Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 10. ЗРЕНИЕ
Мозг через орган зрения получает более 90% сенсорной информации. Фоторецепторы сетчатки глаза из всего спектра электро- магнитных излучений регистрируют только волны длиной от 400 до 800 нм. Физиологическая роль глаза как органа зрения двояка. Во-первых, это оптический инструмент, собирающий свет от объектов внешней среды и проецирующий их изображения на сетчатку. Во-вторых, фоторецепторы сетчатки преобразуют оптические изображения в нервные сигналы, передаваемые в зрительную кору.
Орган зрения (рис. 10-1) включает глазное яблоко, соединённое через зрительный нерв с мозгом, защитный аппарат (в том числе веки и слёзные железы) и аппарат движения (поперечнополосатые глазодвигательные мышцы).
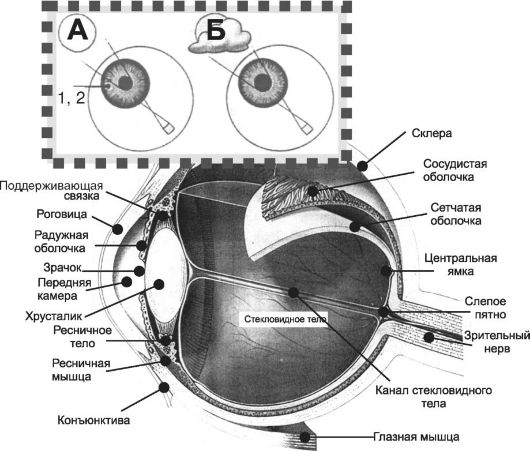
Рис.10-1. Глазное яблоко. На врезке - зрачковый рефлекс (пояснения в тексте).
• Глазное яблоко. Стенка глазного яблока образована оболочками: в передней части расположены конъюнктива и роговица, в задней - сетчатка, сосудистая оболочка и склера. Стекловидное тело занимает полость глазного яблока. Кпереди от стекловидного тела расположен двояковыпуклый хрусталик. Между роговицей и хрусталиком находятся содержащие водянистую влагу перeдняя камера (между задней поверхностью роговицы и радужкой со зрачком) и задняя камера глаза (между радужкой и хрусталиком).
• Защитный аппарат глаза. Длинные ресницы верхнего века предохраняют глаз от попадания пыли; мигательный рефлекс (моргание) осуществляется автоматически. Веки содержат мейбомиевы железы, благодаря которым края век всегда увлажнены. Конъюнктива - тонкая слизистая оболочка - выстилает внутреннюю поверхность век и наружную поверхность глазного яблока. Слёзная железа выделяет слёзную жидкость, которая орошает конъюнктиву.
Сетчатка
Схема зрительного отдела сетчатки представлена на рис. 10-2. У заднего края оптической оси глаза сетчатка имеет округлое жёл- тое пятно диаметром около
• Наружный ядерный слой содержит ядросодержащие части фоторецепторных клеток - колбочек и палочек. Колбочки концентрируются в области жёлтого пятна. Глазное яблоко организовано таким образом, что именно на колбочки падает центральная часть светового пятна от визуализируемого объекта. По периферии от жёлтого пятна расположены палочки.
• Наружный сетчатый. Здесь осуществляются контакты внутренних сегментов палочек и колбочек с дендритами биполярных клеток.
• Внутренний ядерный. Здесь располагаются биполярные клетки, связывающие палочки и колбочки с ганглиозными клетками, а также горизонтальные и амакринные клетки.
• Внутренний сетчатый. В нём биполярные клетки контактируют с ганглиозными клетками, а амакринные клетки выступают в качестве вставочных нейронов.
• Ганглиозный слой содержит тела ганглиозных нейронов.
• Общая схема передачи информации в сетчатке такова: рецепторная клетка - биполярная клетка - ганглиозная клетка и одновременно амакринная клетка - ганглиозная клетка - аксоны ганглиозных клеток. Зрительный нерв выходит из глаза в области, видимой в офтальмоскоп как диск зрительного нерва (см. рис. 10-2, врезка).
Фоторецепторные клетки (рис. 10-3; 10-5, В) - палочки и колбочки. Периферические отростки фоторецепторных клеток состоят из наружного и внутреннего сегментов, соединённых ресничкой.
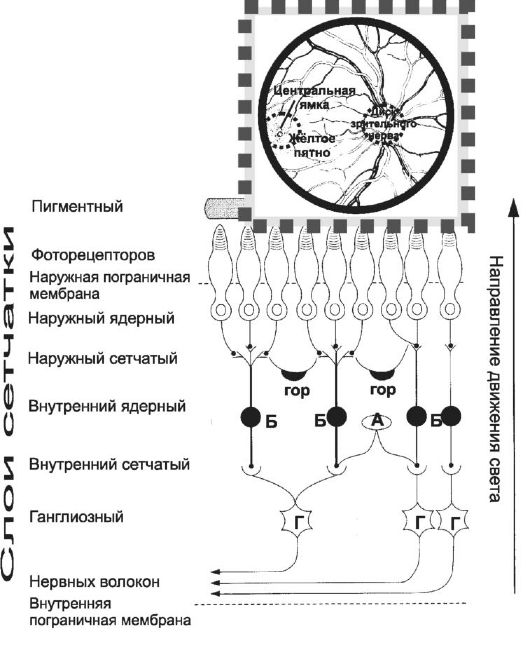
Рис.10-2. Сетчатка. Амакринные (А), биполярные (Б), ганглиозные (Г), горизонтальные (гор) клетки. На врезке - глазное дно.
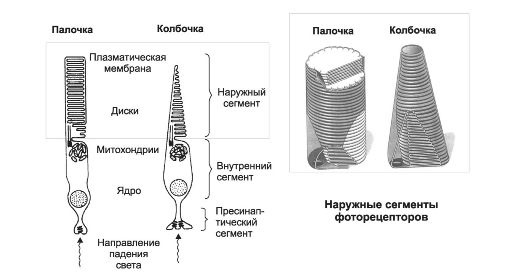
Рис.10-3. Фоторецепторы сетчатки. Наружные сегменты заключены в прямоугольник. Пояснения в тексте.
• Наружный сегмент имеет множество уплощённых замкнутых дисков (дупликатуры клеточных мембран), содержащих зрительные пигменты: родопсин (максимум поглощения 505 нм) - в палочках; красный (570 нм), зелёный (535 нм) и синий (445 нм) пигменты - в колбочках. Наружный сегмент палочек и колбочек состоит из регулярных мембранных образований - дисков (см. рис. 10-3, справа). В каждом фоторецепторе имеется более 1000 дисков.
• Внутренний сегмент заполнен митохондриями и содержит базальное тельце, от которого в наружный сегмент отходит девять пар микротрубочек.
• Центральное зрение, а также острота зрения обеспечиваются колбочками.
• Периферическое зрение, а также ночное зрение и восприятие подвижных объектов - функции палочек.
ОПТИКА ГЛАЗА
Глаз имеет систему линз с различной кривизной и различными показателями преломления световых лучей (рис. 10-4, I), вклю- чающую четыре преломляющие среды между:
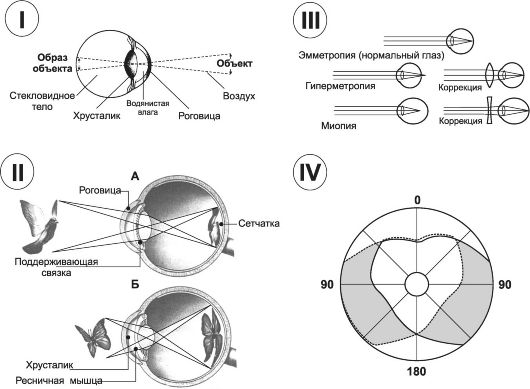
Рис. 10-4. Оптика глаза. I - глаз как оптическая система; II - механизм аккомодации: А - удалённый объект; Б - близко расположенный объект; III - рефракция; IV - поля зрения. Прерывистой линией очерчено поле зрения левого глаза, сплошной линией - поле зрения правого глаза. Светлая (сердцевидная) область в центре - зона бинокулярного зрения. Окрашенные области слева и справа - поля монокулярного зрения.
Φ воздухом и передней поверхностью роговицы; Φ задней поверхностью роговицы и водянистой влагой передней камеры;
• Φ водянистой влагой передней камеры и хрусталиком; Φ задней поверхностью хрусталика и стекловидным телом. Преломляющая сила. Для
практических расчётов преломляющей силы глаза используют понятие о так
называемом редуцированном глазе, когда все преломляющие поверхности
алгебраически складываются и рассматриваются как одна линза. В таком
редуцированном глазу с единственной преломляющей поверхностью,
центральная точка которой располагается на
преломления
составляет 59 диоптрий, когда хрусталик приспособлен для
рассматривания далёких предметов. Преломляющая сила любых оптических
систем выражается в диоптриях (D): 1 диоптрия равна преломляющей силе
линзы с фокусным расстоянием в
• Аккомодация - приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии. Основную роль в процессе аккомодации играет хрусталик, способный изменять свою кривизну. У молодых людей преломляющая сила хрусталика может увеличиваться от 20 до 34 диоптрий. При этом хрусталик изменяет форму от умеренно до значительно выпуклой. Механизм аккомодации проиллюстрирован на рис. 10-4, II. При взгляде на удалённые предметы (А) ресничные мышцы расслабляются, поддерживающая связка растягивает и уплощает хрусталик, придавая ему дискообразную форму. При взгляде на близкие предметы (Б) для полной фокусировки необходима более значительная кривизна хрусталика, поэтому ГМК ресничного тела сокращаются, связки расслабляются, а хрусталик в силу своей эластичности становится более выпуклым.
• Острота зрения - точность, с которой виден объект; теоретически объект должен быть такого размера, чтобы он мог простимулировать одну палочку или колбочку. Оба глаза действуют вместе (бинокулярное зрение), передавая зрительную информацию в зрительные центры коры больших полушарий, где зрительный образ оценивается в трёх измерениях.
• Зрачковый рефлекс. Зрачок
- круглое отверстие в радужной оболочке - очень быстро меняется в
размере в зависимости от количества света, падающего на сетчатку.
Просвет зрачка может изменяться от 1 до
чтобы достаточное количество света могло достичь сетчатки и возбудить фоторецепторы.
• Содружественная реакция зрачков. У здоровых людей зрачки обоих глаз одинакового размера. Освещение одного глаза вызывает сужение зрачка и другого глаза. Такая реакция называется содружественной реакцией зрачков. При некоторых заболеваниях размеры зрачков обоих глаз различны (анизокория).
• Глубина фокуса. Зрачок усиливает чёткость изображения на сетчатке за счёт увеличения глубины резкости. При ярком свете зрачок имеет диаметр
• Контроль фокусировки. Аккомодация хрусталика регулируется механизмом отрицательной обратной связи, автоматически приспосабливая фокусную силу хрусталика к наивысшей остроте зрения. Когда глаза фиксированы на отдалённом объекте и должны немедленно переключиться на фиксацию ближнего предмета, в течение долей секунды происходит аккомодация хрусталика, что улучшает остроту зрения. Если неожиданно изменить точку фиксации, хрусталик всегда изменяет свою преломляющую силу в нужном направлении. Помимо вегетативной иннервации радужки (зрачковый рефлекс), для контроля фокусировки важны следующие моменты.
Φ Хроматическая аберрация. Лучи красного цвета фокусируются позже голубого, поскольку хрусталик преломляет голубые лучи сильнее, чем красные. У глаз появляется возможность определять, какой из этих двух типов лучей находится в лучшем фокусе, и посылать информацию к ак- комодационному механизму с указанием делать хрусталик сильнее или слабее.
Φ Сферическая аберрация. Пропуская только центральные лучи, зрачок устраняет сферическую аберрацию.
Φ Конвергенция глаз при фиксации на близком предмете. Нервный механизм, вызывающий конвергенцию, одновре-
менно сигнализирует об увеличении преломляющей силы хрусталика.
Φ Степень аккомодации хрусталика постоянно, но незначительно колеблется дважды в секунду, что способствует более быстрому реагированию хрусталика для установки фокуса. Зрительный образ становится более чётким, когда осцилляции хрусталика усиливают изменения в нужном направлении; чёткость уменьшается, когда сила хрусталика изменяется в ненужном направлении.
Φ Области мозговой коры, управляющие аккомодацией, взаимодействуют с нервными структурами, которые контролируют фиксацию глаз на движущемся предмете. Окончательная интеграция зрительных сигналов осуществляется в полях 18 и 19 по Бродману, затем двигательные сигналы передаются к ресничной мышце через мозговой ствол и ядра Эдингера-Вестфаля.
Φ Точка ближайшего видения -
способность ясно видеть в фокусе близлежащий предмет - отдаляется в
течение жизни. В десятилетнем возрасте она приблизительно равняется
9-10 см и отдаляется до
• Пресбиопия. Когда человек становится старше, хрусталик разрастается, становится более толстым и менее эластичным. Способность хрусталика изменять свою форму также уменьшается. Сила аккомодации падает с 14 диоптрий у ребёнка до менее 2 диоптрии у человека в возрасте 45-50 лет и до 0 - в возрасте 70 лет. Таким образом, хрусталик утрачивает способность к аккомодации, и это состояние называется пресбиопией (старческая дальнозоркость). Когда человек достигает состояния пресбиопии, каждый глаз остаётся с постоянным фокусным расстоянием, которое зависит от физических характеристик глаз каждого отдельного человека. Поэтому пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами.
• Аномалии рефракции. Эмметропия (нормальное зрение, рис. 10-4, III) соответствует нормальному глазу, если параллельные лучи от отдалённых предметов фокусируются на сет-
чатке, когда ресничная мышца полностью расслаблена. Это значит, что эмметропический глаз может видеть все отдалён- ные объекты очень ясно и легко переходить (посредством аккомодации) на ясное видение близлежащих предметов. Гиперметропия (дальнозоркость) бывает обусловлена слишком коротким глазным яблоком или в более редких случаях тем, что хрусталик глаза слишком малоэластичен. В дальнозорком глазу продольная ось глаза короче, и луч от отдалённых предметов фокусируется за сетчаткой (см. рис. 10-4, III). Дальнозоркий человек компенсирует этот недостаток рефракции аккомодационным усилием. Он напрягает аккомодационную мышцу, рассматривая далёкие объекты. Попытки рассматривать близкие предметы вызывают чрезмерное напряжение аккомодации. Для работы с близко расположенными предметами и чтения дальнозоркие люди должны пользоваться очками с двояковыпуклыми линзами. Миопия (близорукость) представляет тот случай, когда ресничная мышца полностью расслаблена и лучи света от далёкого объекта фокусируются впереди сетчатки (см. рис. 10-4, III). Близорукость возникает либо вследствие слишком длинного глазного яблока, либо в результате большой преломляющей силы хрусталика глаза. Не существует механизма, посредством которого глаз смог бы уменьшить преломляющую силу хрусталика в условиях полностью расслабленной ресничной мышцы. Однако если объект находится рядом с глазами, близорукий человек может использовать механизм аккомодации, чтобы объект чётко фокусировался на сетчатке. Следовательно, у близорукого человека существуют ограничения только в отношении ясной точки «дальнего видения». Для ясного видения вдаль близорукому человеку необходимо использовать очки с двояковогнутыми линзами. Астигматизм - неодинаковое преломление лучей в разных направлениях, вызванное различной кривизной сферической поверхности роговицы. Аккомодация глаза не в силах преодолеть астигматизм, потому что кривизна хрусталика при аккомодации изменяется одинаково. Чтобы компенсировать недостатки рефракции роговицы, применяют специальные цилиндрические линзы.
• Зрительное поле и бинокулярное зрение
Зрительное поле каждого глаза - часть внешнего пространства, видимого глазом. Теоретически оно должно быть круглым, но в действительности оно срезано в медиальном направлении носом и верхним краем глазницы (рис. 10-4, IV). Составление карты зрительного поля важно для неврологической и офтальмологической диагностики. Окружность зрительного поля определяют с помощью периметра. Один глаз закрывается, а другой фиксируется на центральной точке. Передвигая по меридианам в направлении к центру небольшую мишень, отмечают точки, когда мишень становится видимой, описывая таким образом зрительное поле. На рис. 10-4, IV центральные зрительные поля очерчены по касательной линии сплошными и пунктирными линиями. Белые участки за пределами линий являются слепым пятном (физиологическая скотома).
Бинокулярное зрение. Центральная часть зрительных полей двух глаз полностью совпадает; следовательно, любой участок в этом зрительном поле охватывается бинокулярным зрением. Импульсы, идущие от двух сетчаток, возбуждённых световыми лучами от объекта, на уровне зрительной коры сливаются в один образ. Точки на сетчатке обоих глаз, куда должно попадать изображение, чтобы восприниматься бинокулярно как единый предмет, называются корреспондирующими точками. Лёгкое надавливание на один глаз вызывает двоение в глазах, так как нарушается соответствие сетчаток.
Глубина зрения. Бинокулярное зрение играет важную роль в определении глубины зрения, так как основано на относительных размерах объектов, их отражениях, их движении относительно друг друга. На самом деле глубина воспри- ятия является компонентом и монокулярного зрения, но благодаря бинокулярному зрению усиливается чёткость и пропорциональность восприятия глубины.
ФУНКЦИИ СЕТЧАТКИ
Фоторецепция
В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин (рис. 10-5, А)
состоит из белковой части (опсин) и хромофора - 11-цис-ретиналя, под действием фотонов переходящего в транс-ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события (рис. 10-5, Б): активация родопсина в результате фотоизомеризации - каталитическая активация G-белка (Gt, трансдуцин) родопсином - активация фосфодиэстеразы при связывании с
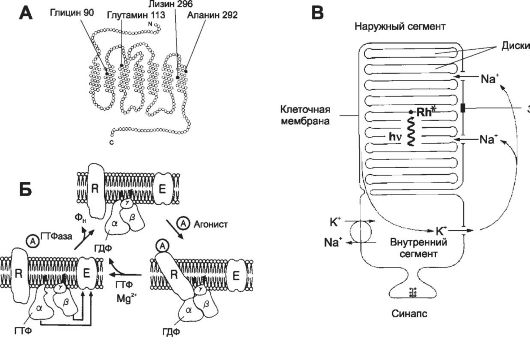
Рис. 10-5. Родопсин и активация ионных каналов [11]. А - молекула опсина содержит семь трансмембранных альфа-спиральных участков. Чёрными кружками отмечена локализация наиболее распространённых молекулярных дефектов. Так, при одной из мутаций глицин во втором трансмембранном участке в 90-м положении заменён на аспарагин, что приводит к врождённой ночной слепоте. Б - трансмембранный белок родопсин и его связь с G-белком (трансдуцин) в плазмолемме фоторецепторной клетки. Возбуждённый фотонами родопсин активирует G-белок. При этом гуанозиндифосфат, связанный с α-СЕ G-белка, заменяется на ГТФ. От- щеплённые α-СЕ и β-СЕ действуют на фосфодиэстеразу и заставляют её превращать цГМФ в гуанозинмонофосфат. Это закрывает Na+-каналы, и ионы Na+ не могут попасть в клетку, что вызывает её гиперполяризацию. R - родопсин; α, β и γ - СЕ G-белка; A - агонист (в данном случае кванты света); E - фермент-эффектор фосфодиэстеразы. В - схема палочки. В наружном сегменте расположена стопка дисков, содержащих зрительный пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет (hv) активирует родопсин (Rh*) в дисках, что закрывает Na+- каналы в клеточной мембране и уменьшает вход Na+ в клетку.
Gta - гидролиз цГМФ цГМФ-фосфодиэстеразой - переход цГМФзависимых Nа+-каналов из открытого состояния в закрытое - гиперполяризация плазмолеммы фоторецепторной клетки - передача сигнала на биполярные клетки.
• Ионные основы фоторецепторных потенциалов В темноте Na+-каналы, входящие в мембрану наружных сегментов палочек и колбочек, открыты и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов (рис. 10-5, В и 10-6, I). Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+-, К+-насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие,
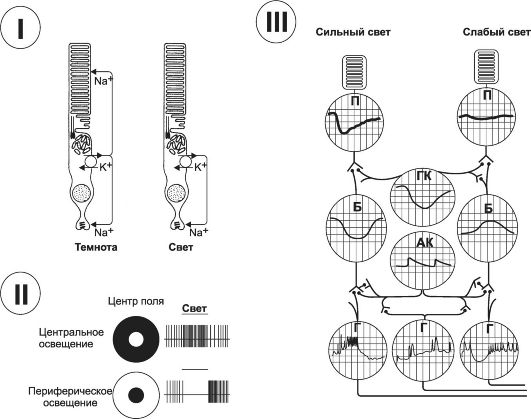
Рис. 10-6. Электрические реакции сетчатки. I - ответ фоторецептора на освещение; II - ответы ганглиозных клеток. Освещённые поля обозначены белым; III - локальные потенциалы клеток сетчатки: П - палочки, ГК - горизонтальные клетки, Б - биполярные клетки, АК - амакринные клетки, Г - ганглиозные клетки. Пояснения в тексте.
компенсируя выход Na+ входом К+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca2+ через открытые каналы обеспечивают появление тока (темновой ток).
Φ На свету, т.е. когда свет возбуждает наружный сегмент, Na+-каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора - глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клеткок. Таким образом, гиперполяризация плазмолеммы - следствие закрытия ионных каналов.
Φ Возврат к исходному состоянию. Свет, вызывающий каскад реакций, понижающих концентрацию внутриклеточного цГМФ и приводящих к закрытию натриевых каналов, уменьшает содержание в фоторецепторе не только Na+, но и Ca2+. В результате понижения концентрации Ca2+ активируется фермент гуанилатциклаза, синтезирующая цГМФ, и в клетке растёт содержание цГМФ. Это приводит к тор- можению функций активированной светом фосфодиэстеразы. Оба этих процесса - повышение содержания цГМФ и торможение активности фосфодиэстеразы - возвраща- ют фоторецептор в исходное состояние и открывают Na+- каналы.
• Световая и темновая адаптация
Φ Световая адаптация. Если человек длительное время находится в условиях яркого освещения, то в палочках и кол- бочках происходит превращение значительной части зрительных пигментов в ретиналь и опсин. Большая часть ретиналя превращается в витамин А. Всё это приводит к соответствующему снижению чувствительности глаза, называемому световой адаптацией.
Φ Темновая адаптация. Напротив, если человек остаётся в темноте продолжительное время, то витамин А вновь превращается в ретиналь, ретиналь и опсин формируют зрительные пигменты. Всё это обусловливает повышение чувствительности глаза - темновую адаптацию.
Электрические ответы сетчатки
Различные клетки сетчатки (фоторецепторы, биполярные, горизонтальные, амакринные, а также дендритная зона ганглиозных нейронов) генерируют локальные потенциалы, но не ПД (рис. 10-6). Из всех клеток сетчатки ПД возникают только в аксонах ганглиозных клеток. Суммарные электрические потенциалы сетчатки - электроретинограмма (ЭРГ). ЭРГ регистрируют так: один электрод накладывают на поверхность роговицы, другой - на кожу лица. ЭРГ имеет несколько волн, связанных с возбуждением различных структур сетчатки, и суммарно отражает интенсивность и длительность действия света. Данные ЭРГ могут использоваться в диагностических целях при заболеваниях сетчатки.
• Нейромедиаторы. Нейроны сетчатки синтезируют ацетилхолин, дофамин, Z-глутаминовую кислоту, глицин, γ-аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды. Палочки и колбочки в синапсах с биполярными клетками секретируют глутамат. Разные амакринные клетки выделяют ГАМК, глицин, дофамин, ацетилхолин и индоламин, оказывающие тормозные эффекты. Нейромедиаторы для биполярных и горизонтальных клеток не идентифицированы.
• Локальные потенциалы. Ответы палочек, колбочек и горизонтальных клеток являются гиперполяризующими (рис. 10-6, II), ответы биполярных клеток либо гиперполяризующие, либо деполяризующие. Амакринные клетки создают деполяризующие потенциалы.
Функциональные особенности клеток сетчатки
• Зрительные образы. Сетчатка вовлечена в формирование трёх зрительных образов. Первый образ формируется под действием света на уровне фоторецепторов, превращается во второй образ на уровне биполярных клеток; в ганглиозных нейронах формируется третий образ. В формировании второго образа принимают также участие горизонтальные клетки, а в образовании третьего задействованы амакринные клетки.
• Латеральное торможение - способ усиления зрительного контраста. Латеральное торможение - важнейший элемент деятельности сенсорных систем, позволяющий в сетчатке усили-
• вать явления контраста. В сетчатке латеральное торможение отмечается во всех нейронных слоях, но для горизонтальных клеток оно является их основной функцией. Горизонтальные клетки латерально синаптически связаны с синаптическими участками палочек и колбочек и с дендритами биполярных клеток. В окончаниях горизонтальных клеток выделяется медиатор, который всегда оказывает тормозное влияние. Таким образом, латеральные контакты горизонтальных клеток обеспечивают возникновение латерального торможения и передачу правильного зрительного паттерна в мозг. Рецептивные поля. В сетчатке на 100 млн палочек и 3 млн колбочек приходится около 1,6 млн ганглиозных клеток. В среднем на одну ганглиозную клетку конвергируется 60 палочек и 2 колбочки. Существуют большие различия между количеством палочек и колбочек, конвергирующих на ганглиозные нейроны, в периферических и центральных отделах сетчатки. На периферии сетчатки фоторецепторы, связанные с одной ганглиозной клеткой, образуют её рецептивное поле. Перекрытие рецептивных полей различных ганглиозных клеток позволяет повышать световую чувствительность при низком пространственном разрешении. По мере приближения к центральной ямке соотношение палочек и колбочек с ганглиозными клетками становится более упорядоченным и на каждое нервное волокно приходится всего лишь несколько палочек и колбочек. В области центральной ямки остаются только колбочки (около 35 000), и количество волокон зрительного нерва, выходящих из этой области, равно количеству колбочек. Так возникает высокая степень остроты зрения по сравнению с относительно слабой остротой зрения на периферии сетчатки. На рис. 10-6, II слева изображены диаграммы рецептивных полей, освещённых по центру и по периферии круга, справа - диаграммы частоты ПД, возникающих в аксонах ганглиозных нервных клеткок в ответ на освещение. При центральном освещении возбуждённое рецептивное поле вызывает латеральное торможение по периферии: на верхнем рисунке справа частота импульсов в центре намного больше, чем по краям. При засветке рецептивного поля по краям круга импульсация имеется по периферии и отсутствует в центре.
• Ганглиозные клетки разных типов. Ганглиозные клетки в состоянии покоя генерируют спонтанные потенциалы частотой от 5 до 40 Гц, на которые накладываются зрительные сигналы. Известно несколько типов ганглиозных нейронов.
W-клетки (диаметр перикариона менее 10 мкм, скорость проведения ПД 8 м/с) составляют 40% общего количества всех ганглиозных клеток. W-клетки имеют обширное рецептивное поле, они получают сигналы от палочек, пере- даваемые биполярными и амакринными клетками, и ответственны за сумеречное зрение.
X-клетки (диаметр 10-15 мкм, скорость проведения около 14 м/с, 55%) имеют небольшое рецептивное поле с дискретной локализацией. Они ответственны за передачу зрительного образа как такового и за все виды цветного зрения.
Y-клетки (диаметр более 35 мкм, скорость проведения более 50 м/с, 5%) - самые крупные ганглиозные клетки - имеют обширное дендритное поле и получают сигналы из различных областей сетчатки. Y-клетки реагируют на быстрые изменения зрительных образов, быстрые движения перед глазами, быстрые изменения интенсивности света. Эти клетки мгновенно сигнализируют в ЦНС, когда в какойлибо части зрительного поля неожиданно появляется новый зрительный образ.
on- и off-ответы. Многие ганглиозные нейроны возбуждаются, когда изменяется интенсивность освещения. Наблюдаются два типа ответов - on-ответ на включение света и off-ответ на выключение света. Эти разные типы ответов появляются соответственно от деполяризованных или гиперполяризованных биполяров.
Цветовое зрение
• Характеристики цвета. Цвет имеет три основных показателя: тон (оттенок), интенсивность и насыщение. Для каждого из цветов существует дополнительный (комплементарный) цвет, который, будучи должным образом перемешан с исходным цветом, даёт ощущение белого цвета. Чёрный цвет является ощущением, создаваемым отсутствием света. Восприятия белого цвета, любого цвета спектра и даже дополнительных цветов спектра можно достичь смешивая в различных
пропорциях красный (570 нм), зелёный (535 нм) и голубой (445 нм) цвета. Поэтому красный, зелёный и голубой - первичные (основные) цвета. Восприятие цвета зависит в какой-то мере от цвета других объектов в поле зрения. Например, красный объект кажется красным, если поле освещается зелёным или голубым цветом, и этот же красный объект будет казаться бледно-розовым или белым, если поле будет освещаться красным цветом.
• Цветовосприятие - функция колбочек. Существуют три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов.
• Трихромазия - возможность различать любые цвета - определяется наличием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего - первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802) и развил Герман Гельмгольц.
НЕРВНЫЕ ПУТИ И ЦЕНТРЫ
Зрительные пути
Зрительные пути подразделяются на старую систему, куда относятся средний мозг и основание переднего мозга, и новую систему (для передачи зрительных сигналов непосредственно в зрительную кору, расположенную в затылочных долях). Новая система фактически отвечает за восприятие всех зрительных образов, цвета и всех форм осознаваемого зрения.
• Основной путь к зрительной коре (новая система). Аксоны ганглиозных клеток в составе зрительных нервов и (после пе- рекрёста) в составе зрительных трактов достигают латеральных коленчатых тел (ЛКТ, рис. 10-7, А). При этом волокна от носовой половины сетчатки в зрительном перекрёсте не переходят на другую сторону. В левом ЛКТ (ипсилатеральном глазу) волокна от носовой половины сетчатки левого глаза и волокна от височной половины сетчатки правого глаза синаптически контактируют с нейронами ЛКТ, аксоны которых образуют коленчато-шпорный тракт (зрительная лучистость). Коленчато-шпорные волокна проходят к первичной зрительной коре той же стороны. Аналогично организованы пути от правого глаза.
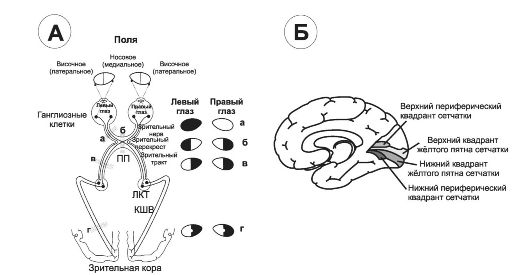
Рис. 10-7. Зрительные пути (А) и корковые центры (Б). А - области перерезки зрительных путей обозначены строчными буквами, а возникающие после перерезки дефекты зрения показаны справа. ПП - перекрёст зрительного нерва, ЛКТ - латеральное коленчатое тело, КШВ - коленчатошпорные волокна; Б - медиальная поверхность правого полушария с проекцией сетчатки в области шпорной борозды.
• Другие пути (старая система). Аксоны ганглиозных нейронов сетчатки проходят также в некоторые древние области мозга: Φ к надперекрёстным ядрам гипоталамуса (контроль и синхронизация циркадных ритмов);
Φ в ядра покрышки (рефлекторные движения глаз при фокусировании объекта, активация зрачкового рефлекса);
Φ в верхнее двухолмие (контроль быстрых направленных движений обоих глаз);
Φ в ЛКТ и окружающие их области (контроль поведенческих реакций).
• Латеральное коленчатое тело (ЛКТ) - часть новой зрительной системы, где оканчиваются все волокна, проходящие в составе зрительного тракта. ЛКТ выполняет функцию передачи информации из зрительного тракта к зрительной коре, в точности сохраняя топологию (пространственное расположение) разного уровня путей из сетчатки (рис. 10-7, Б). Другая функция ЛКТ заключается в контроле над количеством информа-
ции, поступающей к коре. Сигналы для осуществления ЛКТ входного контроля поступают в ЛКТ в виде обратной импуль- сации из первичной зрительной коры и из ретикулярной области среднего мозга.
Зрительная кора
Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды (см. рис. 10-7, Б). Подобно другим частям новой коры, зрительная кора состоит из шести слоёв, волокна коленчато-шпорного пути заканчиваются преимущественно на нейронах слоя IV. Этот слой подразделяется на подслои, принимающие волокна от ганглиозных клеток типа Y и X. В первичной зрительной коре (поле 17 по Бродману) и зри- тельной области II (поле 18) осуществляется анализ трёхмерного расположения объектов, величины объектов, детализация предметов и их окраски, движения объектов и т.д.
• Колонки и полоски. Зрительная
кора содержит несколько миллионов вертикальных первичных колонок,
каждая из которых имеет диаметр от 30 до 50 мкм и содержит около 1000
нейронов. Нейронные колонки формируют переплетённые между собой полоски
шириной в
• Цветовые колонкообразные структуры. Среди первичных зрительных колонок распределяются вторичные области - колонкоподобные образования («цветовые сгустки»). «Цветовые сгустки» получают сигналы от прилежащих колонок и специфически активируются цветовыми сигналами.
• Взаимодействие зрительных сигналов из двух глаз. Зрительные сигналы, поступающие в мозг, остаются раздельными до их вхождения в слой IV первичной зрительной коры. Сигналы из одного глаза входят в колонки каждой полоски, то же самое происходит с сигналами из другого глаза. В ходе взаимодействия зрительных сигналов зрительная кора расшифровывает расположение двух зрительных образов, находит их корреспондирующие точки (точки в одинаковых участках сетчатки обоих глаз) и использует расшифрованную информацию, чтобы определить расстояние до объектов.
• Специализация нейронов. В колонках зрительной коры имеются нейроны, выполняющие совершенно определённые функ-
ции (например, анализ контраста, в том числе цветового, границ и направлений линий зрительного образа и др.
СВОЙСТВА ЗРИТЕЛЬНОЙ СИСТЕМЫ
Движения глаз
• Наружные мышцы глазного яблока. Движения глаз осуществляются шестью парами поперечнополосатых мышц (рис. 10-8, А), координируемых головным мозгом посредством III, IV, VI пар черепных нервов. Если прямая латеральная мышца одного глаза сокращается, то прямая медиальная мышца другого глаза сокращается на такую же величину. Прямые верхние мышцы работают вместе и перемещают глаза назад, чтобы можно было смотреть вверх. Прямые нижние мышцы дают возможность смотреть вниз. Косая верхняя мышца вращает глаз вниз и наружу, а косая нижняя мышца - вверх и наружу.
Конвергенция. Одновременное и содружественное движение обоих глаз позволяет, рассматривая близкие предметы, сводить их (конвергенция).
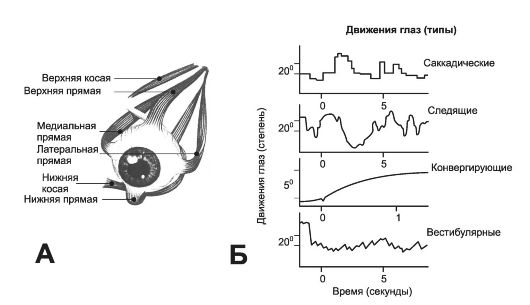
Рис. 10-8. Наружные глазные мышцы. А - глазные мышцы левого глаза; Б - типы движений глаз.
Дивергенция. Рассматривание далёких объектов приводит к разведению зрительных осей обоих глаз (дивергенция).
Диплопия. Поскольку основная часть зрительного поля би- нокулярна, ясно, что необходима высокая степень координации движений обоих глаз, чтобы удержать зрительный образ на корреспондирующих точках обеих сетчаток и тем самым избежать двоения в глазах (диплопия).
• Типы движений. Выделяют четыре типа движений глаз (рис. 10-8, Б).
Саккады - неощущаемые быстрые скачки (в сотые доли секунды) глаза, прослеживающие контуры изображения. Саккадические движения позволяют удерживать изображение на сетчатке, что достигается периодическим смещением изображения по сетчатке, в результате чего активизируются новые фоторецепторы и новые ганглиозные клетки.
Плавные следящие движения глаза за движущимся объектом.
Конвергирующие движения - сведение зрительных осей навстречу друг другу при рассматривании объекта вблизи от наблюдателя. Каждый тип движений контролируется нервным аппаратом раздельно, но в конечном счёте все влияния заканчиваются на мотонейронах, иннервирующих наружные мышцы глаза.
Вестибулярные движения глаза - регулирующий механизм, появляющийся при возбуждении рецепторов полукружных каналов и поддерживающий фиксацию взора во время движений головы.
• Физиологический нистагм. Даже в условиях, когда субъект пытается фиксировать взглядом неподвижный предмет, глазное яблоко продолжает осуществлять скачкообразные и другие движения (физиологический нистагм). Другими словами, нервно-мышечный аппарат глаза берёт на себя функцию удержания зрительного образа на сетчатке, так как попытка удержать зрительный образ неподвижно на сетчатке приводит к его исчезновению из поля зрения. Именно поэтому необходимость постоянного удержания объекта в поле зрения требует постоянного и быстрого смещения зрительного образа по сетчатке.
Критическая частота мельканий. Глаз сохраняет следы световой стимуляции в течение некоторого времени (150-250 мс) после выключения света. Иными словами, глаз воспринимает прерывистый свет как непрерывный при определённых интервалах между вспышками. Минимальная частота следования световых стимулов, при которой отдельные ощущения мелькания сливаются в ощущение непрерывного света, - критическая частота слияния мельканий (24 кадра в секунду). На этом явлении основано телевидение и кино: человек не замечает промежутков между отдельными кадрами, так как зрительное ощущение одного кадра длится до появления другого. Тем самым создаётся иллюзия непрерывности изображения и его движения.
Водянистая влага
Водянистая влага непрерывно образуется и реабсорбируется. Баланс между образованием и реабсорбцией водянистой влаги ре- гулирует объём и давление внутриглазной жидкости.
Каждую
минуту образуется 2-3 мкл водянистой влаги. Эта жидкость вытекает между
связками хрусталика и далее через зрачок в переднюю камеру глаза.
Отсюда жидкость поступает в угол между роговицей и радужкой, проникает
между сетью трабекул в шлеммов канал и изливается в наружные вены
глазного яблока. Нормальное внутриглазное давление в среднем равняется
Обобщение главы
Зрительная функция глаза определяется структурами, которые формируют и приспосабливают изображения, и структурами, трансформирующими изображения в нервные сигналы.
Сетчатка содержит определённое число слоёв и несколько типов клеток, выполняющих специфическую роль в процессе зрительного восприятия.
Палочковые клетки в сетчатке наделены высокой чувствительностью к свету, но воспроизводят неясные бесцветные изображе-
ния, в то время как колбочки обеспечивают яркое цветное зрение с меньшей чувствительностью к свету.
Процесс зрительного восприятия состоит из многих этапов, начинаясь с поглощения света и заканчиваясь электрическими нервными импульсами, передающими образы внешнего мира.