Нормальная физиология: учебник / Орлов Р.С., Ноздрачев А.Д. - 2-е изд., исправл. и доп. 2010. - 832 с.
|
|
|
|
ГЛАВА 12. ОБОНЯНИЕ И ВКУС
Обоняние и вкус относятся к висцеральным чувствам, поскольку они в значительной степени связаны с функцией пищеварения (например, ароматы пищи обычно сочетаются с её вкусом) и дыхания. Воспринимающие структуры органов обоняния и вкуса - хеморецепторы, они возбуждаются молекулами вкусовых веществ и одорантами.
Обоняние
Обонятельные пути начинаются от рецепторных клеток слизистой оболочки обонятельной области и проецируются в обонятель- ный мозг, они не имеют ни переключения в таламусе, ни прямого представительства в коре больших полушарий.
Обонятельная слизистая оболочка
Воспринимающие обонятельные структуры (периферическая часть обонятельного анализатора, орган обоняния) образуют в слизистой оболочке носа специализированную область - парную обонятельную выстилку (обонятельное поле), расположенную под решётчатой пластинкой. Суммарная площадь обонятельного поля (обонятельного эпителия) около 5 см2.
Обонятельный эпителий (рис. 12-1) содержит опорные эпителиальные клетки и 10-20 млн расположенных между ними рецеп- торных обонятельных клеток - биполярных обонятельных нейронов. Короткий и толстый дендрит (периферический отросток, направленный к поверхности обонятельной выстилки) каждого рецепторного нейрона имеет расширенный конец - обонятельную булаву. От булавы отходит 8-40 тонких обонятельных волосков - ресничек. Именно в эти погружённые в слизь волоски «вмонтированы» обонятельные молекулярные рецепторы. От ба-
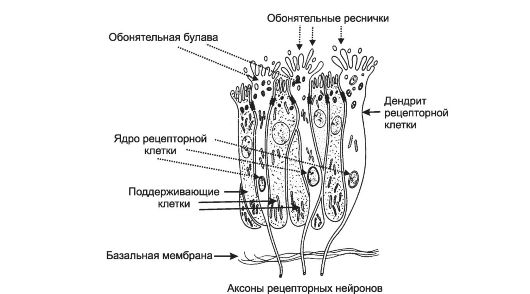
Рис. 12-1. Строение обонятельного эпителия.
зальной части рецепторного нейрона отходит его центральный отросток - аксон. Пучки этих аксонов в виде обонятельных нитей (fila olfactoria) пронизывают подэпителиальную соединительную ткань, костную решётчатую пластинку и входят в обонятельные луковицы. Под эпителием и в самой эпителиальной выстилке расположено множество обонятельных (боуменовых) желёз. Секретируемая ими слизь покрывает поверхность обонятельной выстилки. Слизь содержит воду, гликозаминогликаны, АТ, связывающие молекулы одорантов белки, ферменты и полностью обновляется в течение 10 мин.
Втягивание воздуха. Область, содержащая обонятельные рецепторы, плохо вентилируется. Обычно воздух спокойно движется над носовыми раковинами во время каждого дыхательного движения. Количество воздуха, достигающего обонятельной области, заметно возрастает при интенсивном втягивании воздуха (принюхива- ние). Это происходит за счёт сокращения нижней части ноздрей, прижимающихся ближе к носовой перегородке и помогающих отклонять струю воздуха вверх. Втягивание воздуха - полурефлекторный акт, возникающий в случаях, когда внимание привлечено новым запахом.
Болевые рецепторы. В обонятельной слизистой оболочке расположено много свободных нервных окончаний, имеющихся в волокнах тройничного нерва. Эти ноцирецепторы стимулируются раздражающими веществами, и этот раздражающий компонент - часть характеристики «запаха» ряда веществ (аммиак, хлорная из- весть и др.). Другими словами, эти рецепторы боли ответственны за чиханье, слёзы, задержку дыхания и другие рефлексы, вызванные раздражением слизистой оболочки носа.
Восходящие пути и обонятельный мозг
Обонятельные нервы (пучки аксонов рецепторных обонятельных клеток) в виде 18-20 тонких ветвей вступают в обонятельную луковицу. Последняя переходит в обонятельный тракт, заканчивающийся в первичных обонятельных центрах (обонятельный треугольник, переднее продырявленное вещество, прозрачная перегородка). От первичных обонятельных центров сигналы направляются к корковым центрам обоняния своей (ипсилатеральной) и противоположной (контралатеральной) стороны - к извилине гиппокампа и крючку.
Обонятельный мозг (rhinencephalon) образуют обонятельная луковица, обонятельный тракт, обонятельный треугольник, переднее продырявленное вещество, гиппокампова борозда, зубчатая извилина и ряд прилежащих структур.
Обонятельная луковица
В обонятельной луковице (рис. 12-2) аксоны рецепторных клеток образуют синапсы с дендритами митральных и пучковых клеток, формируя характерные комплексы - обонятельные клубочки. В каждый клубочек входит (конвергирует) в среднем 25 000 аксонов рецепторных клеток, но не любых, а только тех, что имеют идентичные обонятельные рецепторы. В каждом обонятельном клубочке с аксонами рецепторных клеток контактируют дендриты примерно 25 митральных и 60 пучковых клеток. В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют реципрокные синапсы. Эти синаптические связи осуществляют контроль за ис- ходящей из обонятельной луковицы информацией, вероятно, закодированной в спектрах ПД.
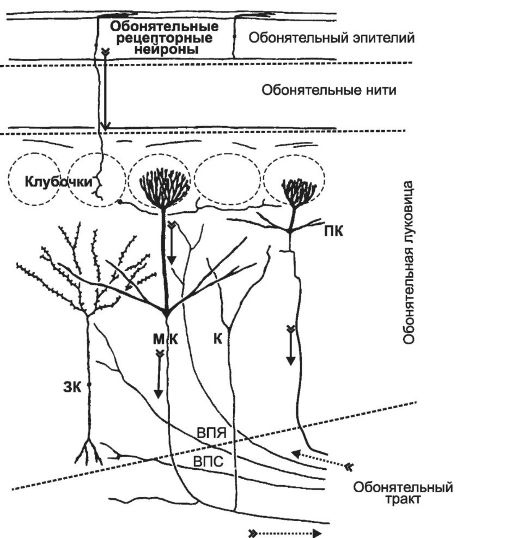
Рис. 12-2. Архитектоника нейронов и связей в обонятельной луковице. ПК -
пучковая клетка, МК - митральная клетка с возвратными коллатералями (К), ЗК - зернистая клетка, ВПЯ - нисходящие в обонятельную луковицу нервные волокна из переднего обонятельного ядра, ВПС - комиссуральные нервные волокна передней спайки мозолистого тела (commissura anterior). Прерывистые линии - границы между анатомическими структурами.
Одоранты
Молекулы, создающие запахи (одоранты), исчисляются миллионами. Они имеют небольшие размеры и содержат от 3-4 до 18-20 атомов углерода. Молекулы однородного химического состава, но с неодинаковой конфигурацией обладают различным запахом. Принято различать следующие основные запахи: мятный, едкий, гнилостный, эфирный, мускусный, камфорный и цветочный. В реальной жизни человек встречается со смесями запахов. Обо- нятельные рецепторы контактно реагируют только на вещества, растворённые в тонком слое слизи на поверхности обонятельного эпителия. У порогов восприятия разных одорантов значителен
диапазон величин - от 5,8 мг в литре воздуха (этиловый эфир) до 0,5 нг/л (метилмеркаптан). Это значит, что чувствительность различается в 10 млн раз. Человек различает от 2000 до 4000 тыс. различных запахов, но хуже регистрирует их концентрацию (чтобы обнаружилась разница, концентрация пахучего вещества должна измениться не менее чем на 30%).
Регистрация и преобразование обонятельного сигнала
Рецепторные клетки обонятельной выстилки регистрируют небольшое количество первичных запахов, но их комбинации фор- мируют ощущение многих и многих воспринимаемых запахов. Возникает вопрос: каким образом обонятельная система может различать множество различных запахов? Варианты ответа таковы: или рецепторные обонятельные нейроны имеют множество различных молекулярных рецепторов (в том числе и каждый нейрон), или/и они содержат один или несколько типов молекулярных обонятельных рецепторов, но посылают в ЦНС различные спектры ПД. Наконец, ощущение конкретного запаха может формироваться в зависимости от фиксированных связей между нервными клетками обонятельной системы.
Обонятельные рецепторные белки кодируют гены, расположенные практически во всех хромосомах, кроме аутосомы 20 и половой хромосомы Y. В геноме человека идентифицировано более 900 генов, кодирующих белки обонятельных рецепторов, что практически равно одной тридцатой всего генома. Эти белки относятся к семейству связанных с G-белком рецепторов.
Последовательность событий при регистрации обонятельного сигнала (рис. 12-3) можно представить следующим образом: взаимодействие пахучего вещества с белком-рецептором в плазмолемме обонятельных волосков - активация G-белка - повышение активности аденилатциклазы - увеличение уровня цАМФ - активация цАМФ-зависимых воротных катионных каналов - деполяризация рецепторных нейронов - генерация ПД и его проведение по аксону.
Механизмы адаптации. Адаптация к обонятельному сигналу на уровне рецепторных нейронов (регуляция чувствительности к обонятельному сигналу) происходит достаточно быстро (50% в течение 1 с). Некоторые механизмы такой быстрой адаптации известны и осуществляются при помощи внутриклеточных вторых
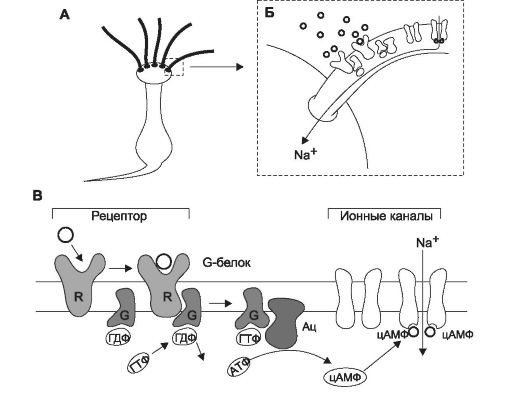
Рис. 12-3. Трансформация сигнала в обонятельном рецепторе. А - рецеп- торный обонятельный нейрон; Б - вход Na+ в клетку; В - пахучие молекулы соединяются с рецептором (R). Рецептор активирует G-белок (G), G-белок активирует аденилатциклазу (Ац), образующийся цАМФ открывает Na+-каналы.
посредников и β-аррестинов - веществ, десенситизирующих адренорецепторы и тормозящих функцию рецепторов, которые связаны с G-белками.
Центральные механизмы
Пребывание в условиях воздействия даже очень неприятного запаха уменьшает и может полностью подавить восприятие запаха. Этот феномен - результат быстрой адаптации или десенситизации на уровне рецепторных нейронов (см. выше). При этом порог для других запахов не изменяется. В центральных обонятельных структурах также существует чёткий контроль обонятельной афферентной импульсации. Так, возбуждение нервных клеток, вызванное афферентной импульсацией, сменяется последующим торможением электрической активности нейронов, что и объясняет суще-
ствование ритмической активности в обонятельной коре. Пример, демонстрирующий центральную регуляцию потоков импульсов от обонятельной луковицы, приведён выше. Обонятельная импульсация направляется в лимбическую систему (здесь осуществляется подсознательное восприятие обоняния), к передним обонятельным ядрам, миндалевидному телу (формирование эмоционального ответа на обонятельные стимулы) и обонятельной борозде (формирование так называемой обонятельной памяти).
Обоняние, половое поведение и память. Между запахом и половой функцией у многих видов животных существует тесная взаимосвязь [скорее всего, реализуемая через cошниково-носовой (вомероназальный) орган Якобсона - участок слизистой оболочки носовых ходов, аналогичный обонятельной выстилке], а использование духов даёт достаточные основания считать, что подобная взаимосвязь существует и у людей (орган Якобсона у человека отсутствует). Обоняние у женщин развито сильнее, и оно обостряется ещё больше в период овуляции. Запах и (в меньшей степени) вкус обладают уникальной способностью оживлять воспоминания, заложенные в долговременной памяти. Этот факт отмечен писателями и экспериментально доказан психологами.
Комбинаторное кодирование. Эта концепция подразумевает, что каждый из миллионов отдельных запахов (одорантов) имеет уникальный код; кодирование происходит на уровне органа обоняния, а декодирование - в обонятельных центрах.
Вкус
Рецепторы вкуса - контактные, а пути проведения от них проходят через ствол мозга к таламусу и проецируются вдоль постцен- тральной извилины. Периферическая часть вкусового анализатора - вкусовые почки - расположена в слизистой оболочке полости рта, переднего отдела глотки, пищевода и гортани. Основная масса вкусовых почек (>90% их общего количества - до 10 000) находится в хемочувствительных сосочках языка (рис. 12-4) - листовидных, грибовидных и желобоватых.
Вкусовые почки (вкусовые луковицы) - тельца яйцевидной формы со средним размером 50-70 мкм, состоят из клеток различных типов. На верхушечной (апикальной) части большинства клеток расположены микроворсинки - вкусовые волоски, занимаю-
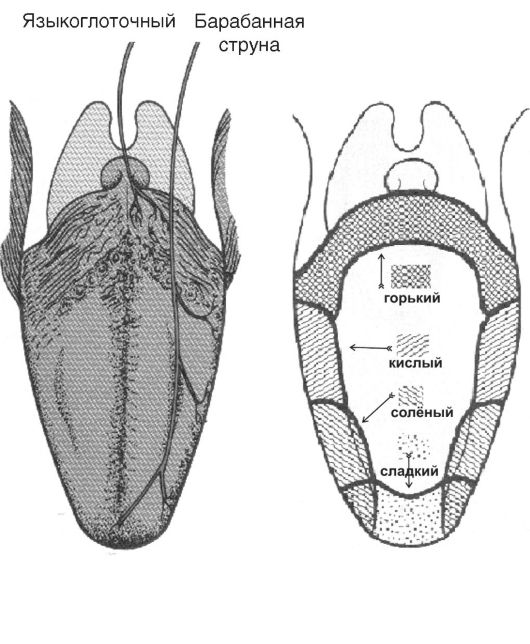
Рис. 12-4. Вкусовые области языка. А - иннервация языка; Б - вкусовые зоны языка. Вкус сладкого регистрируется преимущественно на кончике языка, солёного - ближе к кончику языка, кислого - на боковых сторонах языка, горького - в задней части языка и в мягком нёбе.
щие вкусовой канал, который открывает на поверхность эпителия вкусовой порой. В микроворсинки «вмонтированы» рецепторные вкусовые белки. В базальной части вкусовой луковицы рецепторные клетки образуют синапсы с терминальными разветвлениями периферических отростков первичных чувствительных нейронов. Каждая вкусовая почка иннервирована примерно 59 нервными волокнами, а каждый чувствительный нейрон получает сигналы при- мерно от пяти вкусовых луковиц.
Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1) первичного чувствительного нейрона; 2) нервной клетки
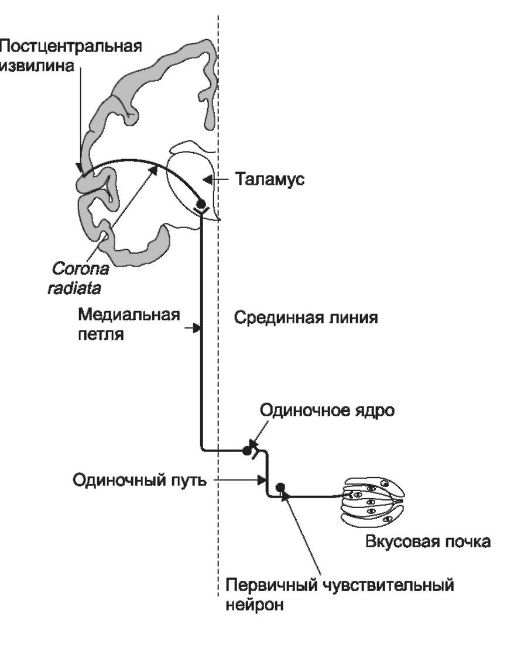
Рис. 12-5. Восходящие пути вкусовой чувствительности.
Часть аксонов одиночного пути направляется к верхнему и нижнему слюнным ядрам, что обеспечивает выполнение некоторых рефлексов, связанных со слюноотделением.
ядра одиночного пути (одиночное ядро, nucleus tractus solitarius); 3) нейрона вентромедиального ядра таламуса (рис. 12-5).
Основные вкусы. Человек различает четыре первичных вкуса (сладкий, кислый, горький и солёный), а также «умами» (от японского «изысканный», вкус глутамата натрия). Существуют карты топографической привязанности преимущественного восприятия основных вкусов (см. рис. 12-4, А), но необходимо иметь в виду, что такие карты действительны только для низких концентраций тестируемых вкусовых веществ. Так, при низких концентрациях деполяризуются и формируют рецепторный (генераторный) потенциал лишь отдельные вкусовые клетки. Некоторые вкусовые клетки и вкусовые луковицы отвечают преимущественно на горькие стимулы, в то время как другие - на сладкие, кислые или солёные. Часть вкусовых клеток и луковицы в целом отвечают на две или три, а иные даже на все вкусовые модальности. Если же концентрация вкусовых веществ увеличивается, происходит воз-
буждение практически всех вкусовых луковиц самой различной локализации.
Классы вкусовых веществ и основные вкусы
Существует некоторая связь между ионами и молекулами вкусовых веществ и вызываемыми ими первичными вкусовыми ощу- щениями сладкого, кислого, горького и солёного.
• Ощущение кислого создают кислоты. Интенсивность ощущения пропорциональна логарифму концентрации водородных ионов (H+).
• Ощущение солёного вкуса формируется при воздействии катионов ионизированных солей.
• Ощущение сладкого вызывают сахара, гликоли, спирты, альдегиды и некоторые другие вещества органической природы. Сахароза, мальтоза, лактоза и глюкоза - типичные примеры. Однако существует большой круг веществ иной природы (белок мореллин, искусственные подсластители - сахарин, аспартам и цикламат, а также хлороформ, соли бериллия), вызывающих ощущение сладкого, причём в некоторых случаях в концентрациях, которые в десятки и сотни тысяч раз меньше, чем у классической глюкозы.
• Ощущение горького характерно для длинноцепочечных азотсодержащих органических соединений и алкалоидов (например, хинина, кофеина, никотина, стрихнина), а также для некоторых неорганических соединений (соли магния, кальция, аммония).
Вкусовой порог. Наиболее
значительные различия наблюдаются между веществами, создающими
ощущение горького и всех остальных первичных вкусов. Так, ощущение
солёного возникает при воздействии вещества в концентрации
кий, 10% раствор поваренной соли - как максимально солёный и т.д.
Адаптация к воздействию вкусового вещества развивается мед- ленно (минуты) и пропорциональна его концентрации. Адаптация к сладкому и солёному развивается быстрее, чем к горькому и кислому.
Механизмы возбуждения вкусовых клеток
Последовательность событий между воздействием вкусового вещества на вкусовые рецепторные клетки и генерацией ПД в пери- ферических нервных отростках первичного чувствительного нейрона в общем виде можно представить следующим образом:
вкусовое вещество - молекулярный вкусовой рецептор вкусовых волосков - вторые посредники - деполяризация клеточной мембраны и генерация рецепторного потенциала - поступление Ca2+ в цитоплазму из межклеточного пространства и/или внутриклеточных кальциевых депо - экзоцитоз синаптических пузырьков из базальной части клетки - постсинаптический потенциал - генерация ПД.
Промежуточные и финальные этапы процесса (от деполяризации клеточной мембраны до генерации ПД в чувствительных нервных волокнах) практически одинаковы для всех вкусовых клеток. В то же время начальные этапы значительно различаются при воздействии веществ, вызывающих формирование различных первичных вкусов. Более того, отдельные этапы процесса восприятия отсутствуют или даже не известны. Ниже рассмотрены последова- тельные этапы хемовосприятия и электрогенеза, происходящие в рецепторных вкусовых клетках при формировании разных вкусовых ощущений.
• Солёное. Основное действующее начало - Na+. Молекулярный рецептор не обнаружен. В клеточной мембране вкусовой клетки имеются многочисленные ионные каналы, в том числе селективные натриевые (чувствительные к амилориду) и потенциалзависимые кальциевые каналы. При воздействии ионизированных солей, т.е. при увеличении содержания внеклеточного Na+, этот катион входит в рецепторную клетку через натриевые каналы. Результат увеличения внутриклеточной концентрации Na+ - деполяризация клеточной мембраны
(появление рецепторного потенциала). Следствие деполяризации клеточной мембраны - активация кальциевых каналов, вход в цитоплазму рецепторной клетки внеклеточного Ca2+, который и запускает слияние синаптических пузырьков с пресинаптической мембраной и дальнейшие события по приве- дённой выше схеме.
• Кислое. Действующее начало - протоны (H+). Молекулярный рецептор не обнаружен. В клеточной мембране вкусовой клетки имеются многочисленные амилоридчувствительные натриевые каналы и К+-селективные каналы. Протоны H+ входят в клетку через натриевые каналы (что вызывает деполяризацию клеточной мембраны) и одновременно блокируют калиевые каналы (что усиливает степень деполяризации мембраны). Дальнейшие события развиваются по приведённой выше схеме.
• Сладкое на примере глюкозы как действующего начала. В мембрану вкусовой клетки (особенно в мембрану вкусовых волосков) «вмонтированы» молекулярные вкусовые рецепторы, регистрирующие наличие молекул сладкого. Эти рецепторы относятся к связанным с G-белком рецепторам. Взаимодействие глюкозы с белком-рецептором активирует G-белок; повышается активность аденилатциклазы и увеличивается содержание цАМФ, что приводит к блокаде К+-селективных каналов и деполяризации клеточной мембраны. Дальнейшие события развиваются по приведённой выше схеме.
• Горькое. Некоторые молекулы: непосредственно блокируют К+-селективные каналы (в результате происходит деполяризация клеточной мембраны). Другие молекулы горького взаимодействуют с молекулярными вкусовыми рецепторами. В мембрану вкусовой клетки вмонтировано несколько типов таких рецепторов. Эти рецепторы относятся к связанным с G-белком рецепторам. В части G-белков их α-субъединица представлена специфичным для вкусовых клеток гастдуцином. При активации такого G-белка происходит активация фосфодиэстеразы, что влечёт за собой уменьшение внутриклеточной концентрации цАМФ, в итоге приводящее к деполяризации клеточной мембраны, входу Ca2+ в клетку и к выбросу содержимого синаптических пузырьков. При взаимодействии молекул горького со вкусовыми рецепторами
• другого типа активируется G-белок, что вызывает активацию фосфолипазы C, увеличение внутриклеточной концентрации второго посредника ИТФ, его взаимодействие с мембраной внутриклеточных кальциевых депо, выброс из них Ca2+ и т.д. Изысканное. Действующее начало - глутамат. В мембране вкусовых клеток имеется множество ионотропных рецепторов глутамата (неселективный катионный канал). При взаимодействии глутамата с такими рецепторами в цитоплазму вкусовой клетки входят Na+ и Ca2+, что приводит к деполяризации клеточной мембраны. Дальнейшие события развиваются по вышеприведённой схеме.
Обобщение главы
Запах обнаруживается нервными клетками обонятельной слизистой оболочки.
Тысячи различных запахов могут определяться и различаться.
Вкус опосредуется сенсорными эпителиальными клетками во вкусовых луковицах.
Имеется пять фундаментальных вкусовых ощущений: сладкого, горького, солёного, кислого и изысканного.