Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА 22. ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Пищеварительный тракт (или желудочно-кишечный тракт - ЖКТ) - мышечная трубка, выстланная слизистой оболочкой, просвет трубки - внешняя среда. Слизистая оболочка содержит лимфатические фолликулы и может включать простые экзокринные железы (например, в желудке). Подслизистая оболочка некоторых отделов пищеварительного тракта (пищевод, двенадцатиперстная кишка) имеет сложные железы. Выводные протоки всех экзокринных желёз пищеварительного тракта (включая слюнные, печень и поджелудочную) открываются на поверхности слизистой оболочки. ЖКТ имеет собственный нервный аппарат (энтеральная нервная система) и собственную систему эндокринных клеток (энтероэндокринная система). ЖКТ вместе с его большими железами формирует пищеварительную систему, ориентированную на обработку поступающей пищи (переваривание) и поступление питательных веществ, электролитов и воды во внутреннюю среду организма (всасывание).
Каждая часть ЖКТ выполняет конкретные функции: ротовая полость - жевание и смачивание слюной, глотка - глотание, пищевод - прохождение пищевых комков, желудок - депонирование и начальное переваривание, тонкая кишка - переваривание и всасывание (через 2-4 часа от поступления пищи в ЖКТ), толстая и прямая кишка - подготовку и удаление каловых масс (дефекация происходит от 10 часов до нескольких дней после приёма пищи). Таким образом, пищеварительная система обеспечивает: -о перемещение пищи, содержимого тонкого кишечника (химус) и каловых масс от ротового до анального отверстия; -о секрецию пищеварительных соков и переваривание пищи; -о всасывание переваренных продуктов, воды и электролитов; - движение крови через органы пищеварения и перенос всосавшихся веществ; -о выделение каловых масс; -о гуморальный и нервный контроль всех этих функций.
Нервная регуляция функций ЖКТ
Энтеральная нервная система - совокупность собственных нервных клеток (интрамуральные нейроны общим числом около 100 млн) ЖКТ, а также отростков вегетативных нейронов, расположенных за пределами ЖКТ (экстрамуральные нейроны). Регуляция двигательной и секреторной активности ЖКТ - главная функции энтеральной нервной системы. Стенка ЖКТ содержит мощные сети нервных сплетений.
• Сплетения (рис. 22-1). Собственный нервный аппарат пищеварительного тракта представлен подслизистым и межмышечным сплетениями.
Межмышечное нервное сплетение (Ауэрбаха) расположено в мышечной оболочке пищеварительного тракта, состоит из содержащей ганглии сети нервных волокон. Количество нейронов в ганглии варьирует от единиц до сотен. Межмышечное нервное сплетение необходимо в первую очередь для управления моторикой пищеварительной трубки.
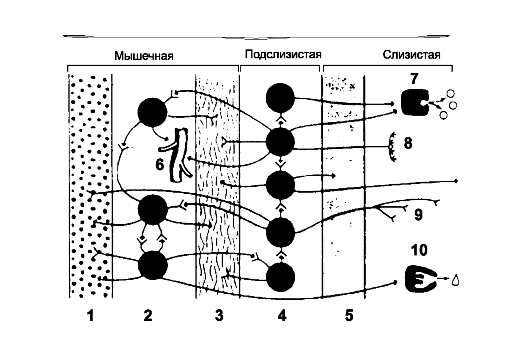 Рис. 22-1. Энтеральная нервная система. 1
- продольный слой мышечной оболочки; 2 - межмышечное (Ауэрбаха) нервное
сплетение; 3 - циркулярный слой мышечной оболочки; 4 - подслизистое
(Майсснера) нервное сплетение; 5 - мышечный слой слизистой оболочки; 6 -
кровеносные сосуды; 7 - эндокринные клетки; 8 - механорецепторы; 9 -
хеморецепторы; 10 - секреторные клетки
Рис. 22-1. Энтеральная нервная система. 1
- продольный слой мышечной оболочки; 2 - межмышечное (Ауэрбаха) нервное
сплетение; 3 - циркулярный слой мышечной оболочки; 4 - подслизистое
(Майсснера) нервное сплетение; 5 - мышечный слой слизистой оболочки; 6 -
кровеносные сосуды; 7 - эндокринные клетки; 8 - механорецепторы; 9 -
хеморецепторы; 10 - секреторные клетки
0 Подслизистое нервное сплетение (Майсснера) расположено в подслизистой оболочке. Это сплетение управляет сокращениями ГМК мышечного слоя слизистой оболочки, а также секрецией желёз слизистой и подслизистой оболочек.
• Иннервация ЖКТ
0 Парасимпатическая иннервация. Возбуждение парасимпатических нервов стимулирует кишечную нервную систему, увеличивая активность пищеварительного тракта. Парасимпатический двигательный путь состоит из двух нейронов.
0 Симпатическая иннервация. Возбуждение симпатической нервной системы тормозит активность пищеварительного тракта. Нейронная цепочка содержит два либо три нейрона.
0 Афференты. Чувствительные хемо- и механорецепторы в оболочках ЖКТ образуют терминальные разветвления собственных нейронов энтеральной нервной системы (клетки Догеля 2-го типа), а также афферентные волокна первичных чувствительных нейронов спинномозговых узлов.
• Гуморальные регуляторные факторы. Помимо классических нейромедиаторов (например, ацетилхолин и норадреналин), нервные клетки энтеральной системы, а также нервные волокна экстрамуральных нейронов секретируют множество биологически активных веществ. Некоторые из них выступают в качестве нейромедиаторов, но большинство - как паракринные регуляторы функций ЖКТ.
• Местные рефлекторные дуги. В стенке пищеварительной трубки присутствует простейшая рефлекторная дуга, состоящая из двух нейронов: чувствительного (клетки Догеля 2-го типа), терминальные разветвления отростков которого регистрируют ситуацию в разных оболочках пищеварительного тракта; и двигательного (клетки Догеля 1-го типа), концевые разветвления аксонов которых образуют синапсы с мышечными и железистыми клетками и регулируют активность этих клеток.
• Желудочно-кишечные рефлексы. Энтеральная нервная система принимает участие во всех рефлексах, контролирующих ЖКТ. По уровню замыкания эти рефлексы подразделяют на местные (1), замыкающиеся на уровне симпатического ствола (2) или на уровне спинного мозга и стволовой части ЦНС (3).
0 1. Местные рефлексы контролируют секрецию желудка и кишечника, перистальтику и другие виды активности ЖКТ.
0 2. К рефлексам, вовлекающим симпатический ствол, относятся желудочно-кишечный рефлекс, вызывающий при активации желудка эвакуацию содержимого толстой кишки; кишечно-желудочный рефлекс, тормозящий секрецию и моторику желудка; ки-
шечно-кишечный рефлекс (рефлекс с ободочной кишки на подвздошную), тормозящий опорожнение содержимого подвздошной кишки в ободочную. 0 3. К рефлексам, замыкающимся на уровне спинного мозга и ствола, относятся рефлексы из желудка и двенадцатиперстной кишки с путями следования в ствол мозга и обратно к желудку через блуждающий нерв (контролируют моторную и секреторную активность желудка); болевые рефлексы, вызывающие общее торможение пищеварительного тракта, и рефлексы дефекации с путями, идущими из ободочной и прямой кишки в спинной мозг и обратно (вызывают необходимые для дефекации сильные сокращения ободочной и прямой кишки и мышц живота).
Гуморальная регуляция функций ЖКТ
Гуморальную регуляцию разнообразных функций ЖКТ осуществляют различные биологически активные вещества информационного характера (нейромедиаторы, гормоны, цитокины, факторы роста и др.), т.е. паракринные регуляторы. К клеткам-мишеням ЖКТ молекулы этих веществ (вещество Р, гастрин, гастрин-рилизинг гормон, гистамин, глюкагон, желудочный ингибирующий пептид, инсулин, метионин-энкефалин, мотилин, нейропептид Y, нейротензин, относящийся к кальцитониновому гену пептид, секретин, серотонин, соматостатин, холецистокинин, эпидермальный фактор роста, VIP, урогастрон) поступают из энтероэндокринных, нервных и некоторых других клеток, расположенных как в стенке ЖКТ, так и за её пределами.
• Энтероэндокринные клетки находятся в слизистой оболочке и особенно многочисленны в двенадцатиперстной кишке. При поступлении пищи в просвет ЖКТ различные эндокринные клетки под действием растяжения стенки, под влиянием самой пищи или изменения pH в просвете ЖКТ начинают выделять гормоны в ткани и в кровь. Активность энтероэндокринных клеток находится под контролем вегетативной нервной системы: стимуляция блуждающего нерва (парасимпатическая иннервация) способствует высвобождению гормонов, усиливающих пищеварение, а повышение активности чревных нервов (симпатическая иннервация) оказывает противоположный эффект.
• Нейроны. Из окончаний нервных волокон секретируется гастринрилизинг гормон; из окончаний нервных волокон, из крови и из собственных (интрамуральных) нейронов ЖКТ поступают пептидные гормоны: нейропептид Y (секретируется вместе с норадреналином), относящийся к кальцитониновому гену пептид.
• Другие источники. Гистамин секретируют тучные клетки, из разных источников поступают серотонин, брадикинин, простагландин Е.
Функции биологически активных веществ в пищеварительном тракте
• Адреналин и норадреналин подавляют перистальтику кишечника и моторику желудка, сужают просвет кровеносных сосудов.
• Ацетилхолин стимулирует все виды секреции в желудке, двенадцатиперстной кишке, поджелудочной железе, а также моторику желудка и перистальтику кишечника.
• Брадикинин стимулирует моторику желудка. Вазодилататор.
• VIP стимулирует моторику и секрецию в желудке, перистальтику и секрецию в кишечнике. Мощный вазодилататор.
• Вещество Р вызывает незначительную деполяризацию нейронов в ганглиях межмышечного сплетения, сокращение ГМК.
• Гастрин стимулирует секрецию слизи, бикарбоната, ферментов, соляной кислоты в желудке, подавляет эвакуацию из желудка, стимулирует перистальтику кишечника и секрецию инсулина, стимулирует рост клеток в слизистой оболочке.
• Гастрин-рилизинг гормон стимулирует секрецию гастрина и гормонов поджелудочной железы.
• Гистамин стимулирует секрецию в железах желудка и перистальтику.
• Глюкагон стимулирует секрецию слизи и бикарбоната, подавляет перистальтику кишечника.
• Желудочный ингибирующий пептид подавляет желудочную секрецию и моторику желудка.
• Мотилин стимулирует моторику желудка.
• Нейропептид Y подавляет моторику желудка и перистальтику кишечника, усиливает вазоконстрикторный эффект норадреналина во многих сосудах, включая чревные.
• Относящийся к кальцитониновому гену пептид подавляет секрецию в желудке, вазодилататор.
• Простагландин Е стимулирует секрецию слизи и бикарбоната в желудке.
• Секретин подавляет перистальтику кишечника, активирует эвакуацию из желудка, стимулирует секрецию сока поджелудочной железы.
• Серотонин стимулирует перистальтику.
• Соматостатин подавляет все процессы в пищеварительном тракте.
• Холецистокинин стимулирует перистальтику кишечника, но подавляет моторику желудка; стимулирует поступление жёлчи в кишечник и секрецию в поджелудочной железе, усиливает высвобожде-
ние инсулина. Холецистокинин имеет значение для процесса медленной эвакуации содержимого желудка, расслабления сфинктера Одди.
• Эпидермальный фактор роста стимулирует регенерацию клеток эпителия в слизистой оболочке желудка и кишечника.
Влияние гормонов на основные процессы в пищеварительном тракте
• Секреция слизи и бикарбоната в желудке. Стимулируют: гастрин, гастрин-рилизинг гормон, глюкагон, простагландин Е, эпидермальный фактор роста. Подавляет соматостатин.
• Секреция пепсина и соляной кислоты в желудке. Стимулируют ацетилхолин, гистамин, гастрин. Подавляют соматостатин и желудочный ингибирующий пептид.
• Моторика желудка. Стимулируют ацетилхолин, мотилин, VIP. Подавляют соматостатин, холецистокинин, адреналин, норадреналин, желудочный ингибирующий пептид.
• Перистальтика кишечника. Стимулируют ацетилхолин, гистамин, гастрин (подавляет эвакуацию из желудка), холецистокинин, серотонин, брадикинин, VIP. Подавляют соматостатин, секретин, адреналин, норадреналин.
• Секреция сока поджелудочной железы. Стимулируют ацетилхолин, холецистокинин, секретин. Подавляет соматостатин.
• Желчеотделение. Стимулируют гастрин, холецистокинин.
МОТОРНАЯ ФУНКЦИЯ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
• Электрические свойства миоцитов. Ритм сокращений желудка и кишечника определяется частотой медленных волн гладких мышц (рис. 22-2А). Эти волны - медленные, волнообразные изменения МП, на гребне которых генерируются потенциалы действия (ПД), которые и вызывают мышечное сокращение. Сокращение возникает тогда, когда МП уменьшается до -40 мВ (МП гладких мышц в покое колеблется от -60 до -50 мВ).
0 Деполяризация. Факторы, деполяризующие мембрану ГМК: ♦ растяжение мышцы, ♦ ацетилхолин, ♦ парасимпатическая стимуляция, ♦ гастроинтестинальные гормоны.
0 Гиперполяризация мембраны миоцитов. Её вызывают адреналин, норадреналин и стимуляция постганглионарных симпатических волокон.
• Виды моторики. Различают перистальтику и перемешивающие движения.
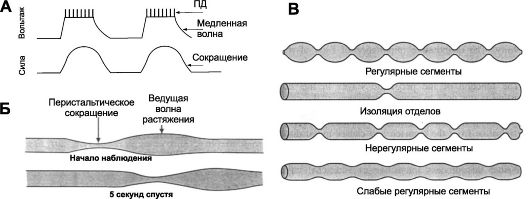 Рис. 22-2. Перистальтика. А. Сверху - медленные волны деполяризации с многочисленными ПД, внизу - запись сокращений [5]. Б. Распространение волны перистальтики. В. Сегментации тонкой кишки
Рис. 22-2. Перистальтика. А. Сверху - медленные волны деполяризации с многочисленными ПД, внизу - запись сокращений [5]. Б. Распространение волны перистальтики. В. Сегментации тонкой кишки
^ Перистальтические движения - продвигающие (пропульсивные) движения. Перистальтика - основной вид двигательной активности, продвигающей пищу (рис. 22-2Б,В). Перистальтическое сокращение - результат выполнения местного рефлекса - перистальтический рефлекс, или миоэнтеральный рефлекс. В норме волна перистальтики продвигается в анальном направлении. Перистальтический рефлекс вместе с анальным направлением движения перистальтики получил название закон кишки. ^ Перемешивающие движения. В некоторых отделах перистальтические сокращения выполняют функцию перемешивания, особенно там, где продвижение пищи задерживается сфинктерами. Могут возникать локальные чередующиеся сокращения, пережимающие кишку от 5 до 30 сек, затем новые пережатия в другом месте и т.д. Перистальтические и пережимающие сокращения приспособлены для продвижения и перемешивания пищи в различных частях пищеварительного тракта. ЖЕВАНИЕ - комбинированное действие жевательных мышц, мышц губ, щёк и языка. Движения этих мышц координируют черепные нервы (V, VII, IX-XII пары). В контроле жевания участвуют не только ядра ствола мозга, но и гипоталамус, миндалина и кора больших полушарий.
• Жевательный рефлекс участвует в произвольно контролируемом акте жевания (регуляция растяжения жевательных мышц).
• Зубы. Передние зубы (резцы) обеспечивают режущее действие, задние зубы (коренные) - перемалывающее. Жевательные мышцы развивают при сжатии зубов силу для резцов в 15 кг и для коренных зубов в 50 кг.
ГЛОТАНИЕ подразделяют на произвольную, глоточную и пищеводную фазы.
• Произвольная фаза начинается с завершения жевания и определения момента готовности пищи к проглатыванию. Пищевой комок продвигается в глотку, надавливая сверху на корень языка и имея сзади мягкое нёбо. С этого момента глотание становится непроизвольным, почти полностью автоматическим.
• Глоточная фаза. Пищевой комок стимулирует рецепторные зоны глотки, нервные сигналы поступают в ствол мозга (центр глотания), вызывая последовательный ряд сокращений мышц глотки.
• Пищеводная фаза глотания отражает основную функцию пищевода - быстрое проведение пищи из глотки в желудок. В норме пищевод имеет два вида перистальтики - первичную и вторичную.
-Ф- Первичная перистальтика - продолжение волны перистальтики, которая начинается в глотке Волна проходит от глотки до желудка в течение 5-10 с. Жидкость проходит быстрее.
-Ф- Вторичная перистальтика. Если первичная перистальтическая волна не может продвинуть всю пищу из пищевода в желудок, то возникает вторичная перистальтическая волна, вызванная растяжением стенки пищевода оставшейся пищей. Вторичная перистальтика продолжается до тех пор, пока вся пища не перейдёт в желудок.
-Ф- Нижний сфинктер пищевода (желудочно-пищеводный гладкомышечный сфинктер) располагается около места соединения пищевода с желудком. В норме происходит тоническое сокращение, предотвращающее попадание содержимого желудка (рефлюкса) в пищевод. В момент движения перистальтической волны по пищеводу сфинктер расслабляется (рецептивное расслабление).
Моторика желудка
В стенке всех отделов желудка сильно развита мышечная оболочка, особенно в привратниковой (пилорической) части. Циркулярный слой мышечной оболочки в месте перехода желудка в двенадцатиперстную кишку образует пилорический сфинктер, постоянно находящийся в состоянии тонического сокращения. Мышечная оболочка обеспечивает двигательные функции желудка - накопление пищи, перемешивание пищи с желудочными секретами и превращение её в полура- створённую форму (химус) и опорожнение химуса из желудка в двенадцатиперстную кишку.
• Голодные сокращения желудка возникают, когда желудок в течение нескольких часов остаётся без пищи. Голодные сокращения - рит-
мические перистальтические сокращения тела желудка - могут сливаться в непрерывное тетаническое сокращение, которое продолжается 2-3 мин. Выраженность голодных сокращений увеличивается при низком уровне сахара в плазме крови.
• Депонирование пищи. Пища поступает в кардиальный отдел отдельными порциями. Новые порции оттесняют предыдущие, что оказывает давление на стенку желудка и вызывает ваго-вагальный рефлекс, уменьшающий тонус мышечной оболочки. В результате создаются условия для поступления новых и новых порций, вплоть до полного расслабления стенки желудка, которое наступает при объёме полости желудка от 1,0 до 1,5 л.
• Перемешивание пищи. В наполненном пищей и расслабленном желудке на фоне медленных спонтанных колебаний МП гладких мышц возникают слабые перистальтические волны - перемешивающие волны. Они распространяются по стенке желудка в направлении привратниковой части каждые 15-20 с. Эти медленные и слабые перистальтические волны на фоне появления ПД сменяются более мощными сокращениями мышечной оболочки (перистальтические сокращения), которые, проходя до пилорического сфинктера, также перемешивает химус.
• Опорожнение желудка. В зависимости от степени переваривания пищи и формирования жидкого химуса перистальтические сокращения становятся всё более сильными, способными не только перемешивать, но и продвигать химус в двенадцатиперстную кишку (рис. 22-3). По мере прогрессирующего опорожнения желудка перистальтические выталкивающие сокращения начинаются от верхних отделов тела и дна желудка, добавляя их содержимое к химусу привратникового отдела. Интенсивность этих сокращений в 5-6 раз больше, чем сила сокращений перемешивающей перистальтики. Каждая сильная волна перистальтики выдавливает несколько
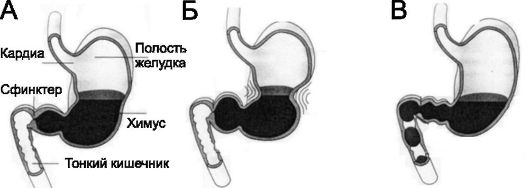 Рис. 22-3. Последовательные фазы опорожнения желудка. А, Б - пилорический сфинктер закрыт. В - пилорический сфинктер открыт
Рис. 22-3. Последовательные фазы опорожнения желудка. А, Б - пилорический сфинктер закрыт. В - пилорический сфинктер открыт
миллилитров химуса в двенадцатиперстную кишку, оказывая пропульсивное насосное действие (пилорический насос).
• Регуляция опорожнения желудка
❖ Темп опорожнения желудка регулируется сигналами из желудка и двенадцатиперстной кишки.
❖ Увеличение объёма химуса в желудке способствует интенсивному опорожнению. Это происходит не из-за повышения давления в желудке, а вследствие реализации местных рефлексов и усиления активности пилорического насоса.
❖ Гастрин, выделяющийся при растяжении стенки желудка, усиливает работу пилорического насоса и потенцирует перистальтическую активность желудка.
❖ Эвакуация содержимого желудка тормозится кишечно-желудочными рефлексами из двенадцатиперстной кишки.
❖ Факторы, вызывающие тормозные кишечно-желудочные рефлексы: кислотность химуса в двенадцатиперстной кишке, растяжение стенки и раздражение слизистой оболочки двенадцатиперстной кишки, возрастание осмоляльности химуса, увеличение концентрации продуктов расщепления белков и жиров.
❖ Холецистокинин, желудочный ингибирующий пептид тормозят опорожнение желудка.
Моторика тонкой кишки
Сокращения гладких мышц тонкого кишечника перемешивают и продвигают химус в просвете кишечника по направлению к толстому кишечнику.
• Перемешивающие сокращения (рис. 22-2В). Растяжение тонкой кишки вызывает перемешивающие сокращения (сегментации). Периодически сдавливая химус с частотой от 2 до 3 раз в минуту (частоту задают медленные электрические волны), сегментации обеспечивают перемешивание пищевых частиц с пищеварительными секретами.
• Перистальтика. Перистальтические волны перемещаются по кишке со скоростью от 0,5 до 2,0 см/сек. Каждая волна затухает через 3- 5 см, поэтому движение химуса происходит медленно (около 1 см/ мин): для его прохождения от пилорического сфинктера до илеоцекальной заслонки требуется от 3 до 5 часов.
• Контроль перистальтики. Поступление химуса в двенадцатиперстную кишку усиливает перистальтику. Такой же эффект оказывают желудочно-кишечный рефлекс, возникающий при растяжении желудка и распространяющийся по межмышечному сплетению от желудка, а также гастрин, холецистокинин, инсулин и серотонин. Секретин и глюкагон тормозят моторику тонкой кишки.
• Илеоцекальный сфинктер (циркулярное утолщение мышечной оболочки) и илеоцекальная заслонка (полулунные складки слизистой оболочки) предотвращают рефлюкс - попадание содержимого толстой кишки в тонкую. Складки заслонки плотно закрываются при повышении давления в слепой кишке, выдерживая давление в 50- 60 см. вод.ст. В нескольких сантиметрах от заслонки мышечная оболочка утолщена, это илеоцекальный сфинктер. Сфинктер в норме не полностью перекрывает просвет кишки, что обеспечивает медленное опорожнение тощей кишки в слепую. Вызванное желудочно-кишечным рефлексом быстрое опорожнение расслабляет сфинктер, что существенно увеличивает перемещение химуса. В норме около 1500 мл химуса ежедневно поступает в слепую кишку.
• Контроль функции илеоцекального сфинктера. Рефлексы из слепой кишки контролируют степень сокращения илеоцекального сфинктера и интенсивность перистальтики тощей кишки. Растяжение слепой кишки усиливает сокращение илеоцекального сфинктера и тормозит перистальтику тощей кишки, задерживая её опорожнение. Эти рефлексы реализуются на уровне энтерального сплетения и экстрамуральных симпатических ганглиев.
Моторика толстого кишечника
В проксимальном отделе толстой кишки происходит преимущественно всасывание (в основном, абсорбция воды и электролитов), в дистальном - накопление каловых масс. Любое раздражение толстой кишки может вызывать интенсивную перистальтику.
• Перемешивающие сокращения. Гладкая мышца продольного слоя мышечной оболочки от слепой до прямой кишки группируется в виде трёх полос, называемых лентами (taenia coli), что придаёт толстой кишке вид сегментарных мешкообразных расширений. Чередование мешкообразных расширений вдоль толстой кишки обеспечивает медленное продвижение, перемешивание и плотный контакт содержимого со слизистой оболочкой. Маятникообразные сокращения происходят преимущественно посегментно, развиваются в течение 30 с и медленно расслабляются.
• Передвигающие сокращения - пропульсивная перистальтика в виде медленных и постоянных маятникообразных сокращений. Требуется не менее 8-15 ч для передвижения химуса от илеоцекальной заслонки через ободочную кишку, чтобы химус превратился в фекальную массу.
• Массированное передвижение. От начала поперечной ободочной к сигмовидной кишке от 1 до 3 раз в день проходит усиленная перистальтическая волна - массированное передвижение, продвигаю-
щее содержимое по направлению к прямой кишке. Во время усиленной перистальтики маятникообразные и сегментарные сокращения толстой кишки временно исчезают. Полная серия усиленных перистальтических сокращений продолжается от 10 до 30 минут. Если фекальные массы продвинуты в прямую кишку, то возникает позыв к дефекации. Возникновение массированного передвижения фекальных масс после приёма пищи ускоряют желудочно-кишечный и дуодено-кишечный рефлексы. Эти рефлексы возникают в результате растяжения желудка и двенадцатиперстной кишки и осуществляются вегетативной нервной системой.
• Другие рефлексы также влияют на моторику толстой кишки. Брюшино-кишечный рефлекс возникает при раздражении брюшины, он сильно тормозит кишечные рефлексы. Почечно-кишечный и пузырно-кишечный рефлексы, возникающие при раздражении почек и мочевого пузыря, тормозят моторику кишечника. Сомато-кишечные рефлексы тормозят моторику кишечника при раздражении кожи поверхности живота.
Дефекация
• Функциональный сфинктер. Обычно прямая кишка свободна от каловых масс. Это результат напряжения функционального сфинктера, расположенного в месте перехода сигмовидной кишки в прямую и наличия острого угла в месте этого перехода, создающего дополнительное сопротивление для наполнения прямой кишки.
• Анальные сфинктеры. Постоянное вытекание фекалий через анальное отверстие предотвращается тоническим сокращением внутреннего и наружного анальных сфинктеров (рис. 22-4А). Внутренний анальный сфинктер - утолщение циркулярной гладкой мышцы, расположенное внутри анального отверстия. Наружный анальный сфинктер состоит из поперечно-полосатых мышц, окружающих внутренний сфинктер. Наружный сфинктер иннервируется соматическими нервными волокнами полового нерва и находится под сознательным контролем. Безусловно-рефлекторный механизм постоянно держит сфинктер сокращённым до тех пор, пока сигналы из коры больших полушарий не затормозят сокращение.
• Рефлексы дефекации. Регуляцию акта дефекации осуществляют рефлексы дефекации.
❖ Собственный ректо-сфинктерный рефлекс возникает при растяжении каловыми массами стенки прямой кишки. Афферентные сигналы через межмышечное нервное сплетение активируют перистальтические волны нисходящей, сигмовидной и прямой кишки, форсируя движение каловых масс к анальному отверстию.
Одновременно расслабляется внутренний анальный сфинктер. Если в это же время поступают сознательные сигналы к расслаблению наружного анального сфинктера, то начинается акт дефекации
❖ Парасимпатический рефлекс дефекации, вовлекающий сегменты спинного мозга (рис. 22-4А), усиливает собственный ректо-сфин- ктерный рефлекс. Сигналы от нервных окончаний в стенке прямой кишки поступают в спинной мозг, обратная импульсация идёт к нисходящей ободочной, сигмовидной и прямой кишке и анальному отверстию по парасимпатическим волокнам тазовых нервов. Эти импульсы значительно усиливают перистальтические волны и расслабление внутреннего и наружного анальных сфинктеров.
❖ Афферентные импульсы, поступающие в спинной мозг во время дефекации, активируют ряд других эффектов (глубокий вдох, смыкание голосовой щели и сокращение мышц передней брюшной стенки).
ГАЗЫ ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА. Источники газов в просвете ЖКТ: заглатывание воздуха (аэрофагия), деятельность бактерий, диффузия газов из крови.
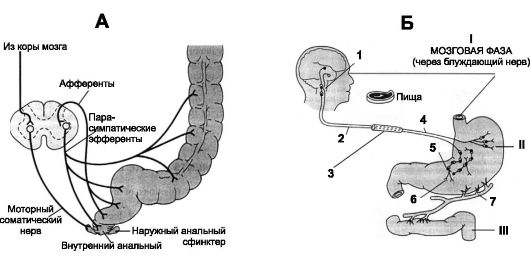
Рис. 22-4. РЕГУЛЯЦИЯ МОТОРИКИ (А) И СЕКРЕЦИИ (Б). А - Парасимпатический механизм рефлекса дефекации. Б - Фазы желудочной секреции. II. Желудочная фаза (местные и вагальные рефлексы, стимуляция выделения гастрина). III. Кишечная фаза (нервный и гуморальный механизмы). 1 - центр блуждающего нерва (продолговатый мозг); 2 - афференты; 3 - ствол блуждающего нерва; 4 - секреторные волокна; 5 - нервные сплетения; 6 - гастрин; 7 - кровеносные сосуды
• Желудок. Газы в желудке - смесь азота и кислорода из заглатываемого воздуха, удаляемого при отрыгивании.
• Тонкий кишечник содержит мало газов, поступающих из желудка. В двенадцатиперстной кишке CO2 накапливается вследствие реакции между соляной кислотой желудочного сока и бикарбонатами поджелудочного сока.
• Толстый кишечник. Основное количество газов (CO2, метан, водород и др.) создаётся деятельностью бактерий. Некоторые виды пищи вызывают значительное выделение газов из анального отверстия: горох, бобы, капуста, огурцы, цветная капуста, уксус. В среднем в толстой кишке каждый день образуется от 7 до 10 л газов и около 0,6 л выталкивается через анальное отверстие. Остающиеся газы абсорбируются слизистой кишечника и выделяются через лёгкие.
СЕКРЕТОРНАЯ ФУНКЦИЯ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Экзокринные железы пищеварительной системы секретируют пищеварительные ферменты от ротовой полости до дистального отдела тощей кишки и выделяют слизь на всём протяжении ЖКТ. Секрецию регулируют вегетативная иннервация и многочисленные гуморальные факторы. Парасимпатическая стимуляция, как правило, стимулирует секрецию, а симпатическая - подавляет.
СЕКРЕЦИЯ СЛЮНЫ. Три пары слюнных желёз (околоушные, нижнечелюстные, подъязычные), а также множество щёчных желёз ежедневно секретируют от 800 до 1500 мл слюны. Гипотоничная слюна содержит серозный компонент (в том числе α-амилазу для переваривания крахмала) и слизистый компонент (в основном муцин для обволакивания пищевого комка и предохранения слизистой оболочки от механического повреждения). Околоушные железы выделяют серозный секрет, нижнечелюстные и подъязычные - слизистый и серозный, щёчные железы - только слизистый. pH слюны колеблется от 6,0 до 7,0. Слюна содержит большое количество факторов, подавляющих рост бактерий (лизоцим, лактоферрин, ионы тиоцианата) и связьвающих Аг (секреторный IgA). Слюна смачивает пищу, обволакивает пищевой комок для более лёгкого прохождения по пищеводу, осуществляет начальный гидролиз крахмала (a-амилаза) и жиров (лингвальная липаза). Стимуляцию секреции слюны осуществляет импульсация, поступающая по парасимпатическим нервным волокнам от верхнего и нижнего слюноотделительных ядер ствола мозга. Эти ядра возбуждаются вкусовыми и тактильными стимулами от языка и других областей ротовой полости и глотки, а также рефлексами, возникающими в желудке и верхнем отделе кишечника. Парасимпатичес-
кая стимуляция также усиливает кровоток в слюнных железах. Симпатическая стимуляция влияет на кровоток в слюнных железах двухфазно: вначале снижает, вызывая сужение сосудов, а затем увеличивает его.
СЕКРЕТОРНАЯ ФУНКЦИЯ ПИЩЕВОДА. Стенка пищевода содержит простые слизистые железы по всему протяжению; а ближе к желудку и в начальной части пищевода - сложные слизистые железы кардиального типа. Секрет желёз предохраняет пищевод от повреждающего действия поступающей пищи и от переваривающего действия забрасываемого в пищевод желудочного сока.
Секреторная функция желудка
Внешнесекреторная функция желудка направлена на защиту стенки желудка от повреждений (в том числе самопереваривания) и на переваривание пищи. Поверхностный эпителий слизистой оболочки желудка вырабатывает муцины (слизь) и бикарбонат, тем самым защищая слизистую оболочку формированием слизисто-бикарбонатного барьера. Слизистая оболочка в различных отделах желудка содержит кардиальные, фундальные и пилорические железы. Кардиальные железы вырабатывают преимущественно слизь, фундальные (80% всех желёз желудка) - пепсиноген, соляную кислоту, внутренний фактор Касла и некоторое количество слизи; пилорические железы секретируют слизь и гастрин.
Слизисто-бикарбонатный барьер
Слизисто-бикарбонатный барьер защищает слизистую оболочку от действия кислоты, пепсина и других потенциальных повреждающих агентов.
• Слизь постоянно секретируется на внутреннюю поверхность стенки желудка.
• Бикарбонат (ионы HCO3-), секретируемый поверхностными слизистыми клетками (рис. 22-5,1), имеет нейтрализующее действие.
• pH. Слой слизи имеет градиент pH. На поверхности слоя слизи pH равен 2, а в примембранной части более 7.
• Н+. Проницаемость плазмолеммы слизистых клеток желудка для Н+ различна. Она незначительна в клеточной мембране, обращён- ной в просвет органа (апикальной), и достаточно высока в базальной части. При механическом повреждении слизистой оболочки и при воздействии на неё продуктов окисления, алкоголя, слабых кислот или жёлчи концентрация H+ в клетках возрастает, что приводит к гибели клеток и разрушению барьера.
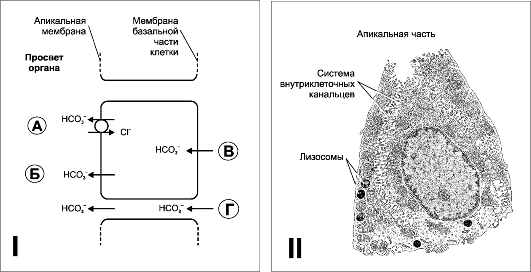 Рис. 22-5. ЖЕЛУДОЧНАЯ СЕКРЕЦИЯ. I -. Механизм секреции НС03~ эпителиальными клетками слизистой оболочки желудка и двенадцатиперстной кишки: А - выход НС03~ в обмен на С1~ стимулируют некоторые гормоны (например, глюкагон), и подавляет блокатор транспорта С1~ фуросемид. Б - активный транспорт НС03~, не зависящий от транспорта С-. В и Г - транспорт НС03~
через мембрану базальной части клетки внутрь клетки и по межклеточным
пространствам (зависит от гидростатического давления в подэпителиальной
соединительной ткани слизистой оболочки). II - Париетальная клетка. Система внутриклеточных канальцев значительно увеличивает площадь поверхности плазматической мембраны. В многочисленных митохондриях вырабатывается АТФ для обеспечения работы ионных насосов плазматической мембраны
Рис. 22-5. ЖЕЛУДОЧНАЯ СЕКРЕЦИЯ. I -. Механизм секреции НС03~ эпителиальными клетками слизистой оболочки желудка и двенадцатиперстной кишки: А - выход НС03~ в обмен на С1~ стимулируют некоторые гормоны (например, глюкагон), и подавляет блокатор транспорта С1~ фуросемид. Б - активный транспорт НС03~, не зависящий от транспорта С-. В и Г - транспорт НС03~
через мембрану базальной части клетки внутрь клетки и по межклеточным
пространствам (зависит от гидростатического давления в подэпителиальной
соединительной ткани слизистой оболочки). II - Париетальная клетка. Система внутриклеточных канальцев значительно увеличивает площадь поверхности плазматической мембраны. В многочисленных митохондриях вырабатывается АТФ для обеспечения работы ионных насосов плазматической мембраны
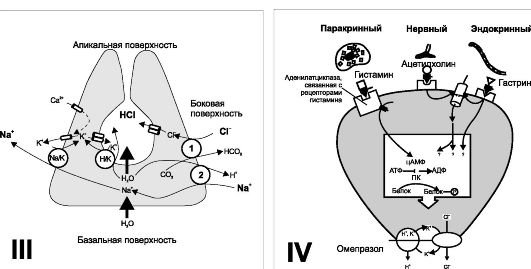 Рис. 22-5. Продолжение. III - Париетальная клетка: транспорт ионов и секреция НС1. Na+,К+-АТФаза участвует в транспорте К+ внутрь клетки. С1~ входит в клетку в обмен на НС03~ через мембрану боковой поверхности (1), а выходит через апикальную мембрану; 2 - обмен Na+ на Н+. Одно из важнейших звеньев - выход Н+ через апикальную мембрану по всей поверхности внутриклеточных канальцев в обмен на К+ при помощи Н+,К+-АТФазы. IV - Регуляции активности париетальных клеток. Стимулирующее влияние гистамина опосредуется через цАМФ, тогда как эффекты ацетилхолина и гастрина - через увеличение притока Са2+
в клетку. Простагландины снижают секрецию НС1, ингибируя
аденилатциклазу, что приводит к уменьшению уровня внутриклеточного цАМФ.
Блокатор Н+,К+-АТФазы (например, омепразол)
снижает выработку НС1. ПК - протеинкиназа, активируемая цАМФ;
фосфорилирует мембранные белки, усиливая работу ионных насосов. [11]
Рис. 22-5. Продолжение. III - Париетальная клетка: транспорт ионов и секреция НС1. Na+,К+-АТФаза участвует в транспорте К+ внутрь клетки. С1~ входит в клетку в обмен на НС03~ через мембрану боковой поверхности (1), а выходит через апикальную мембрану; 2 - обмен Na+ на Н+. Одно из важнейших звеньев - выход Н+ через апикальную мембрану по всей поверхности внутриклеточных канальцев в обмен на К+ при помощи Н+,К+-АТФазы. IV - Регуляции активности париетальных клеток. Стимулирующее влияние гистамина опосредуется через цАМФ, тогда как эффекты ацетилхолина и гастрина - через увеличение притока Са2+
в клетку. Простагландины снижают секрецию НС1, ингибируя
аденилатциклазу, что приводит к уменьшению уровня внутриклеточного цАМФ.
Блокатор Н+,К+-АТФазы (например, омепразол)
снижает выработку НС1. ПК - протеинкиназа, активируемая цАМФ;
фосфорилирует мембранные белки, усиливая работу ионных насосов. [11]
• Регуляция. Секрецию бикарбоната и слизи усиливают глюкагон, простагландин Е, гастрин, эпидермальный фактор роста. Для предупреждения повреждения и для восстановления повреждённого барьера применяют антисекреторные агенты (например, блокаторы гистаминовых рецепторов), простагландины, гастрин, аналоги сахаров (например, сукральфат).
• Разрушение барьера. При неблагоприятных условиях барьер разрушается в течение нескольких минут, происходят гибель клеток эпителия, отёк и кровоизлияния в собственном слое слизистой оболочки. Известны факторы, неблагоприятные для поддержания барьера: -Фнестероидные противовоспалительные препараты (например, аспирин, индометацин); -Фэтанол; -Фсоли жёлчных кислот; -Ф- Helicobacter pylori - грамотрицательная бактерия, выживающая в кислой среде желудка. H. pylori поражает поверхностный эпителий желудка и разрушает барьер, способствуя развитию гастрита и язвенного дефекта стенки желудка. Этот микроорганизм выделяют у 70% больных язвенной болезнью желудка и 90% больных язвой двенадцатиперстной кишки.
• Регенерация эпителия, формирующего слой бикарбонатной слизи, происходит за счёт стволовых клеток, расположенных на дне желудочных ямок; время обновления клеток - около 3 суток. Стимуляторы регенерации: о гастрин из эндокринных клеток желудка; о гастрин-рилизинг гормон из эндокринных клеток и окончаний волокон блуждающего нерва; о эпидермальный фактор роста, поступающий из слюнных, пилорических желёз, желёз двенадцатиперстной кишки и других источников.
Слизь. Помимо поверхностных клеток слизистой оболочки желудка, слизь секретируют клетки практически всех желёз желудка.
Пепсиноген. Главные клетки фундальных желёз синтезирует и секретирует предшественники пепсина (пепсиноген), а также небольшое количество липазы и амилазы. Пепсиноген не обладает пищеварительной активностью. Под влиянием соляной кислоты и особенно ранее образованного пепсина пепсиноген превращается в активный пепсин. Пепсин - протеолитический фермент, активный в кислой среде (оптимум pH от 1,8 до 3,5). При pH около 5 он практически не имеет протеолитической активности и за короткое время полностью инактивируется.
Внутренний фактор. Для всасывания витамина B12 в кишечнике необходим (внутренний) фактор Касла, синтезируемый париетальными клетками желудка. Фактор связывает витамин B12 и защищает его от разрушения ферментами. Комплекс внутреннего фактора с витамином B12 в присутствии ионов Ca2+ взаимодействует с рецепторами эпите-
лиальной клетки дистального отдела подвздошной кишки. При этом витамин B12 поступает в клетку, а внутренний фактор высвобождается. Отсутствие внутреннего фактора приводит к развитию анемии.
Соляная кислота
Соляную кислоту (HCl) вырабатывают париетальные клетки, имеющие мощную систему внутриклеточных канальцев (рис. 22-5,11), существенно увеличивающих секреторную поверхность. Клеточная мембрана, обращенная в просвет канальцев, содержит протонный насос (Н+,К+-ЛТФаза), выкачивающий из клетки Н+ в обмен на К+. Хлорно-бикарбонатный анионообменник встроен в мембрану боковой и базальной поверхности клеток: Cl- входит в клетку в обмен на HCO3- через этот анионообменник и выходит в просвет канальцев. Таким образом, в просвете канальцев оказываются оба компонента соляной кислоты: и Cl-, и H+. Все остальные молекулярные компоненты (ферменты, ионные насосы, трансмембранные переносчики) направлены на сохранение ионного баланса внутри клетки, прежде всего - на поддержание внутриклеточного pH.
Регуляция секреции соляной кислоты представлена на рис. 22-5,IV. Париетальная клетка активируется через мускариновые холинорецепторы (блокатор - атропин), Н2-рецепторы гистамина (блокатор - циметидин) и рецепторы гастрина (блокатор - проглюмид). Указанные блокаторы или их аналоги, а также ваготомия используются для подавления секреции соляной кислоты. Существует ещё один способ понизить выработку соляной кислоты - блокада Н+,К+-АТФазы.
Желудочная секреция
Клинические понятия «желудочная секреция», «желудочный сок» подразумевают секрецию пепсина и секрецию соляной кислоты, т.е. сочетанную секрецию пепсина и соляной кислоты.
• Стимуляторы секреции желудочного сока: о пепсин (оптимум ферментной активности при кислых значениях pH); о Cl- и H+ (соляная кислота); о гастрин; о гистамин; о ацетилхолин.
• Ингибиторы и блокаторы секреции желудочного сока: о желудочный ингибирующий пептид; о секретин; о соматостатин; о блокаторы рецепторов гастрина, секретина, гистамина и ацетилхолина.
Фазы желудочной секреции
Желудочная секреция осуществляется в три фазы - мозговую, желудочную и кишечную (рис. 22-4Б).
• Мозговая фаза начинается до поступления пищи в желудок, в момент приёма пищи. Вид, запах, вкус пищи усиливают секрецию
желудочного сока. Нервные импульсы, вызывающие мозговую фазу, происходят из коры больших полушарий и центров голода в гипоталамусе и миндалине. Они передаются через моторные ядра блуждающего нерва и затем через его волокна к желудку. Секреция желудочного сока в эту фазу составляет до 20% секреции, связанной с приемом пищи.
• Желудочная фаза начинается с момента поступления пищи в желудок. Поступившая пища вызывает ваго-вагальные рефлексы, местные рефлексы энтеральной нервной системы, выделение гастрина. Гастрин в течение нескольких часов пребывания пищи в желудке стимулирует секрецию желудочного сока. Количество сока, выделяющегося в желудочную фазу, составляет 70% от общей секреции желудочного сока (1500 мл).
• Кишечная фаза связана с поступлением пищи в двенадцатиперстную кишку, что вызывает небольшой подъём секреции желудочного сока (10%) за счёт выделения гастрина из слизистой оболочки кишки под влиянием растяжения и действия химических стимулов.
Регуляция желудочной секреции кишечными факторами
• Пища, поступившая из желудка в тонкую кишку, тормозит секрецию желудочного сока. Нахождение пищи в тонкой кишке вызывает тормозный кишечно-желудочный рефлекс, осуществляемый через энтеральную нервную систему, симпатические и парасимпатические волокна. Рефлекс инициируется растяжением стенки тонкой кишки, присутствием кислоты в краниальном отделе тонкой кишки, наличием продуктов белкового расщепления и раздражением слизистой оболочки кишки. Этот рефлекс - часть комплексного рефлекторного механизма, замедляющего переход пищи из желудка в двенадцатиперстную кишку.
• Присутствие кислоты, продуктов жирового и белкового расщепления, гиперили гипоосмотических жидкостей или любых других раздражающих факторов в краниальных отделах тонкой кишки вызывает выделение нескольких кишечных пептидных гормонов - секретина, желудочного ингибирующего пептида и VIP. Секретин - важнейший фактор стимуляции секреции поджелудочной железы - тормозит секрецию желудка. Желудочный ингибирующий пептид, VIP и соматостатин оказывают умеренное тормозное влияние на желудочную секрецию. В результате торможение желудочной секреции кишечными факторами приводит к замедлению поступления химуса из желудка в кишечник, когда тот уже заполнен. Секреция желудка после приёма пищи. Секреция желудка через некоторое время после приёма пищи (2-4 часа) составляет несколько
миллилитров желудочного сока за каждый час «межпищеварительного периода». В основном секретируется слизь и следы пепсина, практически без соляной кислоты. Однако эмоциональные стимулы часто увеличивают секрецию до 50 мл и более в час с высоким содержанием пепсина и соляной кислоты.
Секреторная функция поджелудочной железы
Ежедневно поджелудочная железа секретирует около 1 л литра сока. Сок поджелудочный железы (ферменты и бикарбонаты) в ответ на опорожнение желудка оттекает по длинному выводному протоку. Этот проток, соединившись с общим жёлчным протоком, формирует печё- ночно-поджелудочную ампулу, которая открывается на большом дуоденальном (фатеровом) сосочке в двенадцатиперстную кишку, будучи окружена жомом из ГМК (сфинктер Одди). Поступающий в просвет кишки сок поджелудочной железы содержит пищеварительные ферменты, необходимые для переваривания углеводов, белков и жиров, и большое количество бикарбонатных ионов, обеспечивающих нейтрализацию кислого химуса.
• Протеолитические ферменты - трипсин, химотрипсин, карбоксипептидаза, эластаза, а также расщепляющие макромолекулы ДНК и РНК нуклеазы. Трипсин и химотрипсин расщепляют белки до пептидов, а карбоксипептидаза расщепляет пептиды на отдельные аминокислоты. Протеолитические ферменты находятся в неактивной форме (трипсиноген, химотрипсиноген и прокарбоксипептидаза) и становятся активными только после поступления в просвет кишки. Трипсиноген активирует энтерокиназа из клеток слизистой оболочки кишечника, а также трипсин. Химотрипсиноген активируется трипсином, а прокарбоксипептидаза - карбоксипептидазой.
• Липазы. Жиры расщепляет панкреатическая липаза (гидролизует триглицериды, ингибитор липазы - соли жёлчных кислот), холестерол-эстераза (гидролизует эфиры холестерола) и фосфолипаза (отщепляет жирные кислоты от фосфолипидов).
• α-Амилаза (панкреатическая) расщепляет крахмал, гликоген и большинство углеводов до ди- и моносахаридов.
• Бикарбонат ные ионы секретируют эпителиальные клетки мелких и средних протоков. Механизм секреции HCO3- рассмотрен на рис.
22-5,I.
• Фазы секреции поджелудочной железы такие же, как и желудочной секреции - мозговая (20% всей секреции), желудочная (5-10%) и кишечная (75%).
• Регуляция секреции. Секрецию сока поджелудочной железы стимулируют ацетилхолин и парасимпатическая стимуляция, холецистокинин, секретин (особенно при очень кислом химусе) и прогестерон. Действие стимуляторов секреции обладает эффектом мультипликации, то есть эффект одновременного действия всех стимулов намного больше, чем сумма эффектов каждого стимула по отдельности.
Секреция жёлчи
Одной из многообразных функций печени является желчеобразующая (от 600 до 1000 мл в день). Жёлчь - сложный водный раствор, состоящий из органических соединений и неорганических веществ. Основные компоненты жёлчи - холестерин, фосфолипиды (главным образом лецитин), соли жёлчных кислот (холаты), жёлчные пигменты (билирубин), неорганические ионы и вода. Жёлчь (первая порция жёлчи) постоянно секретируется гепатоцитами и по системе протоков (здесь к жёлчи добавляется стимулируемая секретином вторая порция, содержащая много ионов бикарбоната и натрия) поступает в общий печёночный и далее в общий жёлчный проток. Отсюда печё- ночная жёлчь опорожняется непосредственно в двенадцатиперстную кишку или же поступает в пузырный проток, ведущий в жёлчный пузырь. Жёлчный пузырь накапливает и концентрирует жёлчь. Из жёлчного пузыря концентрированная жёлчь (пузырная жёлчь) по пузырному и далее по общему жёлчному протоку порциями выбрасывается в просвет двенадцатиперстной кишки. В тонком кишечнике жёлчь участвует в гидролизе и всасывании жиров.
• Концентрирование жёлчи. Объём жёлчного пузыря - от 30 до 60 мл,
но за 12 часов в жёлчном пузыре может депонироваться до 450 мл печёночной жёлчи, так как вода, натрий, хлориды и другие электролиты постоянно всасываются через слизистую оболочку пузыря. Основный механизм абсорбции - активный транспорт натрия с последующим вторичным транспортом ионов хлора, воды и других компонентов. Жёлчь концентрируется в 5 раз, максимально - в 20 раз.
• Опорожнение жёлчного пузыря за счёт ритмических сокращений его стенки происходит тогда, когда пища (особенно жирная) поступает в двенадцатиперстную кишку. Эффективное опорожнение жёлчного пузыря наступает при одновременном расслаблении сфинктера Одди. Прием значительного количества жирной пищи стимулирует полное опорожнение жёлчного пузыря в течение 1 часа. Стимулятор опорожнения жёлчного пузыря - холецистокинин, дополнительные стимулы поступают от холинергических волокон блуждающего нерва.
• Функции жёлчных кислот. Ежедневно гепатоциты синтезируют около 0,6 г гликохолевых и таурохолевых жёлчных кислот. Жёлчные кислоты - детергенты, они уменьшают поверхностное натяжение жировых частиц, что приводит к эмульгированию жира. Более того, жёлчные кислоты способствуют всасыванию жирных кислот, моноглицеридов, холестерола и других липидов. Без жёлчных кислот более 40% липидов пищи теряется с калом.
• Кишечно-печёночная циркуляция жёлчных кислот. Жёлчные кислоты всасываются из тонкой кишки в кровь и через воротную вену поступают в печень. Здесь они почти полностью абсорбируются гепатоцитами и снова секретируются в жёлчь. Таким способом жёлч- ные кислоты циркулируют до 18 раз, прежде чем они постепенно не будут удалены с калом. Этот процесс называется кишечно-печё- ночной циркуляцией.
Секреторная функция тонкой кишки
Ежедневно в тонком кишечнике образуется до 2 л секрета (кишечный сок) с pH от 7,5 до 8,0. Источники секрета - железы подслизистой оболочки двенадцатиперстной кишки (бруннеровы железы) и часть эпителиальных клеток ворсинок и крипт.
• Бруннеровы железы секретируют слизь и бикарбонаты. Слизь, выделяемая бруннеровыми железами, защищает стенку двенадцатиперстной кишки от действия желудочного сока и нейтрализует соляную кислоту, поступающую из желудка.
• Эпителиальные клетки ворсинок и крипт. Бокаловидные клетки секретируют слизь, а энтероциты выделяют в просвет кишки воду, электролиты и ферменты.
• Ферменты. На поверхности энтероцитов в ворсинках тонкой кишки находятся пептидазы (расщепляют пептиды до аминокислот), дисахаридазы сукраза, мальтаза, изомальтаза и лактаза (расщепляют дисахариды на моносахариды) и кишечная липаза (расщепляет нейтральные жиры до глицерина и жирных кислот).
• Регуляция секреции. Секрецию стимулируют механическое и химическое раздражение слизистой оболочки (местные рефлексы), возбуждение блуждающего нерва, гастроинтестинальные гормоны (особенно холецистокинин и секретин). Секрецию тормозят влияния со стороны симпатической нервной системы.
Секреторная функция толстой кишки. Крипты толстой кишки выделяют слизь и бикарбонаты. Величину секреции регулируют механическое и химическое раздражение слизистой оболочки и локальные рефлексы энтеральной нервной системы. Возбуждение парасимпатических волокон тазовых нервов вызывает увеличение отделения сли-
зи с одновременной активацией перистальтики толстой кишки. Сильные эмоциональные факторы могут стимулировать акты дефекации с периодическим выделением слизи без фекального содержимого («медвежья болезнь»).
ПЕРЕВАРИВАНИЕ ПИЩИ
Белки, жиры и углеводы в пищеварительном тракте превращаются в продукты, способные всасываться (пищеварение, переваривание). Продукты пищеварения, витамины, минералы и вода проходят сквозь эпителий слизистой оболочки и поступают в лимфу и кровь (всасывание). Основу пищеварения составляет химический процесс гидролиза, осуществляемый пищеварительными ферментами.
• Углеводы. В пище содержатся дисахариды (сахароза и мальтоза) и полисахариды (крахмалы, гликоген), а также другие органические соединения углеводного характера. Целлюлоза в пищеварительном тракте не переваривается, так как у человека нет ферментов, способных её гидролизовать.
о Ротовая полость и желудок. α-Амилаза расщепляет крахмал до дисахарида - мальтозы. За короткое время пребывания пищи в ротовой полости переваривается не более 5% всех углеводов. В желудке углеводы продолжают перевариваться в течение часа, прежде чем пища полностью перемешается с желудочным соком. За этот период до 30% крахмалов гидролизуется до мальтозы.
о Тонкая кишка. α-Амилаза панкреатического сока заканчивает расщепление крахмалов до мальтозы и других дисахаридов. Содержащиеся в щёточной каёмке энтероцитов лактаза, сахараза, мальтаза и α-декстриназа гидролизуют дисахариды. Мальтоза расщепляется до глюкозы; лактоза - до галактозы и глюкозы; сахароза - до фруктозы и глюкозы. Образовавшиеся моносахариды всасываются в кровь.
• Белки
о Желудок. Пепсин, активный при pH от 2,0 до 3,0, превращает 10-20% белков в пептоны и некоторое количество полипептидов. о Тонкая кишка
♦ Ферменты поджелудочной железы трипсин и химотрипсин в просвете кишки расщепляют полипептиды на ди- и трипептиды, карбоксипептидаза отщепляет аминокислоты от карбоксильного конца полипептидов. Эластаза переваривает эластин. В целом образуется немного свободных аминокислот.
♦ На поверхности микроворсинок каёмчатых энтероцитов в двенадцатиперстной и тощей кишке находится трёхмерная густая сеть - гликокаликс, в котором расположены многочисленные
пептидазы. Именно здесь эти ферменты осуществляют так называемое пристеночное пищеварение. Аминополипептидазы и дипептидазы расщепляют полипептиды на ди- и трипептиды, а ди- и трипептиды превращают в аминокислоты. Затем аминокислоты, дипептиды и трипептиды легко транспортируются внутрь энтероцитов через мембрану микроворсинок.
♦ В каёмчатых энтероцитах имеется множество пептидаз, специфичных для связей между конкретными аминокислотами; в течение нескольких минут все оставшиеся ди- и трипептиды превращают в отдельные аминокислоты. В норме более 99% продуктов переваривания белков всасывается в виде отдельных аминокислот. Очень редко всасываются пептиды.
• Жиры находятся в пище преимущественно в виде нейтральных жиров (триглицеридов), а также фосфолипидов, холестерола и эфиров холестерола. Нейтральные жиры входят в состав пищи животного происхождения, их значительно меньше в растительной пище. о Желудок. Липазы расщепляют менее 10% триглицеридов. о Тонкая кишка
♦ Переваривание жиров в тонкой кишке начинается с превращения крупных жировых частиц (глобул) в мельчайшие глобулы - эмульгирование жиров (рис. 22-7А). Этот процесс начинается в желудке под влиянием перемешивания жиров с желудочным содержимым. В двенадцатиперстной кишке жёл- чные кислоты и фосфолипид лецитин эмульгируют жиры до размеров частиц в 1 мкм, увеличивая общую поверхность жиров в 1000 раз.
♦ Панкреатическая липаза расщепляет триглицериды на свободные жирные кислоты и 2-моноглицериды и способна в течение 1 минуты переварить все триглицериды химуса, если они находятся в эмульгированном состоянии. Роль кишечной липазы в переваривании жиров невелика. Накопление моноглицеридов и жирных кислот в местах переваривания жиров останавливает процесс гидролиза, но этого не происходит, потому что мицеллы, состоящие из нескольких десятков молекул жёл- чных кислот, удаляют моноглицериды и жирные кислоты в момент их образования (рис. 22-7А). Мицеллы холатов транспортируют моноглицериды и жирные кислоты к микроворсинкам энтероцитов, где они всасываются.
♦ Фосфолипиды содержат жирные кислоты. Эфиры холестерола и фосфолипиды расщепляются специальными липазами поджелудочного сока: холестерол-эстераза гидролизует эфиры холестерола, а фосфолипаза Л2 расщепляет фосфолипиды.
ВСАСЫВАНИЕ В ПИЩЕВАРИТЕЛЬНОМ ТРАКТЕ
Всасывание - движение воды и растворённых в ней веществ - продуктов пищеварения, а также витаминов и неорганических солей из просвета кишечника через однослойный каёмчатый эпителий в кровь и лимфу. Реально всасывание происходит в тонком и отчасти в толстом кишечнике, в желудке всасываются только жидкости, в том числе алкоголь и вода.
Всасывание в тонкой кишке
• В слизистой оболочке тонкого кишечника имеются циркулярные складки, ворсинки и крипты. За счёт складок площадь всасывания увеличивается в 3 раза, за счёт ворсинок и крипт - в 10 раз и за счёт микроворсинок каёмчатых клеток - в 20 раз. Суммарно складки, ворсинки, крипты и микроворсинки обеспечивают увеличение площади всасывания в 600 раз, а общая всасывающая поверхность тонкой кишки достигает 200 м2. Однослойный цилиндрический каёмчатый эпителий содержит каёмчатые, бокаловидные, энтероэндокринные, панетовские и камбиальные клетки. Всасывание происходит через каёмчатые клетки. Каёмчатые клетки (энтероциты) имеют более 1000 микроворсинок на апикальной поверхности. Именно здесь присутствует гликокаликс. Эти клетки всасывают расщеплённые белки, жиры и углеводы. о Микроворсинки образуют всасывательную, или щёточную каём- ку на апикальной поверхности энтероцитов. Через всасывательную поверхность происходит активный и избирательный транспорт из просвета тонкого кишечника через каёмчатые клетки, через базальную мембрану эпителия, через межклеточное вещество собственного слоя слизистой оболочки, через стенку кровеносных капилляров в кровь, а через стенку лимфатических капилляров (тканевые щели) - в лимфу. о Межклеточные контакты. Поскольку всасывание аминокислот, сахаров, глицеридов и т.д. происходит через клетки, и внутренняя среда организма далеко не безразлична к содержимому кишечника (напомним, что просвет кишечника - внешняя среда), возникает вопрос, каким образом предупреждается проникновение содержимого кишечника во внутреннюю среду по пространствам между клетками эпителия. «Закрытие» реально существующих межклеточных пространств осуществляется за счёт специализированных межклеточных контактов, перекрывающих щели между эпителиальными клетками. Каждая клетка в пласте эпителия по всей окружности в апикальной области имеет сплошной пояс плотных контактов, предупреждающих поступление содержимого кишечника в межклеточные щели.
о Вода. Гипертоничность химуса вызывает движение воды из плазмы в химус, само же трансмембранное перемещение воды происходит посредством диффузии, подчиняясь законам осмоса. Каёмчатые клетки крипт выделяют в просвет кишки Cl-, что инициирует поток Na+, других ионов и воды в том же направлении. В то же время клетки ворсинок «накачивают» Na+ в межклеточное пространство и таким образом компенсируют перемещение Na+ и воды из внутренней среды в просвет кишечника. Микроорганизмы, приводящие к развитию диареи, вызывают потерю воды путём угнетения процесса поглощения Na+ клетками ворсинок и усиления гиперсекреции Cl- клетками крипт. Ежедневный оборот воды в пищеварительном канале - приход равен расходу - составляет 9 л.
о Натрий. Ежедневное поступление от 5 до 8 г натрия. От 20 до 30 г натрия секретируется с пищеварительными соками. Для предотвращения потерь натрия, выделяемого с калом, кишечнику необходимо всасывать от 25 до 35 г натрия, что примерно равно 1/7 общего содержания натрия в организме. Большая часть Na+ всасывается посредством активного транспорта (рис. 22-6). Активный транспорт Na+ связан с всасыванием глюкозы, некоторых аминокислот и ряда других веществ. Присутствие глюкозы в кишечнике облегчает реабсорбцию Na+. Это является физиологической основой для восстановления потерь воды и Na+ при диарее путём питья подсолённой воды с глюкозой. Обезвоживание увеличивает секрецию альдостерона. Альдостерон в течение 2-3 часов активирует все механизмы усиления всасывания Na+. Повышение абсорбции Na+ влечёт за собой увеличение всасывания воды, Cl- и других ионов.
о Хлор. Ионы Cl- секретируются в просвет тонкой кишки через ионные каналы, активируемые цАМФ. Энтероциты всасывают Cl- вместе с Na+ и K+, и натрий служит переносчиком (рис. 22- 6,III). Движение Na+ через эпителий создаёт электронегативность химуса и электропозитивность в межклеточных пространствах. Ионы Cl- движутся вдоль этого электрического градиента, «следуя» за ионами Na+.
о Бикарбонат. Всасывание бикарбонатных ионов ассоциировано с всасыванием ионов Na+. В обмен на всасывание Na+, ионы H+ секретируются в просвет кишечника, соединяются с бикарбонатными ионами и образуют H2CO3 которая диссоциирует на H2O и CO2. Вода остаётся в химусе, а углекислый газ всасывается в кровь и выделяется лёгкими.
о Калий. Некоторое количество ионов K+ секретируются вместе со слизью в полость кишечника; большая часть ионов K+ всасыва-
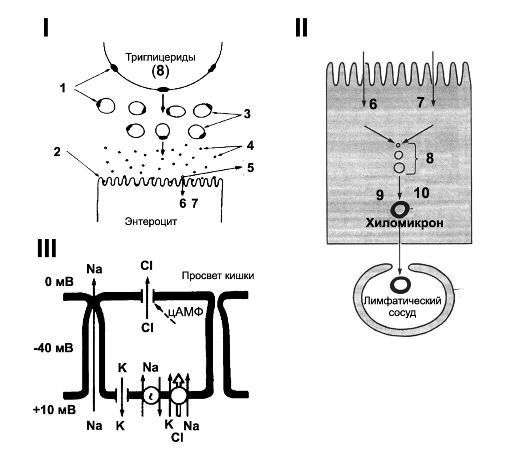 Рис. 22-6. ВСАСЫВАНИЕ В ТОНКОМ КИШЕЧНИКЕ. I - Эмульгация, расщепление и поступление жиров в энтероцит. II - Поступление и выход жиров из энтероцита. 1
- липаза; 2 - микроворсинки; 3 - эмульсия; 4 - мицеллы; 5 - соли
жёлчных кислот; 6 - моноглицериды; 7 - свободные жирные кислоты; 8 -
триглицериды; 9 - белок; 10 - фосфолипиды; 11 - хиломикрон. III - Механизм секреции HCO3- эпителиальными клетками слизистой оболочки желудка и двенадцатиперстной кишки. А - выход HCO3- в обмен на Cl- стимулируют некоторые гормоны (например, глюкагон), и подавляет блокатор транспорта Cl- фуросемид. Б - активный транспорт HCO3-, не зависящий от транспорта Cl-. В и Г - транспорт HCO3-
через мембрану базальной части клетки внутрь клетки и по межклеточным
пространствам (зависит от гидростатического давления в подэпителиальной
соединительной ткани слизистой оболочки). [11]
Рис. 22-6. ВСАСЫВАНИЕ В ТОНКОМ КИШЕЧНИКЕ. I - Эмульгация, расщепление и поступление жиров в энтероцит. II - Поступление и выход жиров из энтероцита. 1
- липаза; 2 - микроворсинки; 3 - эмульсия; 4 - мицеллы; 5 - соли
жёлчных кислот; 6 - моноглицериды; 7 - свободные жирные кислоты; 8 -
триглицериды; 9 - белок; 10 - фосфолипиды; 11 - хиломикрон. III - Механизм секреции HCO3- эпителиальными клетками слизистой оболочки желудка и двенадцатиперстной кишки. А - выход HCO3- в обмен на Cl- стимулируют некоторые гормоны (например, глюкагон), и подавляет блокатор транспорта Cl- фуросемид. Б - активный транспорт HCO3-, не зависящий от транспорта Cl-. В и Г - транспорт HCO3-
через мембрану базальной части клетки внутрь клетки и по межклеточным
пространствам (зависит от гидростатического давления в подэпителиальной
соединительной ткани слизистой оболочки). [11]
ется через слизистую оболочку путём диффузии и активного транспорта.
о Кальций. От 30 до 80% поглощенного кальция всасывается в тонкой кишке путём активного транспорта и диффузии. Активный транспорт Ca2+ усиливает 1,25-дигидроксикальциферол. Белки активируют абсорбцию Ca2+, фосфаты и оксалаты тормозят её.
о Другие ионы. Ионы железа, магния, фосфаты активно всасываются из тонкой кишки. С пищей железо поступает в виде Fe3+, в желудке железо переходит в растворимую форму Fe2+ и всасывается в краниальных отделах кишечника.
о Витамины. Водорастворимые витамины всасываются очень быстро; всасывание жирорастворимых витаминов A, D, E и K зависит от всасывания жиров. Если отсутствуют ферменты поджелудочной железы или жёлчь не поступает в кишечник, то всасывание этих витаминов нарушается. Большинство витаминов всасывается в краниальных отделах тонкой кишки, за исключением витамина B12 Этот витамин соединяется с внутренним фактором (белком, секретируемым в желудке), и образовавшийся комплекс всасывается в подвздошной кишке.
о Моносахариды. Всасывание глюкозы и фруктозы в щёточной каёмке энтероцитов тонкого кишечника обеспечивает белок-переносчик GLUT5. GLUT2 базолатеральной части энтероцитов реализует выход сахаров из клеток. 80% углеводов всасываются преимущественно в виде глюкозы - 80%; 20% приходится на фруктозу и галактозу. Транспорт глюкозы и галактозы зависит от количества Na+ в полости кишечника. Высокая концентрация Na+ на поверхности слизистой кишечника облегчает, а низкая - тормозит движение моносахаридов внутрь эпителиальных клеток. Это объясняется тем, что глюкоза и Na+ имеют общий переносчик. Na+ движется внутрь кишечных клеток по градиенту концентрации (вместе с ним перемещается глюкоза) и высвобождается в клетке. Далее Na+ активно перемещается в межклеточные пространства, а глюкоза за счёт вторичного активного транспорта (энергия этого транспорта обеспечивается косвенно за счёт активного транспорта Na+) поступает в кровь.
о Аминокислоты. Всасывание аминокислот в кишечнике реализуется при помощи переносчиков, кодируемых генами SLC. Нейтральные аминокислоты - фенилаланин и метионин - всасываются посредством вторичного активного транспорта за счёт энергии активного транспорта натрия Na+-независимые переносчики осуществляют перенос части нейтральных и щелочных аминокислот. Специальные переносчики транспортируют дипептиды и трипеп-
• тиды в энтероциты, где они расщепляются до аминокислот и затем путём простой и облегчённой диффузии поступают в межклеточную жидкость. Приблизительно 50% переваренных белков поступают из пищи, 25% - из пищеварительных соков и 25% - из отторгаемых клеток слизистой оболочки. Жиры (рис. 22-6,II). Моноглицериды, холестерол и жирные кислоты, доставленные мицеллами к энтероцитам, всасываются в зависимости от их размера. Жирные кислоты, содержащие менее 10-12 углеродных атомов, проходят сквозь энтероциты непосредственно в воротную вену и оттуда поступают в печень в виде свободных жирных кислот. Жирные кислоты, содержащие более 10-12 углеродных атомов, в энтероцитах превращаются в триглицериды. Некоторое количество всосавшегося холестерола превращается в эфиры холестерола. Триглицериды и эфиры холестерола покрываются пластом из белков, холестерола и фосфолипида, образуя хиломикроны, которые покидают энтероцит и входят в лимфатические сосуды. Всасывание в толстой кишке. Каждый день через илеоцекальную заслонку проходит около 1500 мл химуса, но ежедневно толстая кишка всасывает от 5 до 8 л жидкости и электролитов. Большая часть воды и электролитов всасывается в толстой кишке, оставляя не более 100 мл жидкости и немного Na+ и Cl- в составе кала. Всасывание происходит преимущественно в проксимальной части толстой кишки, дистальный отдел служит для накопления отходов и формирования кала. Слизистая оболочка толстой кишки активно всасывает Na+ и вместе с ним Cl-. Всасывание Na+ и Cl- создаёт осмотический градиент, который вызывает движение воды через слизистую оболочку кишечника. Слизистая оболочка толстой кишки секретирует бикарбонаты в обмен на эквивалентное количество абсорбируемого Cl-. Бикарбонаты нейтрализуют кислотные конечные продукты деятельности бактерий толстой кишки.
Формирование кала. В состав кала входит 3/4 воды и 1/4 плотного вещества. В плотном веществе содержится 30% бактерий, от 10 до 20% жира, 10-20% неорганических веществ, 2-3% белка и 30% непереваренных остатков пищи, пищеварительных ферментов, слущенного эпителия. Бактерии толстой кишки участвуют в переваривании небольшого количества целлюлозы, образуют витамины K, B12 ,тиамин, рибофлавин и различные газы (углекислый, водород и метан). Коричневый цвет кала определяют производные билирубина - стеркобилин и уробилин. Запах создаётся деятельностью бактерий и зависит от бактериальной флоры каждого индивидуума и состава принимаемой пищи. Вещества, придающие калу характерный запах, - индол, скатол, меркаптаны и сероводород.