Нормальная физиология : Учебник. - Р.С. Орлов, А.Д. Ноздрачёв, 2009. - 688 с
|
|
|
|
ГЛАВА13. ДВИГАТЕЛЬНЫЕ ФУНКЦИИ СПИННОГО МОЗГА
Спинной мозг покрыт мягкой, паутинной и твёрдой мозговой оболочкой, омывается спинномозговой жидкостью и состоит из двух симметричных половин, соединённых узкой перемычкой, содержащей центральный канал. На поперечном разрезе легко различимы лежащее снаружи белое вещество и расположенная глубже тём- ная масса - серое вещество. В спинной мозг входят проходящие в составе задних корешков центральные отростки чувствительных нейронов спинномозговых узлов. В передних рогах находятся мотонейроны, аксоны которых через передние корешки иннервируют скелетные мышцы. Передний и задний корешки соединяются и образуют 31 пару спинномозговых нервов.
ОРГАНИЗАЦИЯ ДВИГАТЕЛЬНЫХ ФУНКЦИЙ
• Спинной мозг является интегративной областью для осуществления рефлексов спинного мозга. Каждый сегмент спинного мозга (на уровне каждого нерва) содержит несколько миллионов нервных клеток, в том числе мотонейроны и вставочные нейроны. Мотонейроны. Различают крупные, или α-мотонейроны, и мелкие, или γ-мотонейроны.
α-Мотонейроны входят в состав медиальных и латеральных ядер передних рогов. Это наиболее крупные клетки спинного мозга. Их аксоны в составе двигательных нервных волокон типа Aa образуют нервно-мышечные синапсы с экстрафузальными мышечными волокнами (МВ) скелетной мышцы (см. рис. 6-2) и участвуют в формировании нейромоторных единиц (см. главу 7). Аксоны α-мотонейронов посылают коллатерали к вставочным нейронам Реншоу, образующим тормозные синапсы с α-мотонейронами (см. рис. 6-7, 2).
γ-Мотонейроны располагаются вместе с α-мотонейронами в передних рогах спинного мозга. Они значительно меньше α-мотонейронов и передают ПД по нервных волокнам Ag, осуществляя двигательную иннервацию интрафузальных МВ в составе мышечных веретён (см. ниже).
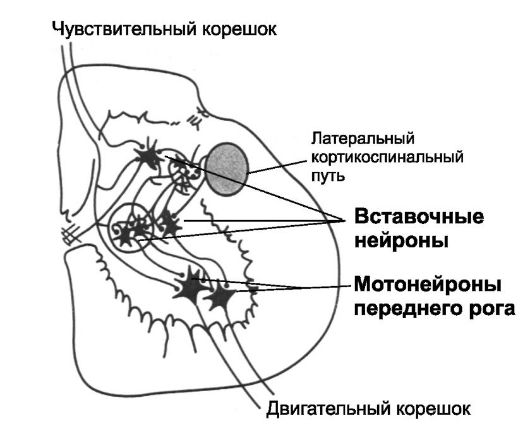 Рис. 13-1. Взаимодействие афферентных, вставочных и моторных нейронов в спинном мозге
Рис. 13-1. Взаимодействие афферентных, вставочных и моторных нейронов в спинном мозге
• Вставочные нейроны получают информацию от одних нейронов и передают её другим. Аксоны вставочных нейронов участвуют также в образовании проводящих путей. Классические примеры вставочных нейронов - клетки Реншоу и энкефалинергические нервные клетки, модулирующие передачу болевых импульсов (см. рис. 9-6). Количество вставочных нейронов в 30 раз превышает количество мотонейронов. Вставочные нейроны небольшого размера, их возбудимость высокая, часто они
спонтанно активны, генерируют импульсы частотой до 1500 Гц, имеют много связей друг с другом, часть из них синаптически контактирует с мотонейронами (рис. 13-1). Особые свойства вставочных нейронов в организации нейронных пулов: дивергенция, конвергенция, повторные разряды, латеральное торможение, торможение через клетки Реншоу имеют непосредственное отношение к моторным функциям спинного мозга. Только некоторые сигналы, поступающие из спинномозговых нервов, или сигналы из мозга сразу передаются к мотонейронам, практически все сигналы попадают к вставочным нейронам и только после того, как они интегрируются в пулах интернейронов с сигналами из других источников, они окончательно конвергируют на мотонейроны.
• Чувствительные нейроны расположены в спинномозговых узлах, их периферические отростки образуют чувствительные нервные окончания (в том числе в мышцах, сухожилиях и капсуле суставов), а центральные отростки по задним корешкам входят в спинной мозг (рис. 13-1). После вхождения в спинной мозг сигналы проходят в двух направлениях. Часть центральных отростков заканчивается почти сразу в сером веществе спинного мозга и вызывает локальные, сегментарные спинномозговые рефлексы и другие местные эффекты. Другие отростки передают сигналы к более высоким уровням спинного мозга, мозгового ствола и коре больших полушарий (см. главу 9).
Проприоцептивная иннервация скелетных мышц
Функцию скелетных мышц контролирует не только импульсация α-мотонейронов, но и сенсорная обратная связь от каждой мышцы к спинному мозгу. Поступающая в спинной мозг сенсорная информация характеризует длину мышцы, её напряжение, темп изменений её длины или напряжения. Эта информация поступает от нервно-мышечных веретён, расположенных в мышечном брюшке, от сухожильных органов Гольджи и чувствительных нервных окончаний в капсуле суставов.
• Нервно-мышечные веретёна (рис. 13-2А) имеют длину от 3 до 10 мм. Их количество в различных мышцах значительно варьирует, но они присутствуют практически во всех мышцах. Основные элементы мышечного веретена - интрафузальные МВ, нервные волокна и капсула.
• Сухожильные органы Гольджи (рис. 13-2Б) расположены в концевой части сухожилия на границе с мышцей, а также в связках капсулы суставов. Рецептор имеет веретеновидную форму и ок- ружён плотной капсулой. Терминали афферентных миелиновых волокон ветвятся среди пучков спиралевидных коллагеновых волокон, расположенных в заполненном жидкостью пространстве.
• Чувствительные нервные окончания в капсуле суставов - важный элемент проприоцептивной системы организма.
ФИЗИОЛОГИЯ ДВИГАТЕЛЬНЫХ ФУНКЦИЙ
Мышечные веретёна
Рецепторная часть нервно-мышечного веретена - его центральная область (13-3А1 и 13-3А2), где отсутствует сократительный аппарат МВ. Как первичные, так и вторичные чувствительные нервные окончания этой области возбуждаются при растяжении веретена, т.е. либо при удлинении всей мышцы, либо при сокращении концевых участков интрафузального МВ (за счёт поступающей по γ-эфферентам импульсации).
В каждый момент времени различная степень длины мышцы и характер её изменения сказываются на функции мышечных вере- тён, что видно при регистрации статических и динамических ответов (рис. 13-4).
Статические и динамические ответы
Статические ответы. Медленное растяжение мышечного веретена вызывает появление импульсов, частота которых пропорциональ-
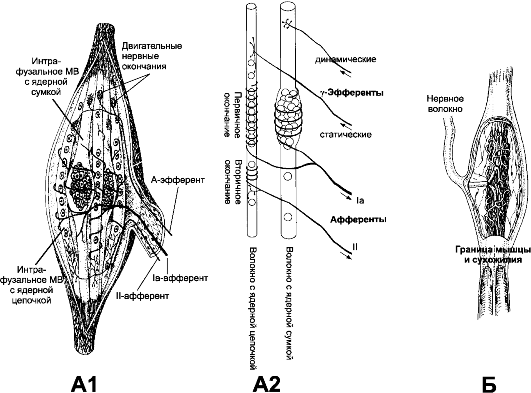 Рис. 13-2. А - нервно-мышечное веретено. А1. Интрафузальные МВ с компактным скоплением ядер - МВ с ядерной сумкой, в интрафузальных МВ с ядерной цепочкой ядра распределены по длине волокна более равномерно. А2. К
веретену подходят афферентные и эфферентные нервные волокна.
Аннулоспиральные (первичные) сенсорные окончания образованы
безмиелиновыми терминалями афферентных Ια-волокон в экваториальной зоне
обоих типов интрафузальных МВ. Ближе к концам интрафузальных МВ (чаще МВ
с ядерной цепочкой) расположены терминали тонких афферентных II-волокон
- вторичные окончания. Эфферентные Ау-волокна образуют нервно-мышечные
синапсы с интрафузальными МВ в концевой их части. В центральной части
каждого интрафузального МВ отсутствует сократительный аппарат, т.е.
центральная часть МВ не сокращается, когда сокращаются концы МВ. Б - сухожильный орган Гольджи. Рецептор
окру- жён капсулой, через которую проходит миелиновое нервное волокно,
образующее терминальное сплетение среди коллагеновых волокон
Рис. 13-2. А - нервно-мышечное веретено. А1. Интрафузальные МВ с компактным скоплением ядер - МВ с ядерной сумкой, в интрафузальных МВ с ядерной цепочкой ядра распределены по длине волокна более равномерно. А2. К
веретену подходят афферентные и эфферентные нервные волокна.
Аннулоспиральные (первичные) сенсорные окончания образованы
безмиелиновыми терминалями афферентных Ια-волокон в экваториальной зоне
обоих типов интрафузальных МВ. Ближе к концам интрафузальных МВ (чаще МВ
с ядерной цепочкой) расположены терминали тонких афферентных II-волокон
- вторичные окончания. Эфферентные Ау-волокна образуют нервно-мышечные
синапсы с интрафузальными МВ в концевой их части. В центральной части
каждого интрафузального МВ отсутствует сократительный аппарат, т.е.
центральная часть МВ не сокращается, когда сокращаются концы МВ. Б - сухожильный орган Гольджи. Рецептор
окру- жён капсулой, через которую проходит миелиновое нервное волокно,
образующее терминальное сплетение среди коллагеновых волокон
на степени растяжения. Если рецептор остаётся растянутым, то импульсация продолжается несколько минут. Этот эффект - статический ответ - связан с активацией интрафузальных МВ типа ядерной цепочки (рис. 13-3А).
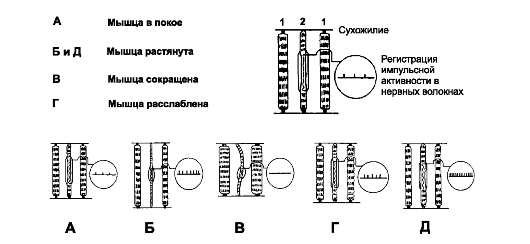 Рис. 13-3. Имульсация от нервно-мышечных веретён в
зависимости от различных состояний мышцы и активации
гамма-мотонейронов. 1 - экстрафузальное МВ; 2 - интрафузальное МВ и
нервно-мышечное веретено. В состояниях Г и Д увеличена импульсация
гамма-эфферентов
Рис. 13-3. Имульсация от нервно-мышечных веретён в
зависимости от различных состояний мышцы и активации
гамма-мотонейронов. 1 - экстрафузальное МВ; 2 - интрафузальное МВ и
нервно-мышечное веретено. В состояниях Г и Д увеличена импульсация
гамма-эфферентов
Динамические ответы
• Быстрое растяжение мышечного веретена стимулирует первичные окончания, вызывая ответ, намного превышающий реакцию на стимулы, вызывающие статический ответ (рис. 13-3Б). Эта стимуляция первичного окончания - динамический ответ - свидетельствует о том, что первичное окончание отвечает на быстрое изменение длины мышечного веретена. Достаточно в доли секунды и на доли микрометра изменить длину мышечного веретена, чтобы в Ia-волокнах возник разряд импульсов. В момент прекращения изменения длины мышечного веретена частота импульсации возвращается к уровню, но меньшему, чем при статическом растяжении.
• Укорочение мышечного веретена мгновенно уменьшает частоту импульсов в первичном окончании; если вскоре мышечное веретено вновь укорачивается, то через доли секунды меняется и импульсация в I α-волокнах (рис. 13-4В).
• Таким образом, первичные окончания посылают очень сильные положительные или отрицательные сигналы в спинной мозг с оценкой любых изменений длины мышечного веретена.
γ-Контроль статических и динамических ответов. γ-Моторные аксоны, идущие к мышечным веретёнам, подразделяются на два типа - γ-динамичные и γ-статичные (рис. 13-2А2). Разряды γ-динамичных аксонов возбуждают интрафузальные МВ с ядер-
ной сумкой, вызывая значительное увеличение динамического ответа, в то же время статический ответ едва заметен (рис. 13-3Г). Напротив, стимуляция γ-статичных аксонов, которые возбуждают интрафузальные МВ с ядерной цепочкой, усиливает статический ответ (рис. 13-3Д), оказывая слабое влияние на динамический ответ.
Рефлексы растяжения
Мышечные веретёна и сухожильные органы Гольджи участвуют в реализации рефлексов на растяжение, возникающих в ответ на резкое растяжение мышцы. При этом возбуждение мышечных рецепторов вызывает рефлекторное сокращение как этой, так и мышц-синерги- стов. На рис. 13-4 показаны рефлекторные дуги рефлексов растяжения, как моносинаптического (I), так и полисинаптического (II).
• Моносинаптическая дуга. Ia-проприоцептивные нервные волокна, отходящие от мышечного веретена, входят в задний корешок спинного мозга и сразу направляются в передний рог, где и образуют синапсы с a-мотонейронами, посьлающими сигналы к мышце.
• Полисинаптическая дуга дополнительно включает вставочный нейрон. На рис. 13-5(II) представлена дуга тормозного рефлекса, возникающего при растяжении сухожильных рецепторов Гольджи.
Динамический и статический рефлексы растяжения. Различают динамический и статический компоненты рефлекса растяжения.
• Динамический рефлекс растяжения осуществляется при неожиданном быстром удлинении мышцы, что приводит к столь же быстрому сокращению. Очевидно, что функция рефлекса направлена против неожиданных изменений в длине мышцы, поскольку мышца сокращается, преодолевая растяжение.
• Статический рефлекс растяжения. Динамичес-
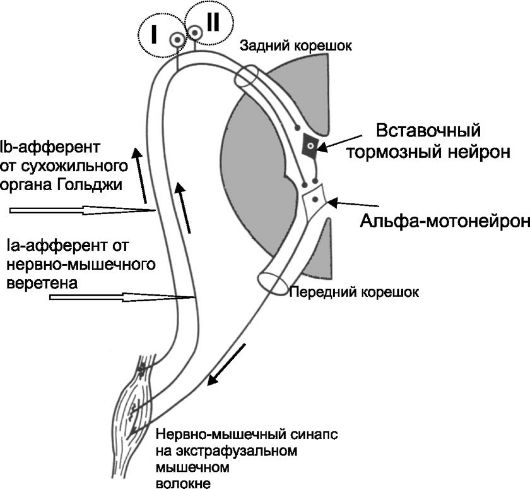 Рис. 13-4. Рефлексы растяжения моносинаптический (I, от мышечных веретён, приводит к сокращению той же мышцы) и полисинаптический (II)
Рис. 13-4. Рефлексы растяжения моносинаптический (I, от мышечных веретён, приводит к сокращению той же мышцы) и полисинаптический (II)
кий рефлекс растяжения совершается в доли секунды. После того как мышца растянута до её новой длины, следует слабый статический рефлекс растяжения. Его важность состоит в том, что он продолжается в течение всего времени, пока длина мышц изменена. Следовательно, функция статического рефлекса растяжения также направлена против сил, вызывающих превышение исходной длины мышцы. Сигналы, поступающие к скелетным мышцам от спинного мозга, как правило, носят дискретный характер (например, увеличить интенсивность в течение нескольких миллисекунд, изменить уровень интенсивности, уменьшить силу сокращения и т.д.). То обстоятельство, что в норме даже самые быстрые движения осуществляются плавно, как раз и связано с наличием динамического и статического компонентов рефлексов растяжения. Другими словами, динамический и статический компоненты рефлекса растяжения - регуляторы плавности сокращений.
Клиническое значение рефлексов растяжения
В повседневной клинической практике рефлексы растяжения используют для определения состояния уровня возбуждения, или «тонуса» ЦНС. На рис. 13-5 приведены некоторые рефлексы спинного мозга. Быстрое растяжение мышцы механическим ударом по её сухожилию вызывает сокращение всей мышцы. Например, легкий удар по связке надколенника (рис. 13-5А) вызывает динамический рефлекс растяжения - сокращение мышц бедра и разгибание голени.
Дуга моносинаптического коленного рефлекса такова: мышечные веретёна четырехглавой мышцы бедра - спинномозговой узел - задние корешки - задние рога III поясничного сегмента - α-мотонейроны передних рогов того же сегмента - экстрафузальные МВ четырехглавой мышцы бедра. Подобные рефлексы могут быть получены почти у всех мышц тела. Усиление рефлексов растяжения расценивается как повышение облегчающих, активирующих влияний верхних уровней ЦНС. Напротив, депрессия облегчающих влияний со стороны ЦНС ослабляет рефлексы растяжения. Эти рефлексы применяют для оценки наличия или отсутствия спастичности мышц. Так, при инсульте или опухоли мозга наблюдается значительное усиление рефлексов растяжения на поражённой стороне тела.
Функции рецепторов Гольджи
Сухожильный орган Гольджи (см. рис. 13-2Б) регистрирует напряжение, развиваемое МВ в области прикрепления к сухожилию. Таким образом, мышечные веретена определяют длину мышцы и изменения её длины, а сухожильные органы - мышечное напряжение. Подобно мышечным веретёнам, сухожильный орган отвечает на
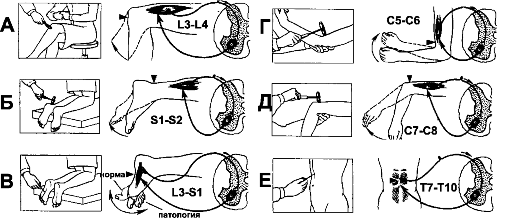 Рис. 13-5. Рефлексы спинного мозга. А - коленный рефлекс. Б - ахиллов рефлекс. В - подошвенный рефлекс (рефлекс Бабинского). Г - сгибательный рефлекс предплечья. Д - разгибательный рефлекс предплечья. Е - брюшной рефлекс
Рис. 13-5. Рефлексы спинного мозга. А - коленный рефлекс. Б - ахиллов рефлекс. В - подошвенный рефлекс (рефлекс Бабинского). Г - сгибательный рефлекс предплечья. Д - разгибательный рефлекс предплечья. Е - брюшной рефлекс
быстрое увеличение мышечного напряжения динамической реакцией, а на медленные изменения напряжения - статической реакцией. Сигналы от сухожильного органа идут по миелиновым и быстропроводящим волокнам Ib. Сигналы поступают в задний рог соответствующего сегмента спинного мозга (рис. 13-4,II) и полисинаптически (через тормозный вставочный нейрон) переключаются на α-мотонейроны. Кроме того, информация от сухожильных органов передаётся по спинномозжечковому тракту, а также по другим афферентным путям, идущим к коре больших полушарий.
Аутогенное торможение. Так как сухожильный орган Гольджи (в отличие от мышечных веретён) связан с экстрафузальными МВ, он реагирует как на растяжение, так и на сокращение мышцы. Сильное быстрое растяжение вызывает интенсивное сокращение. В тот момент, когда напряжение, развиваемое мышцей, становится достаточно высоким, сокращение мышцы внезапно прекращается, и она расслабляется. Это расслабление в ответ на сильное растяжение - возвратный рефлекс растяжения, или аутогенное торможение. Такого же рода тормозное влияние рецептор Гольджи оказывает на мышцу в момент сильного сокращения.
Таким образом, орган Гольджи функционирует как преобразователь в цепи обратной связи, регулируя силу мышечного сокращения.
Полисинаптические рефлексы
• Сгибательный рефлекс возникает в ответ на сильные раздражения болевых рецепторов конечности и заключается в сокращении сги-
бательных мышц с одновременным расслаблением разгибательных мышц, чтобы удалить конечность от источника раздражения (рис. 13-6). Болевое раздражение, наносимое на какой-либо участок тела, также вызывает реакцию ухода от раздражения и носит более общее название - рефлекс отдёргивания. На рис. 13-6А показаны пути осуществления сгибательного рефлекса в ответ на болевой стимул, приложенный к кисти. Импульсы от болевого раздражения кисти поступают в пул интернейронов, где взаимодействуют с большим числом вставочных нейронов, активируя несколько нейронных цепей до того, как сигнал достигнет мотонейронов. Необходима активация минимально трёх нейронных цепей: О дивергенция импульсов для вовлечения различных мышц в рефлекс отдергивания О торможение антагонистических мышц - реципрокное торможение О следовые разряды после действия стимула. Перекрестный разгибательный рефлекс. Возникновение болевого сгибательного рефлекса одной конечности через 200-500 мс вызывает разгибание конечности на противоположной стороне. Это перекрестный разгибательный рефлекс. Разгибание противоположной конечности направлено на оттягивание тела от источника боли, вызвавшего рефлекс от-
дергивания. На рис. 13- 6Б показаны пути реализации перекрестного разгибательного рефлекса. Из нейронного пула сгибательного рефлекса сигналы поступают на противоположную сторону спинного мозга и интегрируются здесь со вставочными нейронами для осуществления реципрокного разгибательного рефлекса. Реципрокные взаимоотношения - те случаи, когда возбуждение одной группы мышц тормозит другую. Например, рефлекс растяжения одной мышцы часто одновре-
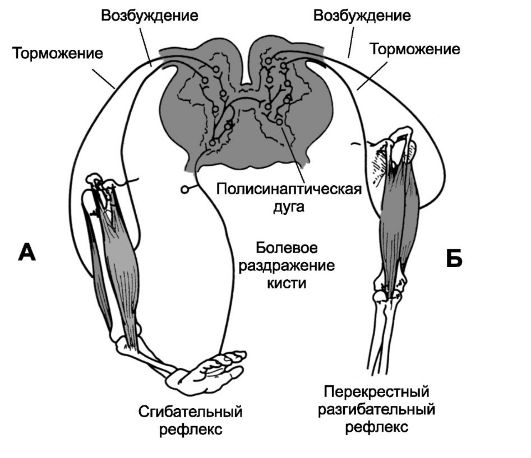 Рис. 13-6. Реципрокное торможение. А -
Рис. 13-6. Реципрокное торможение. А -
сгибательный рефлекс. Б - перекрестный разгибательный рефлекс
• менно тормозит антагонистические мышцы. Это явление - феномен реципрокного торможения, а иннервация, осуществляющая эти взаимоотношения, называется реципрокной иннервацией. Подобные же отношения существуют между мускулатурой двух сторон тела. Постуральные рефлексы и рефлексы движения. Постуральные (позотонические, статические) рефлексы обеспечивают поддержание в пространстве определённого положения всего тела или его части (например, конечности). Так, давление на подушечки стопы спинального животного вызывает реакцию вытягивания конечности, направленного против давления. Это положительная поддерживающая реакция. Если спинальных животных (с перерезкой шейного отдела спинного мозга ниже уровня С4) укладывать на бок, они производят ряд некоординированных движений для восстановления положения стоя. Указанные явления получили название спинномозговой вращательный рефлекс. Спинальные животные совершают ритмические шагательные движения. Это свидетельствует о существовании на уровне спинного мозга закреплённых нейронных пулов, осуществляющих рефлекторный сложнокоординированный акт ходьбы.
Вегетативные рефлексы спинного мозга
В спинном мозге располагаются двигательные нейроны вегетативной нервной системы. На уровне сегментов спинного мозга возникает множество местных вегетативных рефлексов. К ним относятся: О расширение сосудов кожи при локальном нагреве; О потоотделение при нагреве участка кожи; О кишечно-кишечные рефлексы, контролирующие функцию кишечника; О перитонеоинтестинальные рефлексы, тормозящие моторику ЖКТ; О эвакуационные (из мочевого пузыря и толстой кишки) рефлексы.
Масс-рефлекс. В некоторых случаях происходит неожиданная активация большого числа сегментов спинного мозга. Причинами, вызывающими это явление, служат либо сильное болевое раздражение кожи, либо чрезмерное переполнение жидкостью полых органов (например, перерастяжение мочевого пузыря или кишки). В результате возникает масс-рефлекс. Масс-рефлекс проявляется: О спазмом сгибательных мышц тела; О полной эвакуацией кишечника и мочевого пузыря; О подъёмом АД до высоких значений (систолическое давление иногда превышает 200 мм рт.ст.); О профузным потоотделением с больших участков тела. Рефлекс продолжается несколько минут и заканчивается полным восстановлением функций. В основе механизма масс-рефлекса лежит возникновение реверберирующих цепей возбуждения нейронов значительного числа сегментов спинного мозга.