами, довольно стабильна. Изменения величины МП (рис. 5-2) описываются специальными терминами:
• деполяризация - уменьшение величины МП;
• реполяризация - возвращение величины МП к исходному уровню;
• гиперполяризация - увеличение МП по сравнению с исходным уровнем.
Все изменения МП в дендритной зоне нервной клетки, формируемые при участии воротных каналов, которые управляются лигандами, известны как локальные ответы, затухающие на протяжении нескольких микрометров от места возникновения. В этой зоне постоянно возникают десятки и сотни локальных ответов (постсинаптические потенциалы), суммирование которых в виде изменений МП достигает области начала аксона (аксонного хол- мика). Помимо локальных ответов, возбудимые мембраны нервных клеток в области аксона за счет быстрой активации множества потенциалзависимых натриевых каналов могут генерировать потенциалы действия (ПД, спайки, нервные импульсы, см. рис. 5-2). Существенное значение при генерации ПД имеет представление о пороге раздражения.
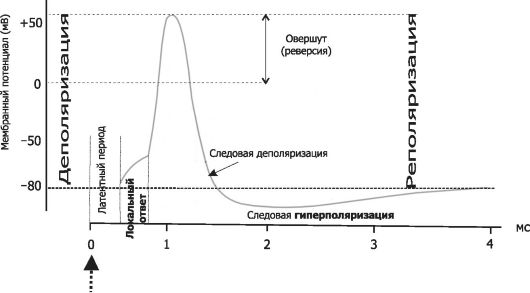
Рис. 5-2. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка - момент появления раздражающего стимула, на отметке -80 мВ - исходный уровень МП.
Локальные ответы, порог раздражения и потенциал действия
Нервная клетка на раздражающие (в том числе электрические) стимулы отвечает изменениями МП: либо реагирует локальными (подпороговыми) реакциями, либо генерирует ПД.
• Локальные ответы. Стимулы, величина которых меньше опре- делённого порога, вызывают локальные ответы, затухающие вблизи места раздражения. Эти локальные ответы могут быть как деполяризующими (возбуждающие) мембрану, так и гиперполяризующими (тормозные).
• Потенциал действия. Деполяризующие стимулы, достигшие порогового значения, вызывают развитие возбуждения в виде ПД. ПД распространяется по мембране нейрона без затухания (без декремента).
Законы раздражения нервной клетки
Генерация ПД подчиняется законам «всё или ничего», «сила- время» и аккомодации.
• Закон «всё или ничего». При достижении пороговой силы раздражающего стимула дальнейшее увеличение его интенсивности или продолжительности раздражения не изменяет ха-
рактеристик ПД. Такая форма реакции нервной клетки на раздражение получила название «всё или ничего» (либо ПД возникает, либо нет).
• Закон силы-времени. Эффект раздражителя на нервную клетку зависит не только от силы раздражителя, но и от времени, в течение которого он действует; так, чем больше сила тока, тем меньше времени он должен действовать, чтобы возник процесс возбуждения, и наоборот. Соотношение силы и длительности действия раздражителя может быть выражено в виде гиперболической кривой (рис. 5-3).
• Закон аккомодации. Раздражающее действие тока зависит не только от амплитуды электрического сигнала, но и от скорости (крутизны) его нарастания во времени: чем быстрее это нарастание, тем сильнее выражено раздражающее действие тока. При действии медленно нарастающей силы раздражителя генерации ПД не происходит, так как возбудимая клетка
в
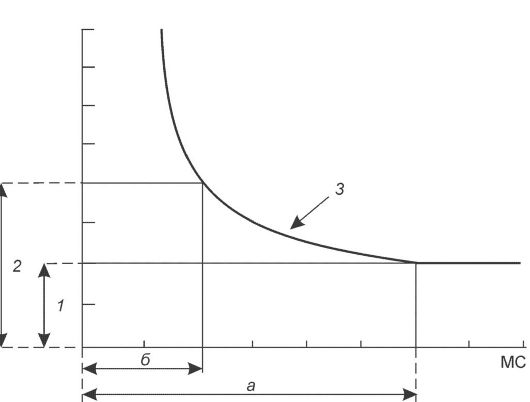
Рис. 5-3. Кривая зависимости силы раздражения возбудимой ткани от продолжительности действия раздражителя «сила-длительность» [21]. 1 - реобаза (минимальная сила раздражителя, способная вызвать ПД); 2 - удвоенная реобаза; 3 - кривая «сила-длительность»; а - полезное время (минимальное время, в течение которого ток, равный одной реобазе, вызывает генерацию ПД); б - хронаксия (минимальное время, в течение которого ток, равный двум реобазам, вызывает ПД).
«приспосабливается» к действию этого раздражителя, что и получило название аккомодации.
Потенциал действия
Потенциал действия (ПД) - быстрое колебание МП - самораспространяющийся процесс, связанный с изменениями ионной проводимости мембраны, которые вызваны функционированием потенциалзависимых натриевых каналов. ПД распространяется без затухания, т.е. практически без уменьшения амплитуды. Проведение ПД по мембране можно сравнить с поджиганием пороховой дорожки: вспыхнувший порох немедленно воспламеняет впереди лежащие частицы, и пламя движется вперёд до конца дорожки.
Временной ход потенциала действия. Продолжительность ПД нервной клетки измеряется единицами миллисекунд (мс). ПД, зарегистрированные двумя электродами, один из которых находится внутри клетки, а другой - в окружающем растворе, представлены на рис. 5-2, 5-4.
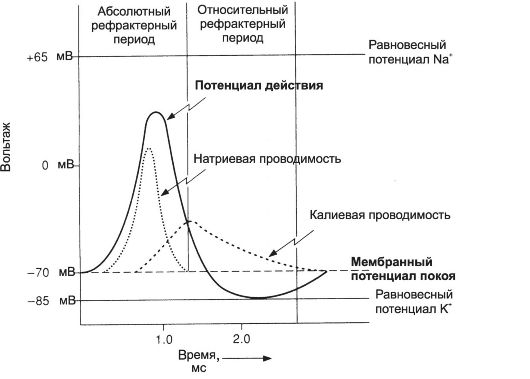
Рис. 5-4. Потенциал действия и ионные токи нервной клетки [5]. По оси
ординат отложены значения МП (мВ), по оси абсцисс - время (мс).
Между моментом, когда наносится раздражение, и первым проявлением ПД существует задержка - латентный период. Латент- ный период соответствует времени, когда ПД движется по мембране нервной клетки от места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны - локальный ответ. При достижении критического уровня деполяризации, который в среднем составляет -55 мВ, начинается фаза деполяризации. В этой фазе уровень МП падает до нуля и даже приобретает положительное значение (овершут), а затем возвращается к исходному уровню (фаза реполяризации). Фазы деполяризации, овершута и реполяризации образуют спайк (пик) ПД. Длительность спайка составляет 1-2 мс. После спайка наблюдается замедление скорости спада потенциала - фаза следовой деполяризации. После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации. Эти следовые потенциалы могут длиться десятки и сотни миллисекунд.
Ионные механизмы потенциала действия. В течении ПД происходят изменения МП, обусловленные ионными механизмами. На рис. 5-4 представлены суммарные ионные токи, протекающие через мембрану нервной клетки в ходе ПД.
• Локальный потенциал. Раздражение клетки обусловливает открытие части потенциалзависимыхNa+-каналов и появление локального (нераспространяющегося) потенциала.
• Фаза деполяризации. При достижении критического уровня деполяризации мембраны (соответствует порогу активации потенциалзависимых Na+-каналов) начинается лавинообразный процесс открытия множества Na+-каналов. В фазе деполяризации происходит массивный вход в клетку ионов Na+ по концентрационному и электрохимическому градиенту.
• Овершут. Деполяризация мембраны приводит к реверсии мембранного потенциала (МП становится положительным). В фазе овершута Na+-ток начинает стремительно спадать, что связано с инактивацией потенциалзависимых Na+-каналов (время открытого состояния этих каналов - доли миллисекунды) и дополнительным открытием К+-каналов.
• Фаза реполяризации. Помимо инактивацииNa+-каналов, развитию реполяризации способствует открытие потенциалзависимых К+-каналов. Этот процесс происходит медленнее, чем
открытие Na+-каналов, но К+-каналы остаются открытыми более продолжительное время. Выход К+ наружу способствует полному завершению фазы реполяризации.
• Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов, что способствует выходу ионов К+. Восстановление исходного уровня МП приводит Na+- и К+-каналы в состояние покоя.
Изменения возбудимости во время потенциала действия. В ходе развития ПД изменяется возбудимость мембраны нервной клетки.
• Абсолютно рефрактерный период. Во время фазы деполяризации и частично фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинстваNa+-каналов. Абсолютно рефрактерный период ограничивает частоту возникновения ПД и возможность его проведения в неверном направлении по аксону.
• Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов.
• Лабильность - максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с.
Проведение возбуждения по нервным волокнам
Нервные волокна - аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских клеток в периферических нервах. Нервные волокна подразделяются на два типа - безмиелиновые и миелиновые. Основная функция нервных волокон - проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5-5) и в значительной мере зависит от диаметра нервных волокон.
• Безмиелиновые нервные волокна (рис. 5-6, А). В покое мембрана аксона (осевого цилиндра) поляризована - положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется и наружная поверхность мембраны при-
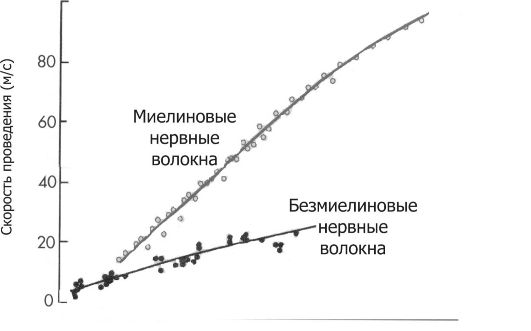
Рис. 5-5. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
обретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны. Такое прове- дение известно как электротоническое, а проведение ПД - своего рода «эстафета», в которой каждый участок мембраны является сначала раздражаемым, а затем раздражающим. ПД возникает за счёт увеличивающейся проводимости через потенциалзависимые Na+-каналы, встроенные в аксолемму с плотностью около 110-120 каналов на 1 мкм2. Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении. Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5-2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения
ПД (см. рис. 5-5).
Миелиновое нервное волокно (рис.
5-6, Б) состоит из осевого цилиндра (аксона), вокруг которого
шванновские клетки образуют миелин за счёт концентрического наслаивания
собственной плазматической мембраны. Миелин прерывается через
регулярные промежутки (от 0,2 до
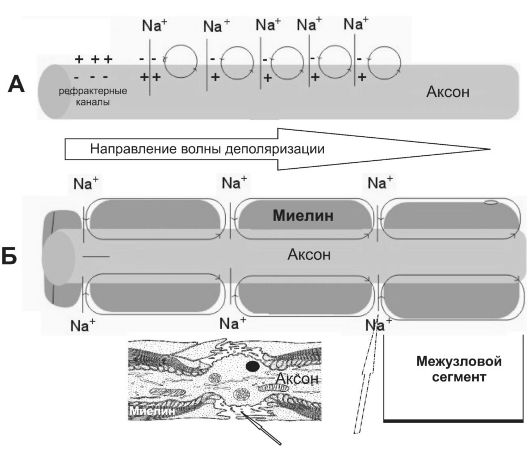
Рис. 5-6. Проведение возбуждения в нервных волокнах [7]. А - безмиелиновое волокно (электротоническое проведение); Б - миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье - проводник.
Законы проведения возбуждения
• Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, т.е. проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени.
• Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена:
Φ наличием оболочек, окружающих отдельные нервные волокна и их пучки (в результате образуется барьер, предупреждающий переход возбуждения с волокна на волокно);
Φ сопротивлением межклеточной жидкости (жидкость, находящаяся между волокнами, имеет гораздо меньшее сопротивление току, чем мембрана аксонов; поэтому ток шунтируется по межволоконным пространствам и не доходит до соседних волокон).
Типы нервных волокон и их функции
При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в
Таблица 5-1. Классификация нервных волокон по диаметру и скорости проведения [3]
Структуры

Окончание табл. 5-1
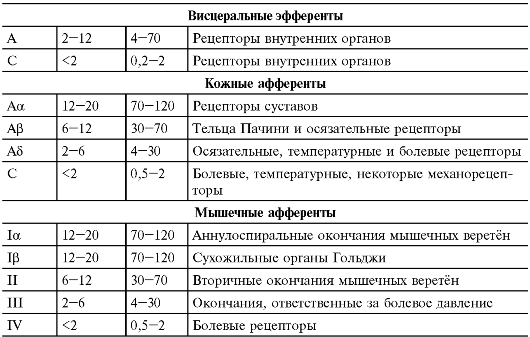
Примечание. Классификация Эрлангера-Гассера (латинские буквы) и альтернативные добавления (римские цифры).
Обобщение главы
Нейроны имеют разветвлённую систему дендритов и, как правило, один протяжённый аксон.
Локализация ионных каналов носит избирательный характер: на нейроне каналы, не имеющие ворот, распределены на нейрональной мембране; потенциалзависимые каналы, в основном, располагаются в аксоне и его отростках.
Потенциал действия представляет собой преходящее изменение мембранного потенциала, характеризующееся быстрой деполяризацией и последующей реполяризацией.
Фаза деполяризации - результат быстрой активации потенциалзависимых Na+-каналов; фаза реполяризации - результат инактивации потенциалзависимых Na+-каналов и задержанной активации потенциалзависимых К+-каналов.
Потенциал действия в нейроне возникает тогда, когда аксонный холмик деполяризуется до порогового уровня быстрой активацией большого количества потенциалзависимых натриевых каналов.
Распространение потенциала действия происходит за счёт постоянного его возникновения в соседних участках мембраны вследствие локального тока, возникающего между возбуждённым и невозбуждённым отделами.
Скорость проведения потенциала действия зависит от диаметра аксона и толщины миелиновой оболочки.
Антероградный аксональный транспорт обеспечивает терминали аксона органеллами, синаптическими везикулами, метаболическими факторами и рецепторами.
Ретроградный аксональный транспорт переносит отработанные вещества, захватывает питательные материалы и сигнальные по- средники из области терминали для сомы нейрона и служит механизмом для входа в ЦНС токсинов и микроорганизмов.