Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА 9. СОМАТИЧЕСКАЯ И ВИСЦЕРАЛЬНАЯ ЧУВСТВИТЕЛЬНОСТЬ
Сенсорные ощущения подразделяются на 3 физиологических класса: механорецептивные, температурные и болевые. Механорецептивные ощущения включают тактильные (прикосновение, давление, вибрация) и проприоцептивные (постуральные) - ощущение позы, статического положения и положения при движении.
По месту возникновения ощущений чувствительность классифицируется, как экстероцептивная (ощущения, возникающие с поверхности тела), висцеральная (ощущения, возникающие во внутренних органах) и глубокая (ощущения поступают от глубоколежащих тканей - фасций, мышц, костей).
• Соматические сенсорные сигналы передаются с большой скоростью, высокой точностью локализации и определения минимальных градаций интенсивности или изменений силы сенсорного сигнала.
• Висцеральные сигналы характеризуются более низкой скоростью проведения, менее развитой системой пространственной локализации восприятия сигнала, менее развитой системой градации силы раздражения и меньшей способностью передавать быстрые изменения сигнала.
СОМАТОСЕНСОРНЫЕ СИГНАЛЫ
Тактильная чувствительность
Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений, но воспринимаются одними и теми же рецепторами.
• Ощущение прикосновения - результат стимуляции чувствительных нервных окончаний кожи и подлежащих тканей.
• Ощущение давления возникает в результате деформации глубоких тканей.
• Вибрационное ощущение возникает в результате быстрых повторных сенсорных стимулов, наносимых на те же рецепторы, что и рецепторы, воспринимающие прикосновение и давление.
Тактильные рецепторы
Кожа имеет богатую чувствительную иннервацию.
Свободные нервные окончания (рис. 8-1А). Эти рецепторы расположены как в различных слоях кожи, так и во многих других тканях. В эпидермисе свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы болевой (ноцицептивной) чувствительности. Терморецепторы подразделяют на рецепторы холодовой (25-30 °C) и тепловой (40-42 °C) чувствительности.
Тельца Майсснера (рис. 8-1В) присутствуют под эпидермисом (в сосочковом слое кожи), они особенно многочисленны в безволосой части кожи - коже ладонной и подошвенной поверхностей пальцев рук, ног, а также в коже губ, век, наружных половых органов и сосков молочной железы. Эти рецепторы особо чувствительны к движению объектов по поверхности кожи и низкочастотной вибрации. Они адаптируются за доли секунды.
Клетки Меркеля (рис. 8-1Б) расположены в базальном слое кожи ладоней и подошв. Диски Меркеля - медленно адаптируемые рецепторы. Механический стимул вызывает вначале быстрый разряд импульсов в течение 1-2 мс, затем наступает неполная адаптация к стимулу, и в течение достаточно продолжительного времени этот рецептор реагирует на продолжающееся соприкосновение предмета с кожей.
Палисадный аппарат волосяных фолликулов является рецептором прикосновения. Он быстро адаптируется и, подобно тельцам Майсснера, является детектором движения объекта по поверхности тела или начального контакта с предметом.
Тельца Пачини (рис. 8-1Г) - самые крупные инкапсулированные рецепторы - расположены глубоко в коже и в подкожной клетчатке преимущественно в пальцах, наружных половых органах и молочной железе. Тельца Пачини регистрируют вибрацию и других быстрые изменения окружающих тканей. Они стимулируются только быстрыми смещениями тканей и адаптируются в течение сотых долей секунды.
Тельца Руффини чаще встречаются в коже подошвенной поверхности стопы, а также в более глубоких тканях (например, в капсуле суставов). Эти рецепторы очень медленно адаптируются и сигнализируют о постоянной, вызванной тяжёлым и длительным прикосновением или давлением.
Колбы Краузе. В сетчатом слое кожи располагаются концевые колбы Краузе - инкапсулированные механорецепторы, имеющие сходную структуру с тельцем Пачини, но меньшие размеры. Они
реагируют на начало деформации кожи и последующие повторные стимулы с умеренно высокой скоростью. Импульсация от них может продолжаться в течение многих секунд после прекращения действия механического стимула.
Восприятие сенсорных сигналов требует тонкой дифференцировки (например, определения точной локализации на коже, минимальных градаций в интенсивности или очень быстрых изменений силы сигнала). Передача таких тактильных ощущений от специализированных сенсорных рецепторов происходит по миелинизированным нервным волокнам со скоростью от 30 до 70 м/с. Свободные нервные окончания передают сигналы со скоростью от 5 до 30 м/с. Грубые виды сигналов (например, сильное давление, прикосновение без чёткой локализации и в особенности щекотание) передаются нервными волокнами малого диаметра с небольшой скоростью (так, от нервных окончаний, формирующих ощущение щекотки, сигналы идут по безмиелиновым нервным волокна С-типа со скоростью до 2 м/с).
Проприоцептивное чувство
Чувство положения тела (проприоцептивное чувство) складывается из регистрации статического положения (осознанное восприятие различных частей тела по отношению друг к другу) и ощущения скорости движения (кинестезия, или динамическая проприоцепция).
• Для определения статического и динамического положения тела необходима информация о степени сгибания всех суставов во всех направлениях и о скорости этих изменений. Эту функцию выполняют многие рецепторы: кожные тактильные и собственные (глубокие) рецепторы суставов. Для пальцев важнее кожные тактильные рецепторы, посредством которых определяется их положение. Для больших суставов тела важнее глубокие рецепторы.
• Для определения степени сгибания конечностей в суставах во время умеренного движения наиболее значимы мышечные веретёна. Когда угол сгибания изменяется, некоторые мышцы растягиваются, а другие расслабляются. Информация от рецепторов растяжения - мышечных веретён - проходит в «компьютерные» системы спинного мозга и по задним канатикам поднимается к более высоким структурам для расшифровки комплексных взаимоотношений между различными суставами.
• Расположение углов сгибания суставов в конечностях, растяжение связок и окружающих суставы тканей - дополнительные факторы для определения положения тела. В этих структурах дат-
чиками служат тельца Пачини, тельца Руффини, сухожильные органы Гольджи.
Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела (см. рис. 9-6) поступает в спинной мозг через проходящие в составе задних корешков центральные отростки чувствительных нейронов спинномозговых узлов (рис. 9-2). Войдя в спинной мозг, центральные отростки чувствительных нейронов либо прямо направляются к продолговатому мозгу (лемнисковая система: тонкий, или нежный пучок Голля и клиновидный пучок Бурдаха), либо заканчиваются на вставочных нейронах, аксоны которых идут к таламусу в составе вентрального, или переднего и латерального, или бокового спиноталамических восходящих путей.
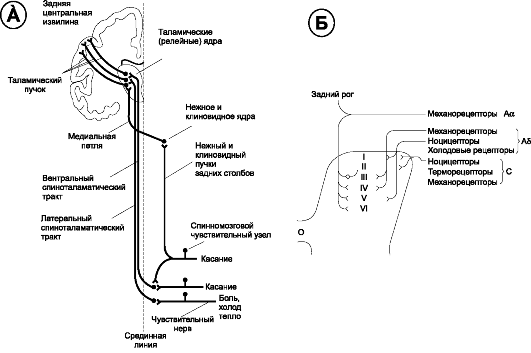 Рис. 9-2. Восходящие пути чувствительности. А. Путь
от чувствительных нейронов спинномозговых узлов (первый, или первичный
чувствительный нейрон) через вторые нейроны (вставочные нейроны спинного
мозга или нервные клетки клиновидного и тонкого ядра продолговатого
мозга) к третьим нейронам пути - таламическим. Аксоны этих нейронов
направляются к коре головного мозга. Б. Расположение нейронов, передающих разные модальности, в пластинах (римские цифры) спинного мозга
Рис. 9-2. Восходящие пути чувствительности. А. Путь
от чувствительных нейронов спинномозговых узлов (первый, или первичный
чувствительный нейрон) через вторые нейроны (вставочные нейроны спинного
мозга или нервные клетки клиновидного и тонкого ядра продолговатого
мозга) к третьим нейронам пути - таламическим. Аксоны этих нейронов
направляются к коре головного мозга. Б. Расположение нейронов, передающих разные модальности, в пластинах (римские цифры) спинного мозга
• Тонкий и клиновидный пучки - проводящие пути проприоцептивной и тактильной чувствительности - проходят в составе заднего канатика той же стороны спинного мозга и заканчиваются в тонком и клиновидном ядрах продолговатого мозга. Аксоны нейронов этих ядер по медиальной петле (отсюда и название - лемнисковая система) переходят на противоположную сторону и направляются к таламусу.
• Спиноталамический путь вентральный - проекционный афферентный путь, проходящий в переднем канатике противоположной стороны. Периферические отростки первых нейронов, расположенных в спинномозговых узлах, проводят тактильные и прессорные ощущения от механорецепторов кожи. Центральные отростки этих нейронов вступают через задние корешки в задние канатики, где поднимаются на 2-15 сегментов и образуют синапсы с вставочными нейронами задних рогов. Аксоны этих нейронов переходят на противоположную сторону и проходят далее в передней периферической зоне переднебоковых канатиков. Отсюда волокна пути восходят к заднелатеральному вентральному ядру таламуса вместе с латеральным спиноталамическим путём.
• Спиноталамический путь латеральный - проекционный афферентный путь, проходящий в боковом канатике. Периферическими рецепторами являются свободные нервные окончания кожи. Центральные отростки псевдоуниполярных нейронов спинномозговых узлов входят в противоположную часть спинного мозга через латеральные отделы задних корешков и, поднявшись в спинном мозге на 1-2 сегмента, образуют синапсы с нейронами роландова студенистого вещества. Аксоны этих нейронов фактически образуют латеральный спиноталамический путь. Они идут на противоположную сторону и поднимаются в латеральных отделах боковых канатиков. Спиноталамические пути проходят через ствол мозга и заканчиваются в вентро-латеральных ядрах таламуса. Это главный путь проведения болевой и температурной чувствительности.
Задний канатик состоит из толстых миелиновых нервных волокон, проводящих сигналы со скоростью от 30 до 110 м/с; спиноталамические пути состоят из тонких миелиновых волокон, проводящих ПД со скоростью от нескольких метров до 40 м/с.
Соматосенсорная кора
Кора больших полушарий человека подразделяется на 50 различных областей, называемых полями Бродманна (карта функциональных полей, рис. 9-2). Сенсорные сигналы всех модальностей (за
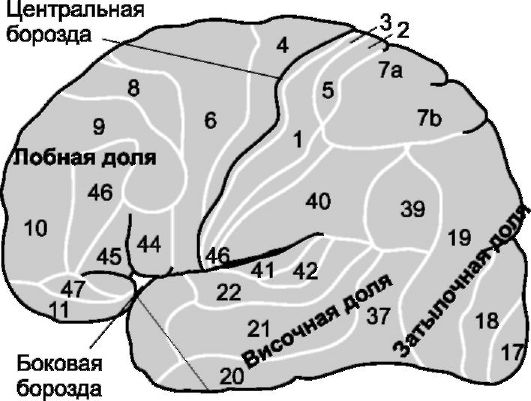 Рис. 9-2. Наружная поверхность больших полушарий головного мозга. Цифры - поля коры по Брод-
Рис. 9-2. Наружная поверхность больших полушарий головного мозга. Цифры - поля коры по Брод-
манну: 1, 2, 3 - первичная соматосенсорная область I, 5, 7 - соматосенсорная ассоциативная область
исключением зрительных и слуховых) поступают в кору мозга позади центральной борозды (на рис. 9-2 поля 1-3, 5, 7). Таким образом, передняя половина теменной доли почти полностью занята рецепцией и интерпретацией соматосенсорных сигналов. Задняя половина теменной доли осуществляет высший уровень интерпретации.
Зрительные сигналы заканчиваются в затылочной области, слуховые сигналы - в височной доле. Область мозговой коры, лежащая кпереди от центральной борозды (задняя половина лобной доли, 4 на рис. 9-2) контролирует мышечные сокращения и движения тела. Важно, что область моторного контроля получает прямые сообщения из сенсомоторной области коры о положении и движениях различных частей тела.
В соматосенсорной коре выделены 2 области - соматосенсорная область I и соматосенсорная область II (рис. 9-3А).
• Соматосенсорная область I располагается непосредственно позади центральной борозды, занимая поля 1, 2 и 3 по Бродманну. Часто употребляемое понятие «соматосенсорная кора» подразумевает именно соматосенсорную область I. В ней плотно расположены точки восприятия различных частей тела - корковое представительство (рис. 9-3Б).
• Соматосенсорная область II. Импульсы поступают в эту область из ствола мозга, передающего сигналы с обеих сторон туловища и конечностей. Кроме того, сюда поступают сигналы из соматосенсорной области I, а также от других сенсорных областей (например, слуховой и зрительной коры).
Соматотопические карты. Соматосенсорная кора каждого полушария получает информацию от противоположной стороны тела (исключением является небольшое количество информации, иду-
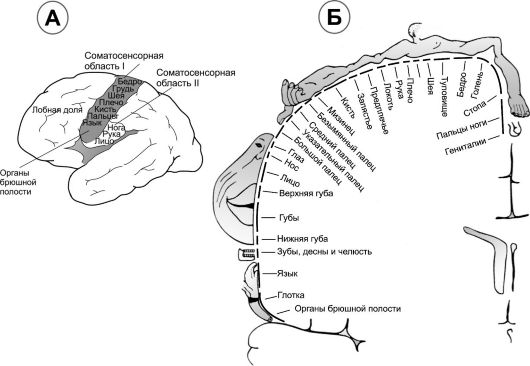 Рис. 9-3. Соматосенсорная кора. А. Локализация соматосенсорных областей I и II на поверхности большого мозга. Б. Соматотопическое
представительство различных областей тела в сенсомоторной области I
(фронтальное сечение мозга на уровне постцентральной извилины)
Рис. 9-3. Соматосенсорная кора. А. Локализация соматосенсорных областей I и II на поверхности большого мозга. Б. Соматотопическое
представительство различных областей тела в сенсомоторной области I
(фронтальное сечение мозга на уровне постцентральной извилины)
щее от той же стороны лица). На рис. 9-3Б показано распределение сенсорных сигналов от различных участков тела в соматосенсорной области I. Самыми большими участками коры представлены губы, затем следуют лицо и большой палец руки. В то же время туловище и нижняя часть тела представлены относительно небольшими участками. Размер участка в соматосенсорной коре прямо пропорционален количеству специализированных рецепторов в каждой периферической области тела. Так, в губах и большом пальце кисти найдено значительное количество специализированных нервных окончаний, а в коже туловища обнаружены лишь отдельные специализированные нервные окончания.
Колонки нейронов - функциональные единицы соматосенсорной коры. Соматосенсорная кора организована в виде функциональных единиц - колонок нейронов, перпендикулярных её поверхности. Каждая колонка имеет диаметр от 0,3 до 0,5 мм и содержит приблизительно 10 тыс. тел нервных клеток. Каждая колонка обслуживает отдельную специфическую сенсорную модальность. Неко-
торые колонки отвечают за рецепторы растяжения вокруг суставов, другие - за стимуляцию волос, третьи - за дискретную локализацию точек давления на коже и так далее. Только в слое IV соматосенсорной коры, где сенсорные сигналы входят в кору, колонки нейронов функционируют отдельно друг от друга. На всех остальных уровнях колонки взаимодействуют, осуществляя начальный анализ значения сенсорных сигналов.
Функции соматосенсорной коры. После двустороннего удаления соматосенсорных областей I наблюдаются следующие изменения:
❖ теряется способность точной локализации ощущений в различных частях тела; ❖ утрачивается способность критически оценивать степень давления; ❖ исчезает способность судить о массе вещей наощупь; ❖ теряется способность оценивать вид или форму предметов наощупь (утрата этой способности называется астереогнозис). Болевое и температурное чувство после удаления соматосенсорной коры не претерпевает значительных изменений в отношении интенсивности или качества ощущений, но становится затруднительной их точная локализация.
Соматосенсорные ассоциативные области и их функции. Поля 5 и 7 по Бродманну (рис. 9-2), лежащие позади соматосенсорной коры, принимают участие в расшифровке информации, поступающей в соматосенсорную кору. Отсюда их название - соматосенсорные ассоциативные области. Электрическая стимуляция соматосенсорной области вызывает у человека комплекс соматических ощущений (например, чувство мяча в руке). По-видимому, соматосенсорная ассоциативная область интегрирует информацию, поступающую из различных точек тела в соматосенсорную кору для расшифровки её значения. Это согласуется с анатомией проводящих путей, входящих в ассоциативную область. Ассоциативная область получает сигналы из: ❖ соматосенсорной области I; ❖ вентробазальных ядер таламуса;
❖ других областей таламуса; ❖ зрительной коры; ❖ слуховой коры. Удаление соматосенсорной ассоциативной области вызывает у человека потерю способности узнавать наощупь комплекс предметов и их форму на противоположной стороне. Кроме того, человек теряет чувство формы своего тела или части тела на противоположной стороне (такая комплексная потеря ощущений называется аморфосинтез).
Обработка сигналов в восходяших проекционных путях
На примере передачи тактильной информации по восходящим проекционным путям лемнисковой системы рассмотрим точность коркового представительства и феномен латерального торможения.
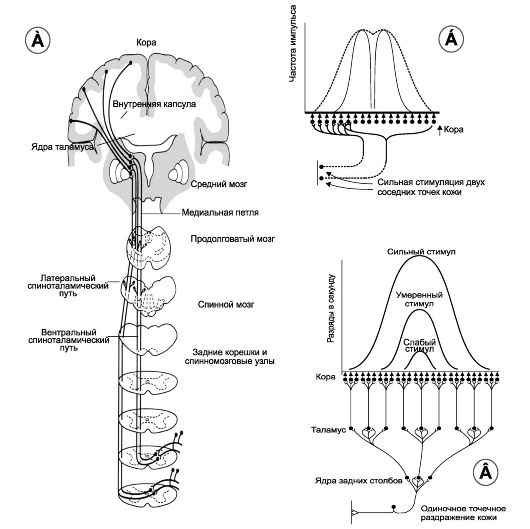 Рис. 9-4. Обработка информации в проекционных путях соматосенсорной чувствительности. А. Проекционный путь. Б. Корковые ответы при раздражении в 2 точках кожи. В. Дивергенция в проекционных путях и корковые ответы при одноточечном раздражении кожи
Рис. 9-4. Обработка информации в проекционных путях соматосенсорной чувствительности. А. Проекционный путь. Б. Корковые ответы при раздражении в 2 точках кожи. В. Дивергенция в проекционных путях и корковые ответы при одноточечном раздражении кожи
• Путь передачи возбуждения по нейронной цепи от раздражения рецептора кожи до соматосенсорной коры показан на рис. 9-4А. Одиночное раздражение. На рис. 9-4В видно, что на каждом этапе синаптического контактирования происходит дивергенция импульсов. Верхняя часть рис. 9-4В демонстрирует, что нейроны
• коры наиболее интенсивно возбуждаются в центральной части коркового рецептивного поля. Так, слабый стимул возбуждает только центрально расположенные нейроны. Умеренное раздражение вовлекает в активность большее число нейронов коры и, наконец, сильное раздражение возбуждает ещё большее количество нейронов. Однако и в этих случаях нейроны с наибольшей скоростью возбуждения (частотой импульсов в секунду) располагаются в центре соматосенсорного коркового поля. Стимуляция в двух точках. На рис. 9-4Б показана передача информации в кору больших полушарий при одновременной стимуляции в коже двух точек. Интенсивная одновременная стимуляция двух соседних точек кожи вызывает в соматосенсорной коре участок возбуждения, пространственный размер которого показан пунктирной кривой. В центральной части возбуждённо- го участка коры частота разрядов самая большая, что отражено в виде двух зубцов, разделённых промежутком. Эти зубцы являются результатом выделения сенсорной корой двух стимулируемых точек. Способность органа чувств находить различие между двумя точками стимуляции в значительной мере зависит от механизма, называемого латеральным торможением (в принципе этот механизм выделения сигнала подходит для любых сенсорных путей).
Раздражение в двух точках применяется в практической медицине, как тест для выявления тактильной дискриминационной способности. Две иглы одновременно слегка надавливают на кожу, и испытуемый должен определить, чувствует ли он раздражение в одной точке или в двух. На кончиках пальцев испытуемый может различить две точки, когда иглы находятся на расстоянии 1- 2 мм. На спине этот интервал составляет от 30 до 70 мм. Естественно, что разница объясняется неодинаковым количеством специализированных тактильных рецепторов в этих областях. Латеральное торможение. Любой сенсорный тракт в состоянии возбуждения одновременно вызывает возникновение латерального торможения. Другими словами, возбуждение, распространяющееся в каком-нибудь направлении, тормозит прилежащие нейроны. Например, возбуждённые нейроны в ядрах задних столбов по коротким коллатералям передают тормозные сигналы к окружающим нервным клеткам. Значение латерального торможения заключается в том, что оно блокирует распространение возбуждающих сигналов в стороны и тем самым увеличивает степень контраста сигналов, воспринимаемых соматосенсорной корой. Возбуждение сенсорных путей задних столбов сопровождается латеральным торможением на
каждом синаптическом уровне (например, в ядрах продолговатого мозга, вентробазальных ядрах таламуса и самой коре (рис. 9-4А). На каждом из этих уровней латеральное торможение блокирует возможность движения возбуждающего сигнала в сторону. В результате возникает устойчивый возбуждённый нейронный ансамбль и блокируются окружающие диффузные стимулирующие сигналы. Этот эффект демонстрируется двумя полностью разделёнными зубцами (рис. 9-4Б: огибающая кривая прерывистая линия отражает возбуждённый участок коры без латерального торможения, а две сплошные кривые отражают эффект возбуждения двух точек кожи с латеральным торможением).
БОЛЕВАЯ ЧУВСТВИТЕЛЬНОСТЬ
Боль - неприятное сенсорное и эмоциональное ощущение, связанное с истинным или потенциальным повреждением ткани или описываемое в терминах такого повреждения. Боль для организма является защитным сигнальным механизмом и может возникнуть в любой ткани, где появились признаки повреждения. Боль подразделяют на быструю и медленную, острую и хроническую.
• Быстрая боль ощущается через 0,1 сек после нанесения болевого стимула. Быструю боль описывают под многими наименованиями: режущая, колющая, острая, электрическая и др. От болевых рецепторов в спинной мозг болевые сигналы передаются по волокнам небольшого диаметра Ad со скоростью от 6 до 30 м/с.
• Медленная боль возникает в течение 1 сек и более, а затем медленно нарастает в течение многих секунд или минут (например, медленная жгучая, тупая, пульсирующая, распирающая, хроническая боль). Боль медленного хронического типа передаётся по С-волокнам со скоростью от 0,5 до 2 м/с.
Существование двойной системы передачи болевых сигналов приводит к тому, что сильное резкое раздражение часто вызывает двойное болевое ощущение. Быстрая боль передаётся немедленно, а через секунду или чуть позже передаётся медленная боль.
Рецепция боли
Боль вызывают многие факторы: механические, температурные и химические болевые стимулы. Быструю боль порождают преимущественно механические и температурные стимулы, медленную - все виды стимулов. Некоторые вещества известны как химические стимуляторы боли: гистамин, брадикинин, серотонин, ионы калия, молочная кислота, ацетилхолин, протеолитические ферменты. Простагландины повышают чувствительность болевых окончаний, но сами
непосредственно не возбуждают их. Болевыми рецепторами (ноцицепторы) являются свободные нервные окончания (рис. 8-1А). Они широко распространены в поверхностных слоях кожи, надкостнице, суставах, стенке артерий. В других глубоких тканях свободных нервных окончаний меньше, но обширные тканевые повреждения могут вызвать боль практически во всех областях организма. Болевые рецепторы практически не адаптируются.
Передача болевых сигналов
Быстрой и медленной боли соответствуют собственные нервные пути проведения: путь проведения быстрой боли и путь проведения медленной хронической боли.
Проведение быстрой боли
Проведение быстрой боли (рис. 9-5А) от рецепторов осуществляют волокна типа Ad, вступающие в спинной мозг по задним корешкам и синаптически контактирующими с нейронами заднего рога этой же стороны. После образования синапсов с нейронами второго порядка на этой же стороне нервные волокна переходят на противоположную сторону и поднимаются вверх к мозговому ство-
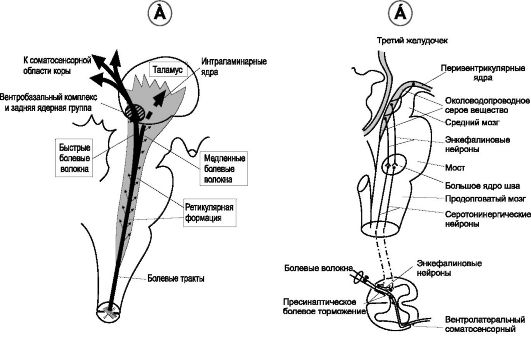
Рис. 9-5. Пути передачи болевой чувствительности (А) и антиноцицептивная система (Б).
лу в составе спиноталамического тракта в переднебоковых канатиках. В стволе мозга часть волокон синаптически контактирует с нейронами ретикулярной формации, основная же масса волокон проходит к таламусу, оканчиваясь в вентро-базальном комплексе вместе с волокнами лемнисковой системы, несущими тактильную чувствительность. Небольшая часть волокон оканчивается в задних ядрах таламуса. Из этих таламических областей сигналы передаются в другие базальные структуры мозга и в соматосенсорную кору (рис. 9-5А).
• Локализация быстрой боли в различных частях тела более чёткая, чем медленной хронической боли.
• Передача болевых импульсов (рис. 9-5Б, 9-6). Глутамат и вещество Р участвует в передаче болевых стимулов в качестве возбуждающего нейромедиатора в синапсах между центральными отростками чувствительных нейронов спинномозгового узла и перикарионами нейронов спиноталамического пути. Блокирование секреции вещества Р и снятие болевых ощущений реализуются через рецепторы опиоидных пептидов, встроенных в мембрану терминали центрального отростка чувствительного нейрона (пример феномена пресинаптического торможения). Источник опиоидного пептида энкефалина - вставочный нейрон.
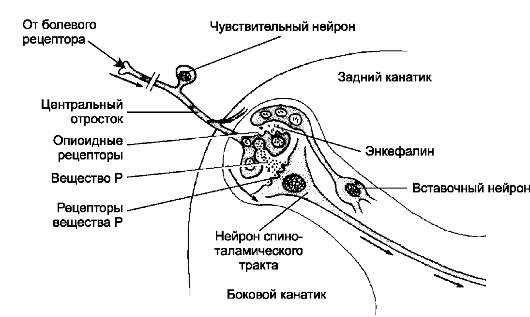 Рис. 9-6. Путь проведения болевых импульсов (стрелки).
Вещество Р пере- даёт возбуждение с центрального отростка
чувствительного нейрона на нейрон спиноталамического тракта. Через
опиоидные рецепторы энкефалин из вставочного нейрона тормозит секрецию
вещества Р из чувствительного нейрона и проведение болевых сигналов.
[11]
Рис. 9-6. Путь проведения болевых импульсов (стрелки).
Вещество Р пере- даёт возбуждение с центрального отростка
чувствительного нейрона на нейрон спиноталамического тракта. Через
опиоидные рецепторы энкефалин из вставочного нейрона тормозит секрецию
вещества Р из чувствительного нейрона и проведение болевых сигналов.
[11]
Проведение медленной хронической боли
Центральные отростки чувствительных нейронов оканчиваются на нейронах пластин II и III. Длинные аксоны вторых нейронов переходят на другую сторону спинного мозга и в составе переднебокового канатика поднимаются в головной мозг. Эти волокна, проводящие сигналы медленной хронической боли в составе палеоспиноталамического тракта, имеют обширные синаптические связи в стволе мозга, оканчиваясь в ретикулярных ядрах продолговатого мозга, моста и среднего мозга, в таламусе, в области покрышки и в сером веществе, окружающем сильвиев водопровод. Из мозгового ствола болевые сигналы поступают к внутрипластинчатым и вентролатеральным ядрам таламуса, гипоталамусу и другим структурам основания мозга (рис. 9-5Б).
• Локализация медленной хронической боли. Медленная хроническая боль локализуется не в отдельных точках тела, а в его больших частях, таких как рука, нога, спина и т.д. Это объясняется полисинаптическими, диффузными связями путей, проводящих медленную боль.
• Центральная оценка медленной боли. Полное удаление соматосенсорной коры у животных не нарушает у них способности ощущать боль. Следовательно, болевые импульсы, входящие в мозг через ретикулярную формацию мозгового ствола, таламус и другие нижележащие центры, могут вызывать осознанное восприятие боли. Соматосенсорная кора участвует в оценке качества боли.
• Нейромедиатор медленной боли в окончаниях C-волокон - вещество P. Болевые волокна типа C, входящие в спинной мозг, в своих окончаниях выделяют нейромедиаторы глутамат и вещество P. Глутамат действует в течение нескольких миллисекунд. Вещество P выделяется медленнее, его действующая концентрация достигается в течение секунд и даже минут.
Система подавления боли
Организм человека не только ощущает и определяет силу и качество болевых сигналов, но и способен понижать и даже подавлять активность болевых систем. Диапазон индивидуальной реакции на боль необыкновенно широк, и ответная реакция на боль в немалой степени зависит от способности мозга подавлять поступающие в нервную систему болевые сигналы при помощи антиноцицептивной (аналгезирующая, антиболевая) системы. Антиноцицептивная система (рис. 9-5Б) состоит из трёх основных компонентов.
1. Комплекс торможения боли, расположенный в задних рогах спинного мозга. Здесь боль блокируется до того, как она достигнет воспринимающих отделов мозга.
2. Большое ядро шва, расположенное по срединной линии между мостом и продолговатым мозгом; ретикулярное парагигантоклеточное ядро, расположенное в боковом отделе продолговатого мозга. Из этих ядер сигналы поступают по заднебоковым столбам в спинной мозг.
3. Околоводопроводное серое вещество и перивентрикулярная область среднего мозга и верхнего отдела моста, окружающие сильвиев водопровод и частично третий и четвёртый желудочки. Нейроны из этих аналгезирующих областей посылают сигналы к большому ядру шва и ретикулярному парагигантоклеточному ядру.
• Нейромедиаторы антиноцицептивной системы. Медиаторами, выделяющимися в окончаниях нервных волокон обезболивающей системы, являются энкефалины и серотонин. Различные отделы аналгезирующей системы чувствительны к морфину, опиатам и опиоидам (b-эндорфину, энкефалинам, динорфину). В частности, энкефалины и динорфин были найдены в структурах аналгезирующей системы мозгового ствола и спинного мозга.
Отражённая боль
Раздражение внутренних органов часто вызывает боль, которая ощущается не только во внутренних органах, но и в некоторых соматических структурах, находящихся достаточно далеко от места вызова боли. Такая боль называется отражённой (иррадиирующей).
Наиболее известным примером отражённой боли является сердечная боль, иррадиирующая в левую руку. Однако будущий врач должен знать, что участки отражения боли не являются стереотипными, а необычные области отражения наблюдаются довольно часто. Сердечная боль, например, может быть чисто абдоминальной, она может иррадиировать в правую руку и даже в шею.
Правило дерматомеров. Афферентные волокна от кожи, мышц, суставов и внутренних органов входят в спинной мозг по задним корешкам в определённом пространственном порядке. Кожные афферентные волокна каждого заднего корешка иннервируют ограниченную область кожи, называемую дерматомером (рис. 9-7). Отражённая боль обычно возникает в структурах, развивающихся из одного и того же эмбрионального сегмента, или дерматомера. Этот принцип называется «правилом дерматомера». Например, сердце и левая рука имеют одну и ту же сегментарную природу, а яичко мигрировало со своим нервным снабжением из урогениталь-
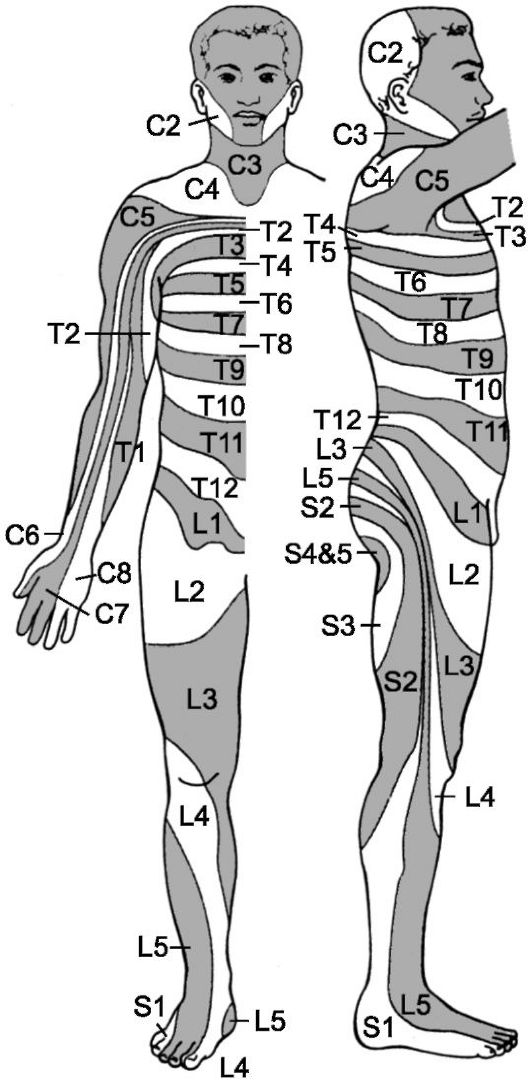 Рис. 9-7. Дерматомеры
Рис. 9-7. Дерматомеры
ного валика, из которого возникли почки и мочеточники. Поэтому не удивительно, что боль, возникшая в мочеточниках или почках, иррадиирует в яичко.
Конвергенция и облегчение в механизме возникновения отражённой боли
В развитии отражённой боли принимают участие не только висцеральные и соматические нервы, входящие в нервную систему на одном сегментарном уровне, но и большое количество сенсорных нервных волокон, проходящих в составе спиноталамических путей. Это создаёт условия для конвергенции периферических афферентных волокон на таламических нейронах, т.е. соматические и висцеральные афференты конвергируют на одних и тех же нейронах (рис. 9-10).
• Теория конвергенции. Большая скорость, постоянство и частота информация о соматической боли способствует закреплению мозгом информации о
• том, что сигналы, поступающие в соответствующие нервные пути, вызваны болевыми стимулами в определённых соматических областях тела. Когда те же нервные пути возбуждаются активностью висцеральных болевых афферентных волокон, то сигнал, достигающий мозга, не дифференцируется, и боль проецируется на соматическую область тела. Теория облегчения. Другая теория происхождения отражённой боли (так называемая теория облегчения) основывается на предположении, что импульсация от внутренних органов понижает порог спиноталамических нейронов к воздействиям афферентных болевых сигналов из соматических областей. В условиях облегчения даже минимальная болевая активность из соматической области проходит в мозг.
ВИСЦЕРАЛЬНАЯ БОЛЬ. В практической медицине боль, возникающая во внутренних органах, является важным симптомом воспаления, инфекционных болезней и других нарушений. Любой сти-
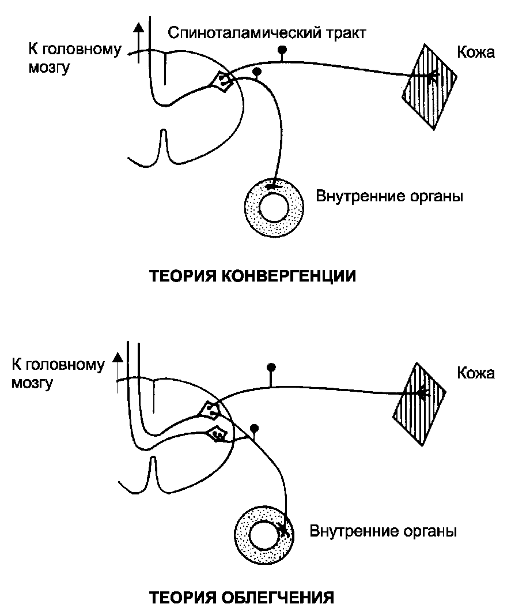 Рис. 9-8. Отражённая боль
Рис. 9-8. Отражённая боль
мул, который чрезмерно возбуждает нервные окончания во внутренних органах, вызывает боль. К ним относятся ишемия висцеральной ткани, химическое повреждение поверхности внутренних органов, спазм гладкой мускулатуры полых органов, растяжение полых органов и растяжение связочного аппарата. Все виды висцеральной боли передаются через болевые нервные волокна, проходящие в составе вегетативных нервов, преимущественно симпатических. Болевые волокна представлены тонкими С-волокнами, проводящими хроническую боль.
ТЕМПЕРАТУРНАЯ ЧУВСТВИТЕЛЬНОСТЬ
Температурные рецепторы. Градации температуры определяются рецепторами холода, рецепторами тепла и болевыми рецепторами. Болевые рецепторы стимулируются экстремальными значениями тепла и холода. Холодовые и тепловые точки кожи располагаются непосредственно под эпидермисом и распределены в виде отдельных точек, имеющих площадь около 1 мм2. В большинстве областей тела холодовых точек (по сравнению с тепловыми) примерно в 5-10 раз больше.
• Тепловые рецепторы - свободные нервные окончания, сигналы от них передают волокнами типа С со скоростью от 0,4 до 2 м/с.
• Холодовые рецепторы - терминальные окончания миелиновых волокон Ad, передающих сигнал со скоростью 20 м/с. Действие перепада температуры. Неожиданное понижение температуры в первый момент сильно стимулирует холодовые окончания. Несколько секунд спустя эффект снижается и продолжает мед-
ленно уменьшаться в течение 30 мин и более, но полной адаптации к изменениям температуры не происходит. Температурные ощущения связаны как с изменениями температуры, так и со стабильным поддержанием температуры. Это значит, что когда температура кожи активно понижается, то человек чувствует больший холод, чем если та же холодная температура остаётся на том же уровне. То же самое можно сказать и о повышении температуры.
Механизм возбуждения температурных рецепторов связан с влиянием на скорость метаболических процессов в них. Так, сдвиг температуры на 10 °С изменяет скорость метаболизма в рецепторах более чем в два раза. Следовательно, детектирование температуры не является результатом прямого физического действия холода или тепла на специальные температурные датчики, а происходит вследствие вызванной изменением температуры химической стимуляции окончаний.
Центральные механизмы. Сигналы от терморецепторов передаются в ЦНС по нервным путям, расположенным параллельно путям передачи боли, и заканчиваются в преоптической области гипоталамуса, ретикулярной формации мозгового ствола и вентробазальном комплексе таламуса.