Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 3. ФИЗИОЛОГИЯ КЛЕТКИ
Функции организма выполняют органы и системы органов, состоящие из тканевых элементов. Главный тканевый элемент - клетка. Другими словами, выполнение функций организма определяется функциями клеток. Любую функцию клетки осуществляют при помощи конкретных белков, информация о химической структуре которых записана в эндогенной клеточной программе - генах (сумма генов - клеточный геном).
В этой главе описаны функции отдельных органоидов клетки, а также функции клеток, осуществляемые при помощи сочетанной работы разных органелл, а именно: реализация генетической информации (от гена до белка) и её регуляция, внутриклеточная подвижность (молекулярные моторы, или хемомеханические преобразователи), деградация внутриклеточного и поглощённого клеткой материала (внутриклеточное пищеварение), клеточный цикл и его регуляция.
Органеллы
Клетка состоит из трёх основных компартментов: плазматической мембраны (её функции рассмотрены в гл. 2), ядра (включая ядерный геном) и цитоплазмы (цитозоль с находящимися в нём структури- рованными субклеточными единицами - органеллами). Цитозоль содержит органеллы, цитоскелет и включения. Органелла (органоид) - специализирующийся на выполнении конкретной функции и метаболически активный компонент цитоплазмы (рис. 3-1). К органеллам относят свободные рибосомы, гранулярную эндо- плазматическую сеть (шероховатый эндоплазматический ретикулум), гладкую эндоплазматическую сеть (гладкий эндоплазматический ретикулум), митохондрии, комплекс Гольджи, центриоли,
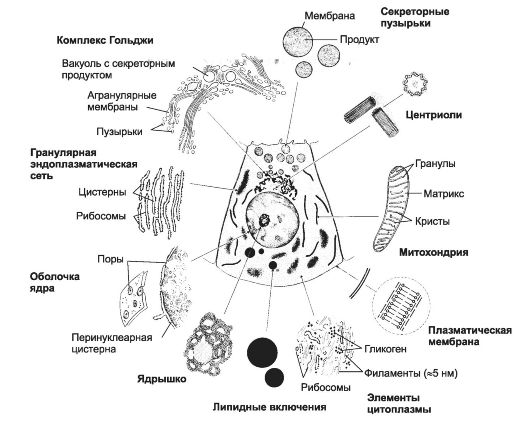
Рис. 3-1. Органеллы и включения.
окаймлённые пузырьки, лизосомы и пероксисомы. Относительное расположение компонентов внутри клетки не хаотично; каждый компонент занимает положение, оптимальное для выполнения его специализированной функции.
Ядро
23 пары хромосом (диплоидный набор: 22 x2 + XY у мужчин и 22 x 2 + 2X у женщин) ядра соматической клетки содержат ядерный геном. Реализация генетической информации (транскрипция и процессинг, рис. 3-2) и другие функции ядра происходят при участии ДНК и разных видов РНК.
• Хроматин. Термином «хроматин» обозначают комплекс ядерной ДНК с белками (гистоны, негистоновые белки).
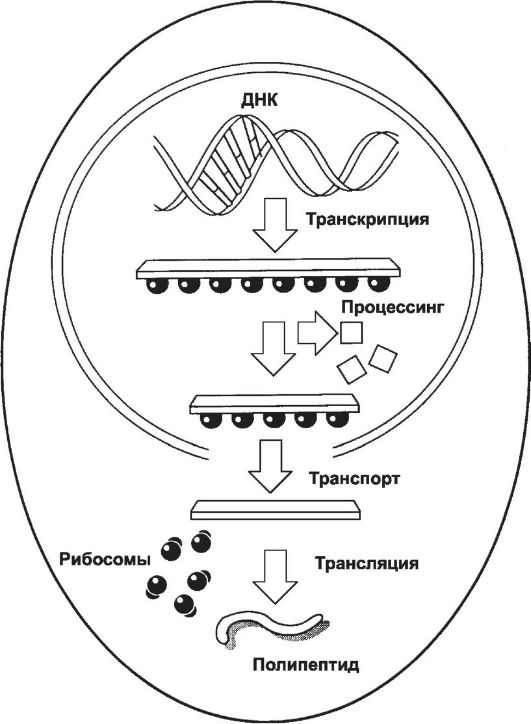
Рис. 3-2. Этапы реализации генетической информации [11, подробнее см. на рис. 3-13]. В ходе транскрипции на ДНК-матрице синтезируется комплементарная ДНК - длинная молекула РНК (первичный транскрипт), содержащая последовательности экзонов и интронов. По завершении синтеза РНК-транскрипта последовательности интронов удаляются (процессинг), что делает молекулу РНК значительно короче. Эта мРНК выходит из ядра в цитоплазму и соединяется с рибосомами. мРНК про- двигается сквозь рибосому, и её нуклеотидная последовательность транслируется в соответствующую последовательность аминокислот создаваемой полипептидной цепи.
• ДНК. Молекула ДНК построена из двух полинуклеотидных цепей, образованных из нуклеотидов.
• Ген - участок молекулы ДНК, кодирующий последовательность аминокислот в полипептидной цепи. Экспрессия гена (см. рис. 3-2) протекает по схеме: транскрипция (синтез первичного транскрипта на матрице ДНК) - процессинг (об-
разование мРНК) - трансляция (считывание информации с мРНК) - сборка полипептидной цепи (включение аминокислот в полипептидную цепь на рибосомах) - посттрансляционная модификация [добавление к полипептиду разных химических группировок, например фосфатных (фосфорилирование), карбоксильных (карбоксилирование) и т.д.].
• РНК - полинуклеотид, сходный по химическому составу с ДНК, но содержащий в нуклеотидах рибозу вместо дезоксирибозы и азотистое основание урацил (U) вместо тимина (T). Различают РНК матричную (мРНК), транспортную (тРНК) и рибосомную (рРНК). Катализирует синтез полимеров рРНК, мРНК и тРНК на матрице ДНК соответственно РНКполимераза I, II и III.
• Геном. Различают ядерный и митохондриальный геномы. Ядерный геном - полный комплект генов (примерно 60 000 на ядро диплоидной клетки, в гаплоидном наборе - около 24 000 генов) в хромосомах. Кариотип описывает количество и структуру хромосом. Гаплоидный набор - 23 хромосомы (3,5χ109 пар нуклеотидов) - характерен для гамет (зрелых половых клеток), а диплоидный набор - стандарт хромосом (23 χ 2) - для соматических клеток. Ядерный геном содержит гены, кодирующие полипептиды (примерно 2% генома), РНК (около 20% генома) и повторяющиеся последовательности (более 50% генома) неясного назначения.
• Ядрышко - компактная структура в ядре интерфазных клеток. Основные функции ядрышка - синтез рРНК (транскрипция и процессинг рРНК) и образование СЕ рибосом. Транскрипция рРНК происходит в хромосомах 13, 14, 15, 21 и 22.
• Нуклеоплазма содержит рибонуклеопротеины, ядерные рецепторы, ферменты (в том числе АТФазы, ГТФазы, ДНК- и РНК-полимеразы) и множество других молекул (онкогены, факторы транскрипции, белки теплового шока, вирусные ДНК, влияющие на транскрипцию и процессинг РНК).
Рибосомы
Рибосома состоит из большой и малой СЕ, содержащих различные типы рРНК и белки. Функция рибосом - трансляция (считы- вание кода мРНК и сборка полипептидов).
Гранулярная эндоплазматическая сеть
Гранулярная эндоплазматическая сеть (шероховатый эндоплазматический ретикулум) - система плоских мембранных цистерн с находящимися на их наружной поверхности рибосомами. Рибосомы связываются с мембранами сети при помощи рибофоринов. В шероховатой эндоплазматической сети происходит синтез белков для плазматической мембраны, лизосом, пероксисом, а также синтез белков на экспорт, т.е. предназначенных для экзоцитоза.
Гладкая эндоплазматическая сеть
Гладкий ретикулум - система анастомозирующих мембранных каналов, пузырьков и трубочек - не содержит рибофоринов и по этой причине не связан с рибосомами. Функции гладкой эндоплазматической сети многообразны: это синтез стероидных гормонов, детоксикация (например, фенобарбитала при помощи оксидаз в гепатоцитах), депонирование ионов кальция.
Митохондрии
Митохондрии чаще имеют форму цилиндра диаметром 0,2-1 мкм и длиной до 7 мкм (в среднем около 2 мкм). У митохондрий две мембраны - наружная и внутренняя; последняя образует кристы. Между наружной и внутренней мембранами находится межмембранное пространство. Внемембранный объём митохондрии - матрикс.
Митохондрии выполняют в клетке множество функций: окисление в цикле Кребса, транспорт электронов, хемиосмотическое сопряжение, фосфорилирование АДФ, сопряжение окисления и фосфорилирования, функцию контроля внутриклеточной концентрации кальция, синтез белков, образование тепла.
Комплекс Гольджи
Комплекс Гольджи образован стопкой из 3-10 уплощённых и слегка изогнутых цистерн с расширенными концами. Цистерны комплекса Гольджи образуют три основных компартмента: циссторону, транс-сторону, промежуточный компартмент. Комплекс Гольджи - основная органелла клетки, где осуществляется биохимическая модификация веществ, большую часть которой составляет прикрепление углеводных комплексов к белкам и липидам, их упаковка и сортировка для дальнейшего транспорта.
Лизосомы
Лизосомы - окружённые мембраной округлые пузырьки - образуются из эндосом путём включения в них лизосомных гидролаз и лизосомных мембранных белков после их синтеза в гранулярной эндоплазматической сети и процессинга в комплексе Гольджи. Известно более 50 лизосомных ферментов: рибонуклеазы, дезоксирибонуклеазы, катепсины, сульфатазы, β-глюкуронидазы, фосфолипазы, гликозидазы, липазы и др. Лизосомные ферменты наиболее активны в кислой среде (pH-5,0), для поддержания которой в мембрану лизосомы встроен протонный насос (H+-, K+- АТФаза).
Лизосомы участвуют во внутриклеточном пищеварении - в переваривании материала внутриклеточных компонентов (аутофагия) или частиц, различными путями попавших в клетку (гетерофагия). При нарушении функции лизосом развиваются так называемые лизосомные болезни накопления.
Пероксисомы
Пероксисомы - мембранные пузырьки размером 0,1-1,5 мкм с электронно-плотной сердцевиной. Они особенно многочисленны в клетках печени и почек. В составе мембраны органеллы находятся специфичные для пероксисом белки - пероксины, а в матриксе - матричные белки, в том числе более 40 ферментов (например, каталаза и пероксидаза), катализирующие анаболические (биосинтез желчных кислот) и катаболические (β-окисление длинных цепей жирных кислот, Н2О2-зависимое дыхание, деградация ксенобиотиков) процессы. Все компоненты пероксисом поступают из цитозоля. Продолжительность жизни пероксисом 5-6 сут. Новые органеллы возникают из предсуществующих путём их деления. Пероксисомы увеличиваются в размерах по мере поступления в них белков из цитозоля, и от них отделяются новые пероксисомы.
Цитозоль
Жидкая часть цитоплазмы (цитозоль) составляет около половины объёма клетки. Помимо воды, в цитозоле присутствуют ионы, множество химических соединений разной природы, макромолекулы. К макромолекулярным комплексам относятся апоптосомы - активаторы каспаз при регулируемой гибели клеток, а также протеосомы - комплексы нелизосомных протеаз, осуществляющие вместе с убиквитинами деградацию короткоживущих белков.
Включения
К разнородной группе включений относят находящиеся в цитозоле единичные или группы гранул и капель: гликогена, липидов, пигментов, липофусцина. Так, липиды содержатся в цитоплазме как капли самых различных размеров и плотности, они не связаны с клеточными мембранами, содержат триглицериды как источник энергии и холестерол, используемый для синтеза стероидов. В каком-то смысле включением, фактически занимающим весь объём эритроцита, можно назвать гемоглобин.
Цитоскелет
Трёхмерная цитоплазматическая сеть волокнистых и трубчатых структур различного типа формирует цитоскелет. К элементам цитоскелета относят микротрубочки, промежуточные филаменты, микрофиламенты (тонкие, или актиновые нити, а также толстые, или миозиновые нити). Цитоскелет придаёт клетке определённую форму и выполняет множество других функций (например, подвижность клетки, внутриклеточный транспорт).
Микротрубочки
• Микротрубочки состоят из 13 тубулиновых протофиламентов (нитей), идущих по спирали; нити имеют диаметр 24 нм и длину несколько микрометров. Каждая нить собрана из субъединиц (СЕ) - чередующихся димеров α- и β-тубулина. Микротрубочки участвуют в поддержании формы клетки, в транспорте макромолекул и органелл, обеспечивают расхождения хромосом при делении клеток, подвижность жгутиков и ресничек, движение сперматозоидов (см. ниже раздел «Молекулярные моторы»). Для микротрубочек характерна полярность, т.е. микротрубочки - динамичные структуры, постоянно растущие с одного конца (полимеризация) и деполимеризующиеся с другого конца. Микротрубочки входят в состав базального тельца, центриоли и аксонемы. Органеллы, содержащие микротрубочки Базальное тельце (рис. 3-3, I) состоит из девяти триплетов микротрубочек, расположенных в основании реснички или жгутика; служит матрицей при организации аксонемы. Центриоли имеют цилиндрическую форму, диаметр 150 нм и длину до 500 нм; их стенка состоит из девяти триплетов
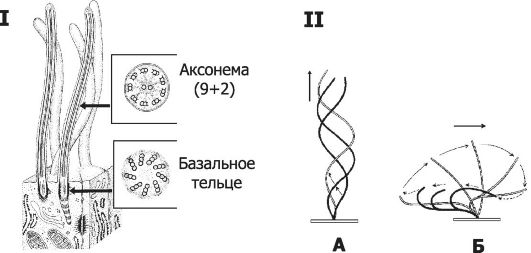
Рис. 3-3. I. Ресничка - тонкий вырост на поверхности клетки [11]. Стержень реснички образован аксонемой - системой микротрубочек 9+2. В основании реснички расположено базальное тельце, служащее матрицей для формирования аксонемы. II. Характер движения жгутика (А) и
реснички (Б). Несмотря на то что молекулярная основа подвижности у жгутиков и ресничек одинакова, характер их движения различен. Жгутики обычно длиннее ресничек, и для них характерно синусоидальное движение, в отличие от циклических волнообразных изгибов ресничек.
микротрубочек. Центриоль - центр организации митотического веретена - участвует в делении клетки. Аксонема (см. рис. 3-3, I) состоит из девяти периферических пар микротрубочек и двух расположенных центрально одиночных микротрубочек. В состав ручек периферических пар входит белок динеин, содержащий 2-3 глобулярные головки, соединённые с гибкой фибриллярной частью молекулы. Аксонема - основной структурный элемент реснички и жгутика.
• Ресничка - вырост клетки длиной 5-10 мкм и шириной 0,2 мкм, содержащий аксонему (см. рис. 3-3, I). Реснички имеются в эпителиальных клетках воздухопроводящих и половых путей, перемещают слизь с инородными частицами и остатками отмерших клеток и создают ток жидкости около клеточной поверхности.
• Жгутик в сперматозоиде имеет длину 50-55 мкм и толщину 0,2-0,5 мкм и содержит аксонему.
Промежуточные нити
Промежуточные нити (филаменты) состоят из белков, специфичных для определённых клеточных типов; имеют диаметр 8-11 нм.
Различают цитокератины эпителия, десмины мышц, виментины мезенхимы, ламины ядерной оболочки, а также нейрофиламенты, глиофиламенты и чётковидные волокна хрусталика. Промежуточные нити создают внутриклеточный каркас, обеспечивают упругость клетки, поддерживают упорядоченность расположения компонентов цитоплазмы. Иммуноцитохимические реакции с АТ конкретных типов промежуточных нитей нашли применение в цитодиагностике генеза опухолей.
Микрофиламенты
Две переплетённые нити фибриллярного актина (F-актин, рис. 3-4, А), составленные из глобулярного актина (G-актин), фор- мируют микрофиламенты (тонкие нити) диаметром 6 нм. Актиновые микрофиламенты образуют скопления по периферии клетки и связаны с плазмолеммой посредством белков (α-актинин, винкулин, талин). Эти тонкие нити могут также взаимодействовать с миозином толстых нитей, образуя актомиозиновый хемомеханический преобразователь (см. ниже раздел «Молекулярные моторы»).
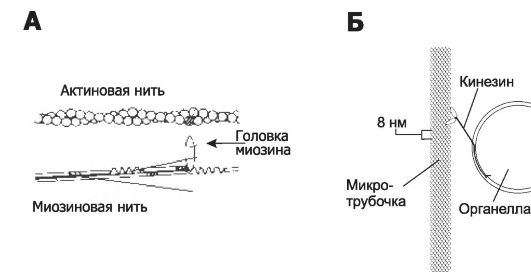
Рис. 3-4. Хемомеханические преобразователи [11]. А. В актомиозиновом молекулярном моторе происходит расщепление АТФ при взаимодействии актина тонких нитей (тонкие филаменты) с головкой миозина, отходящей от миозиновой (толстой) нити. Освобождённая энергия используется для взаимного перемещения относительно друг друга актиновых и миозиновых нитей. Б. Кинезин обеспечивает транспорт органелл из одной части клетки в другую вдоль микротрубочек. Шаг перемещения кинезина по поверхности микротрубочки составляет 8 нм.
• Полярность микрофиламентов. Как и микротрубочки, микрофиламенты полярны; присоединение (полимеризация) СЕ G-актина происходит на (+)-конце. Известны токсины, которые связываются с актином и блокируют его полимеризацию, нарушая тем самым подвижность клеток, фагоцитоз и цитокинез. К таким токсинам относятся цитохалазины и фаллоидин.
• Функции микрофиламентов
Изменение консистенции цитозоля, переход золя в гель и обратно (например, для изменения вязкости примембранной цитоплазмы при образовании псевдоподии).
Подвижность немышечных клеток (например, нейтрофилов и макрофагов) связана с изменением формы клеточной поверхности вследствие регулируемой полимеризации актина.
Стабилизация локальных выпячиваний плазматической мембраны связана с пучками поперечно сшитых актиновых филаментов (например, в микроворсинках эпителиальных клеток кишки, где пучок параллельных микрофиламентов образует их сердцевину).
Молекулярные моторы
Многие внутриклеточные процессы связаны с перемещением различных объектов (например, органелл, макромолекул). Реа- лизацию таких процессов осуществляют так называемые молекулярные моторы, или хемомеханические преобразователи. При работе молекулярных моторов энергия, накопленная в макроэргических связях АТФ или ГТФ, расходуется на выполнение механической работы. В хемомеханических преобразователях, использующих энергию расщепления АТФ, происходит взаимодействие, как минимум, 2 белков; один из этих белков - АТФаза. К таким АТФазам относятся миозин, динеин и кинезин. Входящий в состав клатриновых пузырьков динамин расщепляет ГТФ.
• Актомиозиновый хемомеханический преобразователь расщепляет АТФ при взаимодействии актина и миозина (рис. 3-4, А). Актомиозиновый мотор осуществляет мышечное сокращение, а также различные перемещения в немышечных клетках.
• Тубулинкинезиновый молекулярный мотор. Кинезины и тубулины микротрубочек образуют молекулярный мотор (рис. 3-4, Б), который обеспечивает внутриклеточный транспорт орга-
нелл и перемещение хромосом вдоль микротрубочек в ходе клеточного деления. Перемещение органелл вдоль микротру- бочек с участием кинезинов осуществляется в направлении (+)-конца микротрубочек с шагом 3 нм при развиваемой силе 0,15 пН.
• Тубулиндинеиновый мотор входит в состав аксонемы ресничек, жгутиков и других органоидов (см. рис. 3-3) и функционирует за счёт расщепления АТФ динеином. При расщеплении АТФ глобулярная головка динеина скользит по поверхности микротрубочки по направлению к её (-)-концу. Этот механизм аналогичен скольжению элементов актомиозинового хемомеханического преобразователя в мышце.
• Динамин. Присоединение к динамину ГТФ инициирует формирование механохимической молекулярной пружины из плотно упакованных колец динамина с шагом в 11 нм вокруг шейки эндоцитозного пузырька (см. рис. 2-11). При гидролизе ГТФ расстояние между смежными кольцами увеличивается до 22 нм. Таким образом, ГТФаза динамина катализирует расщепление ГТФ и тем самым растяжение пружины с последующим отрывом окружённого клатрином эндоцитозного пузырька от плазмолеммы.
Динеинкинезиновый молекулярный мотор принимает участие в аксональном транспорте. Движение органелл от тела нервной клетки направляется кинезином, а движение от нервного окончания к телу клетки направляется динеином.
Депо кальция
Цистерны гладкой эндоплазматической сети многих клеток специализированы для накопления в них Ca2+ путём постоянного откачивания Ca2+ из цитозоля, где нормальное содержание Ca2+ находится на уровне 10-7 М. Подобные депо кальция функционируют в любой клетке (гладкая эндоплазматическая сеть). Особенно велика роль кальциевых депо в скелетной и сердечной мышцах, нейронах, хромаффинных клетках, яйцеклетке, эндокринных клетках и т.д. Различные сигналы (например, гормоны, нейромедиаторы, факторы роста) влияют на функции клеток и их пролиферацию, измененяя концентрацию в цитозоле внутриклеточного посредника - Ca2+. Например, условием сокращения мышечных элементов является резкое повышение концентрации Ca2+ в цито-
золе. Для этого необходимо постоянно откачивать ионы кальция из цитозоля и накапливать их в специальных депо, образованных Са2+-запасающими цистернами гладкой эндоплазматической сети (в мышечных элементах их называют саркоплазматическим ретикулумом - СР). Внутри цистерн находятся Са2+-связывающие белки (кальсеквестрин, кальретикулин и др.) В мембрану цистерндепо Са2+ встроены Са2+-насосы (Са2+-АТФаза), постоянно закачивающие Ca2+ внутрь цистерн, и Са2+-каналы, через которые происходит выброс Ca2+ из депо при поступлении сигнала (см. подробнее в гл. 7 и на рис. 7-4).
Клеточный цикл
Клеточный цикл (рис. 3-5) состоит из митоза (М-фаза, см. рис. 3-12) и интерфазы. В интерфазе последовательно различают фазы G1, S и G2.
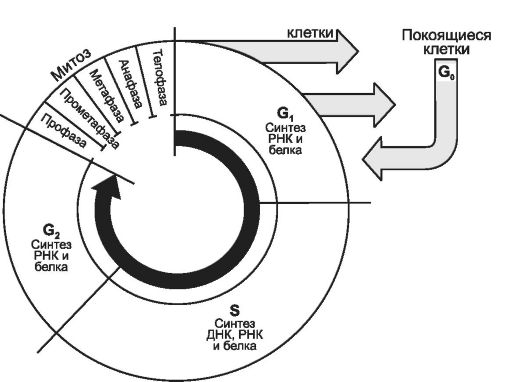
Рис. 3-5. Стадии клеточного цикла [11]. В клеточном цикле различают митоз - сравнительно короткую фазу M и более длительный период - интерфазу. Фаза M состоит из профазы, прометафазы, метафазы, анафазы и телофазы; интерфаза складывается из фаз G1, S и G2. Клетки, выходящие из цикла, более не делятся и обычно специализируются. Клетки в фазе G0 обычно не возвращаются в цикл.
Пролиферация клеток, происходящая путём митоза, жёстко регулируется множеством молекулярных сигналов. Регуляторы кле- точного цикла и митоза подразделяются на внутриклеточные и межклеточные.
• Внутриклеточные молекулярные сигналы многочисленны, среди них в первую очередь следует назвать собственно регуляторы клеточного цикла (циклины, циклинзависимые протеинкиназы, их активаторы и ингибиторы) и онкосупрессоры.
• Межклеточные сигналы. Регуляция клеточного цикла и пролиферация (или блок пролиферации) клеток регулируются внеклеточными сигналами в виде гормонов (например, гормон роста, эстрогены, ФСГ), цитокинов (например, ИЛ и интерфероны), факторов роста (например, фактор роста эпидермиса). Некоторые из таких молекулярных сигналов расцениваются как стимулирующие митогенную активность факторы (митогены). Существенное влияние на пролиферативную активность клеток оказывают и контакты с элементами межклеточного матрикса (например, с ламинином и фибронектином). После взаимодействия с соответствующими рецепторами такие митогенные сигналы (в значительном числе случаев при помощи связанных с G-белками протеинкиназ) передаются на соответствующие участки генома, активируя транскрипцию контролирующих фазы клеточного цикла генов (например, циклинзависимых протеинкиназ). Взаимодействие лигандов с рецепторами и дальнейший внутриклеточный перенос информационного сигнала рассмотрены в главе 4 («Межклеточные взаимодействия»).
Экспрессия генов
В этом разделе рассмотрена экспрессия генов и её регуляция. Экспрессия генов происходит в несколько этапов: транскрипция - процессинг - трансляция - посттрансляционная модификация. На каждом из этих этапов на макромолекулы в виде гена, первичного транскрипта, мРНК, полипептида и белка воздействуют различные регуляторные факторы.
Транскрипция
Транскрипция - первый этап реализации генетической информации - приводит к появлению на матрице ДНК первичного
транскрипта (РНК), состоящего из линейной последовательности транскрибированных интронов и экзонов. ДНК служит только матрицей для синтеза РНК и в ходе транскрипции не изменяется. В основе механизма транскрипции (полимеразная реакция, катализируемая РНК-полимеразами) находится принцип комплементарного спаривания оснований. Синтез РНК начинается в определённых последовательностях (сайтах) ДНК - промоторах и завершается в сайтах терминации.
Процессинг
Первичные транскрипты подвергаются ряду ковалентных модификаций (модификации концов пре-мРНК и сплайсинг), в ре- зультате осуществления которых формируется способная к трансляции зрелая мРНК.
Трансляция
Для синтеза полипептидной цепи необходимы следующие основные компоненты: аминокислоты, тРНК (около 50 разных тРНК), мРНК, рибосомы, источники энергии (АТФ и ГТФ), белковые факторы инициации, элонгации и терминации трансляции.
Посттрансляционные изменения
Полипептидные цепи могут подвергаться структурным модификациям в виде удаления части цепи (частичный протеолиз), присоединения одного или нескольких низкомолекулярных лигандов (ковалентные модификации), приобретения белком естественной для него (нативной) конформации (фолдинг полипептида, формирование белка из отдельных полипептидов - СЕ белка).
Ингибиторы матричного биосинтеза
Большая группа веществ ингибирует матричный синтез ДНК (репликацию), РНК (транскрипцию), полипептида (трансляцию). К таким веществам относятся антибиотики, токсины, интерфероны, а также вирусы.
• Антибиотики применяются в качестве противоопухолевых (курсы хемотерапии) и антибактериальных ЛС.
• Вирусы. Генетический материал вирусов - ДНК или РНК. После заражения клеток синтез вирусных ДНК, РНК и белков происходит за счёт ферментов, субстратов и источников энергии клетки-хозяина, но прекращается собственный синтез.
• Токсины (α-аманитин, рицин, бактериальные).
• Интерфероны секретируются некоторыми клетками в ответ на заражение вирусами и препятствуют распространению вирусной инфекции, стимулируя синтез ферментов, способных разрушать мРНК вирусов, и прекращая синтез белков на рибосомах инфицированных вирусов, препятствуя тем самым экспрессии вирусных генов в заражённых клетках.
Регуляция экспрессии генов
• In vivo не более 1% ядерных генов доступно для транскрипции. Все потенциально доступные для экспрессии гены (следовательно, синтезируемые полипептиды) подразделяются на конститутивные (транскрибируются постоянно и во всех клетках, например, гены ферментов гликолиза, разных РНК, альбумина и некоторых других белков) и клеточно-специфические (например, гены α- и β-цепей глобина транскрибируются в эритроидных клетках-предшественницах). Эти последние, а также некоторые другие гены подвергаются адаптивной регуляции и являются объектами индуцибельных воздействий (положительного контроля) или репрессии (негативного контроля). Другими словами, транскрибируемые гены и экспрессируемые полипептиды подразделяются на конститутивные, индуцируемые и репрессируемые. Все остальные структурные гены in vivo репрессированы необратимо.
Φ Конститутивный синтез происходит в любой клетке всегда и в относительно постоянных количествах, независимо от влияний на клетку.
Φ Регулируемый синтез (индуцируемый и репрессируемый) контролируется влияниями на клетку и её ядерный геном.
• In vitro в принципе может транскрибироваться весь геном, что доказано клонированием организмов из соматических клеток.
• Регуляция экспрессии конкретных генов в конкретных клетках определяется множеством внутриклеточных и межклеточных факторов, оперирующих на уровне межклеточных взаимодействий, отдельных клеток, их хромосом и генов, а также на разных этапах экспрессии гена.
Генетическая изменчивость
Отдельные гены, хромосомы и геном в целом (как под влиянием внешних факторов, так и в результате сбоев различных меха-
низмов) постоянно претерпевают разнообразные изменения. Хотя существуют механизмы репарации (восстановления) ДНК, часть повреждений или ошибок сохраняется. Изменения в последовательности и числе нуклеотидов в ДНК, не исправленные ферментами репарации, - «мутации».
Различают мутации в соматических клетках (фенотипически проявляются только в мутировавшей клетке или её потомстве) и мутации в половых клетках. Только последние потенциально могут передаваться по наследству и проявляться в фенотипе потомства, в том числе и в виде наследственных заболеваний.
• Мутагены. Причинами мутаций могут быть различные факторы. Мутагены (равно и вызываемые ими мутации) классифицируют по происхождению (источнику) на эндогенные и экзогенные, а по природе на физические, химические и биологические.
• Частота мутаций. Средняя частота возникновения мутаций в структурных локусах оценена в пределах от 10-5 до 10-6 на одну гамету за каждое поколение. Весь геном содержит 3х109 пар
оснований. Другими словами, каждое последующее поколение приобретает
несколько десятков мутаций. В широко известном научному и медицинскому
сообществу и ежедневно пополняемом «Каталоге наследственных заболеваний
человека» (OMIM, Online Mendelian Inheritance in Man <http://
www.ncbi.nlm.nih.gov/Omim/>) по состоянию на август
• Типы генных мутаций
Φ По характеру изменений в составе гена различают делеции, дупликации, инверсии, вставки, транзиции, миссенс- и нонсенс-мутации.
Φ По последствиям генные мутации подразделяют на нейтральные, регуляторные и динамические, а также на мис- сенс- и нонсенс-мутации.
• Хромосомные мутации (аберрации) характеризуются изменением структуры отдельных хромосом. При них последовательность нуклеотидов в генах обычно не меняется, но изменение
количества или положения генов при аберрациях может привести к генетическому дисбалансу, что пагубно сказывается на нормальном развитии организма. Различают внутрихромосомные, межхромосомные и изохромосомные аберрации.
• Изменения генома. Геномные мутации характеризуются изменением количества хромосом. У человека известны полиплоидия (в том числе тетраплоидия и триплоидия) и анеуплоидия.
• Типы наследования. Для любого моногенного заболевания существенной характеристикой является тип наследования: аутосомно-доминантный, аутосомно-рецессивный, сцепленный с хромосомой X (доминантный и рецессивный), голандрический (сцепленный с хромосомой Y) и митохондриальный.
Обобщение главы
Все клетки человеческого организма обладают сходным основным набором структур, осуществляющих главные функции клетки. Эти структуры входят в состав трёх основных компартментов клетки: плазматической мембраны, ядра (включая ядерный геном) и цитоплазмы с органеллами.
Клеточное ядро вместе с ядрышком и ядерным комплексом является местом расположения хромосом и ДНК-направляемого синтеза РНК.
Все органеллы окружены одинаковым в основных чертах типом мембраны, белково-липидным бимолекулярным слоем.
Каждый тип органелл имеет специфические вариации в липидном и белковом составе, определяющие особенности их функций.
Доставка молекул к различным отделам внутри клетки и во внеклеточное пространство осуществляется потоком белков или по- током везикул.