Биохимия: учебник для вузов/ под ред. Е.С.Северина - 5-е изд., - 2009. - 768 с.
|
|
|
|
РАЗДЕЛ 15 БИОХИМИЯ МЕЖКЛЕТОЧНОГО МАТРИКСА
У многоклеточных организмов большинство клеток окружено внеили межклеточным мат-риксом. Межклеточный матрикс - сложный комплекс связанных между собой макромолекул. Эти макромолекулы (белки и гетерополиса-хариды), как правило, секретируются самими клетками, а в межклеточном матриксе из них строится упорядоченная сеть. Межклеточный матрикс, окружающий клетки, влияет на их прикрепление, развитие, пролиферацию, организацию и метаболизм.
Межклеточный матрикс вместе с клетками разного типа, которые в нём находятся (фиброб-ласты, хондро- и остеобласты, тучные клетки и макрофаги), часто называют соединительной тканью.
Межклеточный матрикс выполняет в организме самые разнообразные функции:
• образует каркас органов и тканей;
• является универсальным «биологическим» клеем;
• участвует в регуляции водно-солевого обмена;
• образует высокоспециализированные структуры (кости, зубы, хрящи, сухожилия, базальные мембраны).
Основные компоненты межклеточного мат-рикса - структурные белки коллаген и эластин, гликозаминогликаны, протеогликаны, а также неколлагеновые структурные белки (фибронек-тин, ламинин, тенасцин, остеонектин и др.).
I. КОЛЛАГЕН
Коллаген - основной структурный белок межклеточного матрикса. Он составляет от 25 до 33% общего количества белка в организме, т.е. ~6% массы тела. Название «коллаген» объединяет семейство близкородственных фибриллярных белков, которые являются основным белковым элементом кожи, костей, сухожилий, хряща, кровеносных сосудов, зубов. В разных тканях преобладают разные типы коллагена, а это, в свою очередь, определяется той ролью, которую
коллаген играет в конкретном органе или ткани. Например, в пластинчатой костной ткани, из которой построено большинство плоских и трубчатых костей скелета, коллагеновые волокна имеют строго ориентированное направление: продольное - в центральной части пластинок, поперечное и под углом - в периферической. Это способствует тому, что даже при расслоении пластинок фибриллы одной пластинки могут продолжаться в соседние, создавая таким образом единую волокнистую структуру кости. Поперечно ориентированные коллагеновые волокна могут вплетаться в промежуточные слои между костными пластинками, благодаря чему достигается прочность костной ткани. В сухожилиях коллаген образует плотные параллельные волокна, которые дают возможность этим структурам выдерживать большие механические нагрузки. В хрящевом матриксе коллаген образует фибриллярную сеть, которая придаёт хрящу прочность, а в роговице глаза коллаген участвует в образовании гексагональных решёток десцеметовых мембран, что обеспечивает прозрачность роговицы, а также участие этих структур в преломлении световых лучей. В дерме фибриллы коллагена ориентированы таким образом, что формируют сеть, особенно хорошо развитую в участках кожи, которые испытывают сильное давление (кожа подошв, локтей, ладоней), а в заживающей ране они агрегированы весьма хаотично. Аминокислотный состав и конформация коллагена описаны в подразделе «Фибриллярные белки» раздела 1.
Здесь будут разобраны синтез и созревание коллагена, структуры, которые он образует, и их функции, а также заболевания, связанные с нарушением этих процессов.
Полиморфизм коллагена
Коллаген - ярко выраженный полиморфный белок. В настоящее время известно 19 типов коллагена, которые отличаются друг от друга по первичной структуре пептидных цепей, функциям и локализации в организме. Вариантов а-цепей,
образующих тройную спираль, гораздо больше 19 (около 30). Для обозначения каждого вида коллагена пользуются определённой формулой, в которой тип коллагена записывается римской цифрой в скобках, а для обозначения а-цепей используют арабские цифры: например коллагены II и III типа образованы идентичными а-цепями, их формулы, соответственно [α1(II)]3 и [α1 (III)]3; коллагены I и IV типов являются гетеротримерами и образуются обычно двумя разными типами а-цепей, их формулы, соответственно [α1(I)]2α2(I) и [α1(IV)]2α2(IV). Индекс за скобкой обозначает количество идентичных а-цепей. Распределение коллагенов по органам и тканям представлено в табл. 15-1.
Гены коллагена называются соответственно типам коллагена и записываются арабскими цифрами, например COL1 - ген коллагена I типа, COL2 - ген коллагена II типа, COL7 -
ген коллагена VII типа и т.д. К этому символу приписываются буква А (обозначает а-цепь) и арабская цифра (обозначает вид а-цепи). Например, COL1A1 и COL1A2 кодируют, соответственно, α1 и α2-цепи коллагена I типа.
А. ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Синтез и созревание коллагена - сложный многоэтапный процесс, начинающийся в клетке, а завершающийся в межклеточном матриксе. Синтез и созревание коллагена включают в себя целый ряд посттрансляционных изменений
(рис. 15-1):
• гидроксилирование пролина и лизина с образованием гидроксипролина (Hyp) и гидроксилизина (Hyl);
• гликозилирование гидроксилизина;
Таблица 15-1. Распределение коллагена в тканях и органах
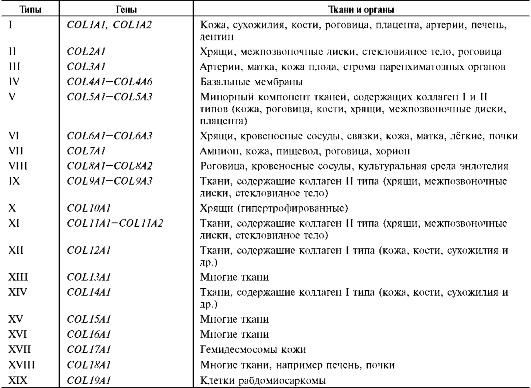
Рис. 15-1. Синтез и созревание коллагена.
• частичный протеолиз - отщепление «сигнального» пептида, а также N- и С-концевых пропептидов;
• образование тройной спирали.
Синтез полипептидных цепей коллагена
Полипептидные цепи коллагена синтезируются на полирибосомах, связанных с мембранами ЭР, в виде более длинных, чем зрелые цепи, предшественников - препро-а-цепей. У этих предшественников имеется гидрофобный «сигнальный» пептид на N-конце, содержащий около 100 аминокислот.
Основная функция сигнального пептида - ориентация синтеза пептидных цепей в полость ЭР. После выполнения этой функции сигнальный пептид сразу же отщепляется. Синтезированная молекула проколлагена содержит дополнительные участки - N- и С-концевые пропептиды, имеющие около 100 и 250 аминокислот, соответственно. В состав пропептидов входят остатки цистеина, которые образуют внутри- и межцепочечные (только в С-пептидах) S-S-связи. Концевые про-пептиды не образуют тройную спираль, а формируют глобулярные домены. Отсутствие N- и С-концевых пептидов в структуре проколлагена нарушает правильное формирование тройной спирали.
Посттрансляционные модификации коллагена
Гидроксилирование пролина и лизина. Роль витамина С
Гидроксилирование пролина и лизина начинается в период трансляции коллагеновой мРНК на рибосомах и продолжается на растущей полипептидной цепи вплоть до её отделения от рибосом. После образования тройной спирали дальнейшее гидроксилирование пролиловых и лизиловых остатков прекращается.
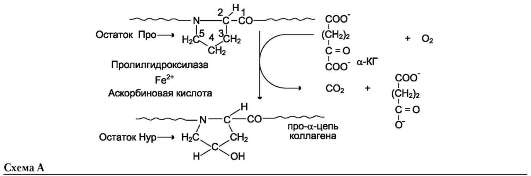
Реакции гидроксилирования катализируют оксигеназы, связанные с мембранами микросом. Пролиловые и лизиловые остатки в Y-положе-нии пептида (Гли-х-у)n подвергаются действию, соответственно, пролил-4-гидроксилазы и лизил-5-гидроксилазы. Пролил-3-гидроксила-за действует на некоторые остатки пролина в Х-положениях. Необходимыми компонентами этой реакции являются а-кетоглутарат, О2 и витамин С (аскорбиновая кислота). Донором атома кислорода, который присоединяется к С-4 пролина, является молекула О2, второй атом О2 включается в сукцинат, который образуется при декарбоксилировании а-кетоглутарата, а из карбоксильной группы а-кетоглутарата образуется СО2 (см. схему А на с. 691).
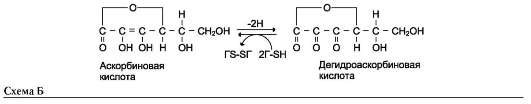
Гидроксилазы пролина и лизина содержат в активном центре атом железа Fe2+. Для сохранения атома железа в ферроформе необходим восстанавливающий агент. Роль этого агента выполняет кофермент гидроксилаз - аскорбиновая кислота, которая легко окисляется в дегидроаскорбиновую кислоту. Обратное превращение происходит в ферментативном процессе за счёт восстановленного глутатиона (см. схему Б на с. 691).
Гидроксилирование пролина необходимо для стабилизации тройной спирали коллагена, ОН-группы гидроксипролина (Hyp) участвуют в образовании водородных связей. А гидрок-силирование лизина очень важно для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл. При цинге - заболевании, вызванном недостатком витамина С, нарушается гид-роксилирование остатков пролина и лизина. В результате этого образуются менее прочные и стабильные коллагеновые волокна, что приводит к большой хрупкости и ломкости кровеносных сосудов с развитием цинги. Клиническая картина цинги характеризуется возникновением множественных точечных кровоизлияний под кожу и слизистые оболочки, кровоточивостью дёсен, выпадением зубов, анемией.
Гликозилирование гидроксилизина
После завершения гидроксилирования при участии специфических гликозилтрансфераз в состав молекулы проколлагена вводятся угле-
водные группы. Чаще всего этими углеводами служат галактоза или дисахарид галактозилглю-коза (рис. 15-2).
Они образуют ковалентную О-гликозидную связь с 5-ОН-группой гидроксилизина. Гли-козилирование гидроксилизина происходит в коллагене, ещё не претерпевшем спирализа-ции, и завершается после образования тройной спирали. Число углеводных единиц в молекуле коллагена зависит от вида ткани. Так, например, в коллагене сухожилий (тип I) это число равно 6, а в коллагене капсулы хрусталика (тип IV) - 110. Роль этих углеводных групп неясна; известно только, что при наследственном заболевании, причиной которого является дефицит лизилгидроксилазы (синдром Элерса-Данло-Ру-сакова, тип VI), содержание гидроксилизина и углеводов в образующемся коллагене снижено; возможно, это является причиной ухудшения механических свойств кожи и связок у людей с этим заболеванием.
Образование проколлагена и его секреция в межклеточное пространство
После гидроксилирования и гликозилирова-ния каждая про-α-цепь соединяется водородными связями с двумя другими про-а-цепями, образуя тройную спираль проколлагена. Эти процессы происходят ещё в просвете ЭР и начинаются после образования межцепочечных дисульфидных мостиков в области С-концевых пропептидов. Из ЭР молекулы проколлагена перемещаются в аппарат Гольджи, включаются
Рис. 15-2. Углеводные компоненты коллагена.
в секреторные пузырьки и секретируются в межклеточное пространство. Образование тропоколлагена. Болезни, связанные с нарушениями этого процесса
В межклеточном матриксе концевые пропеп-тиды коллагенов I, II и III типов отщепляются специфическими проколлагенпептидазами, в результате чего образуются молекулы тропокол-лагена, которые и являются структурной единицей коллагеновых фибрилл. При снижении активности этих ферментов (синдром Элер-са-Данло-Русакова, тип VII) концевые пропеп-тиды проколлагена не отщепляются, вследствие чего нарушается образование тропоколлагена и далее нарушается образование нормальных кол-лагеновых фибрилл. Нити коллагена видны под микроскопом в виде дез-организованных пучков. Клинически это проявляется малым ростом, искривлением позвоночника, привычными вывихами суставов, высокой растяжимостью кожи.
У коллагенов некоторых типов (IV, VIII, X) концевые пропептиды не отщепляются. Это связано с тем, что такие коллагены образуют не фибриллы, а сетеподобные структуры, в формировании которых важную роль играют концевые N- и С-пептиды.
Б. ОСОБЕННОСТИ СТРУКТУРЫ И ФУНКЦИИ РАЗНЫХ ТИПОВ КОЛЛАГЕНОВ
19 типов коллагена подразделяют на несколько классов в зависимости от того, какие структуры они могут образовывать. Эти структуры представлены в табл. 15-2.
Фибриллообразующие (I, II, III, V и XI) типы
95% всего коллагена в организме человека составляют коллагены I, II и III типов, ко-
Таблица 15-2. Классификация коллагенов по видам структур, которые они образуют
торые образуют очень прочные фибриллы. Значительное содержание именно этих типов коллагена объясняется тем, что они являются основными структурными компонентами органов и тканей, которые испытывают постоянную или периодическую механическую нагрузку (кости, сухожилия, хрящи, межпозвоночные диски, кровеносные сосуды), а также участвуют в образовании стромы паренхиматозных органов. Поэтому коллагены I, II и III типов часто называют интерстициальными. К классу фибриллообразующих относят также минорные коллагены V и XI типов.
Структура фибрилл коллагена и их формирование
Основа структурной организации коллаге-новых фибрилл - ступенчато расположенные параллельные ряды молекул тропоколлагена, которые сдвинуты на 1/4 относительно друг друга (рис. 15-3).
На схеме хорошо видно, что молекулы коллагена не связаны между собой «конец в конец», а между ними имеется промежуток в 35-40 нм. Предполагается, что в костной ткани эти промежутки выполняют роль центров минерализации, где откладываются кристаллы фосфата кальция. При электронной микроскопии фиксированные и контрастированные фибриллы коллагена выглядят поперечно исчерченными с периодом 67 нм, который включает одну тёмную и одну светлую полоски. Считают, что такое строение максимально повышает сопротивление всего агрегата растягивающим нагрузкам.
Фибриллы
коллагена образуются спонтанно, путём самосборки. Но эти фибриллы ещё
не являются зрелыми, так как не обладают достаточной прочностью
(известно, что зрелое колла-геновое волокно толщиной в
Образовавшиеся коллагеновые фибриллы укрепляются внутри- и межцепочечными кова-лентными сшивками (они встречаются только в коллагене и эластине). Эти сшивки образуются следующим образом:
• внеклеточный медьсодержащий фермент лизилоксидаза осуществляет окислительное дезаминирование ε-аминогрупп в некоторых остатках лизина и гидроксилизина с образованием реактивных альдегидов (аллизина
Рис. 15-3. Схема ступенчатого расположения молекул коллагена в коллагеновой фибрилле.
и гидроксиаллизина). Для этих реакций необходимо присутствие витаминов РР и
В6 (рис. 15-4).
• образовавшиеся реактивные альдегиды участвуют в формировании ковалентных связей между собой, а также с другими остатками лизина или гидроксилизина соседних молекул тропоколлагена, и в результате возникают поперечные «Лиз-Лиз-сшивки», стабилизирующие фибриллы коллагена
(рис. 15-5).
Количество поперечных связей в фибриллах коллагена зависит от функции и возраста ткани. Например, между молекулами коллагена ахиллова сухожилия сшивок особенно много, так как для этой структуры важна большая прочность. С возрастом количество поперечных связей в фибриллах коллагена возрастает, что приводит к замедлению скорости его обмена у пожилых и старых людей.
При снижении активности лизилоксидазы, а также при недостатке меди или витаминов РР или В6 нарушается образование поперечных сшивок и, как следствие, снижаются прочность и упругость коллагеновых волокон. Такие структуры, как кожа, сухожилия, кровеносные сосуды, становятся хрупкими, легко разрываются. Подробнее эти вопросы рассматриваются ниже в подразделе, посвящённом эластину.
Коллагены, ассоциированные с фибриллами
Этот класс объединяет коллагены, которые выполняют очень важную функцию: они ограничивают размер фибрилл, образуемых интерстициальными коллагенами (прежде всего, I и II типов), и участвуют в организации межклеточного матрикса в костях, коже, хрящах, сухожилиях. К этим коллагенам относят коллагены IX, XII, XIV и XVI типов. Коллагены этого класса сами фибрилл не формируют, но непосредственно связаны с фибриллами, которые образуют интерстициальные коллагены. Функционирование таких типов коллагенов можно рассмотреть на примере коллагена IX типа, который в хряще связан с фибриллами коллагена II типа, он присоединяется к ним антипараллельно с периодичностью ~67 нм (рис. 15-6).
Коллаген IX типа состоит из трёх коллагеновых (фибриллярных) доменов (Кол1 → Кол3) и четырёх неколлагеновых (глобулярных) доменов (НК1 → НК4) (нумерация с С-конца) (рис. 15-7).
Коллаген IX типа связан с фибриллами коллагена II типа поперечными «Лиз-Лиз-мостиками» в области доменов Кол1 и Кол2, а также и НК1,
НК2 и НК3.
НК4-домен не связан с фибриллами коллагена II типа; к его особенностям относят наличие большого количества положительно заряженных
Рис. 15-4. Образование поперечных связей в коллагене. А - образование альдольной поперечной сшивки из двух боковых цепей лизина; Б - образование шиффовых оснований из боковых цепей лизина и аллизина.
групп, поэтому к нему могут присоединяться отрицательно заряженные гликозаминогликаны, например, гиалуроновая кислота и хондроитин-сульфат. Эти особенности обеспечивают участие коллагена IX типа в организации межклеточного матрикса в хряще.
Коллагены, образующие сетеподобные структуры
К этому классу относят коллагены IV, VIII, X типов. Особенности строения и функционирования таких белков можно рассмотреть на примере наиболее изученных к настоящему времени коллагенов IV и VIII типов.
Коллаген IV типа является ключевым структурным компонентом базальных мембран, которые представляют собой особую форму межклеточного матрикса. Его секретируют различные типы клеток: эпителиальные, эндотелиальные, мышечные, нервные, жировые. Особенностью коллагена IV типа является то, что повторяющиеся спирализованные участки с последовательностью (Гли-х-у)n часто прерываются короткими неспиральными сегментами. Это, вероятно, увеличивает гибкость коллагена IV типа и способствует образованию на его основе сетчатых структур (рис. 15-8).
Молекулы этого коллагена не могут ассоциироваться латерально с образованием фибрилл, так как N- и С-концевые пропептиды у него не отщепляются. Но именно эти фрагменты участвуют в образовании олигомерных форм коллагена, так как они имеют ряд потенциальных мест связывания (остатки цистеина и лизина). Дисульфидные мостики и поперечные лизиновые связи стабилизируют образующиеся олигомеры. Кроме этого, возможны латеральные взаимодействия спирализованных участков разных молекул с образованием суперспиралей. В базальной мембране из этих компонентов формируется сетчатая структура с гексагональными ячейками размером 170 нм.
Коллагены VIII и X типов относят к так называемым короткоцепочечным коллагенам. Каждая их молекула состоит из короткого коллагенового домена, который составляет ~1/2 длины ин-терстициальных коллагенов, и неколлагеновых фрагментов на N- и С-концах.
Коллаген VIII типа - главный компонент десцеметовых мембран эндотелия роговицы. Молекулы этого коллагена собираются анти-параллельно с образованием тетрамеров, из которых образуются гексагональные решётки, обеспечивающие прозрачность роговицы (рис. 15-9).
Кроме роговицы, коллаген VIII типа присутствует во многих других тканях, но ещё одна его преимущественная локализация - кровеносные
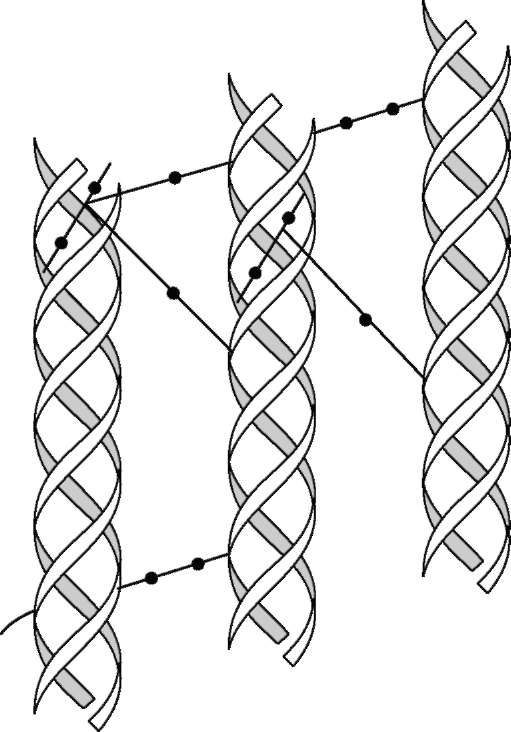
Рис. 15-5. Внутри- и межмолекулярные поперечные связи в коллагене.
сосуды, в которых он в основном находится в мат-риксе под эндотелиальными клетками. Образует ли этот коллаген и здесь гексагональные решётки, неизвестно. Возможно, что в сосудах коллаген VIII типа образует сетевидные структуры, подоб-
IX тип
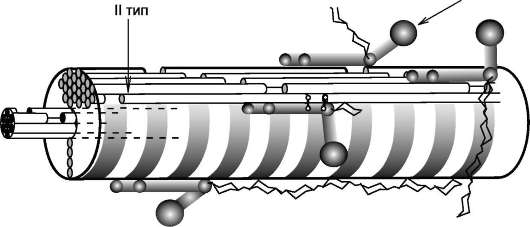
Рис. 15-6. Структура коллагеновых фибрилл II типа и ассоциированного с ним коллагена IX типа.

Рис. 15-7. Модель структуры коллагена IX типа.
КОЛ1-КОЛ3 - коллагеновые домены; НК1 - НК4 - неколлагеновые структуры.
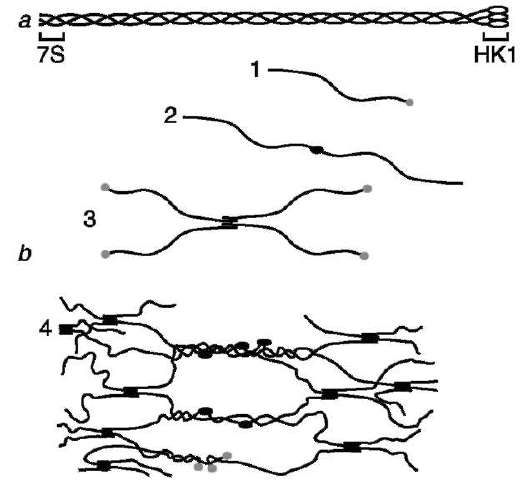
Рис. 15-8. Организация коллагена IV типа. А. Тройная спираль мономера коллагена: 7S - N-конец; НК1 - С-конец. Б. Полимеризация коллагена IV типа: 1 - мономер; 2 - димеры, образованные соединением мономеров в области НК1-доменов; 3 - тетраме-ры, образованные соединением мономеров в области 7S-сегментов в параллельном и антипараллельном направлениях; 4 - образование сетчатой структуры из олигомерных форм коллагена IV типа.
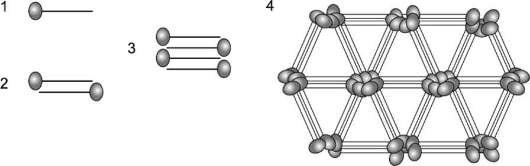
Рис. 15-9. Возможный механизм образования гексагональных решёток молекулами коллагена VIII типа.
1 - мономер; 2 - димер; 3 - тетрамер; 4 - гексагональные решётки.
ные тем, которые формирует коллаген IV типа в базальных мембранах.
Коллагены, образующие микрофибриллы
К этому классу относят коллаген VI типа, который является короткоцепочечным белком. Он образует микрофибриллы, которые располагаются между крупными фибриллами интерстициальных коллагенов. Этот коллаген
широко представлен в хрящевом матриксе, но больше всего его содержится в межпозвоночных дисках: в nucleus pulposus он составляет ~20% общего коллагена. Две молекулы этого коллагена соединяются антипараллельно с образованием димера. Из димеров образуются тетрамеры, которые секретируются из клетки, и вне клетки связываются «конец в конец» с образованием микрофибрилл (рис. 15-10).
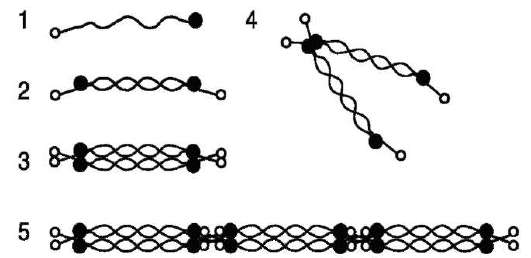
Рис. 15-10. Организация коллагена VI типа. 1 -
мономер; 2 - димер; 3 - тетрамер, соединённый полностью; 4 - тетрамер, соединённый частично; 5 - микрофибриллы, соединённые «конец в конец».
Функции коллагена VI типа пока полностью не ясны, хотя известно, что его микрофибриллы могут связываться со многими компонентами межклеточного матрикса: фибриллами интерс-тициальных коллагенов, гиалуроновой кислотой, протеогликанами. Молекула этого коллагена содержит многочисленные последовательности Арг-Гли-Асп (RGD), поэтому возможно его участие в клеточной адгезии через присоединение к мембранным адгезивным молекулам, например интегринам α1β1 и α2β1.
Коллагены, образующие «заякоренные» фибриллы
К этому классу относят коллагены VII и XVII типов, которые называют также коллагенами, связанными с эпителием, так как они обычно находятся в местах соединения эпителия с субэпителиальными слоями.
Коллаген VII типа - основной структурньгй компонент «заякоренных» фибрилл. Каждая молекула этого белка содержит два неколла-геновых домена (НК1 - у С-конца, НК2 - у N-конца) и один коллагеновый домен между ними. Из мономеров образуются димеры, при этом молекулы соединяются в области НК2-доме-нов антипараллельно по отношению друг к другу. Затем НК2-домены отщепляются, и димеры соединяются между собой «бок о бок» с образованием фибрилл (рис. 15-11).
Эти фибриллы играют важную роль в присоединении эпидермиса к дерме, так как одним концом они могут присоединяться к lamina densa, на которой лежит кожный эпителий, а другой их конец проникает в более глубокие субэпи-
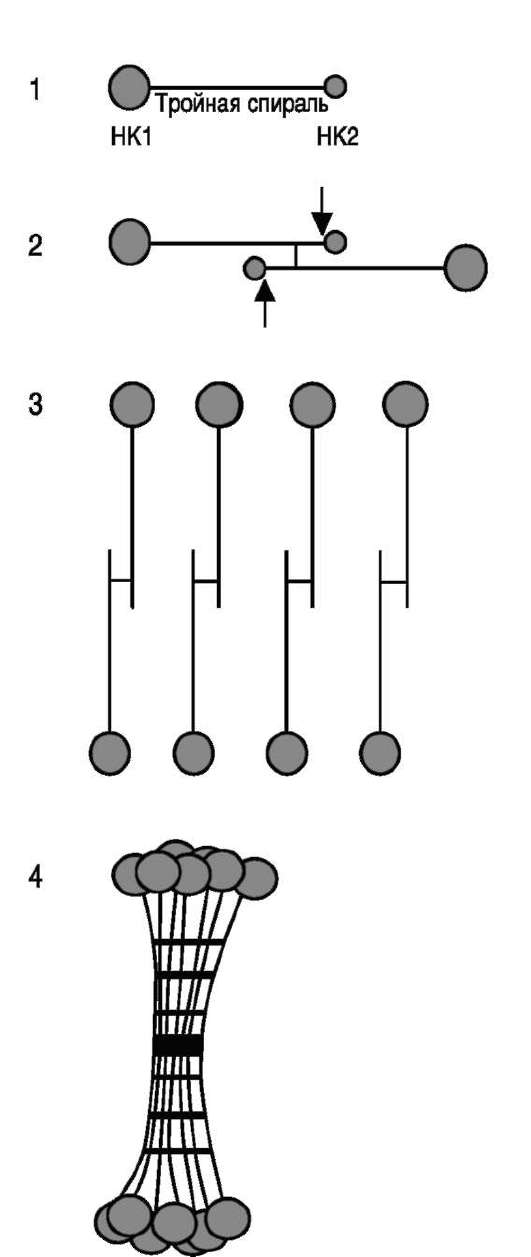
Рис. 15-11. Организация коллагена VII типа.
1 - мономер коллагена VII типа, НК1 и НК2 - не-коллагеновые домены у С- и N-конца; 2 - димер коллагена VII типа, молекулы собраны антипараллельно с перекрытиями на N-конце; 3 - димеры коллагена VII типа после удаления НК2-доменов; 4 - фибрилла, образованная димерами коллагена VII типа, соединёнными «бок о бок».
дермальные слои кожи и связывается там со структурами, называемыми «якорные диски».
Коллаген XVII типа представляет собой трансмембранный белок и обычно находится в гемидесмосомах эпидермиса. Предполагают, что этот коллаген взаимодействует с другими молекулами гемидесмосом и таким образом участвует в процессе присоединения эпидермиса к дерме.
В. КАТАБОЛИЗМ КОЛЛАГЕНА
Как и любой белок, коллаген функционирует в организме определённое время. Его относят к медленно обменивающимся белкам; T1/2 составляет недели или месяцы. Разрушение коллагеновых волокон осуществляется активными формами кислорода и/или ферментативно (гидролитически).
Коллагеназы, особенности их функционирования
Нативный коллаген не гидролизуется обычными пептидгидролазами. Основной фермент
его катаболизма - коллагеназа, которая расщепляет пептидные связи в определённых участках спирализованных областей коллагена. Известны 2 типа коллагеназ.
Тканевая коллагеназа присутствует у человека в различных органах и тканях. В норме она синтезируется клетками соединительной ткани, прежде всего, фибробластами и макрофагами. Тканевая коллагеназа - металлозависимый фермент, который содержит Zn2+ в активном центре. В настоящее время известно 4 изофор-мы этого фермента. Активность коллагеназы зависит от соотношения в межклеточном мат-риксе её активаторов и ингибиторов. Среди активаторов особую роль играют плазмин, калликреин и катепсин В (см. раздел 14). Тканевая коллагеназа обладает высокой специфичностью, она перерезает тройную спираль коллагена в определённом месте, примерно на 1/4 расстояния от С-конца, между остатками глицина и лейцина (или изолейцина) (см. схему ниже).
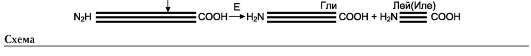
Образующиеся фрагменты коллагена растворимы в воде, при температуре тела они спонтанно денатурируются и становятся доступными для действия других протеолитических ферментов. Нарушение катаболизма коллагена ведёт к фиброзу органов и тканей (в основном печени и лёгких). А усиление распада коллагена происходит при аутоиммунных заболеваниях (ревматоидном артрите и системной красной волчанке) в результате избыточного синтеза коллагеназы при иммунном ответе.
Бактериальная коллагеназа синтезируется некоторыми микроорганизмами. Например, Clostridium histolyticum (возбудитель газовой гангрены) выделяет коллагеназу, расщепляющую пептидную цепь коллагена более чем в 200 местах. Этот фермент гидролизует следующую связь -Х-Гли-Про-Умежду звеньями Х и Гли.
Таким образом разрушаются соединительнотканные барьеры в организме человека, что обеспечивает проникновение (или инвазию) этого микроорганизма и способствует возникновению и развитию газовой гангрены. Сам
возбудитель не содержит коллагена и поэтому не подвержен действию коллагеназы.
Применение коллагеназ в медицине
Коллагеназа используется в медицинской практике для лечения ожоговой болезни в хирургии и для лечения гнойных заболеваний глаз в офтальмологии.
Определение гидроксипролина в физиологических жидкостях человека как показатель скорости распада коллагена
В результате распада коллагена в крови и моче появляется свободный гидроксипролин. Большая часть этой аминокислоты катаболи-зируется под действием фермента гидроксипро-лин-оксидазы, а часть её выводится с мочой, и поэтому гидроксипролин является маркёрной аминокислотой, по которой судят о скорости распада коллагена.
При некоторых заболеваниях, связанных с поражением соединительной ткани, экскреция
гидроксипролина увеличивается вследствие ускоренного распада коллагена. Это наблюдается при болезни Педжета, гиперпаратиреозе, коллагенозах, некоторых инфекционных заболеваниях. При нарушении катаболизма гидрокси-пролина, причиной которого обычно выступает дефект фермента гидроксипролиноксидазы, выделение гидроксипролина может превышать 1 г/сут.
Особенности обмена коллагена
У молодых людей обмен коллагена протекает интенсивно, с возрастом (и особенно в старости) заметно снижается, так как у пожилых и старых людей увеличивается количество поперечных сшивок, что затрудняет доступность коллагена для действия коллагеназы. Поэтому, если у молодых людей в возрасте 10-20 лет содержание гидроксипролина в моче может достигать 200 мг/сут, то с возрастом экскреция гидроксипро-лина снижается до 15-20 мг/сут.
В некоторых ситуациях синтез коллагена заметно увеличивается. Например, фибробласты мигрируют в заживающую рану и начинают активно синтезировать в этой области основные компоненты межклеточного матрикса. Результат этих процессов - образование на месте раны соединительнотканного рубца, содержащего большое количество хаотично расположенных фибрилл коллагена. Сходным образом происходит замещение погибающих клеток соединительной тканью в печени при циррозе, в стенках артерий при атеросклерозе, в мышцах при их дистрофии.
Г. РЕГУЛЯЦИЯ ОБМЕНА КОЛЛАГЕНА
Синтез коллагена регулируется разными способами. Прежде всего, сам коллаген и N-пропептиды после своего отщепления тормозят трансляцию коллагена по принципу отрицательной обратной связи. Аскорбиновая кислота стимулирует синтез коллагена и протеогликанов, а также пролиферацию фибробластов.
Особую роль в регуляции синтеза коллагена играют гормоны. Глюкокортикоиды тормозят синтез коллагена, во-первых, путём снижения уровня мРНК проколлагена, а во-вторых - ингибированием активности ферментов пролил- и лизилгидроксилазы. Недостаточное гидроксилирование остатков пролина и лизина
повышает чувствительность коллагена к действию коллагеназы и неспецифических протеаз. Макроскопически угнетающее действие глюко-кортикоидов на синтез коллагена проявляется уменьшением толщины дермы, а также атрофией кожи в местах продолжительного парентерального введения этих гормонов.
На синтез коллагена влияют также половые гормоны, рецепторы к которым обнаружены не только в строме половых органов, но и в фибробластах других органов и тканей. Обмен коллагена в матке находится под контролем половых гормонов. Синтез коллагена кожи зависит от содержания эстрогенов, что подтверждает тот факт, что у женщин в менопаузе снижается содержание коллагена в дерме.
Д. ЗАБОЛЕВАНИЯ, СВЯЗАННЫЕ С НАРУШЕНИЕМ СИНТЕЗА И СОЗРЕВАНИЯ
КОЛЛАГЕНА
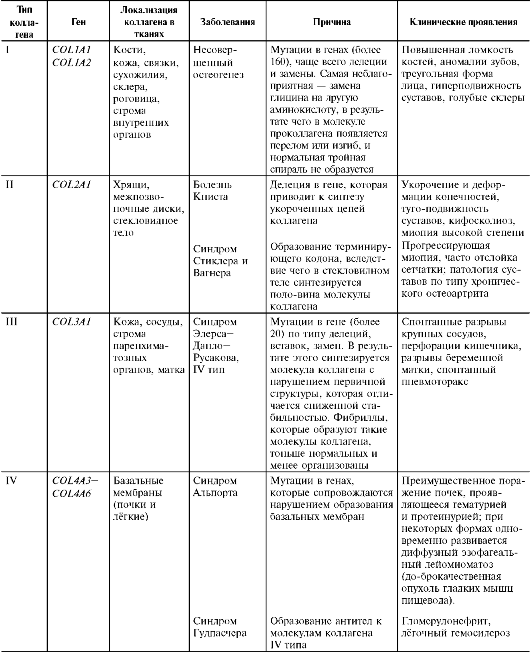
Существует ряд заболеваний, связанных с нарушением структуры или синтеза коллагена. Основная причина - мутации в генах коллагена, которые широко представлены в разных хромосомах. Они очень большие, имеют много коротких экзонов, между которыми располагаются большие интроны.
Так как около 50% всех коллагеновых белков содержится в тканях скелета, около 40% - в коже и 10% - в строме внутренних органов, клиническая картина заболеваний, вызванных дефектами синтеза и созревания коллагена, будет крайне полиморфной. При многих заболеваниях наблюдают не только костно-суставную патологию или изменения со стороны кожи, но и ярко выраженные висцеральные проявления (поражения кишечника, почек, лёгких, сердца, сосудов).
К настоящему времени описано много наследственных заболеваний, причинами которых являются дефекты коллагенов разных типов (см. ниже табл. 15-3).
ii. эластин
Эластин - основной белок эластических волокон, которые в больших количествах содержатся в межклеточном веществе таких тканей, как кожа, стенки кровеносных сосудов, связки, лёгкие. Эти ткани обладают очень важными
свойствами: они могут растягиваться в несколько раз по сравнению с исходной длиной, сохраняя при этом высокую прочность на разрыв, и возвращаться в первоначальное состояние после снятия нагрузки. Резиноподобные свойства названных тканей обеспечиваются особенностями состава и строения эластина - гликопротеина с молекулярной массой 70 кД.
А. СТРУКТУРА ЭЛАСТИНА
1. Аминокислотный состав и особенности конформации эластина описаны в 1-м разделе учебника.
Значение десмозина и лизиннорлейцина
В межклеточном пространстве молекулы эластина образуют волокна и слои, в которых отдельные пептидные цепи связаны множеством жёстких поперечных сшивок в разветвлённую сеть. В образовании этих сшивок участвуют остатки лизина двух, трёх или четырёх пептидных цепей. Структуры, образующиеся при этом, называются десмозинами (десмозин или изодесмозин). Предполагают, что эти гетероциклические соединения формируются следующим образом: вначале 3 остатка лизина окисляются до соответствующих ε-альдегидов, а затем происходит их соединение с четвёртым остатком лизина с образованием замещённого пиридинового кольца. Окисление остатков лизина в ε-альдегиды осуществляется медьзависимой ли-зилоксидазой, активность которой зависит также от наличия пиридоксина (см. подразд. I, Б).
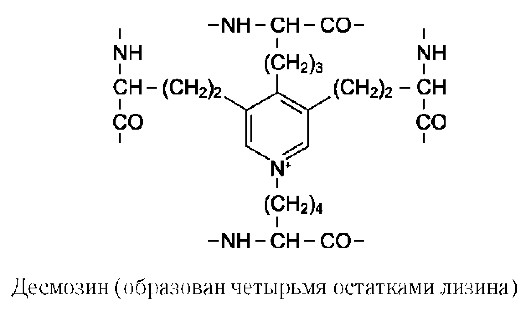
Кроме десмозинов, в образовании поперечных сшивок может участвовать лизиннорлейцин, который образуется двумя остатками лизина.
Наличие ковалентных сшивок между пептидными цепочками с неупорядоченной, случайной конформацией позволяет всей сети волокон эластина растягиваться и сжиматься в разных направлениях, придавая соответствующим тканям свойство эластичности (рис. 15-12).
Следует отметить, что эластин синтезируется как растворимый мономер, который называется «тропоэластин». После образования поперечных сшивок эластин приобретает свою конечную внеклеточную форму, которая характеризуется нерастворимостью, высокой стабильностью и очень низкой скоростью обмена.
Нарушения структуры эластина и их последствия
При снижении образования десмозинов (или их отсутствии) поперечные сшивки образуются в недостаточном количестве или не образуются вообще. Вследствие этого у эластических тканей снижается предел прочности на разрыв и появляются такие нарушения, как истончён-ность, вялость, растяжимость, т.е. утрачиваются их резиноподобные свойства. Клинически такие нарушения могут проявляться кардиоваску-лярными изменениями (аневризмы и разрывы аорты, дефекты клапанов сердца), частыми пневмониями и эмфиземой лёгких.
Причины нарушений структуры эластина
• снижение активности лизилоксидазы, вызванное дефицитом меди или пиридок-сина;
• дефицит лизилоксидазы при наследственных заболеваниях;
• синдром Менкеса - нарушение всасывания меди.
Б. КАТАБОЛИЗМ ЭЛАСТИНА Переваривание эластина
Нативный эластин, содержащийся в пище, не гидролизуется трипсином и химотрипсином, но медленно расщепляется пепсином при рН 2,0. Эластаза поджелудочной железы гидролизует
Таблица 15-3. Заболевания, связанные с нарушением синтеза и созревания коллагена
Окончание таблицы 15-3.

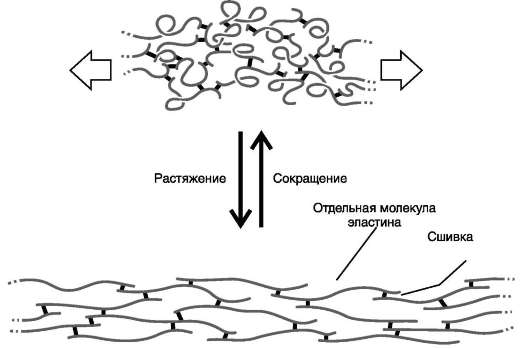
Рис. 15-12. Молекулы эластина связаны ковалентными сшивками в обширную сеть.
эластин после выраженного лаг-периода. Это эндопептидаза, которая преимущественно расщепляет связи, образованные карбоксильными группами алифатических аминокислот.
Разрушение эластина
Катаболизм эластина происходит при участии эластазы нейтрофилов. Это очень активная протеаза, которая выделяется во внеклеточное пространство нейтрофилами и разрушает эластин и другие структурные белки. Особое значение это имеет в лёгких. Поскольку лёгочная ткань не регенерирует, разрушение эластина в альвеолярных стенках ведёт к потере эластичных свойств, разрушению альвеол и развитию
эмфиземы лёгких (растяжение лёгких воздухом или образовавшимся в тканях газом).
В норме этого не происходит, так как эласта-зу нейтрофилов и другие протеазы ингибирует белок, называемый α1-антитрипсином (α1-АТ). Основное количество α1-АТ синтезируется печенью и находится в крови. В лёгких α1-АТ синтезируется альвеолярными макрофагами, что и обеспечивает защиту альвеол от действия эла-стазы (рис. 15-13). При дефиците α1-АТ, который может быть следствием различных мутаций в гене этого белка, повышается риск развития эмфиземы лёгких. В настоящее время это состояние поддаётся профилактике и лечению еженедельным внутривенным введением α1-АТ.
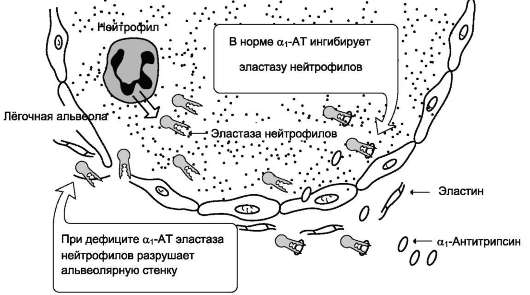
Рис. 15-13. Разрушение лёгочных альвеол эластазой нейтрофилов.
iii. гликозаминогликаны и протеогликаны
Гликозаминогликаны - линейные отрицательно заряженные гетерополисахариды. Раньше их называли мукополисахаридами, так как они обнаруживались в слизистых секретах (мукоза) и придавали этим секретам вязкие, смазочные свойства. Эти свойства обусловлены тем, что гликозаминогликаны могут связывать большие количества воды, в результате чего межклеточное вещество приобретает желеобразный характер.
Протеогликаны - высокомолекулярные соединения, состоящие из белка (5-10%) и гли-козаминогликанов (90-95%). Они образуют основное вещество межклеточного матрикса соединительной ткани и могут составлять до 30% сухой массы ткани.
Белки в протеогликанах представлены одной полипептидной цепью разной молекулярной массы. Полисахаридные компоненты у разных протеогликанов разные. Протеогликаны отличаются от большой группы белков, которые называют гликопротеинами. Эти белки тоже содержат олигосахаридные цепи разной длины, ковалентно присоединённые к полипептидной основе. Углеводный компонент гликопротеинов гораздо меньше по массе, чем у протеогликанов,
и составляет не более 40% от общей массы. Гли-копротеины выполняют в организме человека разные функции и присутствуют во всех классах белков - ферментах, гормонах, транспортных, структурных белках и др. Представители гликопротеинов - коллаген и эластин, иммуноглобулины, ангиотензиноген, трансферрин, церулоплазмин, внутренний фактор Касла, тиреотропный гормон.
Гликозаминогликаны и протеогликаны, являясь обязательными компонентами межклеточного матрикса, играют важную роль в межклеточных взаимодействиях, формировании и поддержании формы клеток и органов, образовании каркаса при формировании тканей.
Благодаря особенностям своей структуры и физико-химическим свойствам, протеогликаны и гликозаминогликаны могут выполнять в организме человека следующие функции:
• они являются структурными компонентами межклеточного матрикса;
• протеогликаны и гликозаминогликаны специфически взаимодействуют с коллагеном, эластином, фибронектином, ламинином и другими белками межклеточного матрикса;
• все протеогликаны и гликозаминогликаны, являясь полианионами, могут присоединять, кроме воды, большие количества катионов (Na+, K+, Ca2+) и таким образом участво-
вать в формировании тургора различных тканей;
• протеогликаны и гликозаминогликаны играют роль молекулярного сита в межклеточном матриксе, они препятствуют распространению патогенных микроорганизмов;
• гиалуроновая кислота и протеогликаны выполняют рессорную функцию в суставных хрящах;
• гепарансульфатсодержащие протеогликаны способствуют созданию фильтрационного барьера в почках;
• кератансульфаты и дерматансульфаты обеспечивают прозрачность роговицы;
• гепарин - антикоагулянт;
• гепарансульфаты - компоненты плазматических мембран клеток, где они могут функционировать как рецепторы и участвовать в клеточной адгезии и межклеточных взаимодействиях. Они также выступают компонентами синаптических и других пузырьков.
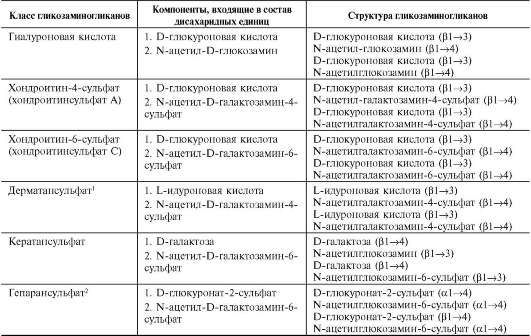
А. СТРОЕНИЕ И КЛАССЫ ГЛИКОЗАМИНОГЛИКАНОВ
Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополи-
сахаридов. Они построены из повторяющихся дисахаридных единиц. Одним мономером этого дисахарида является гексуроновая кислота (D-глюкуроновая кислота или L-идуроновая), вторым мономером - производное аминосахара (глюкоз- или галактозамина). NH2-группа ами-носахаров обычно ацетилирована, что приводит к исчезновению присущего им положительного заряда. Кроме гиалуроновой кислоты, все гли-козаминогликаны содержат сульфатные группы в виде О-эфиров или N-сульфата.
В настоящее время известна структура шести основных классов гликозаминогликанов, которые представлены в табл. 15-4.
Гиалуроновая кислота находится во многих органах и тканях. В хряще она связана с белком и участвует в образовании протеогликановых агрегатов, в некоторых органах (стекловидное тело глаза, пупочный канатик, суставная жидкость) встречается и в свободном виде. Предполагается, что в суставной жидкости гиалуроновая кислота выполняет роль смазочного вещества, уменьшая трение между суставными поверхностями.
Повторяющаяся дисахаридная единица в гиалуроновой кислоте имеет следующую структуру:
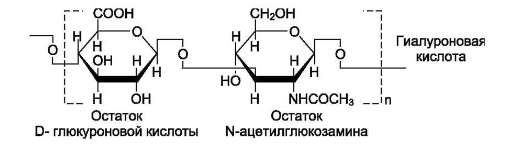
Гиалуроновая кислота содержит несколько тысяч дисахаридных единиц, молекулярная масса её достигает 105-107 Д.
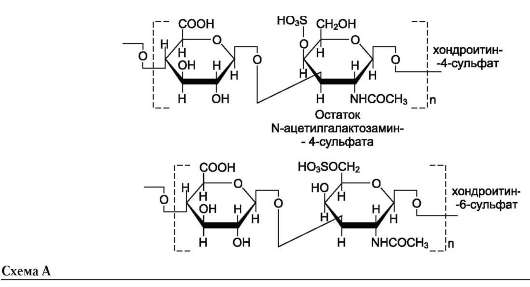
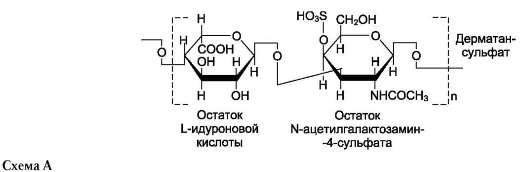
Хондроитинсульфаты - самые распространённые гликозаминогликаны в организме человека; они содержатся в хряще, коже, сухожилиях, связках, артериях, роговице глаза. Хондроитин-сульфаты являются важным составным компонентом агрекана - основного протеогликана хрящевого матрикса. В организме человека встречаются 2 вида хондроитинсульфатов: хон-дроитин-4-сульфат и хондроитин-6-сульфат. Они построены одинаковым образом, отличие касается только положения сульфатной группы в молекуле N-ацетилгалактозамина (см. схему А).
Одна полисахаридная цепь хондроитинсуль-фата содержит около 40 повторяющихся диса-харидных единиц и имеет молекулярную массу 104-106 Д.
Кератансульфаты - наиболее гетерогенные гликозаминогликаны; отличаются друг от друга по суммарному содержанию углеводов и распределению в разных тканях. Кератансульфат I находится в роговице глаза и содержит кроме повторяющейся дисахаридной единицы L-фу-козу, D-маннозу и сиаловую кислоту. Кера-тансульфат II был обнаружен в хрящевой ткани, костях, межпозвоночных дисках. В его состав помимо сахаров дисахаридной единицы входят N-ацетилгалактозамин, L-фукоза, D-манноза
Таблица 15-4. Структура различных классов гликозаминогликанов
1 В состав дисахаридной единицы может входить D-глюкуроновая кислота.
2 Может содержать N-сульфопроизводное глюкозамина вместо N-ацетилглюкозамина и различное количество идуроновой и глюкуроновой кислот.
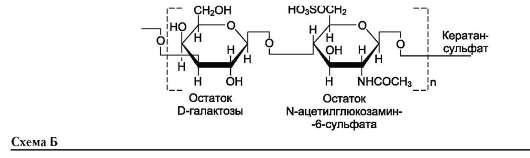
и сиаловая кислота. Кератансульфат II входит в состав агрекана и некоторых малых проте-огликанов хрящевого матрикса. В отличие от других гликозаминогликанов, кератансульфаты вместо гексуроновой кислоты содержат остаток галактозы (см. схему Б выше).
Молекулярная масса одной цепи кератансуль-фата колеблется от 4х103 до 20х103 Д.
Дерматансульфат широко распространён в тканях животных, особенно он характерен для кожи, кровеносных сосудов, сердечных клапанов.
В составе малых протеогликанов (бигликана и декорина) дерматансульфат содержится в межклеточном веществе хрящей, межпозвоночных дисков, менисков. Повторяющаяся дисахаридная единица дерматансульфата имеет следующую структуру (см. схему А).
Молекулярная масса одной цепи дерма-тансульфата колеблется от 15х103 до 40х103 Д.
Гепарин - важный компонент противосвёр-тывающей системы крови (его применяют как антикоагулянт при лечении тромбозов). Он синтезируется тучными клетками и находится в гранулах внутри этих клеток. Наибольшие количества гепарина обнаруживаются в лёгких, печени и коже. Дисахаридная единица гепарина похожа на дисахаридную единицу гепа-рансульфата. Отличие этих гликозаминоглика-нов заключается в том, что в гепарине больше N-сульфатных групп, а в гепарансульфате больше N-ацетильных групп. Молекулярная масса гепарина колеблется от 6х103 до 25х103 Д (см. схему Б).
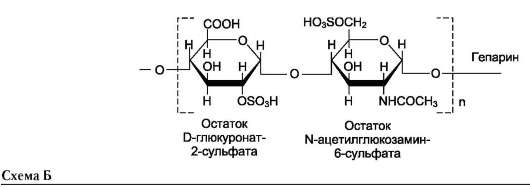
Гепарансульфат находится во многих органах и тканях. Он входит в состав протеогликанов базальных мембран. Гепарансульфат является постоянным компонентом клеточной поверхности. Структура дисахаридной единицы гепа-рансульфата такая же, как у гепарина. Молекулярная масса цепи гепарансульфата колеблется от 5х103 до 12х103 Д.
Б. СИНТЕЗ И РАЗРУШЕНИЕ ГЛИКОЗАМИНОГЛИКАНОВ
Метаболизм гликозаминогликанов зависит от соотношения скорости их синтеза и распада.
Синтез гликозаминогликанов
Полисахаридные цепи гликозаминогликанов практически всегда связаны с белком, который называется ко'ровым, или сердцевинным. Присоединение полисахарида к белку осуществляется через связующую область, в состав которой чаще всего входит трисахарид галактоза-галактоза-ксилоза (рис. 15-14).
Олигосахариды связующей области присоединяются к кóровому белку ковалентными связями 3 типов:
1. О-гликозидной связью между серином и ксилозой;
2. О-гликозидной связью между серином или треонином и N-ацетилгалактозамином;
3. N-гликозиламиновой связью между амид-ным азотом аспарагина и N-ацетилглюко-замином.
Полисахаридные цепи гликозаминоглика-нов синтезируются путём последовательного присоединения моносахаридов. Донорами моносахаридов обычно являются соответствующие нуклеотид-сахара. Реакции синтеза гликозамино-гликанов катализируют ферменты семейства трансфераз, обладающие абсолютной субстратной специфичностью. Эти трансферазы локализованы на мембранах аппарата Гольджи. Сюда по каналам ЭР поступает коровый белок, синтезированный на полирибосомах, к которому присоединяются моносахариды связующей области и затем наращивается вся полисахаридная цепь. Сульфатирование углеводной части происходит здесь с помощью сульфотрансферазы, донором сульфатной группы выступает ФАФС (см. раздел 12).
Аминосахара синтезируются из глюкозы; в соединительной ткани ~20% глюкозы используется таким образом. Непосредственным предшественником N-ацетилглюкозамина, N-аце-тилгалактозамина и сиаловой кислоты является фруктозо-6-фосфат. Источником NH2-группы в этих сахарах служит глутамин. Аминосахар далее ацетилируется с помощью ацетил-КоА. Активированными формами этих аминосахаров служат их УДФ-производные (схема на след. стр., рис. 15-15).
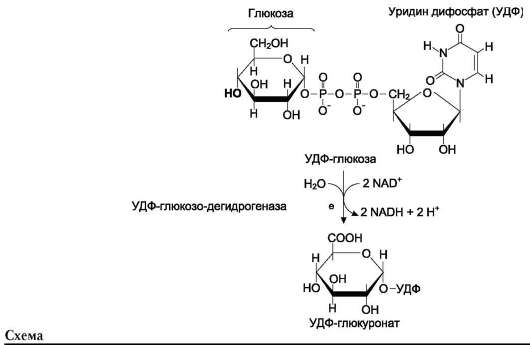
Источниками глюкуроновой кислоты в организме человека могут быть пища, внутриклеточное лизосомальное разрушение гликозами-ногликанов и синтез глюкуроновой кислоты. Активированная форма глюкуроновой кислоты
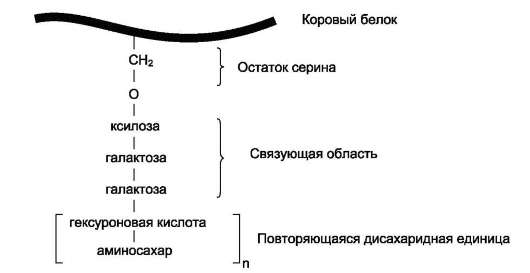
Рис. 15-14. Связующая область гликозаминогликанов.
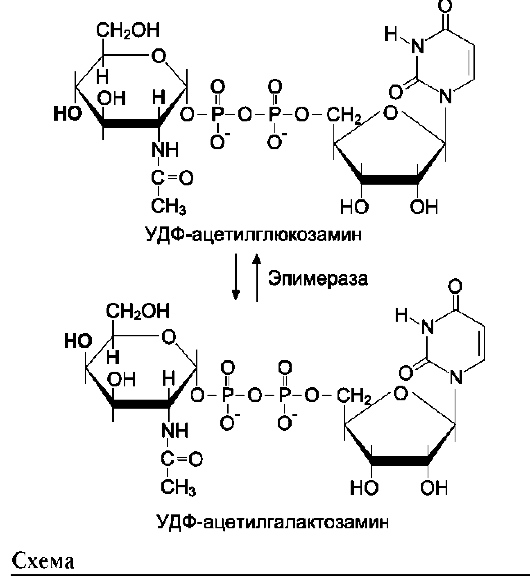
(УДФ-глюкуронат) образуется при окислении УДФ-глюкозы (см. схему ниже).
L-идуроновая кислота образуется после включения D-глюкуроновой кислоты в углеводную цепь в результате реакции эпимеризации.
На синтез гликозаминогликанов влияют глю-кокортикоиды: они тормозят синтез гиалуроно-
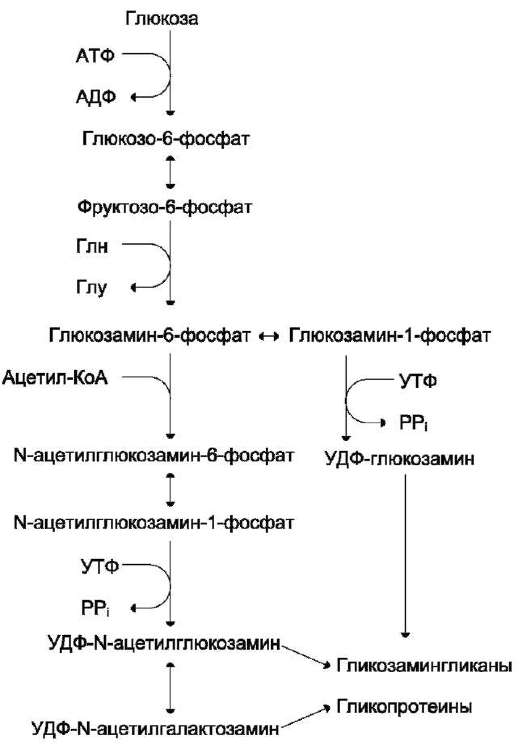
Рис. 15-15. Схема синтеза аминосахаров.
вой кислоты и сульфатированньгх гликозаминогликанов. Показано также тормозящее действие половых гормонов на синтез сульфатированных гликозаминогликанов в органах-мишенях.
Разрушение гликозаминогликанов
Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т1/2) многих из них составляет от 3 до 10 дней (только для кератансульфата Т1/2 =120 дней). Разрушение полисахаридных цепей осуществляется экзо- и эндогликозидазами и сульфатазами, к которым относят гиалуронидазу, глюкуронидазу, галак-тозидазу, идуронидазу и др. Из внеклеточного пространства гликозаминогликаны поступают в клетку по механизму эндоцитоза и заключаются в эндоцитозные пузырьки, которые затем сливаются с лизосомами. Лизосомальные гидролазы обеспечивают постепенное полное расщепление гликозаминогликанов до мономеров.
Мукополисахаридозы - наследственные тяжёлые заболевания, проявляющиеся значительными нарушениями в умственном развитии детей, поражениями сосудов, помутнением роговицы, деформациями скелета, уменьшением продолжительности жизни. В основе мукопо-лисахаридозов лежат наследственные дефекты каких-либо гидролаз, участвующих в катаболизме гликозамино-гликанов. Эти заболевания характеризуются избыточным накоплением гликозаминогликанов в тканях, приводящим к деформации скелета и увеличению органов, содержащих большие количества внеклеточного матрикса. Обычно поражаются ткани, в которых
Таблица 15-5. Типы мукополисахаридозов
в норме синтезируются наибольшие количества гликозаминогликанов. В лизосомах при этом накапливаются не полностью разрушенные гликозаминогликаны, а с мочой выделяются их олигосахаридные фрагменты. Известно несколько типов мукополисахаридозов, вызванных дефектами разных ферментов гидролиза глико-заминогликанов. Основные типы мукополиса-харидозов приведены в табл. 15-5.
Для постановки диагноза конкретного заболевания обычно определяют активность лизосомальных гидролаз. Так как эти болезни в настоящее время не поддаются лечению, необходимо проводить пренатальную диагностику при подозрении на носительство дефектных генов.
В. СТРОЕНИЕ И ВИДЫ ПРОТЕОГЛИКАНОВ
В межклеточном матриксе присутствуют разные протеогликаны. Среди них есть очень крупные - например агрекан и версикан. Кроме них, в межклеточном матриксе имеется целый набор так называемых малых протеогликанов, которые широко распространены в разных видах соединительной ткани и выполняют там самые разнообразные функции.
Основной протеогликан хрящевого матрикса называется агрекан, он составляет 10% по весу исходной ткани и 25% сухого веса хрящевого мат-рикса. Это очень большая молекула, в которой к одной полипептидной цепи присоединены до 100 цепей хондроитинсульфатов и около 30 цепей кератансульфатов. По форме молекула агрекана напоминает бутылочный «ёршик» (рис. 15-16).
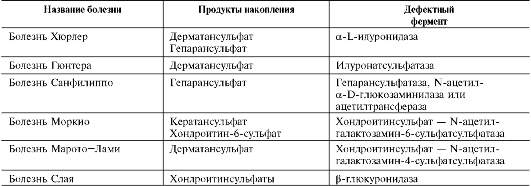
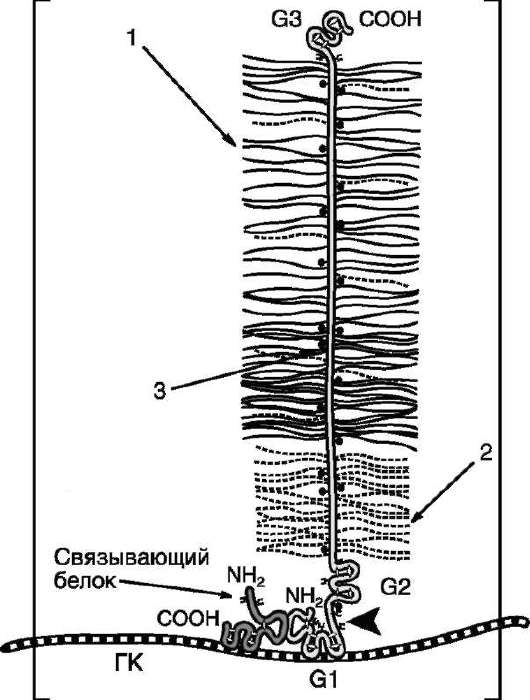
Рис. 15-16. Строение агрекана. ГК - гиалуроновая кислота;
1 - хондроитинсульфат; 2 - кератансульфат; 3 - сердцевинный белок. В центре молекулы находится сердцевинный белок (молекулярная масса ~220 кД), имеющий три глобулярных домена: G1, G2, G3, выполняющих разные функции. N-концевой домен G1 обеспечивает связывание агрекана с гиалуроновой кислотой и низкомолекулярным связывающим белком; функция домена G2 пока неизвестна; С-концевой домен G3 обеспечивает присоединение агрекана к другим молекулам межклеточного матрикса и, возможно, участвует в межклеточных взаимодействиях. Между доменами G2 и G3 находятся области, в которых к белку присоединяются кератансульфаты и хондроитинсуль-фаты. В этих областях в коỷровом белке имеются пептидные участки, состоящие из 6 и 19 аминокислотных остатков, которые повторяются от 10 до 20 раз.
В хрящевой ткани молекулы агрекана собираются в агрегаты с гиалуроновой кислотой и небольшим связывающим белком. Оба компонента присоединяются к агрекану нековалентными связями в области домена G1. Домен G1 взаимодействует примерно с пятью дисахаридными единицами гиалуроновой кислоты, далее этот
комплекс стабилизируется связывающим белком; домен G1 и связывающий белок вместе занимают 25 дисахаридных единиц гиалуроновой кислоты. Конечный агрегат с молекулярной массой более 200х106 Д состоит из одной молекулы гиалуроновой кислоты и ~100 молекул агрекана (и такого же количества связывающего белка). Координация сборки этих агрегатов является центральной функцией хондроцитов. Агрекан и связывающий белок продуцируются этими клетками в необходимых количествах. Эти компоненты могут взаимодействовать друг с другом внутри клетки, но процесс агрегации полностью завершается в межклеточном матриксе. Показано, что гиалуроновая кислота образуется на поверхности хондроцитов специфической синтетазой и «выталкивается» в межклеточное пространство, чтобы связаться с агреканом и связывающим белком. Созревание функционально активного тройного комплекса составляет около 24 ч.
Катаболизм агрекана изучен в настоящее время недостаточно. Имеются данные о наличии в хрящевом межклеточном матриксе фермента агреканазы. Местом действия этого фермента является интерглобулярная область между доменами G1 и G2. Кроме того, в зоне присоединения цепей хондроитинсульфата в коровом белке имеются ещё 3 места протеолитического расщепления агрекана. Конечный продукт расщепления агрекана представляет собой комплекс домена G1, связывающего белка и гиалуроновой кислоты. Он поступает в хондроцит по механизму эндоцитоза и подвергается расщеплению лизосомальными гидроксилазами.
Малые протеогликаны
Малые протеогликаны - протеогликаны с низкой молекулярной массой. Они содержатся в хрящах, сухожилиях, связках, менисках, коже и других видах соединительной ткани.
Эти протеогликаны имеют небольшой ко'ровый белок, к которому присоединены одна или две цепи гликозаминогликанов. Наиболее изучены декорин, бигликан, фибромодулин, люмикан, перлекан.
Кóровые белки бигликана и декорина похожи по размерам и структуре (молекулярная масса 36 000 и 38 000 Д, соответственно). Они имеют несколько тандемных повторов, богатых лейцином, которые образуют α-спирали или β-структуры. На N- и С-концах этих бел-
ков имеются домены, содержащие S-S-связи. Кóровые белки значительно различаются по первичной структуре в N-концевых областях, что определяет различия в присоединении гли-козамино-гликанов. Бигликан содержит серин в положениях 5 и 11, что обеспечивает присоединение двух полисахаридных цепей. Декорин содержит один серин в положении 4, поэтому к нему присоединяется одна полисахаридная цепь. У этих протеогликанов полисахаридные цепи представлены дерматансульфатом с молекулярной массой ~30 000 Д (рис. 15-17).
Ко'ровый белок фибромодулина (молекулярная масса ~40 000 Д) тоже имеет области тандемных повторов, богатые лейцином, но его N-концевая область отличается тем, что не содержит серина, а имеет несколько сульфатированных остатков тирозина, поэтому одна или две цепи кератансуль-
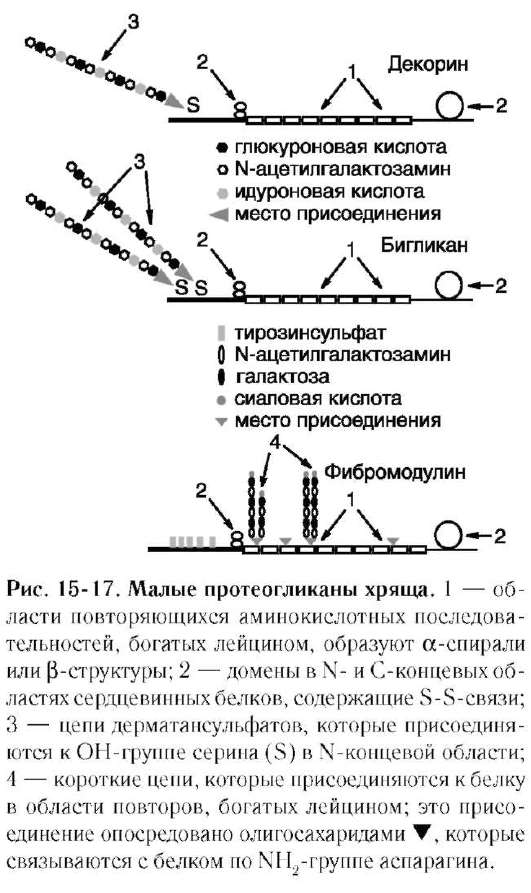
фата присоединяются к кóровому белку фибро-модулина не в N-концевой, а в области, богатой лейцином, через NH2-группу аспарагина.
Малые протеогликаны являются мультифунк-циональными макромолекулами. Они могут связываться с другими компонентами соединительной ткани и оказывать влияние на их строение и функции. Например, декорин и фибромодулин присоединяются к фибриллам коллагена II типа и ограничивают их диаметр (т.е. препятствуют образованию толстых фибрилл). Декорин и биг-ликан, присоединяясь к фибронектину, подавляют клеточную адгезию, а присоединяясь к фактору роста опухолей β, снижают его митогенную активность. Кроме этого, имеется большое количество данных о том, что малые протеогликаны играют важную регуляторную роль в процессах развития и восстановления соединительной ткани.
Протеогликаны базальных мембран
Протеогликаны базальных мембран отличаются значительной гетерогенностью. Это преимущественно гепарансульфатсодержа-щие протеогликаны (ГСПГ), представленные двумя разновидностями: высокой и низкой плотности (рис. 15-18).
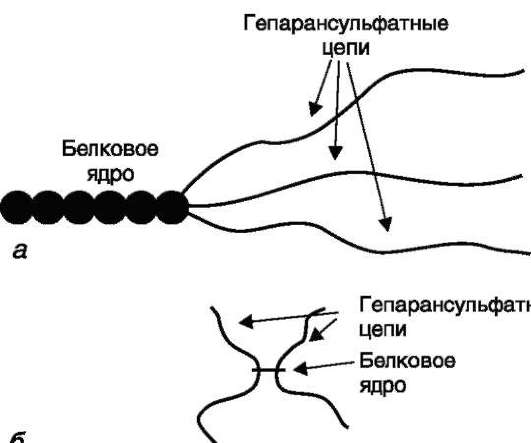
Рис. 15-18. Гепарансульфатсодержащие протеогликаны низкой (А) и высокой (Б) плотности. Гепарансуль-фатсодержащие протеогликаны высокой плотности имеют звёздообразную форму и состоят из четырёх коротких гепарансульфатных цепей, связанных с небольшим белковым ядром. Гепарансульфатсодержа-щие протеогликаны низкой плотности имеют большое многодоменное белковое ядро, представленное одной полипептидной цепью. К одному из полюсов ядра прикреплены три длинные гепарансульфатные цепи.
iv. специализированные белки межклеточного матрикса
Белки межклеточного матрикса выполняют различные функции, но их можно разделить на две большие группы по одному весьма важному признаку: 1) белки, обладающие адгезивными свойствами; 2) белки, подавляющие адгезию клеток.
А. АДГЕЗИВНЫЕ БЕЛКИ
К первой группе белков с выраженными адгезивными свойствами относят фибронектин, ламинин, нидоген, фибриллярные коллагены и коллаген IV типа; их относят к белкам «зрелой» соединительной ткани.
Фибронектин
Фибронектин - один из ключевых белков межклеточного матрикса, неколлагеновый структурный гликопротеин, синтезируемый и выделяемый в межклеточное пространство многими клетками. Он построен из двух идентичных полипептидных цепей, соединённых дисульфидными мостиками у своих С-концов (рис. 15-19).
Полипептидная цепь фибронектина содержит 7-8 доменов, на каждом из которых расположены специфические центры для связывания разных веществ. Фибронектин может связывать коллаген, протеогликаны, гиалуроновую кислоту, углеводы плазматических мембран, гепарин, фермент трансглутаминазу. Благодаря
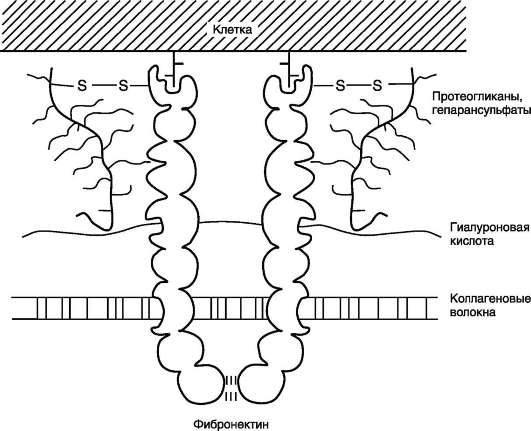
Рис. 15-19. Строение фибронектина.
своей структуре фибронектин может выполнять интегрирующую роль в организации межклеточного вещества, а также способствовать адгезии клеток.
Существует несколько форм фибронектина, которые синтезируются разными клетками. Растворимый, или плазменный, фибронектин синтезируется гепатоцитами. Нерастворимый, или тканевый фибронектин синтезируется в основном фибробластами или эндотелиоцитами, глиоцитами и эпителиальными клетками.
Обе формы фибронектина вовлекаются в разнообразные процессы: способствуют адгезии и распространению эпителиальных и мезенхи-мальных клеток, стимулируют пролиферацию и миграцию эмбриональных и опухолевых клеток, контролируют дифференцировку и поддержание цитоскелета клеток, активно участвуют в воспалительных и репаративных процессах. Это связано с тем, что каждая субъединица фибронектина содержит последовательность Арг-Гли-Асп (RGD), с помощью которой он может присоединяться к клеточным рецепторам (интегринам). Эти рецепторы опосредованно взаимодействуют с актиновыми микрофиламентами, которые находятся в цитозоле. В этом процессе участвуют так называемые белки прикрепления (от англ. attach - прикреплять proteins): талин, винкулин, α-актинин (рис. 15-20).
С помощью таких белок-белковых взаимодействий информация может передаваться из межклеточного матрикса внутрь клетки, а также в обратном направлении - из клетки наружу, таким образом влияя на протекающие в клетке процессы.
Известно также, что фибронектин участвует в миграции клеток, которые могут присоединяться к его RGD-участкам, и, таким образом, фибронектин как бы помогает им перемещаться в межклеточном матриксе.
В межклеточном матриксе, окружающем трансформированные (или опухолевые) клетки, количество фибронектина заметно снижено, что может быть одной из причин появления метастазов.
Ламинин - наиболее распространённый неколлагеновый гликопротеин базальных мембран. Он состоит из трёх полипептидных цепей: А, В1 и В2. Молекула ламинина имеет крестообразную форму с тремя одноцепо-чечными ветвями и одной трёхцепочечной ветвью (рис. 15-21). Каждая цепь ламинина
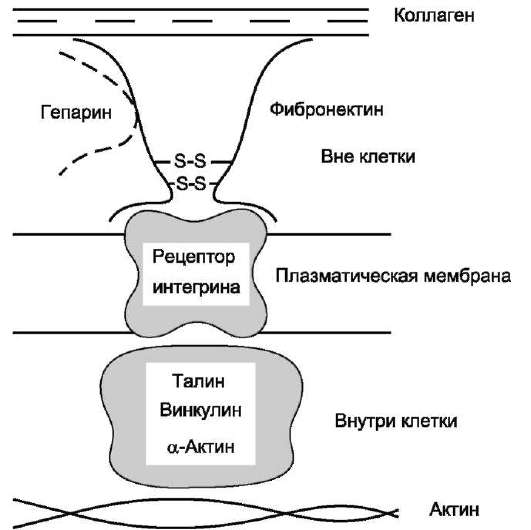
Рис. 15-20. Схема взаимодействия фибронектина интегрином.
содержит несколько глобулярных и стержне-видных доменов, на которых имеются специфические центры связывания для различных веществ. Ламинин взаимодействует со всеми структурными компонентами базальных мембран, включая коллаген IV типа, нидоген, фибронектин, ГСПГ. Кроме того, молекула ламинина имеет несколько центров связывания с клетками. Главные функции ламинина определяются его способностью связывать клетки и модулировать клеточное поведение. Он может влиять на рост, морфологию, дифференцировку и подвижность клеток.
Ламинин выполняет роль адгезивного белка для различных эпителиальных и мезенхималь-ных клеток.
Нидоген - сульфатированный гликопротеин базальных мембран, образует с ламинином плотный, нековалентно связанный комплекс; сила связывания нидогена с коллагеном IV типа гораздо меньше, чем с ламинином. Этот белок представлен одной полипептидной цепью, содержащей три глобулярных домена (рис. 15-21). Один из доменов нидогена имеет центр связывания ламинина, в области другого домена находится центр связывания коллагена IV типа.
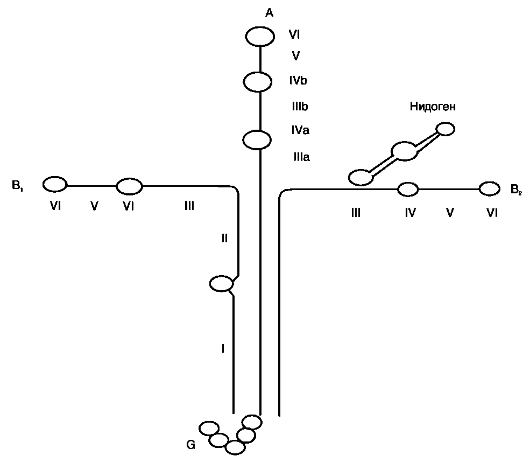
Рис. 15-21. Строение комплекса ламинин-нидоген.
Таким образом, нидоген может выступать в качестве одного из связывающих мостов между различными компонентами межклеточного матрикса и участвовать в образовании тройных комплексов ламинин-нидоген-коллаген. Кроме этого, нидоген содержит RGD-последователь-ность и поэтому может присоединяться к клеточной поверхности.
Б. АНТИАДГЕЗИВНЫЕ БЕЛКИ
Ко второй группе белков, обладающих антиадгезивными свойствами, относят такие гликопро-теины, как остеонектин, тенасцин и тромбоспондин. Эти белки появляются и играют заметную роль в эмбриогенезе и морфогенезе, развитии клеточного ответа на повреждение. Их концентрация в матриксе повышается при некоторых опухолевых заболеваниях.
Остеонектин (синонимы: ВМ-40, SPARC, от англ. secreted protein acidic and rich in cysteine) состоит из 4 доменов, к 2 из которых могут присоединяться ионы Са2+. Остеонектин - кислый
белок, богатый цистеином. Показано, что он может ингибировать G1-S'-фазу роста эндоте-лиальных клеток.
Тенасцин (антиген мышечных сухожилий) - олигомерный гликопротеин, состоящий, подобно фибронектину, из 2 субъединиц, соединённых ди-сульфидной связью. Эту большую молекулу, похожую на осьминога, называют ещё «гексабрахион», так как она имеет 6 «рук», отходящих радиально от одного участка. Благодаря такому строению, тенасцин может взаимодействовать с большим количеством лигандов, к которым относят различные молекулы межклеточного матрикса.
Тенасцин обладает как адгезивными, так и антиадгезивными свойствами, синтезируется в различных тканях эмбриона (наиболее интенсивно - в зонах эпителиально-мезинхимальных контактов и в развивающейся нервной ткани). В зрелых тканях небольшие количества тенасци-на находятся в сухожилиях и хрящах, его синтез увеличивается в заживающих ранах.
Тромбоспондин, как и другие белки межклеточного матрикса, может взаимодействовать со
многими лигандами: коллагеном, фибронекти-ном, ламинином, протеогликанами, ионами Са2+ и др. В клетках роговицы глаза и тромбоцитах тромбоспондин проявляет адгезивные свойства, а в клетках эндотелия и фибробластах он функционирует как антиадгезивный белок.
Таким образом, функции этих белков определяются их локализацией и окружением.
v. структурная организация межклеточного матрикса
Как уже говорилось, межклеточный матрикс представляет собой супрамолекулярный комплекс, образованный сложной сетью связанных между собой макромолекул. В организме человека межклеточный матрикс формирует такие высокоспециализированные структуры, как хрящ, сухожилия, базальные мембраны, а также (при вторичном отложении фосфата кальция) кости и зубы.
Эти структуры различаются между собой как по молекулярному составу, так и по способам организации основных компонентов (белков и полисахаридов) в различных формах межклеточного матрикса.
А. МЕЖКЛЕТОЧНЫЙ МАТРИКС КОСТНОЙ И ЗУБНОЙ ТКАНИ
Костная и зубная ткань - специализированный тип соединительной ткани. Эти ткани выполняют в организме человека следующие важные функции:
• из костей образуется скелет организма;
• кости защищают и поддерживают внутренние органы;
• кости служат местом депонирования кальция и неорганического фосфата;
• костный мозг входит в состав кроветворной и иммунной систем;
• зубы как часть жевательного аппарата входят в состав пищеварительной системы;
• зубы - часть речевого аппарата человека. Замечательным свойством костей является
сочетание в них таких качеств, как высокая прочность на разрыв с очень лёгким весом. Костная и зубная ткань отличаются высокой минерализацией (или кальцификацией) межклеточного матрикса и содержат по массе ~50%
неорганических соединений, 25% органических компонентов и 25% воды.
Неорганическая часть
В состав костей входит 99% всего кальция организма, 87% фосфора, ~60% магния и ~25% натрия. Кальций в костях находится в форме минерала гидроксиапатита, примерный состав которого Ca10(PO4)6(OH)2. Гидроксиапатит образует кристаллы, имеющие обычно размер 20x5x1,5 нм. В костной ткани содержится много микроэлементов, таких как медь, стронций, барий, цинк, фтор и др., которые играют важную роль в обмене веществ в организме. Минеральная часть костей включает также карбонаты, гидроксиды и цитраты.
Минеральный состав зуба различен в разных его частях. Твёрдые части зуба (эмаль, дентин и цемент) содержат от 70% (цемент и дентин) до 96-97% (эмаль) неорганических веществ. Основную часть этих веществ составляют фосфат кальция, входящий в состав кристаллов гидроксиапатита (75%), а также карбонат и фторид кальция.
Мягкие части зуба (пульпа и периодонт) не относят к тканям с высокой степенью минерализации. Пульпа состоит из рыхлой волокнистой соединительной ткани (такая ткань находится практически во всех органах и образует их стро-му, или каркас), а периодонт образован плотной волокнистой соединительной тканью, которая также входит в состав сухожилий и связок.
Органическая часть
Органические вещества костного матрикса представлены белками, липидами и небольшим количеством протеогликанов.
Основной белок костной ткани - коллаген I типа (90-95%). Кроме него, в матриксе костей присутствуют такие белки, как коллаген V типа, остеонектин, остеокальцин, так называемые морфогенетические белки кости (ВМР) и ферменты - щелочная фосфатаза (в остеобластах) и кислая фосфатаза (в остеокластах). Оба эти фермента служат маркёрами соответствующих клеток костной ткани. Углеводная часть про-теогликанов костного матрикса представлена дерматан- и кератансульфатами.
Главный компонент органических веществ зубной ткани - коллаген I типа. Углеводы и
липиды присутствуют в небольших количествах. Содержание органических веществ в твёрдых частях зуба варьирует от 2% (эмаль) до 30% (дентин и цемент). Содержание органических веществ в мягких частях зуба такое же, как в соответствующих видах соединительной ткани.
Минерализация кости
Механизмы минерализации кости пока ещё недостаточно понятны, хотя известно, что в этом процессе важную роль играют все основные компоненты костной ткани, в том числе костеобразу-ющие клетки (остеобласты) и клетки, разрушающие костную ткань (остеокласты). Определяющий фактор минерализации - взаимное расположение молекул тропоколлагена со смещением на 1/4 длины молекулы (см. рис. 15-3). Считается, что промежутки между молекулами тропоколлагена являются центрами минерализации кости, в которых начинается отложение фосфата кальция сначала в аморфном виде с последующим образованием кристаллов гидроксиапатита.
Остеобласты контролируют минерализацию посредством регуляции транспорта ионов кальция и фосфата через свои мембраны. Присутствующая в них щелочная фосфатаза высвобождает неорганический фосфат из органических фосфорсодержащих соединений. Освобождающаяся фосфорная кислота реагирует с солями кальция с образованием Са3(РО4)2. Гликопротеин остеонек-тин имеет высокое сродство к коллагену I типа и к гидроксиапатиту. Он содержит Са2+-связы-вающие домены и способствует осаждению Са2+ и РО43- в присутствии коллагена. Определённую роль в процессе минерализации играют также кислые фосфопротеины, специфичные для костной ткани. Они содержат последовательности поли-Асп и поли-Глу, к которым присоединяется кальций, это может быть пусковым моментом в процессе минерализации.
Костная ткань, несмотря на высокую степень минерализации, находится в динамическом состоянии, в ней постоянно происходят процессы обновления входящих в её состав веществ, адаптационные перестройки к изменяющимся условиям окружающей среды.
Регуляция метаболизма костной ткани
Формирование матрикса кости регулируется биомеханическими, гормональными и другими
факторами. Остеобласты, которые являются клетками-мишенями для паратгормона, реагируют на повышение содержания этого гормона в крови снижением синтеза коллагена, а также повышением активности коллагеназ. Кальцит-риол, как и паратгормон, вызывает резорбцию кости опосредованно через остеобласты, так как остеокласты не имеют к нему рецепторов. По-видимому, стимуляция остеокластов происходит при их контактном взаимодействии с остеобластами или в результате синтеза остеобластами активирующего остеокласты фактора.
Простагландины (А, В, Е1, Е2 и F) и некоторые цитокины (эпидермальный фактор роста, фактор некроза опухолей, ИЛ-1) стимулируют резобцию кости и перестройку костной ткани, воздействуя на остеобласты, которые выделяют фактор, активирующий остеокласты.
Глюкокортикоиды тормозят пролиферацию остеобластов, подавляя в них синтез ДНК, РНК и белков.
Определённую роль в регуляции функционального состояния костной ткани играют половые гормоны. Известно, что в менопаузе у женщин постепенно развивается остеопороз и что предотвратить его можно заместительной терапией эстрогенами, которые, по-видимому, снижают резорбцию кости. Параллельно с этим были получены данные, что эстрогены тормозят также и формирование кости, в результате чего общее количество коллагена не изменяется, а в итоге замедляется смена костной ткани.
Кальцитонин действует непосредственно на остеокласты, которые имеют к нему рецепторы.
Б. МЕЖКЛЕТОЧНЫЙ МАТРИКС СУСТАВНОГО ХРЯЩА
Основные компоненты межклеточного хрящевого матрикса - коллаген II типа, агрекан, гиалуроновая кислота и вода. Кроме них, в мат-риксе находятся малые протеогликаны, коллагены VI, IX, XI типов, связывающий белок, другие неколлагеновые белки (фибронектин, анкорин, хрящевой олигомерный белок, хондроадгерин), разнообразные ростовые факторы. «Эндоскелет» хрящевого матрикса образован фибриллярной сетью, которая состоит из коллагенов II, IX и XI типов и придаёт хрящу прочность (рис. 15-22).
Коллаген XI типа находится внутри фибрилл, образованных коллагеном II типа, он играет
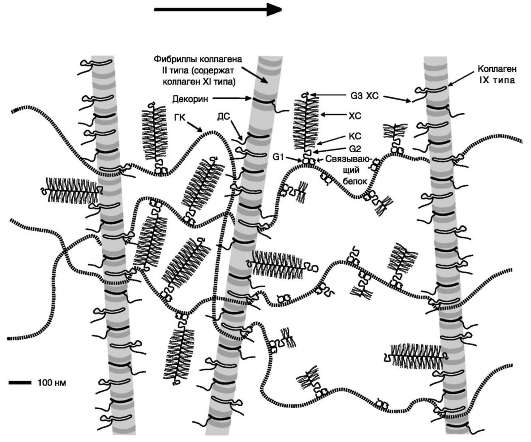
Рис. 15-22. Организация межклеточного матрикса в суставном хряще. ГК - гиалуроновая кислота; ДС дерматансульфат; ХС - хондроитинсульфат.
определённую роль в сборке этих фибрилл. Коллаген IX типа антипараллельно присоединяется к фибриллам коллагена II типа. Его глобулярный НК4-домен - основный, он не связан с фибриллами коллагена II типа, и поэтому к нему может присоединяться такой компонент матрикса, как гиалуроновая кислота. Микрофибриллы, которые образуются тетрамерами коллагена VI типа, присоединяются к фибриллам коллагена II типа и к гиалуроновой кислоте. Кроме того, они могут присоединяться к клеткам, поэтому коллаген VI типа называют «мостовой» молекулой между поверхностью клетки и фибриллами коллагена во внеклеточном матриксе.
Высокомолекулярные агрегаты, состоящие из агрекана и гиалуроновой кислоты, являются полианионами, так как содержат большое количество кислых групп. Это способствует высокой гидратации хрящевого матрикса и обеспечивает выполнение им рессорных функций. Содержание воды в суставном хряще непостоянно: при нагрузке жидкость вытесняется, пока давление набухания не уравновесит внешнюю нагрузку. Когда нагрузка прекращается, вода вновь возвращается в хрящ (рис. 15-23). Очень наглядно это проявляется в межпозвоночных дисках. Утром, после ночного сна, на долю воды приходится около 75% массы диска. При внешней нагрузке на диски в течение дня содержание воды
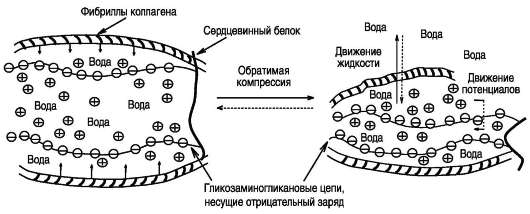
Рис. 15-23. Изменения, происходящие в суставном хряще при его сжатии и при снятии нагрузки. Е - ионы Na+, K+, Ca2+.
уменьшается
примерно на 20%. Вследствие этого рост человека к вечеру на 1-2 см
меньше, чем утром. У космонавтов в условиях невесомости отмечается
увеличение роста даже на
Малые протеогликаны, например, декорин, присоединяются к фибриллам коллагена II типа; они влияют на фибриллогенез, так как ограничивают диаметр этих фибрилл.
Важную роль в организации хрящевого межклеточного матрикса играет также фибро-нектин. Биологическое значение этих и других минорных компонентов хрящевого матрикса заключается в том, что они участвуют в сборке и организации высокомолекулярных компонентов межклеточного вещества и в регуляции функции хондроцитов.
В. МЕЖКЛЕТОЧНЫЙ МАТРИКС КОЖНОЙ ТКАНИ
Основной организующий компонент матрик-са кожной ткани - коллаген VII типа. Пучки фибрилл, образованные димерами этого коллагена, своими С-концами могут присоединяться к lamina densa базальной мембраны (как бы «за-якориваться» в ней) и образовывать петли в субэпидермисе. Такие «заякоренные» фибриллы могут соединять lamina densa базальной мембраны с «якорными дисками», которые находятся в более глубоких субэпителиальных слоях и по своему составу похожи на базальные мембраны (содержат коллаген IV типа). «Заякоренные» фибриллы также захватывают фибриллы коллагена I и III типов (рис. 15-24).
Таким способом «заякоренные» фибриллы коллагена VII типа обеспечивают присоединение эпидермиса к дерме.
Г. БАЗАЛЬНЫЕ МЕМБРАНЫ
Базальные мембраны - специализированная форма межклеточного матрикса. Они синтезируются различными клетками: эндотелиальны-ми, эпителиальными, мышечными, нервными, жировыми. Базальные мембраны представляют собой тонкие слои, которые обычно отделяют клетки и клеточные слои от окружающей соединительной ткани. Например, они окружают отдельные мышечные волокна, жировые и шванновские клетки. В таких структурах, как почечные клубочки и лёгочные альвеолы, ба-зальные мембраны расположены между двумя различными слоями клеток и играют роль высокоселективного фильтрационного барьера.
С помощью электронной микроскопии выявлена двухслойная структура базальных мембран: lamina rara, которая находится со стороны клеточной мембраны, и lamina densa, которая соединена с подлежащей соединительной тканью (рис. 15-25). Основными компонентами базальных мембран являются коллаген IV типа, ламинин, гепарансульфатсодержащие протеог-ликаны (ГСПГ).
Нерастворимость и механическую стабильность базальных мембран обеспечивают молекулы коллагена IV типа, которые организуются в специальную опорную сеть. Эта эластичная
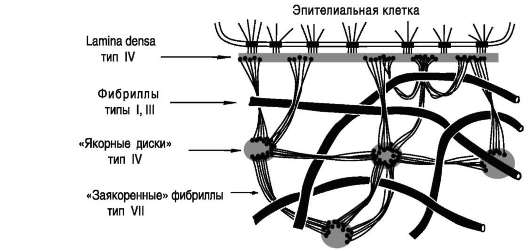
Рис. 15-24. Организация «заякоренных» фибрилл в субэпителиальных слоях.
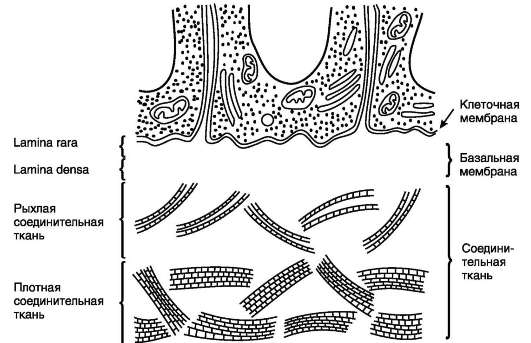
Рис. 15-25. Строение типичной базальной мембраны.
трёхмерная сеть образует структурнsй остов, к которому прикрепляются другие компоненты базальных мембран.
Ламинин взаимодействует практически со всеми структурными компонентами базаль-
ных мембран: коллагеном IV типа, нидогеном, ГСПГ.
Нидоген формирует с ламинином нековален-тно связанный комплекс. Кроме этого, нидоген имеет центр связывания коллагена IV типа и,
таким образом, может играть роль «мостовой» молекулы между различными компонентами базальной мембраны.
ГСПГ базальных мембран могут образовывать олигомеры, соединяясь концевыми доменами белкового ядра, а также связываться с ламини-ном и коллагеном IV типа.
Базальные мембраны выполняют разнообразные и сложные функции. В почечных клубочках базальная мембрана служит полупроницаемым фильтром, препятствующим переходу макро-
молекул из плазмы в первичную мочу. Большое значение в этом процессе имеет высокий отрицательный заряд протеогликанов, который препятствует прохождению через базальную мембрану других отрицательно заряженных молекул (например, белков), а также отрицательно заряженных эритроцитов. Кроме этого, базаль-ные мембраны играют важную роль в прикреплении и ориентации клеток в пространстве, в процессах эмбрионального развития и тканевой регенерации.