Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА 18. ЭНДОКРИННАЯ СИСТЕМА
Для эндокринной системы ключевым является понятие «гормон». Гормоны - межклеточные гуморальные химические регуляторы - секретируются во внутреннюю среду организма (преимущественно в кровь) из специализированных (эндокринных) клеток и действуют на клетки-мишени, содержащие молекулы рецепторов к конкретным гормонам. Такое дистантное (через кровоток) взаимодействие между клетками-продуцентами гормонов и клетками-мишенями известно как эндокринная регуляция. Паракринная регуляция подразумевает эффекты гормонов, влияющих путём диффузии на соседние клетки-мишени, а аутокринная - непосредственно на секретирующие эти гормоны клетки (см. рис. 4-7).
Информационные межклеточные взаимодействия, реализуемые гормонами, укладываются в следующую последовательность событий: «сигнал (гормон) - рецептор - (второй посредник) - физиологический ответ». Физиологические концентрации гормонов, осуществляющих гуморальную регуляцию функций, колеблются в пределах 10-7-10-12 М, т.е. гормоны эффективны в крайне низких концентрациях.
Разнообразные гормоны и гормональные системы регулируют практически все функции организма, включая метаболизм, репродукцию, рост и развитие, водный и электролитный баланс и поведение. Регуляцию активности многих эндокринных желёз осуществляют при помощи механизмов обратной связи гипофиз и гипоталамус. Синтез некоторых гормонов (адреналин, норадреналин и др.) непосредственно не зависит от регулирующего влияния гипофиза и контролируется симпатической нервной системой.
Химия гормонов
По химической структуре гормоны, а также другие биологически активные вещества регуляторного характера (например, факторы роста, интерлейкины, интерфероны, хемокины, ангиотензины, Пг и ряд других) подразделяют на пептидные, стероидные, производные аминокислот и арахидоновой кислоты.
• Пептидные гормоны относятся к полярным веществам, которые не могут непосредственно проникать через биологические мембраны. Поэтому для их секреции используется механизм экзоцитоза. По этой же причине рецепторы пептидных гормонов встроены в плазматическую мембрану клетки-мишени, а передачу сигнала к внутриклеточным структурам осуществляют вторые посредники.
• Стероидные гормоны: минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол. Эти вещества - производные холестерола - относятся к неполярным, поэтому они свободно проникают через биологические мембраны. По этой причине секреция стероидных гормонов происходит без участия секреторных пузырьков. По этой же причине рецепторы неполярных молекул расположены внутри клетки-мишени. Такие рецепторы в общем виде называют ядерными.
• Производные аминокислот - тирозина (йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и дофамин), гистидина (гистамин), триптофана (мелатонин и серотонин)
• Производные арахидоновой кислоты (эйкозаноиды, или простаноиды). Эйкозаноиды (от греч. eikosi - двадцать) состоят (как и арахидоновая кислота) из 20 атомов углерода. К ним относятся простагландины (Пг), тромбоксаны, простациклины, лейкотриены, гидроксиэйкозотетраеноевая (HETE, от англ. hydroxyeicosatetraenoic) и эпоксиэйкозотриеноевая кислоты, а также производные этих кислот. Все эйкозаноиды обладают высокой и разносторонней физиологической активностью, многие из них функционируют только внутри клетки.
Механизмы действия гормонов на клетки-мишени
Информационные межклеточные взаимодействия, реализуемые в эндокринной системе, предусматривают такую последовательность событий:
гормон - рецептор клетки-мишени - (второй посредник) - ответ
клетки-мишени
Каждый гормон оказывает на клетку-мишень регулирующий эффект тогда и только тогда, когда он в качестве лиганда связывается со специфичным для него белком-рецептором в составе клеткимишени.
Циркуляция в крови. Гормоны циркулируют в крови либо свободно, либо в комплексе со связывающими их белками (T4, Т3, стероидные гормоны, инсулиноподобные факторы роста, гормон
роста). Связывание с такими белками существенно увеличивает время полужизни гормонов.
Рецепторы и вторые посредники рассмотрены в главе 4 «Межклеточные взаимодействия».
гормоны и их физиологические
эффекты
В этом разделе дана физиологическая характеристика различных гормонов, синтезируемых и секретируемых клетками эндокринной системы.
ГИПОТАЛАМО-ГИПОФИЗАРНАЯ СИСТЕМА
Часть промежуточного мозга - гипоталамус - и отходящий от его основания гипофиз анатомически и функционально составляют единое целое - гипоталамо-гипофизарную эндокринную систему (см. рис. 16-3,В,Г).
Гипоталамус
В нейросекреторных нейронах гипоталамуса синтезируются нейропептиды, поступающие как в переднюю (рилизинг-гормоны), так и в заднюю (окситоцин и вазопрессин) доли гипофиза.
Рилизинг-гормоны
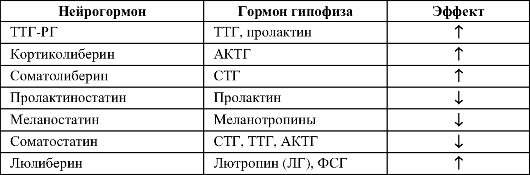
Гипоталамические рилизинг-гормоны (от англ. releasing hormone) - группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза. С функциональной точки зрения рилизинг-гормоны подразделяют на либерины (рилизинг-гормоны, способствующие усилению синтеза и секреции соответствующего гормона в эндокринных клетках передней доли гипофиза) и статины (рилизинг-гормоны, подавляющие синтез и секрецию гормонов в клетках-мишенях). К гипоталамическим либеринам относятся соматолиберин, гонадолиберин, тиреолиберин и кортиколиберин, а статины представлены соматостатином и пролактиностатином.
• Соматостатин - мощный регулятор функций эндокринной и нервной систем, ингибирует синтез и секрецию множества гормонов и секретов.
• Соматолиберин. Гипоталамический соматолиберин стимулирует секрецию гормона роста в передней доле гипофиза.
• Гонадолиберин (люлиберин) и пролактиностатин. Ген LHRH кодирует аминокислотные последовательности для гонадолибери-
на и пролактиностатина. Гонадолиберин - ключевой нейрорегулятор репродуктивной функции, стимулирует синтез и секрецию ФСГ и ЛГ в продуцирующих гонадотрофы клетках, а пролактиностатин подавляет секрецию пролактина из лактотрофных клеток передней доли гипофиза. Люлиберин - декапептид.
• Тиреолиберин - трипептид, синтезируется многими нейронами ЦНС (в том числе нейросекреторными нейронами паравентрикулярного ядра). Тиреолиберин стимулирует секрецию пролактина из лактотрофов и тиреотропина из тиреотрофов передней доли гипофиза.
• Кортиколиберин синтезируется в нейросекреторных нейронах паравентрикулярного ядра гипоталамуса, плаценте, Т-лимфоцитах. В передней доле гипофиза кортиколиберин стимулирует синтез и секрецию АКТГ и других продуктов экспрессии гена проопиомеланокортина.
• Меланостатин подавляет образование меланотропинов. Либерины и статины по аксонам гипоталамических нейронов
достигают срединного возвышения, где секретируются в кровеносные сосуды портальной системы кровотока, далее по воротным венам гипофиза эти нейрогормоны поступают в переднюю долю гипофиза и регулируют активность её эндокринных клеток (табл. 18-1, рис. 16-3,В,Г).
Таблица 18-1. Эффекты гипоталамических нейрогормонов на секрецию гормонов аденогипофиза [11]
 Роль дофамина. Промежуточный
продукт метаболизма тирозина и предшественник норадреналина и
адреналина - катехоловый амин дофамин (3-гидрокситирамин), поступающий к
клеткам передней доли гипофиза через кровь - ингибирует секрецию ФСГ,
лютропина (ЛГ), ТТГ и пролактина.
Роль дофамина. Промежуточный
продукт метаболизма тирозина и предшественник норадреналина и
адреналина - катехоловый амин дофамин (3-гидрокситирамин), поступающий к
клеткам передней доли гипофиза через кровь - ингибирует секрецию ФСГ,
лютропина (ЛГ), ТТГ и пролактина.
Гормоны задней доли гипофиза
Нанопептиды вазопрессин и окситоцин синтезируются в перикарионах нейросекреторных нейронов паравентрикулярного и супраоптического ядер гипоталамуса, транспортируются по их аксонам в составе гипоталамо-гипофизарного пути в заднюю долю гипофиза, где и секретируются в кровь (см. рис. 16-3Г). Сигналом к секреции является импульсная активность этих же самых нейросекреторных нейронов.
• Вазопрессин (аргинин вазопрессин, антидиуретический гормон - АДГ) оказывает антидиуретический (регулятор реабсорбции воды в канальцах почки) и сосудосуживающий (вазоконстриктор) эффекты (эти эффекты гормона приводят к повышению системного АД). Главная функция АДГ - регуляция обмена воды (поддержание постоянного осмотического давления жидких сред организма), что происходит в тесной связи с обменом натрия.
-ФСекрецию АДГ стимулируют: гиповолемия через барорецепторы каротидной области, гиперосмоляльность через осморецепторы гипоталамуса, переход в вертикальное положение, стресс, состояние тревоги.
-ФСекрецию АДГ подавляют: алкоголь, α-адренергические агонисты, глюкокортикоиды.
• Окситоцин стимулирует сокращение ГМК миометрия в родах, при оргазме, в менструальную фазу, секретируется при раздражении соска и околососкового поля и стимулирует сокращение миоэпителиальных клеток альвеол лактирующей молочной железы (рефлекс молокоотделения).
Передняя доля гипофиза
В передней доле синтезируются и секретируются так называемые тропные гормоны и пролактин. Тропными называют гормоны, мишенями которых являются другие эндокринные клетки. По химической структуре гормоны аденогипофиза являются либо пептидными гормонами, либо гликопротеинами.
• Гликопротеины: тиреотропный гормон и гонадотропины (лютеинизирующий гормон - ЛГ и фолликулостимулирующий гормон - ФСГ).
• Полипептидные гормоны: гормон роста, адренокортикотропный гормон (АКТГ) и пролактин. При экспрессии гена проопиомеланокортина - помимо АКТГ - происходит синтез и секреция ряда других пептидов (β- и γ-липотропины, меланокортины [α-, β- и γ-меланотропины], β-эндорфин, АКТГ-подобный пептид),
из которых гормональная функция установлена для меланотропинов; функции остальных пептидов изучены недостаточно.
Гормоны роста
Гормон роста (соматотрофный гормон - СТГ, соматотропин) нормально синтезируется только в ацидофильных клетках (соматотрофы) передней доли гипофиза. Другой гормон роста - хорионический соматомаммотрофин (плаацентарный лактоген) синтезируется в клетках синцитиотрофобласта. Эффекты гормонов роста опосредуют инсулиноподобные факторы роста - соматомедины. Гормоны роста - анаболики, они стимулируют рост всех тканей.
• Регуляторы экспрессии (см. таблицу 18-2).
• Суточная периодичность секреции. СТГ поступает в кровь циклически - «взрывами секреции», чередующимися с периодами прекращения секреции (продолжительность такого цикла измеряется минутами). Пик секреции СТГ приходится на третью и четвёртую фазы сна.
• Возрастные изменения секреции СТГ. Содержание СТГ в плазме крови наибольшее в раннем детском возрасте, постепенно уменьшается с возрастом и составляет: от 5 до 20 лет - 6 нг/мл (с пиком на стадии пубертата), от 20 до 40 лет - 3 нг/мл, после 40 лет - 1 нг/мл.
• Функции
-ФСТГ - анаболический гормон, стимулирующий рост всех клеток за счёт увеличения поступления в клетки аминокислот и усиления синтеза белка. Наиболее очевидны долговременные эффекты СТГ на рост костей. При этом мишенями СТГ явля-
Таблица 18-2. Стимулирующие и подавляющие влияния на секрецию гормона роста
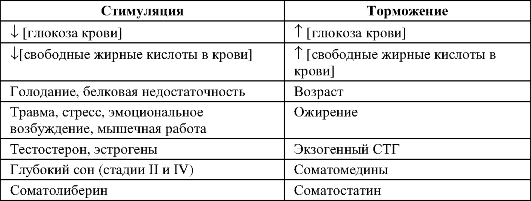 ются клетки эпифизарной хрящевой пластинки длинных трубчатых костей и остеобласты периоста и эндоста. -Φ- Метаболические эффекты СТГ двуфазны, направлены на поддержание уровня глюкозы в крови и обеспечение энергетических затрат организма.
ются клетки эпифизарной хрящевой пластинки длинных трубчатых костей и остеобласты периоста и эндоста. -Φ- Метаболические эффекты СТГ двуфазны, направлены на поддержание уровня глюкозы в крови и обеспечение энергетических затрат организма.
♦ Начальная фаза (инсулиноподобный эффект). СТГ увеличивает поглощение глюкозы мышцами и жировой тканью, а также поглощение аминокислот и синтез белка мышцами и печенью. Одновременно СТГ угнетает липолиз в жировой ткани. Через несколько минут развивается отсроченная фаза эффектов СТГ.
♦ Отсроченная фаза (антиинсулиноподобный, или диабетогенный эффект). Через несколько десятков минут происходят угнетение поглощения и утилизации глюкозы (содержание глюкозы в крови увеличивается) и усиление липолиза (содержание свободных жирных кислот в крови увеличивается).
♦ Обмен белка. СТГ стимулирует поступление аминокислот и синтез белка в клетках (анаболический эффект).
♦ Жировой обмен. СТГ усиливает липолиз, освобождающиеся при этом жирные кислоты используются для пополнения энергетических затрат клеток.
♦ В итоге под влиянием СТГ происходит перераспределение в оче- рёдности использования веществ для получения энергии: используются жиры, а не углеводы или белки. Поскольку СТГ имеет анаболический эффект, это приводит к увеличению массы тела без накопления жиров.
• Циркуляция в крови. Время полужизни СТГ в крови около 25 минут. Около 40% выделившегося СТГ образует комплекс с СТГ- связывающим белком, при этом продолжительность полужизни СТГ значительно возрастает.
• Рецептор СТГ относится (вместе с рецептором пролактина, ряда интерлейкинов и эритропоэтина) к семейству цитокиновых рецепторов (связанные с тирозинкиназой рецепторы). СТГ связывается также с рецептором пролактина.
• Соматомедины C и A (полипептиды из 70 и 67 аминокислотных остатков соответственно) опосредуют эффекты СТГ, выступая в качестве аутокринных факторов роста. Оба соматомедина имеют выраженную структурную гомологию с проинсулином, почему их также называют инсулиноподобными факторами роста. Рецепторы соматомединов, как и рецептор инсулина, относятся к рецепторным тирозинкиназам. Соматомедин C, связываясь с его рецепторами, стимулирует синтез гипофизарного СТГ и гипота-
ламического соматостатина и подавляет синтез гипоталамического соматолиберина.
Адренокортикотропный гормон
Адренокортикотропный гормон (АКТГ, кортикотропин). Структуру АКТГ кодирует ген проопиомеланокортина.
• Суточный ритм. Секреция АКТГ начинает расти после засыпания и достигает пика при пробуждении.
• Функции. АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (преимущественно глюкокортикоида кортизола).
• Рецепторы АКТГ (АКТГ связывается с рецептором меланокортина типа 2) относятся к мембранным, связанным с G-белком (активирует аденилатциклазу, что при помощи цАМФ в конечном итоге активирует многочисленные ферменты синтеза глюкокортикоидов).
Меланокортины (меланотропины) контролируют пигментацию кожи и слизистых оболочек. Экспрессия АКТГ и меланокортинов в значительной степени сочетаются. Меланостатин подавляет секрецию меланотропинов (вероятно, и АКТГ). Известно несколько типов рецепторов меланокортинов, через тип 2 этих рецепторов происходит также реализация эффектов АКТГ.
Гонадотропные гормоны
К этой группе относят гипофизарные фоллитропин (фолликулостимулирующий гормон - ФСГ) и лютропин (ЛГ, лютеинизирующий гормон), а также хорионический гонадотропин (ХГТ) плаценты.
• Фолликулостимулирующий гормон (ФСГ, фоллитропин) у женщин вызывает рост фолликулов яичника, у мужчин регулирует сперматогенез (мишени ФСГ - клетки Сертоли).
• Лютеинизирующий гормон (ЛГ, лютропин) стимулирует синтез тестостерона в клетках Ляйдига яичек (у мужчин ЛГ иногда называют «стимулирующий интерстициальные клетки гормон»), синтез эстрогенов и прогестерона в яичниках, стимулирует овуляцию и образование жёлтого тела в яичниках.
• Хорионический гонадотропин (ХГТ) синтезируется клетками трофобласта с 10-12 дней развития. При беременности ХГТ взаимодействует с клетками жёлтого тела и стимулирует синтез и секрецию прогестерона.
Тиреотропный гормон. Гликопротеиновой природы тиреотропный гормон (ТТГ, тиреотропин) стимулирует синтез и секрецию йодсодержащих гормонов щитовидной железы (T3 и T4). Тиреотропин стимулирует дифференцировку эпителиальных клеток щито-
видной железы (кроме так называемых светлых клеток, синтезирующих тиреокальцитонин) и их функциональное состояние (включая синтез тиреоглобулина и секрецию Т3 и Т4).
Пролактин ускоряет развитие молочной железы и стимулирует секрецию молока. Синтез пролактина происходит в ацидофильных аденоцитах (лактотрофы) передней доли гипофиза. Количество лактотрофов составляет не менее трети всех эндокринных клеток аденогипофиза. При беременности объём передней доли удваивается за счёт увеличения числа лактотрофов (гиперплазия) и увеличения их размеров (гипертрофия). Главная функция пролактина - стимуляция функции молочной железы.
ШИШКОВИДНОЕ ТЕЛО
Шишковидное тело (corpus рineale) - небольшой (5-8 мм) вырост промежуточного мозга, соединённый ножкой со стенкой третьего желудочка (рис. 18-1). Из паренхиматозных клеток этой железы - пинеалоцитов - в ликвор и в кровь секретируется производное триптофана - мелатонин. Орган снабжён многочисленными постганглионарными нервными волокнами от верхнего шейного симпатического узла. Железа принимает участие в реализации циркадианных (околосуточных) ритмов.
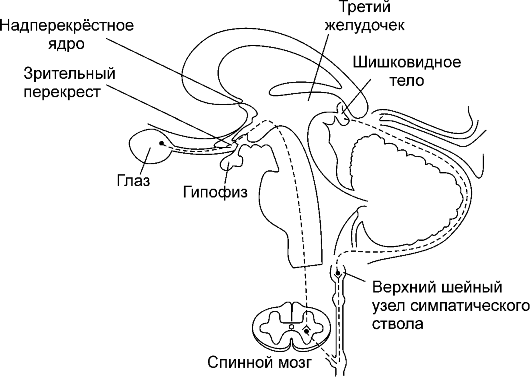 Рис. 18-1. Топография и иннервация шишковидной железы
Рис. 18-1. Топография и иннервация шишковидной железы
• Околосуточный ритм. Циркадианный ритм - один из биологических ритмов (суточная, помесячная, сезонная и годовая ритмика), скоординированный с суточной цикличностью вращения Земли; несколько не соответствует 24 часам. Многие физиологические процессы, в том числе гипоталамическая нейросекреция, подчиняются околосуточному ритму.
• Мелатонин (N-ацетил-5-метокситриптамин) секретируется в ликвор и в кровь преимущественно в ночные часы. Содержание мелатонина в плазме в ночное время составляет у детей в возрасте от 1 до 3 лет 250 пг/мл, у подростков - 120 пг/мл и у лиц в возрасте 50-70 лет - 20 пг/мл. В то же время в течение дня содержание мелатонина составляет всего около 7 пг/мл у лиц любого возраста.
-Ф- Регуляция экспрессии мелатонина происходит при взаимодействии норадреналина с α- и β-адренорецепторами пинеалоцитов: связанный с рецепторами G-белок (активация аденилатциклазы) в конечном итоге приводит к увеличению транскрипции гена арилалкиламин-N-ацетилтрансферазы - ключевого фермента синтеза мелатонина. Полная же цепочка событий - от сетчатки до пинеалоцитов - такова (рис. 18-1).
♦ Изменения освещённости сетчатки через зрительный тракт и дополнительные пути оказывают влияние на разряды нейронов надперекрёстного ядра (ростро-вентральная часть гипоталамуса).
■ Сигналы от сетчатки к гипоталамусу возникают не в палочках и колбочках, а в других клетках (возможно, амакринных) сетчатки, содержащих фотопигменты группы криптохромов.
■ Супраоптическое ядро содержит так называемые эндогенные часы - неизвестной природы генератор биологических ритмов (включая околосуточный), контролирующий продолжительность сна и бодрствования, пищевое поведение, секрецию гормонов и т.д. Сигнал генератора - гуморальный фактор, секретируемый из супраоптического ядра (в том числе. в ликвор).
♦ Сигналы от супраоптического ядра через нейроны паравентрикулярного ядра (n. paraventricularis) активируют преганглионарные симпатические нейроны боковых столбов спинного мозга (columna lateralis).
♦ Симпатические преганглионарные нервные волокна активируют нейроны верхнего шейного узла симпатического ствола.
♦ Постганглионарные симпатические волокна от верхнего шейного узла секретируют норадреналин, взаимодействующий с адренорецепторами плазмолеммы пинеалоцитов. -Φ- Эффекты мелатонина изучены плохо, но известно, что мелатонин в гипоталамусе и гипофизе инициирует транскрипцию гена Period-1 (один из генов, имеющих отношение к так называемым эндогенным часам).
• Рецепторы мелатонина - трансмембранные гликопротеины, связанные с G-белком (активация аденилатциклазы) - обнаружены в гипофизе, надперекрестном ядре (n. suprachiasmaticus) гипоталамуса, сетчатке, некоторых областях ЦНС и в ряде других органов.
ЩИТОВИДНАЯ ЖЕЛЕЗА
В клетках щитовидной железы происходит синтез двух химически и функционально разных классов гормонов - йодсодержащих (синтезируются в эпителиальных фолликулах железы) и продуктов экспрессии кальцитониновых генов (синтезируются в так называемых светлых клетках - C-клетки - фолликулов).
• Йодсодержащие гормоны железы - производные тирозина. Тироксин (T4) и трийодтиронин (Т3) увеличивают обменные процессы, ускоряют катаболизм белков, жиров и углеводов, необходимы для нормального развития ЦНС, увеличивают ЧСС и сердечный выброс.
• Кальцитонин (32-аминокислотный пептид) и катакальцин (21-ами- нокислотный пептид). Их функции антагонистичны эффектам ПТГ - гормона паращитовидной железы: кальцитонин уменьшает [Са2+] в крови, стимулирует минерализацию кости, усиливает почечную экскрецию Са2+, фосфатов и Na+ (уменьшается их реабсорбция в канальцах почки).
• Относящиеся к кальцитониновому гену пептиды α и β (37 аминокислот) экспрессируются в ряде нейронов ЦНС и на периферии (особенно в связи с кровеносными сосудами). Их функции - участие в ноцицепции, пищевом поведении, а также в регуляции тонуса сосудов. Рецепторы к этим пептидам найдены в ЦНС, сердце, плаценте.
Йодсодержащие гормоны
Синтез и секреция йодсодержащих гормонов происходит в эпителиальных фолликулах щитовидной железы. Эти фолликулы имеют различную величину и форму (преимущественно округлую), состоят из стенки (образована одним слоем фолликулярных кле-
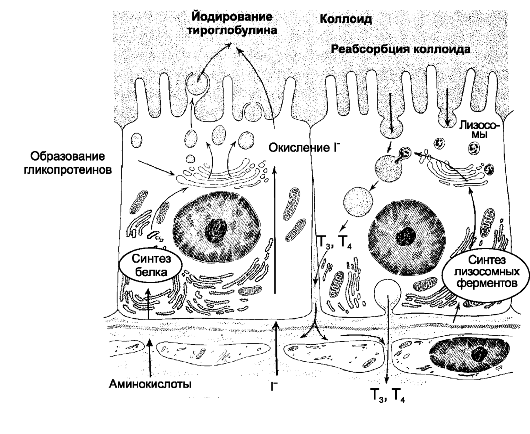
Рис. 18-2. Этапы синтеза и секреции йодсодержащих гормонов [11]. В левой части рисунка направление процессов снизу вверх (из просвета кровеносных капилляров в фолликулярные клетки и далее в коллоид), в правой части рисунка направление процессов сверху вниз (из коллоида в фолликулярные клетки и далее в просвет капилляров)
ток) и полости фолликула, содержащей так называемый коллоид. Функцию фолликулярных клеток стимулирует тиреотропин. Фолликулярные клетки могут иметь различную высоту (от низкокубической до цилиндрической), что зависит от интенсивности их функционирования: высота клеток пропорциональна напряжённости осуществляемых в них процессов. Полный цикл синтеза и секреции йодсодержащих гормонов происходит между фолликулярными клетками и коллоидом (рис. 18-2).
Синтез йодсодержащих гормонов
Синтез и секреция T4 и T3 - многоэтапный процесс, находящийся под активирующим влиянием ТТГ.
• Поглощение йода. Йод в виде органических и неорганических соединений поступает в ЖКТ с пищей и питьевой водой. Транспорт йода из кровеносных капилляров в железу происходит за счёт встроённых в плазматическую мембрану базальной части фолликулярных клеток молекул трансмембранного переносчика ионов натрия и йода (так называемая «йодная ловушка»). Из верхушечной части фолликулярных клеток I- поступает в коллоид при помощи анионного транспортёра (пендрин).
❖ Суточная потребность организма в йоде - 150-200 мкг. Дефицит йода развивается при недостаточном поступлении йода с пищей и водой. Уменьшение синтеза тареоидных гормонов возникает при уменьшении потребления йода ниже 10 мкг/сут.
❖ Соотношение концентраций I- в железе к концентрации I- в сыворотке крови в норме составляет 25:1.
• Окисление йода (I- - I+) происходит при помощи йодидпероксидазы (тиреопероксидаза) сразу после поступления в коллоид. Этот же фермент катализирует присоединение окисленного йода к остаткам тирозина в составе молекул тиреоглобулина.
• Тиреоглобулин. Этот содержащий 115 остатков тирозина гликопротеин синтезируется в фолликулярных клетках и секретируется в коллоид. Это так называемый незрелый тиреоглобулин.
• Йодирование тиреоглобулина
❖ Созревание тиреоглобулина происходит примерно в течение 2 сут на верхушечной поверхности фолликулярных клеток путём его йодирования при помощи тиреопероксидазы.
❖ Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием монойодтирозинов и дийодтирозинов. Гормональной активностью моно- и дийодтирозины не обладают; оба соединения выделяются из фолликулярных клеток, но быстро захватываются обратно и дейодинируются. Две молекулы дийодтирозина конденсируются с образованием йодтиронина (Т4), а монойодтирозин и дийодтирозин - йодтиронина (Т3).
❖ Зрелый тиреоглобулин (полностью йодированный) - прогормон йодсодержащих гормонов, форма их хранения в коллоиде.
• Эндоцитоз и расщепление тиреоглобулина
❖ По мере необходимости зрелый тиреоглобулин поступает (интернализуется) из коллоида в фолликулярные клетки путём опосредованного рецепторами N-ацетилглюкозамина эндоцитоза.
❖ Содержащие тиреоглобулин эндоцитозные пузырьки сливаются с лизосомами, где происходит расщепление тиреоглобулина и освобождение Т3 и Т4.
• Секреция T3 и T4
❖ Образовавшиеся при расщеплении тиреоглобулина аминокислоты используются для новых синтезов, а Т3 и Т4 из базальной части фолликулярных клеток поступают в кровь.
❖ В норме щитовидная железа секретирует 80-100 мкг Т4 и 5 мкг Т3 в сутки. Ещё 22-25 мкг Т3 образуется в результате дейодирования Т4 в периферических тканях, преимущественно в печени.
Регуляция синтеза йодтиронинов
Синтез и секрецию йодтиронинов регулирует гипоталамо-гипофизарная система по механизму обратной связи (рис. 18-3).
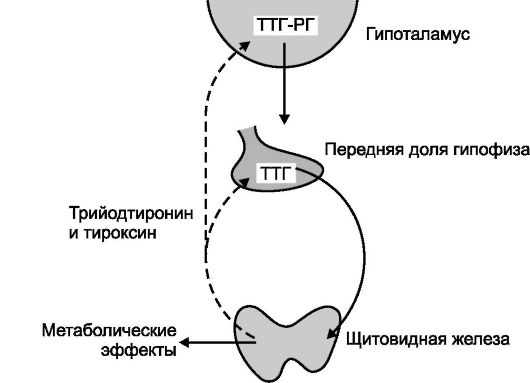 Рис. 18-3. Регуляторные взаимоотношения между гипоталамусом, аденогипофизом и щитовидной железой. Активирующие
влияния - сплошная линия, тормозящие влияния - пунктирная линия. ТТГ-РГ
- тиреотропин рилизинг гормон. Стимул для повышения секреции ТТГ-РГ и
ТТГ - снижение концентрации йодтиронинов в крови
Рис. 18-3. Регуляторные взаимоотношения между гипоталамусом, аденогипофизом и щитовидной железой. Активирующие
влияния - сплошная линия, тормозящие влияния - пунктирная линия. ТТГ-РГ
- тиреотропин рилизинг гормон. Стимул для повышения секреции ТТГ-РГ и
ТТГ - снижение концентрации йодтиронинов в крови
Тироксин
Тироксин {р-[(3,5-дийодо-4-гидроксифенокси)-3,5-дийодофенил]ала- нин, или 3,5,3',5'-тетрайодтиронин, C15H11I4NO4, Т4, молек. масса 776,87} образуется из пары дийодтирозинов. Тироксин - основной йодсодержащий гормон, на долю T4 приходится не менее 90% всего содержащегося в крови йода.
• Транспорт в крови. Не более 0,05% Т4 циркулирует в крови в свободной форме, практически весь тироксин находится в связанной с белками плазмы форме. Главный транспортный белок - тироксин-связывающий глобулин (связывает 80% Т4); на долю тироксин-связывающего преальбумина, а также альбумина приходится 20% Т4.
• Время циркуляции в крови (время полужизни) T4 около 7 дней, при гипертиреозе - 3-4 дня, при гипотиреозе - до 10 дней.
• L-форма тироксина физиологически примерно вдвое активнее рацемической (DL-тироксин), D-форма гормональной активности не имеет.
• Дейодирование наружного кольца тироксина, частично происходящее в щитовидной железе, осуществляется преимущественно в печени и приводит к образованию Т3.
• Реверсивный трийодтиронин. Дейодирование внутреннего кольца тироксина происходит в щитовидной железе, преимущественно в печени и частично в почке, в результате образуется реверсивный (обратный) Т3 (3,3',5'-трийодтиронин, rT3 [от англ. reverse]), имеющий после рождения незначительную физиологическую активность. Содержание rT3 в амниотической жидкости много выше rT3 крови матери. Это обстоятельство означает, что преобладающий йодсодержащий гормон плода - rT3.
Трийодтиронин
Трийодтиронин [3,5,3'-трийодтиронин, или 4-(3-йодо-4-гидрокси- фенокси)-3,5-дийодофенилаланин, C15H12I3NO4, молек. масса 650,98] образуется из монойодтиронина и дийодтиронина (в щитовидной железе синтезируется около 15% циркулирующего в крови T3, остальной трийодтиронин образуется при монодейодировании наружного кольца тироксина, происходящем преимущественно в печени). На долю T3 приходится лишь 5% содержащегося в крови йода, но Т3 существенно важен для организма и для реализации эффектов йодсодержащих гормонов.
• Транспорт в крови. Не более 0,5% Т3 циркулирует в крови в свободной форме, практически весь трийодтиронин находится в связанной форме.
• Время циркуляции в крови (время полужизни) T3 около 1,5 дней.
• Физиологическая активность T3 примерно в четыре раза выше, чем тироксина, но время полужизни много меньше. Биологическая активность и T3 и T4 обусловлена несвязанной фракцией.
• Катаболизм йодтиронинов. Т3 и Т4 конъюгируются в печени с глюкуроновой или серной кислотами и секретируются с желчью, вса-
сываются в кишечнике, дейодируются в почках и выделяются с мочой.
Рецепторы ТИРЕОИДНЫХ ГОРМОНОВ. Ядерные рецепторы тиреоидных гормонов - факторы транскрипции. Известно не менее трёх подтипов этих рецепторов: α1, α2 и β. α1 и β-подтипы - трансформирующие гены ERBA1 и ERBA2 соответственно.
Функции йодсодержащих гормонов
Функции йодсодержащих гормонов многочисленны. Т3 и Т4 увеличивают интенсивность обменных процессов, ускоряют катаболизм белков, жиров и углеводов, они необходимы для нормального развития ЦНС, увеличивают ЧСС и сердечный выброс. Крайне разнообразные эффекты йодсодержащих гормонов на клетки-мишени (ими практически являются все клетки организма) объясняют увеличением синтеза белков и потребления кислорода.
• Синтез белков увеличивается в результате активации транскрипции в клетках-мишенях, в том числе гена гормона роста. Йодтиронины расценивают как синергисты гормона роста. При дефиците Т3 клетки гипофиза теряют способность к синтезу СТГ.
• Потребление кислорода возрастает в результате увеличения активности Na+,К+-АТФазы.
• Печень. Йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т3 повышает чувствительность клеток к эффектам адреналина (стимуляция липолиза в жировой ткани и мобилизация гликогена в печени).
• Мышцы. Т3 увеличивает потребление глюкозы, стимулирует синтез белков и увеличение мышечной массы, повышает чувствительность к действию адреналина.
• Теплопродукция. Йодтиронины участвуют в формировании ответной реакции организма на охлаждение увеличением теплопродукции, повышая чувствительность симпатической нервной системы к норадреналину и стимулируя секрецию норадреналина.
• Гиперйодтиронинемия. Очень высокие концентрации йодтиронинов тормозят синтез белков и стимулируют катаболические процессы, что приводит к развитию отрицательного азотистого баланса.
• Оценка функций щитовидной железы
❖ Радиоиммунологический анализ позволяет прямо измерять содержание Т3, Т4, ТТГ.
❖ Поглощение гормонов смолами - непрямой метод определения связывающих гормоны белков.
❖ Индекс свободного тироксина - оценка свободного Т4.
❖ Тест стимуляции ТТГ тиреолиберином определяет секрецию в кровь тиреотропина в ответ на внутривенное введение тиреолиберина.
❖ Тесты выявления АТ к рецепторам ТТГ выявляют гетерогенную группу Ig, связывающихся с рецепторами ТТГ эндокринных клеток щитовидной железы и изменяющих её функциональную активность.
❖ Сканирование щитовидной железы при помощи изотопов технеция (99mТс) позволяет выявить области пониженного накопления радионуклида (холодные узлы), обнаружить эктопические очаги щитовидной железы или дефект паренхимы органа. 99mТс накапливается только в щитовидной железе, период полувыведения составляет всего 6 часов.
❖ Исследование поглощения радиоактивного йода при помощи йода-123 (123I) и йода-131 (131I).
❖ Содержание йода в питьевой воде. Проводится йодирование воды на водопроводных станциях.
❖ Пищевая соль. В России запрещено вьпускать нейодированную пищевую соль.
• Тиреоидный статус определяет эндокринную функцию щитовидной железы. Эутиреоидия - отсутствие отклонений. Заболевание щитовидной железы можно предположить при появлении симптомов недостаточности эндокринной функции (гипотиреоз), избыточных эффектов тиреоидных гормонов (гипертиреоз) либо при очаговом или диффузном увеличении щитовидной железы (зоб).
Кальцитонин и катакальцин
С-клетки (произносят «си-клетки», от англ. calcitonin - кальцитонин) в составе фолликулов называют также парафолликулярными. Ген CALC1 кодирует пептидные гормоны кальцитонин, катакальцин и относящийся к кальцитониновому гену пептид α. В щитовидной железе синтезируются регуляторы обмена Са2+ - кальцитонин и катакальцин, пептид α в нормальной щитовидной железе не экспрессируется.
• Кальцитонин - пептид, содержащий 32 аминокислотных остатка, мол. масса 3421.
❖ Регулятор экспрессии - [Са2+] плазмы крови, внутривенное введение кальция хлорида существенно увеличивает секрецию кальцитонина. β-Адренергические агонисты, дофамин, эстрогены, гастрин, холецистокинин, глюкагон и секретин также стимулируют секрецию кальцитонина.
❖ Функции кальцитонина многообразны. Кальцитонин - один из регуляторов кальциевого обмена, функции кальцитонина антагонистичны функциям гормона паращитовидной железы.
♦ Уменьшение содержания Са2+ в крови (паратиреокрин увеличивает содержание Са2+).
♦ Стимуляция минерализации кости (ПТГ усиливает резорбцию кости).
♦ Усиление почечной экскреции Са2+, фосфатов и Na+ (уменьшается их реабсорбция в канальцах почки).
♦ Желудочная и панкреатическая секреция. Кальцитонин уменьшает кислотность желудочного сока и содержание амилазы и трипсина в соке поджелудочной железы.
♦ Гормональная регуляция состояния костной ткани (см. ниже).
❖ Рецептор кальцитонина относится к семейству рецепторов секретина, при связывании кальцитонина с рецептором в клет- ках-мишенях (например, остеокластах) происходит увеличение содержания цАМФ.
• Катакальцин - пептид, состоящий из 21 аминокислотного остатка, - имеет те же функции, что и кальцитонин.
ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Четыре небольшие паращитовидные железы расположены на задней поверхности и под капсулой щитовидной железы.
• Поскольку паращитовидные железы топографически связаны с щитовидной железой, при хирургической резекции последней существует опасность удаления паращитовидных желёз. При этом развиваются гипокальциемия, тетания, судороги, возможна смерть.
• При запущенном раке лёгкого возможно поражение опухолевым процессом левого или обоих возвратных гортанных нервов, развивается паралич голосовой связки, приводящий к осиплости или полной потере голоса.
Функция околощитовидных желёз - синтез и секреция Са2+- регулирующего пептидного гормона паратиреокрина (ПТГ). ПТГ вместе с кальцитонином и катакальцином щитовидной железы, а также витамином D регулирует обмен кальция и фосфатов.
Гормоны
В паращитовидной железе синтезируются и секретируются в кровь как паратиреокрин (ПТГ), так и относящийся к ПТГ белок. Эти гормоны кодируют разные гены, но физиологическое значение относящегося к ПТГ белка значительно шире.
Паратиреокрин
Паратиреокрин (паратирин, паратгормон, гормон паращитовидной железы, паратиреоидный гормон, ПТГ) - полипептид из 84 аминокислотных остатков.
• Регуляторы экспрессии ПТГ
❖ [Са2+] сыворотки - главный регулятор секреции ПТГ. Ионы Са2+ взаимодействуют с рецепторами Са2+ (Са2+-сенсор) главных клеток паращитовидных желёз.
♦ Гипокальциемия (↑[Са2+] в крови) усиливает секрецию ПТГ.
♦ Гиперкальциемия (↑[Са2+] в крови) уменьшает секрецию ПТГ. ■ Са2+-сенсор - трансмембранный гликопротеин, обнаруженный в главных клетках околощитовидных желёз, а также в эпителии почечных канальцев. Связывание Са2+ с рецептором стимулирует фосфолипазу C, что приводит к освобождению ИТФ и диацилглицерола с последующим выбросом Са2+ из его внутриклеточных депо. Увеличение внутриклеточного [Са2+] активирует протеинкиназу C. Конечный результат - подавление секреции ПТГ.
❖ Витамин D - вспомогательный регулятор экспрессии гена ПТГ. Рецепторы витамина D (кальцитриола) относятся к ядерным факторам транскрипции. Связывание комплекса «кальцитри- ол-рецептор кальцитриола» с ДНК угнетает транскрипцию гена ПТГ.
❖ Ионы магния. Пониженное содержание Mg2+ стимулирует секрецию ПТГ, избыток Mg2+ оказывает на неё тормозящий эффект.
❖ Секреция ПТГ увеличивается под влиянием активации b-ад- ренергических рецепторов и цАМФ.
• Рецепторы ПТГ - трансмембранные гликопротеины, связанные с G-белком - в значительном количестве содержатся в костной ткани (остеобласты) и корковой части почек (эпителий извитых канальцев нефрона). Известно 2 типа рецепторов ПТГ: тип I связывает ПТГ и относящийся к ПТГ белок, тип II - только ПТГ. При связывании лигандов с рецептором в клетках-мишенях происходит не только увеличение внутриклеточного содержания цАМФ, но и активация фосфолипазы C (освобождение ИТФ и диацилглицерола, выброс Са2+ из его внутриклеточных депо, активация Са2+-зависимых протеинкиназ).
• Функции. ПТГ поддерживает гомеостаз кальция и фосфатов.
❖ ПТГ увеличивает содержание кальция в крови, усиливая резорбцию кости и вымывание кальция из костей, а также усиливая канальцевую реабсорбцию кальция в почках.
❖ ПТГ стимулирует образование кальцитриола в почках, кальцитриол же усиливает всасывание кальция и фосфатов в кишечнике.
❖ ПТГ уменьшает реабсорбцию фосфатов в канальцах почки и усиливает их вымывание из костей.
Обмен минералов и костная ткань
Кости формируют скелет организма, защищают и поддерживают жизненно важные органы, выполняют функцию депо кальция для нужд всего организма. В кости присутствуют две линии клеток - созидающая (остеогенные клетки - остеобласты - остеоциты) и разрушающая (многоядерные остеокласты). Клетки костной ткани окружает костный матрикс. Различают незрелый (неминерализованный) костный матрикс - остеоид и зрелый (обызвествлённый, или кальцифицированный) костный матрикс
Костный матрикс
Зрелый костный матрикс составляет 50% сухого веса кости и состоит из неорганической (50%) и органической (25%) частей и воды
(25%).
• Органическая часть. Органические вещества костного матрикса синтезируют остеобласты. К макромолекулам органического матрикса относятся коллагены (коллаген типа I - 90-95% и коллаген типа V) и неколлагеновые белки (остеонектин, остеокальцин, протеогликаны, сиалопротеины, морфогенетические белки, протеолипиды, фосфопротеины), а также гликозаминогликаны (хондроитинсульфат, кератансульфат).
• Неорганическая часть в значительном количестве содержит два химических элемента - кальций (35%) и фосфор (50%), образующие кристаллы гидроксиапатита - [Ca10(PO4)6(OH)2H2O]. В состав неорганической части кости также входят бикарбонаты, цитраты, фториды, соли Mg2+, К+, Na+.
❖ Кристаллы гидроксиапатита соединяются с молекулами коллагена через остеонектин. Такая связка делает кости исключительно устойчивыми к растяжению и сжатию.
❖ В организме взрослого человека содержится около 1000 г кальция. 99% всего кальция находится в костях. Около 99% кальция костей входит в состав кристаллов гидроксиапатита. Лишь 1% кальция костей находится в виде фосфатных солей, именно эти соли легко обмениваются между костью и кровью и играют роль буфера («обменный кальций») при изменениях концентрации кальция в плазме крови.
Минерализация остеоида. Остеоид - неминерализованный органический костный матрикс вокруг остеобластов, синтезирующих и секретирующих его компоненты. В дальнейшем остеоид минерализуется за счёт активности щелочной фосфатазы. Этот фермент осуществляет гидролиз эфиров фосфорной кислоты с образованием ортофосфата, который взаимодействует с Ca2+, что приводит к образованию осадка в виде аморфного фосфата кальция Ca3(PO4)2 с последующим формированием из него кристаллов гидроксиапатита. Для нормальной минерализации остеоида особенно необходим 1а,25-дигидроксихолекальциферол (активная форма витамина D3 - кальцитриол). Способствуя всасыванию кальция и фосфора в кишечнике, кальцитриол обеспечивает их необходимую концентрацию для запуска кристаллизационных процессов в костном матриксе. Прямо воздействуя на остеобласты, кальцитриол повышает активность щелочной фосфатазы в этих клетках, способствуя минерализации костного матрикса.
Клетки костной ткани
• Остеобласты активно синтезируют и секретируют вещества костного матрикса практически через всю поверхность клетки, что даёт возможность остеобласту окружить себя матриксом со всех сторон. По мере снижения синтетической и секреторной активности остеобласты становятся остеоцитами, замурованными в костный матрикс. И остеобласты, и остеоциты экспрессируют рецепторы ПТГ и кальцитриола.
• Остеоциты - зрелые неделящиеся клетки, расположенные в костных полостях, или лакунах. Тонкие отростки остеоцитов находятся в канальцах, отходящих в разные стороны от костных полостей (лакунарно-канальцевая система). Остеоциты поддерживают структурную целостность минерализованного матрикса, участвуют в регуляции обмена Ca2+ в организме. Эта функция остеоцитов находится под контролем со стороны Ca2+ плазмы крови и различных гормонов. Лакунарно-канальцевая система заполнена тканевой жидкостью, через которую осуществляется обмен веществ между остеоцитами и кровью. В канальцах постоянно циркулирует жидкость, что поддерживает диффузию метаболитов и обмен между лакунами и кровеносными сосудами надкостницы. По химическому составу лакунарно-канальцевая жидкость отличается от плазмы крови или жидкости в матриксе других тканей. Концентрация Ca2+ и PO43- в лакунарно-канальцевой жидкости превышает критический уровень для спонтанного осаждения солей Ca2+, что указывает на присутствие секретируемых костны-
ми клетками ингибиторов осаждения, контролирующих процесс минерализации.
• Остеокласты - крупные многоядерные клетки системы мононуклеарных фагоцитов. Предшественники остеокластов - моноциты. Для дифференцировки остеокластов необходимы колониестимулирующий фактор макрофагов (M-CSF) и кальцитриол, а для их активизации интерлейкин-6 и продуцируемый остеобластами фактор дифференцировки остеокластов (лиганд остеопротегерина). Остеокласты расположены в области резорбции (разрушения) кости (рис. 18-4,I).
Гофрированная каёмка остеокласта (рис. 18-4,II) - многочисленные цитоплазматические выросты, направленные к поверхности кости. Через мембрану выростов из остеокласта выделяется большое количество H+ и Cl-, что создаёт и поддерживает в замкнутом пространстве лакуны кислую среду (pH около 4), оптимальную для растворения солей кальция костного матрикса. Образование H+ в цитоплазме остеокласта катализирует карбоангидраза II. Остеокласты содержат многочисленные лизосомы, ферменты которых (кислые гидролазы, коллагеназы, катепсин К) разрушают органическую часть костного матрикса.
Гормональная регуляция
Регуляция роста. Синтез макромолекул костного матрикса стимулируют кальцитриол, ПТГ, соматомедины, трансформирующий фактор роста β, полипептидные факторы роста из кости.
• Соматомедины стимулируют анаболические процессы в скелетных тканях (синтез ДНК, РНК, белка, включая протеогликаны), а также сульфатирование гликозаминогликанов. Активность соматомединов определяет гормон роста (соматотропин).
• Витамин C необходим для образования коллагена. При дефиците этого витамина замедляются рост костей и заживление переломов.
• Витамин A поддерживает образование и рост кости. Недостаток витамина тормозит остеогенез и рост костей. Избыток витамина A вызывает зарастание эпифизарных хрящевых пластинок и замедление роста кости в длину.
Регуляция минерализации. Кальцитриол, необходимый для всасывания Ca2+ в тонком кишечнике, поддерживает процесс минерализации. Кальцитриол стимулирует минерализацию на уровне транскрипции, усиливая экспрессию остеокальцина. Дефицит витамина D3 приводит к нарушению минерализации кости, что и наблюдают при рахите у детей и остеомаляции у взрослых.
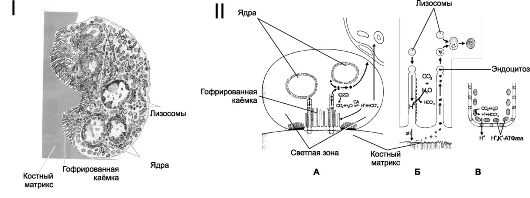 Рис. 18-4. КОСТНАЯ ТКАНЬ. I. Остеокласт [11].
Цитоплазматические выросты гофрированной каёмки направлены к
поверхности костного матрикса. В цитоплазме содержатся многочисленные
лизосомы. П. Остеокласт и резорбция кости [11]. При
взаимодействии остеокласта с поверхностью минерализованного костного
матрикса карбоангидраза II (СА II) катализирует образование Н+ и НС03". Н+ при помощи протонной Н+,К+-АТФазы
активно выкачивается из клетки, что приводит к закислению замкнутого
пространства лакуны. Гидролитические ферменты лизосом расщепляют
фрагменты костного матрикса. А - остеокласт на поверхности кости. Б - часть гофрированной каёмки. В - часть клеточной мембраны остеокласта в области гофрированной каёмки
Рис. 18-4. КОСТНАЯ ТКАНЬ. I. Остеокласт [11].
Цитоплазматические выросты гофрированной каёмки направлены к
поверхности костного матрикса. В цитоплазме содержатся многочисленные
лизосомы. П. Остеокласт и резорбция кости [11]. При
взаимодействии остеокласта с поверхностью минерализованного костного
матрикса карбоангидраза II (СА II) катализирует образование Н+ и НС03". Н+ при помощи протонной Н+,К+-АТФазы
активно выкачивается из клетки, что приводит к закислению замкнутого
пространства лакуны. Гидролитические ферменты лизосом расщепляют
фрагменты костного матрикса. А - остеокласт на поверхности кости. Б - часть гофрированной каёмки. В - часть клеточной мембраны остеокласта в области гофрированной каёмки
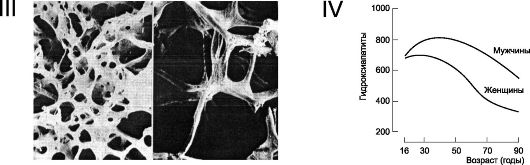 Рис. 18-4. Продолжение. III. Трабекулы костной ткани. Слева - в норме, справа - остеопороз. IV. Возрастная динамика костной массы. Для гидроксиапатита даны относительные значения
Рис. 18-4. Продолжение. III. Трабекулы костной ткани. Слева - в норме, справа - остеопороз. IV. Возрастная динамика костной массы. Для гидроксиапатита даны относительные значения
Регуляция резорбции
• Резорбцию кости усиливают ПТГ, интерлейкины 1 и 6, трансформирующий фактор роста а, Пг. Резорбцию кости поддерживают йодсодержащие гормоны щитовидной железы.
• Усиление резорбции под действием ПТГ не связано с прямым влиянием этого гормона на остеокласты, т.к. эти клетки не имеют рецепторов ПТГ. Активирующее влияние ПТГ и кальцитриола на остеокласты осуществляется опосредованно - через остеобласты. ПТГ и кальцитриол стимулируют образование фактора дифференцировки остеокластов - лиганда остеопротегерина.
• Резорбцию кости и активность остеокластов подавляют кальцитонин (через рецепторы в плазмолемме остеокластов) и γ-интерферон.
• Эстрогены ингибируют выработку ретикулярными клетками костного мозга колониестимулирующего фактора макрофагов (M- CSF), необходимого для образования остеокластов, что тормозит резорбцию кости.
НАДПОЧЕЧНИКИ
Надпочечники - парные органы, расположенные ретроперитонеально у верхних полюсов почки на уровне Th12 и L1. Формально это две железы - кора и мозговая часть, - имеющие разное происхождение (кора надпочечников развивается из мезодермы, хромаффинные клетки мозговой части - производные клеток нервного гребня). Различна и химическая структура синтезируемых гормонов: клетки коры надпочечников синтезирует стероидные гормоны (минералокортикоиды, глюкокортикоиды и предшественники андрогенов), хромаффинные клетки мозговой части - катехоловые амины. В то же время с функциональной точки зрения каждый надпочечник входит в состав единой системы быстрого реагирования на стрессовую ситуацию, обеспечивающую выполнение поведенческой реакции «беги или нападай». В этом контексте важны следующие обстоятельства, функционально обеспечивающие связь между симпатическим отделом нервной системы, хромаффинными клетками и глюкокортикоидами.
• Гуморальным эффектором реакции «беги или нападай» является выбрасываемый в кровоток из мозговой части надпочечников адреналин.
• Хромаффинные клетки образуют синапсы с преганглионарными симпатическими нейронами и расцениваются как постганглионарные клетки эфферентной симпатической иннервации, выбрасывающие в кровь адреналин в ответ на синаптическую секре-
цию ацетилхолина и его связывание с никотиновыми холинорецепторами.
• В мозговую часть надпочечников поступает содержащие глюкокортикоиды кровь из корковой части органа. Другими словами, синтез и секреция адреналина из хромаффинных клеток находятся под контролем глюкокортикоидов.
Кора надпочечников
Эпителиальные стероидогенные клетки коры надпочечников - в зависимости от их функции и морфологии - выглядят по-разному. Непосредственно под капсулой органа расположены клетки клубочковой зоны (занимают 15% от общего объёма коры), глубже лежат клетки пучковой зоны (70% объёма коры), а на границе с мозговым веществом - клетки сетчатой зоны. В разных зонах коры надпочечников синтезируются разные группы стероидных гормонов: минералокортикоиды, глюкокортикоиды и предшественники андрогенов.
• Минералокортикоиды (клубочковая зона). Альдостерон - основной минералокортикоид, его функция - поддержание баланса электролитов в жидкостях организма. В почке альдостерон увеличивает реабсорбцию Na+ (задержка натрия приводит к увеличению содержания воды в организме и повышению АД), увеличивает экскрецию ионов калия (потеря калия вызывает гипокалиемию), увеличивает реабсорбцию хлора, бикарбоната и экскрецию ионов водорода. Синтез альдостерона стимулирует ангиотензин II.
• Глюкокортикоиды (пучковая и сетчатая зоны). Кортизол - основной глюкокортикоид, на его долю приходится 80% всех глюкокортикоидов. Остальные 20% - кортизон, кортикостерон, 11-дезоксикортизол и 11-дезоксикортикостерон. Глюкокортикоиды контролируют метаболизм белков, углеводов и жиров, подавляют иммунные реакции, а также имеют противовоспалительный эффект. Синтез глюкокортикоидов стимулирует тропный гормон аденогипофиза - АКТГ.
• Предшественники андрогенов (пучковая и сетчатая зоны). Дегидроэпиандростерон и андростендион - предшественники андрогенов, их дальнейшие превращения происходят вне надпочечника и рассмотрены в главе 19. Гонадотропные гормоны гипофиза не влияют на секрецию половых гормонов в сетчатой зоне.
Глюкокортикоиды
Основной естественный глюкокортикоид, секретируемый надпочечниками, - кортизол (объём секреции - от 15 до 20 мг/сут,
• концентрация кортизола в крови около 12 мкг/100 мл). Для кортизола, а также для регулирующих его синтез и секрецию кортиколиберина и АКТГ характерна выраженная суточная периодичность. При нормальном ритме сна увеличение секреции кортизола наступает после засыпания и достигает максимума при пробуждении. В качестве ЛС в клинической практике обычно применяют синтетические глюкокортикоиды (дексаметазон, преднизолон, метилпреднизон и др.). Практически все глюкокортикоиды имеют одновременно и эффекты минералокортикоидов. Регуляция секреции глюкокортикоидов (рис. 18-5).
-Ф- Активирующие (нисходящие) влияния. Непосредственный активатор синтеза и секреции кортизола - АКТГ. АКТГ выделяется клетками передней доли гипофиза под действием кортиколиберина, поступающего в кровь гипоталамо-гипофизарной воротной системы из гипоталамуса. Стрессовые стимулы ак-
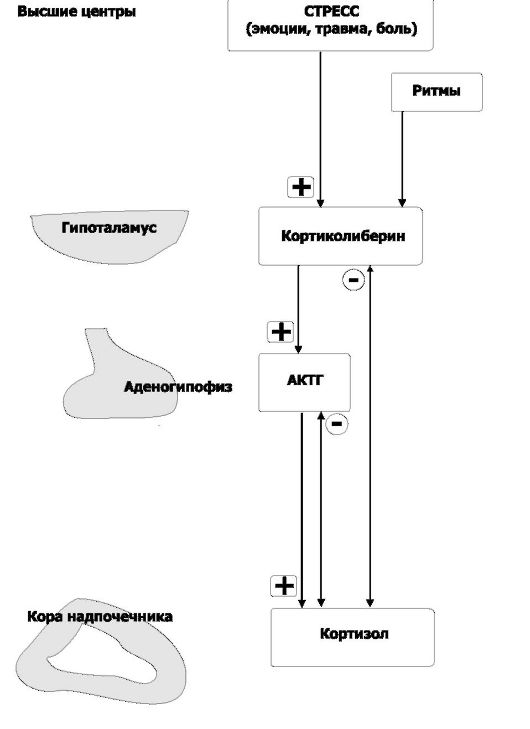 Рис. 18-5. Регуляторные контуры в системе «Гонадо- либерин-АКТГ-кортизол».
Рис. 18-5. Регуляторные контуры в системе «Гонадо- либерин-АКТГ-кортизол».
Символами «+» и «-» обозначены стимулирующие и тормозящие влияния
тивируют всю нисходящую систему влияний, вызывая быстрое выделение кортизола. Кортизол вызывает различные метаболические эффекты, направленные на снятие повреждающей природы состояния стресса. -Φ- Восходящие (тормозные) влияния по принципу отрицательной обратной связи реализует кортизол, подавляя секрецию АКТГ в передней доле гипофиза и кортиколиберина в гипоталамусе. Это уменьшает концентрацию кортизола в плазме в то время, когда организм не подвергается воздействию стресса.
• Метаболизм
-Ф- Связанные и свободные формы. Более 90% глюкокортикоидов циркулирует в крови в связи с белками - альбумином и связывающим кортикоиды глобулином (транскортин). Около 4% кортизола плазмы - свободная фракция.
-Ф- Время циркуляции определяется прочностью связывания с транскортином (время полужизни кортизола - до 2 часов, кортикостерона - менее 1 часа).
-Ф- Водорастворимые формы. Модификация липофильного кортизола осуществляется преимущественно в печени, формируются конъюгаты с глюкуронидом и сульфатом. Модифицированные глюкокортикоиды - водорастворимые соединения, способные к экскреции.
-Ф- Экскреция. Конъюгированные формы глюкокортикоидов секретируются с жёлчью в ЖКТ, из них 20% теряется с калом, 80% всасывается в кишечнике. Из крови 70% глюкокортикоидов экскретируется с мочой.
• Функции глюкокортикоидов разнообразны - от регуляции метаболизма до модификации иммунологического и воспалительного ответов.
-Ф- Углеводный обмен. Основные события разворачиваются между скелетными мышцами, жировыми депо организма и печенью. Основные пути метаболизма - стимуляция глюконеогенеза, синтез гликогена и уменьшение потребления глюкозы внутренними органами (кроме головного мозга). Основной эффект - увеличение концентрации глюкозы в крови. ♦ Глюконеогенез - синтез глюкозы за счёт аминокислот, лактата и жирных кислот, т.е. неуглеводных субстратов.
■ В скелетных мышцах глюкокортикоиды усиливают распад белков. Образующиеся аминокислоты поступают в печень.
■ В печени глюкокортикоиды стимулируют синтез ключевых ферментов обмена аминокислот - субстратов глюконеогенеза.
♦ Синтез гликогена усиливается за счёт активации гликогенсинтетазы. Запасаемый гликоген легко превращается в глюкозу путём гликогенолиза.
❖ Липидный обмен. Кортизол увеличивает мобилизацию жирных кислот - источник субстратов для глюконеогенеза.
♦ Липолиз усиливается в конечностях.
♦ Липогенез усиливается в других частях тела (туловище и лицо).
❖ Белки и нуклеиновые кислоты
♦ Анаболический эффект в печени.
♦ Катаболический эффект в других органах (в особенности в скелетных мышцах).
❖ Иммунная система. В высоких дозах глюкокортикоиды выступают как иммунодепрессанты (применяют для предупреждения отторжения трансплантированных органов, при тяжёлой псевдопаралитической миастении (myasthenia gravis), возникающей в результате появления аутоантител к никотиновым рецепторам ацетилхолина).
❖ Воспаление. Глюкокортикоиды имеют выраженный противовоспалительный эффект.
❖ Синтез коллагена. Глюкокортикоиды при длительном применении ингибируют синтетическую активность фибробластов и остеобластов, в результате развиваются истончение кожи и остеопороз.
❖ Скелетные мышцы. Длительное применение глюкокортикоидов поддерживает катаболизм мышц, что приводит к их атрофии и мышечной слабости.
❖ Воздухоносные пути. Введение глюкокортикоидов может уменьшить отёк слизистой оболочки, развивающийся, например, при бронхиальной астме.
❖ Физиологические реакции органов и систем организма, вызываемые кортизолом, приведены в таблице 18-6.
Альдостерон
Альдостерон - основной минералокортикоид. Нормальная концентрация альдостерона в крови около 6 нг на 100 мл, объём секреции - от 150 до 250 мкг/день. Другие стероиды надпочечников, расцениваемые как глюкокортикоиды (кортизол, 11-дезоксикорти- зол, 11-дезоксикортикостерон, кортикостерон), имеют и минералокортикоидную активность, хотя - сравнительно с альдостероном - их суммарный вклад в минералокортикоидную активность не столь велик.
Таблица 18-3. Физиологические реакции на кортизол
Органы и системы | Эффекты |
Гипоталамус | Прекращение секреции кортиколиберина и вазопрессина |
Гипофиз | Подавление образования и выделения АКТГ |
Сердце и сосуды | Потенцирование сосудосуживающего действия катехоламинов и их лигандов |
Дыхательная система | Ускорение образования сурфактанта |
Почки | Повышение скорости клубочковой фильтрации |
Мышцы | Понижение чувствительности к инсулину, усиление катаболизма белков |
Система иммунитета | Подавление иммунных реакций (иммуносупрессия) |
Костная система | Усиление резорбции кости, подавление остеогенеза |
Соединительная ткань | Уменьшение синтеза коллагена |
Жировая ткань | Блокирование поглощения глюкозы липоцитами |
• Регуляторы синтеза и секреции (рис. 18-6).
❖ Ангиотензин II - компонент ренин-ангиотензиновой системы - главный регулятор синтеза и секреции альдостерона. Этот пептид стимулирует выброс альдостерона.
❖ Сердечный натрийуретический фактор (атриопептин) ингибирует синтез альдостерона.
❖ Na+. Эффекты гипо- и гипернатриемии реализуются через ренин-ангиотензиновую систему.
❖ К+. Эффекты ионов калия не зависят от содержания в крови Na+ и ангиотензина II.
♦ Гиперкалиемия стимулирует секрецию альдостерона.
♦ Гипокалиемия тормозит секрецию альдостерона.
❖ Простагландины
♦ E1 и E2 стимулируют синтез альдостерона.
♦ F1a и F2a тормозят секрецию минералокортикоидов.
❖ Травмы и стрессовые состояния увеличивают секрецию альдостерона за счёт активирующего влияния АКТГ на кору надпочечников.
• Метаболизм. Альдостерон практически не связывается с белками плазмы крови, по этой причине время его циркуляции в крови (время полужизни) не превышает 15 минут. Альдостерон из крови удаляется печенью, где он трансформируется в экскретируемый почками тетрагидроальдостерон-3-глюкуронид.
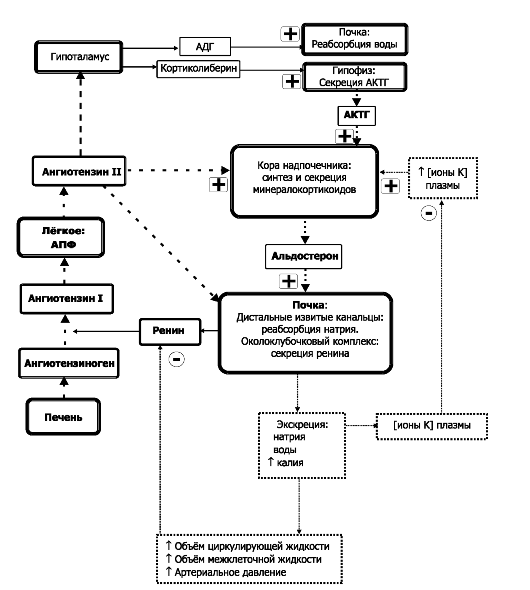 Рис. 18-6. Поддержание баланса электролитов в жидкостях организма. Символами «+» и «-» обозначены стимулирующие и тормозящие влияния. АПФ - ангиотензин-превращающий фермент
Рис. 18-6. Поддержание баланса электролитов в жидкостях организма. Символами «+» и «-» обозначены стимулирующие и тормозящие влияния. АПФ - ангиотензин-превращающий фермент
• Рецептор альдостерона - внутриклеточный (ядерный) полипептид - связывает альдостерон и активирует транскрипцию генов, в первую очередь генов Na+,К+-АТФазы и сочетанного трансмембранного переносчика Na+, К+ и Cl-. Рецепторы альдостерона обнаружены в эпителиальных клетках почечных канальцев, слюнных и потовых железах. Высокоаффинный рецептор в системах in vitro также связывает кортизол, но in vivo взаимодействие кортизола и рецептора практически не происходит, так как внутриклеточная 11β-гидроксистероид дегидрогеназа превращает кортизол в кортизон, плохо связывающийся с рецептором минералокортикоидов. Следовательно, глюкокортикоид кортизол не проявляет в клетках-мишенях эффекта минералокортикоида.
• Функция минералокортикоидов - поддержание баланса электролитов в жидкостях организма - осуществляется посредством влияния на реабсорбцию ионов в почечных канальцах (дистальные извитые канальцы и начальный отдел собирательных трубочек).
❖ Na+. Альдостерон увеличивает реабсорбцию ионов натрия. Задержка натрия приводит к увеличению содержания воды в организме и повышению АД.
❖ К+. Альдостерон увеличивает экскрецию ионов калия. Потеря калия вызывает гипокалиемию.
❖ Cl-, HCO3-, Н+. Альдостерон увеличивает реабсорбцию хлора, бикарбоната и почечную экскрецию ионов водорода.
Хромаффинная ткань
Эндокринную функцию мозговой части надпочечника выполняют происходящие из нервного гребня хромаффинные клетки, формирующие также параганглии. Мелкие скопления и одиночные хромаффинные клетки находят также в сердце, почках, симпатических ганглиях. Для хромаффинных клеток характерны содержащие либо адреналин (их большинство), либо норадреналин гранулы с электроноплотным содержимым, которое с бихроматом калия даёт хромаффинную реакцию. В гранулах также содержатся АТФ и хромогранины.
Катехоловые амины
• Синтез. Катехоламины синтезируются из тирозина по цепочке: тирозин (превращение тирозина катализирует тирозин гидроксилаза) - ДОФА (ДОФА-декарбоксилаза) - дофамин (дофамин- β-гидроксилаза) - норадреналин (фенилэтаноламин-N-метилт- рансфераза) - адреналин.
❖ ДОФА (диоксифенилаланин). Эта аминокислота выделена из бобов Vicia faba L, как антипаркинсоническое средство применяется её L-форма - леводопа (L-ДОФА, леводофа, 3-гидро- кси-L-тирозин, L-дигидроксифенилаланин).
❖ Дофамин - 4-(2-аминоэтил)пирокатехол.
❖ Норадреналин [2-амино-1-(3,4-дигидроксифенил)этанол] - деметилированный предшественник адреналина. Фермент синтеза норадреналина (дофамин--гидроксилаза) секретируется из хромаффинных клеток и норадренергических терминалей вместе с норадреналином.
❖ Адреналин - /-1-(3,4-дигидроксифенил)-2-(метиламино)этанол - только гуморальный фактор, в синаптической передаче не участвует.
• Секреция. При активации симпатической нервной системы хромаффинные клетки выбрасывают в кровь катехоловые амины (преимущественно адреналин). Вместе с катехоламинами из гранул выделяются АТФ и белки. Адреналин-содержащие клетки содержат также опиоидные пептиды (энкефалины) и секретируют их вместе с адреналином.
• Метаболизм адреналина и других биогенных аминов происходит под влиянием катехол-О-метилтрансферазы и моноаминооксидаз. В результате образуются экскретируемые с мочой метанефрины и ванилилминдальная кислота соответственно. Время полураспада катехоламинов в плазме около 2 минут. У здорового мужчины в положении лёжа содержание в крови норадреналина составляет около 1,8 нмоль/л, адреналина - 16 нмоль/л и дофамина - 0,23 нмоль/л.
• Эффекты. Катехоламины имеют широкий спектр эффектов (воздействие на гликогенолиз, липолиз, глюконеогенез, существенное влияние на сердечно-сосудистую систему). Вазоконстрикция, параметры сокращения сердечной мышцы и другие эффекты катехоловых аминов реализуются через α- и β-адренергические рецепторы на поверхности клеток-мишеней (ГМК, секреторные клетки, кардиомиоциты).
• Рецепторы катехоловых аминов - адренергические.
❖ Адренорецепторы клеток-мишеней (включая синаптические) связывают норадреналин, адреналин и различные адренергические ЛС (активирующие - агонисты, адреномиметики, блокирующие - антагонисты, адреноблокаторы). Адренергические рецепторы подразделяют на α- и β-подтипы. Среди α- и β-адренорецепторов различают: α1- (например, постсинаптические в симпатическом отделе вегетативной нервной систе-
мы), α2- (например, пресинаптические в симпатическом отделе вегетативной нервной системы и постсинаптические в головном мозге), β 1- (например, кардиомиоциты), β2- и β3-адренорецепторы. Адренорецепторы связаны с G-белком. ❖ Эффекты, опосредуемые разными подтипами адренергических рецепторов (см. также главу 15).
αι
Гликогенолиз. Усиление.
ГМК сосудов и мочеполовой системы. Сокращение. α2
ГМК ЖКТ. Расслабление.
Липолиз. Подавление.
Инсулин, ренин. Подавление секреции.
Кардиомиоциты. Увеличение силы сокращения. Липолиз. Усиление.
β2
Инсулин, глюкагон, ренин. Усиление секреции. ГМК бронхов, ЖКТ, кровеносных сосудов, мочеполовой системы. Расслабление. Печень. Усиление гликогенолиза и глюконеогенеза. Мышцы. Усиление гликогенолиза.
β3
Липолиз. Усиление. Аварийная функция симпатоадреналовой системы
«Аварийная функция симпатоадреналовой системы» («реакция борьбы», ситуация «беги или нападай»), как часто называют разнообразные эффекты внезапного повышенного выброса адреналина в кровь, представлена в таблице 18-3.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
Поджелудочная железа содержит от 0,5 до 2 млн мелких скоплений эндокринных клеток - островков Лангерханса. В островках идентифицировано несколько типов эндокринных клеток, синтезирующих и секретирующих пептидные гормоны: инсулин (β-клетки, 70% от всех островковых клеток), глюкагон (α-клетки, 15%), соматостатин (δ-клетки), панкреатический полипептид (PP-клетки, seu F-клетки) и у детей младшего возраста - гастрины (G-клетки, seu D-клетки).
• Инсулин - главный регулятор энергетического обмена в организме - контролирует обмен углеводов (стимуляция гликолиза и
Таблица 18-4. Физиологические изменения при реакции «борьбы»
Органы | Результат воздействия |
ЦНС | Увеличение кровотока и потребления глюкозы |
Сердце и сосуды | Рост частоты и силы сердечных сокращений Сужение артериол в висцеральных органах (исключая мозг, сердце и скелетные мышцы). Повышение АД |
Система дыхания | Расширение средних и мелких бронхов. Увеличение глубины и частоты дыхания |
Мышечная система | Увеличение кровотока. Повышение гликогенолиза. Повышение сократимости |
ЖКТ | Уменьшение кровотока, секреции, синтеза белков |
Печень | Стимуляция образования глюкозы (усиление глюконеогенеза и гликогенолиза, подавление синтеза гликогена) |
Жировая ткань | Повышение липолиза, уровня жирных кислот и глицерина в плазме |
Почки | Снижение кровотока, уменьшение образования мочи |
Скелет | Снижение поглощения и усвоения глюкозы |
подавление глюконеогенеза), липидов (стимуляция липогенеза), белков (стимуляция синтеза белка), а также стимулирует пролиферацию клеток (митоген). Основные органы-мишени инсулина - печень, скелетные мышцы и жировая ткань.
• Глюкагон - антагонист инсулина - стимулирует гликогенолиз и липолиз, что ведёт к быстрой мобилизации источников энергии (глюкоза и жирные кислоты). Ген глюкагона кодирует также структуру так называемых энтероглюкагонов - глицентина и глюкагоноподобного пептида 1 - стимуляторов секреции инсулина.
• Соматостатин подавляет в островках поджелудочной железы секрецию инсулина и глюкагона.
• Панкреатический полипептид состоит из 36 аминокислотных остатков. Его относят к регуляторам пищевого режима (в частности, этот гормон угнетает секрецию экзокринной части поджелудочной железы). Секрецию гормона стимулируют богатая белком пища, гипогликемия, голодание, физическая нагрузка.
• Гастрины I и II (идентичные 17-аминокислотные пептиды отличаются наличием сульфатной группы у тирозила в положении 12) стимулирует секрецию соляной кислоты в желудке. Стимулятор секреции - гастрин-освобождающий гормон, ингибитор сек-
реции - соляная кислота. Рецептор гастрина/холецистокинина обнаружен в ЦНС и слизистой оболочке желудка.
Инсулин
Транскрипция гена инсулина приводит к образованию мРНК препроинсулина, содержащей последовательности A, C и B, а также нетранслируемые 3'- и 5'-концы. После трансляции образуется полипептидная цепочка проинсулина, состоящая с N-конца из последовательных доменов B, C и A. В комплексе Гольджи протеазы расщепляют проинсулин на 3 пептида: A (21 аминокислота), B (30 аминокислот) и C (31 аминокислота). Пептиды A и B, интегрируясь при помощи дисульфидных связей, образуют димер - инсулин. Секреторные гранулы содержат эквимолярные количества гормонально активного инсулина и не имеющего гормональной активности C-пептида, а также следы проинсулина.
Секреция инсулина
Количество инсулина, секретируемого на фоне относительного голодания (например, утром до завтрака), составляет около 1 Ед/ час; оно возрастает в 5-10 раз после приёма пищи. В среднем в течение дня взрослый здоровый мужчина секретирует 40 Ед (287 ммоль) инсулина.
Содержимое секреторных гранул β-клеток поступает в кровь в результате экзоцитоза, наступающего в результате увеличения содержания внутриклеточного Ca2+. Именно внутриклеточный кальций (точнее ↑[Ca2+]) является непосредственным и главным сигналом к секреции инсулина. Способствуют экзоцитозу также активированные ↑[цАМФ] протеинкиназа A и активированная Т[диацилглице- рол] протеинкиназа C, фосфорилируя некоторые белки, принимающие участие в экзоцитозе.
• Регуляторы секреции инсулина
❖ СТИМУЛИРУЮТ секрецию инсулина гипергликемия (повышенное содержание глюкозы в плазме крови), гиперкалиемия, некоторые аминокислоты, ацетилхолин, глюкагон и некоторые другие гормоны, приём пищи, а также производные сульфонилмочевины.
♦ Глюкоза - главный регулятор секреции инсулина
1. При повышении содержания глюкозы в плазме крови (более 5 мМ, см. таблицу 18-5) молекулы этого сахара, а также молекулы галактозы, маннозы, b-кетокислоты входят в β-клетки путём облегченной диффузии через трансмембранный переносчик (импортёр) глюкозы GLUT2.
2. Вошедшие в клетку молекулы сахара подвергаются гликолизу, в результате чего в цитоплазме возрастает содержание АТФ.
3. Увеличенное содержание внутриклеточного АТФ закрывает чувствительные к АТФ и к [K+] калиевые каналы плазматической мембраны, что неизбежно приводит к её деполяризации.
4. Деполяризация плазматической мембраны β-клеток открывает потенциалочувствительные кальциевые каналы плазматической мембраны, в результате в клетку из межклеточного пространства входят ионы кальция.
5. Увеличение [Ca2+] в цитозоле стимулирует экзоцитоз секреторных гранул, инсулин этих гранул оказывается вне β- клеток.
♦ Гиперкалиемия. Повышение содержания К+ во внутренней среде организма блокирует чувствительные к [K+] калиевые каналы плазматической мембраны, что приводит к её деполяризации. Дальнейшие события разворачивается так, как описано выше (см. позиции 4 и 5).
♦ Аминокислоты (в особенности аргинин, лейцин, аланин и лизин) поступают в β-клетки при помощи трансмембранного переносчика аминокислот и метаболизируют в митохондриальном цикле трикарбоновых кислот. В результате в клетке возрастает содержание АТФ. Дальнейшие события разворачивается так, как описано выше (см. позиции 3, 4 и 5).
♦ Производные сульфонилмочевины блокируют калиевые каналы в плазмолемме β-клеток, взаимодействуя с рецептором сульфонилмочевины в составе К+ и АТФ-чувствительных калиевых каналов плазматической мембраны, что приводит к её деполяризации. Дальнейшие события разворачивается так, как описано выше (см. позиции 4 и 5).
♦ Ацетилхолин, секретируемый из окончаний нервных волокон правого блуждающего нерва, взаимодействует с мускариновыми холинергическими рецепторами плазматической мембраны, связанными с G-белком. G-белок активирует фосфолипазу C, что приводит к отщеплению от фосфоинозитолбифосфата фосфолипидов клеточной мембраны двух вторых посредников - цитозольного ИТФ и мембранного диацилглицерола.
■ ИТФ, связываясь с его рецепторами, стимулирует выброс Ca2+ из цистерн гладкой эндоплазматической сети, что приводит к экзоцитозу секреторных гранул с инсулином.
■ Диацилглицерол активирует протеинкиназу C, что приводит к фосфорилированию некоторых белков, принимающие участие в экзоцитозе, в результате происходит секреция инсулина.
♦ Холецистокинин взаимодействует с его рецепторами (связанные с G-белком рецепторы). G-белок активирует фосфолипазу C. Дальнейшие события происходят так, как описано выше для ацетилхолина.
♦ Гастрин связывается с рецептором холецистокинина типа B. Дальнейшие события происходят так, как описано выше для холецистокинина и ацетилхолина.
♦ Гастрин-рилизинг гормон также стимулирует секрецию инсулина.
♦ Глюкагоноподобный пептид 1 (см. ниже) - самый мощный стимулятор секреции инсулина.
❖ ИНГИБИТОРЫ СЕКРЕЦИИ ИНСУЛИНА
♦ Адреналин и норадреналин (через α2-адренорецепторы и уменьшение содержания цАМФ) подавляют секрецию инсулина. Через β-адренорецепторы (происходит увеличение содержания цАМФ) эти агонисты стимулируют секрецию инсулина, но в островках Лангерханса преобладают α-адренорецепторы, в результате наблюдается угнетение секреции инсулина.
■ Физическая нагрузка сопровождается активацией симпатического отдела нервной системы и поглощением глюкозы (как источника энергии) скелетными мышцами, что при одновременном гипогликемическом (понижающем содержание сахара в плазме крови) эффекте инсулина может привести к выраженной гипогликемии (что в первую очередь сказывается на функциях мозга). В этом контексте подавляющий секрецию инсулина эффект адреналина и норадреналина представляется весьма уместным.
■ Стресс. Подавляющая секрецию инсулина роль адреналина особенно велика во время развития стресса, когда симпатическая система возбуждена. Адреналин одновременно повышает концентрацию глюкозы и жирных кислот в плазме крови. Смысл этого двойного эффекта следующий: адреналин вызывает мощный гликогенолиз в печени, вызывая в течение нескольких минут выделение значительного количества глюкозы в кровь, и в это же время оказывает прямое липолитическое действие на клетки жировой ткани, повышая в крови концентрацию жирных кислот.
Следовательно, адреналин создаёт условия для использования жирных кислот в условиях стресса. ♦ Соматостатин и нейропептид галанин, связываясь со своими рецепторами, приводят к уменьшению внутриклеточного содержания цАМФ и подавляют секрецию инсулина. ❖ Пищевой режим имеет исключительное значение как для секреции инсулина и содержания глюкозы в плазме крови, так и для состояния зависимого от инсулина обмена белка, жиров и углеводов в органах-мишенях инсулина (см. таблицу 18-5). Метаболизм инсулина. Инсулин и C-пептид в крови циркулируют в свободной форме от 3 до 5 минут. Более половины инсулина расщепляется в печени сразу же по поступлении в этот орган по портальным венам. C-пептид не разрушается в печени, а выделяется через почки. По этим причинам достоверным лабораторным показателем секреции инсулина является не сам гормон (инсулин), а именно C-пептид.
Физиологические эффекты инсулина
• Органы-мишени инсулина. Основные мишени инсулина - печень, скелетные мышцы, клетки жировой ткани. Поскольку инсулин является главным регулятором метаболизма молекул, -
Таблица 18-5. Влияние голодания и приёма пищи на содержание и эффекты инсулина [4]
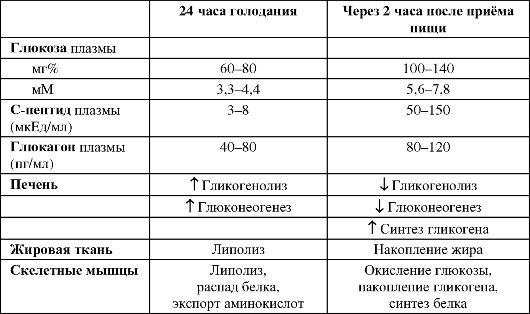 источников
энергетического обмена в организме - то именно в этих органах
развёртываются основные физиологические эффекты инсулина на обмен
белков, жиров и углеводов.
источников
энергетического обмена в организме - то именно в этих органах
развёртываются основные физиологические эффекты инсулина на обмен
белков, жиров и углеводов.
• Функции инсулина разнообразны (регуляция обмена источников энергии - углеводов, липидов и белков). В клетках-мишенях инсулин стимулирует трансмембранный перенос глюкозы и аминокислот, синтез белка, гликогена и триглицеридов, гликолиз, а также рост и пролиферацию клеток, но подавляет протеолиз, липолиз и окисление жиров (см. подробнее ниже).
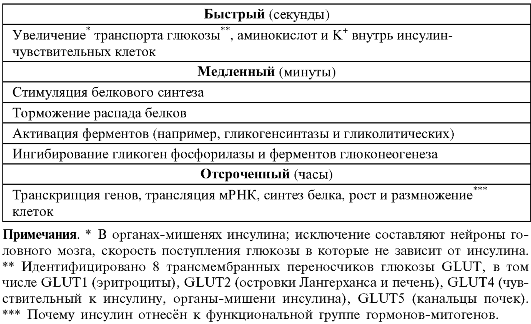
• Скорость проявления эффектов инсулина. Физиологические эффекты инсулина по скорости их наступления после взаимодействия гормона с его рецепторами подразделяют на быстрые (развиваются в течение секунд), медленные (минуты) и отсроченные (табл. 18-6).
Таблица 18-6. Долговременность эффектов инсулина [2]
 Влияние инсулина на обмен углеводов
Влияние инсулина на обмен углеводов
• Печень. Инсулин оказывает на гепатоциты следующие эффекты: ❖ глюкоза постоянно поступает в клетки печени через трансмембранный переносчик GLUT2; инсулин мобилизует дополнительный трансмембранный переносчик GLUT4, способствуя его встраиванию в плазматическую мембрану гепатоцитов.
❖ способствует синтезу гликогена из поступающей в гепатоциты глюкозы путём увеличения транскрипции гена глюкокиназы и активирования гликоген синтазы;
❖ предупреждает распад гликогена путём ингибирования активности гликоген фосфорилазы и глюкозо-6-фосфатазы;
❖ стимулирует гликолиз и окисление углеводов путём активирования глюкокиназы, фосфофруктокиназы и пируват киназы;
❖ активирует метаболизм глюкозы через гексозомонофосфатный шунт;
❖ ускоряет окисление пирувата путём активирования пируватдегидрогеназы;
❖ подавляет глюконеогенез путём ингибирования активности фосфоенолпируват карбоксикиназы, фруктозо-1,6-бифосфатазы и глюкозо-6-фосфатазы;
• Скелетные мышцы. В скелетных мышцах инсулин:
❖ активирует поступление глюкозы в саркоплазму посредством трансмембранного переносчика GLUT4, способствуя его встраиванию в плазматическую мембрану.
❖ способствует синтезу гликогена из поступающей в гепатоциты глюкозы путём увеличения транскрипции гена гексокиназы и активирования гликоген синтазы;
❖ стимулирует гликолиз и окисление углеводов путём активирования гексокиназы, фосфофруктокиназы и пируват киназы;
• Жировая ткань. Инсулин влияет на метаболизм адипоцитов следующим образом:
❖ активирует поступление глюкозы в саркоплазму посредством трансмембранного переносчика GLUT4, способствуя его встраиванию в плазматическую мембрану.
❖ стимулирует гликолиз, что приводит к образованию α-глицерофосфата, идущего на построение триглицеридов;
❖ ускоряет окисление пирувата путём активирования пируватдегидрогеназы и ацетил-КоА-карбоксилазы, что благоприятствует синтезу свободных жирных кислот;
• ЦНС. Инсулин практически не оказывает влияния ни на транспорт глюкозы в нервные клетки, ни на их метаболизм. Нейроны головного мозга отличаются от клеток других органов тем, что они используют как основной источник энергии преимущественно глюкозу, но не жирные кислоты. Более того, нервные клетки не приспособлены синтезировать глюкозу. Именно поэтому бесперебойное поступление глюкозы в головной мозг столь важно для функционирования и выживания нейронов.
• Другие органы. Как и ЦНС, многие органы (например, почка и кишечник) не чувствительны к инсулину.
Гомеостаз глюкозы
Содержание глюкозы во внутренней среде организма должно находиться в строго ограниченных пределах. Так, натощак концентрация глюкозы в плазме крови колеблется в пределах 60-90 мг% (нормогликемия), увеличивается до 100-140 мг% (гипергликемия) в течение одного часа после еды и обычно в течение 2 часов возвращается к нормальным значениям. Существуют ситуации, когда концентрация глюкозы в плазме крови уменьшается до 60 мг% и ниже (гипогликемия). Необходимость поддержания постоянной концентрации глюкозы в крови диктуется тем, что мозг, сетчатка и некоторые другие клетки и органы в качестве источника энергии используют преимущественно глюкозу. Так, в периоды между при- ёмами пищи основная часть глюкозы, находящаяся во внутренней среде организме, используется для метаболизма мозга.
• Гомеостаз глюкозы поддерживают следующие механизмы.
❖ Печень демпфирует колебания концентрации глюкозы. Так, когда содержание глюкозы в крови повышается до высоких концентраций после приёма пищи и объём секреции инсулина увеличивается, то более 60% глюкозы, всосавшейся из кишечника, депонируется в печени в форме гликогена. В последующие часы, когда концентрация глюкозы и секреция инсулина снижаются, печень выделяет глюкозу в кровь.
❖ Инсулин и глюкагон реципрокно регулируют нормальное содержание глюкозы в крови. Повышение содержания глюкозы выше нормы посредством механизма обратной связи действует на β-клетки островков Лангерханса и вызывает повышенную секрецию инсулина. что приводит концентрацию глюкозы к норме. Понижение содержания глюкозы ниже нормы тормозит образование инсулина, но стимулирует секрецию глюкагона, что приводит содержание глюкозы к норме.
❖ Гипогликемия оказывает прямое влияние на гипоталамус, который возбуждает симпатическую нервную систему. В результате адреналин секретируется из надпочечников и увеличивает выделение глюкозы печенью.
❖ Продолжительная гипогликемия стимулирует выделение СТГ и кортизола, которые уменьшают скорость потребления глюкозы большинством клеток организма, что помогает возвращению концентрации глюкозы в крови к нормальному уровню.
• После приёма пищи всосавшиеся в кишечнике моносахариды, триглицериды и аминокислоты по воротным венам поступают в печень, где различные моносахариды превращаются в глюкозу. Глюкоза в печени хранится в виде гликогена (синтез гликогена происходит также в мышцах), в печени окисляется лишь малая часть глюкозы. Глюкоза, не захваченная гепатоцитами, оказывается в системе общей циркуляции и поступает в различные органы, где окисляется до воды и C02 и обеспечивает энергетические потребности этих органов.
❖ Инкретины. При поступления химуса в кишечник из эндокринных клеток его стенки во внутреннюю среду организма выделяются так называемые инкретины (желудочный ингибирующий пептид, энтероглюкагон глицентин и глюкагоноподобный пептид 1), потенцирующие вызванную глюкозой секрецию инсулина.
❖ Всасывание глюкозы из просвета кишечника происходит при помощи встроенных в апикальную плазматическую мембрану энтероцитов Na+-зависимых транспортёров сочетанного переноса ионов натрия и глюкозы, требующих (в отличие от переносчиков глюкозы GLUT) затрат энергии. Напротив, выход глюкозы из энтероцитов во внутреннюю среду организма, происходящий через плазмолемму их базальной части, происходит путём облегчённой диффузии.
❖ Выделение глюкозы через почки
♦ Фильтрация молекул глюкозы из просвета кровеносных капилляров почечных телец в полость капсулы Боумена-Шум- лянского осуществляется пропорционально концентрации глюкозы в плазме крови.
♦ Реабсорбция. Обычно вся глюкоза реабсорбируется в первой половине проксимальных извитых канальцев со скоростью 1,8 ммоль/мин (320 мг/мин). Реабсорбция глюкозы происходит (как и её всасывание в кишечнике) при помощи сочетанного переноса ионов натрия и глюкозы.
♦ Секреция. Глюкоза у здоровых лиц не секретируется в просвет канальцев нефрона.
♦ Глюкозурия. Глюкоза появляется в моче при её содержании в плазме крови свыше 10 мМ.
• Между приёмами пищи глюкоза поступает в кровь из печени, где образуется за счёт гликогенолиза (распад гликогена до глюкозы) и глюконеогенеза (образование глюкозы из аминокислот, лактата, глицерола и пирувата. Из-за малой активности глюкозо-6- фосфатазы глюкоза не поступает в кровь из мышц.
❖ В покое содержание глюкозы в плазме крови составляет 4,5- 5,6 мМ, а общее содержание глюкозы (расчёты для взрослого здорового мужчины) в 15 л межклеточной жидкости - 60 ммоль (10,8 г), что примерно соответствует ежечасному расходу этого сахара. Следует помнить, что ни в ЦНС, ни в эритроцитах глюкоза не синтезируется и не хранится в виде гликогена и в то же время является крайне важным источником энергии.
❖ Между приёмами пищи преобладают гликогенолиз, глюконеогенез и липолиз. Даже при непродолжительном голодании (24- 48 часов) развивается обратимое состояние, близкое к сахарному диабету - голодный диабет. При этом нейроны начинают использовать в качестве источника энергии кетоновые тела.
• При физической нагрузке потребление глюкозы возрастает в несколько раз. При этом увеличиваются гликогенолиз, липолиз и глюконеогенез, регулируемые инсулином, а также функциональными антагонистами инсулина (глюкагон, катехоламины, СТГ, кортизол).
❖ Глюкагон. Эффекты глюкагона (см. ниже).
❖ Катехоламины. Физическая нагрузка через гипоталамические центры (гипоталамический глюкостат) активирует симпатоадреналовую систему. В результате уменьшается выброс инсулина из β-клеток, увеличивается секреция глюкагона из α-клеток, возрастает поступление в кровь глюкозы из печени, усиливается липолиз. Катехоламины также потенцируют вызванное T3 и T4 увеличение потребления кислорода митохондриями.
❖ Гормон роста способствует увеличению содержания глюкозы в плазме крови за счёт усиления гликогенолиза в печени, уменьшения чувствительности мышц и жировых клеток к инсулину (в результате уменьшается поглощение ими глюкозы), а также за счёт стимуляции выброса глюкагона из α-клеток.
❖ Глюкокортикоиды стимулируют гликогенолиз и глюконеогенез, но подавляют транспорт глюкозы из крови в разные клетки.
• Глюкостат. Регуляция содержания глюкозы во внутренней среде организма имеет целью поддержание гомеостаза этого сахара в пределах нормальных значений (концепция глюкостата) и осуществляется на разных уровнях. Выше рассмотрены механизмы поддержания гомеостаза глюкозы на уровне поджелудочной железы и органов-мишеней инсулина (периферический глюкостат). Считают, что центральную регуляцию содержания глюкозы (цен-
тральный глюкостат) осуществляют чувствительные к инсулину нервные клетки гипоталамуса, посылающие далее сигналы активации симпатоадреналовой системы, а также к синтезирующим кортиколиберин и соматолиберин нейронам гипоталамуса. Отклонения содержания глюкозы во внутренней среде организма от нормальных значений, о чём судят по содержанию глюкозы в плазме крови, приводят к развитию гипергликемии или гипогликемии.
❖ Гипогликемия - снижение содержания глюкозы в крови менее 3,33 ммоль/л. Гипогликемия может возникать у здоровых лиц через несколько дней голодания. Клинически гипогликемия проявляется при снижении уровня глюкозы ниже 2,4-3,0 ммоль/л. Ключ к диагностике гипогликемии - триада Уиппла: нервно-психические проявления при голодании, глюкоза крови менее 2,78 ммоль/л, купирование приступа пероральным или внутривенным введением раствора декстрозы (40-60 мл 40% раствора глюкозы). Крайнее проявление гипогликемии - гипогликемическая кома.
❖ Гипергликемия. Массовое поступление глюкозы во внутреннюю среду организма приводит к увеличению её содержания в крови - гипергликемии (содержание глюкозы в плазме крови превышает 6,7 мМ.). Гипергликемия стимулирует секрецию инсулина из β-клеток и подавляет секрецию глюкагона из α- клеток островков Лангерханса. Оба гормона блокируют в печени образование глюкозы как в ходе гликогенолиза, так и глюконеогенеза. Гипергликемия - так как глюкоза является осмотически активным веществом - может привести к обезвоживанию клеток, развитию осмотического диуреза с потерей электролитов. Гипергликемия может вызвать повреждение многих тканей, в особенности кровеносных сосудов. Гипергликемия - характерный симптом сахарного диабета.
Влияние инсулина на жировой обмен
• Печень. Инсулин в гепатоцитах:
❖ способствует синтезу жирных кислот из глюкозы путём активирования ацетил-КоА-карбоксилазы и синтазы жирных кислот. Жирные кислоты, присоединяя α-глицерофосфат, превращаются в триглицериды.
❖ подавляет окисление жирных кислот вследствие увеличенного превращения ацетил-КоА в малонил-КоА. Малонил-КоА ингибирует активность карнитин ацилтрансферазы (транспортирует жирные кислоты из цитоплазмы в митохондрии для их
β-окисления и превращения в кетокислоты. Другими словами, инсулин оказывает антикетогенный эффект.
• Жировая ткань. В липоцитах инсулин способствует превращению свободных жирных кислот в триглицериды и их отложению в виде жира. Этот эффект инсулина осуществляется несколькими путями. Инсулин:
❖ увеличивает окисление пирувата путём активирования пируватдегидрогеназы и ацетил-КоА-карбоксилазы, что благоприятствует синтезу свободных жирных кислот;
❖ увеличивает транспорт глюкозы в липоциты, последующее превращение которой приводит к появлению α-глицерофосфата.
❖ способствует синтезу триглицеридов из α-глицерофосфата и свободных жирных кислот;
❖ предупреждает расщепление триглицеридов на глицерол и свободные жирные кислоты, ингибируя активность гормон-чувствительной триглицерид липазы;
❖ активирует синтез липопротеин липазы, транспортируемой к клеткам эндотелия, где этот фермент расщепляет триглицериды хиломикронов и липопротеинов очень низкой плотности.
Эффекты инсулина на белковый обмен и рост организма
Инсулин в печени, скелетных мышцах, а также в других органахмишенях и клетках-мишенях стимулирует синтез белка и подавляет его катаболизм. Другими словами, инсулин - сильный анаболический гормон. Анаболическое влияние инсулина реализуется несколькими путями. Инсулин:
• стимулирует поглощение аминокислот клетками;
• усиливает транскрипцию генов и трансляцию мРНК;
• подавляет распад белков (особенно мышечных) и их освобождение в кровь;
• уменьшает скорость глюконеогенеза из аминокислот. Анаболические эффекты инсулина и гормона роста синергестич-
ны, что не в последнюю очередь определяется тем обстоятельством, что эффекты гормона роста реализуются посредством инсулиноподобного фактора роста - соматомедина C.
Глюкагон и глюкагоноподобные пептиды
Ген глюкагона содержит последовательности, кодирующие структуру нескольких физиологически родственных гормонов с эффектами глюкагона. В результате транскрипции образуется мРНК препроглюкагона, но эта мРНК по-разному расщепляется (дифференциальный сплайсинг) в α-клетках островков Лангерханса и эндок-
ринных L-клетках слизистой оболочки верхних отделов тонкого кишечника, приводя к образованию разных мРНК проглюкагона.
• Глицентин состоит из 69 аминокислотных остатков, стимулирует секрецию инсулина и желудочного сока, а также принимает участие в регуляции моторики ЖКТ. Глицентин обнаружен также в нервных клетках гипоталамуса и ствола мозга.
• Глюкагоноподобный пептид 1 (аминокислотные последовательности 7-37) - самый мощный стимулятор вызванной глюкозой секреции инсулина (именно поэтому, в частности, тест на толерантность к глюкозе проводят перорально, а не внутривенно). Это пептид подавляет желудочную секрецию и расценивается как физиологический медиатор чувства насыщения. Пептид синтезируется также в нейронах паравентрикулярного ядра гипоталамуса и нейронах центрального ядра миндалевидного тела. Обе группы нервных клеток принимают непосредственное участие в регуляции пищевого поведения.
• Глюкагоноподобный пептид 2 стимулирует пролиферацию клеток кишечных крипт и всасывание в тонком кишечнике.
Секреция глюкагона
Внутриклеточные события, обеспечивающие секрецию глюкагона из α-клеток, происходят по тем же механизмам, что и секреция инсулина из β-клеток (см. выше раздел «Регуляторы секреции инсулина»), но те же внеклеточные сигналы, запускающие секрецию глюкагона, зачастую (но не всегда!) приводят к противоположным результатам.
• Стимулируют секрецию глюкагона аминокислоты (в особенности аргинин и аланин), гипогликемия, инсулин, гастрин, холецистокинин, кортизол, физическая нагрузка, голодание, b-адре- нергические стимуляторы, приём пищи (особенно богатой белком).
• Подавляют секрецию глюкагона глюкоза, инсулин, соматостатин, секретин, свободные жирные кислоты, кетоновые тела, a- адренергические стимуляторы.
Время полужизни глюкагона в крови - около 5 минут.
Физиологические эффекты глюкагона
Основная мишень глюкагона - печень (гепатоциты), в меньшей степени - адипоциты и поперечно-полосатая мышечная ткань (в том числе кардиомиоциты). Рецептор глюкагона расположен в плазмолемме клеток-мишеней, связывает только глюкагон и посредством G-белка активирует аденилатциклазу. Мутации гена глюка-
гонового рецептора приводят к развитию инсулин-независимого сахарного диабета. Глюкагон расценивают как антагонист инсулина; этот гормон стимулирует гликогенолиз и липолиз, что ведёт к быстрой мобилизации источников энергии (глюкоза и жирные кислоты). В то же время глюкагон обладает кетогенным эффектом, т.е. стимулирует образование кетоновых тел.
• Глюкагон увеличивает содержание глюкозы (способствует гипергликемии) в плазме крови. Этот эффект реализуется несколькими путями.
❖ Стимуляция гликогенолиза. Глюкагон, активируя гликоген фосфорилазу и ингибируя гликоген синтазу в гепатоцитах, приводит к быстрому и выраженному распаду гликогена и освобождению глюкозы в кровь.
❖ Подавление гликолиза. Глюкагон ингибирует ключевые ферменты гликолиза (фосфофруктокиназа, пируваткиназа) в печени, что приводит увеличению содержания глюкозо-6-фос- фата в гепатоцитах, его дефосфорилированию и освобождению глюкозы в кровь.
❖ Стимуляция глюконеогенеза. Глюкагон усиливает транспорт аминокислот из крови в гепатоциты и одновременно активирует ключевые ферменты глюконеогенеза (пируват карбоксилаза, фруктозо-1,6-дифосфатаза), что приводит к увеличению содержания глюкозы в цитоплазме клеток и её поступлению в кровь.
• Глюкагон способствует образованию кетоновых тел путём стимуляции окисления жирных кислот: ингибирование активности ацетил-КоА-карбоксилазы приводит к уменьшению содержания ингибитора карнитин ацилтрансферазы - малонил-КоА, что приводит к усиленному поступлению жирных кислот из цитоплазмы в митохондрии для их β-окисления и превращения в кетокислоты. Другими словами, в отличие от инсулина, глюкагон оказывает кетогенный эффект.
ЯИЧКИ
В яичках синтезируются стероидные андрогены и α-ингибин. Их физиологическое значение рассмотрено в главе 19, здесь приведены краткие характеристики гормонов.
• Стероидные андрогены вырабатываются интерстициальными клетками Ляйдига (тестостерон и дигидротестостерон) и клетками сетчатой зоны коры надпочечников (дегидроэпиандростерон и андростендион, обладающие слабой андрогенной активностью.
❖ Тестостерон - основной циркулирующий андроген. В эмбриогенезе андрогены контролируют развитие плода по мужскому
типу. В период полового созревания они стимулируют становление признаков мужского пола. С наступлением половой зрелости тестостерон необходим для поддержания сперматогенеза, вторичных половых признаков, секреторной активности предстательной железы и семенных пузырьков.
❖ Дигидротестостерон. 5α-Редуктаза катализирует превращение тестостерона в дигидротестостерон в клетках Ляйдига, простате, семенных пузырьках.
• α-Ингибин. Этот гликопротеидный гормон синтезируется в клетках Сертоли извитых семенных канальцев и блокирует синтез гипофизарного ФСГ.
ЯИЧНИКИ
В яичниках синтезируются стероидные женские половые гормоны, гликопротеиновые гормоны ингибины и пептидной природы релаксины. Их физиологическое значение рассмотрено в главе 19, здесь приведены краткие характеристики гормонов.
• Женские половые гормоны эстрогены (эстрадиол, эстрон, эстриол) и прогестины (прогестерон) - стероиды.
❖ Эстрогены в период полового созревания стимулируют становление признаков женского пола. У женщин детородного возраста эстрогены активируют пролиферацию фолликулярных клеток, а в эндометрии контролируют пролиферативную фазу менструального цикла.
♦ Эстрадиол (17β-эстрадиол, Е2) - 17β-эстра-1,3,5(10)-триен- 3,17-диол - образуется из тестостерона путём его ароматизации, обладает выраженной эстрогенной активностью. Образование ароматических С18-эстрогенов из С19-андрогенов катализирует ароматаза, называемая также эстроген синтаза. Синтез этого фермента в яичнике индуцирует ФСГ.
♦ Эстрон (Е1) - 3-гидроксиэстра-1,3,5(10)-триен-17-он - метаболит 17β-эстрадиола, образуется путём ароматизации андростендиона, имеет небольшую эстрогенную активность, выделяется с мочой беременных.
♦ Эстриол - 16α,17β-эстри-1,3,5(10)-триен-3,16,17-триол - образуется из эстрона. Этот слабый эстроген экскретируется с мочой беременных, в значительном количестве присутствует в плаценте.
♦ Рецептор эстрогенов относится к ядерным, полипептид из 595 аминокислотных остатков, имеет выраженную гомологию с протоонкогеном v-erbA.
❖ Прогестерон относится к прогестинам, его синтезируют клетки жёлтого тела яичника в лютеиновую стадию овариально- менструального цикла, а также клетки хориона при наступлении беременности. Прогестерон в эндометрии контролирует секреторную фазу менструального цикла и существенно увеличивает порог возбудимости ГМК миометрия. Стимулируют синтез прогестерона ЛГ и ХГТ. Рецептор прогестинов относится к ядерным факторам транскрипции, генный дефекты рецептора приводит к отсутствию характерных для секреторной фазы менструального цикла изменений эндометрия.
• Релаксины - пептидные гормоны из семейства инсулинов, синтезируется клетками жёлтого тела и цитотрофобластом, при беременности оказывают расслабляющий эффект на ГМК миометрия, а перед родами приводят к размягчению лонного сочленения и шейки матки.
• Ингибины, синтезируемые в яичнике, подавляют синтез и секрецию гипоталамического гонадолиберина и гипофизарного ФСГ.
ПЛАЦЕНТА
Плацента синтезирует множество гормонов и других биологически активных веществ, имеющих важное значение для нормального течения беременности и развития плода.
• Пептидные гормоны (в том числе нейропептиды и рилизинг-гор- моны): хорионический гонадотропин (ХГТ), плацентарный вариант гормона роста, соматомаммотропины хорионические 1 и 2 (плацентарные лактогены), тиреотропин (ТТГ), тиреолиберин (ТТГ-РГ), кортиколиберин (АКТГ-РГ), гонадолиберин, соматолиберин, соматостатин, вещество P, нейротензин, нейропептид Y, пептид, относящийся к АКТГ, гликоделин A (белок, связывающий инсулиноподобные факторы роста), ингибины.
• Стероидные гормоны: прогестерон, эстрон, эстрадиол, эстриол.
ПОЧКИ
Разные клетки почек синтезируют значительное количество веществ, обладающих гормональными эффектами.
• Ренин не является гормоном; этот фермент (протеаза, субстратом которой является ангиотензиноген, см. рис. 18-14) - начальное звено в системе «ренин-ангиотензиноген-ангиотензины» (ренинангиотензиновая система), важнейшего регулятора системного АД. Ренин синтезируется в видоизменённых (эпителиоидньгх) ГМК стенки приносящих артериол почечных телец, входящих в состав околоклубочкового комплекса и секретируется в кровь. Регуляторы
синтеза и секреции ренина: 1. опосредуемая β-адренорецепторами симпатическая иннервация (стимуляция секреции ренина); 2. ангиотензины (по принципу отрицательной обратной связи); 3. рецепторы плотного пятна в составе околоклубочкового комплекса (регистрация содержания NaCl в дистальных канальцах нефрона); 4. барорецепторы в стенке приносящей артериолы почечных телец.
• Кальцитриол (1α,25-дигидроксихолекальциферол) - активная форма витамина D3 - синтезируется в митохондриях проксимальных извитых канальцев, способствует всасыванию кальция и фосфатов в кишечнике, стимулирует остеобласты (ускоряет минерализацию костей). Образование кальцитриола стимулируют ПТГ и гипофосфатемия (пониженное содержание фосфатов в крови), подавляет - гиперфосфатемия (повышенное содержание фосфатов в крови).
• Эритропоэтин - содержащий сиаловую кислоту белок - синтезируется интерстициальными клетками, стимулирует эритропоэз на стадии формирования проэритробластов. Основной стимул для выработки эритропоэтина - гипоксия (снижение рО2 в тканях, в т.ч. зависящее от числа циркулирующих эритроцитов).
• Вазодилататоры - вещества, расслабляющие ГМК стенки кровеносных сосудов, расширяющие их просвет и тем самым снижающие АД. В частности, брадикинин и некоторые простагландины (Пг) синтезируется в интерстициальных клетках мозгового вещества почки.
❖ Брадикинин - нонапептид, образующийся из декапептида каллидина (лизил-брадикинин, кининоген, брадикининоген), который в свою очередь отщепляется от α2-глобулина под действием пептидаз - калликреинов (кининогенины).
❖ Простагландин E2 расслабляет ГМК кровеносных сосудов почки, тем самым уменьшая сосудосуживающие эффекты симпатической стимуляции и ангиотензина II.
СЕРДЦЕ
Натрийуретические факторы (предсердный фактор - атриопептин) синтезируют кардиомиоциты правого предсердия и некоторые нейроны ЦНС. Мишени натриуретических пептидов - клетки почечных телец, собирательных трубочек почки, клубочковой зоны коры надпочечников, ГМК сосудов. Функции натриуретических факторов - контроль объёма внеклеточной жидкости и гомеостаза электролитов (угнетение синтеза и секреции альдостеро-
на, ренина, вазопрессина). Эти пептиды оказывают мощный сосудорасширяющий эффект и снижают АД.
ЖЕЛУДОК И КИШЕЧНИК
В стенке трубчатых органов ЖКТ присутствует огромное количество секретирующих гормоны разнообразных эндокринных клеток (энтероэндокринные клетки). Вместе с вырабатывающими различные нейропептиды клетками собственной нервной системой ЖКТ (энтеральная нервная система) энтероэндокринная система регулирует множество функций пищеварительной системы (рассмотрены в главе 21). Здесь же для примера назовём пептидные гормоны гастрин, секретин и холецистокинин.
• Гастрин стимулирует секрецию HCl париетальными клетками слизистой оболочки желудка.
• Секретин стимулирует выделение бикарбоната и воды из секреторных клеток желёз двенадцатиперстной кишки и поджелудочной железы.
• Холецистокинин стимулирует сокращения жёлчного пузыря и выделение ферментов из поджелудочной железы
РАЗНЫЕ ОРГАНЫ
Клетки различных органов вырабатывают множество химических веществ регуляторного характера, формально не относящихся к гормонам и эндокринной системе (например, Пг, интерфероны, интерлейкины, факторы роста, гемопоэтины, хемокины и др.).
• Эйкозаноиды влияют на сократимость ГМК сосудов и бронхов, изменяют порог болевой чувствительности и участвуют в регуляции многих функций организма (поддержание гемостаза, регуляция тонуса ГМК, секреция желудочного сока, поддержание иммунного статуса и т.д.). Например, в лёгких ПгD2 и лейкотриен С4 - мощные сократительные агонисты для ГМК воздухоносных путей, их эффекты соответственно в 30 и в 1000 раз сильнее гистамина. В то же время ПгЕ2 - вазодилататор, а лейкотриены D4 и Е4 - вазоконстрикторы, они также увеличивают проницаемость стенки кровеносных сосудов.
❖ Пг при физиологических значениях pH плохо проникают через биологические мембраны. Их трансмембранный транспорт осуществляют специальные белки-транспортёры, встроенные в клеточные мембраны.
❖ Рецепторы Пг встроены в плазматическую мембрану клетокмишеней и связаны с G-белками.
• Гистамин - мощный стимулятор секреции соляной кислоты в желудке, важнейший медиатор немедленных аллергических реакций и воспаления, вызывает сокращение ГМК воздухоносных путей и бронхоконстрикцию, но в то же время является сосудорасширяющим агентом для мелких сосудов.
• Интерфероны - гликопротеины, имеющие антивирусную активность; выделяют, по крайней мере, 4 типа интерферонов (α, β, γ, ω).
• Интерлейкины (не менее 31) - цитокины, действующие как факторы роста и дифференцировки лимфоцитов и др. клеток.
• Факторы роста стимулируют рост и дифференцировку, а иногда и трансформацию (озлокачествление) различных клеток. Известно несколько десятков факторов роста: эпидермальный, фибробластов, гепатоцитов, нервов и др.
• Хемокины (несколько десятков) - небольшие секреторные белки, в первую очередь регулирующие перемещения лейкоцитов. Примеры наименований хемокинов: фракталкин, лимфотактин, фактор хемотаксиса моноцитов, интерлейкин-18, эутактин и множество других.
• Колониестимулирующие факторы - белковые факторы, необходимые для выживания, пролиферации и дифференцировки гемопоэтических клеток. Они носят имя клеток, на которые оказывают стимулирующее влияние: колониестимулирующий фактор гранулоцитов (G-CSF), колониестимулирующий фактор гранулоцитов и макрофагов (GM-CSF), колониестимулирующий фактор макрофагов (M-CSF) и колониестимулирующий фактор для многих клеточных типов (интерлейкин-3). Эти факторы вырабатывают макрофаги, T-лимфоциты, эндотелий, фибробласты.