Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 14. КОНТРОЛЬ ПОЗЫ И ДВИЖЕНИЙ
Мотонейроны спинного мозга и нейроны двигательных ядер черепных нервов - общий конечный путь к скелетным мышцам. Именно на этих мотонейронах конвергируются сигналы из множества источников. Нервные пути, конвергирующие к мотонейронам, выполняют три взаимосвязанные функции:
• сигнализируют о произвольной активности;
• подготавливают позу тела к устойчивому состоянию, позволяющему двигаться;
• координируют действия различных мышц, содействуя плавным и точным движениям
Программа произвольных движений формируется в структурах мозга, а команды поступают к мышцам по пирамидному пути (корково-спинномозговые, корково-ядерные, корковоретикулярные и другие нервные волокна). На рис. 14-1 показано взаимодействие основных структур ЦНС в ходе произвольных движений.
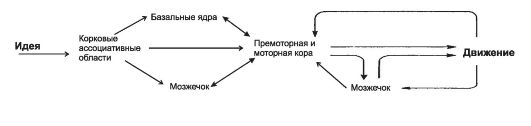
Рис. 14-1. Общая схема контроля над произвольными движениями.
КОРТИКО-СПИНАЛЬНАЯ
И КОРТИКО-БУЛЬБАРНАЯ СИСТЕМЫ
Моторная кора
В моторной коре различают первичную моторную кору, а также премоторную и дополнительную моторную области (рис. 14-2, А).
• Первичная моторная кора. На рис. 14-2, Б дана грубая топография представительства различных мышц, начиная от мышц лица (вблизи от сильвиевой борозды) и кончая мышцами ноги (область в глубине продольной щели). В первичной моторной коре картированы участки, стимуляция которых вызывает сокращения отдельных мышц, но чаще возбуждаются мышечные группы.
• Премоторная область располагается кпереди от первичной моторной коры, её топографическая организация подобна организации первичной коры: наиболее латерально располагаются зоны лица и рта, далее вверх к продольной щели - кисть, рука, туловище и нижние конечности. В премоторной области генерируются сложные спектры движений (например, движения плеча, руки, особенно кисти).
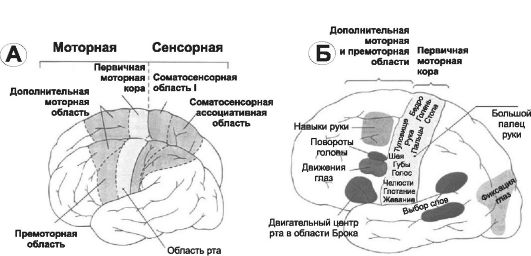
Рис. 14-2. Двигательная кора большого мозга. А - моторная и соматосен- сорная функциональные области; Б - в первичной моторной коре представлены сверху вниз (на рисунке) области тела - от стопы до головы; представительство различных мышц в моторной коре и локализация корковых областей, отвечающих за специальные движения.
• Дополнительная моторная область располагается в продольной щели и функционирует вместе с премоторной областью, обеспечивая движения, поддерживающие осанку, фиксацию движений различных сегментов тела, позиционные движения головы и глаз и базу для тонкого моторного контроля над кистями рук, осуществляемого премоторной областью и первичной моторной корой.
• Специализированные области. На рис. 14-2, Б показано несколько относящихся к двигательной функции специализированных центров (например, центры формирования речи, произвольного движения глаз, вращения головы, целевого движения кисти) коры больших полушарий, впервые обнаруженных нейрохирургами Пенфилдом и Джаспером.
• Сенсорные входы в моторную кору. Функции моторной коры контролируются множеством сигналов из соматосенсорной коры, других сенсорных систем (в частности, зрения и слуха) и иных источников. Основные нервные пути, входящие в моторную кору:
афферентные волокна из соматосенсорной области теменной коры, передних отделов фронтальной коры, слуховой и зрительной коры;
афферентные волокна, проходящие через мозолистое тело из противоположного полушария мозга;
соматосенсорные волокна, идущие непосредственно из вентробазальных таламических ядер (тактильные сигналы и сигналы из мышц и суставов);
нервные волокна вентролатерального и вентромедиального ядер таламуса, передающие информацию из мозжечка и базальных ядер;
аксоны внутрипластинчатых ядер таламуса, контролирующие уровень возбудимости нейронов моторной коры.
• Организация моторной коры. Моторная кора организована в вертикальные колонки диаметром около
Пирамидный путь
Значительная часть аксонов, выходящих из моторной коры, образует пирамидный путь (рис. 14-3, А), проходящий по краю внутренней капсулы и образующий пирамиды продолговатого мозга. В каждом пирамидном пути проходит более миллиона нервных волокон, направляющихся к спинному мозгу (корковоспинномозговые волокна) и к глубоким структурам головного мозга (корково-ядерные волокна).
• Латеральный корково-спинномозговой путь. В перекресте пирамид 80% нервных волокон переходит на другую сторону и об-
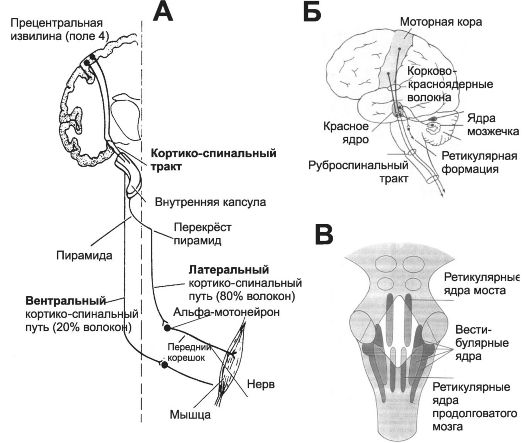
Рис. 14-3. Двигательные пути мозга. А - кортико-спинальный (пирамидный) тракт; Б - корково-красноядерные волокна (кортико-руброспинальный путь) моторного контроля; В - расположение ретикулярных и вестибулярных ядер в стволе мозга.
разует латеральный кортико-спинальный тракт. Волокна этого пути оканчиваются на вставочных нейронах, находящихся в промежуточных областях серого вещества спинного мозга, и на сенсорных релейных (переключательных) нейронах заднего рога. Лишь очень немногие аксоны непосредственно контактируют с α-мотонейронами спинного мозга.
• Передний корково-спинномозговой путь содержит около 20% аксонов пирамидного пути. Эти аксоны в пирамидах не переходят на противоположную сторону и спускаются ипсилатерально, но в шейном или в верхнем грудном отделах спинного мозга большинство волокон этого тракта переходит на другую сторону. Эти волокна участвуют в контроле дополнительной моторной области над регулирующими позу движениями.
• Связи моторной коры с глубокими структурами головного мозга. От моторной коры отходит большое количество тонких нервных волокон к глубоким структурам мозга и мозгового ствола (корково-ядерные волокна). От этих ядер сигналы по множеству путей поступают к другим ядрам стволовой части мозга, мозжечку и к спинному мозгу, в том числе:
к хвостатому ядру и скорлупе, а от них в мозговой ствол и спинной мозг;
к красному ядру и далее к спинному мозгу (красноядерноспинномозговой путь);
к ретикулярной формации и вестибулярным ядрам мозгового ствола и далее к спинному мозгу (через бульборетикулоспинномозговые и преддверно-спинномозговые пути) и мозжечку (в составе ретикуломозжечкового и вестибуломозжечкового трактов);
к ядрам моста, откуда мостомозжечковые пути и мосторетикулоспинномозговые пути проводят сигналы к полушариям мозжечка и спинному мозгу;
к ядрам нижней оливы и оттуда по оливомозжечковым волокнам к центральным областям мозжечка;
аксоны гигантских клеток Беца посылают короткие возвратные коллатерали к участкам коры, прилежащим к гигантским пирамидным клеткам. Эти коллатерали оказывают тормозящее влияние на соседние с клетками Беца нейроны в момент разряда гигантских клеток с целью усиления и выделения их сигналов.
Таким образом, для координации мышечной активности базальные ядра, мозговой ствол и мозжечок постоянно получают сигналы из моторной коры и передают их в спинной мозг.
Красное ядро
Красное ядро среднего мозга функционирует во взаимодействии с кортико-спинальным трактом, имея тесные связи с моторной ко- рой (корково-красноядерные волокна), мозжечком, ретикулярной формацией, оливами (рис. 14-3, Б). От гигантских клеток красного ядра отходят аксоны, образующие красноядерно-спинномозговой путь (руброспинальный тракт). Волокна этого пути оканчиваются преимущественно на вставочных нейронах, находящихся в проме- жуточной зоне серого вещества спинного мозга, но часть аксонов контактирует непосредственно с мотонейронами спинного мозга.
СТВОЛ МОЗГА
Ствол мозга состоит из продолговатого мозга, моста и среднего мозга и содержит двигательные и чувствительные ядра, исполняю- щие моторные и сенсорные функции для лица и головы таким же образом, которым спинной мозг выполняет эти функции по отношению к шее, туловищу и конечностям. В то же время ствол мозга осуществляет множество специальных функций (в том числе функции контроля дыхания, сердечно-сосудистой системы, ЖКТ, многих стереотипных движений тела, равновесия, движения глаз) и служит как бы узловой станцией для «командных сигналов» от вышележащих центров. В контроле над движениями тела и его равновесием важную роль играют вестибулярные и ретикулярные ядра ствола мозга.
• Ретикулярные ядра. На рис. 14-3, В показано расположение ретикулярных ядер. Их подразделяют на ретикулярные ядра моста и ретикулярные ядра продолговатого мозга. Эти две системы ядер функционируют антагонистически по отношению друг к другу: ядра моста возбуждают антигравитационные мышцы, ядра продолговатого мозга тормозят их. Возбуждающие и тормозящие ретикулярные ядра - обязательная часть контролирующей системы, которой управляют сигналы из моторной коры; кроме того, эти ядра создают основной уровень тонического сокращения для противостоя-
ния силам гравитации и могут тормозить отдельные группы мышц, чтобы обеспечить другие функции.
• Вестибулярные ядра функционально связаны с ретикулярными ядрами моста, возбуждая антигравитационные мышцы. Латеральные вестибулярные ядра передают сильные возбуждающие сигналы в латеральный и медиальный преддверноспинномозговой путь. Без участия вестибулярных ядер ретикулярная система моста значительно ослабляет своё возбуждающее влияние на гравитационную мускулатуру шеи, спины, верхних и нижних конечностей. Специфическая роль вестибулярных ядер заключается в селективном контроле возбуждающих сигналов, поступающих из вестибулярного аппарата к различным антигравитационным мышцам для поддержания равновесия (см. гл. 11).
БАЗАЛЬНЫЕ ЯДРА
Термин «базальные ядра» применяют по отношению к пяти структурам: это хвостатое ядро, скорлупа, субталамическое (подбугорное) ядро, бледный шар и чёрное вещество (рис. 14-4, А). Хвостатое ядро и скорлупу называют иногда полосатым телом.
Базальные ядра и их связи с другими структурами мозга - сложный комплекс с огромным количеством связей, обеспечивающий координацию двигательных функций (рис. 14-4, Б). Важное место, определяющее физиологическую роль базальных ядер, занимают две нейронные системы - скорлупы и хвостатого ядра.
• Скорлупа (см. рис. 14-4, Б) имеет входы преимущественно из прилежащих к первичной моторной коре областей, но не из самой первичной моторной коры. Выходы из системы скорлупы осуществляются в основном в первичную моторную кору, премоторную и дополнительную моторную области. Одна из главных задач базальных ядер (в том числе скорлупы) при осуществлении двигательного контроля - контроль за комплексными стереотипами моторной деятельности (например, написание букв алфавита).
• Хвостатое ядро. На рис. 14-5, Б показаны связи между кортикоспинальной двигательной системой и хвостатым ядром. Система хвостатого ядра простирается во все доли мозга, включая лобную, теменную и затылочную. Хвостатое ядро получает
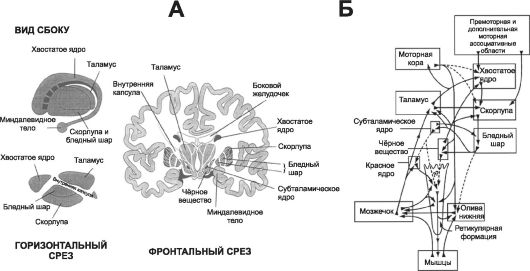
Рис. 14-4. Базальные ядра мозга и их связи с другими системами. А - анатомия базальных ядер; Б - связи базальных ядер с кортико-спинальной и мозжечковой системами, контролирующими движения.
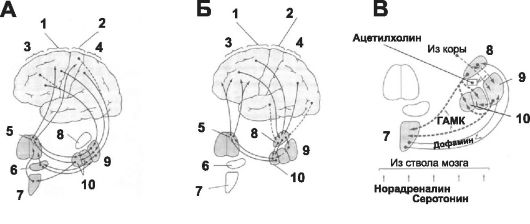
Рис. 14-5. Базальные ядра мозга и контроль двигательных функций.
А - контроль приобретённых моторных навыков; Б - сознательное планирование движений; В - нейромедиаторы: 1 - премоторная и дополнительная моторная области, 2 - первичная моторная кора, 3 - префрон- тальная кора, 4 - соматосенсорная область, 5 - переднемедиальное и переднелатеральное ядра таламуса, 6 - субталамическое ядро, 7 - чёрное вещество, 8 - хвостатое ядро, 9 - скорлупа, 10 - бледный шар.
• много информации из ассоциативных областей коры, которые интегрируют различные виды сенсорной и моторной информации, чтобы формировать программы стереотипных движений. Из коры сигналы: поступают в хвостатое ядро, затем передаются в бледный шар, оттуда в релейные ядра таламуса и снова поступают обратно в префронтальную, премоторную и дополнительную моторные области коры. Анатомические особенности системы хвостатого ядра находят объяснение в его функции: хвостатое ядро играет важную роль в сознательном (когнитивном) контроле двигательной активности. Действительно, большинство наших двигательных актов возникает в результате обдумывания их и сопоставления с информацией, имеющейся в памяти. Нейромедиаторы базальных ядер. Взаимодействие между нейронами базальных ядер осуществляют различные нейромедиаторы (рис. 14-5, В): дофамин, ГАМК, ацетилхолин, норадреналин, серотонин, энкефалин и глутамат.
МОЗЖЕЧОК
Мозжечок располагается, «оседлав» главные сенсорные и моторные системы ствола большого мозга (рис. 14-6, А). Он связан с мозговым стволом на каждой стороне верхней, средней и нижней ножкой. Масса мозжечка только на 10% больше массы коры мозга, но поверхность мозжечка составляет около 75% поверхности коры больших полушарий. С функциональной точки зрения мозжечок подразделяется на три части (рис. 14-6, Б).
• Узел червя и краевые дольки полушарий с каждой стороны образуют клочково-узелковую долю, или vestibulocerebellum
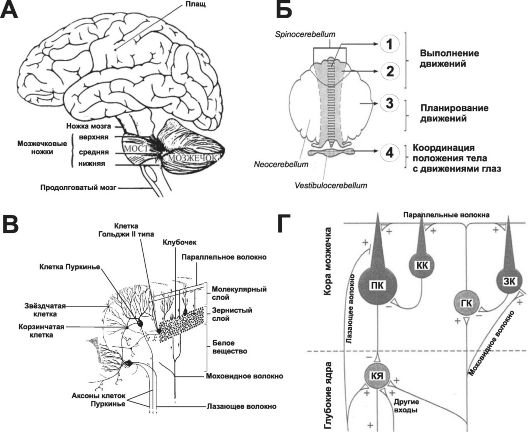
Рис. 14-6. Мозжечок. А - топография; Б - функциональная анатомия: 1 - медиальная нисходящая система, 2 - латеральная нисходящая система, 3 - к моторной и премоторной коре, 4 - к вестибулярным ядрам; В, Г - связи: ПК - Пуркинье клетки, КК - корзинчатые клетки, ЗК - зернистые клетки, ГК - Гольджи клетки, КЯ - клетки глубоких ядер.
(вестибулярный мозжечок). Эта наиболее древняя часть мозжечка (archicerebellum) имеет соединения с вестибулярным аппаратом, связана с равновесием и вызванными научением вестибуломоторными рефлексами.
• Оставшаяся часть червя и прилежащие медиальные порции полушарий образуют переднюю долю - spinocerebellum (paleocerebellum, спинальный мозжечок/ Эта область мозжечка получает проприоцептивную информацию, а также копию «моторного плана» из моторной коры. Сравнивая план с исполнением, spinocerebellum сглаживает и координирует движения, определяя их последовательность. Из области червя в мозговой ствол поступают сигналы в отношении контроля околопозвоночных мышц и проксимальных мышц конечностей. Из медиальных отделов полушарий направляются импульсы в мозговой ствол для контроля дистальных мышц.
• Латеральные отделы: полушарий мозжечка составляют заднюю долю и называются новым мозжечком - neocerebellum. Они взаимодействуют с моторной корой при планировании и программировании движений.
Организация мозжечка
Кора мозжечка отделена белым веществом от глубоких ядер мозжечка (ядро шатра, зубчатое ядро, промежуточное ядро, раз- деляющееся на шаровидное и пробковидное). Первичные афферентные входы (моховидные и лазающие волокна) посылают коллатерали к глубоким ядрам и проходят к коре (рис. 14-6, В, Г). Большинство выходящих путей направляется в ствол мозга, оставшиеся пути от мозжечковой коры проецируются к глубоким ядрам, которые, в свою очередь, имеют связи с мозговым стволом. Таким образом, глубокие ядра принимают сигналы из spinocerebellum и neocerebellum.
• Мозжечковые афферентные пути весьма многочисленны. Мозжечок получает сигналы от мышц, спинного и продолговатого мозга, ретикулярной формации, разных структур среднего мозга, а также от коры больших полушарий.
• Мозжечковые эфферентные пути. Глубокие ядра мозжечка получают сигналы: из двух источников: из коры мозжечка и сенсорных афферентных трактов, направляющихся в мозжечок. Входные сигналы к мозжечку подразделяются и идут в двух
направлениях - непосредственно к одному из глубоких мозжечковых ядер и в соответствующую область коры мозжечка, перекрывающую глубокие ядра. Из этой области коры через доли секунды обратно к глубоким ядрам поступают тормозные сигналы.
Кора мозжечка
Кора мозжечка содержит несколько типов нейронов: клетки Пуркинье, зернистые, корзинчатые, звёздчатые и клетки Гольджи. Кора подразделяется на три слоя (см. рис. 14-6, В): наружный - молекулярный, слой клеток Пуркинье и внутренний - зернистый.
• Афферентные входы. Кора мозжечка имеет два главных входа - моховидные и лазающие волокна (см. рис. 14-6, В), по которым поступают возбуждающие сигналы, конвергирующие от первичных афферентных систем. Лазающие волокна приходят из единственного источника - ядер нижней оливы, получающих проприоцептивные сигналы из всех отделов тела. Каждое лазающее волокно окружает дендриты клеток Пуркинье наподобие вьюнка или плюща. Моховидные волокна обеспечивают прямой проприоцептивный вход от всех частей тела плюс вход от коры больших полушарий через ядра моста. Моховидные волокна оканчиваются на дендритах зернистых клеток в виде синаптического комплекса (клубочек). Клубочки также содержат тормозные окончания, приходящие от клеток Гольджи.
• Функциональные единицы коры мозжечка. Кора
мозжечка имеет около 30 млн функциональных единиц. Центром отдельной
функциональной единицы является клетка Пуркинье. Система нейронных
взаимодействий в функциональной единице схематически представлена на
рис. 14-
заканчивающихся на клетках Пуркинье, глутамата, который
задерживает возникновение разрядов клеток Пуркинье.
Клетки Гольджи возбуждаются импульсами от коллатералей моховидных волокон, коллатералей клеток Пуркинье и параллельных волокон и тормозят передачу импульсов от моховидных волокон к зернистым клеткам. Нейромедиатором, выделяющимся звёздчатыми и корзинчатыми клетками, а также клетками Гольджи и Пуркинье, является ГАМК; зернистые клетки выделяют глутамат.
Клетки Пуркинье в свою очередь оказывают тормозящее влияние на клетки глубоких ядер. К этим ядрам по коллатералям от моховидных и лазающих волокон поступают возбуждающие импульсы. Имея тормозящий вход от клеток Пуркинье, глубокие ядра мозжечка на выходе всегда посылают только возбуждающие сигналы в ствол мозга и таламус. В состоянии покоя клетки Пуркинье и клетки глубоких ядер находятся в состоянии постоянного возбуждения. Клетки Пуркинье разряжаются с частотой 50-100 Гц, клетки глубоких ядер разряжаются с ещё большей частотой. Таким образом, функциональные нейронные единицы мозжечка модулируют и синхронизируют во времени возбуждающие сигналы от глубоких мозжечковых ядер к стволу мозга и таламусу.
Мозжечок в системе двигательного контроля
В системе контроля и координации движений мозжечок принимает участие на трёх уровнях.
• Vestibulocerebellum обеспечивает движения, необходимые для поддержания равновесия.
• Spinocerebellum обеспечивает координацию преимущественно дистальных отделов конечностей (особенно рук и пальцев рук).
• Neocerebellum получает все связи из моторной коры и прилежащих областей премоторной и соматосенсорной зоны мозга. Он передаёт сигналы обратно в большой мозг, планируя последовательность действий вместе с сенсомоторной областью и рассчитывая на десятки секунд вперёд будущие действия.
Функции vestibulocerebellum
• У лиц с вестибуломозжечковыми расстройствами равновесие чаще нарушается при попытках быстрых движений, чем во
время покоя. Особенно это проявляется при попытках изменить направление движения тела. Это свидетельствует, что vestibulocerebellum контролирует баланс между агонистическими и антагонистическими сокращениями мышц позвоночника, бедра и плечевого пояса во время быстрых изменений положений тела.
• Для контроля над равновесием информация от периферии тела и вестибулярного аппарата используется как типичная система обратной связи, обеспечивающая опережающую коррекцию постуральных двигательных сигналов, которые необходимы для поддержания устойчивости во время быстрого движения, включая изменяющееся направление движений.
Функции spinocerebellum
• На рис. 14-7 схематически показано, что каждое из полушарий мозжечка получает два вида информации. В момент начала движения поступает информация из моторной коры и красного ядра, сообщающая мозжечку о последовательности предполагаемого плана движений. В это же время в мозжечок приходит информация от периферических отделов тела (в особенности от проприорецепторов конечностей), говорящая моз-
жечку о характере реального движения. После сравнения планируемого и реального движений клетки глубокого промежуточного ядра посылают корригирующие сигналы в моторную кору через релейные ядра таламуса и в красное ядро, откуда сигналы поступают в руброспинальный тракт для исполнения.
• Spinocerebellum обеспечивает плавность, координированность движений агонистов и антагонистов, сравнивая планируемые корой движения с движения-
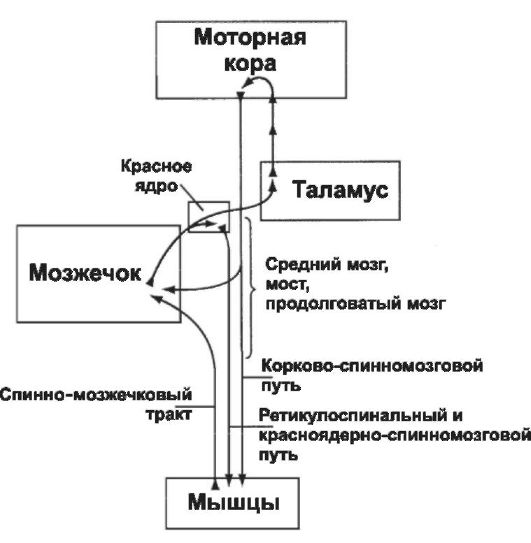
Рис. 14-7. Центральный контроль произ- вольных движений.
ми, реально выполняемыми. Это осуществляется при помощи переднего спинно-мозжечкового тракта, передающего в мозжечок «копии» реальных моторных сигналов.
• Почти все движения нашего тела «маятникообразны». Например, при движении руки имеется инерция выполнения и возможна инерция превышения до того, как движение будет остановлено. В силу инерции все маятникообразные движения имеют тенденцию к превышению. Если превышающий норму размах движений возникает у человека с повреж- дённым мозжечком, то с помощью сознания он распознаёт это и пытается сделать движение в обратном направлении. Но конечность (из-за инерции и нарушенного мозжечкового механизма коррекции) продолжает колебаться вперёд и назад, пока рука не вернётся в исходное положение. Этот феномен - тремор действия, или интенционный тремор. Если же мозжечок не повреждён и соответственно обучен, то подсознательные сигналы точно остановят движение в заданной точке и предотвратят тремор. Эту демпфирующую функцию выполняет spinocerebellum.
• К функции spinocerebellum относится контроль за очень быстрыми короткими движениями, называемыми баллистическими (например, печатание на клавиатуре компьютера или саккадические движения глазного яблока). После удаления мозжечка движения начинаются и заканчиваются медленно, и они слабее, т.е. утрачивается привычный автоматизм баллистических движений.
Функции neocerebellum
• Латеральные зоны полушарий мозжечка осуществляют планирование последовательности движений вместе с премоторной и сенсорной областями коры мозга при постоянной двусторонней связи коры больших полушарий с базальными ядрами. «План» последовательных движений возникает в сенсорной и премоторной зонах коры, оттуда он передаётся в латеральные отделы полушарий мозжечка. Затем по многим двусторонним связям между мозжечком и корой мозга необходимые двигательные сигналы обеспечивают переход от одного движения к следующему. Важно, что в нейронах глубоких зубчатых ядер мозжечка появляются паттерны импульсной активности для
последующих движений в тот момент, когда настоящие движения ещё только начинаются. Следовательно, латеральные отделы полушарий мозжечка оказываются вовлечёнными не в движение, которое в данный момент происходит, а в события, которые произойдут во время следующего последовательного движения, долями секунды позже.
• Важной функцией neocerebellum является расчёт времени для каждого последующего движения. Удаление латеральных отделов полушарий мозжечка приводит к потере подсознательной способности рассчитывать время возникновения тех или иных движений тела. Без этой способности человек не может определить, когда начнётся следующее движение. В результате оно может начинаться или слишком рано, или слишком поздно. Подобные мозжечковые нарушения называются потерей плавного развития движений.
• Neocerebellum играет роль в предсказании временной последовательности не только движений, но и других систем организма. В частности, на основании зрительных наблюдений можно предсказать, как быстро тот или иной движущийся предмет приблизится к какому-нибудь объекту. Удаление части латеральных отделов мозжечка у обезьяны приводит к тому, что она постоянно ударяется головой о стенку коридора, так как утрачивает способность предсказывать время соприкосновения со стенкой.
Мозжечок и обучение движениям
Степень участия мозжечка в координации движений и обучении выявляется при попытках совершить новые моторные акты. Как правило, новые движения вначале неуверенны, неточны, требуют больших усилий. После многократных повторений движения становятся более точными и легковоспроизводимыми. Базой для такого обучения является вход через ядра оливы. Каждая клетка Пуркинье получает на входе от 250 тыс. до 1 млн моховидных волокон и только одно лазающее волокно из нижней оливы, но это лазающее волокно образует 2-3 тыс. синапсов на клетке Пуркинье. Активация лазающего волокна вызывает большой комплексный разряд (спайк) в клетке Пуркинье; а он в свою очередь обусловливает долговременное стойкое изменение спектра активности входа
моховидных волокон в этой же клетке Пуркинье. Активность лазающих волокон увеличивается при обучении новым движениям. Избирательное поражение оливарного комплекса нарушает способность к регулированию двигательных актов.
Клинические проявления нарушений функций мозжечка
Серьёзные и длительные нарушения функций мозжечка возникают при поражении одного или нескольких глубоких ядер и мозжечковой коры. Мозжечковые симптомы определяются местом повреждения.
Дисметрия - неспособность правильно определять расстояние и, как следствие, неспособность сразу взять необходимый предмет.
Атаксия - нарушение координации движений, неспособность выполнять движения в правильном порядке и последовательности. Больным трудно ходить, особенно в темноте, им приходится хвататься за что-нибудь руками; походка напоминает походку пьяного: человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы.
Асинергия - неспособность в определённом порядке активировать мышцы в разных областях тела. Если больной в положении стоя пытается отклонить голову назад, он может упасть.
Адиадохокинез - неспособность быстро вращать ладони вниз и вверх.
Дизартрия - нарушение координации мышц лица и быстрых последовательных движений гортани, губ и дыхательной системы. Речь становится медленной, невыразительной, монотонной, скандированной.
Интенционный тремор - усиливающийся в конце движения, когда больной пытается дотронуться до предмета; его рука дрожит, совершая всё более размашистые движения. Клинически тестируется пальценосовой пробой - с закрытыми глазами дотронуться указательным пальцем до кончика носа.
Мозжечковый нистагм - колебание глазных яблок при попытке фиксировать взгляд на каком-либо предмете при взгляде в сторону.
Мышечная гипотония возникает при поражении глубоких мозжечковых ядер.
Обобщение главы
Спинной мозг является источником рефлексов, которые необходимы для инициации и контроля движений.
Функции спинного мозга находятся под контролем высших центров мозгового ствола.
Наивысший уровень моторного контроля осуществляется корой головного мозга.
Базальные ганглии и мозжечок обеспечивают механизмы обратной связи моторного контроля, осуществляемого из областей коры мозга и мозгового ствола.