Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 15. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Поддержание постоянства и оптимальных показателей внутренней среды (гомеостаз) - задача вегетативного отдела нервной системы. Два подотдела этой системы (симпатический и парасимпатический) контролируют АД, моторику и секреторную функцию ЖКТ, температуру тела, выделение слюны, опорожнение мочевого пузыря и множество других висцеральных (вегетативных, внутренностных, «растительных») функций организма. Работа этого системного регулятора висцеральных функций осуществляется рефлекторно (регуляторные контуры висцеральных рефлексов работают по принципу обратной связи) и независимо (автономно) от сознания, но не от деятельности мозга и других отделов нервной системы. И симпатический, и парасимпатический подотделы представлены в ЦНС и имеют свои эфферентные пути, заканчивающиеся на эффекторах - секреторных и гладкомышечных клетках (ГМК), в миокарде.
К вегетативной нервной системе относится также кишечная (энтеральная) нервная система, расположенная в стенке ЖКТ, функция которой описывается в соответствующем разделе.
Вегетативная нервная система (подобно соматической нервной системе) функционирует посредством рефлексов. В отличие от соматических, рефлексы вегетативной нервной системы являются висцеральными. Подсознательные сенсорные сигналы возникают в висцеральных рецепторах внутренних органов и через афферентные пути поступают в ЦНС. Они интегрируются на различных уровнях вегетативной нервной системы: в ганглиях, мозговом стволе, гипоталамусе, лимбической коре - и возвращаются обратно в виде неосознаваемых рефлекторных ответов.
Висцеральные рецепторы и их афферентные пути рассмотрены в главах 8 и 9, а мышечный эффектор (ГМК) - в главе 7. Предме-
том данной главы является функция эфферентных путей симпатического и парасимпатического подотделов вегетативной нервной системы.
Эфферентные пути вегетативной нервной системы представлены на рис. 15-1.
Симпатический отдел
Каждый симпатический эфферентный путь состоит из двух нейронов: преганглионарного и постганглионарного - и их аксонов в составе нервных пре- и постганглионарных нервных волокон (рис. 15-2). Преганглионарные волокна - миелинизированные, сравнительно медленнопроводящие нервные волокна типа B. Постганглионарные волокна - безмиелиновые нервные волокна типа C.
• Преганглионарные нейроны. Тела преганглионарных нейронов расположены в боковых рогах спинного мозга. Их аксоны проходят через передние рога спинного мозга в составе спинномозговых нервов. Большинство симпатических волокон оставляет нерв и через белые соединительные ветви проходит в один из ганглиев симпатической цепочки. Далее преганглионарные волокна заканчиваются синапсами:
с постганглионарными нейронами в том же ганглии, куда
они вошли из спинного мозга; с нейронами ганглиев симпатической цепочки, расположенных выше и ниже данного узла; с нейронами, лежащими вне симпатической цепочки в периферических симпатических ганглиях. Некоторые постганглионарные нервные волокна проходят из симпатической цепочки в спинномозговой нерв через серые соединительные ветви и направляются к кровеносным сосудам, потовым железам и мышцам, выпрямляющим волосы.
• Постганглионарные нейроны. Их тела расположены либо в ганглиях симпатической цепочки, либо в периферических симпатических ганглиях (в мозговой части надпочечника постганглионарные нейроны - модифицированные нервные, или хромаффинные клетки). Из этих источников их аксоны - постганглионарные симпатические волокна - направляются к различным органам.
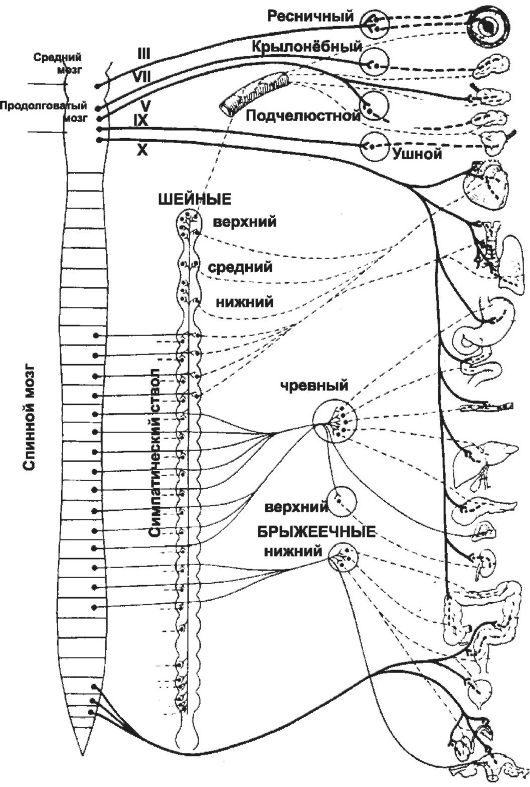
Рис. 15-1. Эфферентные пути вегетативной нервной системы. Слева направо: стволовая часть мозга и спинной мозг, симпатический ствол, ганглии вне ствола, эффекторные структуры. Преганглионарные нейроны - сплошная линия, постганглионарные нейроны - пунктирная линия; толстая линия - парасимпатические пути, тонкая линия - симпатические пути.
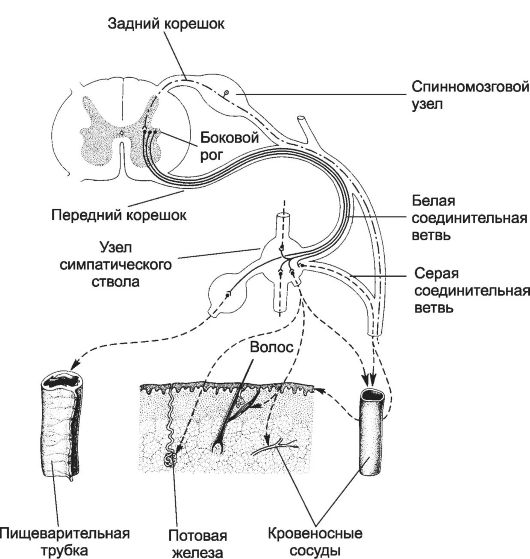
Рис. 15-2. Симпатический отдел вегетативной нервной системы [11]. Пе-
рикарионы первого нейрона расположены в боковых столбах спинного мозга. Их аксоны проходят в составе передних корешков и белой соединительной ветви и заканчиваются на перикарионах второго нейрона в паравертебральных ганглиях симпатической цепочки, превертебральных ганглиях и ганглиях (терминальных), расположенных вблизи иннервируемых ими органов. Аксоны второго нейрона паравертебральных, превертебральных и терминальных ганглиев заканчиваются во внутренних органах, в коже, стенке кровеносных сосудов. Аксоны некоторых нейронов паравертебральных ганглиев проходят через серую соединительную ветвь.
Парасимпатический отдел
• Преганглионарные нейроны. Их тела расположены в среднем и продолговатом мозге, а также в крестцовом отделе спинного мозга (см. рис. 15-1). Аксоны преганглионарных нейронов (преганглионарные волокна) в составе блуждающих и тазовых нервов без перерыва достигают иннервируемых органов, где образуют синапсы с постганглионарными нервными клетками парасимпатического эфферентного пути. Преганглионарные волокна, идущие в составе III, V, VII и IX пар черепных нервов, также образуют синапсы с постганглионарными нервными клетками, но постганглионарные нейроны располагаются не в иннервируемых органах, а вне их - в соответствующих парасимпатических ганглиях.
• Постганглионарные нейроны расположены либо в стенке иннервируемых органов (сердце, лёгкие, пищевод, желудок, кишечник, печень, желчный пузырь, поджелудочная железа, мочеточники, мочевой пузырь, половые органы), либо в нервных узлах (ресничный, крылонёбный, подчелюстной, ушной). Независимо от местоположения тел постганглионарных нейронов, их аксоны иннервируют соответствующие мышечные и секреторные клетки.
Нейромедиаторы
Нейроны вегетативной нервной системы синтезируют и секретируют различные биологически активные вещества, выступающие в качестве нейромедиаторов и нейромодуляторов. Основные нейро- медиаторы - ацетилхолин и норадреналин. Нейроны, синтезирующие ацетилхолин и секретирующие ацетилхолин в терминальных ветвлениях аксона, - холинергические. Нейроны, синтезирующие норадреналин и секретирующие норадреналин в терминальных ветвлениях аксона, - адренергические.
• Холинергические нейроны
Φ Преганглионарные нейроны. Все преганглионарные нейроны холинергические (как в симпатическом, так и в парасимпатическом отделе). Следовательно, ацетилхолин обеспечивает синаптическую передачу между пре- и постганглионарными нейронами как в симпатическом, так и в парасимпатическом отделе вегетативной нервной системы.
Постганглионарные нейроны. Все (или почти все) постганглионарные парасимпатические нейроны холинергические.
• Адренергические нейроны
Преганглионарные нейроны. Преганглионарные адренерги-
ческие нейроны не найдены. Постганглионарные нейроны
♦ Большинство постганглионарных симпатических нейронов адренергические.
♦ Постганглионарные симпатические нервные волокна к потовым железам, к выпрямляющим мышцам волос (пиломоторы) и некоторым кровеносным сосудам - холинергические.
• Варикозные расширения (см. рис. 7-11). Аксоны постганглионарных вегетативных нейронов образуют многочисленные варикозные расширения - локальные утолщения, содержащие синаптические пузырьки. Эти утолщения - места секреции нейромедиатора: норадреналина из адренергических волокон, ацетилхолина - из холинергических волокон.
• Адрено- и холинорецепторы. Норадреналин и ацетилхолин, выделившиеся из пресинаптических окончаний, взаимодействуют с их рецепторами - адренорецепторами и холинорецепторами соответственно. На рис. 15-3 с учётом нейромедиатора и типов рецепторов представлены эффекторные симпатические и парасимпатические цепочки.
Рецепторы и ответы эффекторных структур
Ответ содержащих рецепторы эффекторных структур (нейроны, ГМК, секреторные клетки, клетки проводящей системы сердца и кардиомиоцитов) определяется типом рецептора. В эффекторных путях вегетативной нервной системы существуют два типа холинорецепторов (никотиновые и мускариновые) и несколько типов адренорецепторов: α1 (подтипы α1A, α1B и α1D), α2 (подтипы α2A, α2B,
α2D),β1,β2 и β3
Адренорецепторы
• α1-Адренорецепторы
Φ Локализация и функция: ЦНС - увеличение симпатической активности; слюнные железы - повышение слюноотделе- ния; печень - повышение гликогенолиза; почки - измене-
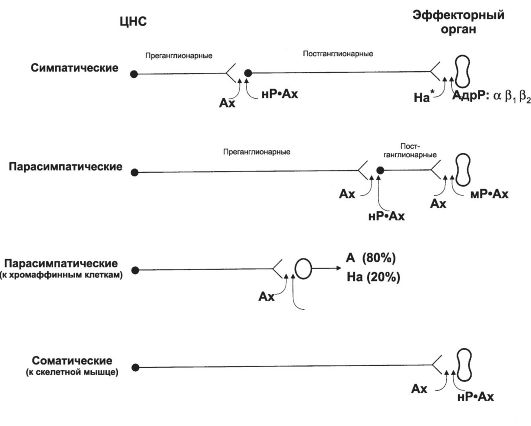
Рис. 15-3. Симпатические и парасимпатические эфферентные пути [5].
Ах - ацетилхолин, На - норадреналин, А - адреналин, АдрР - адренорецептор, нР - никотиновый холинорецептор, мР - мускариновый холинорецептор.
ние порога для высвобождения ренина; гладкие мышцы - сокращение артериол, матки, выносящего протока, бронхиол, мочевого пузыря, желудочно-кишечных сфинктеров и дилататора зрачка. Φ Чувствительность к лигандам: одинаковая к адреналину и норадреналину, в реальных условиях in vivo возбуждает норадреналин.
• Φ Активация агадренорецепторов опосредуется Gq-белками и фосфолипазой C, что обусловливает образование инозитолтрифосфата и увеличение внутриклеточного [Ca2+]. α2-Адренорецепторы
Φ Локализация и функция: ЦНС - уменьшение активности симпатической системы; слюнные железы - снижение са-
ливации; поджелудочная железа - уменьшение секреции инсулина; жировые клетки - уменьшение липолиза; тромбоциты - усиление агрегации; пресинаптические нервные терминали - возникновение гиперполяризации за счет увеличения открытых потенциалзависимых K+- и ингибирования потенциалзависимых Са2+-каналов. Φ Активированные α-рецепторы связываются с Gi-белком, ингибируют активность аденилатциклазы и уменьшают содержание внутриклеточного [цАМФ]. Одновременно с этим увеличивается количество открытых потенциалзависимых К+-каналов (гиперполяризация). Все β-адренорецепторы сцеплены с G..-белком, субъединицы которого высвобождают цАМФ как второй посредник. цАМФ затем активирует протеинкиназу A, которая фосфорилирует различные белки в зависимости от типа клеток-мишеней.
• β1-Адренорецепторы
Φ Локализация и функция: адреналин и норадреналин действуют, открывая Са2+-каналы в мембранах клеток сердца. Они увеличивают содержание внутриклеточного кальция и вызывают положительный хронотропный, дромотропный и инотропный эффекты. В почках увеличивается базальная секреция ренина.
Φ Чувствительность к лигандам: одинаковая к адреналину и норадреналину; чувствительность выше, чем у α-адренорецепторов.
• β2-Адренорецепторы
Φ Локализация и функция: активация β2-адренорецепторов адреналином и норадреналином приводит к снижению внутриклеточного кальция. Это расширяет бронхиолы и кровеносные сосуды в скелетных мышцах и расслабляет гладкие мышцы матки, семявыносящего протока и ЖКТ. Кроме того, увеличивается секреция инсулина, возрастает гликогенолиз в печени и мышцах и уменьшается агрегация тромбоцитов.
Φ Чувствительность к лигандам у адреналина больше, чем у норадреналина; чувствительность к адреналину выше, чем у α-адренорецепторов.
Холинорецепторы
Ацетилхолин присоединяется к постсинаптическим холинергическим рецепторам или холинорецепторам в вегетативных ганглиях
и органах, иннервируемых парасимпатическими волокнами. Холинорецепторы являются никотиновыми (Н) или мускариновыми (М). Н-холинорецепторы могут стимулироваться алкалоидом никотином, в то время как М-холинорецепторы стимулируются ядовитым грибным алкалоидом мускарином.
• Никотиновые холинорецепторы
Φ Локализация и функция: Н-холинорецепторы в ганглиях вегетативной нервной системы отличаются от Н-холинорецепторов нервно-мышечных синапсов скелетной мышцы тем, что они формируются различными субъединицами. Но оба вида Н-холинорецепторов сходны в том, что являются ионотропнымирецепторами, т.е. они действуют как холинорецепторы и катионные каналы в одно и то же время. Ах, взаимодействуя с Н-холинорецептором, приводит к быстрому входу Na+ и Са2+ в клетку и актвирует её.
Φ Чувствительность к лигандам: ганглиоблокаторы (см. табл. 15-3) не блокируют нервно-мышечную передачу.
• Мускариновые холинорецепторы М-холинорецепторы (М1-М3) косвенно влияют на синаптическую передачу через G-белки, являясь метаботропными рецепторами.
Φ Локализация и функция: М1-холинорецепторы встречаются в вегетативных ганглиях, ЦНС и экзокринных железах. Они активируют фосфолипазу через Gq-белок и стимулируют вход Са2+, активируя клетки. М2-холинорецепторы встречаются в сердце и функционируют через Gi-белок, который открывает специфические К+-каналы, расположенные, главным образом, в синоатриальном и атриовен- трикулярном узлах и клетках предсердия. Это вызывает отрицательные хронотропный и дромотропный эффекты на сердце. Gi-белок также ингибирует аденилатциклазу, уменьшая вход Са2+. М3-холинорецепторы встречаются главным образом в ГМК. Подобно М1-холинорецепторам, М3-холинорецепторы вызывают сокращение, стимулируя вход Са2+. Однако они могут также вызывать расслабление ГМК, активируя кальцийзависимый фермент NO-синтазу в клетках эндотелия.
Φ Чувствительность к лигандам: атропин блокирует все М-холинорецепторы.
Физиологические ответы эффекторов
Поступление сигналов по эфферентным путям вегетативного отдела нервной системы вызывает специфические физиологические ответы иннервируемых структур. Характер ответа полностью определяется типом рецепторов, встроенных в клетки-мишени.
Таблица 15-1. Ответы эффекторов на парасимпатическую и симпатиче- скую стимуляцию
Эффектор | Парасимпатический | Симпатический |
Глаз | ||
Зрачок | Сужение | Расширение (α,) |
Цилиарная мышца | Сокращение | Расслабление (β2) |
Слёзная железа | Секреция | Нет |
Носовые железы | Секреция | Торможение (α,) |
Слюнные железы | Секреция | Секреция амилазы (β) |
Кожа | ||
Потовые железы | Нет | Секреция (М-холинергическая) |
Мышцы, поднимающие волосы | Нет | Сокращение (α1) |
Кровеносные сосуды | ||
Кожа (в общем) | Нет | Сужение (α) |
Мышцы лица и шеи | Расслабление | Сужение (α), расслабление (?) |
Скелетные мышцы | Нет | Расслабление (β2) |
Внутренние органы | Нет | Сужение (α,) |
Сердце | ||
Частота | Уменьшается | Увеличивается (β,, β2) |
Сила | Уменьшается | Увеличивается (β,, β2) |
Лёгкие | ||
Бронхиолы | Сужение | Расширение (β2) |
Железы | Секреция | Уменьшение (α,), увеличение (β2) |
ЖКТ | ||
Мышцы стенки | Сокращение | Расслабление (α, β2) |
Сфинктеры | Расслабление | Сокращение (α,) |
Железы | Секреция | Торможение |
Печень | Гликогенолиз и глюконеогенез (α,, β2) |
Окончание табл. 15-1
Эффектор | Парасимпатический | Симпатический |
Поджелудочная железа (инсулин) | Нет | Уменьшение секреции (α2) |
Мозговой слой надпочечников | Нет | Секреция адреналина (Н-холинергическое) |
Мочевыделительная система | ||
Мочеточник | Расслабление | Сокращение (α,) |
Детрузор | Сокращение | Расслабление (β2) |
Сфинктер | Расслабление | Сокращение (α,) |
Репродуктивная система | ||
Матка | Вариабельное | Сокращение (α,) |
Гениталии | Эрекция (Ах и оксид азота) | Эякуляция/сокращения влагалища (α) |
Жировые клетки | Нет | Липолиз (β) |
Агонисты и антагонисты
В клинической практике широко применяют различные ЛС - агонисты и антагонисты адрено- и холинорецепторов разных типов. Агонисты, взаимодействуя с рецептором, воспроизводят эффект естественного рецепторного лиганда (например, норадреналина для адренорецепторов). Антагонисты, взаимодействуя с рецептором, блокируют эффект естественного рецепторного лиганда. В табл. 15-2 перечислены некоторые лиганды адрено- и холинорецепторов, обладающих свойствами агонистов и антагонистов.
Эффекты гуморальных адреналина и норадреналина
В эффекторных путях симпатического отдела вегетативной нервной системы нейромедиатором является норадреналин. В то же время и норадреналин, и главным образом адреналин поступают в кровь из хромаффинных клеток надпочечниково-адреналовой системы. Другими словами, эти химические соединения могут рассматриваться как гормоны (см. подробнее в гл. ,8). Адреналин и норадреналин, выделяющиеся в кровь из хромаффинных клеток (в том числе из мозговой части надпочечников), вызывают практически те же эффекты, что и стимуляция симпатических эффекторов, но действие гуморальных адреналина и норадреналина продолжа-
Таблица 15-2. Агонисты и антагонисты адрено- и холинорецепторов [5]
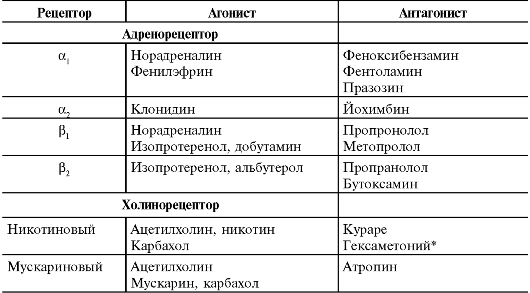
* В ганглиях, но не в нервно-скелегао-мьпнечном синапсе.
ется в 5-10 раз дольше, чем эффект норадреналина в качестве нейромедиатора, поскольку гормоны удаляются из крови в течение долгих 1-3 мин. Адреналин вызывает почти те же самые эффекты, что и норадреналин, но имеются и некоторые отличия.
• Адреналин более интенсивно стимулирует работу сердца, потому что он лучше связывается с β-адренорецепторами.
• Адреналин (в отличие от норадреналина) вызывает некоторое сужение кровеносных сосудов в скелетных мышцах. Это важный момент, потому что сосуды мышц - весьма существенный сегмент сосудов тела, и норадреналин, вызывая вазоконстрикцию, увеличивает периферическое сосудистое сопротивление, в результате чего повышается АД. Адреналин повышает АД в меньшей степени, но зато в большей степени увеличивает производительность сердца.
• Адреналин вызывает метаболический эффект, в 5-Ю раз превышающий эффект норадреналина. Выделяясь из надпочечников, адреналин ускоряет метаболизм и тем самым усиливает активность и возбудимость организма.
• Выброс адреналина и норадреналина из надпочечников совпадает с генерализованной активацией симпатической нервной
системы. Следовательно, одновременно происходит стимуляция эффекторных органов двумя путями: непосредственно симпатическими нервами и опосредованно - гормонами мозговой части надпочечников. Такая двойная адренергическая стимуляция поддерживает и во многих случаях заменяет друг друга.
Понятие о тонусе
Симпатическая и парасимпатическая системы постоянно активны, и величину этой исходной активности рассматривают как симпатический тонус или как парасимпатический тонус соответственно.
• Тонус оценивают по способности одной из систем увеличивать или уменьшать активность стимулируемого органа. Так, симпатический тонус поддерживает артериолы суженными наполовину по сравнению с максимальным диаметром. В ответ на увеличение симпатической стимуляции выше нормы артериолы уменьшают диаметр просвета ещё больше (вазоконстрикция). Напротив, уменьшение симпатической стимуляции ниже нормы вызовет расслабление сосудов - вазодилатацию. Другими словами, если бы постоянный базовый тонус отсутствовал, симпатическая система могла бы вызывать только вазоконстрикцию.
• Другой пример - парасимпатический тонус ЖКТ. Перерезка блуждающего нерва может вызвать длительную атонию желудка и кишечника с нарушением перистальтики и появлением в последующем выраженного запора.
• Поддержание эффекторов в тоническом состоянии обеспечивается редкими (порядка одного импульса на протяжении нескольких секунд) разрядами симпатических или парасимпатических нервов. Для полной активации необходима частота разрядов около 10-20 импульсов в секунду.
• Базальная (спонтанная) секреция адреналина и норадреналина в состоянии покоя составляет около 0,2 мкг/кг/мин адреналина и около 0,05 мкг/кг/мин норадреналина. Такого количества достаточно, чтобы удерживать АД в границах нормы.
Центральная регуляция висцеральных функций
Регуляция висцеральных функций осуществляется в ЦНС по иерархическому принципу (рис. ,5-4).
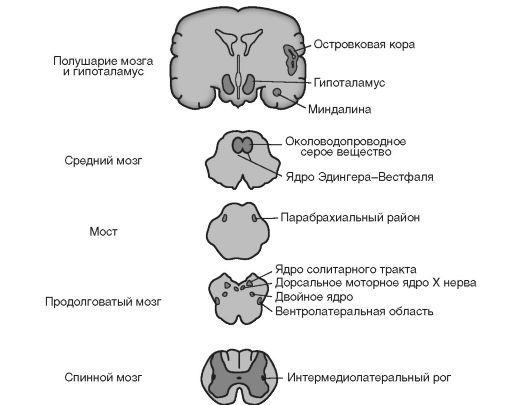
Рис. 15-4. Центры управления вегетативными функциями. Показаны церебральный, гипоталамический, стволовый и спинномозговой отделы.
Рефлексы, координируемые в поясничном и крестцовом отделе спинного мозга, включают мочеиспускание (опорожнение мочевого пузыря), дефекацию (опорожнение прямой кишки) и сексуальные ответы (наполнение кровью эректильной ткани, смазывание влагалища и извержение семенной жидкости). Сенсорные потенциалы от рецепторов мочевого пузыря или толстой кишки сигнализируют о степени растяжения. Симпатические, парасимпа- тические и соматические эфферентные действия требуются, чтобы обеспечить координацию этих ответов. Высшие центры облегчают или тормозят рефлекторные центры спинного мозга. Способность подавлять желание мочиться при переполненном мочевом пузыре является примером того, как высшие центры ЦНС тормозят реф-
лексы спинного мозга. При повреждении шейного или грудного отдела спинного мозга мочеипускание может наблюдаться непро- извольно или провоцироваться малыми объёмами мочи в мочевом пузыре. Участки всех трёх уровней мозгового ствола важны для вегетативных функций (рис. 15-4). Околоводопроводное серое вещество среднего мозга координирует вегетативные реакции на болевые стимулы и может модулировать активность сенсорных трактов, передающих боль. Парабрахиальное ядро моста участвует в дыхательном и сердечно-сосудистом контроле. Продолговатый мозг содержит несколько ключевых для вегетативной нервной системы областей. Ядра солитарного тракта получают афферентные входы от сердечных, дыхательных и желудочно-кишечных рецепторов. Вентролатеральная область продолговатого мозга - это основной центр, предназначенный для контроля преганглионарных симпатических нейронов в спинном мозге.
• Контроль дыхания, частоты сердечных сокращений, артериального давления. Центры продолговатого мозга, осуществляющие рефлекторный контроль кровообращения, дыхания и сердечной деятельности, получили название жизненно важных центров, потому что их повреждение фатально. Афферентные волокна к этим центрам поступают из самых различных участков тела от специализированных рецепторов внутренних органов. Специализированные рецепторы включают не только рецепторы каротидного синуса и аортальных телец, но и рецепторные клетки, которые располагаются в самом продолговатом мозге. Подробно эти рефлексы рассматриваются в разделах, посвя- щённых регуляции дыхания и кровообращения.
• Другие рефлексы продолговатого мозга. Глотание, кашель, чиханье, тошнота и рвота являются рефлекторными ответами, интегрированными в продолговатом мозге. Глотательный рефлекс начинается произвольно с проталкивания содержимого ротовой полости к задней стенке глотки. Кашель начинается с раздражения слизистой оболочки трахеи и вне- лёгочных бронхов. Голосовая щель закрывается, и сильное сокращение дыхательных мышц повышает внутрилёгочное давление; голосовая щель неожиданно открывается, вызывая взрывоподобный выброс воздуха. Подобен этому механизм чиханья, котрое возникает в ответ на раздражение рецепторов эпителия, выстилающего носовую полость.
• Рвота является примером висцерального рефлекса, осуществляемого продолговатым мозгом, с включением координированных соматических и висцеральных компонентов. Рвота начинается со слюноотделения и ощущения тошноты. Обратная перистальтика верхнего отдела кишечника передвигает его содержимое в желудок. Голосовая щель закрывается, предотвращая аспирацию рвотных масс в трахею. Дыхание задерживается на середине вдоха. Мышцы брюшной стенки сокращаются, и в силу того что грудная клетка удерживается в фиксированном положении, это сокращение повышает внутрибрюшное давление. Нижний сфинктер пищевода и пищевод расслабляются, и содержимое желудка удаляется наружу. Центр рвоты, находящийся в ретикулярной формации продолговатого мозга (рис. ,5-5), состоит из нескольких групп нейронов, контролирующих различные компоненты акта рвоты.
Пусковые сигналы рвоты. Раздражение слизистой оболочки верхних отделов ЖКТ вызывает рвоту. Импульсы поступают от слизистой оболочки в продолговатый мозг по висцеральным афферентным путям в составе симпатических нервов и блуждающего нерва. Другие пусковые сигналы достигают центра рвоты из среднего мозга и лимбической системы, поскольку рвота возникает, как известно, и в ответ на эмо- циональные разряды.
Хеморецепторные клетки, расположенные в хеморецепторной триггерной зоне продолговатого мозга (в боковой стенке четвёртого желудочка в области ромбовидной ямки), обладают высокой проницаемостью по отношению к различным веществам, циркулирующим в крови (см. рис. 15-5). Некоторые химические вещества, поступившие в кровь, раздражают эти хеморецептивные клетки и вызывают рвоту. Повреждение этой зоны существенно не сказывается на рвотных ответах на раздражение слизистой оболочки ЖКТ и отрицательных эмоциях, но прерывает рвоту, возникающую при инъекции апоморфина или других рвотных средств.
• Гипоталамус и полушария мозга обеспечивают высшие уровни контроля вегетативных функций. Перивентрикулярная, медиальная и латеральная области гипоталамуса контроли-
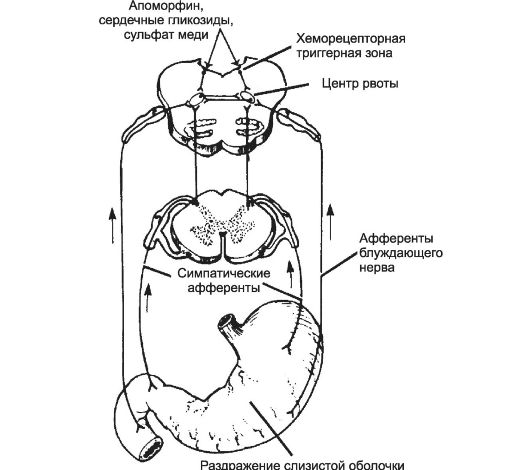
Рис. 15-5. Рефлекторный и хеморецепторный механизмы рвотного рефлекса.
руют циркадные ритмы и гомеостатические функции: терморегуляцию, аппетит и жажду. Островковая и медиально- префронтальная области коры мозга вовлечены в регуляцию моторных и сенсорных вегетативных функций. Миндалина в височной доле координирует вегетативные компоненты эмоциональных ответов. Области полушарий мозга, промежуточного мозга, мозгового ствола и центральные пути к спинному мозгу, вовлечённые в контроль вегетативных функций, составляют центральный комплекс управления вегетативными функциями.
Обобщение главы
Вегетативная нервная система регулирует непроизвольные функции организма.
Вегетативная нервная система имеет три подраздела: симпатический, парасимпатический и кишечный.
Симпатический и парасимпатический подразделы различаются анатомической природой и функциями.
В вегетативной нервной системе используется двухнейронный эфферентный путь.
Характер ответных реакций эффекторных органов на симпатическую или парасимпатическую активацию определяется типом рецепторов.
Центральная нервная система контролирует вегетативные функции посредством иерархии рефлексов и интегративных центров.