Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 19.РЕПРОДУКТИВНАЯ СИСТЕМА
МУЖСКАЯ ПОЛОВАЯ СИСТЕМА
Репродуктивная функция мужчины складывается из трёх взаимосвязанных позиций. К ним относятся герминативная функция яичек - механизмы образования сперматозоидов (сперматогенез и спермиогенез), физиология полового акта и гормональная регуляция репродуктивной функции.
Половое созревание мальчиков
Нормальное половое созревание происходит при переходе от половой незрелости ребёнка к взрослому состоянию половой зрелости через промежуточную стадию пубертата. Половое созревание (пубертат) - процесс, в течение которого у индивидуума появляются физические и поведенческие атрибуты, позволяющие ему воспроизводить потомство. У мальчиков пубертат возникает в результате увеличивающейся секреции андрогенов из яичек, что происходит под действием гонадотропинов передней доли гипофиза. Одновременно в развитии признаков пубертата принимают участие стероидные гормоны коры надпочечника (адренархе, см. ниже). В Европе и России пубертат у мальчиков начинается в 9-14 лет именно с увеличения размеров яичек (рис. 19-1 и сопровождающая рисунок таблица), а также с появления лобкового оволосения (рис. 19-2 и сопровождающая рисунок таблица).
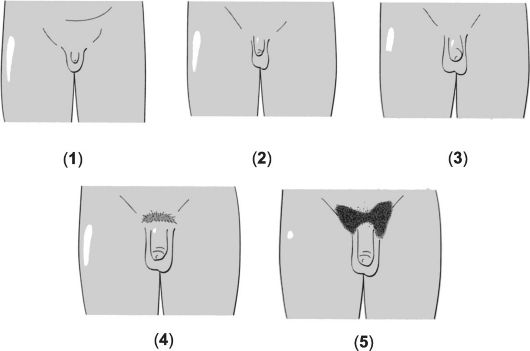
Рис. 19-1. Стадии развития наружных половых органов у мальчиков [18].
Пояснения см. в сопровождающей таблице.
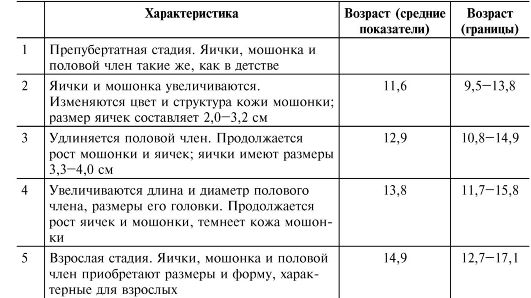
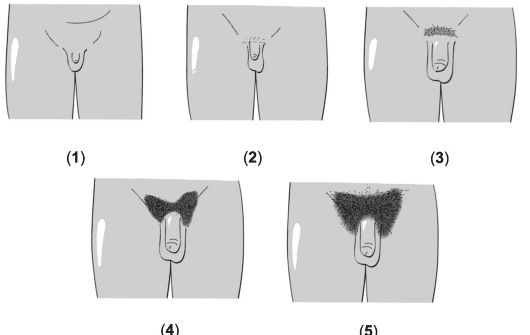
Рис. 19-2. Стадии формирования лобкового оволосения [18]. Цифра в скобках на рисунке указывает на соответствующую стадию. Пояснения см. в сопровождающей таблице.
Созревание яичек
Созревание яичек в пубертатном периоде включает инициацию секреции андрогенов клетками Ляйдига, рост извитых семенных канальцев и начало сперматогенеза. Все эти события находятся под контролем гонадотропинов (ФСГ и ЛГ).
Гонадотропины. У детей концентрации ФСГ и ЛГ как в гипофизе, так и в крови низкие. Амплитуда и частота пульсовой секреции обоих гормонов также низка, что обусловлено медленным и слабым высвобождением гонадолиберина из гипоталамуса. Примерно за год до того, как начинают увеличиваться яички, амплитуда и концентрация пульсовой секреции ФСГ и ЛГ начинает возрастать, особенно во время сна. Этот выраженный суточный ритм в секреции ФСГ и ЛГ - первое эндокринологическое проявление пубертата. Суточные колебания особенно выражены в ходе пубертата и практически исчезают к его окончанию.
Адренархе
Адренархе - отрезок времени, в течение которого надпочечники участвуют в половом созревании. Этот период характеризуется увеличением синтеза и секреции надпочечниками малоактивных андрогенов (андростендион, дегидроэпиандростерон и сульфат дегидроэпиандростерона). Хотя надпочечники образуют только 5% всего количества циркулирующих андрогенов у мальчиков, именно эти гормоны ответственны за начало подмышечного и лобко- вого оволосения. Надпочечниковые андрогены в периферических тканях превращаются в более активные гормоны, такие, как тестостерон и дигидротестостерон, которые и стимулируют лобковое и подмышечное оволосение, а также деятельность подмышечных сальных и потовых желёз. Подмышечное и лобковое оволосение начинается одновременно с увеличением размеров яичек, что делает внешне заметным наступление пубертата. Причина наступления адренархе не известна. Наиболее наглядное доказательство того, что это внутренний программируемый процесс, происходящий в самих надпочечниках, - его независимость от АКТГ.
Вторичные половые признаки
Тестостерон и его метаболиты вызывают характерные соматические изменения у мальчиков в пубертатном периоде. К таким изменениям относятся увеличение размера гортани и «ломка» голоса, увеличение массы костной ткани, массы и силы мышц, утолщение
кожи, увеличение количества и утолщение волос на теле, лобке, в подмышечных впадинах и на лице. Эти вторичные половые при- знаки прогрессивно появляются вслед за началом созревания яичек в последующие 2-2,5 года. Оволосение лица - последний по времени проявления вторичный половой признак - полностью развивается только к 20-25 годам.
Соматический рост
Соматический рост во время пубертата обусловлен совместным действием половых гормонов, СТГ и соматомедина C (инсулиноподобный фактор роста 1). Инсулин и тироксин также необходимы для роста. Отсутствие СТГ или соматомедина C (или рецепторов соматомединов) приводит к карликовости даже при достаточной концентрации половых гормонов в плазме.
Одновременно с изменением пульсовой частоты высвобождения ЛГ, что служит сигналом к началу пубертата, изменяется амплитуда секреции СТГ. Полагают, что эти изменения - результат стимуляции гипофиза эстрогенами (как у девочек, так и у мальчиков). У мальчиков повышение синтеза СТГ инициируется и поддерживается тестостероном, но этого не происходит при введении дигидротестостерона. Кроме того, секрецию СТГ в присутствии тестостерона тормозит тамоксифен - блокатор эстрогеновых рецепторов. Напротив, даже минимальные концентрации эстрогенов весьма стимулируют секрецию СТГ. Эти наблюдения позволяют предположить, что эффект тестостерона на рост костей не является первичным, а, возможно, обусловлен ароматизацией тестостерона в эстрадиол. Если же тестостерон влияет на мышцы, андрогены, напротив, действуют прямо и вызывают увеличение мышечной массы.
Рост костей начинается, когда тестостерон, ароматизированного до эстрадиола, повышает содержание СТГ, что индуцирует параллельное увеличение концентрации соматомедина C. Соматомедин C выступает в качестве мощного анаболического гормона, опосредующего многие метаболические функции СТГ, включая формирование кости. В нормальных условиях СТГ стимулирует синтез соматомедина C, а соматомедин C в свою очередь тормозит высвобождение СТГ по механизму отрицательной обратной связи. Во время пубертата, однако, содержание СТГ продолжает увеличиваться, несмотря на высокую концентрацию соматомедина C, что создаёт условия для быстрого роста. Вне периода пубертата сочетание повышенной
секреции СТГ и соматомедина C наблюдают только при акромегалии (непропорциональный и выраженный рост костей, преиму- щественно конечностей), когда секреция СТГ автономна, т.е. не зависит от различных регуляторных механизмов.
Скорость роста у мальчиков максимальна, когда концентрация тестостерона плазмы достигает 50% таковой взрослого мужчины. Этот рост продолжается до тех пор, пока в кости имеются эпифизарные хрящи. Половые стероиды (возможно, эстрогены) ответственны за закрытие зон роста в области эпифизов, что происходит у подростков в среднем до 20 лет.
Андрогены оказывают прямой анаболический эффект на мышечную массу, существенно возрастающую в пубертатный период.
ГЕРМИНАТИВНАЯ ФУНКЦИЯ
Яички выполняют две различные функции - герминативную (образование сперматозоидов - сперматогенез) и эндокринную (синтез андрогенов и ингибинов).
• Сперматогенез осуществляется в специальных структурах, называемых извитыми семенными канальцами, которые имеют сильно извитый ход и располагаются внутри долек яичка. Извитые канальцы, подходя к средостению яичка, превращаются в прямые канальцы, которые, в свою очередь, переходят в канальцы сети, располагающиеся непосредственно в средостении яичка. Прямые и извитые канальцы служат для выведения сперматозоидов, образующихся исключительно в сперматогенном эпителии извитых семенных канальцев.
• Андрогены синтезируются группами специализированных клеток, лежащих в интерстиции между извитыми семенными канальцами. Эти клетки называются клетками Ляйдига.
• Ингибины синтезируют так называемые поддерживающие клетки сперматогенного эпителия - клетки Сертоли.
Сперматогенный эпителий
Клетки, ответственные за сперматогенез, располагаются в канальцах, окружённых базальной мембраной. Выстилающий их эпителий состоит из развивающихся сперматозоидов и поддерживающих клеток. Этот эпителий называют сперматогенным (рис. 19-3, А). На поперечных срезах яичка видны сперматоциты на различных стадиях созревания.
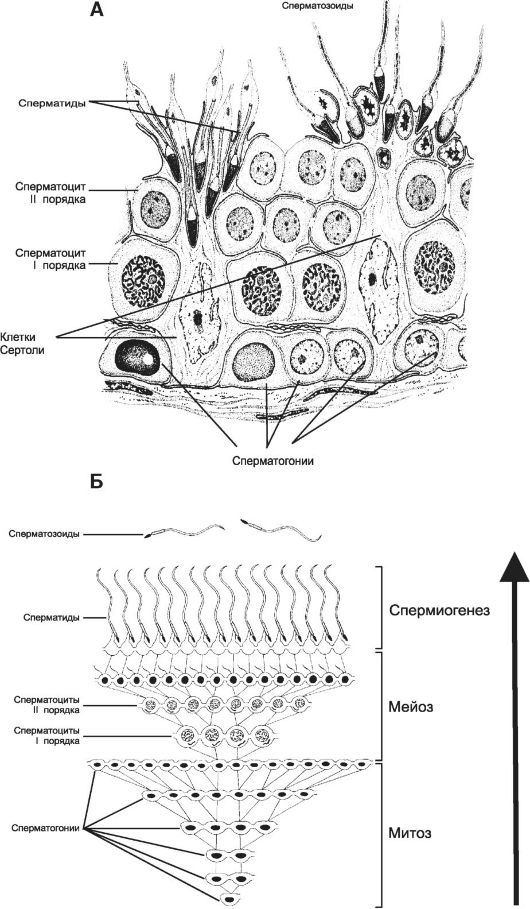
Рис. 19-3. Сперматогенный эпителий (А) [11]. Клон сперматогенных клеток (Б). На базальной мембране располагаются поддерживающие клетки Сертоли, а также сперматогонии. Выше лежат сперматоциты первого и второго порядка, над которыми находятся сперматиды на различных этапах развития и сперматозоиды (А). Практически во всех стадиях (размножение, мейоз, рост и созревание) сперматогенные клетки связаны друг с другом цитоплазматическими мостиками. Только в стадии формирования гамет цитоплазматические мостики разрываются и сперматозоиды оказываются свободными (Б).
Клетки Сертоли
Среди сперматогенных клеток располагаются клетки Сертоли - единственный вид несперматогенных клеток сперматогенного эпителия. Эти клетки, удачно названные «клетками-няньками», ответственны за метаболическую и структурную поддержку развивающихся сперматозоидов. Клетки Сертоли одной стороной (нижней) контактируют с базальной мембраной, а другой стороной (верхней) обращены к развивающимся сперматозоидам. Клетки Сертоли имеют большие и многочисленные пальцевидные выросты, которые могут одновременно контактировать со множеством клеток сперматогенного эпителия. В клетки Сертоли погружены развивающиеся сперматиды и сперматозоиды. Функции этих клеток - трофическая (обеспечение развивающихся гамет питательными веществами), фагоцитоз избыточной цитоплазмы сперматид и дегенерирующих половых клеток, ароматизация андрогенов (превращение тестостерона в эстрогены, что необходимо, чтобы на локальном уровне регулировать функции эндокринных клеток Ляйдига), секреция жидкости и связывающего андрогены белка (необходимы для транспорта сперматозоидов в семенных канальцах) и эндокринная (синтез ингибинов). Важная функция клеток Сертоли - создание гематотестикулярного барьера.
Гематотестикулярный барьер
Поддерживающие клетки Сертоли в период полового созревания образуют друг с другом плотные контакты, предупреждающие диффузию межклеточной жидкости между базальным и более поверхностно расположенным компартментами сперматогенного эпителия. Базальный компартмент содержит сперматогонии, а поверхностный - более зрелые сперматогенные клетки. Благодаря такому барьеру, в поверхностном компартменте сперматогенного эпителия создаётся специфическая гормональная среда с высоким уровнем тестостерона. Избирательная проницаемость барьера изолирует созревающие половые клетки от токсичных веществ и препятствует развитию аутоиммунного ответа против поверхностных Аг, экспрессирующихся на мембране сперматозоидов в ходе их со- зревания.
Сперматогенез
Сперматогенез (см. рис. 19-3) начинается с наступлением половой зрелости. Яички, в отличие от яичников, располагаются вне
полости тела (в мошонке). Это обстоятельство важно для нормального течения сперматогенеза, происходящего при температуре
Сперматогенез состоит из трёх последовательных фаз (рис. 19-3, Б): пролиферация сперматогоний (митозы), созревание спермато- цитов (два деления мейоза) и формирование сперматозоидов из сперматид (спермиогенез).
• Сперматогонии и пролиферация. Исходные сперматогенные клетки (сперматогонии), активизирующиеся к началу пубертата, практически постоянно делятся в базальном отделе извитого семенного канальца. Из этого резерва самообновляющиеся стволовые клетки преобразуются в сперматогонии. Каждая сперматогония претерпевает ограниченное количество митотических делений и формирует клон зародышевых клеток. После финального деления сперматогонии становятся сперматоцитами I порядка. Эта фаза находится под стимулирующим влиянием тестостерона и ФСГ. Сперматогонии - наиболее чувствительные к повреждению клетки яичка. Многие факторы (в том числе ионизирующее излучение, перегревание, приём алкоголя, голодание, местное воспаление и тяжёлые заболевания) могут вызвать их дегенеративные изменения.
• Мейоз. Сперматоциты I порядка проходят два мейотических деления, в результате которых получаются дочерние клетки с уменьшенным в два раза набором хромосом. Сперматоциты I порядка, проходят первое деление мейоза, в результате образуются сперматоциты II порядка, а после второго деления - сперматиды, имеющие гаплоидный набор хромосом.
• Фаза формирования (спермиогенез) находится в тесном контакте сперматид с клетками Сертоли. В сперматидах происходят существенные перестройки, в ходе которых формируются хвост, митохондриальная муфта и акросома (см. рис. 19-3, А). Спермиогенез осуществляется только при наличии эстрогенов и ФСГ. Сперматозоиды отделяются от сперматогенного эпителия под влиянием ЛГ.
Параметры сперматогенеза
• Количество клеточных делений сперматогонии постоянно: у человека происходит четыре митотических деления.
• В каждом небольшом участке извитого семенного канальца сперматогенез происходит циклично. В результате соседние участки канальцев имеют различную морфологию, а сперматозоиды на протяжении извитого семенного канальца продвигаются к поверхности сперматогенного эпителия в виде своеобразных волн.
• Время, необходимое для превращения сперматогонии в сперматозоид, готовый войти в придаток яичка, постоянно: у человека этот процесс занимает 64?4 дня. Окончательная дифференцировка сперматозоидов происходит в протоке придатка яичка в течение следующих 2 нед. Только в области хвоста придатка под влиянием тестостерона и эстрогенов сперматозоиды становятся зрелыми половыми клетками и приобретают способность к самостоятельному передвижению. Зрелые половые клетки хранятся в семявыносящем протоке (vas deferens).
Сперматозоиды и сперма
Жидкость, эякулируемая во время полового акта (эякулят), - сперма, она содержит сперматозоиды и секреторную жидкость до- бавочных желёз мужской половой системы (семенные пузырьки, простата и бульбоуретральные железы). В семенной жидкости на долю сперматозоидов приходится 5% объёма, 95% - на секреты добавочных желёз.
• Семенные пузырьки секретируют вязкий, желтоватого цвета секрет, поступающий в семявыбрасывающий проток во время эякуляции. Секрет семенных пузырьков разжижает семя, содержит фруктозу, соли аскорбиновой и лимонной кислоты, Пг - т.е. вещества, обеспечивающие сперматозоиды энергетическим запасом, повышающие их выживаемость и функциональную активность.
• Предстательная железа. Секрет железы принимает участие в разжижении семени и способствует его прохождению по мочеиспускательному каналу при эякуляции. Секрет железы содержит бикарбонат, липиды, протеолитические ферменты (фибринолизин), кислую фосфатазу. Слабощелочная реакция секрета (pH 7,5) нейтрализует кислотность других компонентов семенной жидкости и, таким образом, увеличивает подвижность и фертильность (оплодотворяющая способность) сперматозоидов.
• Бульбоуретральные железы Купера. Вязкий слизистый секрет, выделяемый в период полового возбуждения, служит для смазки уретры перед эякуляцией.
Эякулят
В конце полового акта сперма выбрасывается из полового члена во влагалище партнёрши. Количество эякулята при каждом сово- куплении составляет 3,5 (2-6) мл, каждый миллилитр содержит примерно 20-50 млн сперматозоидов. Для обеспечения фертильности (оплодотворяющей способности) в каждом миллилитре спермы должно быть не менее 20 млн сперматозоидов (в том чис- ле 60% нормальной морфологии и свыше 50% подвижных). После эякуляции максимальная продолжительность жизни сперматозоидов в половых путях женщины не превышает 48 ч. В то же время при температуре ниже -100 ?C сперматозоиды сохраняют фертильность годами.
Эндокринология мужской половой системы
Многообразные процессы в мужском организме (как непосредственно связанные с репродуктивной функцией, так и определяю- щие мужские соматический, психологический и поведенческий фенотипы) регулируются андрогенами (стероидные мужские половые гормоны), ингибинами, гипоталамическим люлиберином, гипофизарными гонадотропными гормонами (ЛГ и ФСГ), а также эстрадиолом и некоторыми другими биологически активными веществами. Все гормоны мужской половой сферы формируют регуляторный контур, упрощённая схема которого приведена на рис. 19-4.
Андрогены
Известно несколько стероидных гормонов (тестостерон, дигидротестостерон, дегидроэпиандростерон, андростендион и некоторые другие) с андрогенной активностью.
Тестостерон
Тестостерон - основной циркулирующий андроген, суточная секреция - 5 мг (от 2 до 10 мг), синтезируется в клетках Ляйдига. Гормон удаляется из крови в течение 30-60 мин, присоединяясь к клеткам-мишеням или распадаясь на неактивные компоненты.
• Стимулятор синтеза - ЛГ (стимулирующий интерстициальные клетки гормон).
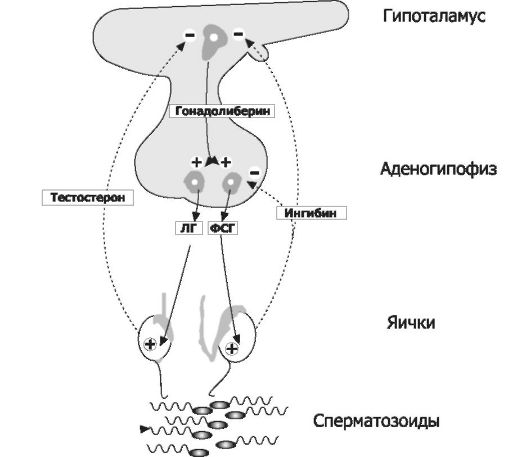
Рис. 19-4. Регуляторные взаимоотношения в системе «гипоталамус-гипофиз- яички» [7]. Непрерывные стрелки и символ «+» - активирующие влияния, прерывистые стрелки и символ «-» - ингибирующие влияния.
• Ароматизация тестостерона ведёт к образованию эстрадиола.
• Транспортные белки
Φ Андрогенсвязывающий белок (см. рис. 19-5) отвечает за поддержание высокого уровня тестостерона в сперматогенном эпителии, накапливая тестостерон в просвете семенных канальцев.
Φ β-Глобулин и альбумин связывают в крови до 99% тестостерона.
• Рецептор андрогенов (рецептор дигидротестостерона) относится к ядерным, содержит ДНК-связывающую область (см. подробнее в подразделе «Рецепторы», гл. 18). Тестостерон свободно проникает в цитоплазму клеток-мишеней. Под влиянием 5а-кеторедуктазы тестостерон превращается в дигидротестостерон, который связывается рецепторным цитоплазматическим белком. Этот комплекс мигрирует в клеточное ядро и активирует транскрипцию ряда генов. Известно множество мутаций, приводящих к полной или частичной нечувствительности мишеней к андрогенам.
• Функции
Φ Половая сфера. Тестостерон, как и остальные андрогены, весьма необходим для половой дифференцировки, полового созревания, поддержания вторичных половых признаков и сперматогенеза (см. ниже).
Φ Тестостерон - анаболический гормон. В этом качестве в разных органах (печень, скелетные мышцы, кости) тестостерон стимулирует синтез белка. В частности, под влиянием тестостерона возрастает мышечная масса, плотность и масса костной ткани. Вследствие стимуляции синтеза эритропоэтина повышается содержание Hb и гематокрит (Ht), а в связи с увеличением синтеза липазы печени в крови в ней уменьшается уровень липопротеинов высокой плотности и увеличивается содержание липопротеинов низкой плотности. Другими словами, тестостерон имеет выраженный атерогенный эффект, т.е. способствует развитию атеросклероза (в том числе венечных сосудов). Дигидротестостерон
Дигидротестостерон образуется в клетках Ляйдига (около 100 мкг/сут) и в ряде других органов (до 300 мкг/сут). Дигидро- тестостерон необходим для дифференцировки наружных половых органов (мошонка, половой член). Этот анаболический стероид запрещён законодательством ряда стран к применению (кроме диагностических целей).
Дегидроэпиандростерон и андростендион и ряд других стероидов обладают слабой андрогенной активностью.
Регуляция сперматогенеза
Гормональная регуляция сперматогенеза многообразна (см. рис. 19-4). Гипоталамо-гипофизарная система при помощи гона- долиберина активирует синтез и секрецию гонадотропных гормонов гипофиза, влияющих на активность клеток Ляйдига (синтез и секреция тестостерона) и Сертоли (синтез и секреция ингибинов). В свою очередь, вырабатываемые в яичке тестостерон и ингибины корректируют эндокринную деятельность гипоталамо- гипофизарной системы.
• Тестостерон. Сперматогенез в яичках поддерживается тестостероном - главным активатором этого процесса.
• ФСГ. Для полноценного осуществления сперматогенеза необходим также ФСГ. Мишенью ФСГ в извитых семенных канальцах
служат клетки Сертоли. ФСГ поступает в интерстиций яичка по мелким артериолам (рис. 19-5), затем диффундирует через базальную мембрану извитых канальцев и связывается со специфическими мембранными рецепторами на клетках Сертоли. Стимуляция рецепторов ФСГ приводит к синтезу внутриклеточных рецепторов андрогенов и андрогенсвязывающего белка (АСБ). Затем клетки Сертоли секретируют АСБ, и он связывает тестостерон, образуемый клетками Ляйдига и диффундирующий внутрь извитых семенных канальцев. АСБ переносит андрогены к сперматогенным клеткам, где они воздействуют на премейоти- ческие клетки (сперматоциты I порядка), имеющие андрогеновые рецепторы. Зависимость функций клеток Сертоли от ФСГ сопоставима с функциями их гомологов (фолликулярные клетки) в яичнике. Как и фолликулярные клетки яичника, клетки Сертоли секретируют ингибины. Ингибины вместе с тестостероном тормозят образование ФСГ у мужчин.
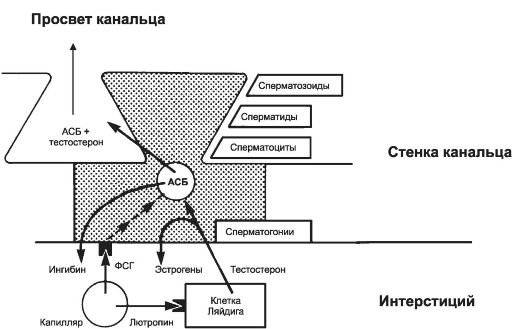
Рис. 19-5. Гормональная регуляция секреторной функции клеток Сертоли
(заштрихована на рисунке). Фоллитропин стимулирует секрецию андрогенсвязывающего белка (АСБ), поддерживающего высокий уровень тестостерона в верхних слоях сперматогенного эпителия. Часть тестостерона в клетках Сертоли путём ароматизации превращается в эстрогены [11].
Гонадолиберин
Гонадолиберин синтезируется в нейросекреторных клетках гипоталамуса. Синтез гонадолиберина подавляется тестостероном и ингибинами (см. рис. 19-4). Гонадолиберин по аксоном нервных клеток транспортируется к срединному возвышению и поступает в кровь из аксонов нейросекреторных клеток в пульсирующем режиме с пиковыми интервалами около 2 ч. Достигая по гипоталамогипофизарной системе кровотока передней доли гипофиза, гона- долиберин активирует синтезирующие ФСГ и ЛГ эндокринные клетки.
Гипофизарные гонадотропины
Гонадотропные гормоны (фолликулостимулирующий - ФСГ и лютеинизирующий - ЛГ), как и гонадолиберин, высвобождаются в кровь в пульсирующем режиме, что особенно характерно для ЛГ. Его концентрации в крови мужчин эпизодически повышаются (с интервалами 90-120 мин).
• Регуляторы синтеза и секреции (см. рис. 19-4). Секреция гонадотропных гормонов контролируется гонадолиберином (активирует) и тестикулярными гормонами (подавляют). Супрессорный эффект тестостерона на секрецию ЛГ в основном проявляется на уровне гипоталамуса (через синтез гонадолиберина), тогда как эстрогены снижают чувствительность гонадотропных клеток к гонадолиберину. В то же время половые стероиды слабо влияют на секрецию ФСГ, тогда как ингибины оказывают выраженное подавляющее действие на гонадотропные клетки, синтезирующие ФСГ.
• Мишени гонадотропных гормонов - яички. Клетки Сертоли имеют рецепторы ФСГ, а клетки Ляйдига - ЛГ.
Φ ФСГ активирует в клетках Сертоли синтез и секрецию андрогенсвязывающего белка, ингибинов, эстрогенов, трансферрина, активаторов плазминогена (см. рис. 19-5).
Φ Лютропин. Клетки Ляйдига имеют рецепторы ЛГ. ЛГ стимулирует в клетках Ляйдига синтез и секрецию тестостеро- на и отчасти эстрогенов.
Эстрогены
В гладком эндоплазматическом ретикулуме клеток Сертоли путём ароматизации происходит превращение тестостерона, син-
тезированного в клетках Ляйдига, в эстрогены. Хотя этот вклад в уровень эстрогенов крови невелик, клетки Сертоли оказывают значительное влияние на синтез тестостерона. Эстрогены связываются с рецепторами в клетках Ляйдига и подавляют синтез тестостерона.
Ингибины
В ответ на стимуляцию ФСГ клетки Сертоли выделяют гликопротеины ингибины (а- и β), блокирующие синтез и секрецию ФСГ (см. рис. 19-5) и гонадолиберина. Структура ингибинов гомологична мюллерову ингибирующему фактору, секретируемому клетками Сертоли у плода.
ЖЕНСКАЯ ПОЛОВАЯ СИСТЕМА
Функция женской половой системы - репродуктивная. Разные органы системы специализируются на выполнении конкретных задач. Так, яичники выполняют герминативную (овогенез, овуляция) и эндокринную (синтез и секреция эстрогенов, прогестерона, релаксинов и ингибинов) функцию, маточные трубы - транспортную (продвижение овулировавшей яйцеклетки в полость матки, оплодотворение), матка - вынашивание плода; канал шейки матки и влагалище - это родовые пути, молочные железы необходимы для вскармливания ребёнка.
Для герминативной функции женщины детородного возраста вне беременности характерна помесячная цикличность процессов (овариально-менструальные циклы).
• Овариальный цикл - продолжение овогенеза (фазы роста и созревания), овуляция, формирование жёлтого тела. Овариальный цикл регулируется гипофизарными гонадотропинами - ФСГ и ЛГ. Маточные трубы, влагалище, молочные железы также подвергаются циклическим изменениям в рамках овариального цикла.
• Менструальный цикл - характерные изменения слизистой оболочки матки (эндометрия), имеющие целью возможность имплантации и без наступления последней заканчивающиеся отторжением части эндометрия (менструация). Все фазы менструального цикла контролируются гормонами яичника - эстрогенами и прогестероном.
Половое созревание девочек
Половое созревание (пубертат) - процесс, в течение которого у девочки появляются физические и поведенческие атрибуты, позволяющие ей воспроизводить потомство. У девочек наступление пубертата обусловлено усиливающейся секрецией яичниками эстрогенов, возникающей под действием гонадотропинов передней доли гипофиза. В Европе и России пубертат у девочек начинается с развития молочных желёз (телархе), что происходит между 8-10-м годом жизни. Другие вторичные половые признаки (лобковое оволосение, увеличение размеров больших и малых половых губ, матки, повышенное отложение жира в нижней части туловища и на бёдрах) появляются в следующие 2-2,5 года. Кульминация пубертата у девочек - появление первой менструации - менархе и регулярных менструаций. Средний возраст менархе 12,8?1,2 года.
Лобковое оволосение
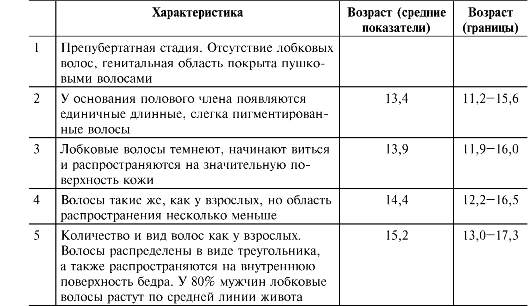
Соматические проявления пубертата у девочек подразделяются на пять стадий. Стадии формирования лобкового оволосения при- ведены на рис. 19-6 и в сопровождающей рисунок таблице.
Адренархе
Адренархе у девочек начинается в 6-8-летнем возрасте. Секреция слабых надпочечниковых андрогенов наступает примерно на 2 года раньше видимого начала пубертата. Дегидроэпиандростерон и дегидроэпиандростерона сульфат обусловливают начало роста волос на лобке и в подмышечных впадинах, активацию работы подмышечных желёз. Появление подмышечного и лобкового оволосения происходит параллельно с началом роста молочных желёз, что делает заметным наступление пубертата у девочек. Пусковой механизм адренархе не известен.
Менархе
Термин «менархе» применяют, чтобы обозначить начало менструальных циклов у девочек, буквально - время наступления первой менструации. Менархе - конечный итог сложной последовательности событий, включающей созревание гипоталамогипофизарно-яичниковой системы. Четыре стадии созревания включают (i) синтез и секрецию гипоталамического гонадолиберина, (ii) усиленный синтез ФСГ и ЛГ в гипофизе, (iii) реакцию яичника на гонадотропины в виде секреции половых гормонов и
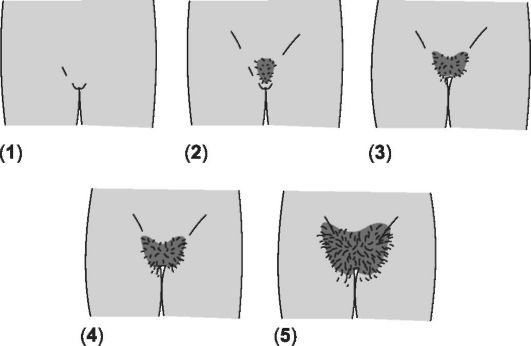
Рис. 19-6. Стадии формирования лобкового оволосения [18]. Цифра в скобках указывает на соответствующую стадию (пояснения см. в сопровождающей таблице).
Характеристика | Возраст (средние показатели) | Возраст (границы) | |
1 | Предпубертатная стадия. Отсутствие лобковых волос, генитальная область покрыта пушковыми волосами | ||
2 | По медиальному краю больших половых губ с двух сторон появляются единичные длинные, слегка пигментированные волосы | 11,7 | 9,3-14,1 |
3 | Лобковые волосы темнеют, начинают виться и распространяются на значительную поверхность кожи лобка | 12,4 | 10,2-14,6 |
4 | Волосы такие же, как у взрослых: пигментированные, жёсткие и вьющиеся. Область распространения несколько меньше, чем у взрослых | 13,0 | 10,8-15,1 |
5 | Взрослая стадия. Количество и вид волос как у взрослых. Волосы распределены в виде треугольника, а также распространяются на внутреннюю поверхность бедра. Волосы вдоль срединной линии живота не растут | 14,4 | 12,2-16,7 |
(iv) установление положительной обратной связи между эстрогенами и ЛГ, что позволяет индуцировать овуляцию (см. ниже).
У девочек концентрации ФСГ и ЛГ в гипофизе и плазме крови низкие, так как гипоталамический «пульсовый генератор» гона- долиберина работает медленно (юношеская пульсация). Первым эндокринологическим проявлением пубертата становится возрастание пульсовой секреции ФСГ и ЛГ во время сна. Предполагают, что начало пубертата связано с устранением тормозящего действия ЦНС на гипоталамический «пульсовый генератор» гонадолиберина. Суточные различия в секреции ФСГ и ЛГ почти полностью исчезают к концу пубертата.
Наступление менархе также связано с достижением определён- ного содержания жировой ткани в организме. В этом отношении важную роль играют соматомедин C и лептин, обеспечивающие связь между состоянием метаболизма и регуляцией ЦНС полового созревания.
Созревание яичников в пубертатном периоде включает начало секреции эстрогенов фолликулярными клетками, окружающими овоцит. С началом пубертата происходит размножение, а затем атрезия фолликулярных клеток одного из фолликулов. Под действием овариальных эстрогенов, выделяемых фолликулярными клетками, происходит созревание яйцеклетки. Итог этого процесса - овуляция (выход первой зрелой яйцеклетки). Одновременно начинается образование прогестерона формирующимся жёлтым телом. Образующиеся в яичнике эстрогены вызывают в эндометрии пролиферативные изменения, которые сменяются секреторными, происходящими под влиянием прогестерона. Всё это даёт возможность имплантации. В начале становления репродуктивной функции, когда девочки только достигают менархе, некоторые циклы могут быть ановуляторными, так как положительная обратная связь между ги- поталамусом и эстрогенами в полной мере ещё не сформировалась. Маточные кровотечения, появляющиеся в результате продолжительного воздействия эстрогенов на эндометрий, в действительности вызваны отторжением пролиферативного или гиперпластического эндометрия, их нельзя считать собственно менструацией. Эти кровотечения весьма непредсказуемы и могут быть достаточно выраженными из-за отсутствия прогестерона, способствующего пре- кращению менструации. Через 5 лет после наступления менархе у 90% девочек устанавливаются регулярные овуляторные циклы.
Телархе
Каждая молочная железа представлена дольками железистой ткани, волокнистой соединительной тканью, соединяющей доли и дольки, и жировой тканью, расположенной между долями. Каждая доля подразделяется на дольки, состоящие в основном из альвеол, кровеносных сосудов и молочных протоков. Ткани молочных желёз весьма чувствительны к гормонам уже с внутриутробного периода. Стадии развития молочных желёз в пубертате показаны на рис. 19-7 и в сопровождающей его таблице.
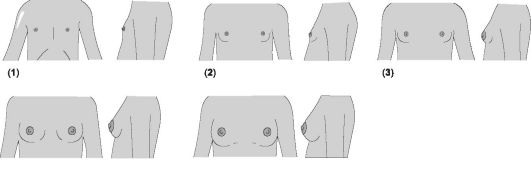
Рис. 19-7. Стадии развития молочных желёз [18]. Цифра в скобках указывает на соответствующую стадию, пояснения см. в сопровождающей рисунок таблице.
Характеристика | Возраст (средние показатели) | Возраст (крайние границы) | |
1 | Препубертатная стадия: выступающий сосок и небольшая плоская ареола | ||
2 | Стадия почки: сосок и ареола приподнимаются в виде небольшого (первичного) возвышения. Диаметр ареолы увеличивается | 11,2 | 9,0-13,3 |
3 | Дальнейшее увеличение размеров железы (с приподнятием соска) и ареолы, ареола не отграничена от железы | 12,2 | 10,0-14,3 |
4 | Ареола и сосок формируют вторичное возвыше- ние, отграничены от железы | 13,1 | 10,8-15,3 |
5 | Взрослая стадия. Ареола возвышается над по- верхностью железы; сосок выступает | 15,3 | 11,9-18,8 |
Ко времени рождения молочная железа почти полностью состоит из молочных протоков с очень небольшим количеством альвеол. Эти рудиментарные молочные железы могут обладать незначительной секреторной функцией в течение нескольких дней после рождения. Наличие такой секреции обусловлено высоким содержанием пролактина, а также воздействием больших концентраций плацентарных эстрогенов и прогестерона во время беременности. Как только плацентарные гормоны исчезают из циркуляции ново- рождённого, молочные железы входят в состояние покоя до момента пубертата.
С наступлением пубертата овариальные эстрогены индуцируют рост молочных протоков. Протоки не только начинают расти, но и ветвятся, их концы заканчиваются плотными сферическими клеточными образованиями, которые позже станут альвеолами долек. Молочная железа и её сосок начинают увеличиваться в размерах. Последующее за наступлением менархе установление регулярного циклического синтеза эстрогенов и прогестерона вызывает дополнительный рост и ветвление протоков и закладку рудиментарных альвеол. Молочная железа продолжает ещё более увеличиваться за счёт отложения жира и развития соединительной ткани. Надпочечниковые кортикостероиды ещё более усиливают развитие системы протоков. Тем не менее конечная дифференцировка и рост молочной железы происходят только во время беременности.
Соматический рост
Пубертатный
скачок роста у девочек обычно начинается на 2 года раньше, чем у
мальчиков. Это обстоятельство частично об- условливает различие роста у
мужчин и женщин (в среднем на
ГЕРМИНАТИВНАЯ ФУНКЦИЯ
Яичники выполняют две основные функции - герминативную (образование способных к оплодотворению женских половых клеток) и эндокринную (синтез половых стероидных гормонов, ре-
лаксинов и ингибинов). В яичниках женщины репродуктивного возраста половые клетки находятся в составе фолликулов, разви- тие которых под влиянием гипофизарных гонадотропинов происходит по схеме: примордиальный - первичный - вторичный - третичный (зрелый фолликул, граафов пузырёк). Суть герминативной функции - процесс овогенеза, в результате митотических и мейотических делений которого из незрелых половых клеток (овогонии) через промежуточные стадии овоцитов I и II формируются зрелые яйцевые клетки.
Овогенез
Овогенез состоит из стадий размножения, роста и созревания. Стадия размножения завершается ещё в плодном периоде, после этого новые овогонии не образуются. Каждый яичник новорож- дённой девочки содержит в составе примордиальных фолликулов около 2 млн овоцитов, находящихся в профазе первого мейотического деления. К началу полового созревания общее количество примордиальных фолликулов в обоих яичниках оценивается в 200-400 тыс. Все остальные стадии овогенеза происходят при развитии фолликулов в ходе овариального цикла, продолжительность которого в среднем составляет 28 сут.
Овариальный цикл
В каждом овариальном цикле ограниченное количество примордиальных фолликулов под влиянием гипофизарных гонадо- тропинов начинает развитие по схеме: примордиальный - первичный - вторичный - третичный (зрелый фолликул, граафов пузырёк). Первая половина цикла - фолликулярная (под влиянием ФСГ происходит развитие части примордиальных фолликулов), вторая половина - лютеиновая (под влиянием ЛГ из клеток овулировавшего граафова пузырька формируется эндокринная железа - жёлтое тело). Овуляция приходится примерно на середину цикла.
• Примордиальный фолликул покрыт одним слоем фолликулярных клеток (гранулёза, клетки гранулёзы) и окружён базальной мембраной.
98% примордиальных фолликулов в течение репродуктивного периода погибает, около 2% достигает стадии первичного и вторичного фолликулов, не более 400 развивается в граафов пузырёк и подвергается овуляции (каждый цикл -
1, намного реже - 2). Все начавшие развитие, но не достигшие стадии овуляции фолликулы подвергаются атрезии (образуются атретические тела).
Ф Риск генных дефектов плода увеличивается с возрастом матери, что не в последнюю очередь объясняется чрезвычайно большой продолжительностью жизни овоцита до его оплодотворения (до 40-50 лет).
Ф Каждый цикл от 3 до 30 примордиальных фолликулов под влиянием ФСГ вступает в фазу роста и формирует первичный фолликул.
• Первичный фолликул. Фолликулярные клетки имеют рецепторы к ФСГ, эстрогенам и тестостерону.
Ф ФСГ индуцирует синтез ароматазы. Из тестостерона и других стероидов образуются эстрогены.
Ф Эстрогены (преимущественно 17р-эстрадиол) стимулируют пролиферацию фолликулярных клеток (количество клеток гранулёзы стремительно увеличивается, фолликул растёт) и экспрессию новых рецепторов ФСГ и половых стероидных гормонов.
Ф Тестостерон угнетает пролиферацию фолликулярных клеток.
Ф Инсулиноподобный фактор роста C также стимулирует рост фолликулов.
• Вторичные фолликулы характеризуются дальнейшим ростом, появляется так называемый доминантный фолликул, в его составе формируется выраженная оболочка (theca), а между фолликулярными клетками (гранулёза) появляются полости. Ф Доминантный фолликул - один из фолликулов, который
опережает в росте остальные.
Ф Лютеинизирующий гормон (ЛГ) стимулирует синтез андрогенов (андростендион и тестостерон) в клетках theca.
Ф Андрогены из theca через базальную мембрану (стекловидная оболочка на более поздних стадиях развития) диффундируют к клеткам гранулёзы, где конвертируются при помощи ароматазы в эстрогены. На этой стадии в крови значительно увеличивается содержание эстрогенов.
Ф Увеличение содержания эстрадиола в крови усиливает секрецию ЛГ в аденогипофизе и подавляет секрецию ФСГ (считают, что при небольшом содержании эстрогенов го-
надолиберин стимулирует клетки, синтезирующие ФСГ, а при значительном содержании эстрогенов - клетки, синтезирующие ЛГ). Ф Ослабление секреции ФСГ и усиление секреции ЛГ в конечном счёте блокирует рост фолликулов.
• Третичный фолликул (преовуляторный, граафов пузырёк) быстро растёт от 200 мкм до 1-2,5 см в диаметре, прежде всего за счёт накопления жидкости в его полости. Непосредственно перед овуляцией на поверхности яичника появляется истон- чённый участок фолликула, называемый стигмой. На поздних стадиях развития фолликула происходят характерные изменения содержания гормонов в крови (см. рис. 19-11).
Ф ФСГ стимулирует транспорт жидкости в полость фолликула, а также индуцирует экспрессию рецепторов ЛГ на клетках гранулёзы.
Ф Лютропин (ЛГ) стимулирует лютеинизацию фолликулярных клеток и синтез прогестерона.
Ф Уровень эстрогенов быстро увеличивается, достигая пика приблизительно за 24-36 ч до овуляции (см. рис. 19-11).
Ф Преовуляторное повышение уровня прогестерона вызывает выброс ФСГ в середине цикла за счёт усиления гипофизарного ответа на гонадолиберин.
Ф Содержание ЛГ увеличивается постепенно до середины цикла, затем происходит резкий подъём его уровня. Это объясняют тем, что при высокой концентрации эстрогенов под влиянием гонадолиберина аденогипофиз усиливает секрецию ЛГ.
• Овуляция происходит примерно через 10-12 ч после достижения пика ЛГ и через 24-36 ч после пика эстрадиола; подъём уровня ЛГ, начинающийся за 28-36 ч до овуляции, - наиболее объективный её признак (рис. 19-8).
Ф Сроки овуляции. Овуляция происходит приблизительно через 2 нед после начала цикла (чаще 11-й, 12-й или 13-й день 28-дневного цикла). При 28-дневном цикле овуляция возможна между 8-м и 20-м днём.
Ф Выброс ЛГ стимулирует также лютеинизацию клеток гра- нулёзы и синтез прогестерона и простагландинов в клетках фолликула. Истончение и разрыв стенки фолликула происходят под влиянием простагландинов и протеолитических ферментов гранулёзы.
• Жёлтое тело синтезирует прогестерон и эстрогены. Прогестерон оказывает выраженные эффекты на эндометрий и миометрий (как в рамках завершающегося менструацией цикла, так и в случае имплантации бластоцисты).
Ф Менструальное жёлтое тело функционирует до завершения цикла (имплантации нет).
♦ Уровень прогестерона достигает пика через 8-9 дней после овуляции, что приблизительно соответствует времени имплантации.
♦ Термогенный эффект прогестерона обусловливает повышение базальной температуры тела как минимум на
Ф Жёлтое тело беременности активно функционирует в течение первой половины беременности, позднее начинается его инволюция.
♦ Хорионический гонадотропин посредством рецепторов обеспечивает расцвет (значительное увеличение размеров) жёлтого тела ЛГ.
♦ Плацента - важный источник прогестерона во время беременности.
• Белое тело - соединительнотканный рубец на месте завершившего функцию и дегенерировавшего жёлтого тела.
Таким образом, каждые 28 дней гонадотропные гормоны передней доли гипофиза вызывают рост 8-12 первичных фолликулов в каждом яичнике. Один из этих фолликулов становится зрелым и овулирует на 14-й день цикла. Во время роста фолликулов выделяется значительное количество эстрогена. После овуляции секреторные клетки овулировавшего фолликула превращаются в жёлтое тело, которое секретирует прогестерон. 2 нед спустя жёл- тое тело дегенерирует, после чего начинается новый овариальный цикл.
Вырабатываемые на протяжении овариального цикла в яичниках эстрогены и прогестерон воздействуют на слизистую оболочку маточных труб, матки и влагалища, а также на ГМК миометрия, вызывая характерные циклические изменения, особенно выраженные в эндометрии (см. рис. 19-8). Эти циклические изменения известны как менструальный цикл.
Менструальный цикл
Изменения гормонального фона (содержание в крови эстрогенов и прогестерона в разные дни овариального цикла, см. рис. 19-11, А, Б и В) прямо влияют на состояние эндометрия, а также слизистой оболочки маточных труб, цервикального канала и влагалища. Слизистая оболочка матки подвергается циклическим изменениям (менструальный цикл, см. рис. 19-8). В каждом цикле эндометрий проходит менструальную, пролиферативную и секреторную фазы. В эндометрии различают функциональный (отпадающий при менструации) и базальный слой (сохраняющийся при менструации).
• Продолжительность цикла (от первого дня наступившей до первого дня следующей менструации) 28?7 дней. Если не происходит оплодотворения и имплантации бластоцисты, текущий цикл завершается менструацией и начинается новый цикл. Такой биоритм сохраняется на протяжении всего репродуктивного периода. Укороченный менструальный цикл (менее 21 дня) - полименорея, удлинённый менструальный цикл (более 35 дней) - олигоменорея. Термины гипоменоррагия и меноррагия означают скудные и обильные профузные кровотечения во время регулярного периода. Метроррагия - кровотечение из матки между менструальными периодами, олигоменорея - уменьшение частоты периодов, дисменорея - болезненные менструации.
• Регулярность цикла. Как правило, менструальный цикл нерегулярен в течение 2 лет после менархе (первая менструация) и в течение 3 лет перед менопаузой. В таких ситуациях нередки ановуляторные циклы (овуляции не происходит). Беременность обрывает (точнее, блокирует) овариально-менструальный цикл.
• Фазы цикла в эндометрии (см. рис. 19-8).
Ф Пролиферативная (фолликулярная) - первая половина цикла - длится от первого дня менструации до момента ову- ляции; в это время под влиянием эстрогенов (в основном эстрадиола) происходит пролиферация клеток базального слоя и восстановление функционального слоя эндометрия. Длительность фазы может варьировать. Базальная температура тела нормальна.
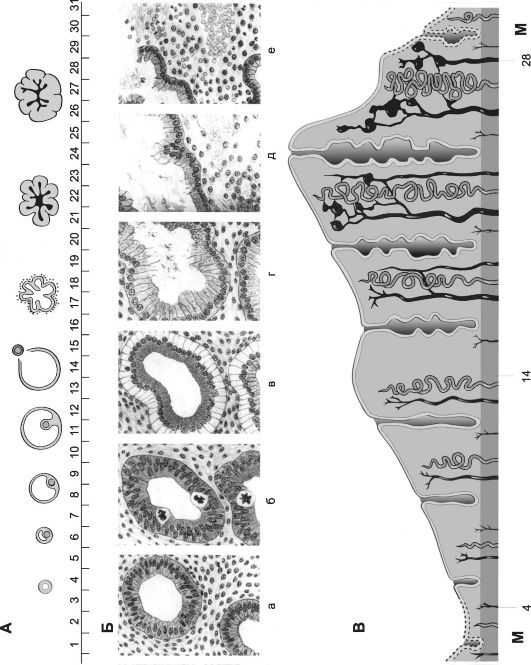
Рис. 19-8. Овариально-менструальный цикл [18]. По абсциссе указаны дни цикла. А - овариальный цикл; Б - эндометрий в различных фазах менструального цикла: а - ранней пролиферативной; б - поздней пролиферативной; в - ранней секреторной; г - средней секреторной; д - поздней секреторной; е - менструальной; В - циклические изменения в эндометрии матки (менструальный цикл).
Ф Секреторная (лютеиновая) фаза - вторая половина - продолжается от овуляции до начала менструации. Высокий уровень секретируемого жёлтым телом прогестерона соз- даёт благоприятные условия для имплантации зародыша. Базальная температура тела выше 37 ?С. Клетки стромы эндометрия в ходе так называемой предецидуализации приобретают черты децидуальных клеток. Если не произошло имплантации, то вследствие вызванного локальным выделением простагландинов (Пг) спазма сосудов ухудшается кровоток в функциональном слое эндометрия (ишемия), что приводит к отторжению функционального слоя и генитальному кровотечению.
• Ф Менструальная фаза - отторжение функционального слоя эндометрия. При длительности цикла 28 дней менструация продолжается 5?2 дня. Менструальный цикл и фертильность
Ф В менструальном цикле, имея в виду возможность оплодотворения, можно выделить три фазы (абсолютной стерильности, относительной стерильности и фертильности). На протяжении этих фаз наиболее существенны для ориентации состояние слизистого отделяемого шейки матки и влагалища, а также базальная температура тела.
♦ Относительная стерильность (фаза 1) длится с последнего дня менструации до овуляции. Продолжительность этой фазы зависит от быстроты ответа фолликула на действие гормонов гипофиза. В этой фазе иногда возникают сложности с контрацепцией, поскольку продолжительность фазы может варьировать от цикла к циклу.
♦ Фаза фертильности (фаза 2) начинается с момента овуляции и заканчивается через 48 ч после неё. Эти 48 ч включают время, в течение которого зрелая яйцеклетка способна к оплодотворению (24 ч); ещё 24 ч отводят на возможную неточность клинического определения времени овуляции. Сперматозоиды, попавшие в шеечную слизь, которая обильно секретируется во время этой фазы, сохраняют способность к оплодотворению яйцеклетки в течение максимум 5 дней. Из практических соображений считают, что фертильная фаза продолжается 6-8 дней.
♦ Абсолютная стерильность (фаза 3) начинается через 48 ч после овуляции и продолжается до конца менструации. Длительность этой фазы вполне постоянна и составляет около 10-16 дней.
Ф На протяжении фаз абсолютной стерильности, относительной стерильности и фертильности наиболее существенны для ориентации состояние слизистого отделяемого шейки матки и влагалища, а также базальная температура тела.
♦ Шеечная слизь. В связи с изменением концентрации эстрогенов и прогестерона изменяется качество шеечной слизи.
■ Фаза 1. Сразу после менструации слизь не образуется (или её очень мало). Если слизь всё-таки есть, то она густая, липкая и непрозрачная. При наличии слизи возникает ощущение клейкости во влагалище. В отсутствие слизи ощущается сухость слизистой оболочки вульвы.
■ Фаза 2. По мере синтеза растущими фолликулами большего количество эстрогенов количество слизи, отделяемой из половых путей, увеличивается.
- Слизь становится более обильной, жидкой, тянущейся, прозрачной и водянистой. В этот период выявляют феномен папоротника (арборизации): при высыхании слизи на предметном стекле образуется узор, напоминающий по форме лист папоротника.
- Появляется ощущение влажности слизистой оболочки вульвы.
- Пик образования шеечной слизи наблюдают при максимальной секреции эстрогенов максимум за 3 дня до овуляции; как правило, пик образования шеечной слизи определяют только задним числом (т.е. после того, как слизь опять изменится и станет густой, липкой и непрозрачной).
- Если считать, что зрелая яйцеклетка может оплодотвориться в течение 24 ч, то четвёртый день после пика образования шеечной слизи означает конец фертильной фазы.
■ Фаза 3. После овуляции с повышением уровня прогестерона быстро изменяется количество и состав слизи.
Количество слизи резко уменьшается, и иногда она исчезает совсем. Если слизь есть, она становится густой, липкой и непрозрачной, сходной со слизью I (стерильной) стадии цикла. ♦ Базальная температура тела (рис. 19-9) - температура тела в состоянии полного покоя, сразу после пробуждения и до начала повседневной активности, включая при- ём пищи. В фолликулярной фазе менструального цикла базальная температура ниже 37 ?С, перед овуляцией она снижается на 0,1-0,2 ?С, затем происходит резкий подъ- ём на 0,3-1 ?С, и в лютеиновой фазе цикла температура бывает выше 37 ?С в течение 8-10 дней.
• Шейка матки. В
фолликулярной фазе секреция шеечной слизи увеличивается в десятки раз,
развивается отёк слизистой оболочки. Непосредственно перед овуляцией
наблюдают «феномен папоротника». При овуляции наружное отверстие
цервикального канала открывается до
• Влагалище. В начале фолликулярной фазы влагалищный эпителий тонкий и бледный. Под влиянием эстрогенов происходит пролиферация эпителия, он утолщается, появляются признаки частичного ороговения. В лютеиновой фазе на поверхности эпителия появляются лейкоциты и роговые чешуйки.
• Молочные железы. Эстрогены вызывают пролиферацию клеток в протоках молочной железы, прогестерон способствует росту долек и альвеол, что известно как нагрубание молочных желёз. Многие женщины отмечают при этом болевые ощущения в дни, предшествующие менструации. Эти явления связаны с растяжением протоков, гиперемией и отёчностью интерстициальной ткани молочной железы.
• Клинические проблемы менструального цикла - дисменорея и предменструальный синдром.
Ф Дисменорея (болезненные менструации) часто возникает у молодых женщин и обычно начинается с наступлением овуляторных циклов. При ановуляторных циклах дисменорея не развивается. Менструальные боли - следствие сокращений миометрия под влиянием простагландинов (Пг),
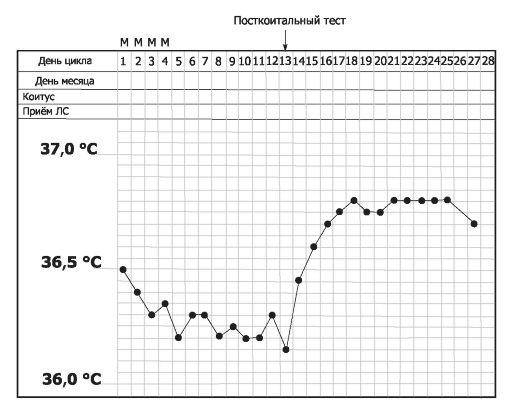
Рис. 19-9. Базальная температура тела в течение менструального цикла [18].
На графике приведена типичная двухфазная кривая с указанием дней менструации (М) и половой близости (-1). Посткоитальная проба - оценка сперматозоидов в шеечной слизи через 2-12 ч после полового сношения и за 1-2 дня до овуляции после трёхдневного полового воздержания. Проведение и оценка: шеечную слизь исследуют под микроскопом, чтобы определить общее количество сперматозоидов, а также степень их подвижности. Пробу считают положительной (удовлетворительной), если в каждом поле зрения при большом увеличении видно более 10 подвижных сперматозоидов. Критерии неудовлетворительного результата: сперматозоидов очень мало или совсем нет (азооспермия); большинство сперматозоидов неподвижно; подвижность сперматозоидов характеризуется в большей степени колебательными, а не поступательными движениями.
синтезируемых в клетках эндометрия. Уровень Пг в первый день менструальной фазы увеличивается в несколько раз по сравнению с лютеиновой фазой. Ф Предменструальный синдром - комплекс симптомов (например, раздражительность, нагрубание молочных желёз, плаксивость, депрессия, утомляемость), возникающий с приближением менструации и исчезающий с началом кровотечения.
Эндокринология женской половой системы
Гормональная регуляция овариально-менструального цикла
Овариально-менструальный цикл контролируют гипофизарные гонадотропины - ФСГ и ЛГ. Эту эндокринную функцию передней доли гипофиза регулирует гипоталамический гонадолиберин - люлиберин. В свою очередь, гормоны яичника (эстрогены, прогестерон, а также ингибины) вовлечены в регуляцию синтеза и секреции гонадотропинов гипофиза и люлиберина гипоталамуса. Таким образом, циклические изменения яичника и эндометрия - иерархическая (гипоталамус - гипофиз - яичники - матка) и саморегулирующаяся (яичники - гипоталамус и гипофиз) система, функционирующая в течение репродуктивного периода (от менархе до наступления климактерических изменений - менопаузы). Иными словами, все гормоны женской половой сферы формируют регуляторный контур, упрощённая схема которого приведена на рис. 19-10.
• Гонадолиберин. Секреция гонадолиберина имеет пульсирующий характер: пики усиленной секреции гормона продолжительностью несколько минут сменяются 1-3-часовыми интервалами относительно низкой секреторной активности. Частоту и амплитуду секреции гонадолиберина регулирует уровень эстрогенов и прогестерона.
• Фолликулярная стадия цикла. Резкое падение содержания эстрогенов и прогестерона в конце каждого цикла (вследствие инволюции менструального жёлтого тела) побуждает нейросекреторные клетки гипоталамуса выделять гонадолиберин с пиками усиленной секреции гормона продолжительностью несколько минут с интервалом в 1 ч. В первую очередь гормон секретируется из пула, запасённого в нейросекреторных
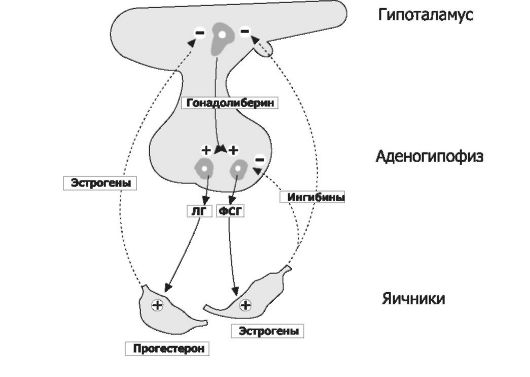
Рис. 19-10. Регуляторные взаимоотношения в системе «гипоталамус- гипофиз-яичники» [7]. Непрерывные стрелки и символ «+» - активирующие влияния; прерывистые стрелки и символ «-» - ингибирующие влияния.
гранулах, а затем - сразу же по окончании их синтеза. Такой режим секреции гонадолиберина активирует гонадотропные клетки аденогипофиза.
• Лютеиновая стадия цикла. Жёлтое тело активно продуцирует половые гормоны. На фоне высокого содержания эстрогенов и прогестерона интервал между пиками усиленной секреции гонадолиберина увеличивается до 2-3 ч, что недостаточно для того, чтобы стимулировать секрецию гонадотропных гормонов.
• Фоллитропин (фолликулостимулирующий гормон, ФСГ).
Ф Секреция. В фолликулярной стадии (в начале цикла) на фоне пониженного содержания в крови эстрогенов и про- гестерона гонадолиберин стимулирует секреторную активность клеток, синтезирующих ФСГ. Эстрогены (с пиком за сутки до овуляции) и ингибины подавляют продукцию
ФСГ.
Ф Мишени фоллитропина - фолликулярные клетки. ФСГ (действуя вместе с эстрадиолом) увеличивает плотность рецепторов фоллитропина в мембране клеток гранулёзы, тем самым усиливая своё действие на мишень.
Ф Функция. ФСГ стимулирует пролиферацию фолликулярных клеток и рост фолликула. Активирует в его клетках ароматазу и усиливает секрецию эстрогенов. Инициирует встраивание рецепторов ЛГ в мембрану фолликулярных клеток и секрецию ингибинов в конце фолликулярной стадии.
• Лютропин (лютеинизирующий гормон, ЛГ)
Ф Секреция. В конце фолликулярной стадии на фоне высокой концентрации эстрогенов блокируется продукция ФСГ и одновременно стимулируется секреция ЛГ. Пик ЛГ наблюдается за 12 ч до овуляции. Сигналом к снижению уровня ЛГ является начало секреции клетками гранулёзы прогестерона.
Ф Мишени. Рецепторы ЛГ имеют клетки theca фолликулов и клетки гранулёзы. После активации фоллитропином в клетках появляются рецепторы ЛГ.
Ф Функция. Лютеинизация фолликулярных клеток и клеток внутренней теки. Стимуляция синтеза андрогенов в клетках theca. Индукция секреции прогестерона клетками грану- лёзы. Активация протеолитических ферментов гранулёзы. На пике ЛГ завершается первое деление мейоза.
• Эстрогены и прогестерон
Ф Секреция эстрогенов клетками гранулёзы постепенно нарастает в фолликулярной стадии и достигает пика за сутки до овуляции. Продукция прогестерона начинается в клетках гранулёзы до овуляции; основной источник прогестерона - жёлтое тело. В лютеиновой стадии овариального цикла синтез эстрогенов и прогестерона значительно усиливается.
Ф Мишени. К половым гормонам чувствительны нейросекреторные клетки гипоталамуса, гонадотропные клетки, фолликулярные клетки яичника, клетки слизистых оболочек матки, яйцевода, влагалища, клетки молочных желёз.
Ф Функции. Если одновременно повышается содержание в крови прогестерона и эстрогенов, интервал между фаза- ми усиленной секреции гонадолиберина увеличивается до 2-3 ч, что блокирует продукцию гонадотропных гормонов, а следовательно, рост и созревание очередного фолликула. Если содержание половых гормонов в крови резко снижается, пики секреции гонадолиберина разделены одночасовым интервалом. На этом фоне в аденогипофизе активируется
секреция ФСГ (начинается фолликулярная стадия цикла). Эстрогены контролируют пролиферативную фазу менструального цикла (восстановление функционального слоя эндометрия), а прогестерон - секреторную фазу (подготовка эндометрия к имплантации). Снижение содержания в крови эстрогенов и прогестерона сопровождается отторжением функционального слоя эндометрия и маточным кровотечением (менструальная фаза). Эстрогены и прогестерон в сочетании с пролактином (и хорионическим соматомаммотропином у беременной) стимулируют дифференцировку секреторных клеток молочной железы.
• Ингибины. Каждый из физиологически активных ингибинов (α и β) состоит из двух субъединиц (одной общей α-СЕ и двух разных β-СЕ). Оба ингибина блокируют синтез гипоталамического гонадолиберина и гипофизарного ФСГ. В то же время димер из 2 β-СЕ (активин) in vitro стимулирует секрецию ФСГ, однако роль активина in vivo не выяснена.
В заключение приводим схему (рис. 19-11), которая иллюстрирует основные положения, относящиеся к гормональной регуля- ции овариально-менструального цикла.
Гормональная контрацепция
Гормоны с целью контрацепции (в том числе и пероральные контрацептивы) предложены на основе понимания схемы гормональной регуляции овариально-менструального цикла. Ключевая позиция - ингибирование секреции гонадолиберина к середине цикла, что предупреждает увеличение содержания гипофизарных гонадотропинов и тем самым предупреждает овуляцию. Другими словами, гормональные контрацептивы приводят к появлению ановуляторных циклов.
Менопауза
Процесс естственного возрастного угасания функций женской половой системы принято подразделять на пременопаузальный период, менопаузу и постменопаузальный период.
• Пременопаузальный период - от 45 лет до наступления менопаузы.
• Менопауза - аменорея - период от первого отсутствия менструации и последующие 12 мес у женщин старше 45 лет; последняя менструация в среднем наступает в возрасте 50,8 года.
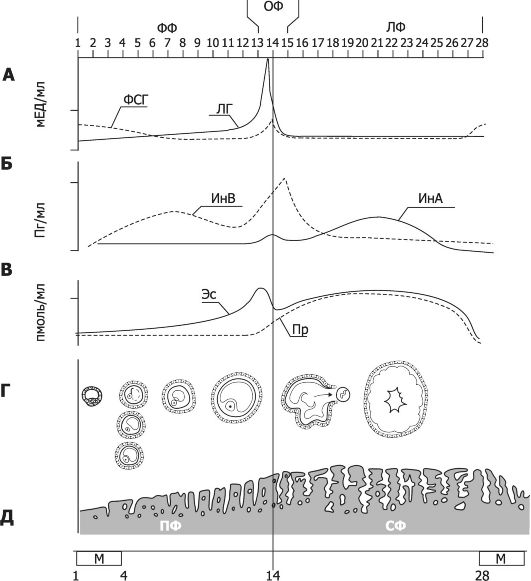
Рис. 19-11. Гормональная регуляция овариально-менструального цикла (по абсциссе указаны дни цикла). А-В - циклические изменения содержания в крови гонадотропных гормонов, ингибинов, эстрогенов и прогестерона; Г - регулируемые гонадотропными гормонами созревание фолликула и овуляция; Д - вызванные эстрогенами и прогестероном изменения эндометрия матки. ФФ - фолликулярная фаза; ОФ - овуляторная фаза; ЛФ - лютеиновая фаза; ФСГ - фолликулостимулирующий гормон; ЛГ - лютеинизирующий гормон; ИнВ - ингибин B; ИнА - ингибин A; Эс - эстрадиол; Пр - прогестерон; ПФ - пролиферативная фаза; СФ - секреторная фаза; М - менструация.
• Постменопаузальный период начинается по завершении менопаузы и длится вплоть до смерти женщины.
Физиология менопаузы определяется массовой атрезией примордиальных фолликулов, изменениями гормональной регуляции и разнообразными последствиями последней.
• Атрезия примордиальных фолликулов. Из-за малого количества созревающих фолликулов увеличиваются интервалы между циклами. Прекращается овуляторный выброс ФСГ и ЛГ, овуляторные циклы сменяются циклами с недостаточностью жёлтого тела, а затем - ановуляторными циклами.
• Изменения гормональной регуляции состоят в том, что синтез эстрогенов значительно уменьшается, если одновременно увеличивается выброс в кровь гипофизарных гонадотропинов и сохраняется прежний уровень синтеза андростендиона и тестостерона.
Φ Сокращение выработки эстрогенов. С возрастом постепенно уменьшается абсолютное количество примордиальных фол- ликулов, к менопаузе они практически исчезают, развитие очередного фолликула замедляется или вовсе не происходит, при этом сокращается или прекращается выработка эстрогенов. Эстрогены в основном поступают из надпочечников, значительно меньшую часть вырабатывает строма яичников. В то же время у полных и тучных женщин в жировой ткани происходит усиленное образование эстрона из андростендиона.
Φ Усиленная выработка гонадотропинов происходит по механизму отрицательной обратной связи (ФСГ с 40 лет, ЛГ с 45 лет). После менопаузы содержание ЛГ возрастает в 3 раза, а содержание ФСГ - в 14 раз.
Φ Андростендион. Гормональная активность стромы яичника не изменяется: происходит секреция андростендиона (пред- шественника эстрона) и небольшого количества тестостерона.
Реакция органов-мишеней на сниженное содержание эстрогенов
• Количества эстрогенов недостаточно для пролиферативных изменений эндометрия. При отсутствии жёлтого тела значительно снижается синтез прогестерона, в результате уменьшается секреторная активность маточных желёз. В силу названных причин прекращаются менструации.
• Эндометрий становится рыхлым, атрофичным, со множеством мелких кровоизлияний; количество эндометриальных желёз значительно уменьшается. Миометрий атрофируется, матка уменьшается в размере.
• Влагалище становится менее растяжимым, особенно в верхних отделах, слизистая оболочка - бледной, истончённой и сухой.
• Малые половые губы выглядят бледными и сухими, уменьшается содержание жировой ткани в больших половых губах.
• Молочные железы теряют упругость и форму вследствие отложения жировой ткани и атрофии железистой ткани.
• Костная ткань постепенно теряет кальций, в результате чего возникает остеопороз, часто сопровождающийся болями, возможно искривление позвоночника и переломы костей.
• Тип оволосения изменяется на мужской за счёт относительного преобладания андрогенов.
Климактерические расстройства
Климактерические расстройства по характеру проявлений подразделяются на вазомоторные, эмоционально-психические, атро- фические и остеопорозные.
• Вазомоторные симптомы у большинства женщин продолжаются 1-2 года, но могут длиться и 5 лет. Наиболее часто встречаются «приливы» в виде неожиданного покраснения кожи лица, шеи и груди, ощущения сильного жара и потоотделения. Продолжительность «приливов» составляет от нескольких секунд до нескольких минут. «Приливы» чаще возникают и более выражены ночью или во время стрессовых ситуаций. Лечение эстрогенами уменьшает частоту и выраженность «приливов». Из других симптомов возможны гипергидроз (увеличенная потливость), скачки АД, головные боли, озноб, сердцебиение.
• Эмоционально-психическая симптоматика в виде раздражительности, сонливости, слабости, беспокойства, депрессии, забывчивости, невнимательности, снижения либидо.
• Остеопороз - дистрофия костной ткани с перестройкой её структуры, характеризующаяся уменьшением числа костных перекладин в единице объёма кости, истончением, искривлением и полным рассасыванием части этих элементов, обусловливающая повышенную склонность к переломам. Усиленная
резорбция кости сохраняется в течение 3-7 лет после менопаузы. Приблизительно у половины женщин старше 60 лет, не получающих заместительной эстрогенной терапии, возникают компрессионные переломы позвоночника, переломы шейки бедра и других костей.
• Атрофические изменения в виде сухости кожи, ломкости ногтей, морщин, сухости и выпадения волос. Возможно развитие атрофического вагинита, зуда вульвовагинальной области, диспареунии (болезненный половой акт), расстройств мочеиспускания.
• Сердечно-сосудистые заболевания. Риск развития инфаркта миокарда у женщин до менопаузы значительно меньше, чем у мужчин того же возраста. После менопаузы этот риск возрастает. Приём эстрогенов во время менопаузы значительно снижает содержание липопротеинов низкой плотности, холестерола сыворотки и увеличивает концентрацию липопротеинов высокой плотности.
• Лечение и профилактика. Лечение климактерических расстройств состоит в применении недостающих эстрогенов и прогестинов (заместительная терапия). Профилактика развития климактерических расстройств - ранняя заместительная гормональная терапия препаратами, содержащими эстрогены и гестагены, а также постоянный приём препаратов кальция и витамина D.
ФИЗИОЛОГИЯ ПОЛОВОГО ВОЗБУЖДЕНИЯ
У человека оплодотворение происходит внутри женских половых путей, и для возникновения естественного зачатия необходим интимный контакт (половой акт) между мужчиной и женщиной. У большинства животных спаривание сезонно, зависит от содержания половых гормонов и стадии цикла, т.е. направлено именно на осуществление репродуктивной функции. Люди же могут вступать в половую связь и в то время, когда зачатие совершенно невозможно. Другими словами, сексуальность человека преследует не только и даже не столько функцию продолжения рода, сколько составляет своеобразное развлечение и удовольствие, высшим проявлением которой является любовь. Эти нерепродуктивные стороны сексуальности человека весьма важны и для человека, и для
общества, они крайне разнообразны, вариабельны и подвержены сильнейшим индивидуальным и социокультурным влияниям. Например, то, что доставляет удовольствие одному человеку, может не быть таковым для другого, а нормы поведения одного общества могут быть неприемлемы для другого. В то же время необходимо назвать и столь характерную для обществ западного типа существенную тенденцию толерантного отношения к разнообразным формам проявления сексуальности человека. С физиологической же точки зрения почти во всех ситуациях физиологическая и психологическая удовлетворённость - главные мотивационные факторы сексуального поведения человека.
Фазы сексуального поведения
На основании тысяч научно документированных наблюдений гетеросексуальных отношений и актов мастурбации процесс полового контакта принято подразделять на фазу полового возбуждения, плато, оргазм и фазу расслабления (рис. 19-12).
Общая характеристика полового поведения
Фаза полового возбуждения наиболее разнообразна. Многие стороны полового влечения, либидо (желания половой близости) и полового возбуждения тесно связаны с особенностями личности и культурными традициями. Например, табу инцеста, т.е. запрет вступать в брак и половую связь близким родственникам, практически универсально в разных культурах, в то время как отношение к женской груди или массе тела значительно варьирует.
• Эротические стимулы могут быть самыми различными, включая фантазии, запахи, звуки и физические проявления (прикосновения, ласки). На основании исследований кровотока в области таза и анкетирования обследуемых было установлено, что половая возбудимость мужчин и женщин практически одинакова, но существенно различается по предпочтениям. Определённые визуальные стимулы (например, изображения нагого тела, новые или непредсказуемые ситуации и позиции) нравятся мужчинам больше, чем женщинам. Женщины в общем предпочитают ситуации с эмоциональным, романтическим или знакомым им контекстом. Физическое состояние и интеллектуальное развитие человека также оказывают
Рис. 19-12. Сексуальная реакция [11]. М - у мужчин; Ж - у женщин.
1 - возбуждение; II - плато; III - оргазм; 1 - рефрактерный период;
2 - расслабление. Сплошные линии - стандартные реакции, прерывистые линии - варианты.
влияние как на порог возбудимости, так и на те или иные эротические стимулы.
• В течение фазы возбуждения сексуальный интерес стимулируют разнообразные психологические и физиологические факторы. Это возбуждённое состояние затем усиливается в течение фазы плато. Если стимуляция достаточна, наступает оргазм, или кульминация, проявляющаяся обычно как взрывоподобное и приятное освобождение от сексуального напряжения. Наконец, в течение фазы релаксации половое возбуждение рассеивается и физиологические параметры возбуждения возвращаются к исходному уровню. Хотя и мужчины, и женщины проходят через все стадии, существуют и различия в продолжительности и интенсивности любого отдельно взятого сексуального контакта. Наиболее значимое физиологическое различие между мужчинами и женщинами - наличие у мужчин рефрактерного периода, в течение которого половое возбуждение после эякуляции (оргазма) какое-то время не возобновляется и повторный оргазм невозможен. У женщин значительная стимуляция может вызвать оргазм или их серии в фазе релаксации.
• В ЦНС половое поведение (за счёт наличия рецепторов к половым стероидным гормонам) обеспечивают лимбическая система, терминальная полоска и преоптическая часть гипоталамической области. У обоих полов либидо поддерживается андрогенами. У женщин либидо обычно усиливается при ову-
ляции (за счёт эстрадиола) или непосредственно перед менструальным кровотечением.
• Половое поведение, включая его психологические и физиологические компоненты), относится не только к гетеросексуальному половому акту. Существует множество эквивалентов и полового поведения, и гетеросексуального полового акта. Например, ношение одежды противоположного пола (трансвестизм), фетишизм - использование фетиша (предмета или вещи, а также части тела, наделяемых эротическими свойствами) для сексуального удовлетворения, мастурбация, гомосексуальные отношения и др.
• Фундамент физиологического ответа человека на сексуальные стимулы образуют две составляющие: первичная реакция - увеличение кровоснабжения, вторичный ответ - мышечное напряжение. Рефлексы, замыкающиеся на уровне спинного мозга и модулируемые ЦНС, контролируют обе составляющие и обеспечивают эрекцию и эякуляцию у мужчин и оргазм у женщин.
Физиологические сексуальные реакции мужчин
Физиологические реакции мужской половой системы при половом акте складываются из эрекции, эмиссии и эякуляции.
Фаза возбуждения
Стандартный физиологический ответ мужчины на сексуальную стимуляцию - эрекция полового члена. В течение фазы возбуждения происходит расслабление ГМК трабекул кавернозных тел и артериол полового члена, что вызывает увеличение его размеров и отвердевание. Даже минимальная степень сексуального возбуждения может сопровождаться этой реакцией. Продолжительность этой фазы значительно варьирует.
Эректильные стимулы могут быть психогенными или соматогенными. В качестве психогенных могут выступать и фантазии, и явные визуальные стимулы (например, эротические изображения недвусмысленного содержания). Эти сигналы интегрируются в лимбической системе мозга и затем передаются по нисходящим путям в спинной мозг, где переходят на вегетативные парасимпатические эфференты, иннервирующие ГМК полового члена и выделяющие оксид азота. Соматогенные стимулы, например при-
косновения к половому члену или прилежащим к нему областям, будут рефлекторно активировать те же самые эфференты, идущие от спинного мозга. Эрекция, возникающая в фазе возбуждения, достаточно чувствительна к внешним сигналам. Эта фаза может прерваться. Так, внезапный громкий звук и изменения в окружающей обстановке могут нарушить эрекцию и прервать фазу воз- буждения. Эрекция может возникать и без реального сексуального возбуждения (например, во время сна, особенно у мальчиков в периоде пубертата).
Эрекция (рис. 19-13) - сложный нейрофизиологический процесс, приводящий к тому, что кровь, быстро притекая в половой член, как бы запирается в его губчатых хранилищах. В возникновении эрекции задействованы (i) кавернозные тела, (ii) вегетативная иннервация, (iii) кровоснабжение полового члена, а также (iv) афферентные и эфферентные нервные пути, спинной мозг и
ЦНС.
• Кавернозные тела. Хотя половой член содержит три эректильных образования, только два из них - кавернозные тела - непосредственно ответственны за отвердение этого органа при эрекции. Губчатое тело не приобретает достаточной твёрдости, хотя также становится более плотным. Последнее обстоятельство способствует перераспределению внутриуретрального давления. При этом просвет мочеиспускательного канала остаётся открытым, что способствует эффективному продвижению спермы при эякуляции. Основы физиологии эрекции станут более понятны, если рассматривать каждое кавернозное тело, как если бы это была одна полая камера (рис. 19-13, А). Завитковые артерии входят внутрь этой камеры, стенки которой содержат ГМК. Из этой полой камеры выходят мелкие венулы, которые собираются в более крупные, расположенные под белочной оболочкой. Эти подоболочечные венулы проходят через белочную оболочку и формируют выносящие вены.
• Иннервация ГМК. Когда половой член находится в спокойном состоянии, ГМК стенок этих полых камер сокращены. Это сокращение поддерживается адренергическими нервными волокнами симпатической нервной системы. При активации парасимпатических нервных волокон, идущих от крестцовой области спинного мозга по тазовыми нервам к половому чле-
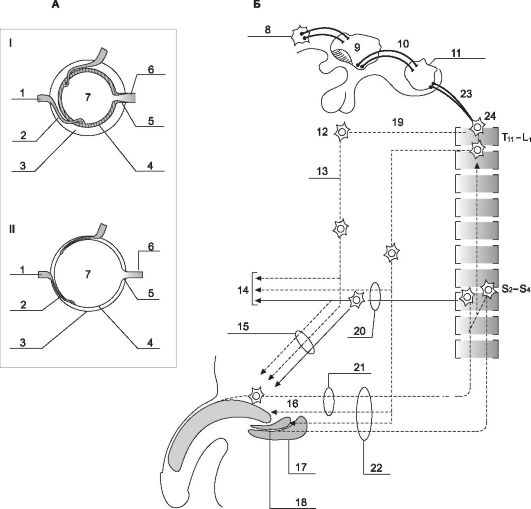
Рис. 19-13. Механизм эрекции [18]. А. Кровоснабжение полового члена и эрекция. I - расслабленное состояние полового члена; II - эрегированное состояние; Б. Нервная регуляция эрекции: 1 - огибающая вена, 2 - подоболочечная венула, 3 - белочная оболочка, 4 - трабекулярная гладкомышечная ткань, 5 - завитковая артерия, 6 - пещеристая артерия, 7 - полости пещеристых тел, 8 - миндалевидное тело, 9 - медиальное предоптическое поле, 10 - секреция дофамина, 11 - ядра шва, 12 - верхнее подчревное сплетение (α-адренергические нейроны), 13 - подчревный нерв, 14 - другие тазовые органы, 15 - пещеристый нерв, 16 - NO (оксид азота), VIP (вазоактивный интестинальный пептид), ацетилхолин, 17 - седалищно-пещеристая мышца, 18 - луковично-губчатая мышца, 19 - узлы симпатического ствола, 20 - тазовый нерв, 21 - дорсальный нерв полового члена, 22 - половой нерв, 23 - секреция серотонина, 24 - спинной мозг.
ну, адренергические влияния блокируются и ГМК стенок камер и артериол расслабляются. В это время по приносящим артериям кровь легко затекает внутрь камеры, что приводит к некоторому растяжению её стенок. В результате подоболочечные венулы и выносящие вены оказываются сдавлены растянутыми стенками камеры. Важно, что дальнейшее поступление крови в камеру приводит к ещё большему растягиванию её стенок и ещё более затрудняет отток крови из неё. Парасимпатические нервные волокна полового члена (в отличие от большинства других парасимпатических волокон), кроме ацетилхолина, выделяют VIP. В составе тазового нерва имеются также неадренергические и нехолинергические волокна, содержащие большое количество NO-синтетазы. Этот фермент катализирует образование мощного сосудорасширяющего вещества - оксида азота (NO). Оксид азота особенно сильно расслабляет артерии полового члена и трабекулярную сеть ГМК в эректильной ткани кавернозных и губчатого тел полового члена.
• Регуляция сократимости ГМК, расположенных в перегородках кавернозных тел и кровеносных сосудах полового члена, занимает центральную позицию в контроле эрекции (рис. 19-13, Б). Синусоидные пространства заполняются кровью при расслаблении ГМК за счёт активации парасимпатической нервной системы и одновременного ингибирования симпатических влияний. Парасимпатические влияния достигают полового члена по тазовым нервам, а симпатические - через подчревный нерв (n. hypogastricus).
• Нейромедиаторы. Несколько нейромедиаторов вовлечено в нервную модуляцию расслабления ГМК кавернозных тел, в то время как их сокращение зависит в основном от α-адренергических эффектов. Оксид азота - главный нейромедиатор, регулирующий эрекцию. Он локализуется вместе с VIP и ацетилхолином в нервных волокнах тазового нерва, заканчивающихся в трабекулах кавернозных тел и завитковых артериях. Норадреналин - главный антиэректильный агент.
• Силденафил (торговое название - «Виагра») - специфический ингибитор цГМФ-фосфодиэстеразы типа 5. Этот препарат применяют при эректильной дисфункции, но для проявления его эффекта необходима сексуальная стимуляция.
Силденафил повышает концентрацию цГМФ, вызывает расслабление ГМК эректильной ткани; усиливает эффект оксида азота, увеличивает кровоток в половом члене и предназначен для перорального приёма за 1-2 ч до полового сношения.
• Рефлекс эрекции вызывают афферентные сигналы: от чувствительных нервных волокон головки полового члена. Этот рефлекс замыкается на уровне спинного мозга. Афферентную часть рефлекса образует внутренний половой нерв, который может быть активирован тактильной стимуляцией кожи области промежности около мошонки. Эрекция может модулироваться супраспинальными влияниями со стороны ЦНС. Серотонинергические структуры ядра шва среднего мозга могут ингибировать эрекцию. Гипоталамическую медиальную преоптическую область и миндалевидное тело рассматривают как важные высшие центры регуляции эрекции.
• Тестостерон. Значение тестостерона в эректильной функции не известно. Ночные эрекции, возникающие в фазе сна с быстрыми движениями глазных яблок, не зависят от тестостерона; в то же время эрекции, появляющиеся в ответ на зрительную стимуляцию, не возникают у мужчин с гипогонадизмом.
• Слизистый секрет. Во время половой стимуляции парасимпатические эфференты (дополнительно к вызываемой ими эрекции) побуждают выделение слизистого секрета железами мочеиспускательного канала и бульбоуретральных желёз. Эта слизь смазывает половые пути, облегчая фрикционные движения пениса во время полового акта. Однако основная часть смазочного материала во время коитуса обеспечивается женскими половыми органами. Без достаточной смазки половой акт бывает редко полноценным из-за сильных болевых ощущений, тормозящих возбуждающие сексуальные ощущения.
Плато
Эрекция. В течение фазы плато эрекция несколько усиливается за счёт ещё большего кровенаполнения пещеристых тел полового члена, внутрисосудистое давление в полостях эректильной ткани достигает значений, характерных для системного систолического АД. Мошонка и яички рефлекторно подтягиваются к промежности. Размеры яичек несколько увеличиваются из-за их возросшего кровенаполнения. В этой фазе также наблюдают возрастание ЧСС
и систолического АД. При достижении фазы плато расслабление полового члена без эякуляции и оргазма у здоровых мужчин происходит очень редко.
Эмиссия. В конце фазы плато рефлекторно возбуждаются центры спинного мозга на уровне Th12-L2. Это возбуждение прово- дится к половым органам через подчревные и тазовые симпатические нервные сплетения и приводит к эмиссии - процессу активации ГМК семенных пузырьков, семявыносящего протока и предстательной железы, в результате чего содержимое этих органов поступает в простатическую часть уретры. Эмиссия опосредуется адренергическими симпатическими нервными волокнами и α-адренорецепторами.
Мужской оргазм
Наполнение внутренней части уретры спермой порождает сенсорные сигналы, передаваемые через половые нервы в крестцовый отдел спинного мозга и вызывающие ощущение переполнения внутренних половых органов. Эти же сенсорные сигналы возбуждают ритмические сокращения внутренних половых органов и вызывают сокращение луковично-пещеристой и седалищно-пещеристой мышц, сдавливающих в основании полового члена эректильные ткани. Непосредственно перед эякуляцией на коже верхней части живота, груди, шеи и лица может появиться гиперемия. Общий тонус мышц тела становится максимальным. В течение оргазма изменения со стороны сердечно-сосудистой системы и общий мышечный тонус достигают максимума. Возможны гипервентиляция и звуковые реакции (вокализация). При сокращении ГМК мочеиспускательного канала, поперечнополосатых луковичнопещеристой и седалищно-пещеристой мышц эякулят выбрасывается из уретры во влагалище. Мышцы тазового дна и сфинктер прямой кишки также могут ритмично сокращаться. Ритмические сокращения мышц туловища вонзающими движениями таза и полового члена помогают сперме глубже проникать во влагалище. Этот момент эякуляции обычно и называют мужским оргазмом.
Релаксация
Эякуляция сопровождается снятием сексуального напряжения и выраженным чувством удовольствия. Половое возбуждение мужчины почти полностью угасает в течение 1-2 мин, эрекция прекращается. Фаза релаксации и размягчение полового члена - это
две раздельные стадии. Сначала достаточно быстро уменьшаются размеры полового члена. При этом половой член уменьшается до размера, превышающего его спокойное состояние примерно на 50%. На этой стадии половой член абсолютно нечувствителен к стимуляции. Затем наступает вторая стадия, во время которой половой член возвращается к своим исходным размерам в нестимулированном состоянии. В это время половой член только частично нечувствителен к стимуляции, но постепенно вновь становится чувствительным к новой, более значительной стимуляции. Пролонгирование первой фазы размягчения полового члена наблюдают, если фазы возбуждения или плато сексуального цикла сознательно удлиняют усилием воли. Так поступают некоторые мужчины для того, чтобы задержать наступление эякуляции до тех пор, пока половая партнёрша не будет удовлетворена.
Физиологические сексуальные реакции женщин
Возбуждение
Во время фазы возбуждения соматогенные и психогенные стимулы приводят женщин в состояние возбуждения по тем же нервным путям, что и мужчин (преимущественно парасимпатическим). Ответ эректильной ткани входа во влагалище (в том числе и клитора) на возбуждение выражен слабее, чем у его мужского гомолога - полового члена. Тактильная стимуляция промежности женщины или головки клитора может вызвать кровенаполнение и набухание тела клитора и его эрекцию только у некоторых женщин.
Реакция влагалища на возбуждение наиболее постоянна и хорошо выражена. Она заключается в увлажнении влагалища за счёт активации секреции бартолиновых желёз и транссудации жидкости через слизистую оболочку. Этот процесс начинается через 10-30 с после начала возбуждающих стимулов и продолжает прогрессировать до оргазма. Пролонгирование фазы возбуждения и фазы плато вызывает более сильную влагалищную секрецию. В фазе возбуждения верхние две трети влагалища удлиняются, что способствует некоторому смещению матки вверх. Это смещение способствует поднятию шейки матки выше сводов влагалища и увеличению диаметра влагалища, в основном на уровне шейки. Наконец в ходе фазы возбуждения малые половые губы становят-
ся заметно
набухшими в результате переполнения их кровью. Это приводит к тому, что
малые половые губы «раздвигают» большие половые губы в стороны от
входа во влагалище. Увеличение размеров малых половых губ добавляет как
минимум
Плато
В течение фазы плато наиболее выраженные изменения в гениталиях женщины - гиперемия малых половых губ в результате их кровенаполнения. Этот свекольно-красный цвет - существенный физический маркёр полового возбуждения. Клитор втягивается внутрь «капюшона», формируемого малыми половыми губами. Гипервентиляция, учащённый пульс и повышенное АД также сопутствуют позднему отрезку фазы плато, но у женщин они не так сильно выражены, как у мужчин. В то же время генерализованная миотония может дойти до спастических сокращений поперечнополосатых мышц предплечий и голеней.
Проникновение полового члена или иного объекта во влагалище во время полового сношения усиливает сексуальное возбуждение женщины путём непрямого стимулирования втягивающегося клитора. Это происходит из-за его подтягивания наполненными кровью малыми половыми губами, спаянный передний сегмент которых образует «капюшон» клитора. Прямой пролонгированный контакт с клитором может вызвать очень сильную стимуляцию, так как головка клитора исключительно чувствительна в стадии возбуждения.
Женский оргазм
Оргазм у женщин, как и у мужчин, включает ритмические сокращения репродуктивных органов с последующим снятием крове- наполнения и мышечного тонуса, возникших в течение фазы возбуждения. В типичных случаях сокращения начинаются с нижней трети влагалища и затем охватывают всё влагалище и матку. Гиперемия кожи и диффузное потоотделение могут распространиться на всё тело. Помимо эякуляции, оргазм у мужчины и женщины имеет два существенных физиологических различия. Во-первых, женщина способна испытать несколько следующих друг за другом оргазмов (если происходит адекватная сексуальная стимуляция в то время, пока женщина не вышла из фазы плато полового воз-
буждения). Во-вторых, оргазм у женщины может продолжаться значительно дольше.
Релаксация
Релаксация включает расслабление половых губ (отток крови), размягчение клитора, а затем и расслабление влагалища.
ОПЛОДОТВОРЕНИЕ
Оплодотворение - слияние мужской и женской гамет, приводящее к образованию зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы; при этом сливаются их ядра (пронуклеусы), объединяются хромосомы и возникает первая диплоидная клетка нового организма - зигота. Начало оплодотворения - момент слияния мембран сперматозоида и яйцевой клетки, окончание оплодотворения - момент объединения материала мужского и женского пронуклеусов. Всё, что происходит до слияния мембран сперматозоида и яйцевой клетки, именуют процессами, предшествующими оплодотворению.
Сперматозоид
В эякуляте человека содержится примерно 3х108 сперматозоидов.
• Подвижность. В женских половых путях сперматозоиды: активно передвигаются со скоростью 1-4 мм/мин и сохраняют способность к перемещению и оплодотворению максимально до 2 сут. Примерно 200 сперматозоидов достигает воронки маточной трубы, где сперматозоид встречается с яйцеклеткой. Сперматозоиды, не участвующие в оплодотворении, удаляются из женских половых путей или перевариваются фагоцитами. Поскольку в клеточной мембране сперматозоидов существуют рецепторы к агентам хемотаксиса - N-формилпептидам, не исключено, что эти подвижные клетки перемещаются направленно, путём хемотаксиса.
• Капацитация. До того как сперматозоид встретится с яйцеклеткой, он в течение нескольких часов продвигается по женским половым путям. При этом на сперматозоид воздействуют факторы женского организма (pH, слизь и др.), не только поддерживающие способность к миграции и оплодотворению,
но и активирующие их. Этот процесс называют капацитацией. В ходе капацитации мембрана сперматозоида становится проницаемой для ионов Ca2+, которые, войдя в сперматозоид, усиливают моторику жгутика, а также вызывают слияние мембраны акросомы и сперматозоида (акросомная реакция, см. ниже).
• Простагландины, содержащиеся в эякуляте, вызывают антиперистальтические сокращения ГМК миометрия и маточных труб, облегчая продвижение сперматозоидов в просвете маточных труб.
Яйцеклетка
Большинство яйцеклеток в яичниках находится на стадии поздней диплотены профазы первого деления мейоза. В ходе овуляции при разрыве стенки зрелого фолликула из яичника в брюшную полость выбрасывается овоцит второго порядка. Его окружает прозрачная оболочка (zona pellucida) и состоящий из фолликулярных клеток лучистый венец (corona radiata).
• Перемещение яйцеклетки. Перед овуляцией увеличение тонуса ГМК маточной трубы приближает воронку маточной трубы к поверхности яичника. Биение ресничек эпителиальных клеток, выстилающих фимбрии ампулы маточной трубы, создаёт ток жидкости, способствующий перемещению яйцеклетки из брюшной полости в ампулу.
• Способность к оплодотворению. Для успешного оплодотворения яйцеклетка должна встретиться со сперматозоидом в течение 1 сут после овуляции. Из практических соображений время, в течение которого овулировавшая яйцевая клетка может оплодотвориться, оценивают максимально в 5 сут.
Механизмы оплодотворения
Для того чтобы произошло оплодотворение, сперматозоид должен последовательно преодолеть три барьера: лучистый венец (corona radiata), состоящий из нескольких слоёв фолликулярных клеток, прозрачную оболочку (zona pellucida) и наконец плазматическую мембрану яйцеклетки, при слиянии которой с плазмолеммой сперматозоида и начинается собственно оплодотворение.
Акросомная реакция
Ферменты акросомы предназначены для разрушения прозрачной оболочки и преодоления сперматозоидом этого барьера. При акросомной реакции мембрана акросомы и клеточная мембрана сливаются и формируют мелкие пузырьки, отделяющиеся от головки сперматозоида. При этом из акросомы освобождаются гиалуронидаза, протеазы, гликозидазы, липазы, нейраминидаза и фосфатазы. Эти ферменты расщепляют молекулы прозрачной обо- лочки, что позволяет сперматозоиду преодолеть этот барьер.
Механизм акросомной реакции. Взаимодействие рецепторов zona pellucida с цитолеммой сперматозоида вызывает массированный транспорт внутрь головки сперматозоида Ca2+ и Na+ в обмен на H+. Увеличение внутриклеточной концентрации Ca2+ активирует Са2+-зависимую фосфолипазу, в результате изменяется уровень вторых посредников - циклических нуклеотидов. Вслед за этим активируется протонная АТФаза, что приводит к увеличению вну- триклеточного рН. Увеличение концентрации Ca2+ в цитозоле и повышение pH в головке сперматозоида запускают акросомную реакцию.
Оплодотворение
В результате акросомной реакции в прозрачной оболочке образуется узкий канал, через который проходит сперматозоид; плазматические мембраны сперматозоида и яйцеклетки вступают в соприкосновение и сливаются. Вскоре слившиеся мембраны разрушаются, а головка сперматозоида оказывается погружённой в цитоплазму яйцеклетки. В течение первых 12 ч после проникновения сперматозоида в яйцеклетку происходит перестройка ядер (пронуклеусов) слившихся гамет. Пронуклеусы мигрируют в центр яйцеклетки и сближаются. Их ядерные оболочки исчезают, а материнские и отцовские хромосомы перемешиваются - образуется синкарион. Этот процесс (сингамия) и есть собственно оплодотворение - появляется диплоидная зигота (новый организм, пока одноклеточный). Роль сперматозоида в оплодотворении не исчерпывается только тем, что половина хромосом диплоидной зиготы - отцовские; новый организм получает часть генетической информации (митохондриальный геном) вместе с митохондриями сперматозоида. Сперматозоид приносит также сигнальный белок дробления. В ходе оплодотворения происходит также завершение
мейоза и детерминируется генетический пол нового организма. Сразу после того как сливаются плазматические мембраны гамет и объединяются ядерные геномы, существенно изменяется внутриклеточный ионный состав, вследствие чего уменьшается объём зиготы, деполяризуется её плазмолемма, развивается кортикальная реакция.
• Уменьшение объёма зиготы. Прямое следствие этого процесса - образование перивителлинового пространства. Перивителлиновое пространство содержит оба полярных тельца и имеет характерный ионный состав (гомеостатическая среда для предэмбриона), создающий дополнительное препятствие для проникновения к зиготе других сперматозоидов.
• Деполяризация плазматической мембраны предупреждает проникновение других сперматозоидов в зиготу.
• Кортикальная реакция. По периферии яйцеклетки расположены кортикальные гранулы. Их диаметр 200-600 нм; они содержат ферменты, в том числе различные гидролазы. Сразу после проникновения сперматозоида в яйцеклетку начинается кортикальная реакция - выделение содержимого кортикальных гранул в перивителлиновое пространство. Сигнал для кортикальной реакции - повышение концентрации Ca2+ в цитозоле. Под действием ферментов кортикальных гранул происходят протеолиз и модификация рецептора сперматозоида, который теряет способность к запуску акросомной реакции. Тем самым блокируется полиспермия. В результате кортикальной реакции прозрачная оболочка подвергается существенным изменениям - происходит её стабилизация (образуется оболочка оплодотворения). В стабилизированном состоянии прозрачная оболочка защищает предэмбрион, проходящий по маточной трубе. Без прозрачной оболочки дробление зиготы невозможно.
• Обобщение главы
В яичках лютеинизирующий гормон (ЛГ) контролирует синтез тестостерона в клетках Лейдига, а фолликулостимулирующий гормон (ФСГ) увеличивает образование андрогенсвязанного белка, ингибина и эстрогена в клетках Сертоли.
Сперматозоиды образуются в извитых семенных канальцах обоих яичек. Сперматозоид развивается из сперматогоний через ряд последовательных стадий, включая сперматоциты и сперматиды.
Сперма созревает и накапливается в придатках яичка. Во время эякуляции за счёт мышечных сокращений придатков и семявыно- сящих протоков сперма продвигается через семявыносящие протоки в мочеиспускательный канал в области простаты. В конечном счёте сперма выбрасывается из организма через уретру в половой член.
Гипоталамический гонадолиберин контролирует секрецию ЛГ и ФСГ передним отделом гипофиза.
Тестостерон в основном уменьшает секрецию ЛГ, а ингибин - секрецию ФСГ. Тестикулярные гормоны, таким образом, осуществляют отрицательную обратную связь с гипоталамо-гипофизарной системой.
Андрогены оказывают влияние на несколько органов-мишеней: мышечную, костную и другие системы - и регулируют развитие вторичных половых признаков, либидо и сексуального поведения.
Циклическая секреция гипоталамического гонадолиберина регулирует секрецию ЛГ и ФСГ, которые способствуют развитию фолликулов, стероидогенезу, овуляции и образованию жёлтого тела.
ЛГ и ФСГ в координации с яичниками регулируют секрецию фолликулярного эстрадиола.
Овуляция происходит в результате положительной обратной связи фолликулярного эстрадиола с гипоталамо-гипофизарной системой, которая стимулирует выход ЛГ и ФСГ.
Развитие фолликула осуществляется в несколько этапов: примордиальном, первичном, вторичном, третичном и стадии граафова пузырька.
Для образования функционирующего жёлтого тела необходимо выделение ЛГ, адекватное количество ЛГ-рецепторов, достаточное количество гранулёзных клеток и значительная секреция прогестерона.
Эстрадиол и прогестерон регулируют маточный цикл. Эстрадиол индуцирует пролиферацию эндометрия матки, в то время как прогестерон индуцирует дифференцировку эндометрия и секрецию различных продуктов.
Во время полового созревания гипоталамус секретирует повышенное количество гонадолиберина, которое увеличивает секре- цию ЛГ и ФСГ, активирует функцию яичников и вызывает первую овуляцию.
Менопауза возникает вследствие истощения яйцеклеток в яичнике и последующего нарушения в развитии фолликулов и паде- ния секреции эстрадиола. Уровень секреции ЛГ и ФСГ возрастает в результате утраты отрицательной обратной связи от эстрадиола.