Патофизиология Новицкого, Е.Д. Гольдберга Тома 1 и 2 - 2009 г.
|
|
|
|
ЧАСТЬ II ТИПОВЫЕ ПАТОЛОГИЧЕСКИЕ ПРОЦЕССЫ ГЛАВА 7 ПАТОФИЗИОЛОГИЯ ИММУНИТЕТА
7.1. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ИММУННОЙ
СИСТЕМЫ
7.1.1. Основные понятия
Иммунитет (от лат. immunio - укреплять, защищать) - это феномен биологической защиты организма от чужеродных патогенов.
Смысл подобной защиты - обеспечение генетической целостности организма в течение всей его жизни путем поддержания баланса между «своим» и «несвоим» в условиях чужеродного окружения. Однако иммунитет напрямую не контролирует генетический гомеостаз организма. Геном как таковой не является объектом действия для факторов иммунитета, распознающих молекулы поверхности клеток и межклеточного матрикса (то, что в классической генетике называют фенотипом).
Иммунная система специализируется на реализации механизмов данного феномена. Чтобы понять основы науки иммунологии, необходимо изучить защитные механизмы, направленные на устранение «несвоего», и регуляторные процессы, которые формируют внутренний гомеостаз «своего» в организме. Иммунитет может быть врожденным и приобретенным (адапативным).
Врожденный иммунитет обусловливается наследственными анатомо-физиологическими особенностями организма и представляет собой совокупность факторов и механизмов неспецифической защиты. К ним относятся:
• механические барьеры (кожа, слизистые);
• нормальная непатогенная микрофлора (аутофлора) кожи и слизистых оболочек дыхательных путей, желудочно-кишечного и мочеполового трактов;
• физико-химические барьеры: а) лизоцим крови, лимфы, слизистых оболочек и секретов (слезная и семенная (эякулят) жидкость, материнское молоко); б) антимикробные пептиды (кателицидины, дефенсины и др.) эпителия кожи, дыхательных путей, мочеполового тракта; в) соляная кислота (HCl) желудочного сока; г) пищеварительные ферменты, альдегиды и жирные кислоты сальных и потовых желез; д) монооксигеназная система печени (цитохром Р450);
• естественные натуральные антитела, представляющие собой IgM, которые постоянно присутствуют в крови и направлены против условно-патогенной флоры; каталитические антитела, обладающие протеиназной и нуклеазной активностью;
• система естественной цитотоксичности (натуральные киллеры - NK, интерфероны α, β, γ), обеспечивающие экстренную защиту против вирусов и других внутриклеточных патогенов без воспалительной реакции, а также индукцию апоптоза HLA I-негативных «своих» инфицированных, опухолевых, деградирующих и поврежденных клеток;
• фагоциты профессиональные (макрофаги, нейтрофилы) и вспомогательные (или факультативные - дендритные клетки, фибробласты и др.);
• система комплемента (подробнее см. раздел 14.4.6);
• белки теплового шока (HPS - heat shock proteins) - белки клеток, повышающие термотолерантность, участвующие в регуляции апоптоза и образовании комплексов с чужеродными (антигенными) пептидами, облегчая связывание и поглощение «несвоего» макрофагами;
• белки острой фазы, образующиеся в печени в ответ на повреждение клеток и тканей (например, С-реактивный белок) (подробнее см. раздел 4.2).
Если патогену удается миновать неспецифический эшелон защиты, включаются механизмы адаптивного иммунитета.
Адаптивный иммунитет вырабатывается в процессе жизни индивида и представляет собой специфическую защитную реакцию организма на конкретный чужеродный агент (антиген) с участием лимфоцитов.
В его основе лежит способность лимфоцитов распознавать антигены и организовывать их деструкцию, в то время как врожденный иммунитет не дифференцирует антигены и реализуется в отношении любого антигена вне зависимости от его природы. Кроме того, для формирования адаптивного иммунитета сначала необходима
предварительная обработка (процессинг) «несвоего», после чего происходит его распознавание и деструкция; факторы врожденного иммунитета (фагоциты, система комплемента и др.) распознают «чужое» в исходной, необработанной форме («паттерны»).
Отказ одного или нескольких компонентов иммунной системы может приводить к развитию иммунодефицитов и утрате способности к защите от «несвоего». Расстройства регуляции в функционировании данной системы обусловливают аутоиммунные и аллергические заболевания (на основе I, II, III, IV и V типов гиперчувствительности), частые, тяжелые, затяжные и рецидивирующие инфекции, рост опухолей. Важность самого существования иммунной системы иллюстрируется появлением в последние 20 лет новой болезни - СПИДа (синдрома приобретенного иммунодефицита), при которой выявляются различные варианты нарушений со стороны иммунной системы.
Антигены - это генетически чужеродные вещества, вызывающие иммунный ответ. Общее число различных антигенов оценивается числом 1018. Основными свойствами антигена, определяющими его иммуногенность, т.е. способность вызывать иммунный ответ, являются макромолекулярность (молекулярная масса не менее 10 кДа) и генетическая чужеродность.
Полные антигены - это макромолекулярные соединения (в основном белки и белоксодержащие соединения). Молекула полного антигена состоит из двух частей - информационной и несущей. Информационная часть (низкомолекулярные антигенные детерминанты, эпитопы) определяет специфичность антигена, несущая (макромолекулярный белок) - способствует проникновению антигена в организм.
Неполные антигены (или гаптены) - это низкомолекулярные небелковые антигены (лекарственные препараты, вирусы и др.), которые имеют в своем составе только информационную часть. Изолированная информационная часть сама по себе не является иммуногенной, т.е. не способна индуцировать иммунный ответ, но обладает конкретной специфичностью - свойством вступать в реакции взаимодействия с предсуществующими (ранее образованными при предыдущих контактах) антителами и лимфоцитами. Гаптены приобретают иммуногенность лишь в соединении с белками-носителями организма.
Антитело (иммуноглобулин) - это один из типов молекул иммунной системы. Антитела и антигенраспознающие рецепторы могут связывать соответствующие антигены.
Паттерны связываются паттернраспознающими молекулами (toll-подобными рецепторами1 и др.).
Лимфоциты являются главными клетками иммунной системы. Посуществу, иммунная система - это иерархическая совокупность лимфоидных клеток (1013). Существуют T-, B- и NK-лимфоциты. Т-клетки дифференцируются в тимусе и играют ключевую роль во всех направлениях специфического иммунного ответа. В-лимфоциты дифференцируются в костном мозгу, являются предшественниками плазмоцитов - антителопродуцентов. NK-клетки (натуральные или естественные киллеры) участвуют в неспецифической цитотоксичности по отношению к внутриклеточно расположенным патогенам. Остальные клетки (макрофаги, дендритные клетки, нейтрофилы, эозинофилы, базофилы, тучные клетки и др.) также участвуют во многих иммунных процессах, но их участие скорее опосредованное, т.е. они привлекаются лимфоцитами для реализации функций иммунной (лимфоидной) системы.
Функции иммунной системы:
1. Защита от «несвоего» (инфекция, трансплантат).
2. Элиминация модифицированного «своего»: трансформированных (в том числе опухолевых), инфицированных, поврежденных, стареющих клеток.
3. Регуляция роста и развития клеток и тканей.
1 Факторы врожденного иммунитета распознают консервативные, присущие только микроорганизмам и отсутствующие у позвоночных (от мыши до человека) молекулярные структуры, обозначаемые в современной литературе термином «патогенассоциированные молекулярные структуры» (patogen associated molecular patterns - PAMP). Эти структуры распознаются специальными рецепторами - паттернраспознающими рецепторами (pattern recognition receptors - PRRs). PRRs (растворимые, внутриклеточные, мембраносвязанные) рассматриваются как носители эволюционной памяти многоклеточных организмов о «своем» и «несвоем», обеспечивающие экстренный неспецифический ответ на чужеродный агент. Разновидностью мембраносвязанных PPRs являются toll-подобные рецепторы (TLRs). TLRs экспрессированы на многих клетках: дендритных клетках, макрофагах, моноцитах, нейтрофилах, В-, Т-лимфоцитах и др. Результатом связывания TLRs с лигандами микроорганизмов (или их продуктов) является активация в TLR-несущих клетках цитоплазматических адапторных молекул (MyD88 и др.), киназ (МАРК), ядерного фактора транскрипции (NF-kB), опосредующих индукцию генов адгезивных молекул, цитокинов и, как следствие, развитие воспаления. PAMP, распознаваемые TLRs, описаны у многих грамположительных и грамотрицательных бактерий, вирусов, грибов и простейших.
Главными «партнерами» иммунной системы являются центральная нервная система, эндокринная система и печень, которые наиболее важны для обеспечения регуляции гомеостаза.
Функциональная организация иммунной системы может быть рассмотрена на органном, клеточном и молекулярном уровнях. Существуют два типа органов иммунной системы - центральные (или первичные) и периферические (или вторичные).
К центральным органам иммунной системы относятся костный мозг и тимус.
К периферическим органам иммунной системы относятся:
1. Лимфатические узлы, лимфатические протоки и селезенка.
2. Лимфоидная ткань, ассоциированная со слизистыми оболочками (Mucous-Associated Lymphoid Tissue - MALT), которая расположена на пяти уровнях. Первые два уровня - евстахиева труба (Tube-Associated Lymphoid Tissue - TALT) и носоглотка (NasalAssociated Lymphoid Tissue - NALT) - представлены различными миндалинами; следующий уровень - бронхи (Bronchus-Associated Lymphoid Tissue - BALT) и грудные железы (у женщин); четвертый уровень - верхние отделы желудочно-кишечного тракта (GutAssociated Lymphoid Tissue - GALT) - содержат лимфоидную ткань желудка и пейеровы бляшки тонкого кишечника; пятый уровень - это нижние отделы желудочно-кишечного тракта и мочеполовая система - аппендикс, солитарные фолликулы толстого кишечника и лимфоидная ткань мочеполовой системы.
3. Лимфоидная ткань, ассоциированная с кожей (Skin-Associated Lymphoid Tissue - SALT).
Центральные органы являются местом лимфопоэза, т.е. образования и антигеннезависимого созревания лимфоцитов. В костном мозгу находятся ранние клетки-предшественницы Т- и В-лимфоцитов (преТ- и преВ-клетки), В-лимфобласты созревают из преВ-клеток, размножаются и дифференцируются в В-лимфоциты. Тимус - орган, в котором Т-лимфобласты размножаются и дифференцируются в Т-лимфоциты. В периферических органах происходит контакт Т- и В-лимфоцитов с антигеном и иммуногенез - антигензависимый этап созревания лимфоцитов. Этот процесс называется иммунным ответом. Его сутью является создание «армии» (клонов) специфически реагирующих лимфоцитов и специфических антител, которые осуществляют эффекторные реакции по уничтожению конкретного антигена. Существуют
Т-клеточный и В-клеточный (гуморальный) пути иммунного ответа.
Большой вклад в развитие иммунологии от Edward Jenner до наших дней внесли L. Pasteur, И.И. Мечников, P. Ehrlich, Н.Ф. Гамалея, F. McFarlane Burnet, N.K. Jerne, Р.В. Петров, S. Tonegawa, Р.М. Хаитов и др.
7.1.2. Клетки иммунной системы
Все клетки, относящиеся к иммунной системе и привлекаемые ею для обеспечения эффекторных реакций, в функциональном отношении условно разделяют на четыре группы.
1. Антигенпрезентирующие клетки: макрофаги, дендритные клетки типов 1 и 2, В-лимфоциты.
2. Регуляторные клетки: T-индукторы, Т-хелперы типов 1, 2 и 3, естественные регуляторные Т-клетки.
3. Эффекторные клетки: плазматические клетки (дифференцирующиеся из В-лимфоцитов), цитотоксические Т-клетки с фенотипом CD8+ (или T-киллеры); эффекторные Т-клетки воспаления с фенотипом CD4+ (или Т-лимфоциты, ответственные за гиперчувствительность замедленного типа); нейтрофилы, эозинофилы, базофилы, тучные клетки, натуральные киллеры (NK-клетки), макрофаги.
4. Клетки памяти: Т-клетки памяти с фенотипом CD8+; Т-клетки памяти с фенотипом CD4+; долгоживующие плазматические клетки; В-клетки памяти.
Номенклатура CD («Cluster of Differentiation»), которая была разработана нобелевскими лауреатами (1984) G.J.F. Kohler (Швейцария) и C. Milstein (Аргентина/Великобритания), основана на моноклональной технологии. Она позволяет идентифицировать клетки в соответствии с их происхождением, стадией дифференцировки, функциональным состоянием и т.д. (см. табл. 7-1). Данная технология, без сомнения, оказалась революционной в иммунологических и смежных областях исследований.
Лимфоциты как главные клетки иммунной системы имеют отличительные особенности:
1. Постоянная «патрульная» рециркуляция по кровотоку, лимфотоку, межтканевым пространствам и секретам.
2. Способность распознавать, т.е. взаимодействовать со «своим» и «чужим» по принципу «лиганд - рецептор».
Taблица 7-1. Главные идентификационные CD-маркеры клеток иммунной системы
Обозначение кластера | Клетки |
CD10, CD34 | Лимфоидная стволовая клетка |
CD3 | T-лимфоцит |
CD4 | Т-хелпер/Т-индуктор |
CD8 | Цитотоксический T-лимфоцит |
CD19, CD72, CD79a/b и др. | В-лимфоцит |
CD16/CD56 | Натуральный киллер (NK-клетка) |
CD64 | Moноцит/макрофаг |
3. Клональная (или групповая) организация (McF. Burnet) и способность формировать сетевые элементы (N.K. Jerne).
4. Способность к непрерывным реаранжировкам в своем геноме в любом возрасте в связи с потребностями формирования специфического ответа на патоген.
5. Умение запоминать антигены и обеспечивать в будущем экспрессный высокоэффективный антигенспецифический ответ.
Клон - это группа лимфоцитов, коммитированных к определенному антигену. До встречи с этим антигеном каждый лимфоцит клона называют наивным. По-видимому, в человеческом организме исходно существуют десятки миллионов клонов Т- и В-лимфоцитов. После контакта с соответствующим антигеном и в результате иммунного ответа коммитированный лимфоцит становится праймированным.
7.1.3. Молекулы иммунной системы
Для осуществления необходимых функций клетки иммунной системы имеют сложную молекулярную организацию своих рецепторов и способны к выработке целого ряда молекул.
1. Антигенпредставляющие, антигенраспознающие и антигенсвязывающие молекулы. Набор этих молекул уникален для каждого организма, для каждого лимфоцитарного клона и для каждого специфического иммунного ответа. К ним относятся:
• свободные иммуноглобулины: IgM, IgG, IgA, IgE, IgD;
• антигенпредставляющие молекулы: лейкоцитарные антигены главного комплекса гистосовместимости (human leukocyte antigens - HLA I и II) и молекулы CD1 (a, b, c, d, e);
• иммуноглобулиновые рецепторы В-клеток для распознавания и связывания антигена (B cellular receptors - BCR);
• рецепторы Т-клеток для распознавания и связывания антигена (T cellular receptors - TCR);
2. Адгезивные молекулы опосредуют взаимодействия между клетками и лигандами при непосредственном контакте:
• суперсемейство иммуноглобулиноподобных молекул;
• интегрины;
• селектины;
• муцины;
• суперсемейство рецепторов к факторам некроза опухоли и фактору роста нервов - TNF/NGF (или молекулы, опосредующие апоптоз);
• линк-семейство (компоненты экстрацеллюлярного матрикса).
3. Иммуноцитокины являются гормонами иммунной системы, действующими чаще с пара- и аутокринным, реже - с эндокринным эффектами:
• интерлейкины (interleukins - ILs);
• колониестимулирующие факторы (colony-stimulating factors -
CSFs);
• интерфероны (interferons - IFNs);
• факторы некроза опухоли (tumor necrosis factors - TNFs);
• трансформирующие факторы роста (transforming grows factors -
TGFs);
• хемокины и др.
4. Сборная группа различных медиаторов воспаления включает белки комплемента, белки острой фазы, простаноиды и лейкотриены, протеолитические ферменты и др.
Иммуноглобулины (immunoglobulin - Ig) M, G, A, E и D являются эффекторными молекулами гуморального иммунного ответа. Молекула иммуноглобулина является гликопротеином; белковые цепи включают сотни aминокислотных последовательностей; углеводный компонент составляет до 12%. Молекула IgG (рис. 7-1) состоит из двух идентичных легких (light - L) и двух идентичных тяжелых (heavy - H) цепей (chains). Легкие цепи бывают двух типов: χ и λ, а тяжелые - пяти: μ, γ, α, ε и δ. Легкие и тяжелые цепи содержат повторяющиеся гомологичные последовательности и об-
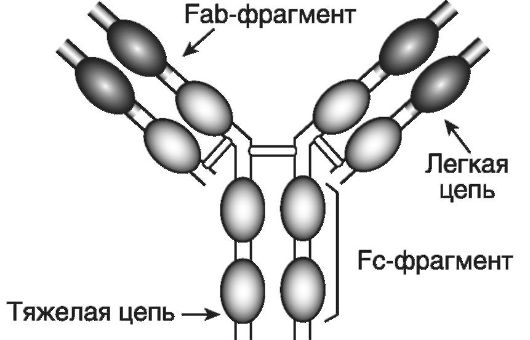 Рис. 7-1. Структура
молекулы IgG. Молекула IgG состоит из двух идентичных легких и двух
идентичных тяжелых цепей, которые образуют три фрагмента: два Fab
(антисвязывающие сайты) и один Fc (неспецифические эффекторные функции).
Цепи укладываются таким образом, что возникают «клубки» - домены,
которые могут быть вариабельными или константными
Рис. 7-1. Структура
молекулы IgG. Молекула IgG состоит из двух идентичных легких и двух
идентичных тяжелых цепей, которые образуют три фрагмента: два Fab
(антисвязывающие сайты) и один Fc (неспецифические эффекторные функции).
Цепи укладываются таким образом, что возникают «клубки» - домены,
которые могут быть вариабельными или константными
разуют своеобразные домены (клубки или глобулы). Существуют константные (constant) (CL, CH1, CH2 и CH3) и вариабельные (variable) домены (VL и VH). Гипервариабельные области вариабельных доменов формируют антигенсвязывающие сайты (активные центры). Участок CL-CH1 определяет различия амонокислотных последовательностей у индивидуумов одного и того же вида. Антигенсвязыгаающие сайты и область CL-CH1 входят в состав так называемых Fab (fragment antigen-binding)-фрагментов (их 2), ответственных за связывание антигена, а участки CH2-CH2 и СН3- СН3 формируют Fc (fragment crystalline)-фрагмент, отвечающий за неспецифические эффекторные функции. Например, область CH2-CH2 участвует в фиксации и активации компонентов комплемента, а область СН3-СН3 - в фиксации антител к клеткам. Между Fab- и Fc-фрагментами тяжелых цепей имеется шарнирная область, обеспечивающая гибкость молекулы антитела.
IgM являются эволюционно наиболее древними антителами и составляют 5-10% всех иммуноглобулинов. Они являются также первыми антителами новорожденных, не проникают через плаценту. вырабатываются только при первичном иммунном ответе. Они появляются в крови в начале иммунного ответа и затем замещаются IgG. служат основными рецепторами для распознавания антигенов на поверхности зрелых В-лимфоцитов. В
сыворотке крови существуют в виде пентамеров (т.е. имеют 5 мономерных субъединиц). связывают комплемент и активируют его по классическому пути.
На IgG в сыворотке крови человека приходится около 75% от общего числа иммуноглобулинов. Вырабатываются они как при первичном, так и при вторичном иммунном ответе вслед за IgM. IgG способны проникать в ткани, через плаценту, связывать комплемент и активировать его по альтернативному пути. Каждая молекула IgG состоит из 2 легких и 2 тяжелых цепей. У человека имеется 4 подкласса IgG, различающихся последовательностью аминокислот в тяжелой цепи.
IgA составляют 7-15% общего количества сывороточных иммуноглобулинов. IgA содержатся в секретах (слюне, слезной жидкости, секретах желудочно-кишечного и мочеполового тракта, слизистой носа и грудной желез), где они находятся в виде секреторного IgA - sIgA - димера, состоящего из 2-х соединенных между собой молекул IgA и гликопротеина, называемого секреторным компонентом (СК). СК образуется в эпителиальных клетках и предохраняет молекулу IgA от расщепления. выходят из кровотока и проникают через эпителиальный слой, соединяясь с СК. Образовавшийся sIgA отщепляется от поверхности эпителиальной клетки и поступает в секреты, где реализует свою защитную функцию - предотвращает проникновение антигена с внешних поверхностей в ткани.
Содержание IgE в сыворотке крови является наиболее низким - около 0,003%. Они накапливаются главным образом в слизистых оболочках и коже, где за счет своего Fс-фрагмента фиксируются на поверхности тучных клеток, базофилов и эозинофилов. При связывании антигена с IgE происходит дегрануляция этих клеток и выброс в кровь биологически активных веществ. Защитная роль IgE до сих пор не выяснена, однако установлено повышение сывороточной концентрации иммуноглобулинов данного типа при инфекциях и аллергических состояниях.
IgD также содержатся в сыворотке в небольших количествах. IgD экспрессируются на поверхностной мембране В-лимфоцитов, формируя вместе с антигенраспознающий В-клеточный рецептор (BCR).
Химическая природа антител была подробно изучена нобелевскими лауреатами (1972) G.M. Edelman (США) и R.R. Porter (Великобритания).
Каждый В-лимфоцит экспрессирует В-клеточный антигенраспознающий рецептор (BCR), который состоит из мономерных IgM и IgD, имеет клональную гетерогенность и ассоциирован с молекулами CD79a и CD79b, необходимыми для проведения сигнала внутрь клетки. Наряду с этими молекулами имеется также корецепторный комплекс (CD19, CD21(CR2), CD81), предназначенный для распознавания HLA II.
Каждый Т-лимфоцит экспрессирует Т-клеточный антигенраспознающий рецептор abTCR), который состоит из двух цепей - a и β, и имеет один из корецепторов - CD4 (у Т-хелперов) или CD8 (у цитотоксических Т-лимфоцитов). Эти инвариантные корецепторы CD4 и CD8 необходимы для распознавания соответственно HLA II или HLA I. Каждая цепь TCR подобно молекуле иммуноглобулина имеет вариабельный и константный домены, что обеспечивает клональную гетерогенность антигенраспознающих рецепторов. Еще одна молекула (CD3) тесно ассоциирована c TCR и служит для проведения сигнала внутрь клетки. CD3 состоит из 5 инвариантных белков (ε, γ, δ, ξ, η). Другой тип антигенраспознающего рецептора - γδTCR - экспрессируется на небольшой субпопуляции Т-клеток. Эти γδT-клетки, роль которых пока до конца не исследована, по-видимому, функционируют подобно NK-клеткам, однако они также имеют определенную клональную гетерогенность.
Синтез и экспрессия на мембране лимфоцитов TCR и ВCR происходят во время лимфопоэза, т.е. при отсутствии антигена. Каждый лимфоцит экспрессирует только один вариант антигенсвязывающего рецептора, т.е. каждый лимфоцит предназначен для распознавания и связывания только одного конкретного антигена.
Молекулы гистосовместимости были открыты нобелевскими лауреатами (1980) В. Benacerraf (США), J. Dausset (Франция) и G.D. Snell (США). Эти молекулы играют решающую роль во многих иммунных процессах, включая загрузку антигенных пептидов и их презентацию. Молекулы HLA разделяются на класс I (A, B, C, E, F, G) и класс II (DR, DP, DQ) в зависимости от их структуры и функции. Экспрессия HLA I имеет место почти на всех клетках (за исключением синцитиотрофобласта), выполняя функцию взаимной информации клеток внутри организма об аутологичности; экспрессия HLA II обнаруживается исключительно на клетках иммунной системы: B-лимфоцитах, макрофагах, эндотелиоцитах, активированных Т-клетках и др.
Молекула HLA I (рис. 7-2) состоит из трехдоменной α-цепи и β2-микроглобулина, связь между которыми стабилизируется калнексином. Между доменами α1 и α2 имеется желобок, предназначенный для загрузки антигена. После того как антиген загружается, происходит высвобождение β2-микроглобулина, возможно, за счет конформационных изменений. Имеется около 180 генов с более чем 500 аллелями (на хромосоме 6), кодирующих синтез молекул HLA I, что обеспечивает их видоспецифический и индивидуальный полиморфизм. Молекула HLA II состоит из двух димеров - α и β. Вариабельные домены α1 и β1 формируют желобок для загрузки антигена. В целом β-цепи HLA-DR, DP, DQ характеризуются меньшей гетерогенностью; известно более 300 аллельных продуктов HLA II (более 20 генов, кодирующих HLA II, также находятся на хромосоме 6). Для контроля укладки при синтезе, протекции, транспорте и загрузке антигенных пептидов на HLA служат особые молекулы - шапероны (калнексин, кальретикулин, инвариантная цепь Ii и др).
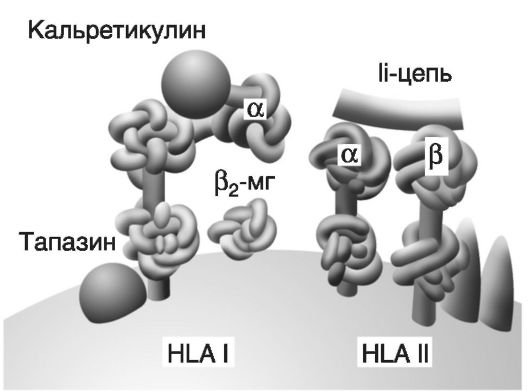 Рис. 7-2. Структура белков гистосовместимости (HLA I и HLA II) и шапероны. Молекула HLA I состоит из α-цепи (три домена) и β2-микроглобулина ф2-МГ).
Молекула HLA II содержит две цепи: α (два домена) и β (тоже два
домена). Шапероны (кальретикулин, тапазин, Ii-цепь и др.) - это белки,
ответственные за правильную укладку белковых молекул; в частности,
шапероны обеспечивают протекцию «желобков» молекул HLA до момента
загрузки на них антигенных пептидов
Рис. 7-2. Структура белков гистосовместимости (HLA I и HLA II) и шапероны. Молекула HLA I состоит из α-цепи (три домена) и β2-микроглобулина ф2-МГ).
Молекула HLA II содержит две цепи: α (два домена) и β (тоже два
домена). Шапероны (кальретикулин, тапазин, Ii-цепь и др.) - это белки,
ответственные за правильную укладку белковых молекул; в частности,
шапероны обеспечивают протекцию «желобков» молекул HLA до момента
загрузки на них антигенных пептидов
Не-HLA молекулы CD1 (a, b, c, d, e), которые состоят по аналогии с HLA I из α-цепи и β2-микроглобулина, также вовлечены в процессы загрузки антигенов, но небелковой природы (фосфолипиды, липополисахариды).
Имеется связь между наследованием некоторых генов HLA и высоким риском развития определенных болезней. Например, более 90% пациентов с анкилозирующим спондилоартритом, тяжелой аутоиммунной патологией позвоночника имеют ген HLA-B27.
Поскольку HLA определяет гистосовместимость, необходимо, чтобы у донора и реципиента при трансплантации органа или ткани было соответствие по HLA. Карта пациента по HLA называется «Jull house» («полный дом») и включает данные относительно двух аллелей каждого вида молекул (например, HLA-A, HLA-B, HLA-DR и др.). Сенсибилизация белками HLA в прошлом (при гемотрансфузиях, трансплантациях или беременности) может приводить к острому отторжению пересаживаемого трансплантата или тромбоцитопении при переливании крови, поэтому необходимо обязательное тестирование на наличие антител против HLA. Типирование HLA может служить дополнительным критерием для диагностики таких болезней, как анкилозирующий спондилоартрит, диабет, целиакия, гемохроматоз, псориаз и нарколепсия, при которых известна высокая степень связи с определенными гаплотипами HLA.
7.2. ИММУННЫЙ ОТВЕТ
Иммунный ответ - это процесс взаимодействия клеток иммунной системы, который индуцируется антигеном и приводит к образованию эффекторных клеток и молекул, уничтожающих данный антиген.
Иммунный ответ является всегда специфическим, но не изолированным процессом, который протекает только в периферических органах иммунной системы. Как правило, он сопровождается такими неспецифическими реакциями, как фагоцитоз, активация комплемента, NK-клеток и т.д.
7.2.1. Стадии иммунного ответа
В начальных стадиях иммунного ответа участвуют, по крайней мере, три вида клеток: макрофаг (или дендритная клетка), Т- и В-лимфоцит. В целом все клетки, вовлеченные в этот процесс,
могут быть разделены, как указывалось выше, на антигенпредставляющие, регуляторные, эффекторные и клетки памяти. Имеются 2 магистральных пути иммунного ответа:
1. Клеточный иммунный ответ (T-клеточный).
2. Гуморальный иммунный ответ (В-клеточный).
Первый из них регулируется Т-хелперами типа 1 (Th1) и приводит к формированию эффекторных CD4+ T-клеток воспаления и цитотоксических CD8+ T-лимфоцитов, а также соответствующих им Т-клеток памяти.
Второй путь регулируется Т-хелперами типа 2 (Th2) и заканчивается образованием плазматических клеток (продуцентов антител) и В-лимфоцитов памяти. Переключение на синтез некоторых изотипов антител частично контролируется Th1. За исключением скрытого индуктивного периода иммунный ответ в среднем продолжается около трех недель с максимальным напряжением на 1-й неделе.
Можно выделить несколько основных стадий иммунного ответа:
1. Эндоцитоз антигена, его процессинг (обработка) и загрузка на молекулы HLA I или HLA II для презентации лимфоцитам.
2. Распознавание комплекса антигенный пептид/HLA I или антигенный пептид/HLA II и других стимулов.
3. Сигнальная трансдукция и активация лимфоцитов.
4. Клональная экспансия (пролиферация) лимфоцитов.
5. Созревание эффекторных лимфоцитов и клеток памяти.
6. Эффекторная активность (деструкция антигена).
Первая стадия иммунного ответа (эндоцитоз, процессинг и презентация антигена). Антигенпредставляющая клетка (макрофаг, дендритная клетка или В-лимфоцит) сталкивается с нативным антигеном и поглощает его. Макрофаги фагоцитируют главным образом патогены, для которых характерно внутриклеточное паразитирование (вирусы, бактерии, грибы, простейшие и др.); дендритные клетки пиноцитируют вирусы; В-клетки интернализируют различные токсины.
Следующее событие, процессинг, представляет собой ферментативный катализ макромолекулы антигена внутри антигенпредставляющей клетки. В результате процессинга происходит высвобождение доминирующей антигенной детерминанты (иммуноактивного пептида или суперантигена), который загружается на желобки собственных молекул HLA I или HLA II и выводится на поверхность
клетки для представления лимфоцитам. В зависимости от происхождения антигена выделяют два пути процессинга.
Экзогенные антигены презентируются в комплексе с молекулами HLA II «наивным» CD4+ T-клеткам (путь, опосредуемый HLA II).
Огачала эти антигены поглощаются и фрагментируются с помощью протеолитических ферментов в эндосомах (лизосомах). В то же самое время молекулы HLA II, связанные с шаперонами (калнексином и инвариантной цепью Ii), синтезируются и собираются в эндоплазматическом ретикулуме. Ii-цепь необходима для защиты желобка молекулы HLA вплоть до того момента, пока не будет загружен антигенный пептид. Затем комплекс HLA II/Ii-цепь транспортируется через аппарат Гольджи в эндосомы, где Ii-цепь теряется, а роль по защите желобка начинают выполнять дополнительные молекулы HLA-DM и, вероятно, HLA-DO. Наконец, антигенный пептид загружается на желобок молекулы HLA II, и этот комплекс экспрессируется на поверхности клетки.
Эндогенные или внутриклеточно расположенные антигены микробного происхождения загружаются на молекулы HLA I (путь, опосредуемый HLA I) для представления «наивным» CD8+ T-клеткам. Сначала, в отличие от экзогенных антигенов, такие цитоплазматические антигены перемещаются в цитозоль, где они расщепляются в крупном протеолитическом комплексе - протеасоме. После этого антигенный пептид транспортируется через «туннель» молекул TAP-1/TAP-2 в эндоплазматический ретикулум. Одновременно здесь происходит сборка молекулы HLA I, желобок которой (по аналогии с Ii-цепью у HLA II) находится «под защитой» шаперонов (сначала калнексина, затем кальретикулина), а укладка всей молекулы HLA I в последующем стабилизируется дополнительными молекулами (тапазином и др.). После загрузки антигенного пептида на желобок HLA I этот комплекс транспортируется на поверхность клетки.
Небелковые антигены загружаются на не-HLA-антигенпредстав- ляющие молекулы CD1.
В целом макрофаги и В-клетки вовлечены, соответственно, в Т-клеточный или гуморальный иммунный ответ по пути, опосредованному HLA II, а дендритные клетки двух типов способны к перекрестной презентации. Дендритная клетка типа 1 осуществляет процессинг эндогенных антигенов по пути с HLA I для запуска Т-клеточного ответа, а дендритная клетка типа 2 процессирует экзогенные антигены по пути с HLA II и включает В-клеточный ответ.
Вторая стадия иммунного ответа (распознавание антигена) протекает в течение нескольких часов. Однако при нарушениях клеточной миграции и межклеточных взаимодействий она может быть более длительной. Возможно, это приводит к замедлению всего иммунного ответа на патоген. Клиническими проявлениями этой стадии являются повышение температуры тела, мышечная слабость, снижение аппетита и сонливость. По большей части они обусловлены системными эффектами цитокинов (см. ниже).
Для того чтобы мог стартовать специфический иммунный ответ на конкретный антиген, необходимо, чтобы Т- и В-лимфоциты соответствующего клона встретились с антигенпредставляющей клеткой. Некоторые антигены бактерий (T-независимые антигены) распознаются с помощью BCR B-клеток и не требуют помощи со стороны Т-хелперов. Большинство нативных антигенов (так называемых T-зависимых антигенов) полностью распознаются «наивными» CD4+ Th1 и CD8+ T-клетками (для включения Т-клеточного ответа или пути Th1), а также «нaивными» CD4+ Th2 (для включения гуморального ответа или пути Th2). Интересно, что для прайминга CD8+ Т-клеток необходимо участие CD4+ Th1.
Третья стадия (сигнальная трансдукция и активация лимфоцитов). Во время распознавания лимфоциты воспринимают три типа обязательных сигналов, один специфический и два неспецифических:
1. Aнтигенный пептид/HLA I или антигенный пептид/HLA II.
2. Цитокины.
3. Костимулирующие молекулы.
Антигенный пептид, загружаемый на HLA I или HLA II в результате процессинга, служит специфическим сигналом. Это одновременное «двойное» распознавание «своего» (белков HLA) и «несвоего» (антигена) было открыто нобелевскими лауреатами (1996) P.C. Doherty (Австралия, США) и R.M. Zinkernagel (Швейцария) и оказалось довольно универсальным явлением. Секретируемые цитокины и экспрессируемые костимулирующие молекулы являются двумя обязательными неспецифическими сигналами. Более того, для обеспечения надежного физического контакта клеток необходимо также взаимодействие таких адгезивных молекул, как LFA-1, ICAM-1, ICAM-2, ICAM-3.
Цитокины играют одну из ключевых ролей в неспецифической регуляции иммунного ответа. Т- и В-лимфоциты получают цитокиновые сигналы от антигенпредставляющих клеток, NK-клеток,
тучных клеток и др. Обратный сигнал от лимфоцитов, например секретируемый IFN-γ, способствует реэкспрессии HLA I/HLA II на антигенпредставляющих клетках. Цитокины, действующие на ранних стадиях иммунного ответа, могут быть разделены на две группы в зависимости от его направления:
1. Th1-цитокины: IL-2, IL-12, IL-18, IFN-γ, TNF-α/β (для пути Т-хелперов типа 1).
2. Th2-цитокины: IL-4, IL-13 (для пути Т-хелперов типа 2). Однако на следующих стадиях иммунного ответа (клональная
экспансия, созревание эффекторов, переключение синтеза изотипов антител) в процесс вовлекаются другие цитокины.
Костимулирующие молекулы также играют важную роль в неспецифической регуляции иммунного ответа (табл. 7-2).
Таблица 7-2. Костимулирующие молекулы при иммунном ответе
Клетка | Молекула | Функция |
Антигенпредставляющие клетки | B7.1, B7.2 | Лиганды для CD28, CTLA-4 |
Т-хелпер 1 | CD28 CTLA-4 (CD152) | Активационный сигнал Ингибирующий сигнал |
Т-хелпер 2 | CD28 CTLA-4 (CD152) | Ингибирующий сигнал Активационный сигнал |
В-лимфоцит | CD40 CD22 | Активационный сигнал на переключение синтеза различных классов антител Ингибирующий сигнал |
Активация клеток является результатом сигнальной трансдукции, которая осуществляется серией сложных внутриклеточных реакций. После распознавания первоначально происходит активация связанных с корецепторами и молекулами CD3 (на Т-клетках) или CD79 (на В-клетках) тирозинкиназ нескольких семейств (Lck, Fyn, Blk, Btk, Lyn, Zap70, Syk и др.), затем через посредничество адаптерных белков включаются сигнальные пути. Один из них связан с активацией фосфолипазы Cγ, образованием инозитолтрифосфата и диацилглицерола, активацией протеинкиназы С и мобилизацией внутриклеточного Ca2+, с транскрипцией гена IL-2. Данный цитокин является ключевым ростовым фактором для лимфоцитов в иммунном ответе. Второй сигнальный путь связан
с обменом арахидоновой кислоты и приводит к транскрипции генов структурных белков, необходимых для осуществления митозов клеток.
Четвертая (клональная экспансия лимфоцитов) и пятая (созревание эффекторных лимфоцитов и клеток памяти) стадии иммунного ответа. Клональная экспансия представляет собой пролиферацию активированных лимфоцитов, которая протекает в периферических органах иммунной системы. Пролиферирующие В-лимфоциты образуют вторичные фолликулы в лимфатических узлах (центробластная стадия), при этом размножение клеток регулируется рядом цитокинов: IL-2, IL-4, IL-5, IL-6, IL-10, IL-13, IL-14, IFN-γ, TNF и др. В последующем центробласты начинают превращаться в центроциты, которые мигрируют на периферию фолликулов (центроцитарная стадия). В этот момент клетки вступают в период соматических гипермутаций, который является своеобразным способом селекции нужной специфичности BCR. Происходит позитивный отбор клеток с высокоспецифичным BCR и негативный отбор В-лимфоцитов с низкоспецифичным рецептором. В процессе созревания В-клетки претерпевают морфологические изменения (плазмобласт (иммунобласт) - проплазмоцит (лимфоплазмоидная клетка) - плазмоцит) и мигрируют в костный мозг и MALT для синтеза антител различных классов. Синтез ранних антител (IgM) регистрируется уже к концу первых суток клинического инфекционного эпизода, а высокоспецифических IgG - на 5-7 сутки.
Клональная экспансия и созревание Т-клеток протекают в паракортикальных зонах лимфатических узлов и периартериолярных пространствах селезенки. Распознавшие антиген клетки вступают в пролиферацию и превращаются в лимфобласты. Клоны CD8+ T-клеток нарастают быстро, а клоны CD4+ Т-лимфоцитов - более медленно. В целом, клональная экспансия и дифференцировка регулируются разными цитокинами (IL-2, IL-7, IL-9, IL-12, IL-15, IFN-γ, TNF и др.) и адгезивными молекулами. В процессе дифференцировки значительно изменяется фенотип Т-лимфоцитов, но, в отличие от В-лимфоцитов, они не меняются морфологически.
Характерным клиническим эквивалентом стадий клональной экспансии и дифференцировки является увеличение периферических лимфатических узлов, миндалин, видимых лимфатических фолликулов и селезенки. Можно наблюдать эти симптомы при респираторной, урогенитальной или значительной системной инфекциях.
В процессе иммунного ответа наряду с эффекторными клетками формируются Т- и В-клетки памяти. В отличие от эффекторных лимфоцитов с небольшими сроками жизни, клетки памяти остаются жизнеспособными в течение длительного времени (пожизненно). Существуют CD4+ и CD8+ T-клетки памяти, В-клетки памяти и долгоживущие плазматические клетки. В отличие от наивных Т-лимфоцитов Т-клетки памяти характеризуются фенотипом CD45RO+, CD44hi, быстрым HLA-независимым циклом и способностью секретировать большие количества цитокинов. Долгоживущие плазматические клетки обеспечивают дополнительный механизм поддержания синтеза иммуноглобулинов без дополнительной антигенной стимуляции в течение 1,5 лет.
Вторичный иммунный ответ протекает в ускоренном режиме за счет клеток памяти (рис. 7-3). Появление IgM в сыворотке крови часто указывает на «свежую» инфекцию или реактивацию персистирующего патогена, а синтез IgG соответствует наличию иммунной памяти к однажды перенесенной инфекции. При таком ускоренном синтезе IgG клинические проявления инфекционной болезни обычно отсутствуют.
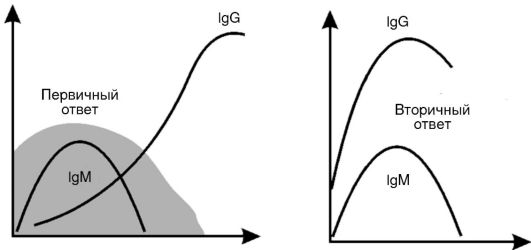 Рис. 7-3. Первичный
и вторичный гуморальный ответ. В первом случае нарастание IgG отстает
от нарастания IgM, при этом, поскольку IgM является низкоспецифичным по
отношению к патогену, отмечаются все симптомы инфекционного заболевания
(выделено серым). При вторичном ответе патоген связывается сразу
высокоспецифичными антителами класса IgG, поэтому клинических проявлений
болезни нет. По горизонтали - время (сутки); по вертикали - содержание
иммуноглобулинов (г/л)
Рис. 7-3. Первичный
и вторичный гуморальный ответ. В первом случае нарастание IgG отстает
от нарастания IgM, при этом, поскольку IgM является низкоспецифичным по
отношению к патогену, отмечаются все симптомы инфекционного заболевания
(выделено серым). При вторичном ответе патоген связывается сразу
высокоспецифичными антителами класса IgG, поэтому клинических проявлений
болезни нет. По горизонтали - время (сутки); по вертикали - содержание
иммуноглобулинов (г/л)
Шестая стадия иммунного ответа (эффекторная активность). Конечной стадией как гуморального, так и клеточного иммунного ответа является деструкция антигена, которая осуществляется с привлечением неспецифических факторов врожденного иммунитета. Известны следующие эффекторные механизмы деструкции антигена.
При иммунном ответе гуморального типа:
1. Простая нейтрализация антигена антителами при образовании иммунных комплексов «антиген + антитело» (АГ+АТ).
2. Комплементзависимый лизис антигена, связанного с антителом. Иммунные комплексы АГ+АТ, фиксированные на поверхности клеток-мишеней, присоединяют и активируют комплемент по классическому пути.
3. Фагоцитоз растворимых иммунных комплексов АГ+АТ с последующим их расщеплением в лизосомах фагоцитов.
4. Антителозависимая клеточная цитотоксичность (АЗКЦ). Реализуется путем разрушения клетками-киллерами (К-клетками) покрытых антителами (IgG) клеток-мишеней через присоединение к Fc-фрагменту IgG. Такими К-киллерами могут быть гранулоциты, макрофаги, тромбоциты, NK-клетки (натуральные киллеры).
При иммунном ответе клеточного типа:
1. Цитолиз и апоптоз клеток-мишеней. Цитотоксические Т-лимфоциты лизируют клетки-мишени с помощью белков - перфоринов. Перфорины - мономерные белки, способные встраивать в клеточную мембрану клетки-мишени и при полимеризации в присутствии Са2+ образовывать в ней каналы (поры), повышая тем самым ее проницаемость для Na+ и воды. В результате клетка-мишень набухает, происходит разрыв ее мембраны и гибель (осмотический лизис). Наряду с этим через поры, образованные перфоринами, в клетку-мишень поступают секретируемые цитотоксическими лимфоцитами TNF-β (лимфотоксин) и гранзимы (сериновые протеазы), запускающие механизм естественной клеточной гибели (перфорин-гранзимовый путь апоптоза). При этом сами цитотоксические Т-лимфоциты за счет синтеза специфических эндогенных ингибиторов сериновых протеаз нечувствительны к воздействию гранзимов. Реализация цитотоксичности Т-лимфоцитов может быть связана также с синтезом IFN-γ (ингибирует репликацию вирусов, активирует экспрессию HLA I/II и процесс распознавания вирусов и вирусинфицированных клеток Т-лимфоцитами) и индукцией рецепторзависимого апоптоза. Его развитие обусловливается лиганд-
рецепторным взаимодействием между Fas-рецептором (CD95), экспрессируемым клеткой-мишенью, и Fas-лигандом (Fas-L) Т-киллера либо посредством секреции Т-киллером TNF-α, активирующего TNF-R-ассоциированный домен смерти (TRADD - TNF-R-associated death domain) при связывании со специфическим рецептором TNF-R1 на клетке-мишени. Кроме того, проапоптотическое действие цитотоксических лимфоцитов может быть опосредовано увеличением проницаемости митохондриальных мембран клеток-мишеней, снижением трансмембранного потенциала митохондрий и выходом в цитоплазму клеток различных апоптогенных факторов, например цитохрома С и апоптозиндуцирующего фактора (AIF - apoptosis induced factor), активирующих каспазы (цистеиновые протеазы). Процедура апоптоза включает фрагментацию ДНК, конденсацию хроматина, блеббинг (образование вздутий - пузырей) мембраны, сокращение клетки, ее дезорганизацию и упаковку в апоптотические тельца. На поверхности апоптотирующих клеток экспрессируются молекулы, распознаваемые фагоцитами (фосфосерин, тромбоспондин, десиалированные мембранные гликоконъюгаты). За счет этого апоптотические клетки и тельца подвергаются фагоцитозу и разрушению лизосомальными факторами фагоцитов.
2. CD4+ Т-лимфоциты, ответственные за гиперчувствительность замедленного типа, с помощью секретируемых цитокинов (прежде всего IFN-γ) инициируют миграцию макрофагов и нейтрофилов в очаг иммунного воспаления и их активацию в очаге. Активированные макрофаги и нейтрофилы разрушают клетки-мишени путем фагоцитоза.
7.2.2. Регуляция иммунного ответа
Иммунный ответ является регулируемым процессом. Регуляция имеет большое значение для достижения нужного уровня специфичности и иммунной памяти, включения именно тех эффекторных механизмов, которые бы наибольшим образом отвечали потребностям организма, а также для исключения нежелательных последствий гиперактивации иммунной системы (например, при чрезмерном иммунном воспалении, аллергии и аутоиммунных расстройствах).
Регуляция иммунных процессов осуществляется с помощью механизмов иммунного самоконтроля, за счет влияния печени, эндо-
кринной системы, ЦНС и в связи с генетическими механизмами контроля.
Система иммунной саморегуляции включает принцип отрицательной обратной связи, баланс Th1/Th2, цитокиновую регуляцию1, контроль со стороны костимулирующих молекул, идиотипантиидиотипическую сеть и др. Антиидиотипы - это антитела к антигенсвязывающим участкам иммуноглобулинов (к идиотипам), т.е. антитела против антител. Являясь поверхностными рецепторами лимфоцитов, антиидиотипы взаимодействуют со своими идиотипами, формируя иммунологические сети и подавляя иммунный ответ.
Одну из ключевых ролей в регуляции иммунного ответа играют регуляторные Т-лимфоциты, которые функционируют, имея разные цитокиновые профили (табл. 7-3).
Таблица 7-3. Регуляторные субпопуляции Т-лимфоцитов
Субпопуляция | Цитокиновый профиль | Основные функции |
Т-хелпер нулевой (Th0) | IL-2 | Активация пролиферации лимфоцитов (митоген) |
T-хелпер 1 типа (Th1) | IFN-γ, IL-2, TNF-β | Регуляция Т-клеточного и гуморального адаптивных ответов (переключение на синтез IgG2, IgG3, IgA), активация макрофагов |
T-хелпер 2 (Th2) | IL-4, IL-5, IL-6, IL-10, IL-13 | Регуляция гуморального адаптивного ответа, включая переключение на синтез антител IgE и IgG4 |
Т-хелпер 3 (Th3) | TGF-β | Ингибиция пролиферации лимфоцитов |
T-регулятор (T-reg) | IL-10, TGF-β | Естественная супрессия иммунного ответа, индукция Fas- зависимого апоптоза |
T-хелпер 17 (Th17) | IL-17 | Развитие аутоиммунных реакций |
1 Образуемая цитокинами система регуляции иммунного ответа - «цитокиновая сеть» - включает клетки-продуценты цитокинов, иммуноцитокины, клеткимишени, экспрессирующие цитокинсвязывающие рецепторы и антагонисты цитокинов и их рецепторов.
Печень является, главным образом, источником иммуносупрессивных факторов (α-фетопротеин, «pit» NK-клетки и др.). Гормоны оказывают дозозависимые разнонаправленные эффекты в отношении иммунных процессов. В физиологических концентрациях соматотропный и тиреотропный гормоны, тироксин, трийодтиронин, инсулин, пролактин стимулируют иммунный ответ, а адренокортикотропный гормон, глюкокортикоиды и половые гормоны - супрессируют. В последние годы интенсивно исследуется влияние на иммунную систему гормона эпифиза мелатонина, который, наряду с иммуностимулирующим действием, имеет также ритморегулирующий, снотворный, антиоксидантный и др. эффекты. ЦНС влияет на иммунный ответ через посредничество нейтротрансмиттерных (допамин-, серотонин-, ГАМК- и пептидергических) систем, проявляя стереотипную координацию иммунных процессов, связывая их с разными сферами психоэмоциональной деятельности мозга.
Генетическая регуляция иммунитета позволяет обеспечивать многообразие антигенраспознающих рецепторов за счет генных реаранжировок в хромосомах 2, 7, 14 и 22 не только в эмбриональном периоде, но и непосредственно в ходе иммунного ответа. Дополнительное разнообразие синтезируемых антител достигается также в связи с соматическими гипермутациями. Индивидуальная сила иммунного ответа регулируется индивидуальным набором HLA-генов (6-я хромосома), белковые продукты которых способны с разной эффективностью загружать различные антигенные пептиды.
Патология иммунного ответа может быть обусловлена гипофункцией (иммунодефициты) или гиперфункцией (реакции гиперчувствительности) иммунной системы.
7.3. ИММУНОДЕФИЦИТНЫЕ СОСТОЯНИЯ
Иммунодефициты - это состояния, обусловленные функциональной недостаточностью иммунной системы вследствие отсутствия либо снижения уровня одного или нескольких факторов иммунной системы.
Одним из наиболее частых проявлений иммунодефицита являются рекуррентные и/или оппортунистические инфекции. Вместе с тем инфекционный синдром в форме острых гипертоксических или повторных, рецидивирующих, затяжных и хронических инфекционных процессов является преобладающим, но не единственным
синдромом, характерным для иммунодефицитов. Другими проявлениями могут быть разнообразные аллергические реакции, аутоиммунные расстройства и даже пролиферативные процессы (в доброкачественном варианте - лимфаденопатии, гипертрофия миндалин, тимомегалия, в злокачественном - развитие лимфом, лимфосарком и др.).
Иммунодефициты подразделяются по происхождению на первичные и вторичные, по клинической выраженности - на манифестные и минорные, по локализации дефекта - на дефициты антител или Т-лимфоцитов, комбинированные дефициты Т- и В-клеток, дефекты NK-клеток, фагоцитоза, дефицитах адгезивных молекул, комплемента.
Диагностика иммунодефицитных состояний основывается на клинико-иммунологических исследованиях (общий анализ крови, определение уровня IgM, IgG, IgA, CD-типирование лимфоцитов, тесты на фагоцитоз, оценка пролиферативной способности Т- и В-клеток, продукции цитокинов, специфических антител и аутоантител, содержания белков комплемента, «острой фазы» в крови и т.д.).
Первичные иммунодефициты достаточно редки. Они обусловлены дефектами в отдельном гене или целых кластерах генов, ответственных за синтез белковых молекул иммунной системы. Из них примерно 50-75% приходится на нарушения образования иммуноглобулинов, 5-10% - на клеточные иммунодефициты, 10-25% - на комбинированные иммунодефициты и около 1-2% - на нарушения фагоцитоза и системы комплемента. Манифестные формы первичных иммунодефицитных состояний проявляются на первом или втором году жизни гнойничковыми поражениями кожи и слизистых оболочек, рецидивирующими инфекциями органов дыхания, мочевыводящих путей, кишечника, которые в ряде случаев (тяжелые формы) при отсутствии адекватной терапии могут приводить к летальному исходу. Однако если рассмотреть проблему более широко, с учетом слабо диагностируемых минорных форм, и принять во внимание неустойчивость равновесия популяционных наборов HLA и наиболее часто встречающихся патогенов в среде обитания данной популяции (от чего зависит сила иммунного ответа), то можно констатировать, что по большей части все иммунодефициты - первичные. Многие из первичных иммунодефицитов являются Х-связанными, т.е. наследуются преимущественно мальчиками от матерей.
Классическими примерами первичных иммунодефицитов являются:
1. Дефициты системы фагоцитов в основном касаются нарушений фагоцитарной функции нейтрофильных гранулоцитов. Выделяют следующие механизмы их формирования:
• нейтропения в связи с дефицитом, нарушением пролиферации и созревания гранулоцитарных клеток-предшественниц в костном мозгу (циклическая нейтропения, болезнь Костманна (или инфантильный агранулоцитоз), семейная доброкачественная нейтропения, ретикулярный дисгенез, синдром Швахмана-Даймонда-Оски, врожденный дискератоз и др.), вследствие нарушения высвобождения нейтрофилов из костного мозга в кровь, укорочения срока жизни качественно неполноценных (с цитогенетическими аномалиями) нейтрофилов (миелокахексия);
• нарушение хемотаксиса нейтрофильных гранулоцитов, обусловленное дисфункцией актина и неспособностью клеток образовывать псевдоподии (синдром «ленивых лейкоцитов»);
• дефицит молекул адгезии (от лат. adhaesio - прилипание) нейтрофилов вследствие дефекта гена β2-цепи интегринов (CD18) или утраты клетками способности синтезировать молекулы селектинов, что приводит, в частности, к нарушению взаимодействий сиалогликопротеина Sgp50 с CD62L (лиганд для L-селектина лейкоцитов) и сиалил-Lewisх-олигосахарида с CD62E (лиганд для Е-селектина эндотелиальных клеток). В результате нейтрофилы утрачивают способность к адгезии и связанным с ней функциям - миграции и эндоцитозу;
• утрата нейтрофилами способности к киллингу при включении в их цитоплазму аномальных гранул (синдром Чедиака-Хигаси, аномалия Альдера-Рейли), дефиците вторичных (специфических) цитоплазматических гранул или нарушениях продукции факторов респираторного взрыва - активных метаболитов кислорода (супероксидного анион-радикала, перекиси водорода и др.) вследствие недостаточности синтеза внутриклеточных ферментов (НАДФН-оксидазы, миелопероксидазы, глюкозо-6-фосфатдегидрогеназы), обусловленной аномалиями генов в Х, 1-й, 7-й или 16-й хромосомах. Примером такого рода иммунодефицита является хроническая гранулематозная болезнь, при которой в результате дефекта киллинговой функции нейтрофилов микробы (Staphylococcus, Serratia, Kleb-
siella, грибы Nocarida, Aspergillus и др.) формируют хронические инфекционные гранулемы в лимфатических узлах, печени и легких.
2. Дефициты системы комплемента описаны практически для всех компонентов комплемента человека (C1q, C1r, C1s, C2-C9, пропердина) и подразделяются на дефициты ранних компонентов классического пути активации комплемента и дефициты поздних компонентов комплемента. Как правило, дефекты системы комплемента являются аутосомно-рецессивными с локализацией в хромосомах 1, 5, 6, 9, 12, 19 (исключение - Х-сцепленный дефицит пропердина) и сопровождаются снижением противоинфекционной резистентности и развитием иммунокомплексных болезней в связи с нарушением элиминации циркулирующих иммунных комплексов АГ+АТ из организма (системная красная волчанка, гломерулонефрит и др.). Так, в основе развития системной красной волчанки лежит системное иммунокомплексное повреждение соединительной ткани с обязательным вовлечением в патологический процесс сосудов микроциркуляторного русла. Развитие иммунокомплексного гломерулонефрита связано с фиксацией и накапливанием иммунных комплексов на субэпителиальной части базальной мембраны капилляров почечных клубочков.
К данной группе нарушений относятся также дефициты ингибиторов комплемента (например, аутосомно-доминантный дефицит С1-ингибитора).
3. Дефициты антител. Диагноз дефицита гуморального звена иммунитета ставится на основании определения уменьшения в сыворотке содержания IgG, IgA, IgM, сопоставления лабораторных и клинических данных, включая анализ частых, особенно инфекционных заболеваний. Спектр иммунологических нарушений и клинических проявлений различных форм недостаточности антител варьирует в значительных пределах. Типичными клиниколабораторными признаками дефицита иммуноглобулинов являются гипоплазия и атрезия лимфоидных органов (миндалин, лимфатических узлов), снижение СОЭ вне связи с инфекцией.
К первичным дефектам гуморального звена иммунитета относятся агаммаглобулинемия с дефицитом В-клеток, общая вариабельная иммунная недостаточность, агаммаглобулинемия с гипер- IgM, редкие виды агаммаглобулинемии (формулы MgA, mgA), транзиторная гипогаммаглобулинемия младенческого возраста, селективный дефицит IgA.
Агаммаглобулинемия с дефицитом В-клеток составляет 61% всех случаев тотального дефицита антител. Основными иммунологическими характеристиками заболевания являются стойкое снижение концентрации IgG в сыворотке (ниже 200 мг/дл), отсутствие IgM, IgA, IgE, IgD и глубокий дефицит В-клеток (CD19-CD22, CD72) - менее 1% от общего числа циркулирующих лимфоцитов. Известно, что наиболее распространен Х-сцепленный тип заболевания (85%). Ген, локализованный на Xq21.3-22, продуктом которого является брутоновская тирозинкиназа, отвечает за развитие этого заболевания у мальчиков. Наряду с этим мутации в генах, кодирующих пре-В-клеточный рецептор, приводят к развитию заболевания и у девочек. Преобладающими в клинической картине являются повторные гнойные инфекции, манифестирующие у детей со второго полугодия жизни. При этом практически в 100% случаев имеют место бронхолегочные заболевания и гнойные инфекции ЛОР-органов. У большинства детей обнаруживается гипоплазия нёбных миндалин и периферических лимфатических узлов.
Общая вариабельная иммунная недостаточность является вторым по частоте встречаемости заболеванием среди тотального дефицита антител, иммунологические нарушения при котором характеризуются стойким снижением суммарной концентрации иммуноглобулинов в сыворотке крови (ниже 300 мг/дл), при этом уровень IgG составляет менее 250 мг/дл при нормальном или умеренно сниженном количестве В-клеток. У части больных В-клетки отсутствуют полностью. Отсутствие общего генетического дефекта, а также полиморфный характер иммунологических проявлений подтверждают гетерогенную природу этой болезни, манифестация которой возможна в любом возрасте у лиц как мужского, так и женского пола. Клиническая картина сходна с проявлениями агаммаглобулинемии. Часто развиваются агранулоцитоз и тромбоцитопения.
Агаммаглобулинемия с гипер-IgM включает аутосомно-рецессивный вариант, связанный с мутацией в гене AID и Х-сцепленную форму, при которой имеет место дефект гена, расположенного на Xq26,27, что приводит к аномалии CD40-лиганда. В 70% случаев заболевание наследуется по Х-сцепленному варианту, в остальных - по аутосомно-рецессивному типу. В сыворотке крови обнаруживается выраженное снижение концентрации IgG, IgA и IgE при резко повышенном уровне IgM, количество В-клеток (CD19- CD22) обычно нормальное или субнормальное. Как правило, ма-
нифестация заболевания приходится на ранний детский возраст. Клинические проявления сходны с другими глубокими дефектами антителопродукции. Часты аутоиммунные цитопении.
Транзиторная младенческая гипогаммаглобулинемия имеет значительно более мягкие клинические проявления с преобладанием в клинической картине «малых» инфекций (повторные ОРЗ, отиты, фурункулез и др.). Иммунологическая диагностика основывается на обнаружении у детей в возрасте от 1 до 5 лет снижения уровня одного или нескольких изотипов иммуноглобулинов: IgG (ниже 500 мг/дл), IgA (ниже 20 мг/дл) и IgM (ниже 40 мг/дл).
Селективный дефицит иммуноглобулина А характеризуется повторными инфекциями дыхательных путей и ЛОР-органов с благоприятной эволюцией. Отмечается повышенная частота развития аллергических и аутоиммунных заболеваний. Критерием для постановки диагноза является изолированное снижение концентрации IgA в сыворотке крови (до 5 мг/дл) при нормальных уровнях других изотипов иммуноглобулинов.
В отличие от синдромов тотального дефицита антител, имеющих ярко выраженную клиническую картину тяжелых повторных инфекций, транзиторная младенческая гипогаммаглобулинемия и селективный дефицит IgA могут не иметь клинических проявлений и выявляться при случайном лабораторном обследовании у асимптоматичных детей.
4. Т-дефициты проявляются количественной и функциональной недостаточностью Т-лимфоцитов. Факторами такого рода иммунодефицитов являются:
• аплазия или дисплазия тимуса на этапе эмбриогенеза (синдром Ди-Джорджи, синдром Незелофа);
• нарушение пролиферации Т-лимфоцитов и Т-клеточной ДНКрепарации, связанное с аномалией генов в хромосомах 11q22- 23 (синдром Луи-Бар или атаксия-телеангиэктазия) или 10р;
• дефицит внутриклеточных ферментов: аденозиндезаминазы (мутация гена в хромосоме 20q13-ter) и пуриннуклеозидфосфорилазы (мутация гена в хромосоме 14q13.1), при недостатке которых в Т-лимфоцитах накапливаются токсичные метаболиты метилирования (S-аденозилгомоцистеин) и пуринового обмена (dATP, dGTP), подавляющие пролиферацию и индуцирующие гибель (апоптоз) Т-клеток;
• дефицит синтеза цитокинов и цитокиновых рецепторов (так называемые цитокиновые дефициты), связанные с мутациями
гена в Х-хромосоме (Xq13), гена, кодирующего белок JAK (Янус-киназы) Т-лимфоцитов, передающий сигнал для γ-цепи рецепторов цитокинов (IL-2R, IL-4R, IL-7R, IL-9R, IL-15R, IL-21R) и гена IL-7Ra, локализованного в хромосоме 5р13;
• снижение экспрессии TCR - поверхностных антигенсвязывающих рецепторов на Т-лимфоцитах при мутациях генов RAG-1 и RAG-2 в хромосоме 11р13, кодирующих рекомбинацию генов V, D, J TCR;
• дефицит CD45 (тирозинспецифическая фосфатаза) вследствие дефекта гена 1q3/PTPRC, что приводит к нарушению связывания TCR с корецепторами CD4 или CD8 Т-лимфоцитов и снижению эффективности прохождения антигенного сигнала внутрь клетки.
5. Комплексные иммунодефициты. К данной группе иммунодефицитов относятся синдром тяжелой комбинированной иммунной недостаточности и синдром Вискотта-Олдрича.
Синдром тяжелой комбинированной иммунной недостаточности -
группа Х-сцепленных или аутосомно-рецессивных генетических дефектов, характеризующихся лимфоцитопенией, обусловленной глубоким дефицитом Т-лимфоцитов (резко снижено содержание клеток субпопуляций CD3, CD4, CD8), анергией Т-клеток in vitro и in vivo (иногда и NK-клеток) и отсутствием антительного ответа при нормальном (иногда) содержании иммуноглобулинов и В-клеток в крови. Клинические проявления начинаются с первых недель или месяцев жизни. В клинической картине выявляются остановка развития и роста, неукротимый понос, рвота, обширная молочница, нарастающая одышка, сухой кашель. Спектр микробов, приводящих больного к гибели, оппортунистический или малопатогенный: Candida albicans, Pneumocystis carinii, вирусы Varicella zoster, парагриппа, респираторно-синцитиальный, адено- и цитомегаловирус, вирус Эпштейна-Барр, БЦЖ.
Синдром Вискотта-Олдрича - Х-сцепленное заболевание (дефектный ген локализуется в Хр11.22), проявляющееся экземой, возвратными инфекциями (как правило, с нетипичным или резистентным к лечению течением), тромбоцитопенией, дефицитом и нарушением пролиферативного ответа Т-лимфоцитов, а также снижением уровня IgM при увеличении содержания и IgE в сыворотке крови.
Вторичные иммунодефициты обусловлены количественным и качественным голоданием (недостатком белков, витаминов, микро-
элементов Fe, Zn, Cu и др.), эндокринопатиями (сахарный диабет, болезнь Иценко-Кушинга), потерей иммунокомпетентных клеток и молекул при травмах, кровотечениях, операциях, ожоговой болезни, утрате органов иммунной системы, воздействием ионизирующей радиации, полихимиотерапии, гипертоксическими инфекциями и инфекциями, к которым иммунная система человека оказалась эволюционно не готовой (ВИЧ и др.).
Оиндром приобретенного иммунодефицита является классическим примером такого рода патологии. Возбудитель болезни, ВИЧ, имеет избирательное сродство одного из своих оболочечных белков (gp120) к молекуле CD4, которая экспрессируется Т-лимфоцитами-хелперами. При СПИДе происходит последовательное выключение Т-клеточного иммунитета, что приводит к активации оппортунистических инфекций и другим расстройствам со стороны иммунной системы, включая опухолевые заболевания (саркома Капоши).
Перспективы лечения иммунодефицитных состояний связаны с замещением недостающих компонентов (введение иммуноглобулинов, пересадка костного мозга и т.д.), назначением иммунотропных препаратов (при минорных формах), генной терапией.
7.4. РЕАКЦИИ ГИПЕРЧУВСТВИТЕЛЬНОСТИ
Иммунная гиперчувствительность - это патологическое состояние, характеризующееся повышенной реактивностью иммунной системы к антигенам. Реакции гиперчувствительности лежат в основе развития аллергии, аутоиммунных расстройств (см. главу 8) и отторжения трансплантата (см. раздел 7.5).
Классификация реакций гиперчувствительности в зависимости от характера иммунного повреждения была предложена P.G.H. Gell и P.R.A. Coombs в 1969 г. и позднее дополнена I.M. Roitt. Согласно этой классификации выделяют реакции гиперчувствительности I, II, III, IV и V типов (см. табл. 7-4, рис. 7-4).
I тип (реагиновый, анафилактический) связан с образованием особого типа антител (IgE, IgG4), имеющих высокое сродство (аффинность) к определенным клеткам (тучным, базофилам), - так называемые цитотропные антитела. Антиген, вступая во взаимодействие с фиксированными на клетках антителами, приводит к секреции предсуществующих и вновь образующихся биологически актив-
Таблица 7-4. Классификация реакций гиперчувствительности в зависимости от типа иммунного повреждения тканей и органов
Тип реакции (иммунного повреждения) | Аллергия | Аутоиммунные заболевания |
I (реагиновый, анафилактический) | Атопическая бронхиальная астма, аллергические риниты, конъюнктивиты, анафилактический шок, аллергическая крапивница, отек Квинке и др. | Не развиваются |
II (цитотоксический или цитолитический) | Лекарственная тромбоцитопеническая пурпура, аллергический лекарственный агранулоцитоз, гемотрансфузионные реакции, возникающие вследствие несовместимости групп крови | Миастения гравис |
III (иммунокомплексный) | Экзогенный аллергический альвеолит, сывороточная болезнь, феномен Артюса | Системная красная волчанка, системные васкулиты и др. |
IV (клеточноопосредованный) | Аллергия, формирующаяся при некоторых инфекционных заболеваниях (туберкулез, лепра, бруцеллез, сифилис), аллергический контактный дерматит, реакция отторжения трансплантата | Ревматоидный артрит, рассеянный склероз |
V (антирецепторный) | Не развивается | Иммунный тип сахарного диабета, аутоиммунные заболевания щитовидной железы, гипофиза и др. |
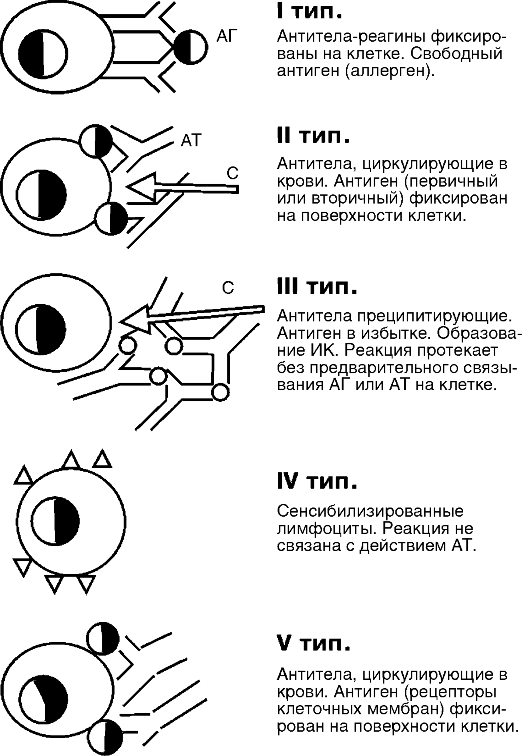 Рис. 7-4. Типы иммунного повреждения. ИК - иммунный комплекс
Рис. 7-4. Типы иммунного повреждения. ИК - иммунный комплекс
ных веществ (медиаторов), которые вызывают повышение проницаемости сосудов, отек ткани, гиперсекрецию слизи, сокращение гладкой мускулатуры. Типичным примером этого типа повреждения являются такие аллергические реакции, как атопическая бронхиальная астма, сезонные аллергические риниты, конъюнктивиты, анафилактический шок, анафилактическая крапивница, отек Квинке и др.
II тип (цитотоксический, или цитолитический) связан с образованием антител классов IgG (кроме IgG4) и IgM к компонентам клеточных мембран или веществам, сорбированным на клеточной поверхности. Образующийся на поверхности клеток комплекс АГ+АТ активирует систему комплемента, в результате чего возникают повреждение и лизис клеток. Примерами такого типа цитотоксического повреждения являются: аллергические реакции на некоторые лекарственные вещества - лекарственная тромбоцитопеническая пурпура, агранулоцитоз; гемотрансфузионные реакции, возникающие вследствие несовместимости групп крови; аутоиммунные заболевания - аутоиммунная гемолитическая анемия, тромбоцитопения, миастения гравис и др.
III тип иммунного повреждения (иммунокомплексный) связан с образованием токсических иммунных комплексов АГ+АТ (с антителами классов IgM, IgG1, IgG3). Примерами являются экзогенный аллергический альвеолит (при попадании ингаляционных антигенов), сывороточная болезнь, феномен Артюса; аутоиммунные заболевания (системная красная волчанка, системные васкулиты и др.).
IV тип иммунного повреждения - клеточно-опосредованный (ГЗТ). К этому типу принадлежат: аллергия, формирующаяся при некоторых инфекционных заболеваниях (туберкулез, лепра, бруцеллез, сифилис), аллергический контактный дерматит, реакция отторжения трансплантата и др.; аутоиммунные заболевания (ревматоидный артрит, рассеянный склероз).
V тип иммунного повреждения (антирецепторный) связан с наличием антител (главным образом IgG) к физиологически важным детерминантам клеточной мембраны - рецепторам (β-адренорецепторам, ацетилхолиновым и инсулиновым рецепторам, рецепторам для тиреотропного гормона). V тип иммунного повреждения играет особую роль при аутоиммунизации, является ведущим в развитии иммуноопосредованного сахарного диабета, иммунных заболеваний щитовидной железы, гипофиза и др. В развитии бронхиальной астмы, атопического дерматита и некоторых других антирецептор-
ный тип повреждения может быть одним из механизмов, осложняющих течение заболевания.
7.5. ОТТОРЖЕНИЕ ТРАНСПЛАНТАТА
Развитие медицины в ХХ веке привело к созданию принципиально новой технологии лечения - пересадке органов и тканей (см. подробнее в разделе 13.4). Трансплантации почек, костного мозга, печени, легких, сердца стали применяться, особенно в развитых странах мира, очень широко, что позволило продлить жизнь миллионам людей.
С другой стороны, пересадка чужеродной ткани стала биологическим явлением, неизвестным в эволюции человеческого организма. Трансплантация органов - ятрогенное действие, не имеющее естественных аналогов в природе. Ятрогенная трансплантация ни в коем случае не может быть сведена к беременности - физиологическому процессу, при котором полуаллогенный плод находится в особых условиях с наличием полупроницаемого «HLA-негативного» барьера (плаценты) и в зоне действия мощных иммуносупрессорных факторов (TGF-β, IL-4, IL-10, хорионический гонадотропин и др.).
Однако оказалось, что в целом механизмы отторжения трансплантата (graft rejection) соответствуют главным направлениям иммунного ответа на чужеродные антигены. Разные клоны наивных Т-лимфоцитов реципиента распознают: 1) HLA трансплантата, 2) антигенные пептиды трансплантата + HLA трансплантата, 3) антигенные пептиды трансплантата + собственные HLA. В последующем развиваются все варианты иммунного ответа, что приводит к формированию цитотоксических Т-лимфоцитов, Т-эффекторов иммунного воспаления, специфических иммуноглобулинов и соответствующих Т- и В-клеток памяти. Существенную роль в инициации иммунного ответа на антигены трансплантата имеют Т-лимфоциты, не отличающие «свои» HLA от «несвоих»1.
1 T-лимфоциты отбираются в процессе позитивной селекции в тимусе для распознавания любого антигена только в комплексе со «своими» молекулами HLA, в то время как на трансплантате экспрессированы «несвои» HLA. При этом 90-99% Т-лимфоцитов реципиента не замечают присутствия несущего «несвои» HLA трансплантата, в то время как 1-10% Т-клеток «ошибаются», принимая чужие HLA- молекулы за «свои» (перекрестная реактивность), что приводит к их активации и формированию иммунного ответа с последующим отторжением трансплантата.
Проблема эффективности пересадок органов и тканей связана прежде всего: 1) с эффективной иммуносупрессивной терапией; 2) с индукцией иммунологической толерантности.
Если пересаживаемая ткань содержит иммунокомпетентные аллогенные клетки, то при наличии у реципиента иммунодефицита может включиться реакция «трансплантат против хозяина». Такое явление наблюдается при пересадке костного мозга и при некоторых иммунодефицитах.
Американские ученые J.E. Murray и E.D. Thomas были удостоены Нобелевской премии (1990) за работы в области трансплантологии, которые стали важным подходом к лечению многих болезней человека.