Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА 15. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Поддержание постоянства и оптимальных показателей внутренней среды (гомеостаз) - задача вегетативного отдела нервной системы. Два подотдела этой системы (симпатический и парасимпатический) контролируют АД, моторику и секреторную функцию ЖКТ, температуру тела, выделение слюны, опорожнение мочевого пузыря и множество других висцеральных (вегетативных, внутренностных, «растительных») функций организма. Работа этого системного регулятора висцеральных функций осуществляется рефлекторно (регуляторные контуры висцеральных рефлексов работают по принципу обратной связи) и независимо (автономно) от сознания, но не от деятельности мозга и других отделов нервной системы. И симпатический, и парасимпатический подотделы имеют вегетативные центры и эфферентные пути, заканчивающиеся на эффекторах - секреторных и гладкомышечных клетках (ГМК), в миокарде.
Вегетативная нервная система (подобно соматической нервной системе) функционирует посредством рефлексов. В отличие от соматических рефлексов, рефлексы вегетативной нервной системы - висцеральные рефлексы. Подсознательные сенсорные сигналы возникают в висцеральных рецепторах внутренних органов и через афферентные пути поступают в ЦНС. Они интегрируются на различных уровнях вегетативной нервной системы: ганглиях, мозговом стволе, гипоталамусе, лимбической коре и возвращаются обратно в виде неосознаваемых рефлекторных ответов.
Висцеральные рецепторы и их афферентные пути рассмотрены в главах 8 и 9, а главный эффектор - ГМК - в главе 7. Содержание данной главы составляет функция эфферентных путей симпатического и парасимпатического подотделов вегетативной нервной системы.
Эффекторные пути вегетативной нервной системы представлены на рис. 15-1.
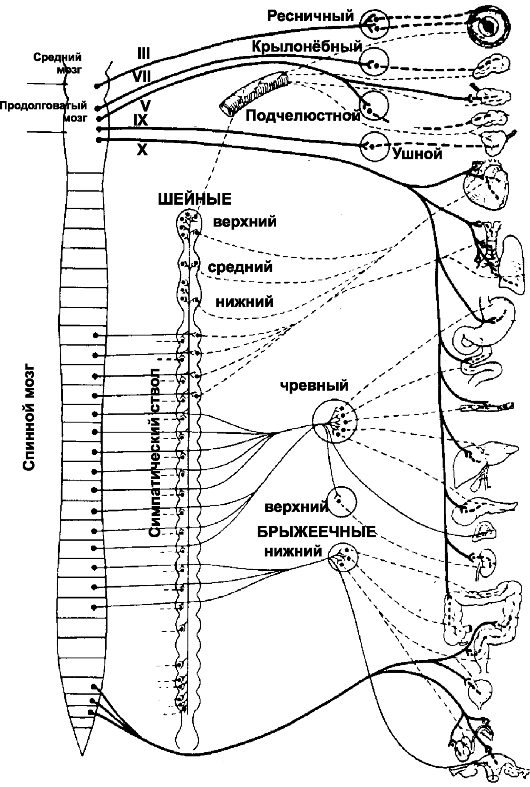 Рис. 15-1. Эфферентные пути вегетативной нервной системы. Слева
направо: стволовая часть мозга и спинной мозг, симпатический ствол,
ганглии вне ствола, эффекторные структуры. Преганглионарные нейроны -
сплошная линия; постганглионарные нейроны - пунктирная линия; толстая
линия - парасимпатические пути; тонкая линия - симпатические пути
Рис. 15-1. Эфферентные пути вегетативной нервной системы. Слева
направо: стволовая часть мозга и спинной мозг, симпатический ствол,
ганглии вне ствола, эффекторные структуры. Преганглионарные нейроны -
сплошная линия; постганглионарные нейроны - пунктирная линия; толстая
линия - парасимпатические пути; тонкая линия - симпатические пути
Симпатический отдел
Каждый симпатический эффекторный путь состоит из двух нейронов - преганглионарного и постганглионарного - и их аксонов в составе нервных пре- и постганглионарных нервных волокон (рис. 15-2). Преганглионары миелинизированы, это сравнительно
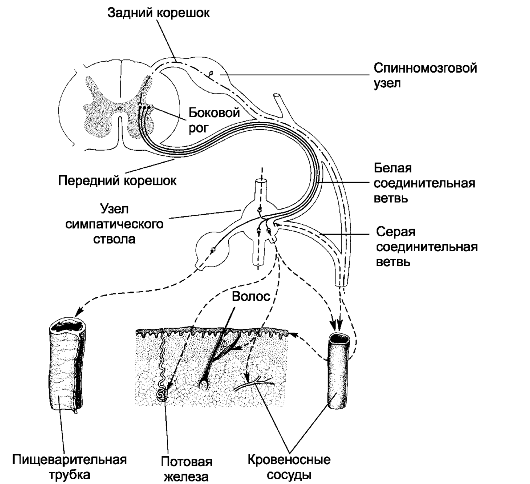 Рис. 15-2. Симпатический отдел вегетативной нервной системы [11].
Перикарионы первого нейрона расположены в боковых столбах спинного
мозга. Их аксоны проходят в составе передних корешков и белой
соединительной ветви и заканчиваются на перикарионах второго нейрона в
паравертебральных ганглиях симпатической цепочки, превертебральных
ганглиях и ганглиях (терминальных), расположенных вблизи иннервируемых
ими органов. Аксоны второго нейрона паравертебральных, превертебральных и
терминальных ганглиев заканчиваются во внутренних органах, коже, стенке
кровеносных сосудов. Аксоны некоторых нейронов паравертебральных
ганглиев проходят через серую соединительную ветвь
Рис. 15-2. Симпатический отдел вегетативной нервной системы [11].
Перикарионы первого нейрона расположены в боковых столбах спинного
мозга. Их аксоны проходят в составе передних корешков и белой
соединительной ветви и заканчиваются на перикарионах второго нейрона в
паравертебральных ганглиях симпатической цепочки, превертебральных
ганглиях и ганглиях (терминальных), расположенных вблизи иннервируемых
ими органов. Аксоны второго нейрона паравертебральных, превертебральных и
терминальных ганглиев заканчиваются во внутренних органах, коже, стенке
кровеносных сосудов. Аксоны некоторых нейронов паравертебральных
ганглиев проходят через серую соединительную ветвь
медленнопроводящие нервные волокна типа B. Постганглионары - безмиелиновые нервные волокна типа C.
• Преганглионарные нейроны. Тела преганглионарных нейронов расположены в боковых рогах спинного мозга. Их аксоны проходят через передние рога спинного мозга в составе спинномозговых нервов. Большинство симпатических волокон оставляет нерв и через белые соединительные ветви проходит в один из ганглиев симпатической цепочки. Далее преганглионарные волокна заканчиваются синапсами: -о с постганглионарными нейронами в том же ганглии, куда они вошли из спинного мозга; -о с нейронами ганглиев симпатической цепочки, расположенных выше и ниже данного узла; - с нейронами, лежащими вне симпатической цепочки в периферических симпатических ганглиях. Некоторые постганглионарные нервные волокна проходят из симпатической цепочки в спинномозговой нерв через серые соединительные ветви и направляются к кровеносным сосудам, потовым железам и мышцам, выпрямляющим волосы.
• Постганглионарные нейроны. Их тела расположены либо в ганглиях симпатической цепочки, либо в периферических симпатических ганглиях (в мозговой части надпочечника постганглионарные нейроны - модифицированные нервные, или хромаффинные клетки). Из этих источников их аксоны - постганглионарные симпатические волокна - направляются к различным органам.
Парасимпатический отдел
• Преганглионарные нейроны. Их тела расположены в среднем и продолговатом мозге, а также в крестцовом отделе спинного мозга (рис. 15-1). Аксоны преганглионарных нейронов (преганглионарные волокна) в составе блуждающих и тазовых нервов без перерыва достигают иннервируемых органов, в которых образуют синапсы с постганглионарными нервными клетками парасимпатического эффекторного пути. Преганглионарные волокна, идущие в составе III, V, VII и IX пар черепных нервов также образуют синапсы с постганглионарными нервными клетками, но постганглионарные нейроны располагаются не в иннервируемых органах, а вне этих органов - в соответствующих парасимпатических ганглиях.
• Постганглионарные нейроны расположены либо в стенке иннервируемых органов (сердце, лёгкие, пищевод, желудок, кишечник, печень, жёлчный пузырь, поджелудочная железа, мочеточники, мочевой пузырь, половые органы), либо в нервных узлах (ресничный, крылонёбный, подчелюстной, ушной). Вне зависи-
мости от местоположения тел постганглионарных нейронов, их аксоны иннервируют соответствующие мышечные и секреторные клетки.
Нейромедиаторы
Нейроны вегетативной нервной системы синтезируют и секретируют различные биологически активные вещества, выступающие в качестве нейромедиаторов и нейромодуляторов. Основные нейромедиаторы - ацетилхолин и норадреналин. Нейроны, синтезирующие ацетилхолин и секретирующие ацетилхолин в терминальных ветвлениях аксона, - холинергические. Нейроны, синтезирующие норадреналин и секретирующие норадреналин в терминальных ветвлениях аксона, - адренергические.
• Холинергические нейроны
О Преганглионары. Все преганглионарные нейроны холинергические (как в симпатическом, так и в парасимпатическом отделах). Следовательно, ацетилхолин обеспечивает синаптическую передачу между пре- и постганглионарными нейронами как в симпатическом, так и в парасимпатическом отделах вегетативной нервной системы.
О Постганглионары. Все (или почти все) постганглионарные парасимпатические нейроны холинергические.
• Адренергические нейроны
О Преганглионары. Преганглионарные адренергические нейроны
не найдены. О Постганглионары
♦ Большинство постганглионарных симпатических нейронов адренергические.
♦ Постганглионарные симпатические нервные волокна к потовым железам, к выпрямляющим мышцам волос (пиломоторы) и некоторым кровеносным сосудам - холинергические.
• Варикозные расширения (см. рис. 7-11). Аксоны постганглионарных вегетативных нейронов образуют многочисленные варикозные расширения - содержащие синаптические пузырьки локальные утолщения. Эти утолщения - места секреции нейромедиатора: адреналина из адренергических волокон, ацетилхолина - из холинергических волокон.
• Адрено- и холинорецепторы. Адреналин и ацетилхолин, выделившиеся из пресинаптических окончаний, взаимодействуют с их рецепторами - адренорецепторами и холинорецепторами соответственно. На рис. 15-3 с учётом нейромедиатора и типов ре-
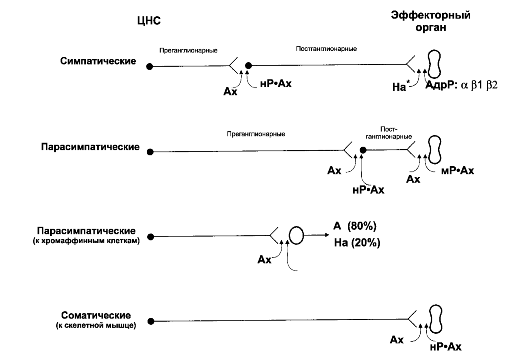 Рис. 15-3. Симпатические и парасимпатические эффекторные пути [5]. Ах -
Рис. 15-3. Симпатические и парасимпатические эффекторные пути [5]. Ах -
ацетилхолин; На - норадреналин; А - адреналин; АдрР - адренорецептор; нР - никотиновый холинорецептор; мР - мускариновый холинорецептор
цепторов представлены эффекторные симпатические и парасимпатические цепочки.
Рецепторы и ответы эффекторных структур
Ответ содержащих рецепторы эффекторных структур (нейронов, ГМК, секреторных клеток, клеток проводящей системы сердца и кардиомиоцитов) определяется типом рецептора. В эффекторных путях вегетативной нервной системы присутствуют 2 типа холинорецепторов (никотиновые и мускариновые) и несколько типов адренорецепторов (α1 ,α2, β1 и β2).
Адренорецепторы
• α1-Адренорецепторы
^ Локализация: ГМК (кроме ГМК бронхов). ^ Эффект: активация.
-Φ- Чувствительность к лигандам: одинаковая к адреналину и норадреналину, в реальных условиях in vivo возбуждает норадреналин.
-ФМеханизм: образование инозитолтрифосфата и увеличение внутриклеточного [Ca2+].
• а2-Адренорецепторы
-ФЛокализация: пресинаптические нервные терминали, ГМК,
жировые клетки. -ФЭффект: зачастую ингибирование.
-ФМеханизм: ингибирование активности аденилатциклазы и уменьшение внутриклеточного [цАМФ].
• β1-Адренорецепторы
-ФЛокализация: стенка сердца. -ФЭффект: активация.
-ФЧувствительность к лигандам: одинаковая к адреналину и норадреналину, чувствительность выше, чем у α-адренорецеп- торов.
-ФМеханизм: увеличение активности аденилатциклазы и внутриклеточного [цАМФ].
• β2-Адренорецепторы
-ФЛокализация: сосудистые ГМК, ГМК бронхов, ЖКТ. -ФЭффект: расслабление ГМК.
-ФЧувствительность к лигандам: адреналин > норадреналин, чувствительность к адреналину выше, чем у α-адренорецепторов.
-ФМеханизм: увеличение активности аденилатциклазы и внутриклеточного [цАМФ].
Холинорецепторы
• Никотиновые холинорецепторы
-ФЛокализация: ганглии вегетативной нервной системы и нервно- скелетномышечный синапс (субъединицы рецепторов разной локализации кодируют разные гены).
-ФЭффект: активация.
-ФЧувствительность к лигандам: ганглиоблокаторы (см. табл. 15-3)
не блокируют нервно-мышечную передачу. -ФМеханизм: никотиновый холинорецептор - ионный канал для
K+ и Na+.
• Мускариновые холинорецепторы
-ФЛокализация: сердце, железы, ГМК (но не сосудистые ГМК).
-ФЭффект: активация ГМК и желёз, подавление в сердце.
-ФЧувствительность к лигандам: атропин блокирует рецепторы.
-Φ- Механизм. Синусно-предсердньй узел: ингибирование активности аденилатциклазы и открытие калиевых каналов; в ГМК: образование инозитолтрифосфата и увеличение внутриклеточного [Ca2+].
Физиологические ответы эффекторов
Поступление сигналов по эффекторным путям вегетативного отдела нервной системы вызывает специфические физиологические ответы иннервируемых структур. Характер ответа полностью определяется типом рецепторов, встроенных в клетки-мишени. Эти ответы рассматриваются в соответствующих главах учебника, в табл. 15-1 и 15-2 приведены некоторые типичные примеры.
Таблица 15-1. Адренергические рецепторы и эффекты норадреналина
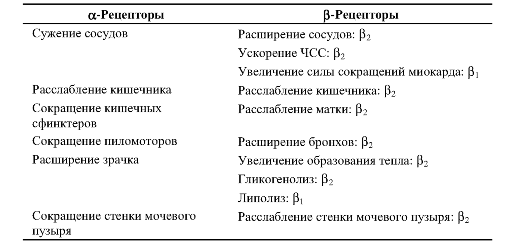 Таблица 15-2. Физиологические ответы эффекторов на симпатическую и парасимпатическую стимуляцию [5]
Таблица 15-2. Физиологические ответы эффекторов на симпатическую и парасимпатическую стимуляцию [5]
 Окончание табл. 15-2.
Окончание табл. 15-2.
 Агонисты и антагонисты
Агонисты и антагонисты
В клинической практике широко применяют различные ЛС - агонисты и антагонисты адрено- и холинорецепторов разных типов. Агонисты, взаимодействуя с рецептором, воспроизводят эффект естественного рецепторного лиганда (например, норадреналина для адренорецепторов). Антагонисты, взаимодействуя с рецептором, блокируют эффект естественного рецепторного лиганда. В табл. 15-2 перечислены некоторые лиганды адрено- и холинорецепторов, обладающих свойствами агонистов и антагонистов.
Эффекты гуморальных адреналина и норадреналина
В эффекторных путях симпатического отдела вегетативной нервной системы нейромедиатором является норадреналин. В то же время и норадреналин, и главным образом адреналин поступают в кровь из хромаффинных клеток надпочечниково-адреналовой системы. Другими словами, эти химические соединения могут рассматриваться как гормоны (см. подробнее в главе 18). Адреналин и норадреналин, выделяющиеся в кровь из хромаффинных клеток (в том числе из мозговой части надпочечников), вызывают практи-
Таблица 15-3. Агонисты и антагонисты адрено- и холинорецепторов [5]
 Примечание: * В ганглиях, но не в нервно-скелетномышечном синапсе
Примечание: * В ганглиях, но не в нервно-скелетномышечном синапсе
чески те же эффекты, что и стимуляция симпатических эффекторов, но действие гуморальных адреналина и норадреналина продолжается в 5-10 раз дольше, чем эффект норадреналина в качестве нейромедиатора, поскольку гормоны удаляются из крови в течение долгих 1-3 мин. Адреналин вызывает почти те же самые эффекты, что и норадреналин, но имеются и некоторые отличия.
• Адреналин более интенсивно стимулирует работу сердца, потому что он лучше связывается с β-адренорецепторами.
• Адреналин (в отличие от норадреналина) вызывает некоторое сужение кровеносных сосудов в скелетных мышцах. Это важный момент, потому что сосуды мышц - существенно большой сегмент сосудов тела, и норадреналин, вызывая вазоконстрикцию, увеличивает периферическое сосудистое сопротивление, в результате чего повышается АД. Адреналин повышает АД в меньшей степени, но зато в большей степени увеличивает производительность сердца.
• Адреналин вызывает метаболический эффект в 5-10 раз больший, чем норадреналин. Выделяясь из надпочечников, адреналин ускоряет метаболизм и тем самым усиливает активность и возбудимость организма.
• Выброс адреналина и норадреналина из надпочечников совпадает с генерализованной активацией симпатической нервной системы. Следовательно, одновременно происходит стимуляция эффекторных органов двумя путями: непосредственно симпатическими
нервами и опосредованно - гормонами мозговой части надпочечников. Такая двойная адренергическая стимуляция поддерживает и во многих случаях заменяет друг друга.
Понятие о тонусе
Симпатическая и парасимпатическая системы постоянно активны, и величину этой исходной активности рассматривают как симпатический тонус или как парасимпатический тонус соответственно.
• Тонус оценивают по способности одной из систем увеличивать или уменьшать активность стимулируемого органа. Так, симпатический тонус поддерживает артериолы суженными на половину от их максимального диаметра. В ответ на увеличение симпатической стимуляции выше нормы артериолы уменьшат диаметр просвета ещё больше (вазоконстрикция). Напротив, уменьшение симпатической стимуляции ниже нормы вызовет расслабление сосудов - вазодилатацию. Другими словами, если бы постоянный базовый тонус отсутствовал, симпатическая система могла бы вызывать только вазоконстрикцию.
• Другой пример - парасимпатический тонус ЖКТ. Перерезка блуждающего нерва может вызвать длительную атонию желудка и кишечника, нарушения перистальтики, появление в последующем выраженных запоров.
• Поддержание эффекторов в тоническом состоянии обеспечивается редкими (порядка 1 импульса на протяжении в несколько секунд) разрядами симпатических или парасимпатических нервов. Для полной активации необходима частота разрядов около 10- 20 импульсов в секунду.
• Базальная (спонтанная) секреция адреналина и норадреналина в состоянии покоя составляет около 0,2 мкг/кг/мин адреналина и около 0,05 мкг/кг/мин норадреналина. Такого количества достаточно, чтобы удерживать АД в границах нормы.
Центральная регуляция висцеральных функций
Регуляция висцеральных функций осуществляется в ЦНС по иерархическому принципу. Простые рефлексы (например, сокращение полного мочевого пузыря или опорожнение прямой кишки) замыкаются на уровне спинного мозга. Более сложные рефлексы (например, рефлексы, регулирующие дыхание и АД; зрачковая реакция на свет и аккомодация глаза) интегрированы на уровне продолговатого и среднего мозга. Комплекс механизмов, поддерживающих химическое постоянство и температуру внутренней среды, интегрирован в гипоталамусе.
• Контроль дыхания, частоты сердечных сокращений, артериального давления. Центры продолговатого мозга, осуществляющие рефлекторный контроль кровообращения, дыхания и сердечной деятельности, получили название жизненно важных центров, потому что их повреждение фатально. Афферентные волокна к этим центрам поступают из самых различных участков тела от специализированных рецепторов внутренних органов. Специализированные рецепторы включают в себя не только рецепторы каротидного синуса и аортальных телец, но также рецепторные клетки, которые располагаются в самом продолговатом мозге. Детали этих рефлексов обсуждаются в разделах, посвящённых регуляции дыхания и кровообращения.
• Другие рефлексы продолговатого мозга. Глотание, кашель, чихание, тошнота и рвота являются рефлекторными ответами, интегрированными в продолговатом мозге. Глотательный рефлекс начинается произвольно с проталкивания содержимого ротовой полости к задней стенке глотки. Кашель начинается с раздражения слизистой оболочки трахеи и внелёгочных бронхов. Голосовая щель закрывается, и сильное сокращение дыхательных мышц повышает внутрилёгочное давление; голосовая щель неожиданно открывается, вызывая взрывоподобный выброс воздуха. Чихание имеет подобный же механизм в ответ на раздражение рецепторов эпителия носовой полости.
• Рвота является примером висцерального рефлекса, осуществляемого продолговатым мозгом, с включением координированных соматических и висцеральных компонентов. Рвота начинается со слюноотделения и ощущения тошноты. Обратная перистальтика верхнего отдела кишечника передвигает его содержимое в желудок. Голосовая щель закрывается, предотвращая аспирацию рвотных масс в трахею. Дыхание задерживается на середине вдоха. Мышцы брюшной стенки сокращаются, и в силу того, что грудная клетка удерживается в фиксированном положении, это сокращение повышает внутрибрюшное давление. Нижний сфинктер пищевода и пищевод расслабляются, и содержимое желудка удаляется наружу.
-Ф- Центр рвоты, находящийся в ретикулярной формации продолговатого мозга (рис. 15-4) состоит из нескольких групп нейронов, контролирующих различные компоненты акта рвоты.
-Ф- Пусковые сигналы рвоты. Раздражение слизистой оболочки верхних отделов ЖКТ вызывает рвоту. Импульсы поступают от слизистой оболочки в продолговатый мозг по висцеральным афферентным путям в составе симпатических нервов и блуж-
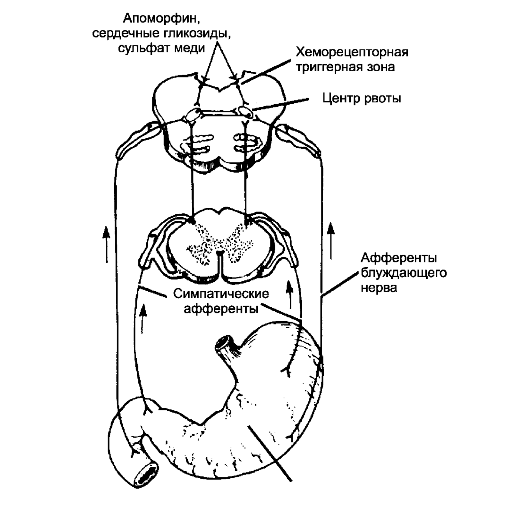 Рис. 15-4. Рефлекторный и хеморецепторный механизмы рвотного рефлекса
Рис. 15-4. Рефлекторный и хеморецепторный механизмы рвотного рефлекса
дающего нерва. Другие пусковые сигналы достигают центра рвоты из среднего мозга и лимбической системы, поскольку рвота возникает, как известно, и в ответ на эмоциональные разряды.
- Хеморецепторные клетки, расположенные в хеморецепторной триггерной зоне продолговатого мозга (в боковой стенке чет- вёртого желудочка в области ромбовидной ямки), обладают высокой проницаемостью к различным веществам, циркулирующим в крови (рис. 15-4). Некоторые химические вещества, поступившие в кровь, раздражают эти хеморецептивные клетки и вызывают рвоту. Повреждение этой зоны существенно не сказывается на рвотных ответах на раздражение слизистой оболочки ЖКТ и отрицательных эмоциях, но прерывает рвоту, возникающую при инъекции апоморфина или других рвотных средств.