Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА 8. РЕГИСТРАЦИЯ И ПЕРЕРАБОТКА СИГНАЛОВ
Внешние раздражители, а также сигналы о состоянии внутренней среды и двигательных систем организма регистрируют воспринимающие структуры - сенсорные рецепторы. Эти регистрирующие элементы (рецепторные клетки органов чувств и воспринимающие структуры чувствительных нервных окончаний) преобразуют воздействующие на них различные формы энергии в ПД нервного волокна. Рецепторные клетки органов чувств (фоторецепторы сетчатки, биполярные нейроны обонятельной выстилки, волосковые клетки Кортиева органа и органа равновесия, рецепторные клетки вкусовых луковиц) рассмотрены в главах 10, 11 и 12, чувствительные нервные окончания представлены на рис. 8-1.
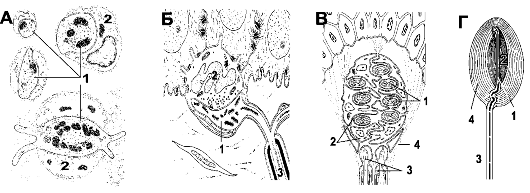 Рис. 8-1. Чувствительные нервные окончания [11]. А - свободные нервные окончания в соединительной ткани. Б - Комплекс клетки Меркеля с нервной терминалью. В - Тельце Майсснера. Г -
инкапсулированное тельце Пачини. 1 - нервная терминаль; 2 -
вспомогательные клетки; 3 - миелиновые нервные волокна; 4 - капсула.
Масштаб между изображениями А-Г не выдержан
Рис. 8-1. Чувствительные нервные окончания [11]. А - свободные нервные окончания в соединительной ткани. Б - Комплекс клетки Меркеля с нервной терминалью. В - Тельце Майсснера. Г -
инкапсулированное тельце Пачини. 1 - нервная терминаль; 2 -
вспомогательные клетки; 3 - миелиновые нервные волокна; 4 - капсула.
Масштаб между изображениями А-Г не выдержан
Основные понятия сенсорной физиологии
Сенсорный рецептор может быть терминалью аксона периферического отростка чувствительного нейрона или контактирующей с терминалью ненервной клеткой (рис. 8-1Б,В), а также рецепторным нейроном (палочки и колбочки сетчатки, обонятельные нейроны). В любом случае в мембране нервной терминали или рецепторном нейроне в ответ на раздражение (сигнал) развивается возбуждение - ответная электрогенная реакция в виде изменения МП (чаще деполяризации - рецепторный потенциал) с последующей генерацией ПД. К видам энергий, преобразуемых сенсорными рецепторами в ПД, относятся механическая (прикосновение, давление), температурная (степени тепла), электромагнитная (свет) и химическая (запах, вкус, содержание O2 или CO2 в крови).
Адекватный стимул. Рецепторные элементы отвечают на изменение конкретной формы энергии низким порогом возбуждения (адекватный стимул). Так, адекватным стимулом для палочек и колбочек сетчатки глаза являются фотоны (ощущение света у человека возникает, когда минимальная интенсивность освещения составляет 10-17-10-18 Вт). Надавливание на глазные яблоки (неадекватный стимул) также может возбудить фоторецепторы и вызвать ощущение световой вспышки, но порог восприятия будет на 13-14 порядков выше адекватного.
Сенсорная единица - чувствительный нейрон и все его периферические разветвления, а также любые ассоциированные с терминалями ненервные клетки, выполняющие функцию преобразователя.
Рецептивное поле - пространственный участок, внутри которого стимулы достаточной величины и адекватные виду раздражаемого рецептора вызовут разряд нервных импульсов в сенсорной единице. Например, рецептивным полем является участок кожи, в которой механические стимулы возбуждают кожные механорецептивные афферентные волокна. Периферические разветвления соседних сенсорных единиц обычно перекрывают друг друга, т.е. рецептивные поля накладываются друг на друга (принцип частичного сме- щённого перекрытия).
Адаптация. Сенсорные волокна различаются характером импульсации на продолжительную стимуляцию. Одни рецепторы отвечают только коротким ответом на включение и выключение стимулов, другие - на постоянную стимуляцию отвечают высокой частотой ПД и поддерживают её всё время стимуляции (подробнее см. на рис. 8-4).
Модальность ощущения. Конкретный сенсорный стимул (например, прикосновение, свет, боль, звук) называют модальностью ощущения. В то же время нервные волокна передают только ПД. Возникает вопрос, как принципиально одинаковые по структуре и выполняемой функции нервные волокна передают в ЦНС различные модальности ощущений? Ответ состоит в том, что каждый нервный путь оканчивается в специальных областях ЦНС, и воспринимаемый вид ощущения детектируется тем участком нервной системы, куда приходят импульсы. Так, если стимулированы нервные волокна, проводящие боль, то человек ощущает боль. Он не различает природы болевых стимулов, которые могут быть электрическими, тепловыми, механическими разрушениями нервного волокна или повреждениями клеток около болевых рецепторов. Точно так же, если рецепторы прикосновения стимулируются электрически или другим путём, человек ощущает прикосновение, потому что нервные волокна оканчиваются в специфической области мозга, интерпретирующей пачки приходящих ПД именно как прикосновение. Подобным же образом волокна от сетчатки оканчиваются в зрительной коре мозга, волокна от уха оканчиваются в слуховой области мозга, температурные волокна оканчиваются в области мозга, определяющей температуру. Такая специфичность нервных волокон, однообразно (в виде ПД) проводящих только одну модальность ощущения, получила название принцип меченой линии.
Классификация рецепторов
Существует несколько классификаций сенсорных рецепторов.
• По модальности: зрение, слух, запах, вкус, ускорение, прикосновение, давление, боль, тепло, холод, напряжение кислорода (pO2), pH, осмотическое давление.
• В зависимости от вида адекватных раздражителей рецепторы классифицируют как механо-, термо-, хеморецепторы, болевые и электромагнитные. Согласно другой классификации, рецепторы подразделяют на дистантные (воспринимающие сигналы на расстоянии); экстерорецепторы (воспринимающие сигналы внешней среды); интерорецепторы, реагирующие на сигналы внутренней среды; проприорецепторы, получающие информацию о положении тела в пространстве.
• В зависимости от наличия рецептирующих (нервных и ненервных) клеток различают первичночувствующие (тактильные, обонятельные, интеро- и проприоцептивные) и вторичночувствующие (слуховые, зрительные, вестибулярные) воспринимающие структуры. Во втором случае рецептирующая клетка формирует
синапс с чувствительным нейроном или со следующим нейроном анализатора (зрение, обоняние).
Трансформация сенсорных раздражений в нервные импульсы
Рецепторный потенциал и генерация потенциалов действия
Вне зависимости от вида раздражающего стимула, который возбуждает сенсорный рецептор, первая его реакция - изменение
МП. Это изменение МП называется рецепторным потенциалом (РП, рис. 8-2).
• Изменения мембранной проницаемости. Различные сенсорные рецепторы могут возбуждаться под влиянием адекватных раздражителей разными путями: открытием ионных каналов (механо- и хеморецепторы), изменением проницаемости мембраны (температурные рецепторы), либо прямым или непрямым изменением характеристик мембраны (воздействие электромагнитных излучений). Во всех случаях главной причиной развития РП являются изменения проницаемости в мембране рецептора.
• Последовательность событий от нанесения адекватного стимула
до возникновения ПД такова: стимул - физико-химические изменения в мембране терминали или преобразования в рецепторной клетке и выделение нейромедиатора - локальные изменения проницаемости - локальная деполяризация (РП) - возникновение и проведение ПД. Эти события рассмотрены на рис. 8-3.
При достижении РП порогового уровня появляется ПД. По мере того как РП превышает свой пороговый уровень, увеличивается и частота ПД. В тельце Пачини (см. рис. 8-1Г) сенсорным элементом является терминаль периферического отростка первичного чувствительного нейрона. Терминаль окружена десятками и сотнями концентрически расположенных клеток, давление на которые (механический стимул) вызывает в терминали изменения ионной проводимости (Na+ входит в терминаль), деполяри-
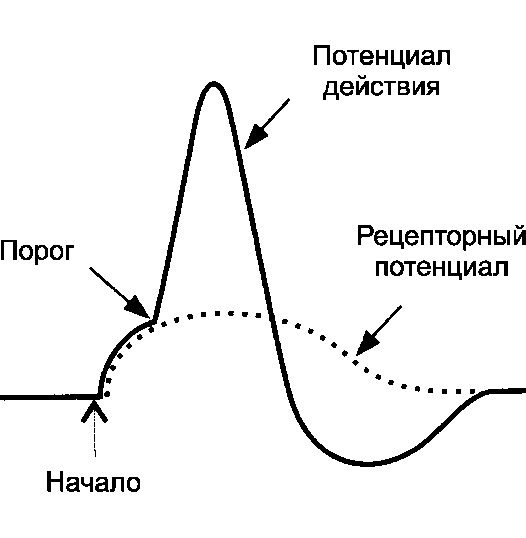
Рис. 8-2. Рецепторный потенциал и потенциал действия в сенсорном рецепторе. Рецепторный потенциал - локальное изменение МП (чаще деполяризация). По достижении критического уровня генерируется ПД
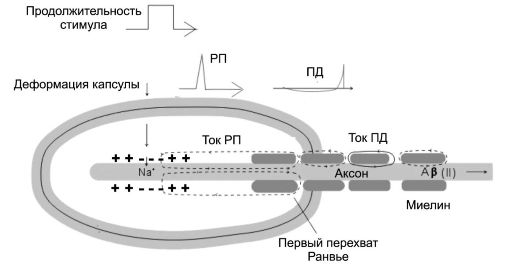 Рис. 8-3. Электрогенез в структурах первичночувствующего механорецептора - тельца Пачини [7]
Рис. 8-3. Электрогенез в структурах первичночувствующего механорецептора - тельца Пачини [7]
зация мембраны приводит к появлению РП, электротонически распространяющегося до первого перехвата Ранвье, где и генерируется ПД.
• Свойства РП. О РП регистрируется в самой нервной терминали, а не в клетках сенсорного органа. О РП изменяется по амплитуде в зависимости от величины механического стимула. О РП - местная реакция, распространяется по мембране электротонически. О Величина РП может увеличиваться посредством временной или пространственной суммации.
Адаптация рецепторов
Все сенсорные рецепторы обладают свойством частично или полностью адаптироваться к длительно действующим раздражителям. Адаптация рецепторов заключается в снижении или прекращении частоты генерируемых ими ПД в ответ на раздражение постоянной силы. Скорость адаптации у различных рецепторов, даже относящихся к одной группе модальности, различна. В зависимости от выраженности процесса адаптации различают быстро- и медленноадаптирующиеся сенсорные рецепторы (рис. 8-4). Так, тельца Пачини адаптируются исключительно быстро, в доли секунды. Рецепторы суставных сумок и рецепторы мышечных веретён адаптируются значительно медленнее. Ещё медленнее адаптируются механорецепторы, расположенные в стенке крупных сосудов (для механорецепторов аорты и каротидного синуса время полной адаптации со-
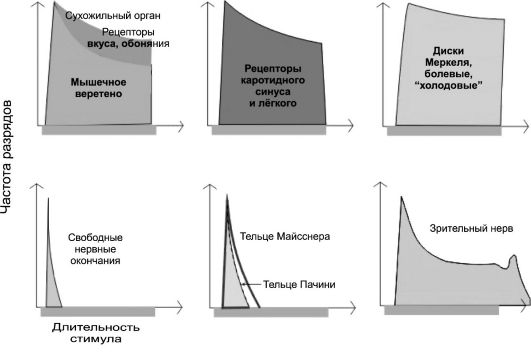 Рис. 8-4. Адаптация сенсорных рецепторов [7]
Рис. 8-4. Адаптация сенсорных рецепторов [7]
ставляет 2 сут - неадаптируемые сенсорные рецепторы). На основании скорости адаптации воспринимающие приборы предложено подразделять на тонические и фазные.
• Тонические рецепторы - медленно адаптируемые рецепторы, постоянно посылающие информацию в мозг о положении тела, его отдельных частей и состоянии внутренней среды. К таким рецепторам относятся мышечные веретёна, сухожильные органы Гольджи, барорецепторы сосудов, болевые рецепторы.
• Фазные рецепторы - быстро адаптируемые рецепторы, реагирующие только в момент действия раздражителя.
НЕЙРОННЫЕ СИСТЕМЫ:
ПРИНЦИПЫ ОРГАНИЗАЦИИ И КОММУНИКАЦИИ
При помощи синапсов нейроны образуют функциональные системы (пулы, центры, ансамбли) различной сложности. Важная функция таких нейронных систем (сетей) - обеспечение коммуникаций: получение, переработка, накопление, извлечение и передача информации, что достигается последовательными процессами кодирования, передачи, расшифровки и нового кодирования. Другая важная функция - поддержание функциональной устойчивости нейронных сетей.
Потоки импульсов через системы нейронов
Потоки информации в функциональных системах формализованы в виде представлений о возбуждаемом поле, конвергенции и дивергенции импульсации, возбуждающих и тормозных цепочках а также о реверберации (самовозбуждении) в нервных центрах.
Возбуждаемое поле. На рис. 8-5А схематически показано функциональное объединение нейронов, имеющее вход слева (1 и 2, афферентные нервные волокна), а выход в виде аксонов (а, б, в, г) справа. Каждое входящее волокно распадается на сотни и тысячи терминалей, которые синаптически контактируют с дендритами или телом нейронов в объединении. Область, иннервируемая каждым входящим волокном, - возбуждаемое, или стимуляционное поле. Из рисунка видно, что значительное количество терминалей от каждого волокна находится в «поле» соседних нейронов, а некоторая часть терминалей направляется к отдалённым нейронам (не показаны).
Пороговые возбуждающие и подпороговые облегчающие стимулы
Одиночный импульс возбуждающей терминали не может вызвать ПД в постсинаптическом нейроне, но большое количество одновременно возбуждаемых терминалей на том же нейроне или их быстрое последовательное раздражение вызовет возбуждение. Если подсчитать количество терминалей от двух афферентов («1» и «2»), оканчивающихся на нейронах, то можно видеть, что входящее во-
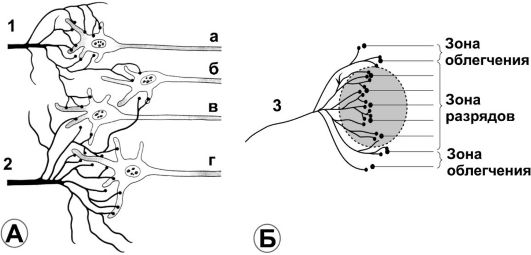 Рис. 8-5. Нейронное объединение (А) и стимуляционное поле (Б). Цифрами обозначены входящие терминали (вход) в нейронное объединение, строчными буквами - выход
Рис. 8-5. Нейронное объединение (А) и стимуляционное поле (Б). Цифрами обозначены входящие терминали (вход) в нейронное объединение, строчными буквами - выход
локно «1» имеет достаточное количество терминалей, чтобы вызвать разряды в нейроне «а». Другими словами, стимулы от афферента «1» к нейрону «а» можно назвать возбуждающими, или надпороговыми стимулами, поскольку они превышают порог, необходимый для возбуждения.
• Входящее волокно «1» распространяет терминали также и к нейронам «б» и «в», но терминалей недостаточно для возбуждения постсинаптических нейронов. Тем не менее суммарное влияние этих терминалей (постсинаптические потенциалы) делают нейроны «б» и «в» более возбудимыми к влияниям, поступающим от других входящих волокон. Такие стимулы можно назвать подпороговыми, а нейроны - облегчающими. Подобным же образом для входящего волокна 2 стимулы, следующие к нейрону «г», будут надпороговыми, а для нейронов «б» и «в» - подпороговыми, но повышающими их возбудимость.
• Рис. 8-5Б детализирует понятие стимуляционного поля. В центральной его части (затемнённый круг) все нейроны стимулируются входящим волокном «3» т.е. эта область является зоной разряда входящего волокна (зона возбуждения, или пороговая зона). Соседние нейроны имеют повышенную возбудимость (облегчение), но они не генерируют разряды на выходе (зона облегчения, или подпороговая зона).
• Аналогично, для тормозных входящих волокон область контакта с нейронами будет называться тормозной зоной. Степень торможения в центре зоны больше, так как здесь имеется значительное количество терминалей.
Дивергенция - свойство нервных клеток контактировать с многими другими нейронами (рис. 8-6, I). Дивергенция нервных импульсов - возможность входящих в нервную систему сигналов возбудить как можно большее количество нервных клеток. Имеется два вида дивергенции: дивергенция усиливающего типа (рис. 8-6, 1А) состоит в том, что входящий сигнал распространяется на увеличивающееся количество нейронов, входящих в состав одной системы. Этот тип дивергенции характерен для кортикоспинальных путей, контролирующих скелетные мышцы. В частности, одна большая пирамидная клетка двигательной коры мозга способна возбудить не менее 10 тыс. скелетных МВ. Дивергенция в различные тракты означает дальнейшую передачу сигнала от получивших импульс нейронов в двух направлениях (рис. 8-6, 1Б). Например, информация, передаваемая по задним столбам спинного мозга, поступает в двух направлениях: в мозжечок, в таламус и далее - кору больших полушарий.
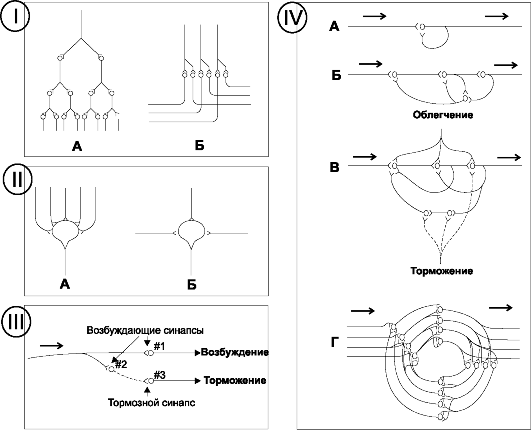 Рис. 8-6. Дивергенция (I), конвергенция (II), тормозная цепочка (III) и реверберирующие круги (IV). IA -
дивергенция внутри одного пути вызывает усиление сигнала. IБ -
дивергенция во многих путях передаёт сигнал в разные области.
Конвергенция многих входов на один нейрон. НА - множественный вход из
одного источника. ПБ - вход из многих источников. Стрелками обозначены направления сигналов
Рис. 8-6. Дивергенция (I), конвергенция (II), тормозная цепочка (III) и реверберирующие круги (IV). IA -
дивергенция внутри одного пути вызывает усиление сигнала. IБ -
дивергенция во многих путях передаёт сигнал в разные области.
Конвергенция многих входов на один нейрон. НА - множественный вход из
одного источника. ПБ - вход из многих источников. Стрелками обозначены направления сигналов
Конвергенция
• Конвергенция - поступление сигналов из многих источников для возбуждения одного нейрона (рис. 8-6, II). Конвергенция из одного источника (рис. 8-6, IIА) происходит в случаях, когда много терминалей из одного волокна оканчиваются на одном и том же нейроне. Нейроны практически никогда не генерируют импульсы от одиночного сигнала, поступающего из одной терминали, но при конвергенции (поступление сигналов по многим терминалям), пространственная суммация доводит уровень МП постсинаптического нейрона до пороговой величины генерации ПД.
• Конвергенция может происходить при поступлении сигналов (возбуждающих и тормозных) из многих источников (рис. 8-6, IIБ). Например, вставочные нейроны спинного мозга получают сигналы от входящих в спинной мозг периферических нервных волокон, от проходящих от одного сегмента спинного мозга в другой проприоспинальных волокон, от кортикоспинальных волокон и некоторых других длинных путей, спускающихся из головного мозга в спинной. Далее сигналы из вставочных нейронов конвергируют на мотонейроны передних рогов спинного мозга. Такого рода конвергенция приводит к суммации информации, поступившей из различных источников, и ответ будет результатом сложения различных видов информации.
Циркуляция возбуждающих и тормозных импульсов. Нередко афферентная импульсация одновременно активирует и возбуждающие, и тормозные нейроны, далее посылающие сигналы к различным эффекторам. Так, в момент когда возбуждающий сигнал передаётся мотонейронам, чтобы вызвать движение ноги вперёд, тормозящий сигнал проходит через отдельную систему нейронов к мышцам задней поверхности ноги, чтобы они не затормозили движение ноги вперёд. Такой тип циркуляции нервных импульсов характерен для контроля антагонистических пар мышц (реципрокное торможение). На рис. 8-6,III проиллюстрирован механизм осуществления реципрокного торможения. Входящее волокно активирует возбуждающий нейрон «1» (посылающий возбуждающие импульсы) и одновременно стимулирует вставочный тормозящий нейрон «2», секреция нейромедиатора из которого блокирует выход сигнала из нейрона «3».
Следовые потенциалы. Электротоническое возбуждение на поверхности дендритов или сомы нейрона может длиться десятки и сотни миллисекунд, особенно когда в процесс вовлечены многие пресинаптические терминали. В течение всего этого времени продолжается возбуждение нейронов и возникновение серии импульсов - синаптический следовой разряд. Одной из важных форм взаимодействия нейронов является реверберация (самовозбуждение), или осцилляторная круговая импульсация. Круговое движение импульсов по цепочке нейронов происходит по принципу положительной обратной связи, в итоге возникают реверберирующие круги возрастающей сложности (рис. 8-6,IV). В самом простом случае в создание реверберирующего круга вовлекается один нейрон (рис. 8-6,IVА), к соме или дендритам которого возбуждение возвращается по коллатерали. Возможно включение нескольких дополнительных нейронов в реверберирующий круг, что увеличивает время между на-
чальным разрядом и сигналом обратной связи. На рис. 8-6,IVБ,В показана ещё более сложная система: облегчающие и тормозные влияния на реверберирующий круг. Облегчающий сигнал усиливает интенсивность и частоту реверберации, а тормозный - уменьшает или останавливает реверберацию. Наконец, многие реверберирующие пути состоят из параллельных волокон (8-6,IVГ). Каждый нейрон, от которого начинается круг, имеет множество широко разветвлённых терминалей. В такой системе общий реверберирующий сигнал может быть слабым или сильным (в зависимости от того, какое количество параллельных волокон вовлечено в реверберацию).
Поддержание функциональной устойчивости сетей
Практически каждая часть мозга прямо или опосредованно соединена с другими его частями, и это создаёт сложные проблемы. Если первая часть возбуждает вторую, вторая - третью и так далее до того момента, когда сигнал снова возбудит первую часть, то мозг может прийти в состояние непрерывного возвратного возбуждения всех частей и окажется бессмысленной массой неконтролируемых реверберирующих сигналов. Такая ситуация наблюдается при эпилептических припадках. Для предупреждения такой генерализации ЦНС использует два основных механизма: тормозные круговые системы и утомление синапсов.
Тормозные круговые системы
Чрезмерное распространение сигналов предотвращают два типа тормозных круговых систем.
1. Тормозные круговые пути, в которых по типу отрицательной обратной связи сигнал возвращается к тому же самому нейрону. Этот тормозной круг наблюдается во всех сенсорных нервных путях, и он тормозит либо вход в сенсорные нейроны тех же самых путей, либо промежуточные нейроны, когда сенсорные пути становятся чрезмерно возбуждёнными.
2. Некоторые отделы мозга (например, базальные ганглии) выполняют специальную тормозную функцию по отношению к большими областями мозга (в частности, оказывают тормозные влияния на системы мозга, контролирующие деятельность мышц). Синаптическое утомление - прогрессивное ослабление синап-
тической передачи в результате интенсивного или продолжительного возбуждения. На рис. 8-7 приведены 3 последовательные записи сгибательного рефлекса, вызванного болевым раздражением внутренней поверхности конечности. Видно, что сила сокраще-
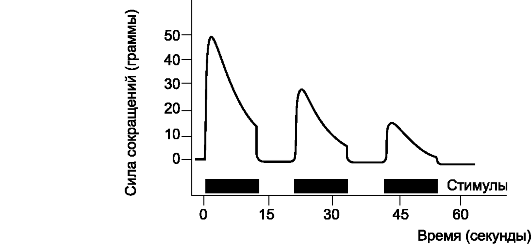 Рис. 8-7. Синаптическое утомление
Рис. 8-7. Синаптическое утомление
ний прогрессивно уменьшается, свидетельствуя о развитии утомления в рефлекторной дуге сгибательного рефлекса. Более того, чем короче интервал между раздражениями, тем меньше сила рефлекторного ответа. Следовательно, чем больше загружен рефлекторный нейронный путь, тем чувствительность системы становится менее выраженной.