Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА 16. ПОВЕДЕНЧЕСКИЕ И ВИСЦЕРАЛЬНЫЕ РЕАКЦИИ
Кора больших полушарий, ствол мозга, гипоталамус и лимбическая система имеют непосредственное отношение к врождённым формам поведения, возникновению эмоций, мотивациям, реакциям внимания и пробуждения, поддержанию состояния бодрствования и сна. Эмоции, являясь субъективным переживанием человека, - сложные комплексы осознанного поведения, памяти и инстинктивных желаний. В самом общем виде под эмоциями понимают чувства и их проявления в поведении и реакциях вегетативной и эндокринной систем.
АКТИВИРУЮЩИЕ СИСТЕМЫ МОЗГА
Сигналы из ствола мозга активируют кору большого мозга двумя путями: постоянной посылкой нервных импульсов в обширные области больших полушарий мозга и стимуляцией нейрогормональных систем.
Ретикулярная формация ствола мозга
Ретикулярная формация - филогенетически древняя сердцевина мозга - занимает медиовентральную область продолговатого мозга, моста и среднего мозга. Она состоит из множества образующих комплексы сетевых структур (например, адренергических, серотонинергических и норадренергических нейронных систем). В ретикулярной формации находятся области, связанные с регуляцией ритма сердца, АД, дыхания и других функций. Некоторые её нисходящие тракты блокируют передачу импульсов в сенсорных проводниках спинного мозга и влияют на систему рефлексов растяжения.
• Неспецифичность системы. Ретикулярная формация - комплекс полисинаптических путей. Аксоны входят в неё не только из про- тяжённых восходящих сенсорных трактов, но и из систем тройничного, слухового, зрительного нервов. Сложность нейронных сетей и степени конвергенции сигналов привели к нарушению
• модальной специфичности. Поэтому большинство нейронов ретикулярной системы активируется с одинаковой степенью лёг- кости различными сенсорными стимулами, в этом отношении ретикулярная формация является неспецифической. В то же время волокна классических сенсорных путей (см. главы 8-12) называются специфическими, поскольку они активируются одним типом сенсорного раздражения. Контроль активности мозга. На рис. 16-1 приведена общая система контроля активности мозга. Центральный компонент этой системы расположен в ретикулярной формации моста и среднего мозга, это бульборетикулярная облегчающая область. Возбуждающие сигналы из этой области поступают прежде всего в таламус, где они возбуждают новую систему нейронов. Через таламус проходят два типа сигналов - быстрые и медленные.
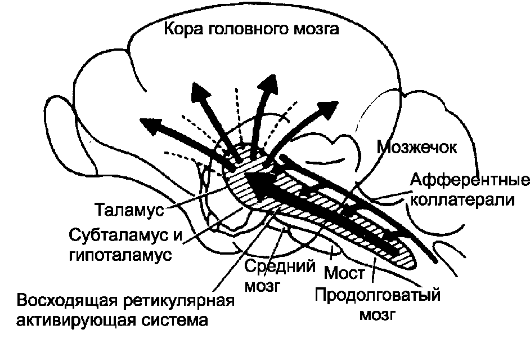 Рис. 16-1. Проекции восходящей ретикулярной активирующей системы
Рис. 16-1. Проекции восходящей ретикулярной активирующей системы
❖ Крупные нейроны, лежащие в глубине мозгового ствола, генерируют быстропередаваемые ПД. Ацетилхолин, выделяющийся из терминальных ветвлений аксонов этих нервных клеток, возбуждает мозг в течение нескольких миллисекунд.
❖ Второй тип возбуждающих сигналов генерируют нейроны небольшого размера, расположенные в ретикулярной формации ствола мозга. Их тонкие и медленно проводящие волокна заканчиваются синапсами на внутрипластинчатых ядрах таламуса и лишь отсюда сигналы распространяются по всей коре больших полушарий. Возбуждающий эффект этой системы может продолжаться секунды, минуты и более.
• Уровень активности ретикулярной формации и, следовательно, уровень активности большого мозга в значительной мере опре-
деляется количеством и характером сенсорных сигналов, поступающих в мозг с периферии. Так, сигналы боли исключительно сильно возбуждают ретикулярную формацию мозгового ствола, повышая реакции внимания и тревоги.
• Обратная связь. Возбуждённое состояние ретикулярной формации усиливают сигналы обратной связи от больших полушарий. Активированная кора мозга посылает сигналы к стволу мозга, который, в свою очередь, посылает ещё более сильные возбуждающие сигналы к коре. Это помогает поддерживать уровень возбуждения коры больших полушарий и даже усиливать его по механизму положительной обратной связи.
• Нейрогормональная активация мозга. Помимо прямого (импульсного) контроля активности головного мозга со стороны ствола мозга, существует и нейрогормональный способ активации. Нейрогормоны разных центров (голубое пятно и норадреналин, чёр- ное вещество и дофамин, ядра шва и серотонин, гигантоклеточные нейроны ретикулярной формации и ацетилхолин) действуют от минут до нескольких часов, обеспечивая длительный эффект.
ЛИМБИЧЕСКАЯ СИСТЕМА
Термин «лимбическая система» применяют по отношению к старой коре (рис. 16-2, врезка) и группе связанных с этой корой и между собой глубоких структур (перегородка, околообонятельное поле, передний таламус, часть базальных ганглиев, гипоталамус, гиппокамп, миндалевидное тело; рис. 16-2А,Б). С физиологической точки зрения гипоталамус - центральная структура лимбической системы.
Гипоталамус
Гипоталамус (рис. 16-2В) - часть промежуточного мозга - содержит ядра и ядерные области, имеющие многочисленные связи с разными структурами, что позволяет гипоталамусу контролировать разнообразные функции.
• Афферентные и эфферентные связи. Гипоталамус соединён со многими отделами ЦНС и в том числе с другими частями лимбической системы, структурами среднего мозга, моста и продолговатого мозга (и через них - с периферическими отделами вегетативной нервной системы). Восходящие влияния гипоталамуса направлены к различным областям промежуточного мозга и больших полушарий, особенно к переднему таламусу и лимбической коре. Большинство нервных волокон в этих путях - без-
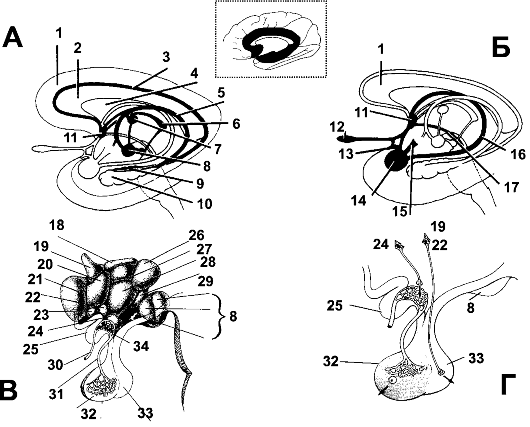 Рис. 16-2. Лимбическая система. На врезке вверху: старая (лимбическая) кора выделена чёрным. А. Система гиппокампа и его связи. Б. Связи обонятельной системы и миндалевидного тела. В. Гипоталамус. Г. Гипоталамо-гипофизарная
система. 1 - лимбическая кора; 2 - мозолистое тело; 3 - медиальный
продольный пучок; 4 - прозрачная перегородка; 5 - свод; 6 - поводок; 7 -
передний таламус; 8 - сосцевидное тело; 9 - зубчатая извилина; 10 -
гиппокамп; 11 - ядра перегородки; 12 - обонятельная луковица; 13 -
латеральная обонятельная полоска; 14 - миндалевидное тело; 15 - волокна
между миндалевидным телом и гипоталамусом; 16 - терминальная полоска; 17
- медиальный пучок конечного мозга; 18 - дорсальная гипоталамическая
область; 19 - паравентрикулярное ядро; 20 - передняя гипоталамическая
область; 21 - предзрительное поле; 22 - супраоптическое ядро; 23 -
надперекрестное ядро; 24 - дугообразное ядро; 25 - зрительный перекрест;
26 - заднее ядро гипоталамуса; 27 - дорсомедиальное ядро; 28 -
вентромедиальное ядро гипоталамуса; 29 - предсосцевидные ядра; 30 -
верхняя гипофизарная артерия; 31 - воротный сосуд; 32 - передняя доля
гипофиза; 33 - задняя доля гипофиза; 34 - первичное сосудистое сплетение
Рис. 16-2. Лимбическая система. На врезке вверху: старая (лимбическая) кора выделена чёрным. А. Система гиппокампа и его связи. Б. Связи обонятельной системы и миндалевидного тела. В. Гипоталамус. Г. Гипоталамо-гипофизарная
система. 1 - лимбическая кора; 2 - мозолистое тело; 3 - медиальный
продольный пучок; 4 - прозрачная перегородка; 5 - свод; 6 - поводок; 7 -
передний таламус; 8 - сосцевидное тело; 9 - зубчатая извилина; 10 -
гиппокамп; 11 - ядра перегородки; 12 - обонятельная луковица; 13 -
латеральная обонятельная полоска; 14 - миндалевидное тело; 15 - волокна
между миндалевидным телом и гипоталамусом; 16 - терминальная полоска; 17
- медиальный пучок конечного мозга; 18 - дорсальная гипоталамическая
область; 19 - паравентрикулярное ядро; 20 - передняя гипоталамическая
область; 21 - предзрительное поле; 22 - супраоптическое ядро; 23 -
надперекрестное ядро; 24 - дугообразное ядро; 25 - зрительный перекрест;
26 - заднее ядро гипоталамуса; 27 - дорсомедиальное ядро; 28 -
вентромедиальное ядро гипоталамуса; 29 - предсосцевидные ядра; 30 -
верхняя гипофизарная артерия; 31 - воротный сосуд; 32 - передняя доля
гипофиза; 33 - задняя доля гипофиза; 34 - первичное сосудистое сплетение
миелиновые. Наконец, гипоталамус контролирует эндокринные функции гипофиза.
• Связь с гипофизом (рис. 16-2В,Г) осуществляется двояко: при помощи гипоталамо-гипофизарного тракта (нервный путь) и по сосудам портальной системы кровотока (гуморальный путь).
Функции гипоталамуса
В отличие от активирующей ретикулярной системы стволовой части мозга, реакции гипоталамуса (от достаточно чётких висцеральных рефлексов до комплексных поведенческих и эмоциональных актов) всегда имеют специфический характер, т.е. возникают как реакции на специфические стимулы. Гипоталамус контролирует множество висцеральных (в том числе эндокринных) и поведенческих функций.
• Висцеральные функции гипоталамуса. В реализации многих висцеральных функций принимают участие конкретные структуры гипоталамуса, во многих случаях выступая в качестве главного регулятора или интегрирующего центра. Так, заднее ядро гипоталамуса ответственно за повышение АД и расширение зрачков, вентромедиальное ядро контролирует насыщение, предсосцевидные ядра - голод, сосцевидное тело - пищедобывательные рефлексы, дугообразное ядро осуществляет нейроэндокринный контроль, надперекрестное ядро ответственно за сокращение мочевого пузыря, снижение ЧСС, уменьшение АД, супраоптическое ядро синтезирует вазопрессин. Предзрительное поле ответственно за регуляцию температуры тела, одышку, потоотделение, а также тормозит выделение ТТГ, паравентрикулярное ядро синтезирует окситоцин и регистрирует задержку воды в организме.
• Поведенческие функции гипоталамуса. Участие гипоталамуса во многих поведенческих функциях установлено экспериментально на подопытных животных, наблюдениями за больными с повреждениями гипоталамических структур, а также в ходе нейрохирургичесих операций (эффекты стимуляции и повреждения).
❖ Эффекты стимуляции гипоталамуса
♦ Латеральный гипоталамус: жажда, аппетит, увеличение активности организма, ярость, агрессия.
♦ Вентромедиальное ядро и окружающие его области: чувство насыщения, снижается аппетит, возникает успокоение.
♦ Перивентрикулярные ядра: страх и боязнь наказания.
♦ Некоторые области переднего и заднего гипоталамуса: усиление поиска полового партнёра.
❖ Эффекты разрушения гипоталамуса противоположны эффектам его стимуляции.
♦ Латеральный гипоталамус: потеря аппетита и жажды, пассивность и малоподвижность.
♦ Вентромедиальная область: неукротимые аппетит и жажда, жестокость и ярость.
❖ Центры поощрения и наказания. Личные оценки могут характеризовать ощущения как приятные или неприятные (поощрительные или наказывающие, или как приносящие удовольствие или вызывающие неудовольствие). Электрическая стимуляция некоторых лимбических областей доставляет удовольствие, раздражение других зон - боль, страх, защиту, реакции нападения или избегания. Степень стимуляции этих двух противоположно отвечающих систем существенно влияет на поведение животных.
❖ Роль поощрения и наказания в поведении, обучении и памяти.
Всё или почти все, что делает человек, имеет отношение к поощрению и наказанию. Следовательно, центры поощрения и наказания являются одними из наиболее важных контролё- ров нашей физической активности, побуждений, антипатий, мотиваций. Центры поощрения и наказания лимбической системы существенно влияют на отбор получаемой информации. Обычно около 99% информации удаляется и для закрепления в памяти остаётся не более 1%.
❖ Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших полушарий. Повторение этих же стимулов приводит к почти полному затуханию корковых ответов (если сенсорное научение не вызывает чувства поощрения).
Гиппокамп
Гиппокамп (рис. 16-2А,Б) вместе с прилежащими к нему структурами лобной и теменной долей имеет хорошо развитую систему связей с корой больших полушарий, обонятельным мозгом, миндалевидным телом, перегородкой, гипоталамусом и сосцевидными телами. Любой вид сенсорного опыта вызывает активацию гиппокампа, который, в свою очередь, посылает сигналы в передний таламус, гипоталамус и другие отделы лимбической системы через свод головного мозга. Таким образом, гиппокамп служит дополнительным каналом, через который входящие сенсорные сигналы могут побуждать к соответствующим поведенческим реакциям. Раздражение различных участков гиппокампа может вызывать разнооб-
разные эмоциональные и поведенческие реакции: удовольствие, ярость, пассивное или чрезмерное половое влечение. Существенной функцией гиппокампа является консолидация следов памяти.
Миндалевидное тело
Миндалевидное тело (рис. 16-2В) - комплекс ядер, имеющих обширные двусторонние связи с гипоталамусом и всеми частями лимбической системы, а также с височной, теменной и затылочной долями новой коры. Особенно велико число связей со слуховой и зрительной ассоциативными зонами. Функция миндалевидного тела заключается в осуществлении поведенческих проявлений на уровне врождённых форм поведения. Миндалевидное тело посылает в лимбическую систему информацию о сиюминутном состоянии организма и его окружения и помогает создавать образцы персональных поведенческих ответов, соответствующих каждому случаю.
Старая кора
Лимбическая кора функционирует как ассоциативная область контроля поведения. Передняя височная кора имеет отношение к вкусовым и обонятельным ассоциациям, парагиппокампальная извилина - к слуховым и речевым ассоциациям, средняя и задняя поясная кора - к сенсомоторным ассоциациям.
КОРА БОЛЬШИХ ПОЛУШАРИЙ И ТАЛАМУС
Кора больших полушарий - тонкий слой нейронов, покрывающий всю поверхность мозга. Толщина коры от 2 до 5 мм, площадь - до четверти квадратного метра, количество нейронов около 100 млрд. Большинство нейронов относится к зернистым, веретенообразным и пирамидным, расположенным в шести слоях (рис. 16-3А). Пирамидные нейроны имеют разветвлённое вертикальное дендритное дерево (рис. 16-3Б), а аксоны - возвратные коллатерали. Афферентные волокна от специфических ядер таламуса заканчиваются в слое 4, неспецифические - в слоях 1-4.
Таламус подразделяют на эпиталамус, дорсальный таламус и вентральный таламус. Эпиталамус имеет связи с обонятельной системой и функционирует самостоятельно. Дорсальный таламус содержит неспецифические проекционные ядра (ядра срединной линии и внутрипластинчатые), которые проецируются ко всей коре, и ядра, проецирующиеся к специфическим участкам коры и лимбической системы (специфические проекционные ядра). Неспецифические проекционные ядра получают информацию из ретикулярной активирующей системы. Активация неспецифических ядер вызывает диф-
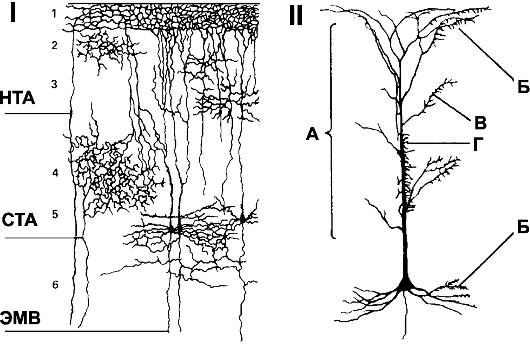 Рис. 16-3. СВЯЗИ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ. I. Афферентные пути из таламуса. СТА
- специфические таламические афференты; НТА - неспецифические
таламические афференты; ЭМВ - эфферентные моторные волокна. Цифрами
обозначены слои коры. II. Пирамидный нейрон и распределение окончаний на нём. А
- неспецифические афферентные волокна из ретикулярной формации и
таламуса. Б - возвратные коллатерали от аксонов пирамидных нейронов. В -
комиссуральные волокна из зеркальных клеток противоположного полушария.
Г - специфические афферентные волокна из сенсорных релейных ядер
таламуса
Рис. 16-3. СВЯЗИ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ. I. Афферентные пути из таламуса. СТА
- специфические таламические афференты; НТА - неспецифические
таламические афференты; ЭМВ - эфферентные моторные волокна. Цифрами
обозначены слои коры. II. Пирамидный нейрон и распределение окончаний на нём. А
- неспецифические афферентные волокна из ретикулярной формации и
таламуса. Б - возвратные коллатерали от аксонов пирамидных нейронов. В -
комиссуральные волокна из зеркальных клеток противоположного полушария.
Г - специфические афферентные волокна из сенсорных релейных ядер
таламуса
фузный электрический ответ в коре, регистрируемый на электроэнцефалограмме (ЭЭГ). Специфические проекционные ядра проецируют свои влияния на специфические участки коры. Эти ядра подразделяются на специфические сенсорные релейные, отвечающие за контроль эфферентных механизмов и имеющие отношение к комплексу интегративных функций.
Электрическая активность мозга
Вызванные корковые потенциалы (рис. 16-4,1). Электрические проявления в коре больших полушарий можно наблюдать после раздражения чувствительного органа. Под электродами, наложенными на соответствующий участок коры, спустя 5-12 мс после раздражения появляется небольшая положительно-отрицательная
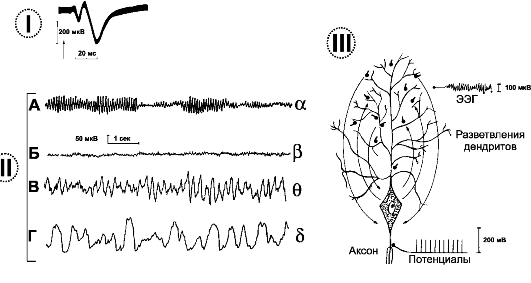
Рис. 16-4. ЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ МОЗГА. I. Вызванный ответ в сенсорной коре. Отклонение вверх - электроотрицательный потенциал. II. Типы ритмов электроэнцефалограммы. А - альфа. Б - бета. В - тета. Г - дельта. III. Ответы аксона и дендритов пирамидного нейрона коры больших полушарий
волна (соответствует первичному вызванному потенциалу) с последующей более продолжительной положительной волной (соответствует диффузному вторичному ответу).
• Первичный вызванный потенциал специфичен по локализации и может наблюдаться только там, где заканчивается специфический афферентный путь.
• Диффузный вторичный ответ не имеет строгой локализации и может одновременно появляться в разных участках коры, возникает за счёт активации неспецифических таламических путей.
Электроэнцефалограмма
Электроэнцефалограмма (ЭЭГ) - регистрация колебаний разности потенциалов с интактной кожи головы. Электрокортикограм-
ма - регистрация потенциалов с электродов, наложенных непосредственно на поверхность коры больших полушарий. ЭЭГ может быть зарегистрирована биполярно или монополярно. Биполярная регистрация показывает колебания потенциалов между двумя электродами на поверхности кожи головы. Монополярно регистрируется разность потенциалов между электродом на кожной поверхности головы и индифферентным электродом, удалённым от поверхности головы.
• Ритмы ЭЭГ. В норме у здорового взрослого человека с закрытыми глазами регистрируется основной α-ритм, при открытии глаз - β-ритм, в состоянии сна - медленные ритмы типа θ и δ- волн (рис. 16-4,II).
Альфа (а)-ритм: частота 8-13 Гц, амплитуда около 50 мкВ. Бета (β)-ритм: частота 14-40 Гц, амплитуда до 15 мкВ. Лучше всего регистрируется в области передних центральных извилин. Тета (θ)-ритм: частота 4-6 Гц, амплитуда волн превышает 40 мкВ, при патологических состояниях может достигать 300 мкВ и более.
Дельта (δ)-ритм: частота 0,5-3 Гц, амплитуда δ-волн такая же, как у θ-ритма.
• Изменения в ЭЭГ. У человека частота доминирующего ритма в покое варьирует с возрастом. У детей может встречаться β-по- добная быстрая активность, но доминирует медленный ритм (0,5- 2 Гц). С возрастом частота ритмов возрастает. Снижение уровня сахара крови, низкая температура тела, пониженный уровень глюкокортикоидных гормонов и высокое содержание CO2 в крови снижают частоту α-ритма. Частота α-ритма возрастает в условиях повышенной температуры, низкого pCO2 повышенного уровня глюкокортикоидных гормонов. Усиленное дыхание (гипервентиляция лёгких), снижая pCO2 в крови, используется в клинике для выявления скрытых нарушений в ЭЭГ.
• Блокада α-ритма. Открывание глаз приводит к замещению α-ритма на β-ритм (блокада α-ритма). Прерывание α-активности происходит при любом виде сенсорной стимуляции, даже при попытках «считать в уме». Общепринятый термин, используемый для описания замены доминирующего α-ритма нерегулярной низкоамплитудной активностью, - десинхронизация ЭЭГ.
• Синхронизирующие механизмы. Чёткий ритмический характер α- волн указывает, что активность множества дендритных единиц синхронна. Два фактора определяют такую синхронизацию: синхронизирующее влияние активности каждой отдельной единицы на соседние единицы и ритмические разряды из таламуса.
• Клиническое применение электроэнцефалографии. ЭЭГ применяют с целью: -о диагностики эпилепсии, -о оценки функционального состояния ЦНС, -о определения тяжести состояния при коматозных явлениях, - оценки последствий черепно-мозговых травм и инсультов, - контроля мозговой активности при сложных операционных вмешательствах. На рис 16-5 представлены регистрируемые на ЭЭГ при судорожном припадке основные типы эпилептиформной активности.
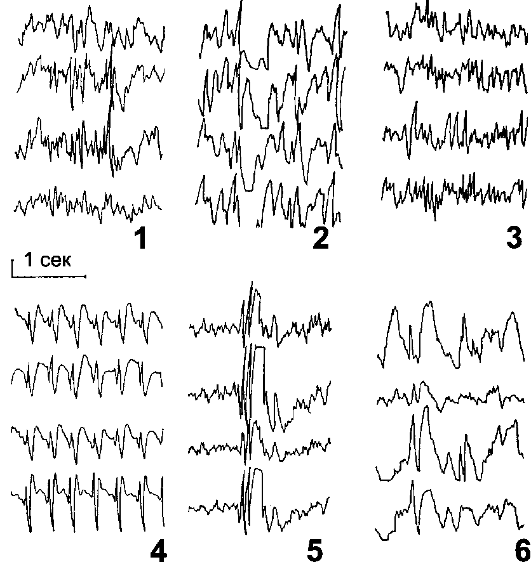 Рис. 16-5. Электроэнцефалограммы при судорожных припадках. 1
- спайки; 2 - острые волны; 3 - острые волны в полосе бета; 4 -
комплекс «спайк-волна»; 5 - множественные «спайки-волна»; 6 - «острая
вол- на-медленная волна». Значение калибровочного сигнала для 1-4 - 100
мкВ, для остальных записей - 50 мкВ
Рис. 16-5. Электроэнцефалограммы при судорожных припадках. 1
- спайки; 2 - острые волны; 3 - острые волны в полосе бета; 4 -
комплекс «спайк-волна»; 5 - множественные «спайки-волна»; 6 - «острая
вол- на-медленная волна». Значение калибровочного сигнала для 1-4 - 100
мкВ, для остальных записей - 50 мкВ
Сон
Сон - бессознательное состояние, из которого человек может быть выведен (разбужен) сенсорным или каким-либо другим стимулом. Различают медленный (медленно-волновой) сон и быстрый (парадоксальный) сон, или сон с быстрыми движениями глаз (REM- сон, от Rapid Eye Movement) парадоксальным быстрый сон называется потому, что в условиях мышечной атонии наблюдаются быстрые движения глаз и нерегулярные мышечные сокращения конечностей и туловища. Большую часть времени сна занимает медленный сон. Он может быть глубоким, успокаивающим сном, который испытывает человек в течение первого часа сна после многочасового бодрствования. Быстрый сон наблюдается эпизодически в течение медленного сна и занимает у молодых людей около 25% общего времени сна. Каждый период быстрого сна нормально повторяется через 90 минут. Быстрый сон не является успокаивающим, поскольку он связан с яркими сновидениями.
Медленный сон характеризуется успокаивающим воздействием на организм: происходит снижение тонуса периферических сосудов, замедление вегетативных функций, снижаются АД, частота дыхания и основной метаболизм тела. Во время медленного сна (вопреки мнению об отсутствии сновидений) появляются сновидения и даже ночные кошмары. Отличие сновидений во время медленного и быстрого сна в том, что сновидения, возникающие во время быстрого сна, остаются в памяти, а сновидения во время медленного сна не запоминаются. Медленный сон подразделяют на 4 стадии (рис. 16-6).
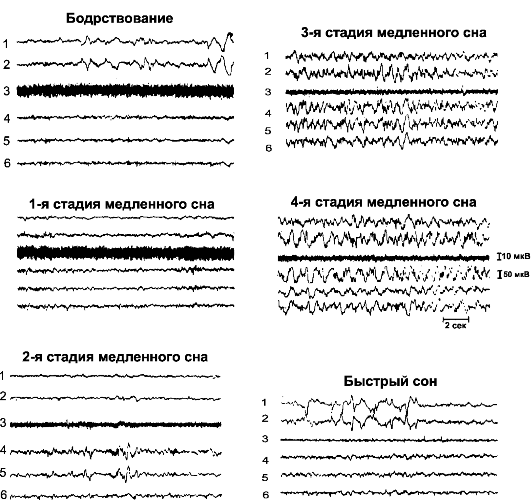 Рис. 16-6. Электроэнцефалограммы и записи мышечной активности на различных стадиях сна. 1, 2 - электроокулограммы; 3 - электромиограммы; ЭЭГ: 4 - париетальная; 5 - лобная; 6 - затылочная
Рис. 16-6. Электроэнцефалограммы и записи мышечной активности на различных стадиях сна. 1, 2 - электроокулограммы; 3 - электромиограммы; ЭЭГ: 4 - париетальная; 5 - лобная; 6 - затылочная
• Первая стадия - дремота и переход ко сну. В ЭЭГ регистрируется низкоамплитудная активность высокой частоты.
• Вторая стадия - появление в ЭЭГ сонных веретён. Это α-подобные волны частотой 10-14 Гц и амплитудой 50 мкВ.
• Третья стадия характеризуется наличием в ЭЭГ волн низкой частоты и высокой амплитуды (δ-волны).
• В четвертой стадии в ЭЭГ отмечаются максимально медленные δ-волны большой амплитуды. Следовательно, глубокий сон характеризуется ритмичными медленными волнами ЭЭГ, указывающими на синхронизацию дендритных потенциалов коры. Быстрый сон. Во время нормального ночного сна периоды: быстрого сна продолжаются от 5 до 30 мин каждые 90 мин. Если человек находится в состоянии глубокого сна, то каждый отрезок быстрого сна становится короче, а иногда он даже отсутствует. И наоборот: если человек хорошо отдохнул днём, то продолжительность быстрого сна возрастает. Быстрый сон характеризуется следующими особенностями: - много мышечных движений и сновидений; -о человека труднее разбудить, чем во время медленного сна; несмотря на это, во время эпизодов быстрого сна человек просыпается спонтанно; - тонус мышц всего тела снижен вследствие сильного торможения областей спинного мозга, контролирующих мускулатуру; - ЧСС и частота дыхания становятся нерегулярными; - наблюдаются нерегулярные мышечные сокращения и быстрые движения глаз; - возможна эрекция полового члена и клитора; - мозг находится в состоянии повышенной активности и общий метаболизм мозга увеличивается до 20%; -о ЭЭГ имеет тот же вид, что и в состоянии бодрствования.
Физиологическое значение сна. Сон оказывает влияние как на нервную систему, так и на деятельность различных органов и систем организма. Длительная инсомния усиливает раздражительность, утомляемость, понижается координация движений. Сон различными путями восстанавливает нормальный уровень активности нервных центров и баланс их деятельности. Во время сна уменьшается активность симпатической нервной системы и возрастает деятельность парасимпатической нервной системы. Восстанавливающее действие сна на вегетативные функции проявляется в снижении АД, расширении сосудов кожи, некотором усилении работы ЖКТ, расслаблении мышечного тонуса, снижении общего обмена на 10-30%, повышение процессов анаболизма на III-IV стадиях медленного сна.