Биохимия: учебник для вузов/ под ред. Е.С.Северина - 5-е изд., - 2009. - 768 с.
|
|
|
|
РАЗДЕЛ 7 ОБМЕН УГЛЕВОДОВ
Углеводы входят в состав живых организмов и вместе с белками, липидами и нуклеиновыми кислотами определяют специфичность их строения и функционирования. К углеводам относят соединения, обладающие разнообразными и зачастую сильно отличающимися функциями. Углеводы участвуют во многих метаболических процессах, но прежде всего они являются основными поставщиками энергии. На долю углеводов приходится примерно 75% массы пищевого суточного рациона и более 50% от суточного количества необходимых калорий. Однако неправильно сводить функцию углеводов только к энергетическому обеспечению процессов жизнедеятельности организма. Следует отметить и структурную роль углеводов. Так, в виде гликозаминогликанов углеводы входят в состав межклеточного матрикса. Большое число белков (ферменты, белки-транспортёры, белки-рецепторы, гормоны) - гликопротеины, углеводная составляющая которых повышает их специфичность. Например, различия в строении олигосахаридных фрагментов клеточной оболочки эритроцитов обеспечивают групповую принадлежность крови. Из углеводов в процессе метаболизма образуется большое число органических соединений, которые служат исходными субстратами для синтеза липидов, аминокислот, нуклеотидов. Производные углеводов - глюкурониды - участвуют в детоксикации ксенобиотиков и инактивации веществ эндогенного происхождения. Углеводы могут быть синтезированы в организме с использованием других метаболитов: некоторых аминокислот, глицерина, молочной кислоты. Углеводы нельзя считать незаменимыми компонентами пищи. Однако если исключить углеводы из диеты, то следствием может быть гипогликемия, для компенсации которой будут расходоваться белки и липиды. Таким образом, углеводы - обязательные пищевые компоненты, потому что помимо их основной энергетической функции (кле-
точные «дрова») углеводы участвуют во многих метаболических клеточных процессах.
I. СТРОЕНИЕ УГЛЕВОДОВ
Термин «углеводы», предложенный в XIX столетии, был основан на предположении, что все углеводы содержат 2 компонента - углерод и воду, и их элементарный состав можно выразить общей формулой Сm(Н20)п. Хотя из этого правила есть исключения и оно не абсолютно точно, тем не менее указанное определение позволяет наиболее просто характеризовать класс углеводов в целом. К тому же попытка, предпринятая Комиссией по химической номенклатуре, заменить термин «углеводы» на «глициды» не удалась. Новый термин не получил широкого признания. Термин «углеводы» укоренился и общепризнан.
Углеводы можно разделить на 3 основные группы в зависимости от количества составляющих их мономеров: моносахариды, олигосаха-риды и полисахариды.
А. МОНОСАХАРИДЫ
Моносахариды - производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от положения в молекуле карбонильной группы моносахариды подразделяют на альдозы и кетозы.
Альдозы содержат функциональную альдегидную группу -НС=О, тогда как кетозы содержат кетонную группу >C=0. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, аль-догексозы, кетогексозы и т.д.
Моносахариды по строению можно отнести к простым углеводам, так как они не гидро-лизуются при переваривании, в отличие от сложных, которые при гидролизе распадаются с образованием простых углеводов. Строение основных представителей моносахаридов показано на рис. 7-1.
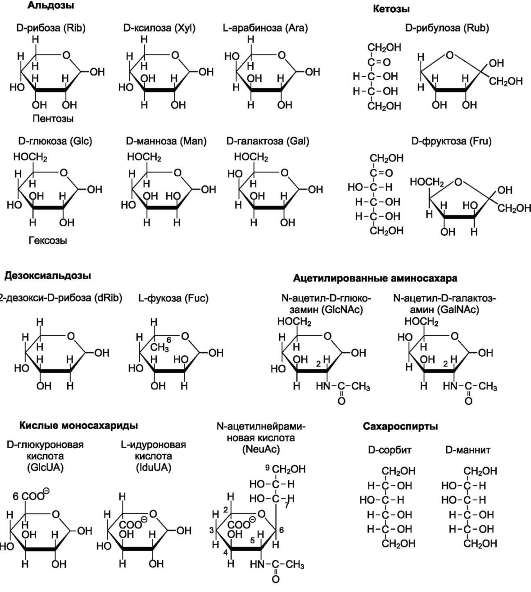
Рис. 7-1. Важнейшие моносахариды.
В пище человека (фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза.
Глюкоза является альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы, предпочтительная в термодинамическом отношении, обусловливает химические свойства глюкозы. Как и все гексозы, глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие стереоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. Эти типы изомеров зеркально отображают друг друга (рис. 7-2).
Расположение Н- и ОН-групп относительно пятого углеродного атома определяет принадлежность глюкозы к D- или L-ряду. В организме млекопитающих моносахариды находятся в D- конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), называемые
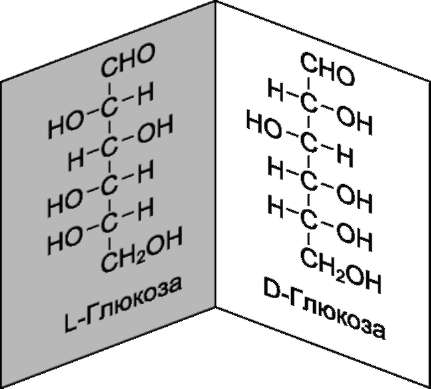
Рис. 7-2. D- и L-изомеры глюкозы.
аномерами, обозначающие определённую кон-формацию Н- и ОН-групп относительно С1 (рис. 7-3). У α-D-глюкозы ОН-группа располагается ниже плоскости кольца, а у β-D-глюкозы, наоборот, над плоскостью кольца.
Фруктоза является кетогексозой (кетогруп-па находится у второго углеродного атома). Фруктоза так же, как и глюкоза, существует в циклической форме, образуя α- и β-аномеры (рис. 7-4).
Б. РЕАКЦИИ МОНОСАХАРИДОВ
Присутствие гидроксильных, альдегидных и кетонных групп позволяет моносахаридам вступать в реакции, характерные для спиртов, альдегидов или кетонов. Эти реакции довольно многочисленны. В данном разделе будут описаны лишь некоторые из них, причём в основном имеющие наибольшее биологическое значение.
В этом разделе основные реакции моносахаридов рассмотрены на примере D-глюкозы (рис. 7-5), хотя надо иметь в виду, что в метаболизме углеводов принимают участие и другие моносахариды, а также их производные.
Мутаротация, или аномеризация - взаимопревращение аномерных форм моносахаридов. α- и β-формы аномеров находятся в растворе в
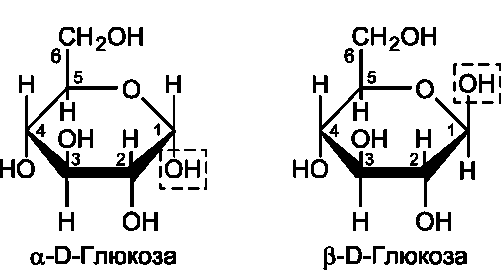
Рис. 7-3. α- и β-аномеры D-глюкозы.
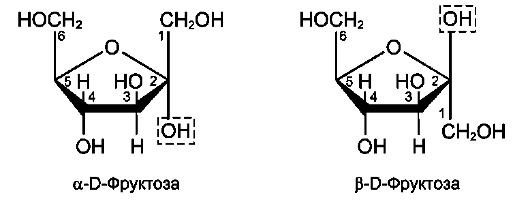
Рис. 7-4.α- и β-аномеры D-фруктозы.
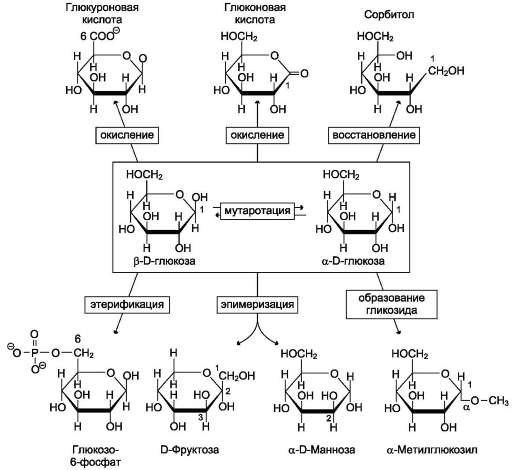
Рис. 7-5. Реакции моносахаридов.
состоянии равновесия. При достижении этого равновесия происходит мутаротация - размыкание и замыкание пиранового кольца и, соответственно, изменение расположения Н- и ОН-групп при первом углероде моносахарида.
Образование гликозидов. Гликозидная связь имеет важное биологическое значение, потому что именно с помощью этой связи осуществляется ковалентное связывание моносахаридов в составе олиго- и полисахаридов. При образовании гликозидной связи аномерная ОН-группа одного моносахарида взаимодействует с ОН-группой другого моносахарида или спирта. При этом происходят отщепление молекулы воды и образование О-гликозидной связи. Все линейные олигомеры (кроме дисахаридов) или полимеры
содержат мономерные остатки, участвующие в образовании двух гликозидных связей, кроме концевых остатков, образующих только одну гликозидную связь. Некоторые гликозидные остатки могут образовывать три гликозидные связи, что характерно для разветвлённых оли-го- и полисахаридов. Олиго- и полисахариды могут иметь концевой остаток моносахарида со свободной аномерной ОН-группой, не использованной при образовании гликозидной связи. В этом случае при размыкании цикла возможно образование свободной карбонильной группы, способной окисляться. Такие олиго- и полисахариды обладают восстанавливающими свойствами и поэтому называются восстанавливающими, или редуцирующими (рис. 7-6).
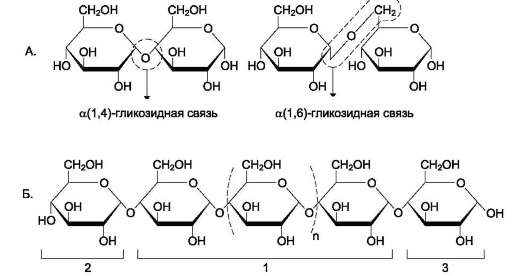
Рис. 7-6. Строение полисахарида. А. Образование α-1,4- и α-1,6-гликозидных связей. Б. Строение линейного полисахарида: 1 - α-1,4-гликозидные связи между мономерами; 2 - невосстанавливающий конец (невозможно образование свободной карбонильной группы у аномерного углерода); 3 - восстанавливающий конец (возможно размыкание цикла с образованием свободной карбонильной группы у аномерного углерода).
Аномерная ОН-группа моносахарида может взаимодействовать с NH2-группой других соединений, что приводит к образованию N-гли-козидной связи. Подобная связь присутствует в нуклеотидах и гликопротеинах (рис. 7-7).
Этерификация. Это реакция образования эфирной связи между ОН-группами моносахаридов и различными кислотами. В метаболизме углеводов важную роль играют фосфоэфиры - эфиры моносахаридов и фосфорной кислоты.В метаболизме глюкозы особое место занимает глюко-зо-6-фосфат. Образование глюкозо-6-фосфата происходит в ходе АТФ-зависимой реакции при участии ферментов, относящихся к группе ки-наз. АТФ в данной реакции выступает как донор фосфатной группы. Фосфоэфиры моносахаридов могут образовываться и без использования АТФ. Например, глюкозо-1-фосфат образуется из гликогена при участии Н3РО4. Физиологическое значение фосфоэфиров моносахаридов заключается в том, что они представляют собой метаболически активные структуры. Реакция фосфорилирования моносахаридов важна для метаболизма ещё и потому, что клеточная мембрана мало проницаема для этих соединений, т.е. клетка удерживает моносахариды благодаря тому, что они находятся в фосфорилированной форме.
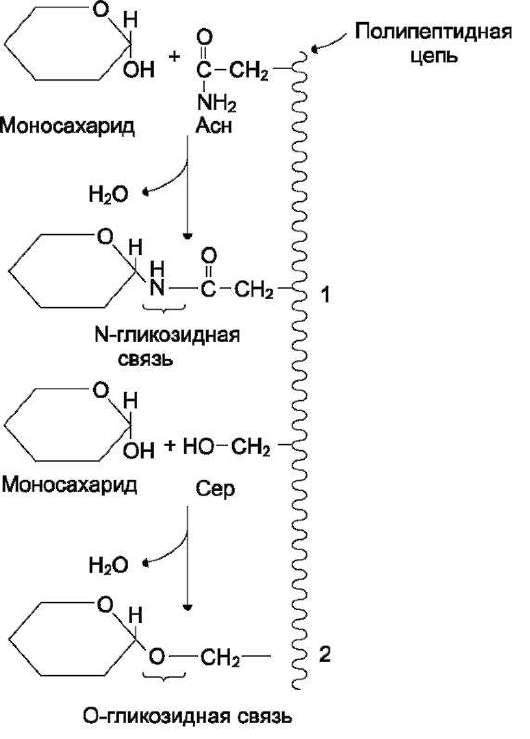
Рис. 7-7. Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.
Окисление и восстановление. При окислении концевых групп глюкозы -СНО и -СН2ОН образуются 3 различных производных. При окислении группы -СНО образуется глюконовая кислота. Если окислению подвергается концевая группа -СН2ОН, образуется глюкуроновая кислота. А если окисляются обе концевые группы, то образуется сахарная кислота, содержащая 2 карбоксильные группы. Восстановление первого углерода приводит к образованию сахароспир-та - сорбитола.
В. ОЛИГОСАХАРИДЫ
Олигосахариды содержат несколько (от двух до десяти) остатков моносахаридов, соединённых гликозидной связью. Дисахариды - наиболее распространённые олигомерные углеводы, встречающиеся в свободной форме, т.е. не связанной с другими соединениями. По химической природе дисахариды представляют собой гликозиды, которые содержат 2 моносахарида, соединённые гликозидной связью в α- или β-конфигурации. В пище содержатся в основном такие дисахариды, как сахароза, лактоза и мальтоза (рис. 7-8).
Сахароза - дисахарид, состоящий из α-D-глюкозы и β-D-фруктозы, соединённых α,β-1,2-гликозидной связью. В сахарозе обе аномерные ОН-группы остатков глюкозы и фруктозы участвуют в образовании гликозидной связи. Следовательно, сахароза не относится к восстанавливающим сахарам. Сахароза - растворимый дисахарид со сладким вкусом. Источником сахарозы служат растения, особенно сахарная свёкла, сахарный тростник. Последнее объяс-
няет возникновение тривиального названия сахарозы - «тростниковый сахар».
Лактоза - молочный сахар; важнейший ди-сахарид молока млекопитающих. В коровьем молоке содержится до 5% лактозы, в женском молоке - до 8%. В лактозе аномерная ОН-груп-па первого углеродного атома остатка D-галак-тозы связана β-гликозидной связью с четвёртым углеродным атомом D-глюкозы (β-1,4-связь). Поскольку аномерный атом углерода остатка глюкозы не участвует в образовании гликозид-ной связи, следовательно, лактоза относится к восстанавливающим сахарам.
Мальтоза поступает с продуктами, содержащими частично гидролизованный крахмал, например, солод, пиво. Мальтоза также образуется при расщеплении крахмала в кишечнике. Мальтоза состоит из двух остатков D-глюкозы, соединённых α-1,4-гликозидной связью.
Изомальтоза - промежуточный продукт, образующийся при расщеплении крахмала в кишечнике. Состоит из двух остатков D-глюкозы, но соединены эти моносахариды α-1,6-глико-зидной связью.
Г. ПОЛИСАХАРИДЫ
Структурные различия между полисахаридами определяются:
• строением моносахаридов, составляющих цепь;
• типом гликозидных связей, соединяющих мономеры в цепи;
• последовательностью остатков моносахаридов в цепи.
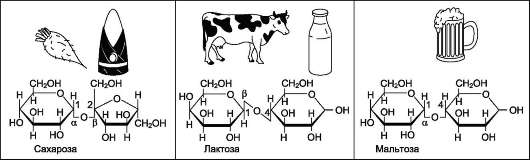
Рис. 7-8. Дисахариды пищи.
В зависимости от строения остатков моносахаридов полисахариды можно разделить на гомополисахариды (все мономеры идентичны) и гетерополисахариды (мономеры различны). Оба типа полисахаридов могут иметь как линейное расположение мономеров, так и разветвлённое.
В зависимости от выполняемых ими функций полисахариды можно разделить на 3 основные группы:
• резервные полисахариды, выполняющие энергетическую функцию. Эти полисахариды служат источником глюкозы, используемым организмом по мере необходимости. Резервная функция этих углеводов обеспечивается их полимерной природой. Полисахариды менее растворимы, чем моносахариды, следовательно, они не влияют на осмотическое давление и поэтому могут накапливаться в клетке, например, крахмал - в клетках растений, гликоген - в клетках животных;
• структурные полисахариды, обеспечивающие клеткам и органам механическую прочность (см. раздел 15);
• полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, а также в пролиферации и дифференцировке клеток. Полисахариды межклеточного матрикса водорастворимы и сильно гидратированы (см. раздел 15).
В пище человека в основном содержатся полисахариды растительного происхождения - крахмал, целлюлоза. В меньшем количестве с пищей поступает полисахарид животных - гликоген.
Крахмал - наиболее важный углеводный компонент пищевого рациона. Это резервный полисахарид растений, содержащийся в наибольшем количестве (до 45% от массы сухого вещества) в зёрнах злаков (пшеница, кукуруза, рис и др.), а также луковицах, стеблях и клубнях растений (в картофеле примерно 65%). Крахмал - разветвлённый полисахарид, состоящий из остатков глюкозы (гомогликан). Он находится в клетках растений в виде гранул, практически нерастворим в воде.
Крахмал состоит из амилозы и амилопектина (рис. 7-9). Амилоза - неразветвлённый полисахарид, включающий 200-300 остатков глюкозы, связанных а-1,4-гликозидной связью. Благодаря α-конфигурации глюкозного остатка, полиса-харидная цепь имеет конформацию спирали. Синяя окраска при добавлении йода к раствору крахмала обусловлена наличием такой спирали. Амилопектин имеет разветвлённую структуру. В местах ветвления остатки глюкозы соединены а-1,6-гликозидными связями. Линейные участки содержат примерно 20-25 остатков глюкозы. При этом формируется древовидная структура, в которой имеется лишь одна ано-мерная ОН-группа. Крахмал - высокомолекулярное соединение, включающее сотни тысяч остатков глюкозы. Его молекулярная масса составляет порядка 105-108 Д.
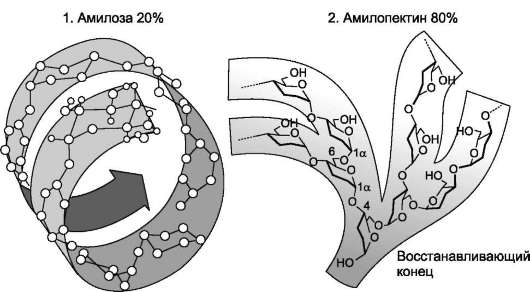
Рис. 7-9. Строение крахмала.
Целлюлоза (клетчатка) - основной структурный полисахарид растений. Это самое распространённое органическое соединение на земле. Доля целлюлозы в клеточных стенках растений составляет 40-50%. Целлюлоза имеет молекулярную массу порядка 106 Д, длина молекулы может доходить до 6-8 мкм.
Целлюлоза - линейный полисахарид гомогли-кан, построенный из остатков глюкозы, соединённых между собой β-1,4-гликозидными связями. Пищеварительная система человека не имеет ферментов, гидролизующих β-связи в полисахаридах. Поэтому целлюлоза - неиспользуемый углевод, но этот пищевой компонент необходим для нормального протекания переваривания.
Гликоген - полисахарид животных и человека. Так же, как крахмал в растениях, гликоген в клетках животных выполняет резервную функцию, но, так как в пище содержится лишь небольшое количество гликогена, он не имеет пищевого значения.
Гликоген представляет собой структурный аналог крахмала, но имеет большую степень ветвления: примерно на каждые 10 остатков глюкозы приходится одна а-1,6-гликозидная связь.
ii. переваривание углеводов
Эпителиальные клетки кишечника способны всасывать только моносахариды. Поэтому процесс переваривания заключается в ферментативном гидролизе гликозидных связей в углеводах, имеющих олигоили полисахаридное строение (рис. 7-10).
А. ПЕРЕВАРИВАНИЕ УГЛЕВОДОВ В РОТОВОЙ ПОЛОСТИ
В ротовой полости пища измельчается при пережёвывании, смачиваясь при этом слюной. Слюна на 99% состоит из воды и обычно имеет рН 6,8. В слюне присутствует гидролитический фермент α-амилаза (α-1,4-гликозидаза), расщепляющая в крахмале α-1,4-гликозидные связи. В ротовой полости не может происходить полное расщепление крахмала, так как действие фермента на крахмал кратковременно. Кроме того, амилаза слюны не расщепляет а-1,6-гли-козидные связи (связи в местах разветвлений), поэтому крахмал переваривается лишь частично
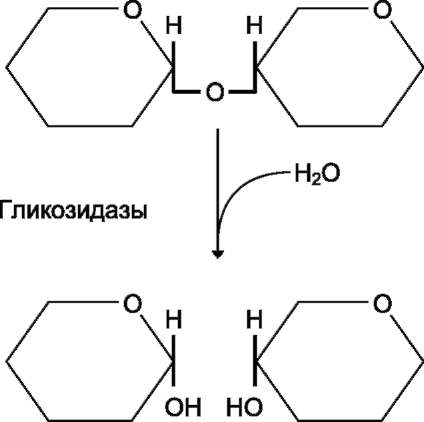
Рис. 7-10. Гидролиз гликозидной связи.
с образованием крупных фрагментов - декстринов и небольшого количества мальтозы. Следует отметить, что амилаза слюны не гидролизует гликозидные связи в дисахаридах.
Действие амилазы слюны прекращается в резко кислой среде содержимого желудка (рН 1,5-2,5). Однако внутри пищевого комка активность амилазы может некоторое время сохраняться, пока рН не изменится в кислую сторону. Желудочный сок не содержит ферментов, расщепляющих углеводы. В желудочном содержимом возможен лишь незначительный кислотный гидролиз гликозидных связей.
Б. ПЕРЕВАРИВАНИЕ УГЛЕВОДОВ В КИШЕЧНИКЕ
Последующие этапы переваривания нерасщеп-лённого или частично расщеплённого крахмала, а также других углеводов пищи происходит в тонком кишечнике в разных его отделах под действием гидролитических ферментов - гликозидаз.
Панкреатическая а-амилаза
В двенадцатиперстной кишке рН среды желудочного содержимого нейтрализуется, так как секрет поджелудочной железы имеет рН 7,5-8,0 и содержит бикарбонаты (НСО3-). С секретом поджелудочной железы в кишечник поступает панкреатическая α-амилаза. Этот фермент гидро-лизует а-1,4-гликозидные связи в крахмале и декстринах.
Продукты переваривания крахмала на этом этапе - дисахарид мальтоза, содержащая 2 остатка глюкозы, связанные а-1,4-связью. Из тех
остатков глюкозы, которые в молекуле крахмала находятся в местах разветвления и соединены α-1,6-гликозидной связью, образуется дисахарид изомальтоза. Кроме того, образуются олигоса-хариды, содержащие 3-8 остатков глюкозы, связанные α-1,4- и α-1,6-связями (рис. 7-11).
а-Амилаза поджелудочной железы, так же, как а-амилаза слюны, действует как эндогликозида-за. Панкреатическая а-амилаза не расщепляет α-1,6-гликозидные связи в крахмале. Этот фермент также не гидролизует β-1,4-гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Целлюлоза, таким образом, проходит через кишечник неизменённой. Тем не менее непереваренная целлюлоза выполняет важную функцию балластного вещества, придавая пище дополнительный объём и положительно влияя на процесс переваривания. Кроме того, в толстом кишечнике целлюлоза может подвергаться действию бактериальных ферментов и частично расщепляться с образованием спиртов, органических кислот и СО2. Продукты бактериального расщепления целлюлозы важны как стимуляторы перистальтики кишечника.
Мальтоза, изомальтоза и триозосахариды, образующиеся в верхних отделах кишечника из крахмала, - промежуточные продукты. Дальнейшее их переваривание происходит под действием специфических ферментов в тонком кишечнике. Дисахариды пищи сахароза и лактоза также гидролизуются специфическими дисахаридазами в тонком кишечнике.
Особенность переваривания углеводов в тонком кишечнике заключается в том, что ак-
тивность специфических олиго- и дисахаридаз в просвете кишечника низкая. Но ферменты активно действуют на поверхности эпителиальных клеток кишечника.
Тонкий кишечник изнутри имеет форму пальцеобразных выростов - ворсинок, покрытых эпителиальными клетками. Эпителиальные клетки, в свою очередь, покрыты микроворсинками, обращёнными в просвет кишечника. Эти клетки вместе с ворсинками образуют щёточную каёмку, благодаря которой увеличивается поверхность контакта гидролитических ферментов и их субстратов в содержимом кишечника. На 1 мм2 поверхности тонкой кишки у человека приходится 80-140 млн ворсинок.
Ферменты, расщепляющие гликозидные связи в дисахаридах (дисахаридазы), образуют ферментативные комплексы, локализованные на наружной поверхности цитоплазматической мембраны энтероцитов.
Сахаразо-изомальтазный комплекс
Этот ферментативный комплекс состоит из двух полипептидных цепей и имеет доменное строение. Сахаразо-изомальтазный комплекс прикрепляется к мембране микроворсинок кишечника с помощью гидрофобного (трансмембранного) домена, образованного N-концевой частью полипептида. Каталитический центр выступает в просвет кишечника (рис. 7-12). Связь этого пищеварительного фермента с мембраной способствует эффективному поглощению продуктов гидролиза клеткой.
Сахаразо-изомальтазный комплекс гидроли-зует сахарозу и изомальтозу, расщепляя а-1,2- и
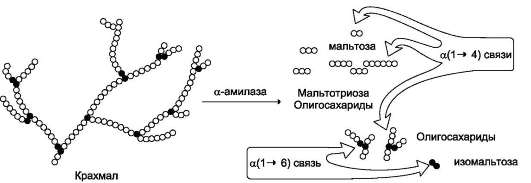
Рис. 7-11. Гидролиз крахмала панкреатической a-амилазой.
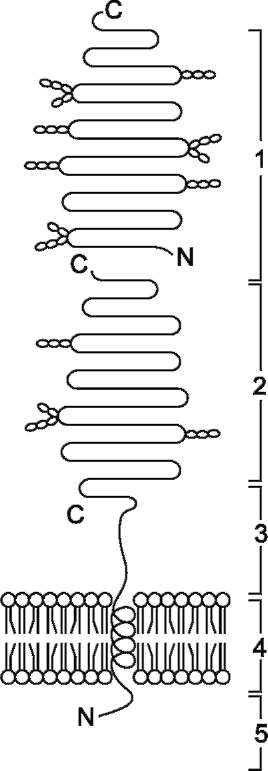
Рис. 7-12. Сахаразо-изомальтазный комплекс.
1 - сахараза; 2 - изомальтаза; 3 - связывающий домен; 4 - трансмембранный домен; 5 - цитоплаз-матический домен.
α-1,6-гликозидные связи. Кроме того, оба ферментных домена имеют мальтазную и мальтотри-азную активности, гидролизуя α-1,4-гликозиднь1е связи в мальтозе и мальтотриозе (трисахарид, образующийся из крахмала). На долю сахара-зо-изомальтазного комплекса приходится 80% от всей мальтазной активности кишечника. Но несмотря на присущую ему высокую мальтазную активность, этот ферментативный комплекс назван в соответствии с основной специфичностью. К тому же сахаразная субъединица - единственный фермент в кишечнике, гидролизующий сахарозу. Изомальтазная субъединица с большей скоростью гидролизует гликозидные связи в изомальтозе, чем в мальтозе и мальтотриозе (рис. 7-13, 7-14).
В тощей кишке содержание сахаразо-изомаль-тазного ферментативного комплекса достаточно высокое, но оно снижается в проксимальной и дистальной частях кишечника.
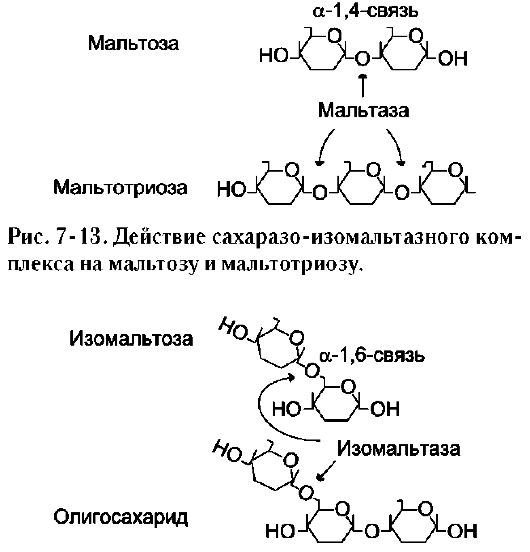
Рис. 7-14. Действие сахаразо-изомальтазного комплекса на изомальтозу и олигосахарид.
Гликоамилазный комплекс
Этот ферментативный комплекс катализирует гидролиз а-1,4-связи между глюкозными остатками в олигосахаридах, действуя с восстанавливающего конца. По механизму действия этот фермент относят к экзогликозидазам. Комплекс расщепляет также связи в мальтозе, действуя как мальтаза. В гликоамилазный комплекс входят две разные каталитические субъединицы, имеющие небольшие различия в субстратной специфичности. Гликоамилазная активность комплекса наибольшая в нижних отделах тонкого кишечника.
β-Гликозидазный комплекс (лактаза)
Лактаза расщепляет β-1,4-гликозидные связи между галактозой и глюкозой в лактозе (рис. 7-15).
Этот ферментативный комплекс по химической природе является гликопротеином. Лактаза, как и другие гликозидазные комплексы, связана с щёточной каемкой и распределена неравномерно по всему тонкому кишечнику. Активность лак-тазы колеблется в зависимости от возраста. Так, активность лактазы у плода особенно повышена
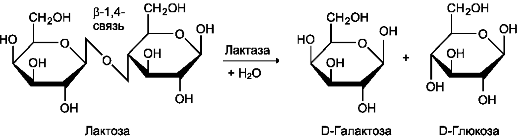
Рис. 7-15. Действие лактазы.
в поздние сроки беременности и сохраняется на высоком уровне до 5-7-летнего возраста. Затем активность фермента снижается, составляя у взрослых 10% от уровня активности, характерного для детей.
Трегалаза - также гликозидазный комплекс, гидролизующий связи между мономерами в трегалозе - дисахариде, содержащемся в грибах. Трегалоза состоит из двух глюкозных остатков, связанных гликозидной связью между первыми аномерными атомами углерода (рис. 7-16).
Совместное действие всех перечисленных ферментов завершает переваривание пищевых олиго- и полисахаридов с образованием моносахаридов, основной из которых - глюкоза. Кроме глюкозы, из углеводов пищи также образуются фруктоза и галактоза, в меньшем количестве - манноза, ксилоза, арабиноза. Общая схема переваривания углеводов представлена на рис. 7-17.
III. МЕХАНИЗМ ТРАНСМЕМБРАННОГО ПЕРЕНОСА ГЛЮКОЗЫ И ДРУГИХ МОНОСАХАРИДОВ В КЛЕТКИ
Моносахариды, образовавшиеся в результате переваривания, всасываются эпителиальными клетками тощей и подвздошной кишок с помощью специальных механизмов транспорта через мембраны этих клеток.
А. ВСАСЫВАНИЕ МОНОСАХАРИДОВ В КИШЕЧНИКЕ
Транспорт моносахаридов в клетки слизистой оболочки кишечника может осуществляться разными способами: путём облегчённой диффузии и активного транспорта. В случае активного транспорта глюкоза и Na+ проходят через мембраны с люминальной стороны, связываясь с разными
Рис. 7-16. Строение трегалозы.
участками белка-переносчика. При этом поступает в клетку по градиенту концентрации, и одновременно глюкоза транспортируется против градиента концентрации (вторично-активный транспорт, см. раздел 5). Следовательно, чем больше градиент Na+, тем больше поступление глюкозы в энтероциты. Если концентрация Na+ во внеклеточной жидкости уменьшается, транспорт глюкозы снижается. Градиент концентрации Na+, являющийся движущей силой активного симпорта, создаётся работой Na+,K+-АТФ-азы. Перенос в клетки слизистой оболочки кишечника по механизму вторично-активного транспорта характерен также для галактозы.
При разной концентрации глюкозы в просвете кишечника «работают» различные механизмы транспорта. Благодаря активному транспорту эпителиальные клетки кишечника могут поглощать глюкозу при её очень низкой концентрации в просвете кишечника. Если же концентрация глюкозы в просвете кишечника велика, то она может транспортироваться в клетку путём облегчённой диффузии. Таким же способом может всасываться и фруктоза. Следует отметить, что скорость всасывания глюкозы и галактозы гораздо выше, чем других моносахаридов. Способы транспорта моносахаридов через мембрану эпителиальных клеток кишечника представлены на рис. 7-18.
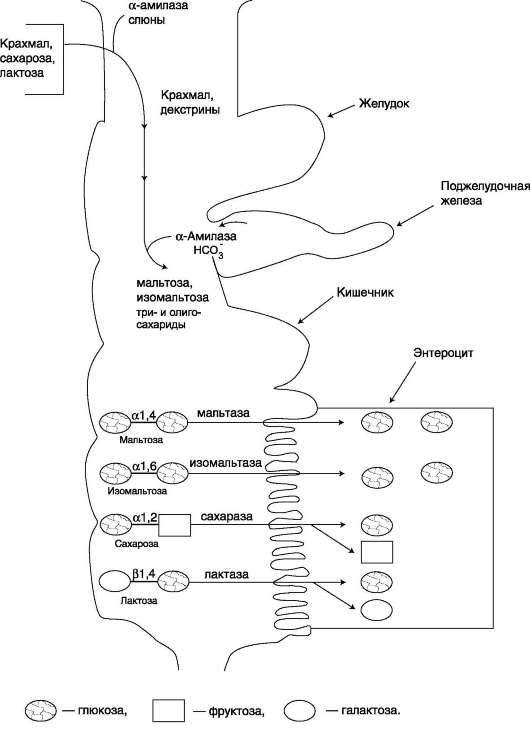
Рис. 7-17. Переваривание углеводов.
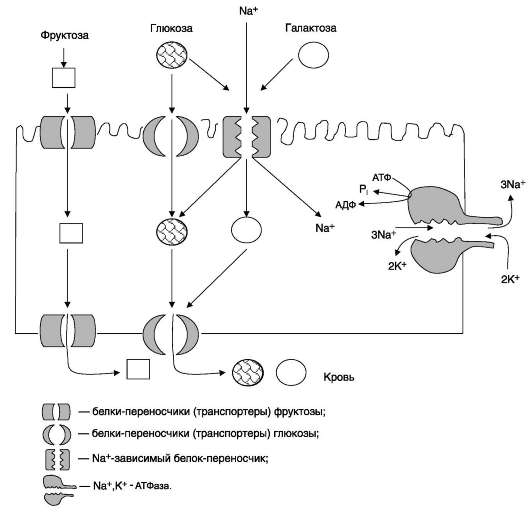
Рис. 7-18. Всасывание углеводов в кишечнике. Всасывание моносахаридов из кишечника происходит путём облегчённой диффузии с помощью специальных белков-переносчиков (транспортёров). Кроме того, глюкоза и галактоза транспортируются в энтероцит путём вторично-активного транспорта, зависимого от градиента концентрации ионов натрия. Белки-транспортёры, зависимые от градиента Na+, обеспечивают всасывание глюкозы из просвета кишечника в энтероцит против градиента концентрации. Концентрация Na+, необходимая для этого транспорта, обеспечивается Na+,К+-АТФ-азой, которая работает как насос, откачивая из клетки Na+ в обмен на К+. В отличие от глюкозы, фруктоза транспортируется системой, не зависящей от градиента натрия.
После всасывания моносахариды (главным образом, глюкоза) покидают клетки слизистой оболочки кишечника через мембрану, обращён-ную к кровеносному капилляру, с помощью облегчённой диффузии. Часть глюкозы (более половины) через капилляры кишечных ворсинок попадает в кровеносную систему и по воротной вене доставляется в печень. Остальное количество глюкозы поступает в клетки других тканей.
б. транспорт глюкозы из крови в клетки
Потребление глюкозы клетками из кровотока происходит также путём облегчённой диффузии. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента её концентрации. Исключение составляют клетки мышц и жировой ткани, где облегчённая диффузия регулируется инсулином (гормон поджелудочной железы). В отсутствие инсулина плазматическая мембрана этих клеток непроницаема для глюкозы, так как она не содержит белки-переносчики (транспортёры) глюкозы. Транспортёры глюкозы называют также рецепторами глюкозы. Например, описан транспортёр глюкозы, выделенный из эритроцитов. Это трансмембранный белок, полипептидная цепь которого построена из 492 аминокислотных остатков и имеет доменную структуру. Полярные домены белка расположены по разные стороны мембраны, гидрофобные располагаются в мембране, пересекая её несколько раз. Транспортёр имеет участок связывания глюкозы на внешней стороне мембраны. После присоединения глюкозы конформация белка изменяется, в результате чего глюкоза оказывается связанной с белком в участке, обращённом внутрь клетки.
Затем глюкоза отделяется от транспортёра, переходя внутрь клетки (см. раздел 5).
Считают, что способ облегчённой диффузии по сравнению с активным транспортом предотвращает транспорт ионов вместе с глюкозой, если она транспортируется по градиенту концентрации.
Глюкозные транспортёры (ГЛЮТ) обнаружены во всех тканях. Существует несколько разновидностей ГЛЮТ (табл. 7-1), они пронумерованы в соответствии с порядком их обнаружения.
Структура белков семейства ГЛЮТ отличается от белков, транспортирующих глюкозу через мембрану в кишечнике и почках против градиента концентрации.
Описанные 5 типов ГЛЮТ имеют сходные первичную структуру и доменную организацию.
• ГЛЮТ-1 обеспечивает стабильный поток глюкозы в мозг;
• ГЛЮТ-2 обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов и печени. ГЛЮТ-2 участвует в транспорте глюкозы в β-клетки поджелудочной железы;
• ГЛЮТ-3 обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей;
• ГЛЮТ-4 - главный переносчик глюкозы в клетки мышц и жировой ткани;
• ГЛЮТ-5 встречается, главным образом, в клетках тонкого кишечника. Его функции известны недостаточно.
Все типы ГЛЮТ могут находиться как в плазматической мембране, так и в цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток.
Таблица 7-1. Распределение белков-транспортёров глюкозы (ГЛЮТ)

Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию транспортёров в мембрану. После чего возможен облегчённый транспорт глюкозы в эти клетки. После снижения концентрации инсулина в крови транспортёры глюкозы снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается (рис. 7-19).
Перемещение глюкозы из первичной мочи в клетки почечных канальцев происходит вторично-активным транспортом, подобно тому, как это осуществляется при всасывании глюкозы из просвета кишечника в энтероциты. Благодаря этому глюкоза может поступать в клетки даже в том случае, если её концентрация в первичной моче меньше, чем в клетках. При этом глюкоза реабсорбируется из первичной мочи почти полностью (99%).
Известны различные нарушения в работе транспортёров глюкозы. Наследственный дефект этих
белков может лежать в основе инсулинонезави-симого сахарного диабета (см. раздел 11). В то же время причиной нарушения работы транспортёра глюкозы может быть не только дефект самого белка. Нарушения функции ГЛЮТ-4 возможны на следующих этапах:
• передача сигнала инсулина о перемещении этого транспортёра к мембране;
• перемещение транспортёра в цитоплазме;
• включение в состав мембраны;
• отшнуровывание от мембраны и т.д.
iv. нарушения переваривания и всасывания углеводов
В основе патологии переваривания и всасывания углеводов могут быть причины двух типов:
• дефекты ферментов, участвующих в гидролизе углеводов в кишечнике;
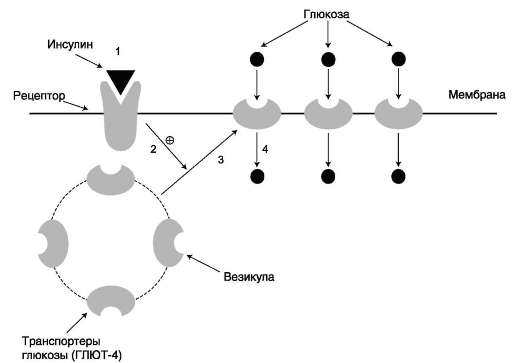
Рис. 7-19. Влияние инсулина на перемещение транспортёров глюкозы из цитоплазмы в плазматическую мембрану. 1 - связывание инсулина с рецептором; 2 - участок инсулинового рецептора, обращённый внутрь клетки, стимулирует перемещение транспортёров глюкозы. 3, 4 - транспортёры в составе содержащих их везикул перемещаются к плазматической мембране клетки, включаются в её состав и переносят глюкозу в клетку.
• нарушение всасывания продуктов переваривания углеводов в клетки слизистой оболочки кишечника.
В обоих случаях возникает осмотическая диарея, которую вызывают нерасщеплённые дисахариды или невсосавшиеся моносахариды. Эти невостребованные углеводы поступают в дистальные отделы кишечника, изменяя осмотическое давление содержимого кишечника. Кроме того, оставшиеся в просвете кишечника углеводы частично подвергаются ферментативному расщеплению микроорганизмами с образованием органических кислот и газов. Всё вместе приводит к притоку воды в кишечник, увеличению объёма кишечного содержимого, усилению перистальтики, спазмам и болям, а также метеоризму.
Термином «мальабсорбция» называют недостаточное всасывание переваренных продуктов углеводов. Но поскольку клинические проявления при недостаточном переваривании и всасывании сходны, то термином «мальабсорбция» называют оба вида нарушений.
А. НАРУШЕНИЯ ПЕРЕВАРИВАНИЯ УГЛЕВОДОВ В КИШЕЧНИКЕ
Нарушения переваривания могут быть связаны как с недостаточной активностью отдельных дисахаридаз, так и с недостаточностью всего ферментативного комплекса, например сахара-зо-изомальтазного.
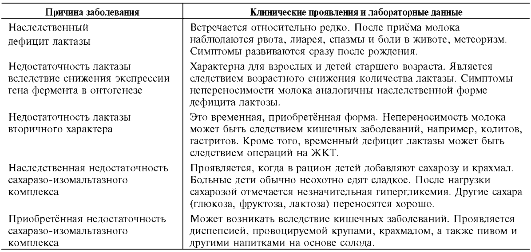
Известны наследственные и приобретённые формы недостаточности активности ферментов. Симптомы врождённых форм проявляются достаточно рано, например после первых кормлений грудным молоком (при дефиците лактазы), после перехода на искусственное вскармливание или при добавлении в рацион сахара и крахмала (при дефиците а-амилазы или специфических дисахаридаз). В случае недостаточного лечения врождённые формы патологии сопровождаются хроническим дисбактериозом и нарушениями физического развития ребёнка.
Приобретённые формы патологии могут наблюдаться при кишечных заболеваниях, например гастритах, колитах, энтеритах. Следует заметить, что в этих случаях особенно заметно снижение активности лактазы. Как уже говорилось, активность лактазы в кишечнике ниже, чем других дисахаридаз, поэтому уменьшение её активности
становится заметным для организма в первую очередь.
Дефицит лактазы у взрослых людей может иметь и другую причину. Возможно снижение экспрессии гена лактазы возрастного характера. Уже упоминалось, что активность лактазы у взрослых людей в норме значительно ниже, чем у детей. Поэтому снижение активности лактазы относительно уже имеющегося низкого уровня у отдельных людей может проявляться непереносимостью молока. Носителями патологии, связанной с дефицитом лактазы, являются чаще всего лица африканского и азиатского происхождения. Средняя частота данной формы патологии в странах Европы составляет 7-12%, в Китае - 80%, в отдельных районах Африки - до 97%. Подобные наблюдения распространения лактазной недостаточности связывают с исторически сложившимся рационом питания и отсутствием молочного скотоводства в упомянутых регионах. Примеры и причины нарушения переваривания дисахаридов перечислены в табл. 7-2.
Существуют редкие формы нарушения переваривания углеводов. Например, известна наследственная недостаточность трегалазы, которая проявляется диспепсией после употребления грибов, содержащих трегалозу.
В отдельных случаях мальабсорбция может быть вызвана несколькими причинами. Например, после операции на желудке возможны ухудшение смешивания пищи с пищеварительными соками, снижение их секреции, ускорение прохождения пищи через кишечник, колонизация бактериями слепой и приводящей петель.
Б. НАРУШЕНИЯ ВСАСЫВАНИЯ
МОНОСАХАРИДОВ
Нарушения всасывания могут быть следствием дефекта какого-либо компонента (белка или фермента), участвующего в системе транспорта моносахаридов через мембрану. Описаны патологии, связанные с дефектом натрийзависимого белка-переносчика глюкозы.
Для диагностики различных нарушений переваривания используют пробы с нагрузкой определёнными углеводами. Недостаточность кишечных дисахаридаз можно диагностировать с помощью введения дисахарида и последующего определения концентрации глюкозы
Таблица 7-2. Нарушения переваривания дисахаридов
в крови. Для большей чувствительности этот тест проводят, вводя сначала дисахарид (
Если тест при нагрузке моносахаридом сопровождается адекватным повышением его концентрации в крови, а нагрузка дисахаридом не даёт нормальной реакции, то это, скорее всего, указывает на дефект кишечной дисахаридазы, а не системы транспорта.
О недостаточности лактазы можно судить, определяя водород в выдыхаемом воздухе (водородный тест). Водород образуется в результате действия бактериальных ферментов на лактозу.
v. метаболизм глюкозы в клетке
После всасывания в кишечнике моносахариды поступают в воротную вену и далее преимущественно в печень. Поскольку в составе основных углеводов пищи преобладает глюкоза, её можно считать основным продуктом переваривания уг-
леводов. Другие моносахариды, поступающие из кишечника в процессе метаболизма, могут превращаться в глюкозу или продукты её метаболизма. Часть глюкозы в печени депонируется в виде гликогена, а другая часть через общий кровоток доставляется и используется разными тканями и органами. При нормальном рационе питания концентрация глюкозы в крови поддерживается на уровне 3,3-5,5 ммоль/л (60-100 мг/дл). А в период пищеварения её концентрация может повышаться примерно до 150 мг/дл (8 ммоль/л).
А. ФОСФОРИЛИРОВАНИЕ ГЛЮКОЗЫ
В дальнейших превращениях в клетках глюкоза и другие моносахариды участвуют только в виде фосфорных эфиров. Фосфорилирование свободных моносахаридов - обязательная реакция на пути их использования, она приводит к образованию более реакционно-способных соединений и поэтому может рассматриваться как реакция активации.
Глюкоза, поступающая в клетки органов и тканей, сразу же подвергается фосфорилированию с использованием АТФ. Эту реакцию во многих тканях катализирует фермент гексокиназа, а в печени и поджелудочной железе - фермент глюкокиназа. Фосфорилирование глюкозы - практически необратимая реакция, так как она
протекает с использованием значительного количества энергии. Образование глюкозо-6-фосфата в клетке - своеобразная «ловушка» для глюкозы, так как мембрана клетки непроницаема для фос-форилированной глюкозы (нет соответствующих транспортных белков). Кроме того, фосфорили-рование уменьшает концентрацию свободной глюкозы в цитоплазме. В результате создаются благоприятные условия для облегчённой диффузии глюкозы в клетки из крови.
Глюкокиназа. Фосфорилирование глюкозы в гепатоцитах в период пищеварения обеспечивается свойствами глюкокиназы, которая имеет высокое значение Кm - 10 ммоль/л. В этот период концентрация глюкозы в воротной вене больше, чем в других отделах кровяного русла и может превышать 10 ммоль/л, а следовательно, активность глюкокиназы в гепатоцитах повышается. Следует отметить, что активность глюкокиназы, в отличие от гексокиназы, не ингибируется продуктом катализируемой реакции - глюкозо-6-фосфатом. Это обстоятельство обеспечивает повышение концентрации глюкозы в клетке в фосфорилированной форме, соответственно её уровню в крови. Как уже упоминалось, глюкоза проникает в гепатоциты путём облегчённой диффузии при участии транспортёра ГЛЮТ-2 (независимого от инсулина). ГЛЮТ-2, так же, как глюкокиназа, имеет высокую Кт, что способствует повышению скорости поступления глюкозы в гепатоциты в период пищеварения, следовательно, ускоряет её фосфорилирование и дальнейшее использование для депонирования.
Хотя инсулин и не влияет на транспорт глюкозы, он усиливает приток глюкозы в гепато-циты в период пищеварения косвенным путём, индуцируя синтез глюкокиназы и ускоряя тем самым фосфорилирование глюкозы.
Преимущественное потребление глюкозы гепатоцитами, обусловленное свойствами глю-кокиназы, предотвращает чрезмерное повышение её концентрации в крови в абсорбтивном периоде. Это, в свою очередь, снижает последствия протекания нежелательных реакций с участием глюкозы, например гликозилиро-вания белков.
Гексокиназа отличается от глюкокиназы высоким сродством к глюкозе (Кm <0,1 ммоль/л). Следовательно, этот фермент, в отличие от глюкокиназы, активен при низкой концент-
рации глюкозы в крови, что характерно для постабсорбтивного состояния. Печень в этот период поглощает гораздо меньше глюкозы, так как скорость её внутриклеточного фосфорили-рования глюкокиназой резко снижается. Тогда как потребление глюкозы мозгом, эритроцитами и другими тканями обеспечивается активной в этих условиях гексокиназой. Фермент гексоки-наза может катализировать фосфорилирование не только D-глюкозы, но и других гексоз, хотя и с меньшей скоростью. Активность гексокиназы изменяется в зависимости от потребностей клетки в энергии. В качестве регуляторов выступают соотношение АТФ/АДФ и внутриклеточный уровень глюкозо-6-фосфата (продукта катализируемой реакции). При снижении расхода энергии в клетке повышается уровень АТФ (относительно АДФ) и глюкозо-6-фосфата. В этом случае активность гексокиназы снижается, и, следовательно, уменьшается скорость поступления глюкозы в клетку.
Следует отметить, что в разных тканях гек-сокиназа присутствует в различных изоформах, отличающихся величиной Кт. Глюкокина-за печени (и почек) является изоформой IV (гексокиназа IV). В клетках мышц содержится гексокиназа II, а в клетках опухолевых тканей преобладает гексокиназа III, с более высоким, чем у гексокиназы II, сродством к глюкозе.
Б. ДЕФОСФОРИЛИРОВАНИЕ
ГЛЮКОЗО-6-ФОСФАТА
Превращение глюкозо-6-фосфата в глюкозу возможно в печени, почках и клетках эпителия кишечника. В клетках этих органов имеется фермент глюкозо-6-фосфатаза, катализирующая отщепление фосфатной группы гидролитическим путём: Глюкозо-6-фосфат +Н2О → Глюкоза + Н3РО4 Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь. В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому дефосфорилирование глюкозо-6-фосфата невозможно. Пример подобного необратимого проникновения глюкозы в клетку - мышцы, где глюкозо-6-фосфат может использоваться только в метаболизме этой клетки.
В. МЕТАБОЛИЗМ ГЛЮКОЗО-6-ФОСФАТА
Глюкозо-6-фосфат может использоваться в клетке в различных превращениях, основными
из которых являются: синтез гликогена, катаболизм с образованием СО2 и Н2О или лактата, синтез пентоз. Распад глюкозы до конечных продуктов служит источником энергии для организма. Вместе с тем в процессе метаболизма глюкозо-6-фосфата образуются промежуточные продукты, используемые в дальнейшем для синтеза аминокислот, нуклеотидов, глицерина и жирных кислот. Таким образом, глюкозо-6-фосфат - не только субстрат для окисления, но и строительный материал для синтеза новых соединений (рис. 7-20).
vi. метаболизм гликогена
Многие ткани синтезируют в качестве резервной формы глюкозы гликоген. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови и создают депо для её использования тканями по мере необходимости.
А. СТРОЕНИЕ И ФУНКЦИИ ГЛИКОГЕНА
Гликоген - разветвлённый гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках α-1,4-гликозидной связью. В точках ветвления мономеры соединены α-1,6-гликозидными связями. Эти связи образуются примерно с каждым десятым остатком глюкозы. Следовательно, точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Так возникает древообразная структура с молекулярной массой >107 Д, что соответствует приблизительно 50 000 остатков глюкозы (рис. 7-21). Таким образом, в молекуле гликогена имеется только одна свободная аномерная ОН-группа и, следовательно, только один восстанавливающий (редуцирующий) конец.
В клетках животных гликоген - основной резервный полисахарид. При полимеризации глюкозы снижается растворимость образующейся молекулы гликогена и, следовательно, её влияние на осмотическое давление в клетке. Это обстоятельство объясняет, почему в клетке депонируется гликоген, а не свободная глюкоза.
Гликоген хранится в цитозоле клетки в форме гранул диаметром 10-40 нм. С гранулами связаны и некоторые ферменты, участвующие в метаболизме гликогена, что облегчает их
взаимодействие с субстратом. Разветвлённая структура гликогена обусловливает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих мономеры при распаде или синтезе гликогена, так как эти ферменты могут одновременно работать на нескольких ветвях молекулы. Гликоген депонируется главным образом в печени и скелетных мышцах.
После
приёма пищи, богатой углеводами, запас гликогена в печени может
составлять примерно 5% от её массы. В мышцах запасается около 1%
гликогена, однако масса мышечной ткани значительно больше и поэтому
общее количество гликогена в мышцах в 2 раза больше, чем в печени.
Гликоген может синтезироваться во многих клетках, например в нейронах,
макрофагах, клетках жировой ткани, но содержание его в этих тканях
незначительно. В организме может содержаться до
Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови в пост-абсорбтивном периоде. Поэтому содержание гликогена в печени изменяется в зависимости от ритма питания. При длительном голодании оно снижается почти до нуля. Гликоген мышц служит резервом глюкозы - источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови. Как уже упоминалось ранее, в клетках мышц нет фермента глюкозо-6-фосфа-тазы, и образование свободной глюкозы невозможно. Расход гликогена в мышцах зависит в основном от физической нагрузки (рис. 7-22).
Б. СИНТЕЗ ГЛИКОГЕНА (ГЛИКОГЕНОГЕНЕЗ)
Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи). Следует отметить, что синтез гликогена из глюкозы (рис. 7-23), как и любой анаболический процесс, является эндергоническим, т.е. требующим затрат энергии.
Глюкоза, поступающая в клетку, фосфори-лируется при участии АТФ (реакция 1). Затем глюкозо-6-фосфат в ходе обратимой реакции превращается в глюкозо-1-фосфат (реакция 2) под действием фермента фосфоглюкомутазы. Глюкозо-1-фосфат по термодинамическому состоянию мог бы служить субстратом для синтеза гликогена. Но в силу обратимости реакции
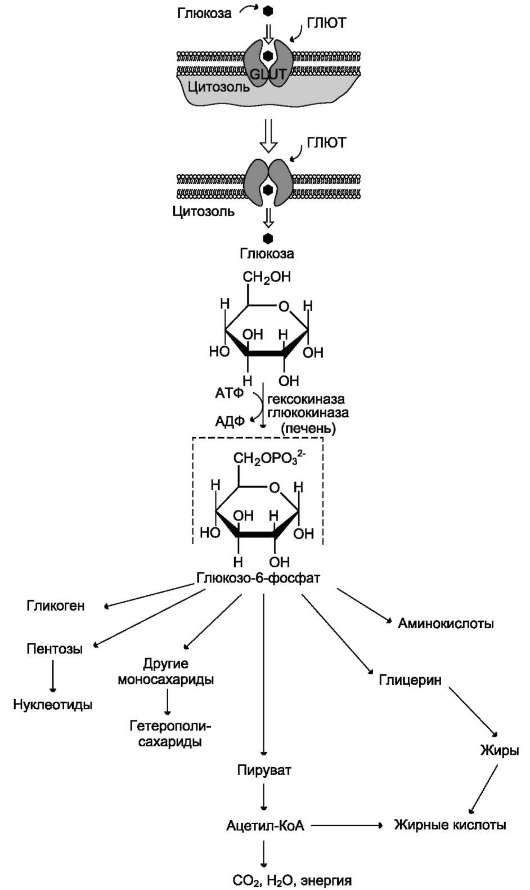
Рис. 7-20. Метаболизм глюкозо-6-фосфата.
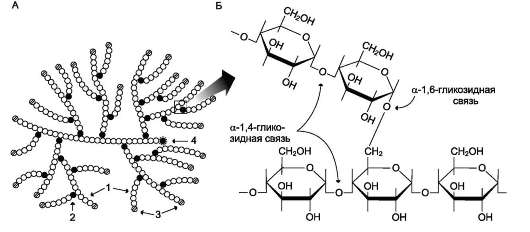
Рис. 7-21. Структура гликогена. А. Строение молекулы гликогена: 1 - остатки глюкозы, соединённые α-1,4-гликозидной связью; 2 - остатки глюкозы, соединённые α-1,6-гликозидной связью; 3 - нередуцирующие концевые мономеры; 4 - редуцирующий концевой мономер. Б. Строение отдельного фрагмента молекулы гликогена.
глюкозо-6-фосфат ↔ глюкозо-1-фосфат синтез гликогена из глюкозо-1-фосфата и его распад оказались бы также обратимыми и поэтому неконтролируемыми. Чтобы синтез гликогена был термодинамически необратимым, необходима дополнительная стадия образования уридинди-фосфатглюкозы из УТФ и глюкозо-1-фосфата (реакция 3). Фермент, катализирующий эту реакцию, назван по обратной реакции: УДФ-глюкопирофосфорилаза. Однако в клетке обратная реакция не протекает, потому что образовавшийся в ходе прямой реакции пирофосфат очень быстро расщепляется пирофосфатазой на 2 молекулы фосфата (рис. 7-24).
Реакция образования УДФ-глюкозы обусловливает необратимость всей серии реакций, протекающих при синтезе гликогена. Этим же объясняется невозможность протекания распада гликогена путём простого обращения процесса его синтеза.
Образованная УДФ-глюкоза далее используется как донор остатка глюкозы при синтезе гликогена (рис. 7-23, реакция 4). Эту реакцию катализирует фермент гликогенсинтаза (глюко-зилтрансфераза). Поскольку в данной реакции не используется АТФ, фермент называют син-тазой, а не синтетазой. Нуклеотидная часть
УДФ-глюкозы играет существенную роль в действии гликоген синтазы, выполняя функцию «рукоятки», при помощи которой фермент располагает глюкозу в полисахаридной цепи в нужном положении. Кроме того, нуклеотидная
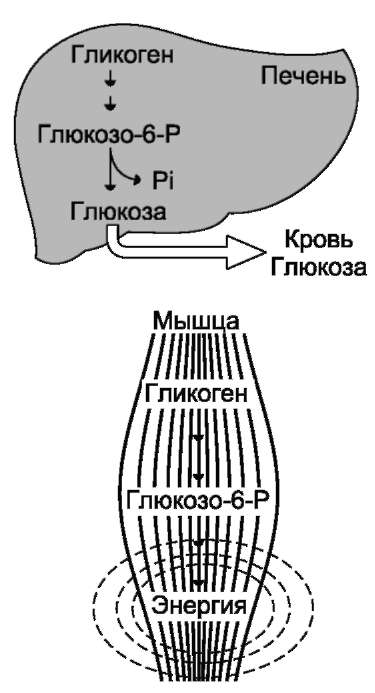
Рис. 7-22. Функции гликогена в печени и мышцах.
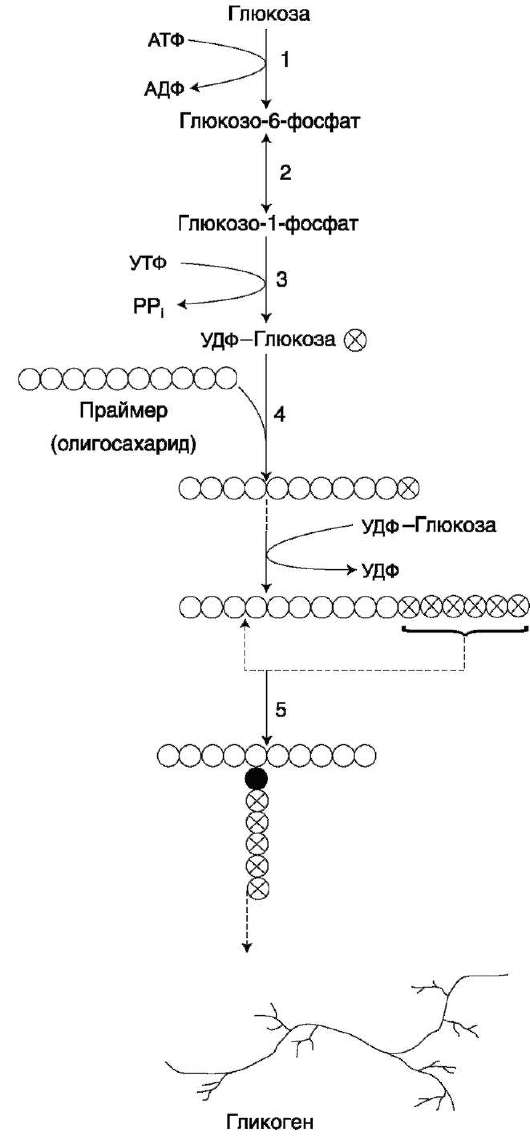
Рис. 7-23. Синтез гликогена. 1 - глюкокиназа или гексокиназа; 2 - фосфоглюкомутаза; 3 - УДФ-глюкопирофосфорилаза; 4 - гликогенсинтаза (глюкозилтрансфераза); 5 - фермент «ветвления» (амило-1,4→ 1,6-глюкозилтрансфераза), светлые и заштрихованные кружки - глюкозные остатки, закрашенные кружки - глюкозные остатки в точке ветвления.
часть УДФ-глюкозы, по-видимому, необходима для узнавания субстрата при катализе.
Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида, называемой «затравка», или «праймер». К «затравке» последовательно присоединяются молекулы глюкозы. Строением молекулы «затравки» как бы предопределяется тип связи, который возникает в реакции трансгликозилирования. Таким образом, синтезируется полисахарид, аналогичный по строению с «затравочным». В состав «затравки» может входить белок гликогенин, в котором к ОН-группе одного из тирозиновых остатков присоединена олигосахаридная цепочка (примерно 8 остатков глюкозы). Глюкозные остатки переносятся гликогенсинтазой на не-редуцирующий конец олигосахаридной цепочки и связываются а-1,4-гликозидными связями. По окончании синтеза гликогенин остаётся включённым в гранулу гликогена.
Разветвлённая структура гликогена образуется при участии амило-1,4→1,6-глюкозилтрансфе-разы, называемой ферментом «ветвления» (от англ. branching enzyme). Как только гликоген-синтаза удлиняет линейный участок примерно до 11 глюкозных остатков, фермент ветвления переносит её концевой блок, содержащий 6- 7 остатков, на внутренний остаток глюкозы этой или другой цепи. В точке ветвления концевой остаток глюкозы олигосахарида соединяется с гидроксильной группой в С6-положении с образованием а-1,6-гликозидной связи. Новая точка ветвления может быть образована на расстоянии не менее 4 остатков от любой уже существующей. Таким образом, по мере синтеза гликогена многократно возрастает число ветвлений. Концы цепей служат точками роста молекулы при её синтезе и началом при её распаде.
В. РАСПАД ГЛИКОГЕНА (ГЛИКОГЕНОЛИЗ)
Распад гликогена или его мобилизация происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени распадается в основном в интервалах между приёмами пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической работы.
Распад гликогена (рис. 7-25) происходит путём последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Глико-зидная связь расщепляется с использованием
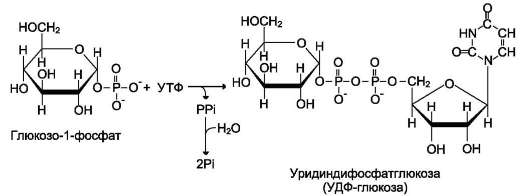
Рис. 7-24. Образование УДФ-глюкозы.
неорганического фосфата, поэтому процесс называется фосфоролизом, а фермент гликоген-фосфорилазой.
Так же как и синтез, расщепление гликогена начинается с нередуцирующего конца поли-сахаридной цепи. При этом наличие разветвлённой структуры гликогена облегчает быстрое высвобождение глюкозных остатков, так как чем больше концов имеет молекула гликогена, тем больше молекул гликогенфосфорилазы могут действовать одновременно.
Гликогенфосфорилаза расщепляет только α-1,4-гликозидные связи (реакция 1). Последовательное отщепление глюкозных остатков прекращается, когда до точки ветвления остаётся 4 мономера. Подобная особенность в действии гликогенфосфорилазы обусловлена размером и строением её активного центра.
Дальнейший распад гликогена требует участия двух других ферментов. Сначала три оставшихся до точки ветвления глюкозных остатка переносятся при участии олигосахаридтрансферазы (реакция 2) на нередуцирующий конец соседней цепи, удлиняя её и таким образом создавая условия для действия фосфорилазы. Оставшийся в точке ветвления глюкозный остаток гидроли-
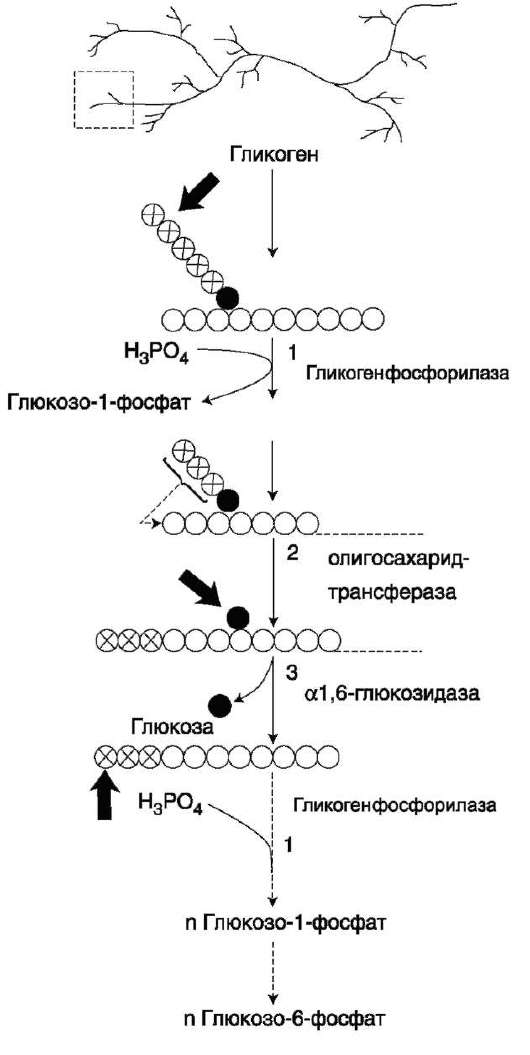
Рис. 7-25. Распад гликогена. В рамке - фрагмент гликогена с точкой ветвления. Закрашенный кружок - глюкозный остаток, связанный α-1,6-гликозидной связью; светлые и заштрихованные кружки - глю-козные остатки в линейных участках и боковых ветвях, связанные а-1,4-гликозидной связью. 1 - гликоген-фосфорилаза; 2 - олигосахаридтрансфераза; 3 - α-1,6-глюкозидаза.
тически отщепляется с помощью а-1,6-глюко-зидазы в виде свободной глюкозы (реакция 3), после чего неразветвлённый участок гликогена может вновь атаковаться фосфорилазой.
Считают, что перенос трёх остатков глюкозы и удаление мономера из точки ветвления (реакции 2 и 3) катализирует один и тот же фермент, который обладает двумя разными ферментативными активностями - трансферазной и гликозидазной. Его называют «деветвящим» ферментом (от англ. debranching enzyme).
Продукт действия гликогенфосфорилазы - глюкозо-1-фосфат - затем изомеризуется в глю-козо-6-фосфат фосфоглюкомутазой. Далее глю-козо-6-фосфат включается в процесс катаболизма или другие метаболические пути. В печени (но не в мышцах) глюкозо-6-фосфат может гидролизо-ваться с образованием глюкозы, которая выделяется в кровь. Эту реакцию катализирует фермент глюкозо-6-фосфатаза. Реакция протекает в про-
свете ЭР, куда с помощью специального белка транспортируется глюкозо-6-фосфат. Фермент локализован на мембране ЭР таким образом, что его активный центр обращён в просвет ЭР. Продукты гидролиза (глюкоза и неорганический фосфат) возвращаются в цитоплазму также с помощью транспортных систем.
Г. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ОБМЕНА ГЛИКОГЕНА В ПЕЧЕНИ И МЫШЦАХ
На рисунке 7-26 приведена общая схема синтеза и распада гликогена и регуляция этих процессов гормонами.
Сравнение этих процессов позволяет сделать следующие выводы:
• синтез и распад гликогена протекают по разным метаболическими путям;
• печень запасает глюкозу в виде гликогена не столько для собственных нужд, сколько
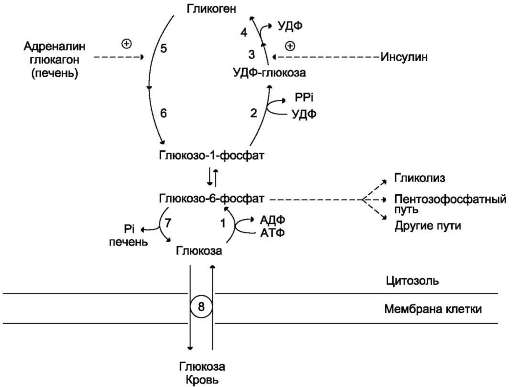
Рис. 7-26. Синтез и распад гликогена. 1 - гексокиназа или глюкокиназа (печень); 2 - УДФ-глюкопирофос-форилаза; 3 - гликогенсинтаза; 4 - амило-1,4→1,6-глюкозилтрансфераза (фермент ветвления); 5 - глико-генфосфорилаза; 6 - «деветвящий» фермент; 7 - глюкозо-6-фосфатаза (печень); 8 - транспортные системы ГЛЮТ.
для поддержания постоянной концентрации глюкозы в крови, и, следовательно, обеспечивает поступление глюкозы в другие ткани. Присутствие в печени глюкозо-6-фосфатазы обусловливает эту главную функцию печени в обмене гликогена;
• функция мышечного гликогена заключается в освобождении глюкозо-6-фосфата, потребляемого в самой мышце для окисления и использования энергии;
• синтез гликогена - процесс эндергоничес-кий. Так на включение одного остатка глюкозы в полисахаридную цепь используется 1 моль АТФ и 1 моль УТФ;
• распад гликогена до глюкозо-6-фосфата не требует энергии;
• необратимость процессов синтеза и распада гликогена обеспечивается их регуляцией.
vii. регуляция метаболизма гликогена
Процессы накопления глюкозы в виде гликогена и его распада должны быть согласованы с потребностями организма в глюкозе как источнике энергии. Одновременное протекание этих метаболических путей невозможно, так как в этом случае образуется «холостой» цикл, существование которого приводит только к бесполезной трате АТФ.
Изменение направления процессов в метаболизме гликогена обеспечивают регуляторные механизмы, в которых участвуют гормоны. Переключение процессов синтеза и мобилизации гликогена происходит при смене абсорбтивного периода на постабсорбтивный или состояния покоя организма на режим физической работы. В переключении этих метаболических путей в печени участвуют гормоны инсулин, глюкагон и адреналин, а в мышцах - инсулин и адреналин.
а. характеристика гормонов, регулирующих обмен гликогена
Первичным сигналом для синтеза и секреции инсулина и глюкагона является изменение уровня глюкозы в крови. В норме концентрация глюкозы в крови соответствует 3,3-5,5 ммоль/л (60-100 мг/дл).
Инсулин - белковый гормон, синтезируется и секретируется в кровь β-клетками островков Лангерханса поджелудочной железы. β-клетки чувствительны к изменениям содержания глюкозы в крови и секретируют инсулин в ответ на повышение её содержания после приёма пищи. Транспортный белок (ГЛЮТ-2), обеспечивающий поступление глюкозы в β-клетки, отличается низким сродством к ней. Следовательно, этот белок транспортирует глюкозу в клетку поджелудочной железы лишь после того, как её содержание в крови будет выше нормального уровня (более 5,5 ммоль/л).
В β-клетках глюкоза фосфорилируется глю-кокиназой, имеющей также высокую Кm для глюкозы - 12 ммоль/л. Скорость фосфорилиро-вания глюкозы глюкокиназой в β-клетках прямо пропорциональна её концентрации в крови.
Синтез инсулина регулируется глюкозой. Глюкоза (или её метаболиты), по-видимому, непосредственно участвуют в регуляции экспрессии гена инсулина. Секреция инсулина и глюкагона также регулируется глюкозой, которая стимулирует секрецию инсулина из β-клеток и подавляет секрецию глюкагона из α-клеток. Кроме того, сам инсулин снижает секрецию глюкагона (см. раздел 11).
Глюкагон - «гормон голода», вырабатываемый α-клетками поджелудочной железы в ответ на снижение уровня глюкозы в крови. По химической природе глюкагон - пептид.
Адреналин выделяется из клеток мозгового вещества надпочечников в ответ на сигналы нервной системы, идущие из мозга при возникновении экстремальных ситуаций (например, бегство или борьба), требующих внезапной мышечной деятельности. Адреналин является сигналом «тревоги». Он должен мгновенно обеспечить мышцы и мозг источником энергии.
б. регуляция активности гликогенфосфорилазы
и гликогенсинтазы
Поскольку синтез и распад гликогена протекают по различным метаболическим путям, эти процессы могут контролироваться реципрокно. Влияние гормонов на синтез и распад гликогена осуществляется путём изменения в противоположных направлениях активности двух ключевых
ферментов: гликогенсинтазы и гликогенфос-форилазы с помощью их фосфорилирования и дефосфорилирования (рис. 7-27).
Гликогенфосфорилаза существует в 2 формах: 1) фосфорилированная - активная (форма а); 2) дефосфорилированная - неактивная (форма в). Фосфорилирование осуществляется путём переноса фосфатного остатка с АТФ на гидроксильную группу одного из сериновых остатков фермента. Следствие этого - кон-формационные изменения молекулы фермента и его активация.
Взаимопревращения 2 форм гликогенфосфо-рилазы обеспечиваются действием ферментов киназы фосфорилазы и фосфопротеинфосфа-тазы (фермент, структурно связанный с моле-
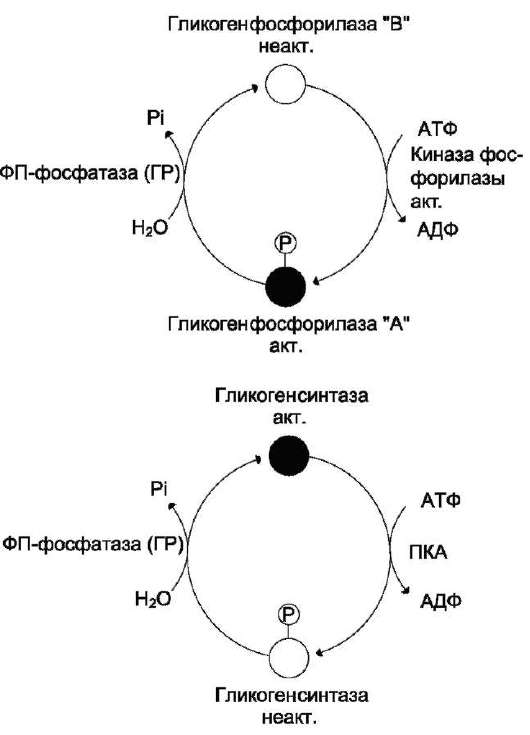
Рис. 7-27. Изменение активности гликогенфосфо-рилазы и гликогенсинтазы. Кружками обозначены молекулы фермента: активные - чёрные, неактивные - белые. ФП-фосфатаза (ГР) - фосфопроте-инфосфатаза гранул гликогена.
кулами гликогена). В свою очередь, активность киназы фосфорилазы и фосфопротеинфосфата-зы также регулируется путём фосфорилирования и дефосфорилирования.
Активация киназы фосфорилазы происходит под действием протеинкиназы А - ПКА (цАМФ-зависимой). цАМФ сначала активирует про-теинкиназу А, которая фосфорилирует киназу фосфорилазы, переводя её в активное состояние, а та, в свою очередь, фосфорилирует глико-генфосфорилазу. Синтез цАМФ стимулируется адреналином и глюкагоном (см. раздел 5).
Активация фосфопротеинфосфатазы происходит в результате реакции фосфорилирования, катализируемой специфической протеинкиназой, которая, в свою очередь, активируется инсулином посредством каскада реакций с участием Ras-белка, а также других белков и ферментов (сигнальный Ras-путь, см. раздел 11). Активируемая инсулином протеинкиназа фосфорилирует и тем самым активирует фосфопротеинфосфа-тазу. Активная фосфопротеинфосфатаза дефос-форилирует и, следовательно, инактивирует киназу фосфорилазы и гликогенфосфорилазу (рис. 7-28).
Активность гликогенсинтазы также изменяется в результате фосфорилирования и дефосфори-лирования (см. выше рис. 7-27). Однако есть существенные различия в регуляции гликоген-фосфорилазы и гликогенсинтазы:
• фосфорилирование гликогенсинтазы катализирует ПК А и вызывает её инактивацию;
• дефосфорилирование гликогенсинтазы под действием фосфопротеинфосфатазы, наоборот, её активирует.
В. РЕГУЛЯЦИЯ МЕТАБОЛИЗМА ГЛИКОГЕНА В ПЕЧЕНИ
Как уже отмечалось, первичный сигнал для синтеза инсулина и глюкагона - изменение концентрации глюкозы в крови. Инсулин и глюкагон постоянно присутствуют в крови, но при смене абсорбтивного периода на пос-табсорбтивный изменяется их относительная концентрация, что является главным фактором, переключающим метаболизм гликогена в печени. Отношение концентрации инсулина в крови к концентрации глюкагона называют «ин-сулин-глюкагоновый индекс». В постабсорбтивном периоде инсулин-глюкагоновый индекс сни-
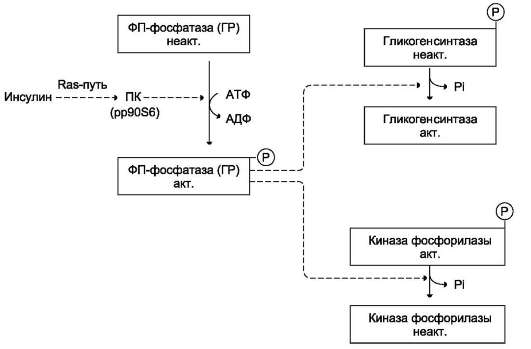
Рис. 7-28. Влияние инсулина на активность гликогенсинтазы и киназы фосфорилазы. ФП-фосфатаза (ГР) фосфопротеинфосфатаза гранул гликогена. ПК (рр90S6) - протеинкиназа, активируемая инсулином.
жается, и решающее значение в регуляции концентрации глюкозы в крови приобретает концентрация глюкагона.
Глюкагон для гепатоцитов служит внешним сигналом о необходимости выделения в кровь глюкозы за счёт распада гликогена (гликогено-лиза) или синтеза глюкозы из других веществ - глюконеогенеза (этот процесс будет изложен позднее). Гормон связывается с рецептором на плазматической мембране и активирует при посредничестве G-белка аденилатциклазу, которая катализирует образование цАМФ из АТФ (см. раздел 5). Далее следует каскад реакций, приводящий в печени к активации гликогенфос-форилазы и ингибированию гликогенсинтазы (рис. 7-29). Этот механизм приводит к высвобождению из гликогена глюкозо-1-фосфата, который превращается в глюкозо-6-фосфат. Затем под влиянием глюкозо-6-фосфатазы образуется свободная глюкоза, способная выйти из клетки в кровь. Таким образом, глюкагон в печени, стимулируя распад гликогена, способствует поддержанию глюкозы в крови на постоянном уровне.
Адреналин стимулирует выведение глюкозы из печени в кровь, для того чтобы снабдить
ткани (в основном мозг и мышцы) «топливом» в экстремальной ситуации. Эффект адреналина в печени обусловлен фосфорилированием (и активацией) гликогенфосфорилазы. Адреналин имеет сходный с глюкагоном механизм действия (рис. 7-29). Но возможно включение и другой эффекторной системы передачи сигнала в клетку печени (рис. 7-30).
Какая система передачи сигнала в клетку будет использована, зависит от типа рецепторов, с которыми взаимодействует адреналин. Так, взаимодействие адреналина с β2-рецепторами клеток печени приводит в действие аденилатциклазную систему. Взаимодействие же адреналина с а1-рецепторами «включает» инозитолфосфатный механизм трансмембранной передачи гормонального сигнала. Результат действия обеих систем - фосфорилирование ключевых ферментов и переключение процессов с синтеза гликогена на его распад. Следует отметить, что тип рецепторов, который в наибольшей степени вовлекается в ответ клетки на адреналин, зависит от концентрации его в крови.
В период пищеварения преобладает влияние инсулина, так как инсулин-глюкагоновый индекс в этом случае повышается. В целом инсулин вли-
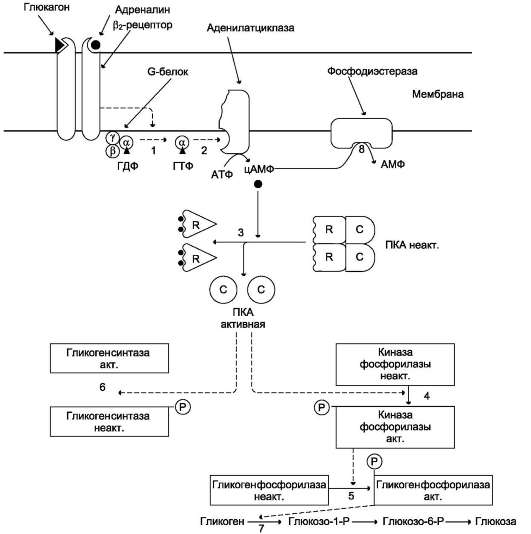
Рис. 7-29. Регуляция синтеза и распада гликогена в печени глюкагоном и адреналином. 1 - глюкагон и адреналин взаимодействуют со специфическими мембранными рецепторами. Комплекс гормон-рецептор влияет на конформацию G-белка, вызывая диссоциацию его на протомеры и замену в a-субъединице ГДФ на ГТФ;
2 - α-субъединица, связанная с ГТФ, активирует аденилатциклазу, катализирующую синтез цАМФ из АТФ;
3 - в присутствии цАМФ протеинкиназа А (цАМФ-зависимая) обратимо диссоциирует, освобождая обладающие каталитической активностью субъединицы С; 4 - протеинкиназа А фосфорилирует и активирует киназу фосфорилазы; 5 - киназа фосфорилазы фосфорилирует гликогенфосфорилазу, переводя её в активную форму; 6 - протеинкиназа А фосфорилирует также гликогенсинтазу, переводя её в неактивное состояние; 7 - в результате ингибирования гликогенсинтазы и активации гликогенфосфорилазы гликоген включается в процесс распада; 8 - фосфодиэстераза катализирует распад цАМФ и тем самым прерывает действие гормонального сигнала. Комплекс а-субъединица-ГТФ затем распадается, а-, β- и γ-субъединицы G-белка реассоциируются.
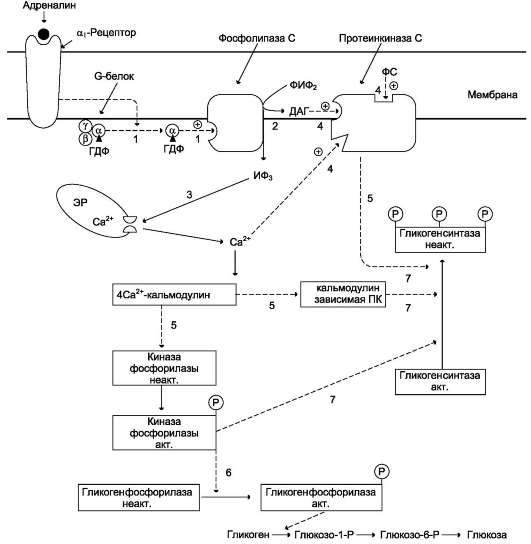
Рис. 7-30. Регуляция синтеза и распада гликогена в печени адреналином и Са2+. ФИФ2 - фосфатидили-нозитолбисфосфат; ИФ3 - инозитол-1,4,5-трифосфат; ДАГ - диацилглицерол; ЭР - эндоплазматический ретикулум; ФС - фосфодитилсерин. 1 - взаимодействие адреналина с α1-рецептором трансформирует сигнал через активацию G-белка на фосфолипазу С, переводя её в активное состояние; 2 - фосфолипаза С гидроли-зует ФИФ2 на ИФ3 и ДАГ; 3 - ИФ3 активирует мобилизацию Са2+ из ЭР; 4 - Са2+, ДАГ и фосфодитилсерин активируют протеинкиназу С. 5 - протеинкиназа С фосфорилирует гликогенсинтазу, переводя её в неактивное состояние; 5 - комплекс 4Са2+-кальмодулин активирует киназу фосфорилазы и кальмодулинзависимые протеинкиназы; 6 - киназа фосфорилазы фосфорилирует гликогенфосфорилазу и тем самым её активирует; 7 - активные формы трёх ферментов (кальмодулинзависимая протеинкиназа, киназа фосфорилазы и протеин-киназа С) фосфорилируют гликогенсинтазу в различных центрах, переводя её в неактивное состояние.
яет на обмен гликогена противоположно глюка-гону. Инсулин снижает концентрацию глюкозы в крови в период пищеварения, действуя на метаболизм печени следующим образом:
• снижает уровень цАМФ в клетках, фосфо-рилируя (опосредованно через Ras-путь) и тем самым активируя фосфодиэстеразу цАМФ - фермент, гидролизующий цАМФ с образованием АМФ. Механизм влияния инсулина на уровень цАМФ в клетке подробнее будет изложен в разделе 11;
• активирует (через Ras-путь) фосфопроте-инфосфатазу гранул гликогена, которая дефосфорилирует гликогенсинтазу и таким образом её активирует. Кроме того, фос-фопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует киназу фос-форилазы и гликогенфосфорилазу;
• индуцирует синтез глюкокиназы, тем самым ускоряя фосфорилирование глюкозы в клетке. Следует напомнить, что регуля-торным фактором в метаболизме гликогена является также величина Кт глюкокиназы, которая много выше, чем Кт гексокиназы. Смысл этих различий понятен: печень не должна потреблять глюкозу для синтеза гликогена, если её количество в крови в пределах нормы.
Всё это вместе приводит к тому, что инсулин одновременно активирует гликогенсинтазу и ингибирует гликогенфосфорилазу, переключая процесс мобилизации гликогена на его синтез.
В печени существует и аллостерическая регуляция гликогенфосфорилазы, обеспечивающая внутриклеточные потребности в глюкозе, но гормональные сигналы имеют приоритет над внутриклеточными и преследуют другие физиологические цели. Ранее (см. раздел 6) рассматривалось значение изменения в клетке уровней АТФ, АДФ и АМФ как показателя, отражающего потребности клетки в энергии. Замедление утилизации АТФ сопровождается снижением активности гликогенфосфорилазы и уменьшением скорости распада гликогена. Напротив, увеличение расходования АТФ ведёт к повышению уровня АМФ, активации гликогенфосфорилазы и ускорению распада гликогена. АТФ и АМФ являются ал-лостерическими эффекторами по отношению к гликогенфосфорилазе. Существует также и мета-
болический контроль активности гликогенфос-форилазы. Так, при повышении концентрации глюкозо-6-фосфата активность этого фермента в клетках печени снижается.
Г. РЕГУЛЯЦИЯ МЕТАБОЛИЗМА ГЛИКОГЕНА В МЫШЦАХ
Регуляция обмена гликогена в мышцах обеспечивает энергетическим материалом как интенсивную работу мышц (бег или борьба), так и энергозатраты в состоянии покоя.
В экстремальных ситуациях в мышечных клетках мобилизация гликогена ускоряется адреналином. Связывание адреналина с β-рецептора-ми, ассоциированными с аденилатциклазной системой, приводит к образованию цАМФ в клетке, а затем фосфорилированию и активации киназы фосфорилазы и гликогенфосфорилазы
(рис. 7-31).
Образование цАМФ, стимулированное адреналином, служит сигналом к увеличению производства энергии в результате ускорения расщепления гликогена. Именно в ходе распада, образованного из гликогена глюкозо-6-фосфата, синтезируется АТФ.
Инактивация гликогенсинтазы под влиянием адреналина в мышечных клетках проходит так же, как и в печени.
В состоянии покоя при низких концентрациях адреналина в крови гликогенфосфорилаза мышц находится в дефосфорилированном - неактивном состоянии (форма В), но распад гликогена всё-таки происходит. Это объясняется тем, что гликогенфосфорилаза активируется способом, не связанным с её фосфорилированием, так как уровень цАМФ в клетке низкий. В данной ситуации происходит аллостерическая активация гликогенфосфорилазы В. Активаторами фермента служат АМФ и Н3РО4, образующиеся в клетке при распаде АТФ (рис. 7-31, путь 1).
При умеренных мышечных сокращениях, т.е. в ситуации, не требующей участия в регуляции цАМФ, аллостерическим способом активируется киназа фосфорилазы (рис. 7-31, путь 2). В данном случае аллостерическими эффекторами служат ионы Са2+, концентрация которых резко возрастает при сокращении мышц в ответ на сигнал от двигательного нерва. Активность фермента снижается сразу же, как только кон-
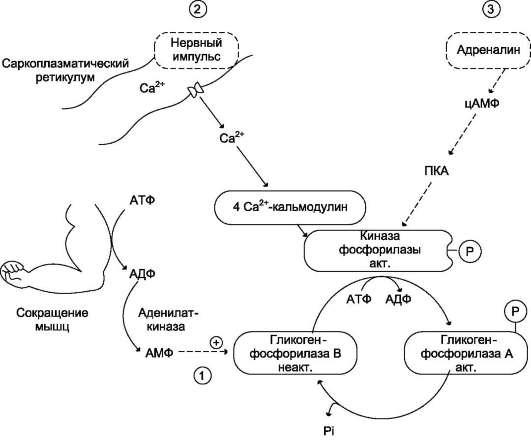
Рис. 7-31. Активация гликогенфосфорилазы мышц. 1 - аллостерическая активация гликогенфосфорилазы В. В процессе мышечного сокращения происходит разрушение АТФ с образованием АМФ, который является аллостерическим активатором гликогенфосфорилазы В; 2 - нервный импульс инициирует освобождение Са2+ из саркоплазматического ретикулума. Са2+ образует комплекс с кальмодулином, способный активировать киназу фосфорилазы; 3 - активация гликогенфосфорилазы адреналином через аденилатциклазную систему.
центрация Са2+ в клетке уменьшается после поступления сигнала к расслаблению мышц. Таким образом, роль ионов Са2+ заключается не только в инициации мышечного сокращения, но также в обеспечении его энергозатрат.
Активация киназы фосфорилазы с помощью ионов Са2+ опосредована кальмодулином. Каль-модулин в данном случае - прочно связанная субъединица фермента (рис. 7-32). Мышечная киназа фосфорилазы состоит из субъединиц 4 типов: α, β, γ и δ, объединённых в комплекс. Фермент включает 4 таких комплекса. Каталитической активностью обладает γ-субъединица. Субъединицы α и β выполняют регуляторную функцию. Они содержат остатки серина, фос-форилируемые ПК А. δ-Субъединица связывает 4 иона кальция; она идентична белку кальмо-
дулину. Связывание ионов кальция вызывает конформационные изменения, что приводит к активации каталитического центра γ-субъеди-ницы, хотя молекула остаётся в дефосфорили-рованном состоянии.
В мышцах в период пищеварения, если он совпадает с состоянием покоя, происходит стимуляция синтеза гликогена. Мышечная работа во время пищеварения замедляет процесс синтеза гликогена, так как при этом мышцы используют для окисления глюкозу крови, поступающую из кишечника.
В переключении мобилизации гликогена на запасание глюкозы участвует инсулин. Как уже говорилось, глюкоза поступает в мышечные и жировые клетки с помощью глюкозо-транс-портёров ГЛЮТ-4. Транспортёры в отсутствие
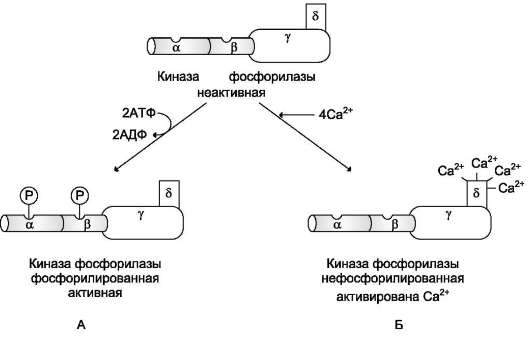
Рис. 7-32. Регуляция активности киназы фосфорилазы. Фермент состоит из 4 идентичных белковых комплексов. Каждый комплекс содержит 4 разных субъединицы α, β, γ, δ. На рисунке показан один из тетрамеров. Каталитической активностью обладает γ-субъединица. а- и β- протомеры выполняют регуляторную функцию, они фосфорилируются при участии ПК А. Кальмодулин - δ-субъединица, прочно связанная с ферментом. А - активация киназы фосфорилазы в результате фосфорилирования; Б - активация киназы фосфорилазы после присоединения Са2+ к кальмодулину.
инсулина находятся в цитоплазме клеток, и глюкоза клетками не используется, так как в мембране нет белков-переносчиков. Инсулин стимулирует перемещение ГЛЮТ-4 и встраивание их в мембрану клеток. Механизм подобного влияния инсулина изучен недостаточно, но определены его основные этапы. Цепь событий при стимуляции инсулином потребления глюкозы мышцами и жировыми клетками выглядит следующим образом:
• рецептор инсулина (IR) - инсулинстимули-руемая тирозиновая протеинкиназа - обязательный посредник всех действий инсулина (см. раздел 5);
• активированный инсулином IR фосфори-лирует специфические цитоплазматические белки - субстраты инсулина (IRS);
• фосфорилированный субстрат (в основном IRS-1) соединяется с фосфатидилинозитол-3-киназой (ФИ-3-киназа) и активирует этот фермент;
• активная ФИ-3-киназа катализирует фосфо-рилирование по позиции 3 ряд компонентов инозитолфосфатной сигнальной системы,
приводящей к стимуляции транслокации ГЛЮТ из цитозоля в плазматическую мембрану;
• глюкоза с помощью ГЛЮТ-4 поступает в мышечные клетки и включается в синтез гликогена.
Влияние инсулина на скорость синтеза гликогена в мышцах осуществляется посредством изменения активности гликогенсинтазы и гли-когенфосфорилазы - ключевых ферментов, о чём уже говорилось при обсуждении влияния инсулина на метаболизм гликогена в печени.
Д. НАРУШЕНИЯ ОБМЕНА ГЛИКОГЕНА
Гликогеновые болезни - группа наследственных нарушений, в основе которых лежит снижение или отсутствие активности ферментов, катализирующих реакции синтеза или распада гликогена, либо нарушение регуляции этих ферментов.
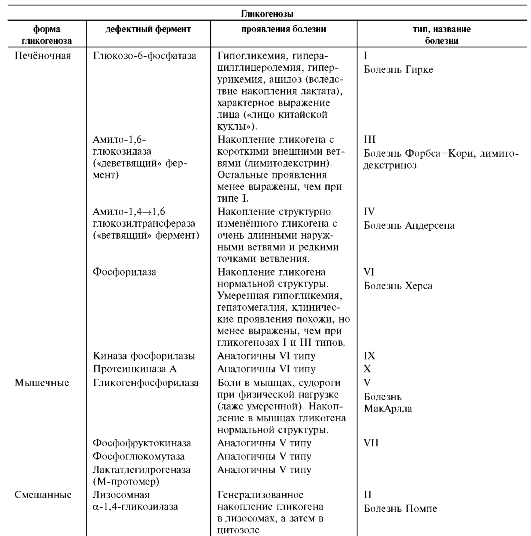
1. Гликогенозы - заболевания, обусловленные дефектом ферментов, участвующих в распаде гликогена. Они проявляются или необычной структурой гликогена, или его избыточным
накоплением в печени, сердечной или скелетных мышцах, почках, лёгких и других органах. В таблице 7-3 описаны некоторые типы глико-генозов, различающихся характером и локализацией ферментного дефекта.
Следует отметить, что термин «гликогеноз» был впервые предложен К.Ф. Кори и Г.Т. Кори.
Они же предложили систему нумерации этих болезней. Однако в настоящее время преобладает деление гликогенозов на 2 группы: печёночные и мышечные.
Печёночные формы гликогенозов ведут к нарушению использования гликогена для поддержания уровня глюкозы в крови. Поэтому общий
Таблица 7-3. Характеристика некоторых гликогеновых болезней
симптом для этих форм - гипогликемия в
постабсорбтивный период.
Болезнь Гирке (тип I) отмечают наиболее часто. Описание основных симптомов этого типа гликогеноза и их причин может служить основанием для понимания симптомов всех остальных типов. Причина этого заболевания - наследственный дефект глюкозо-6-фосфатазы - фермента, обеспечивающего выход глюкозы в кровоток после её высвобождения из гликогена клеток печени. Болезнь Гирке проявляется гипогликемией, гипертриацилглицеролемией (повышением содержания триацилглицеролов), гиперури-кемией (повышением содержания мочевой кислоты).
Гипогликемия - следствие нарушения реакции образования свободной глюкозы из глюкозо-6-фосфата. Кроме того, вследствие дефекта глюкозо-6-фосфатазы происходит накопление в клетках печени субстрата - глюкозо-6-фосфата, который вовлекается в процесс катаболизма, где он превращается в пируват и лактат. В крови повышается количество лактата, поэтому возможен ацидоз. В тяжёлых случаях результатом гипогликемии могут быть судороги. Гипогликемия сопровождается уменьшением содержания инсулина и снижением отношения инсулин/глюкагон, что, в свою очередь, ведёт к ускорению липолиза жировой ткани в результате действия глюкагона и выходу в кровь жирных кислот (см. раздел 8).
Гипертриацилглицеролемия возникает в результате снижения активности ЛП-липазы жировой ткани - фермента, активируемого инсулином и обеспечивающего усвоение ТАГ клетками жировой ткани (см. раздел 8).
Гиперурикемия возникает в результате следующих событий:
• увеличиваются содержание в клетках глюкозо-6-фосфата и его использование в пентозофосфатном пути с образованием рибозо-5-фосфата - субстрата для синтеза пуриновых нуклеотидов;
• увеличивается образование мочевой кислоты вследствие избыточного синтеза, а следовательно, и катаболизма пуриновых нуклеотидов, конечным продуктом которого является мочевая кислота.
• снижается выведение мочевой кислоты вследствие увеличения продукции лактата и изменения рН мочи в кислую сторону, что затрудняет выведение уратов - труднорастворимых солей мочевой кислоты.
При диагностике данной патологии определяют активность глюкозо-6-фосфатазы в биоптатах печени. Кроме того, используют тест со стимуляцией глюкагоном или адреналином, который в случае болезни даёт отрицательный результат, т.е. после инъекции гормона уровень глюкозы в крови изменяется незначительно.
Лечение состоит в ограничении употребления продуктов, содержащих глюкозу. Рекомендуется исключить из диеты продукты, содержащие сахарозу и лактозу, так как образующиеся из них галактоза и фруктоза после превращения в глюкозо-6-фосфат ведут к дальнейшему накоплению гликогена. Для предотвращения гипогликемии используют метод частого кормления. Этим можно предупредить симптомы гипогликемии.
Гликогеноз I типа наследуется по аутосомно-рецессивному типу. Уже в раннем периоде наиболее заметный признак - гепатомега-лия. У больных детей короткое туловище, большой живот, увеличены почки. Больные дети отстают в физическом развитии.
Описанное заболевание иногда обозначают как гликогеноз типа 1а, так как существует его разновидность - тип Ib. Гликогеноз Ib представляет собой редко встречающуюся патологию, которая характеризуется тем, что дефектен фермент транслоказа глюко-зо-6-фосфата, обеспечивающий транспорт фосфорилированной глюкозы в ЭР. Поэтому, несмотря на достаточную активность глюкозо-6-фосфатазы, отщепление неорганического фосфата и выход глюкозы в кровь нарушен. Клиническая картина гликогеноза типа Ib такая же, как при гликогенозе Iа.
Болезнь Кори (тип III) весьма распространена. Она составляет 1/4 всех случаев печёночных гликогенозов. Накапливаемый гликоген аномален по структуре, так как дефектен фермент амило-1,6-глюкозидаза, гидроли-зующий гликозидные связи в местах разветвлений («деветвящий фермент», от англ. debranching enzyme). Недостаток глюкозы в крови проявляется быстро, поскольку гли-
когенолиз возможен, но в незначительном объёме. В отличие от гликогеноза I типа, лактоацидоз и гиперурикемия не отмечаются. Болезнь отличается более лёгким течением.
Болезнь Андерсен (тип IV) - крайне редкое ауто-сомно-рецессивное заболевание, возникающее вследствие дефекта ветвящего фермента - амило-1,4-1,6-глюкозилтрансферазы. Содержание гликогена в печени не сильно увеличено, но структура его изменена, и это препятствует его распаду. Молекула гликогена имеет мало точек ветвления, а также очень длинные и редкие боковые ветви. В то же время гипогликемия выражена умеренно. Болезнь развивается быстро, отягощается ранним циррозом печени и практически не поддаётся лечению. Дефект фермента ветвления обнаруживается не только в печени, но также в лейкоцитах, мышцах, фиброблас-тах, но ранние и преобладающие проявления болезни обусловлены нарушением функции печени.
Болезнь Херса (тип VI) также проявляется симптомами, обусловленными поражением печени. Данный гликогеноз - следствие дефекта гликогенфосфорилазы. В гепато-цитах накапливается гликоген нормальной структуры. Течение болезни сходно с гли-коге-нозом I типа, но симптомы выражены в меньшей степени. Сниженная активность гликогенфосфорилазы обнаруживается также в лейкоцитах. Болезнь Херса - редкий тип гликогеноза; наследуется по аутосомно-рецессивному типу.
Дефект киназы фосфорилазы (тип IX) встречается только у мальчиков, так как этот признак сцеплен с Х-хромосомой.
Дефект протеинкиназы А (тип Х), так же как и дефект киназы фосфорилазы, проявляется симптомами, сходными с болезнью Херса.
Мышечные формы гликогенозов характеризуются нарушением в энергоснабжении скелетных мышц. Эти болезни проявляются при физических нагрузках и сопровождаются болями и судорогами в мышцах, слабостью и быстрой утомляемостью.
Болезнь МакАрдла (тип V) - аутосомно-рецес-сивная патология, при которой полностью отсутствует в скелетных мышцах активность
гликогенфосфорилазы. Поскольку активность этого фермента в гепатоцитах нормальная, то гипогликемия не наблюдается (строение фермента в печени и мышцах кодируются разными генами). Тяжёлые физические нагрузки плохо переносятся и могут сопровождаться судорогами, однако при физических нагрузках гиперпродукция лактата не наблюдается, что подчёркивает значение внемышечных источников энергии для сокращения мышц, например, таких как жирные кислоты, замещающие при данной патологии глюкозу (см. раздел 8). Хотя болезнь не сцеплена с полом, большая частота заболевания характерна для мужчин.
Дефект фосфофруктокиназы характерен для гли-когеноза VII типа. Больные могут выполнять умеренные физические нагрузки. Течение болезни сходно с гликогенозом V типа, но основные проявления менее выражены.
Дефект фосфоглицеромутазы и дефект М-субъеди-ницы ЛДГ (ненумерованные по классификации Кори, см. табл. 7-3) характерны для мышечных форм гликогенозов. Проявления этих патологий аналогичны болезни МакАр-дла. Дефект фосфоглицеромутазы в мышцах описан только у одного больного.
2. Агликогенозы
Агликогеноз (гликогеноз 0 по классификации) - заболевание, возникающее в результате дефекта гликогенсинтазы. В печени и других тканях больных наблюдают очень низкое содержание гликогена. Это проявляется резко выраженной гипогликемией в постабсорбтивном периоде. Характерный симптом - судороги, проявляющиеся особенно по утрам. Болезнь совместима с жизнью, но больные дети нуждаются в частом кормлении.
viii. катаболизм глюкозы
Катаболизм глюкозы - основной поставщик энергии для процессов жизнедеятельности организма.
ОСНОВНЫЕ ПУТИ КАТАБОЛИЗМА ГЛЮКОЗЫ
А. Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6 О2 → 6 СО2 + 6Н2О + 2820 кДж/моль. Этот процесс включает несколько стадий
(рис. 7-33).
• Аэробный гликолиз - процесс окисления глюкозы с образованием двух молекул пи-рувата;
• Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его дальнейшее окисление в цитратном цикле;
• ЦПЭ, сопряжённая с реакциями дегидрирования, происходящими в процессе распада глюкозы.
Б. Анаэробный распад (анаэробный гликолиз)
В определённых ситуациях обеспечение кислородом тканей может не соответствовать их потребностям. Например, на начальных стадиях интенсивной мышечной работы при стрессе сердечные сокращения могут не достигать нужной частоты, а потребности мышц в кислороде для аэробного распада глюкозы велики. В подобных случаях включается процесс, который протекает без кислорода и заканчивается образованием лак-тата из пировиноградной кислоты. Этот процесс называют анаэробным распадом, или анаэробным гликолизом. Анаэробный распад глюкозы энергетически малоэффективен, но именно этот процесс может стать единственным источником энергии для мышечной клетки в описанной ситуации. В дальнейшем, когда снабжение мышц кислородом будет достаточным в результате перехода сердца на ускоренный ритм, анаэробный распад переключается на аэробный. Пути катаболизма глюкозы и их энергетический эффект показаны на рис. 7-34.
АЭРОБНЫЙ ГЛИКОЛИЗ
Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.
1. Этапы аэробного гликолиза
В аэробном гликолизе можно выделить 2 этапа.
1. Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия
реакций протекает с использованием 2 молекул АТФ. 2. Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфот-риозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
2. Реакции аэробного гликолиза
Превращение глюкозо-6-фосфата в 2 молекулы гли-церальдегид-3-фосфата
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с участием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Эта обратимая реакция изомеризации протекает под действием фермента фосфоглюкоизомеразы.
Затем следует ещё одна реакция фосфо-рилирования с использованием фосфатного остатка и энергии АТФ. В ходе этой реакции, катализируемой фосфофруктокиназой, фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Данная реакция, так же, как гексокиназная, практически необратима, и, кроме того, она наиболее медленная из всех реакций гликолиза. Реакция, катализируемая фосфофруктокиназой, определяет скорость всего гликолиза, поэтому, регулируя активность фосфофруктокиназы, можно изменять скорость катаболизма глюкозы.
Фруктозо-1,6-бисфосфат далее расщепляется на 2 триозофосфата: глицеральдегид-3-фосфат и дигидроксиацетонфосфат. Реакцию катализирует фермент фруктозобисфосфатальдолаза, или просто альдолаза. Этот фермент катализирует как реакцию альдольного расщепления, так и аль-дольной конденсации, т.е. обратимую реакцию. Продукты реакции альдольного расщепления - изомеры. В последующих реакциях гликолиза используется только глицеральдегид-3-фосфат, поэтому дигидроксиацетонфосфат превращается с участием фермента триозофосфатизомеразы в глицероальдегид-3-фосфат (рис. 7-35).
В описанной серии реакций дважды происходит фосфорилирование с использованием АТФ. Однако расходование двух молекул АТФ (на одну молекулу глюкозы) далее будет компенсировано синтезом большего количества
АТФ.
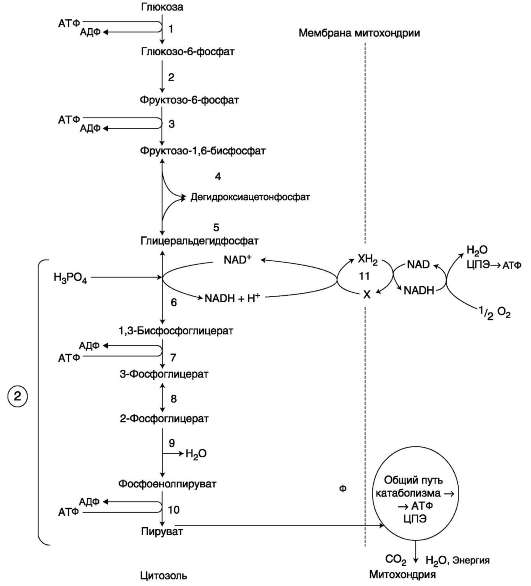
Рис. 7-33. Аэробный распад глюкозы. 1 - 10 - реакции аэробного гликолиза; 11 - малат-аспартатный челночный механизм транспорта водорода в митохондрии; 2 (в кружке) - стехиометрический коэффициент.
Рис. 7-34. Пути катаболизма глюкозы. 1 - аэробный гликолиз; 2, 3 - общий путь катаболизма; 4 - аэробный распад глюкозы; 5 - анаэробный распад глюкозы (в рамке); 2 (в кружке) - стехиометрический коэффициент.
Превращение глицеральдегид-3-фосфата в пируват
Эта часть аэробного гликолиза включает реакции, связанные с синтезом АТФ. Наиболее сложной в данной серии реакций является реакция превращения глицеральдегид-3-фосфата в 1,3-бисфосфоглицерат. Это превращение - первая реакция окисления в ходе гликолиза. Реакцию катализирует глицеральдегид-3-фосфатдегидрогена-за, которая является NАD-зависимым ферментом. Значение данной реакции заключается не только в том, что образуется восстановленный кофермент, окисление которого в дыхательной цепи сопряжено с синтезом АТФ, но также и в том, что свободная энергия окисления концентрируется в макроэргической связи продукта реакции. Глицеральдегид-3-фосфатдегидро-геназа содержит в активном центре остаток цистеина, сульфгидрильная группа которого принимает непосредственное участие в катализе. Окисление глицеральдегид-3-фосфата приводит к восстановлению NAD и образованию с участием Н3РО4 высокоэнергетической ангидридной связи в 1,3-бисфосфоглицерате в положении 1. В следующей реакции высокоэнергетический фосфат передаётся на АДФ с образованием АТФ. Фермент, катализирующий это превращение, назван по обратной реакции
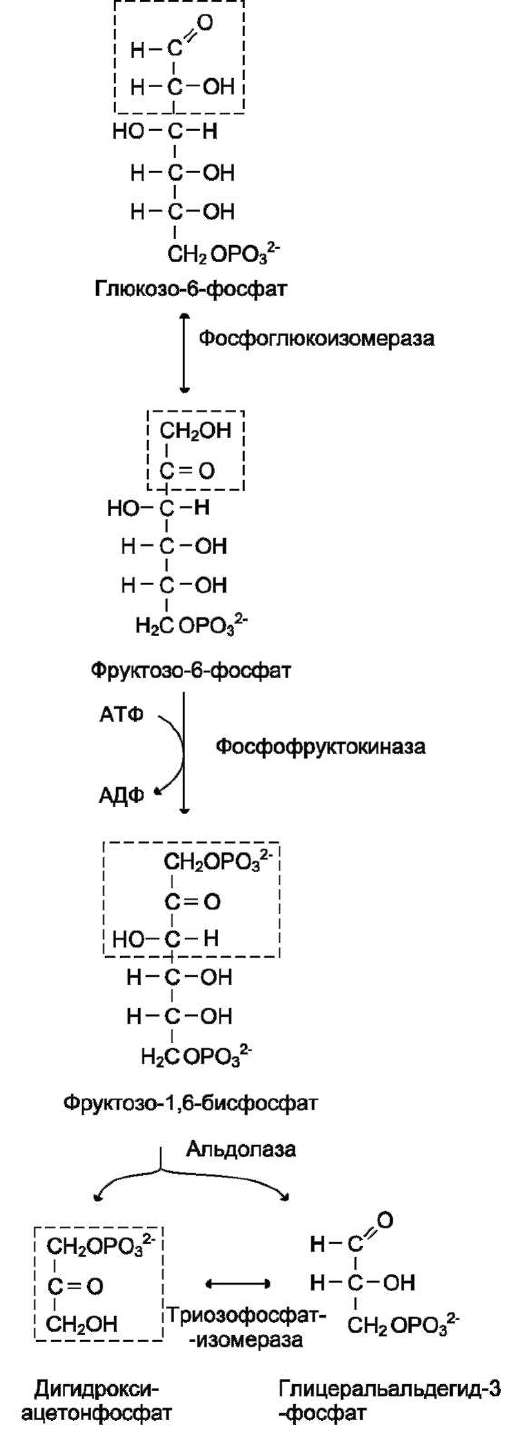
Рис. 7-35. Превращение глюкозо-6-фосфата в три-озофосфаты.
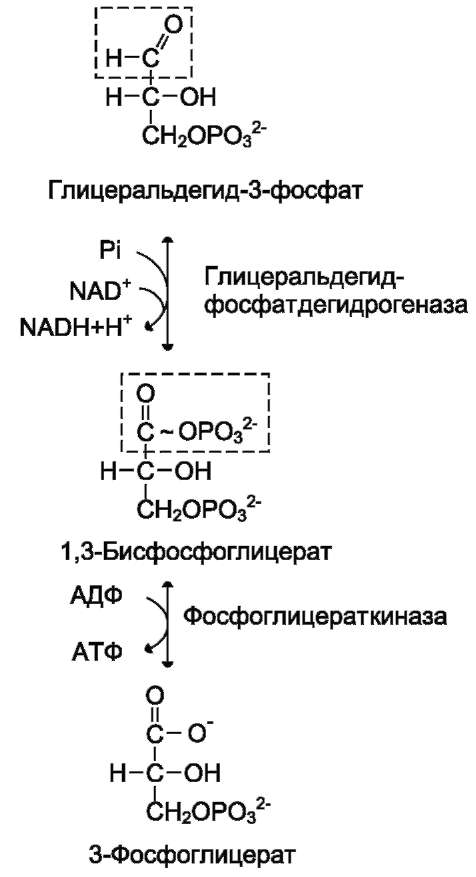
Рис. 7-36. Превращение глицеральдегид-3-фосфата в 3-фосфоглицерат.
фосфоглицераткиназой (киназы называются по субстрату, находящемуся в уравнении реакции по одну сторону с АТФ). Данная серия реакций показана на рис. 7-36.
Образование АТФ описанным способом не связано с дыхательной цепью, и его называют субстратным фосфорилированием АДФ. Образованный 3-фосфоглицерат уже не содержит макроэргической связи. В следующих реакциях происходят внутримолекулярные перестройки, смысл которых сводится к тому, что низкоэнергетический фосфоэфир переходит в соединение, содержащее высокоэнергетический фосфат. Внутримолекулярные преобразования заключаются в переносе фосфатного остатка из положения 3 в фосфоглицерате в положение 2. Затем от образовавшегося 2-фосфоглицерата отщепляется молекула воды при участии фермента енолазы. Название дегидратирующего фермента дано по обратной реакции. В результа-
те реакции образуется замещённый енол - фос-фоенолпируват. Образованный фосфоенолпи-руват - макроэргическое соединение, фосфатная группа которого переносится в следующей реакции на АДФ при участии пируваткиназы (фермент также назван по обратной реакции, в которой происходит фосфорилирование пи-рувата, хотя подобная реакция в таком виде не имеет места).
Превращение фосфоенолпирувата в пиру-ват - необратимая реакция. Это вторая в ходе гликолиза реакция субстратного фосфорилиро-вания. Образующаяся енольная форма пирувата затем неферментативно переходит в более термодинамически стабильную кетоформу. Описанная серия реакций представлена на рис. 7-37.
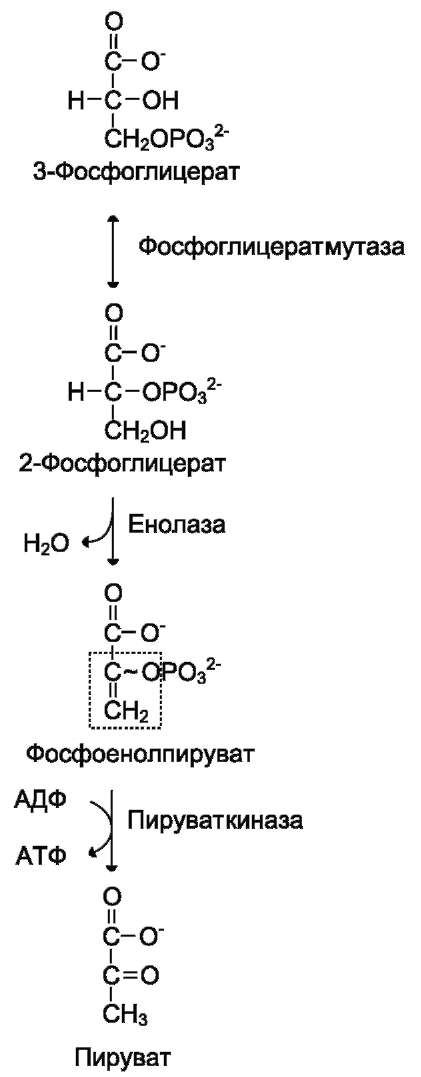
Рис. 7-37. Превращение 3-фосфоглицерата в пируват.
Схема 10 реакций, протекающих при аэробном гликолизе, и дальнейшее окисление пирувата представлены на рис. 7-33.
3. Окисление цитоплазматического NADH в митохондриальной дыхательной цепи. Челночные системы
NADH, образующийся при окислении гли-церальдегид-3-фосфата в аэробном гликолизе, подвергается окислению путём переноса атомов водорода в митохондриальную дыхательную цепь. Однако цитозольный NADH не способен передавать водород на дыхательную цепь, потому что митохондриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных систем, называемых «челночными». В этих системах водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е. с обеих сторон митохондриальной мембраны находится специфическая дегидрогеназа. Известны 2 челночные системы. В первой из этих систем водород от NADH в цитозоле передаётся на дигидроксиа-цетонфосфат ферментом глицерол-3-фосфатде-гидрогеназой (NAD-зависимый фермент, назван по обратной реакции). Образованный в ходе этой реакции глицерол-3-фосфат, окисляется далее ферментом внутренней мембраны митохондрий - глицерол-3-фосфатдегидрогеназой (FAD-зависимым ферментом). Затем протоны и электроны с FADH2 переходят на убихинон и далее по ЦПЭ (рис. 7-38).
Глицеролфосфатная челночная система работает в клетках белых мышц и гепатоцитов. Однако в клетках сердечных мышц митохондриальная глицерол-3-фосфатдегидрогеназа отсутствует. Вторая челночная система, в которой участвуют малат, цитозольная и митохондриальная малат-дегидрогеназы, является более универсальной. В цитоплазме NADH восстанавливает оксалоаце-тат в малат (рис. 7-39, реакция 1), который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат NAD-зависимой малатдегидрогеназой (реакция 2). Восстановленный в ходе этой реакции NAD отдаёт водород в митохондриальную ЦПЭ. Однако образованный из малата оксалоацетат выйти самостоятельно из митохондрий в цитозоль не может, так как мембрана митохондрий для него непроницаема. Поэтому оксалоацетат превращается в аспартат, который и транспортируется в цитозоль, где снова превращается в оксалоацетат. Превращения оксалоацетата в аспартат и обратно связаны с присоединением и отщеплением аминогруппы (реакции трансаминирования, см. раздел 9). Эта челночная система называется малат-аспартат-ной (рис. 7-39). Результат её работы - регенерация цитоплазматического NAD+ из NADH
Обе челночные системы существенно отличаются по количеству синтезированного АТФ. В первой системе соотношение Р/O равно 2, так как водород вводится в ЦПЭ на уровне KoQ. Вторая система энергетически более эффективна, так как передаёт водород в ЦПЭ через митохондриальный NAD+ и соотношение Р/О близко к 3.
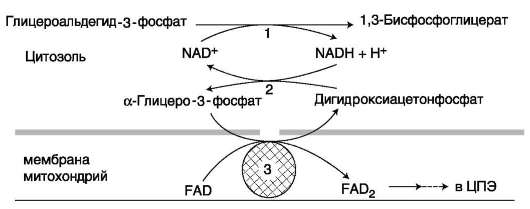
Рис. 7-38. Глицерофосфатная челночная система. 1 - глицеральдегид-3-фосфатдегидрогеназа; 2 - глице-рол-3-фосфатдегидрогеназа (цитозольный фермент, назван по обратной реакции); 3 - глицерол-3-фосфатде-гидрогеназа (митохондриальный флавиновый фермент).
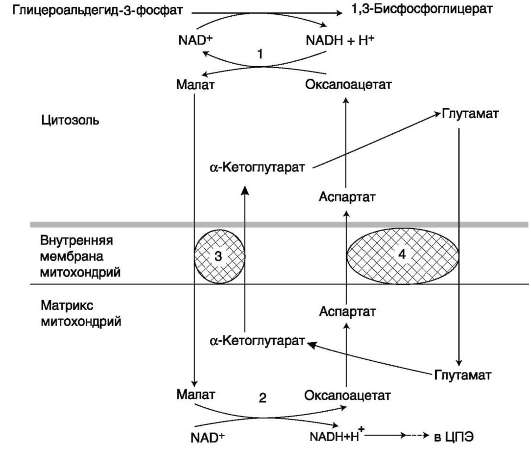
Рис. 7-39. Малат-аспартатная челночная система. 1, 2 - окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ; 3, 4 - транслоказы, обеспечивающие транспорт a-кетоглутарата, аспартата и глутамата и через мембрану митохондрий.
4. Баланс АТФ при аэробном гликолизе и распаде глюкозы до СО2 и Н2О
Выход АТФ при аэробном гликолизе
На образование фруктозо-1,6-бисфосфата из одной молекулы глюкозы требуется 2 молекулы АТФ (реакции 1 и 3 на рис. 7-33). Реакции, связанные с синтезом АТФ, происходят после распада глюкозы на 2 молекулы фосфотриозы, т.е. на втором этапе гликолиза. На этом этапе происходят 2 реакции субстратного фосфорилирования и синтезируются 2 молекулы АТФ (реакции 7 и 10). Кроме того, одна молекула глицеральдегид-3-фосфата дегидрируется (реакция 6), а NADH передаёт водород в митохондриальную ЦПЭ, где синтезируется 3 молекулы АТФ путём окислительного фосфо-рилирования. В данном случае количество АТФ
(3 или 2) зависит от типа челночной системы. Следовательно, окисление до пирувата одной молекулы глицеральдегид-3-фосфата сопряжено с синтезом 5 молекул АТФ. Учитывая, что из глюкозы образуются 2 молекулы фосфотриозы, полученную величину нужно умножить на 2 и затем вычесть 2 молекулы АТФ, затраченные на первом этапе. Таким образом, выход АТФ при аэробном гликолизе составляет (5x2) - 2 =
8 АТФ.
Выход АТФ при аэробном распаде глюкозы до конечных продуктов
В результате гликолиза образуется пируват, который далее окисляется до СО2 и Н2О в ОПК, описанном в разделе 6. Теперь можно оценить энергетическую эффективность гликолиза и
Таблица 7-4. Этапы аэробного распада глюкозы
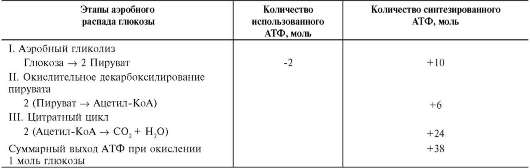
ОПК, которые вместе составляют процесс аэробного распада глюкозы до конечных продуктов (табл. 7-4).
Таким образом, выход АТФ при окислении 1 моль глюкозы до СО2 и Н2О составляет
38 моль АТФ.
В процессе аэробного распада глюкозы происходят 6 реакций дегидрирования. Одна из них протекает в гликолизе и 5 в ОПК (см. раздел 6). Субстраты для специфических NAD-зависимых дегидрогеназ: глицеральдегид-3-фосфат, пируват, изоцитрат, α-кетоглутарат, малат. Одна реакция дегидрирования в цитратном цикле под действием сукцинатдегидрогеназы происходит с участием кофермента FAD. Общее количество АТФ, синтезированное путём окислительного фофорилирования, составляет 17 моль АТФ на 1 моль глицеральдегидфосфата. К этому необходимо прибавить 3 моль АТФ, синтезированных путём субстратного фосфорилирования (две реакции в гликолизе и одна в цитратном цикле).
Учитывая, что глюкоза распадается на 2 фосфотриозы и что стехиометрический коэффициент дальнейших превращений равен 2, полученную величину надо умножить на 2, а из результата вычесть 2 моль АТФ, использованные на первом этапе гликолиза.
АНАЭРОБНЫЙ РАСПАД ГЛЮКОЗЫ (АНАЭРОБНЫЙ ГЛИКОЛИЗ)
Анаэробным гликолизом называют процесс расщепления глюкозы с образованием в качестве конечного продукта лактата. Этот процесс протекает без использования кислорода и поэтому
не зависит от работы митохондриальной дыхательной цепи. АТФ образуется за счёт реакций субстратного фосфорилирования. Суммарное уравнение процесса:
С6Н1206 + 2 Н3РО4 + 2 АДФ = 2 С3Н6О3 + 2 АТФ + 2 Н2О.
1. Реакции анаэробного гликолиза
При анаэробном гликолизе (рис. 7-40) в ци-тозоле протекают все 10 реакций, идентичных аэробному гликолизу. Лишь 11-я реакция, где происходит восстановление пирувата цитозоль-ным NADH является специфической для анаэробного гликолиза (рис. 7-41). Восстановление пирувата в лактат катализирует лактатдегидроге-наза (реакция обратимая, и фермент назван по обратной реакции). С помощью этой реакции обеспечивается регенерация NAD+ из NADН без участия митохондриальной дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от NADН (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию NAD+. К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма. Это вещество выводится в кровь и утилизируется, превращаясь в печени в глюкозу, или при доступности кислорода превращается в пируват, который вступает в общий путь катаболизма, окисляясь до СО2 и Н2О. Строение лактатдегидрогеназы, механизм действия и значение определения активности
Рис. 7-40. Анаэробный гликолиз.
этого фермента для диагностики заболеваний описывались ранее в разделе 2.
Баланс АТФ при анаэробном гликолизе
Анаэробный гликолиз по сравнению с аэробным менее эффективен. В этом процессе катаболизм 1 моль глюкозы без участия мито-хондриальной дыхательной цепи сопровождается синтезом 2 моль АТФ и 2 моль лактата. АТФ образуется за счёт 2 реакций субстратного фосфорилирования. Поскольку глюкоза распадается на 2 фосфотриозы, то с учётом стехиометрического коэффициента, равного 2, количество моль синтезированного АТФ равно 4. Учитывая 2 моль АТФ, использованных на первом этапе гликолиза, получаем конечный энергетический эффект процесса, равный 2 моль АТФ. Таким образом, 10 цитозольных ферментов, катализирующих превращение глюкозы в пируват, вместе с лактатдегидрогеназой обеспечивают в анаэробном гликолизе синтез 2 моль АТФ (на 1 моль глюкозы) без участия кислорода.
Пируват Лактат
Рис. 7-41. Восстановление пирувата в лактат.
ЗНАЧЕНИЕ КАТАБОЛИЗМА ГЛЮКОЗЫ
Основное физиологическое назначение катаболизма глюкозы заключается в использовании энергии, освобождающейся в этом процессе для синтеза АТФ.
Энергия, выделяющаяся в процессе полного распада глюкозы до СО2 и Н2О, составляет 2880 кДж/моль. Если эту величину сравнить с энергией гидролиза высокоэнергетических связей - 38 моль АТФ (50 кДж на моль АТФ), то получим: 50x38 = 1900 кДж, что составляет 65% от всей энергии, выделяющейся при полном распаде глюкозы. Такова эффективность использования энергии распада глюкозы для синтеза АТФ. Необходимо учитывать, что реальная эффективность процесса может быть ниже. Точно оценить выход АТФ можно только при субстратном фосфорилировании, а соотношение между поступлением водорода в дыхательную цепь и синтезом АТФ является приблизительным.
Аэробный распад глюкозы
происходит во многих органах и тканях и служит основным, хотя и не
единственным, источником энергии для жизнедеятельности. Некоторые ткани
находятся в наибольшей зависимости от катаболизма глюкозы как
источника энергии. Например, клетки мозга расходуют до
Анаэробный распад глюкозы происходит в мышцах, в первые минуты мышечной работы, в эритроцитах (в которых отсутствуют митохондрии), а также в разных органах в условиях ограниченного снабжении их кислородом, в том числе в клетках опухолей. Для метаболиз-
ма клеток опухолей характерно ускорение как аэробного, так и анаэробного гликолиза. Но преимущественный анаэробный гликолиз и увеличение синтеза лактата служит показателем повышенной скорости деления клеток при недостаточной обеспеченности их системой кровеносных сосудов.
Кроме энергетической функции, процесс катаболизма глюкозы может выполнять и анаболические функции. Метаболиты гликолиза используются для синтеза новых соединений. Так, фруктозо-6-фосфат и глицеральдегид-3-фосфат участвуют в образовании рибозо-5-фос-фата - структурного компонента нуклеотидов; 3-фосфоглицерат может включаться в синтез аминокислот, таких как серин, глицин, цистеин (см. раздел 9). В печени и жировой ткани ацетил-КоА, образующийся из пирувата, используется как субстрат при биосинтезе жирных кислот, холестерина, а дигидроксиацетонфосфат как субстрат для синтеза глицерол-3-фосфата (см. раздел 8).
регуляция катаболизма глюкозы
Поскольку основное значение гликолиза состоит в синтезе АТФ, его скорость должна коррелировать с затратами энергии в организме.
Большинство реакций гликолиза обратимы, за исключением трёх, катализируемых гексоки-назой (или глюкокиназой), фосфофруктокина-зой и пируваткиназой. Регуляторные факторы, изменяющие скорость гликолиза, а значит и образование АТФ, направлены на необратимые реакции. Показателем потребления АТФ является накопление АДФ и АМФ. Последний образуется в реакции, катализируемой адени-латкиназой:
2 АДФ ↔ АМФ + АТФ
Даже небольшой расход АТФ ведёт к заметному увеличению АМФ. Отношение уровня АТФ к АДФ и АМФ характеризует энергетический статус клетки (см. раздел 6), а его составляющие служат аллостерическими регуляторами скорости как общего пути катаболизма, так и гликолиза. На рисунке 7-42 показана аллостери-ческая регуляция скорости катаболизма глюкозы в скелетных мышцах.
Существенное значение для регуляции гликолиза имеет изменение активности фосфофрук-
токиназы, потому что этот фермент, как упоминалось ранее, катализирует наиболее медленную реакцию процесса.
Фосфофруктокиназа активируется АМФ, но ингибируется АТФ. АМФ, связываясь с аллосте-рическим центром фосфофруктокиназы, увеличивает сродство фермента к фруктозо-6-фосфату и повышает скорость его фосфорилирования. Эффект АТФ на этот фермент - пример гомот-ропного аллостеризма (см. раздел 2), поскольку АТФ может взаимодействовать как с аллостери-ческим, так и с активным центром, в последнем случае как субстрат.
При физиологических значениях АТФ активный центр фосфофруктокиназы всегда насыщен субстратами (в том числе АТФ). Повышение уровня АТФ относительно АДФ снижает скорость реакции, поскольку АТФ в этих условиях действует как ингибитор: связывается с аллосте-рическим центром фермента, вызывает конфор-мационные изменения и уменьшает сродство к его субстратам.
Изменение активности фосфофруктокиназы способствует регуляции скорости фосфори-лирования глюкозы гексокиназой. Снижение активности фосфофруктокиназы при высоком уровне АТФ ведёт к накоплению как фруктозо-6-фосфата, так и глюкозо-6-фосфата, а последний ингибирует гексокиназу. Следует напомнить, что гексокиназа во многих тканях ингибируется глюкозо-6-фосфатом.
При высоком уровне АТФ снижается скорость цикла лимонной кислоты и дыхательной цепи. В этих условиях процесс гликолиза также замедляется. Следует напомнить, что аллостерическая регуляция ферментов ОПК и дыхательной цепи также связана с изменением концентрации таких ключевых продуктов, как NADH, АТФ и некоторых метаболитов. Так, NADH, накапливаясь в том случае, если не успевает окислиться в дыхательной цепи, ингибирует некоторые аллостерические ферменты цитратного цикла (см. раздел 6).
Физиологическая роль гликолиза в печени и жировой ткани несколько иная, чем в других тканях. В печени и жировой ткани гликолиз в период пищеварения функционирует в основном как источник субстратов для синтеза жиров. Регуляция гликолиза в печени имеет свои особенности и будет рассмотрена позже.
Рис. 7-42. Регуляция катаболизма глюкозы в скелетных мышцах.
Е. 2,3-БИСФОСФОГЛИЦЕРАТНЫЙ ЦИКЛ
В гликолитическом пути может протекать дополнительная реакция, катализируемая бисфос-фоглицератмутазой, превращающей 1,3-бисфос-фоглицерат в 2,3-бисфосфоглицерат (2,3-БФГ), который может при участии 2,3-бисфосфогли-цератфосфатазы превращаться в 3-фосфоглице-рат - метаболит гликолиза (рис. 7-43).
В большинстве тканей 2,3-БФГ образуется в небольших количествах. В эритроцитах этот метаболит образуется в значительных количествах и выполняет роль аллостерического регулятора функции гемоглобина. 2,3-БФГ, связываясь с гемоглобином, понижает его сродство к кислороду, способствует диссоциации кислорода и переходу его в ткани (см. раздел 1).
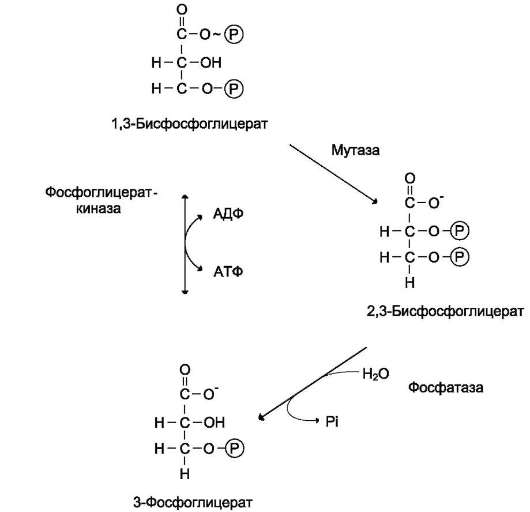
Рис. 7-43. Образование и превращение 2,3-бисфосфоглицерата.
Образование 2,3-БФГ предполагает потерю энергии макроэргической связи в 1,3-бис-фосфоглицерате, которая не переносится на АТФ, а рассеивается в форме теплоты, что означает снижение энергетического эффекта гликолиза.
ix. синтез глюкозы
в печени (глюконеогенез)
Некоторые ткани, например мозг, нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счёт расщепления гликогена в печени. Однако запасы гликогена в печени невелики. Они значительно уменьшаются к 6-10 ч голодания и практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается синтез глюкозы de novo - глюконеогенез. Глюконе-
огенез - процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Это объясняется тем, что клетки мозга не способны, в отличие от других тканей, обеспечивать потребности в энергии за счёт окисления жирных кислот (см. раздел 8).
Кроме мозга, в глюкозе нуждаются ткани и клетки, в которых аэробный путь распада невозможен или ограничен, например эритроциты (они лишены митохондрий), клетки сетчатки, мозгового слоя надпочечников и др.
Первичные субстраты глюконеогенеза - лак-тат, аминокислоты и глицерол. Включение этих
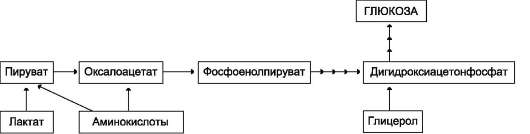
Рис. 7-44. Включение субстратов в глюконеогенез.
субстратов в глюконеогенез зависит от физиологического состояния организма.
• Лактат - продукт анаэробного гликолиза. Он образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
• Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке.
• Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
На рисунке 7-44 показаны пункты включения первичных субстратов в глюконеогенез.
А. РЕАКЦИИ ГЛЮКОНЕОГЕНЕЗА
Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза (рис. 7-45, реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. Однако 3 реакции гликолиза термодинамически необратимы. На этих стадиях реакции глюконеогенеза протекают другими путями.
Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях.
Рассмотрим более подробно те реакции глю-конеогенеза, которые отличаются от реакций гликолиза и происходят в глюконеогенезе с использованием других ферментов. Рассмотрим процесс синтеза глюкозы из пирувата.
1. Образование фосфоенолпирувата из пирувата - первая из необратимых стадий глюконеогенеза
Образование фосфоенолпирувата из пирува-та происходит в ходе двух реакций (рис. 7-45,
реакции 11, 12), первая из которых протекает в митохондриях. Пируват, образующийся из лактата или из некоторых аминокислот, транспортируется в матрикс митохондрий и там карбоксилиру-ется с образованием оксалоацетата (рис. 7-46). Пируваткарбоксилаза, катализирующая данную реакцию, - митохондриальный фермент, ко-ферментом которого является биотин. Реакция протекает с использованием АТФ.
Дальнейшие превращения оксалоацетата протекают в цитозоле. Следовательно, на этом этапе должна существовать система транспорта окса-лоацетата через митохондриальную мембрану, которая для него непроницаема. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием малата (рис. 7-47) при участии NADH (обратная реакция цитратного цикла). Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспар-татного челночного механизма, рассмотренного ранее (рис. 7-39).
В цитозоле малат вновь превращается в окса-лоацетат в ходе реакции окисления с участием кофермента NАD+. Обе реакции: восстановление оксалоацетата и окисление малата катализируют малатдегидрогеназа, но в первом случае это митохондриальный фермент, а во втором - ци-тозольный. Образованный в цитозоле из малата оксалоацетат затем превращается в фосфое-нолпируват в ходе реакции, катализируемой фосфоенолпируваткарбоксикиназой - ГТФ-зависимым ферментом (рис. 7-48). Название фермента дано по обратной реакции.
Схема всех реакций, протекающих на первой необратимой стадии глюконеогенеза, представлена на рис. 7-49.
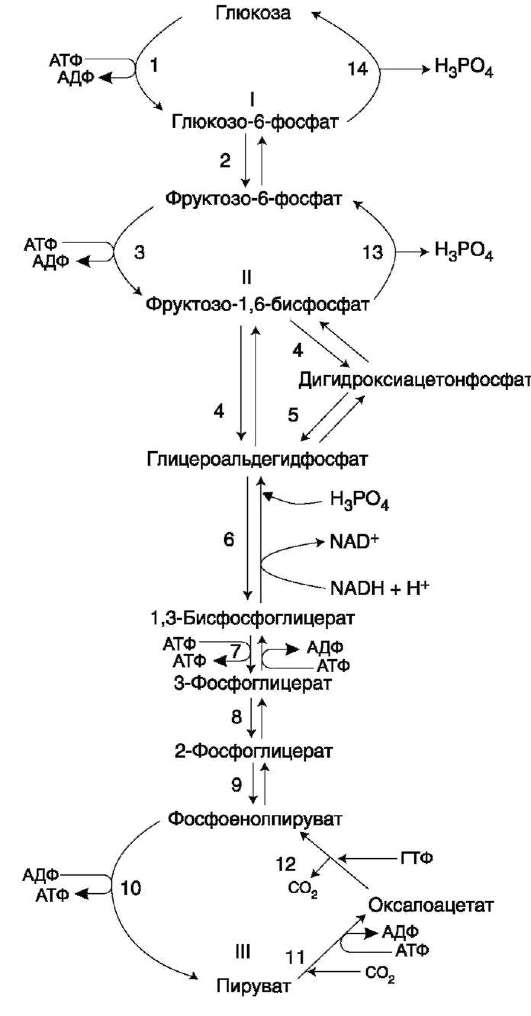
Рис. 7-45. Гликолиз и глюконеогенез. Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 - фосфоглюкоизомераза; 4 - альдолаза; 5 - три-озофосфатизомераза; 6 - глицеральдегидфосфатде-гидрогеназа; 7 - фосфоглицераткиназа; 8 - фосфог-лицератмутаза; 9 - енолаза. Ферменты необратимых реакций глюконеогенеза: 11 - пируваткарбоксилаза; 12 - фосфоенолпируваткарбоксикиназа; 13 - фрук-тозо-1,6-бисфосфатаза; 14 - глюкозо-6-фосфатаза. I-III - субстратные циклы.
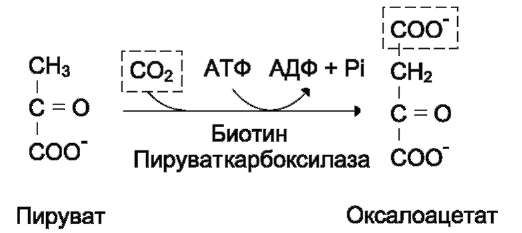
Рис. 7-46. Образование оксалоацетата из пи-рувата.
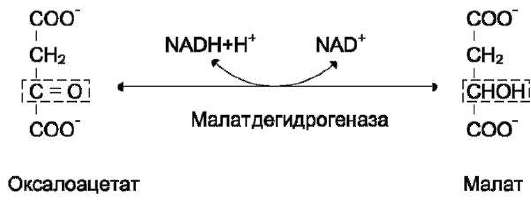
Рис. 7-47. Превращение оксалоацетата в ма-лат.
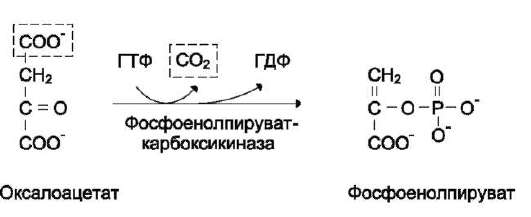
Рис. 7-48. Превращение оксалоацетата в фосфо-енолпируват.
Следует отметить, что этот обходной участок глюконеогенеза требует расхода двух молекул с макроэргическими связями (АТФ и ГТФ) в расчёте на одну молекулу исходного вещества - пирувата. В пересчёте на синтез одной молекулы глюкозы из двух молекул пирувата расход составляет 2 моль АТФ и 2 моль ГТФ или 4 моль АТФ (для удобства рассуждений предлагается считать, что энергозатраты на синтез АТФ и ГТФ равны). После образования фосфоенолпирувата все остальные реакции также протекают в цитозоле вплоть до образования фруктозо-1,6-бисфосфата и катализируются гликолитическими ферментами.
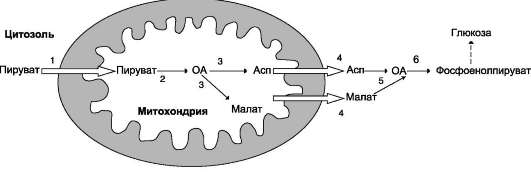
Рис. 7-49. Образование оксалоацетата, транспорт в цитозоль и превращение в фосфоенолпируват. 1 - транс-порт пирувата из цитозоля в митохондрию; 2 - превращение пирувата в оксалоацетат (ОА); 3 - превращение ОА в малат или аспартат; 4 - транспорт аспартата и малата из митохондрии в цитозоль; 5 - превращение аспартата и малата в ОА; 6 - превращение ОА в фосфоенолпируват.
2. Гидролиз фруктозо-1,6-бисфосфата и глюкозо-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфата - также необратимые реакции глюконеогенеза. В ходе гликолиза эти реакции катализируют специфические киназы с использованием энергии АТФ. В глюконеогенезе они протекают без участия АТФ и АДФ и ускоряются не киназами, а фос-фатазами - ферментами, принадлежащими к классу гидролаз. Ферменты фруктозо-1,6-бис-фосфатаза и глюкозо-6-фосфатаза катализируют отщепление фосфатной группы от фруктозо-1,6-бисфосфата и глюкозо-6-фосфата. После чего свободная глюкоза выходит из клетки в кровь. Схема всех реакций глюконеогенеза представлена на рис. 7-45.
Итак, в печени существуют 4 фермента, которые принимают участие только в глюконеогенезе и катализируют обходные реакции необратимых стадий гликолиза. Это - пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза.
3. Энергетический баланс глюконеогенеза из пирувата
В ходе этого процесса расходуются 6 моль АТФ на синтез 1 моль глюкозы из 2 моль пирувата. Четыре моль АТФ расходуются на стадии синтеза фосфоенолпирувата из оксало-ацетата и ещё 2 моль АТФ на стадиях обра-
зования 1,3-бисфосфоглицерата из 3-фосфо-глицерата.
Суммарный результат глюконеогенеза из пи-рувата выражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NАDН+ Н+) + 4 H2O → Глюкоза + 4 АДФ + 2 ГДФ + 6 Н3РО4 + 2 NАD+.
Б. СИНТЕЗ ГЛЮКОЗЫ ИЗ ЛАКТАТА
Лактат, образованный в анаэробном гликолизе, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват. Лактат как источник пирувата важен не столько при голодании, сколько при нормальной жизнедеятельности организма. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата.
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NАDН/NАD+ ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном направлении, т.е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют «глюкозо-лактатным циклом», или «циклом Кори» (рис. 7-50).
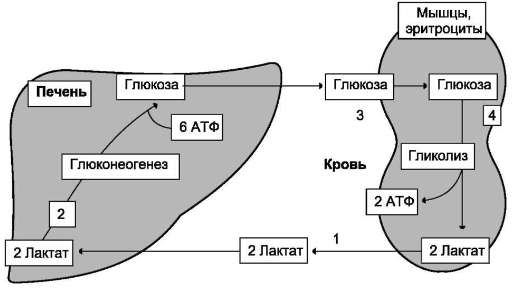
Рис. 7-50. Цикл Кори (глюкозо-лактатный цикл). 1 - поступление лактата из сокращающейся мышцы с током крови в печень; 2 - синтез глюкозы из лактата в печени; 3 - поступление глюкозы из печени с током крови в работающую мышцу; 4 - использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
Цикл Кори выполняет 2 важнейшие функции: 1 - обеспечивает утилизацию лактата; 2 - предотвращает накопление лактата и, как следствие этого, опасное снижение рН (лактоацидоз). Часть пирувата, образованного из лактата, окисляется печенью до СО2 и Н2О. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза.
Лактоацидоз. Термин «ацидоз» обозначает увеличение кислотности среды организма (снижение рН) до значений, выходящих за пределы нормы. При ацидозе либо увеличивается продукция протонов, либо происходит снижение их экскреции (в некоторых случаях и то и другое). Метаболический ацидоз возникает при увеличении концентрации промежуточных продуктов обмена (кислотного характера) вследствие увеличения их синтеза или уменьшения скорости распада или выведения. При нарушении кислотно-основного состояния организма быстро включаются буферные системы компенсации (через 10-15 мин). Лёгочная компенсация обеспечивает стабилизацию соотношения НСО3-/Н2СО3, которая в норме соответствует 1:20, а при ацидозе уменьшается. Лёгочная компенсация достигается увеличением объёма вентиляции и, следовательно, ускорением выведения СО2 из организма. Однако основную роль в компенсации ацидоза играют почечные механизмы с участием амми-
ачного буфера (см. раздел 9). Одной из причин метаболического ацидоза может быть накопление молочной кислоты. В норме лактат в печени превращается обратно в глюкозу путём глюконе-огенеза либо окисляется. Кроме печени, другим потребителем лактата служат почки и сердечная мышца, где лактат может окисляться до СО2 и Н2О и использоваться как источник энергии, особенно при физической работе.
Уровень лактата в крови - результат равновесия между процессами его образования и утилизации. Кратковременный компенсированный лактоацидоз встречается довольно часто даже у здоровых людей при интенсивной мышечной работе. У нетренированных людей лактоацидоз при физической работе возникает как следствие относительного недостатка кислорода в мышцах и развивается достаточно быстро. Компенсация осуществляется путём гипервентиляции.
При некомпенсированном лактоацидозе содержание лактата в крови увеличивается до 5 ммоль/л (в норме до 2 ммоль/л). При этом рН крови может составлять 7,25 и менее (в норме 7,36-7,44).
Повышение содержания лактата в крови может быть следствием нарушения метаболизма пирувата (рис. 7-51).
Так, при гипоксии, возникающей вследствие нарушения снабжения тканей кислородом или
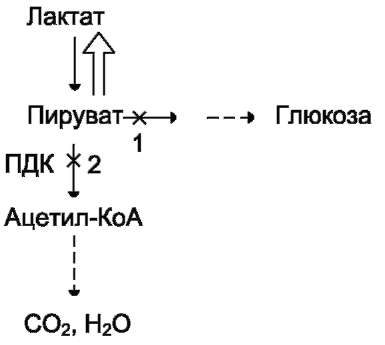
Рис. 7-51. Нарушения метаболизма пирувата при лактоацидозе. 1 - нарушение использования пирувата в глюконеогенезе; 2 - нарушение окисления пирувата.
кровью, уменьшается активность пируватдегид-рогеназного комплекса и снижается окислительное декарбоксилирование пирувата. В этих условиях равновесие реакции пируватолактат сдвинуто в сторону образования лактата. Кроме того, при гипоксии уменьшается синтез АТФ, что следовательно, ведёт к снижению скорости глюконеогенеза - другого пути утилизации лактата. Повышение концентрации лактата и снижение внутриклеточного рН отрицательно влияют на активность всех ферментов, в том числе и пируваткарбоксилазы, катализирующей начальную реакцию глюконеогенеза.
Возникновению лактоацидоза также способствуют нарушения глюконеогенеза при печёночной недостаточности различного происхождения. Кроме того, лактоацидозом может сопровождаться гиповитаминоз В1, так как производное этого витамина (тиаминдифосфат) выполняет кофер-ментную функцию в составе ПДК при окислительном декарбоксилировании пирувата (см. раздел 6). Дефицит тиамина может возникать, например, у алкоголиков с нарушенным режимом питания.
Итак, причинами накопления молочной кислоты и развития лактоацидоза могут быть:
• активация анаэробного гликолиза вследствие тканевой гипоксии различного происхождения;
• поражения печени (токсические дистрофии, цирроз и др.);
• нарушение использования лактата вследствие наследственных дефектов ферментов глюконеогенеза;
• нарушение работы ПДК вследствие дефектов ферментов или гиповитаминозов;
• применение ряда лекарственных препаратов, например бигуанидов (блокаторы глюконеогенеза, используемые при лечении сахарного диабета).
В. СИНТЕЗ ГЛЮКОЗЫ ИЗ АМИНОКИСЛОТ
В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма. Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты цитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных. Например, оксалоацетат, образующийся из аспарагиновой кислоты, является промежуточным продуктом как цитратного цикла, так и глюконеогенеза.
Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюконеогенез. Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах → пируват в мышцах → аланин в мышцах → аланин в печени → глюкоза в печени → глюкоза в мышцах (рис. 7-52). Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз.
Г. СИНТЕЗ ГЛЮКОЗЫ ИЗ ГЛИЦЕРОЛА
Глицерол образуется при гидролизе триа-цилглицеролов, главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицеролкиназа, например печень, почки. Этот АТФ-зависимый фермент катализирует превращение глицерола в α-глицерофосфат (глицерол-3-фосфат). При включении глицерол-3-фосфата в глюконе-
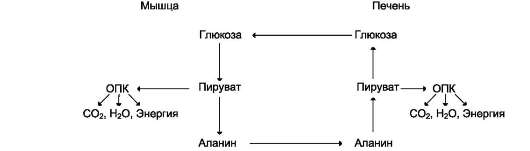
Рис. 7-52. Глюкозо-аланиновый цикл.
огенез происходит его дегидрирование NAD-зависимой дегидрогеназой с образованием дигидроксиацетонфосфата (рис. 7-53), который далее превращается в глюкозу.
Рис. 7-53. Превращение глицерола в дигидрокси-ацетонфосфат.
x. регуляция гликолиза и глюконеогенеза в печени
По сравнению с другими органами печень отличается наиболее сложным обменом глюкозы. Кроме пары противоположных процессов (синтеза и распада гликогена), в печени могут происходить ещё два противоположно направленных процесса - гликолиз и глюконеогенез. В большинстве других органов происходит только гликолиз. Переключение печени с гликолиза на глюконеогенез и обратно происходит с участием инсулина и глюкагона и осуществляется с помощью:
• аллостерической регуляции активности ферментов;
• ковалентной модификации ферментов путём фосфорилирования/дефосфорилирования;
• индукции/репрессии синтеза ключевых ферментов.
Регуляторные воздействия направлены на ферменты, катализирующие необратимые стадии гликолиза и глюконеогенеза, сочетание которых называют «субстратными», или «холостыми» циклами.
А. РЕГУЛЯЦИЯ СКОРОСТИ РЕАКЦИЙ ГЛИКОЛИЗА И ГЛЮКОНЕОГЕНЕЗА, СОСТАВЛЯЮЩИХ СУБСТРАТНЫЕ ЦИКЛЫ
«Субстратные» циклы - парные комбинации процессов синтеза и распада метаболитов. Как уже упоминалось, сочетание процессов синтеза и распада гликогена или необратимых реакций гликолиза и соответствующих им необратимых реакций глюконеогенеза может составить подобный цикл. Название «субстратный цикл» означает объединение реакций синтеза и распада
субстрата. Название «холостой» отражает результат работы подобного цикла, заключающийся в бесполезном расходовании АТФ. Хотя существование «холостых» циклов нелогично, тем не менее они могут функционировать. Более того, эти циклы могут быть мишенью регуляторных воздействий, так как составляющие их реакции катализируют разные ферменты. Реципрокное изменение активности этих ферментов предотвращает одновременное протекание противоположных процессов.
Изменение в печени гликолитического направления на глюконеогенез и обратно при смене абсорбтивного состояния на постабсорб-тивное или при голодании происходит главным образом в результате регуляции активности ферментов, катализирующих реакции субстратных циклов. Эти циклы обозначены цифрами I, II, III на рис. 7-54, представляющем общую картину регуляции гликолиза и глюконеогенеза в печени.
Направление реакции первого субстратного цикла
регулируется главным образом концентрацией глюкозы. При пищеварении концентрация глюкозы в крови повышается (до 8-10 ммоль/л). Активность глюкокиназы в этих условиях максимальна. Вследствие этого ускоряется гликоли-тическая реакция образования глюкозо-6-фос-фата. Кроме того, инсулин индуцирует синтез глюкокиназы и ускоряет тем самым фосфори-лирование глюкозы. Поскольку глюкокиназа печени не ингибируется глюкозо-6-фосфатом (в отличие от гексокиназы мышц), то основная часть глюкозо-6-фосфата в абсорбтивном периоде направляется на синтез гликогена и по гликолитическому пути.
Направление реакций второго субстратного цикла
зависит от активности фосфофруктокиназы и фосфатазы фруктозо-1,6-бисфосфата. Активность этих ферментов зависит от концентрации фруктозо-2,6-бисфосфата. Фруктозо-2,6-бисфос-фат - метаболит, образующийся в незначительных количествах из фруктозо-6-фосфата и выполняющий только регуляторные функции. Образование фруктозо-2,6-бисфосфата путём фосфорилирования фруктозо-6-фосфата катализирует бифункциональный фермент (БИФ), который катализирует также и обратную реакцию (рис. 7-55, А). Однако превращение фруктозо-2,6-бисфосфата в фруктозо-6-фосфат не является обратимым процессом. Образова-
ние фруктозо-2,6-бисфосфата требует затрат АТФ, а при образовании фруктозо-6-фосфата из фруктозо-2,6-бисфосфата гидролитически отщепляется неорганический фосфат.
В реакции фосфорилирования фруктозо-6-фосфата фермент проявляет киназную активность, а при дефосфорилировании образованного фруктозо-2,6-бисфосфата - фосфатазную. Это обстоятельство и определило название фермента «бифункциональный».
Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для абсорбтивного периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфос-фата увеличивается (рис. 7-55, Б).
При низком инсулин-глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результат - снижение количества фруктозо-2,6-бис-фосфата.
Киназную и фосфатазную реакции катализируют разные активные центры БИФ, но в каждом из двух состояний фермента (фосфорилирован-ном и дефосфорилированном) один из активных центров ингибирован. Регуляторное влияние фруктозо-2,6-бисфосфата заключается в том, что он аллостерически активирует фосфофруктоки-назу (фермент гликолиза). При этом фруктозо-2,6-бисфосфат снижает ингибирующее действие АТФ на этот фермент в абсорбтивном периоде и повышает его сродство к фруктозо-6-фосфату. В то же время фруктозо-2,6-бисфосфат инги-бирует фруктозо-1,6-бисфосфатазу (фермент глюконеогенеза). Итак, в абсорбтивном периоде уровень фруктозо-2,6-бисфосфата повышается, что приводит к активации фосфофруктокиназы и ускорению гликолиза.
Результатом уменьшения количества фрукто-зо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктоки-назы, замедление гликолиза и переключение гликолиза на глюконеогенез. Регуляторное влияние фруктозо-2,6-бисфосфата представлено на рис. 7-56.
В регуляции третьего субстратного цикла основная роль принадлежит пируваткиназе, фосфорили-рованная форма которой неактивна, а дефосфо-рилированная - активна (рис. 7-57).
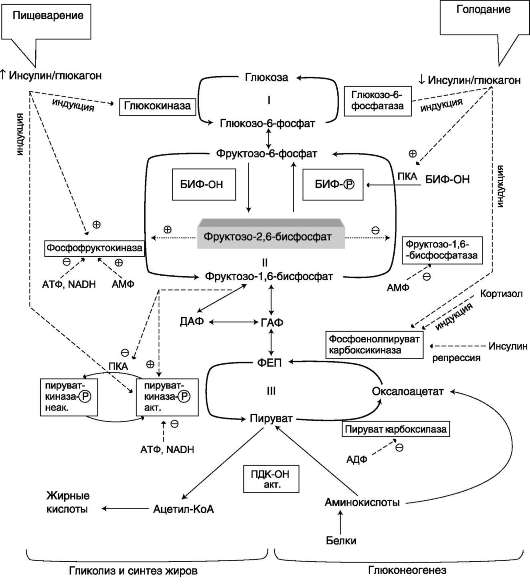
Рис. 7-54. Регуляция метаболизма глюкозы в печени. БИФ - бифункциональный фермент (фруктозо-2,6-бисфосфатаза/фосфофруктокиназа-2); БИФ-ОН - дефосфорилированный фермент; БИФ-Р - фосфори-лированный фермент, ПДК-ОН - дефосфорилированный пируватдегидрогеназный комплекс; ПК-ОН - де-фосфорилированная пируваткиназа; ГАФ - глицеральдегидфосфат; ДАФ - дигидроксиацетонфосфат, ФЕП - фосфоенолпируват. I-III - субстратные циклы: в рамках - регуляторные ферменты глиполиза и глюконеогенеза.
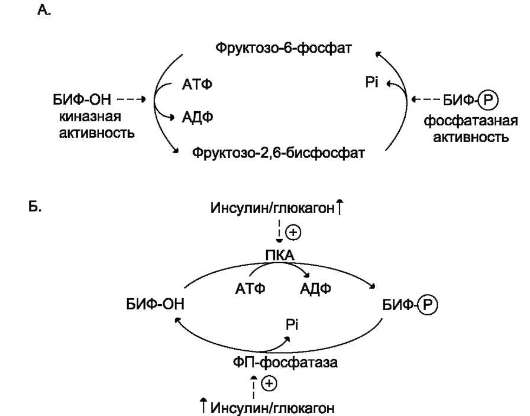
Рис. 7-55. Реакции, катализируемые бифункциональным ферментом (БИФ) в печени (А). Регуляция активности БИФ (Б).
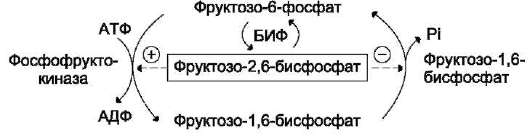
Рис. 7-56. Регуляция реакций II субстратного цикла фрук-тозо-2,6-бисфосфатом.
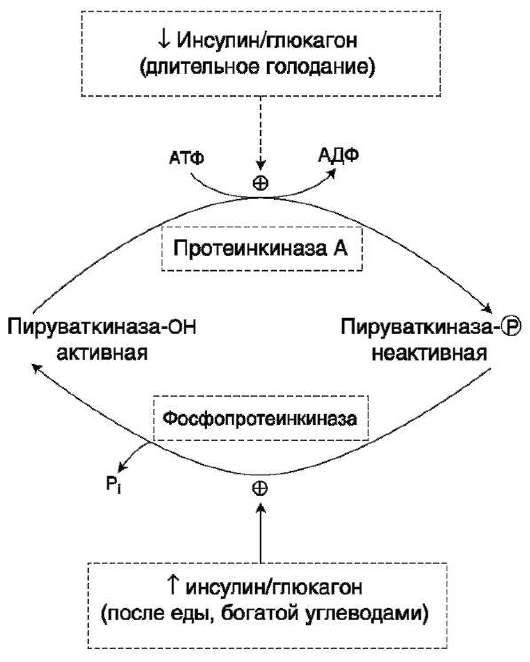
Рис. 7-57. Регуляция пируваткиназы в печени.
В период пищеварения инсулин активирует фосфопротеинфосфатазу, которая дефосфори-лирует пируваткиназу, переводя её в активное состояние. Кроме того, инсулин в печени влияет на количество ферментов, индуцируя синтез пируваткиназы и репрессируя синтез фосфо-енолпируваткарбоксикиназы. Следовательно, гликолитическая реакция фосфоенолпируват -пируват ускоряется при пищеварении. Эта же реакция замедляется в постабсорбтивном состоянии под влиянием глюкагона, который опосредованно через цАМФ-зависимую про-теинкиназу фосфорилирует и инактивирует пируваткиназу.
При длительном голодании глюкагон ускоряет глюконеогенез. Это достигается не только путём фосфорилирования пируваткиназы и снижением скорости гликолиза, но и путём индукции синтеза ферментов глюконеогенеза: фосфоенолпируваткарбоксикиназы, фрукто-зо-1,6-бисфосфатазы и глюкозо-6-фосфатазы. Известно, что глюкагон, фосфорилируя опосредованно транскрипционные факторы, влияет на их активность и таким образом индуцирует синтез этих ферментов глюконеогенеза. Кроме того, синтез фосфоенолпируваткарбоксикина-зы при длительном голодании индуцируется кортизолом, однако это происходит в результате включения другого механизма действия, характерного для стероидных гормонов (см. разделы 5, 11).
Координация скорости реакции II и III субстратных циклов достигается с помощью фруктозо-1,6-бис-фосфата - продукта II субстратного цикла (гли-колитическое направление), который является аллостерическим активатором пируваткиназы. В период пищеварения вследствие ускорения начальных стадий гликолиза концентрация фрук-тозо-1,6-бисфосфата повышается, что приводит к дополнительной активации пируваткиназы.
Общая картина регуляции процессов, составляющих метаболизм глюкозы в печени, представлена на рис. 7-54.
Необходимо отметить, что противоположные реакции каждого из субстратных циклов могут протекать одновременно. Соответственно, гликолиз и глюконеогенез в печени в какой-то мере тоже могут происходить одновременно, хотя их относительные скорости изменяются. Так, при пищеварении преобладает гликолитическое направление, а в постабсорбтивном состоянии - на-
правление глюконеогенеза. Например, реакция глюконеогенеза пируват - оксалоацетат может протекать при любых состояниях организма. Это объясняется необходимостью поддерживать концентрацию оксалоацетата на определённом уровне, потому что оксалоацетат используется не только в глюконеогенезе, но и в других процессах, таких как цитратный цикл, трансмембранный перенос веществ, синтез аминокислот.
б. значение гликолиза в печени для синтеза жиров
Основным значением ускорения гликолиза в печени в период пищеварения является образование дигидроксиацетонфосфата и ацетил-КоА - исходных веществ для синтеза жира. Образование ацетил-КоА из пирувата в ходе реакции, катализируемой ПДК, регулируется разными способами и подробно описывалось в разделе 6.
В абсорбтивном периоде ПДК находится в дефосфорилированной (активной) форме, следовательно, декарбоксилирование пирувата ускоряется. Образуемый ацетил-КоА используется в основном двумя путями: для синтеза жирных кислот и в цитратном цикле. В период пищеварения ускоряются образование ацетил-КоА и его использование для синтеза жирных кислот. Необходимый для синтеза жира α-глицерофосфат образуется в реакции восстановления из дигидроксиацетонфосфата (рис. 7-58). Подробно этот процесс рассматривается в разделе 8.
в. аллостерическая регуляция аэробного распада глюкозы и глюконеогенеза в печени
энергетическим статусом клетки
Аллостерическая регуляция скорости гликолиза, зависимая от изменения соотношения АТФ/АДФ, направлена на изменение скорости использования глюкозы непосредственно клетками печени. Глюкоза в клетках печени используется не только для синтеза гликогена и жиров, но также и как источник энергии для синтеза АТФ. Основными потребителями АТФ в гепа-тоцитах являются процессы трансмембранного переноса веществ, синтез белков, гликогена, жиров, глюконеогенез. От скорости утилизации
Рис. 7-58. Синтез жира из углеводов. 1 - окисление глюкозы до пирувата и окислительное декарбоксили-рование пирувата приводят к образованию ацетил-КоА; 2 - ацетил-КоА является строительным блоком для синтеза жирных кислот; 3 - жирные кислоты и a-глицеролфосфат, образующийся в реакции восстановления дигидроксиацетонфосфата, участвуют в синтезе триацилглицеролов.
АТФ в этих процессах зависит скорость его синтеза. АТФ, АДФ и АМФ, а также NAD+ и NADH служат аллостерическими эффекторами некоторых гликолитических ферментов и ферментов глюконеогенеза. В частности, АМФ активирует фосфофруктокиназу и ингибирует фруктозо-1,6-бисфосфатазу. АТФ и NADH ингибируют пируваткиназу, а АДФ ингибирует пируваткарбоксилазу.
Следовательно, при усилении расходования АТФ и снижении его концентрации с одновременным увеличением концентрации АМФ, активируется гликолиз и образование АТФ, а глюконеогенез при этом замедляется. Кроме того, от соотношения АТФ/АДФ, АМФ и NAD/NADH зависит скорость реакций общего пути катаболизма (см. раздел 6).
xi. регуляция содержания глюкозы в крови
Результат регуляции метаболических путей превращения глюкозы - постоянство концентрации глюкозы в крови.
Концентрация глюкозы в артериальной крови в течение суток поддерживается на постоянном уровне 60-100 мг/дл (3,3-5,5 ммоль/л). После приёма углеводной пищи уровень глюкозы возрастает в течение примерно 1 ч до 150 мг/дл (~8 ммоль/л, алиментарная гипергликемия), а затем возвращается к нормальному уровню (примерно через 2 ч). На рисунке 7-59 представлен график изменений концентрации глюкозы в
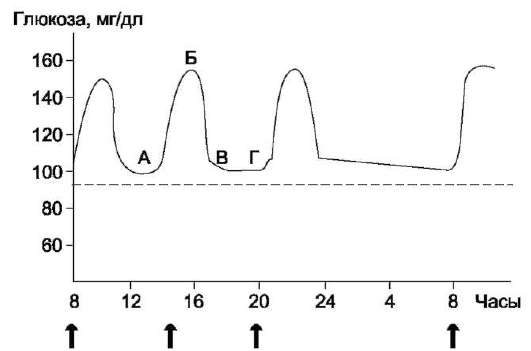
Рис. 7-59. Изменение концентрации глюкозы в крови в течение суток. А, Б - период пищеварения; В, Г - постабсорбтивный период. Стрелкой указано время приёма пищи, пунктиром показана нормальная концентрация глюкозы.
крови в течение суток при трёхразовом приёме пищи.
А. РЕГУЛЯЦИЯ СОДЕРЖАНИЯ ГЛЮКОЗЫ В КРОВИ В АБСОРБТИВНОМ И ПОСТАБСОРБТИВНОМ ПЕРИОДАХ
Для предотвращения чрезмерного повышения концентрации глюкозы в крови при пищеварении основное значение имеет потребление глюкозы печенью и мышцами, в меньшей мере - жировой тканью. Следует напомнить, что более половины всей глюкозы (60%), поступающей из кишечника в воротную вену, поглощается
печенью. Около 2/3 этого количества откладывается в печени в форме гликогена, остальная часть превращается в жиры и окисляется, обеспечивая синтез АТФ. Ускорение этих процессов инициируется повышением инсулин-глюкагонового индекса. Другая часть глюкозы, поступающей из кишечника, попадает в общий кровоток. Примерно 2/3 этого количества поглощается мышцами и жировой тканью. Это обусловлено увеличением проницаемости мембран мышечных и жировых клеток для глюкозы под влиянием высокой концентрации инсулина. Глюкоза в мышцах откладывается в форме гликогена, а в жировых клетках превращается в жиры. Остальная часть глюкозы общего кровотока поглощается другими клетками (инсулинонезависимыми).
При нормальном ритме питания и сбалансированном рационе концентрация глюкозы в крови и снабжение глюкозой всех органов поддерживается главным образом за счёт синтеза и распада гликогена. Лишь к концу ночного сна, т.е. к концу самого большого перерыва между приёмами пищи, может несколько увеличиться роль глюконеогенеза, значение которого будет возрастать, если завтрак не состоится и голодание продолжится (рис. 7-60).
Б. РЕГУЛЯЦИЯ СОДЕРЖАНИЯ ГЛЮКОЗЫ В КРОВИ ПРИ ДЛИТЕЛЬНОМ ГОЛОДАНИИ
При голодании в течение первых суток исчерпываются запасы гликогена в организме, и в дальнейшем источником глюкозы служит только глюконеогенез (из лактата, глицерина и аминокислот). Глюконеогенез при этом ускоряется, а гликолиз замедляется вследствие низкой концентрации инсулина и высокой концентрации глюкагона (механизм этого явления описан ранее). Но, кроме того, через 1-2 сут существенно проявляется действие и другого механизма регуляции - индукции и репрессии синтеза некоторых ферментов: снижается количество гликолитических ферментов и, наоборот, повышается количество ферментов глюконеогенеза. Изменение синтеза ферментов также связано с влиянием инсулина и глюка-гона (механизм действия рассматривается в разделе 11).
Начиная со второго дня голодания достигается максимальная скорость глюконеогенеза из аминокислот и глицерина. Скорость глю-
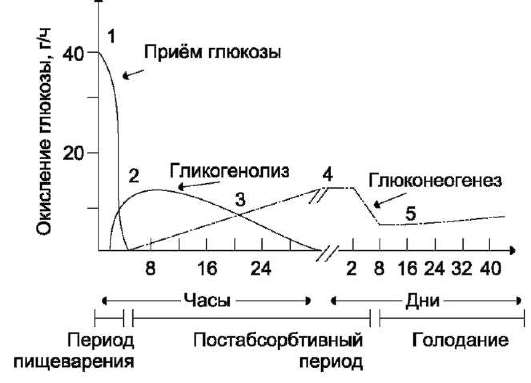
Рис. 7-60. Источники глюкозы в крови в период пищеварения и во время голодания. 1 - в период пищеварения углеводы пищи являются основным источником глюкозы в крови; 2 - в постабсорб-тивный период печень поставляет глюкозу в кровь за счёт процессов гликогенолиза и глюконеогенеза, причём в течение 8-12 ч уровень глюкозы в крови поддерживается в основном за счёт распада гликогена; 3 - глюконеогенез и гликоген в печени участвуют в равной степени в поддержании нормальной концентрации глюкозы; 4 - в течение суток гликоген печени практически полностью исчерпывается, и скорость глюконеогенеза увеличивается; 5 - при длительном голодании (1 нед и более) скорость глюконеогенеза уменьшается, но глюконеогенез остаётся единственным источником глюкозы в крови.
конеогенеза из лактата остаётся постоянной. В результате синтезируется около
Следует отметить, что при голодании глюкоза не используется мышечными и жировыми клетками, поскольку в отсутствие инсулина не проникает в них и таким образом сберегается для снабжения мозга и других глюкозозависи-мых клеток. Поскольку при других условиях мышцы - один из основных потребителей глюкозы, то прекращение потребления глюкозы мышцами при голодании имеет существенное значение для обеспечения глюкозой мозга. При достаточно продолжительном голодании (несколько дней и больше) мозг начинает использовать и другие источники энергии (см. раздел 8).
Вариантом голодания является несбалансированное питание, в частности такое, когда по калорийности рацион содержит мало углеводов - углеводное голодание. В этом случае также активируется глюконеогенез, и для синтеза глюкозы используются аминокислоты и глицерол, образующиеся из пищевых белков и жиров.
В. РЕГУЛЯЦИЯ СОДЕРЖАНИЯ ГЛЮКОЗЫ
В КРОВИ В ПЕРИОД ПОКОЯ И ВО ВРЕМЯ ФИЗИЧЕСКОЙ НАГРУЗКИ
Как
в период покоя, так и во время продолжительной физической работы
сначала источником глюкозы для мышц служит гликоген, запасённый в самих
мышцах, а затем глюкоза крови. Известно, что
Итак, изложенные сведения позволяют сделать вывод о том, что координация скоростей гликолиза, глюконеогенеза, синтеза и распада гликогена с участием гормонов обеспечивает:
• предотвращение чрезмерного повышения концентрации глюкозы в крови после приёма пищи;
• запасание гликогена и его использование в промежутках между приёмами пищи;
• снабжение глюкозой мышц, потребность которых в энергии быстро возрастает при мышечной работе;
• снабжение глюкозой клеток, которые при голодании в качестве источника энергии используют преимущественно глюкозу (нерв-ные клетки, эритроциты, мозговое вещество почек, семенники).
xii. пентозофосфатный путь превращения глюкозы
Пентозофосфатный путь, называемый также гексомонофосфатным шунтом, служит
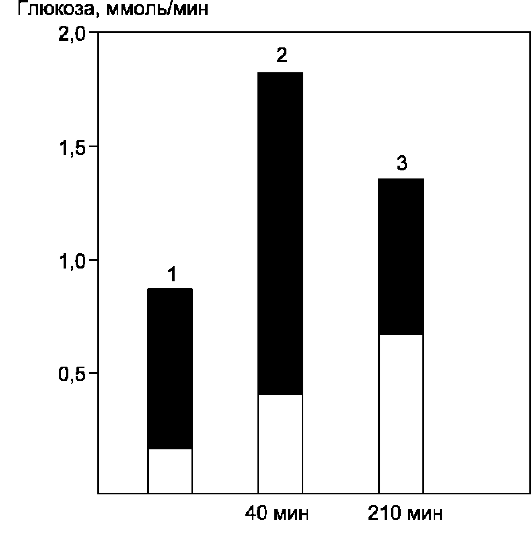
Рис. 7-61. Вклад гликогена печени и глюконеогенеза в поддержание уровня глюкозы крови в период покоя и во время продолжительных физических упражнений. Тёмная часть столбика - вклад гликогена печени в поддержание уровня глюкозы в крови; светлая - вклад глюконеогенеза. При увеличении продолжительности физической нагрузки с 40 мин (2) до 210 мин (3) распад гликогена и глюконеогенез практически в равной степени обеспечивают кровь глюкозой. 1 - состояние покоя (постабсорбтивный период); 2, 3 - физическая нагрузка.
альтернативным путём окисления глюкозо-6-фосфата. Пентозофосфатный путь состоит из 2 фаз (частей) - окислительной и неокислительной.
В окислительной фазе глюкозо-6-фосфат необратимо окисляется в пентозу - рибуло-зо-5-фосфат, и образуется восстановленный NADPH.
В неокислительной фазе рибулозо-5-фосфат обратимо превращается в рибозо-5-фосфат и метаболиты гликолиза.
Пентозофосфатный путь обеспечивает клетки рибозой для синтеза пуриновых и пи-римидиновых нуклеотидов и гидрированным коферментом NADPH, который используется в восстановительных процессах.
Ферменты пентозофосфатного пути, так же, как и ферменты гликолиза, локализованы в цитозоле.
Наиболее активно пентозофосфатный путь протекает в жировой ткани, печени, коре надпочечников, эритроцитах, молочной железе в период лактации, семенниках.
А. ОКИСЛИТЕЛЬНЫЙ ЭТАП
В окислительной части пентозофосфатного пути глюкозо-6-фосфат подвергается окислительному декарбоксилированию, в результате которого образуются пентозы. Этот этап включает 2 реакции дегидрирования.
Первая реакция дегидрирования - превращение глюкозо-6-фосфата в глюконолактон-6-фосфат - катализируется NADP+-зависимой глюкозо-6-фосфатдегидрогеназой и сопровождается окислением альдегидной группы у первого атома углерода и образованием одной молекулы восстановленного кофермента NADPH.
Далее глюконолактон-6-фосфат быстро превращается в 6-фосфоглюконат при участии фермента глюконолактонгидратазы.
Фермент 6-фосфоглюконатдегидрогеназа катализирует вторую реакцию дегидрирования окислительной части, в ходе которой происходит также и декарбоксилирование. При этом углеродная цепь укорачивается на один атом углерода, образуется рибулозо-5-фосфат и вторая молекула гидрированного NADPH (рис. 7-62).
Восстановленный NADPH ингибирует первый фермент окислительного этапа пентозофосфат-ного пути - глюкозо-6-фосфатдегидрогеназу. Превращение NADPH в окисленное состояние NADP+ приводит к ослаблению ингибирования фермента. При этом скорость соответствующей реакции возрастает, и образуется большее количество NADPH.
Суммарное уравнение окислительного этапа пенто-зофосфатного пути можно представить в виде:
Глюкозо-6-фосфат + 2 NADP+ + Н2О → Рибу-лозо-5-фосфат + 2 (NADPH + Н+) + СО2.
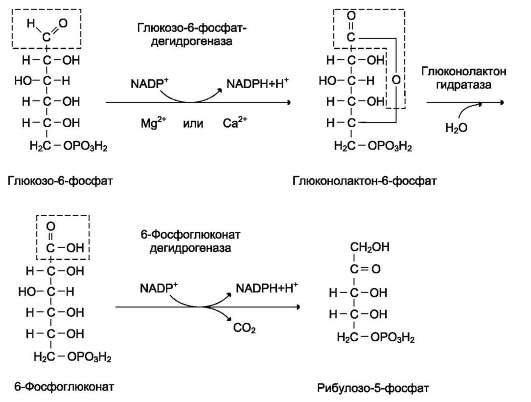
Рис. 7-62. Окислительный этап пентозофосфатного пути.
Реакции окислительного этапа служат основным источником NADPH в клетках. Гидрированные коферменты снабжают водородом биосинтетические процессы, окислительно-восстановительные реакции, включающие защиту клеток от активных форм кислорода. NADPH, как донор водорода участвует в анаболических процессах, например в синтезе холестерина. Это источник восстановительных эквивалентов для цитохрома Р450, катализирующего образование гидроксильных групп при синтезе стероидных гормонов, жёлчных кислот, при катаболизме лекарственных веществ и других чужеродных соединений (см. разделы 8, 11, 12). Высокая активность фермента глюкозо-6-фосфатдегидроге-назы обнаружена в фагоцитирующих лейкоцитах, где NADPH-оксидаза использует восстановленный NADPH для образования супероксидного иона из молекулярного кислорода. Супероксидньгй ион генерирует другие активные формы кислорода, под действием которых и повреждаются молекулы ДНК, белков, липидов бактериальных клеток. Синтез жирных кислот из углеводов в печени является основным путём утилизации NADPH и обеспечивает регенерацию окисленной формы NADP+. В печени глюкозо-6-фосфатдегидро-геназа, как и ключевые ферменты гликолиза и биосинтеза жирных кислот, индуцируется при увеличении соотношения инсулин/глюкагон после приёма богатой углеводами пищи.
Несмотря на то, что NADPH образуется также при окислении малата до пирувата и диоксида углерода (при участии NADP+-зависимой малатдегидрогеназы) и дегидрировании изо-цитрата (при участии NADP+-зависимой изо-цитратдегидрогеназы), в большинстве случаев потребности клеток в восстановительных эквивалентах удовлетворяются за счёт пентозофос-фатного пути.
Реакции окислительного пути протекают только в том случае, если восстановленный кофермент NADPH возвращается в исходное окисленное состояние NADP+ при участии NADPH-зависимых дегидрогеназ (т.е. при условии использования гидрированного NADPH в восстановительных процессах). Если потребности клетки в NADPH незначительны, рибо-зо-5-фосфат образуется в результате обратимых реакций неокислительного этапа пентозофос-фатного пути, используя в качестве исходных
веществ метаболиты гликолиза - глицеральде-гид-3-фосфат и фруктозо-6-фосфат.
б. неокислительный этап
Неокислительный этап пентозофосфатного пути включает серию обратимых реакций, в результате которых рибулозо-5-фосфат превращается в рибозо-5-фосфат и ксилулозо-5-фосфат, и далее за счёт переноса углеродных фрагментов в метаболиты гликолиза - фруктозо-6-фосфат и глицеральдегид-3-фосфат. В этих превращениях принимают участие ферменты: эпимераза, изомераза, транскетолаза и трансальдолаза. Транскетолаза в качестве кофермента использует тиаминдифосфат. Неокислительный этап пентозофосфатного пути не включает реакции дегидрирования и поэтому используется только для синтеза пентоз.
Рибулозо-5-фосфат служит субстратом для двух ферментов. Фермент рибулозо-5-фосфат-3-эпиме-раза изменяет стехиометрическое положение одной ОН-группы у третьего атома углерода, превращая рибулозо-5-фосфат в ксилулозо-5-фосфат. Другой фермент - рибулозо-5-фосфат-изомераза - катализирует превращение рибулозо-5-фосфата в рибозо-5-фосфат (рис. 7-63). Рибозо-5-фосфат, образующийся в неокислительной фазе, обеспечивает клетки рибозой, необходимой для синтеза нуклеотидов, которые служат предшественниками и структурными компонентами коферментов де-гидрогеназ и нуклеиновых кислот.
Ферменты транскетолаза и трансальдолаза катализируют перенос двух- и трёхуглеродных фрагментов, соответственно используя в качестве донора углеродных фрагментов кетозу, а альдо-зу - в качестве акцептора. Эти реакции протекают в 2 этапа: сначала происходит отщепление углеродного фрагмента от молекулы-донора, а затем - перенос этого фрагмента на молекулу, выполняющую роль акцептора. Транскетолаза в неокислительной фазе пентозофосфатного пути катализирует 2 реакции. В первой реакции (рис. 7-64) транскетолаза расщепляет связь С-С между кетогруппой и соседним атомом углерода в молекуле ксилулозо-5-фосфат, в результате чего кетосахар превращается в альдозу, глицеральде-гид-3-фосфат, содержащую на 2 атома углерода меньше. Образующийся после расщепления двух-углеродный фрагмент остаётся ковалентно связанным в каталитическом центре фермента
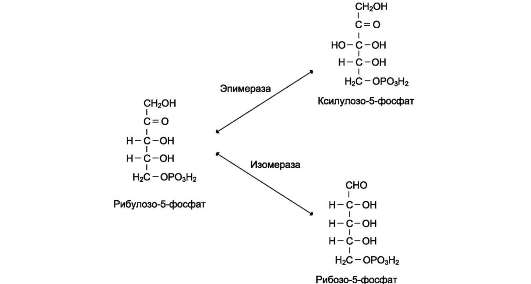
Рис. 7-63. Превращения рибулозо-5-фосфата.
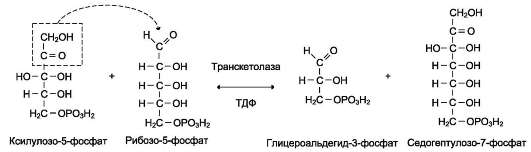
Рис. 7-64. Реакция переноса двухуглеродного фрагмента, катализируемая транскетолазой.
с коферментом тиаминдифосфатом. Далее фермент переносит двухуглеродный фрагмент на альдегидную группу альдосахара, образуя новую кетозу - седогептулозо-7-фосфат.
Трансальдолаза переносит трёхуглеродный фрагмент от седогептулозо-7-фосфата на глице-ральдегид-3-фосфат, образуя эритрозо-4-фосфат и фруктозо-6-фосфат (рис. 7-65).
Эта реакция подобна реакции альдольного расщепления гликолитического пути, за исключением того, что в данном случае трёхуглерод-ный фрагмент, содержащий кетогруппу, переносится на альдосахар глицеральдегид-3-фосфат,
а в гликолитическом пути кетофрагмент высвобождается в виде дигидроксиацетонфосфата.
В следующей реакции, катализируемой транс-кетолазой, происходит перенос двухуглеродного фрагмента от ксилулозо-5-фосфата на эритрозо-4-фосфат. Продуктами этой реакции являются фруктозо-6-фосфат и глицеральдегид-3-фосфат (рис. 7-66).
Так как все реакции неокислительного этапа обратимы, образование рибозо-5-фосфата может происходить не только в результате изомерного превращения продукта окислительной фазы пентозофосфатного пути рибулозо-
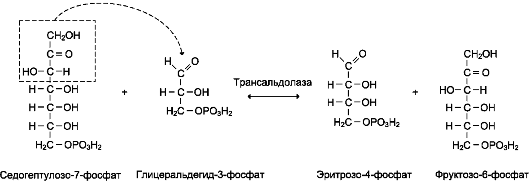
Рис. 7-65. Реакция, катализируемая трансальдолазой.
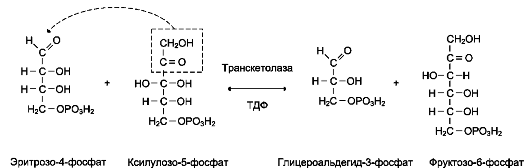
Рис. 7-66. Реакция, катализируемая транскетолазой.
5-фосфата в рибозо-5-фосфат под действием изомеразы, но также и из промежуточных продуктов гликолиза - фруктозо-6-фосфата и глицеральдегид-3-фосфата. Последовательность превращений, приводящих к образованию ри-бозо-5-фосфата из таких продуктов гликолити-ческого пути, можно представить в виде: 2 Фруктозо-6-фосфат + Глицеральдегид-3-фосфат
→ 2 Ксилулозо-5-фосфат + Рибозо-5-фосфат 2 Ксилулозо-5-фосфат → 2 Рибулозо-5-фосфат 2 Рибулозо-5-фосфат → 2 Рибозо-5-фосфат.
Суммарный результат метаболизма 3 молекул рибулозо-5-фосфата в неокислительной фазе пентозофосфатного пути - образование 2 молекул фруктозо-6-фосфата и 1 молекулы глице-ральдегид-3-фосфата. Далее фруктозо-6-фосфат и глицеральдегид-3-фосфат могут превратиться в глюкозу. С учётом стехиометрического коэф-
фициента, равного 2, для образования 5 молекул глюкозы (содержащих 30 атомов углерода) потребуются 4 молекулы фруктозо-6-фосфата и 2 молекулы глицеральдегид-3-фосфата (в сумме содержащие также 30 атомов углерода) или, соответственно, 6 молекул рибулозо-5-фосфата. Таким образом, неокислительный путь можно представить как процесс возвращения пентоз в фонд гексоз.
В. ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ
Окислительный этап образования пентоз и неокислительный этап (путь возвращения пен-тоз в гексозы) составляют вместе циклический процесс.
Такой процесс можно описать общим уравнением:
6 Глюкозо-6-фосфат + 12 NADP+ + 2 H2O →
5 Глюкозо-6-фосфат + 12 NADPH +12 H+ +
6 CO2.
Это означает, что из 6 молекул глюкозы образуются 6 молекул рибулозо-5-фосфат (пентозы) и 6 молекул СО2. Ферменты неокислительной фазы превращают 6 молекул рибулозо-5-фосфат в 5 молекул глюкозы (гексозы). При последовательном проведении этих реакций единственным полезным продуктом является NADPH, образующийся в окислительной фазе пенто-зофосфатного пути. Такой процесс называют пентозофосфатным циклом (рис. 7-67).
Протекание пентозофосфатного цикла позволяет клеткам продуцировать NADPH, необходимый для синтеза жиров, не накапливая пентозы.
Энергия, выделяющаяся при распаде глюкозы, трансформируется в энергию высокоэнергетического донора водорода - NADPH. Гидрированный NADPH служит источником водорода для восстановительных синтезов, а энергия NADPH преобразуется и сохраняется во вновь синтезированных веществах, например жирных кислотах, высвобождается при их катаболизме и используется клетками.
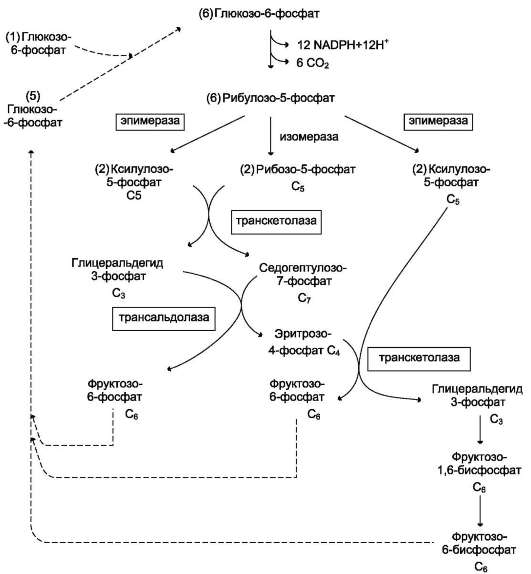
Рис. 7-67. Пентозофосфатный цикл в жировой ткани.
Г. ДЕФЕКТ ГЛЮКОЗО-6-ФОСФАТ ДЕГИДРОГЕ-НАЗЫ В ЭРИТРОЦИТАХ
Неферментативное окисление гемоглобина (Fe2+) в метгемоглобин (Fe3+) приводит к од-ноэлектронному восстановлению кислорода и появлению реакционно-способного анион-радикала - супероксида О2-, который служит предшественником других активных форм кислорода: пероксида водорода Н2О2 и гидроксиль-ного радикала ОН-. Активные формы кислорода являются сильнейшими окислителями и поэтому способны вызывать серьёзные повреждения молекул ДНК, белков, ненасыщенных липидов.
В эритроцитах, как и в большинстве клеток, присутствует тиолсодержащий трипептид - глу-татион (γ-глутамил-цистенил-глицин). Восстановленная форма глутатиона (Г-SH) содержит SH-группу (рис. 7-68), которая может служить донором электронов в реакциях восстановления. Под действием фермента глутатионпероксидазы восстановленный глутатион превращает молекулу пероксида водорода в молекулу воды, а сам переходит в окисленное состояние (Г-SS-Г). Регенерацию восстановленного глутатиона обеспечивает глутатионредуктаза, используя в качестве донора водорода гидрированньгй NADPH. Для эритроцитов единственным источником получения NADPH служит пентозофосфатный путь, для других тканей существует альтернативный способ - при участии NADH-зависимой малат-дегидрогеназы (малик-фермент).
Взаимодействие восстановленного глутатиона с пероксидом водорода в эритроцитах предохраняет цистеиновые остатки в протомерах гемоглобина от окисления. При генетическом дефекте
глюкозо-6-фосфатдегидрогеназы концентрация восстановленного кофермента NADPH уменьшается, в результате чего резко снижается концентрация восстановленного глутатиона, а в клетке, соответственно, увеличивается количество активных форм кислорода. В этом случае окисление SH-групп молекул гемоглобина в эритроцитах приводит к образованию перекрёстных дисульфидных связей и агрегации про-томеров гемоглобина с формированием телец Хайнца (см. раздел 14). В присутствии телец Хайнца пластичность мембраны нарушается, и она теряет способность к деформации при прохождении эритроцитов через капилляры. Это вызывает нарушение целостности мембраны, что приводит к гемолизу эритроцитов. Некоторые лекарственные вещества, например антималярийный препарат примахин, сульфаниламиды, также снижают способность эритроцитов бороться с активными формами кислорода.
xiii. метаболизм фруктозы и галактозы
Метаболизм фруктозы и галактозы включает пути использования их для синтеза других веществ (гетерополисахаридов, лактозы и др.) и участие в энергообеспечении организма. В последнем случае фруктоза и галактоза превращаются в печени либо в глюкозу, либо в промежуточные продукты её метаболизма. Таким образом, в результате фруктоза и галактоза наряду с глюкозой могут быть окислены до СО2 и Н2О или использованы на синтез гликогена и триацилглицеролов.
Рис. 7-68. Восстановление глутатиона под действием глутатионредуктазы. А - строение глутатиона; Б - восстановление глутатиона.
Причиной нарушения метаболизма фруктозы и галактозы может быть дефект ферментов, катализирующих промежуточные реакции их обмена. Эти нарушения встречаются относительно редко, но могут представлять достаточно серьёзную опасность, так как накапливаемые промежуточные метаболиты фруктозы и галактозы обладают токсичностью.
А. МЕТАБОЛИЗМ ФРУКТОЗЫ
Значительное количество фруктозы, образующееся при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Дру-гая часть фруктозы всасывается с помощью белка-переносчика, т.е. путём облегчённой диффузии.
Метаболизм фруктозы (рис. 7-69) начинается с реакции фосфорилирования (реакция 1), катализируемой фруктокиназой с образованием фруктозо-1-фосфата. Фермент обнаружен в печени, а также в почках и кишечнике. Этот фермент обладает абсолютной специфичностью, поэтому, в отличие от глюкокиназы, инсулин не влияет на его активность. Последнее обстоятельство объясняет, почему уровень выведения фруктозы в моче у больных сахарным диабетом и здоровых не отличается. Фруктозо-1-фосфат не может превращаться во фруктозо-6-фосфат из-за отсутствия соответствующего фермента. Вместо этого фруктозо-1-фосфат далее расщепляется фруктозо-1-фосфатальдолазой (альдолаза В) на глицеральдегид и дигидрок-сиацетон-3-фосфат (реакция 2). Последний
Рис. 7-69. Метаболизм фруктозы. а - превращение фруктозы в дигидроксиацетон-3-фосфат и глицераль-дегид-3-фосфат; б - путь включения фруктозы в гликолиз и глюконеогенез; в - путь включения фруктозы в синтез гликогена.
является промежуточным продуктом гликолиза и образуется в ходе реакции, катализируемой фруктозо-1,6-бисфосфосфатальдолазой (аль-долаза А). Глицеральдегид может включаться в гликолиз после его фосфорилирования с участием АТФ (реакция 3). Две молекулы триозофос-фатов либо распадаются по гликолитическому пути, либо конденсируются с образованием фруктозо-1,6-бисфосфата и далее участвуют в глюконеогенезе (реакции 8, 7, 5, 9). Фруктоза в печени включается главным образом во второй путь. Часть дигидроксиацетон-3-фосфата может восстанавливаться до глицерол-3-фосфата и участвовать в синтезе триацилглицеролов.
Следует отметить, что включение фруктозы в метаболизм через фруктозо-1-фосфат минует стадию, катализируемую фосфофруктокиназой (реакция 6), которая является пунктом мета-болитического контроля скорости катаболизма глюкозы. Этим обстоятельством можно объяснить, почему увеличение количества фруктозы ускоряет в печени процессы, ведущие к синтезу жирных кислот, а также их этерификацию с образованием триацилглицеролов.
Б. НАРУШЕНИЯ МЕТАБОЛИЗМА ФРУКТОЗЫ
Нарушения метаболизма фруктозы, причиной которых является дефект ферментов, отражены в табл. 7-5.
Недостаточность фруктокиназы клинически не проявляется. Фруктоза накапливается в крови и выделяется с мочой, где её можно обнаружить лабораторными методами. Очень важно не перепутать эту безвредную аномалию с сахарным диабетом. Данное заболевание известно как
доброкачественная эссенциальная фруктозурия и встречается с частотой 1:130 000.
Наследственная непереносимость фруктозы, возникающая при генетически обусловленном дефекте фруктозо-1-фосфатальдолазы, не проявляется, пока ребёнок питается грудным молоком, т.е. пока пища не содержит фруктозы. Симптомы возникают, когда в рацион добавляют фрукты, соки, сахарозу. Рвота, боли в животе, диарея, гипогликемия и даже кома и судороги возникают через 30 мин после приёма пищи, содержащей фруктозу. У маленьких детей и подростков, продолжающих принимать фруктозу, развиваются хронические нарушения функций печени и почек. Непереносимость фруктозы - достаточно частая аутосомно-рецессивная форма патологии.
Дефект альдолазы фруктозо-1-фосфата сопровождается накоплением фруктозо-1-фосфа-та, который ингибирует активность фосфоглю-комутазы, превращающей глюкозо-1-фосфат в глюкозо-6-фосфат и обеспечивающей включение продукта гликогенфосфорилазной реакции в метаболизм. Поэтому происходит торможение распада гликогена на стадии образования глю-козо-1-фосфата, в результате чего развивается гипогликемия. Как следствие, ускоряется мобилизация липидов и окисление жирных кислот. Следствием ускорения окисления жирных кислот и синтеза кетоновых тел, замещающих энергетическую функцию глюкозы, может быть метаболический ацидоз (см. раздел 8), так как кетоновые тела являются кислотами и при высоких концентрациях снижают рН крови.
Результатом торможения гликогенолиза и гликолиза является снижение синтеза АТФ.
Таблица 7-5. Нарушения метаболизма фруктозы

Кроме того, накопление фосфорилированной фруктозы ведёт к нарушению обмена неорганического фосфата и гипофосфатемии.
Для пополнения внутриклеточного фосфата ускоряется распад адениловых нуклеотидов. Продукты распада этих нуклеотидов включаются в катаболизм, проходя стадии образования гипоксантина, ксантина и, наконец, мочевой кислоты. Повышение количества мочевой кислоты и снижение экскреции уратов в условиях метаболического ацидоза проявляются в виде гипер-урикемии. Следствием гиперурикемии может быть подагра даже в молодом возрасте (см. раздел 10).
В. МЕТАБОЛИЗМ ГАЛАКТОЗЫ
Галактоза образуется в кишечнике в результате гидролиза лактозы. Чтобы превратить галактозу в глюкозу, необходимо изменить оптическую конфигурацию Н- и ОН-групп С4 атома в галактозе, т.е. провести реакцию эпимеризации. Эта реак-
ция в клетке возможна только с УДФ-производ-ным галактозы. УДФ-галактоза образуется из УДФ-глюкозы (метаболит в синтезе гликогена) в ходе реакции, катализируемой уридилфосфат-4-эпимеразой (рис. 7-70, 7-71).
Однако включению галактозы в описанную реакцию эпимеризации предшествует её фосфорилирование с образованием галакто-зо-1-фосфата (реакция 1 на рис. 7-70). Далее галактозо-1-фосфат замещает остаток глюкозы в УДФ-глюкозе с образованием УДФ-галактозы (реакция 2), т.е. прямая реакция фосфорилированной галактозы с УТФ не происходит.
Реакцию 2 можно рассматривать как перенос уридильного остатка с УДФ-глюкозы на галактозу, поэтому фермент назван галактозо-1-фосфатуридилтрансферазой (ГАЛТ).
Затем галактоза в составе нуклеотида включается в реакцию эпимеризации, в которой участвует эпимераза - NAD-зависимый фермент, катализирующий окисление и восстановление галактозы по С4 углеродному атому (реакция 3).
Рис. 7-70. Обмен галактозы.
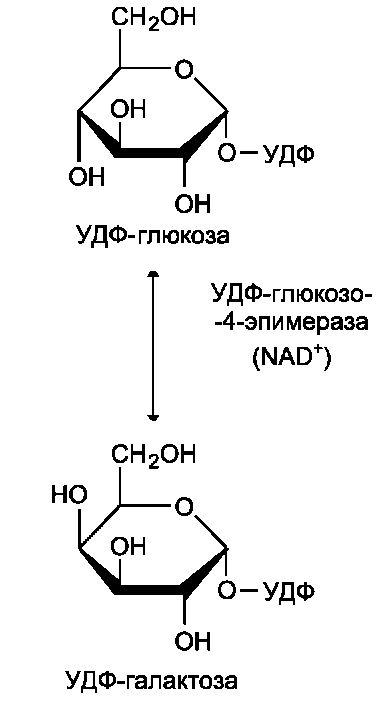
Рис. 7-71. Реакция эпимеризации УДФ-глюкозы в УДФ-галактозу.
Эпимераза может работать и в другом направлении, преобразуя УДФ-глюкозу в УДФ-галактозу. Эта обратная эпимеризация важна для синтеза галактозильных остатков в гликолипидах и гликопротеинах. Кроме того, галактоза необходима для синтеза лактозы в грудных железах. В период лактации галактоза не является неза-
менимым компонентом пищи, так как может образовываться из глюкозы.
Глюкозо-1-фосфат, образованный в реакции 2, может включаться в разные метаболические пути: 1) синтез гликогена после реакции с УДФ и образования УДФ-глюкозы; 2) превращение в печени в свободную глюкозу и поддержание её концентрации в крови; 3) катаболизм, сопряжённый с синтезом АТФ, и т.д. (см. рис. 7-70). Г. Нарушения метаболизма галактозы Обмен галактозы особенно интересен в связи с наследственным заболеванием - галактозе-мией.
Галактоземия возникает при нарушении обмена галактозы, обусловленном наследственным дефектом любого из трёх ферментов, включающих галактозу в метаболизм глюкозы (табл. 7-6).
Галактоземия, вызванная недостаточностью галактозо-1-фосфатуридилтрансферазы (ГАЛТ), наиболее хорошо изучена. Это заболевание проявляется очень рано, и особенно опасно для детей, так как основным источником углеводов для них служит материнское молоко, содержащее лактозу. Ранние симптомы дефекта ГАЛТ: рвота, диарея, дегидратация, уменьшение массы тела, желтуха. Они появляются вскоре после рождения, как только ребёнок начинает получать молоко. В крови, моче и тканях повышается концентрация галактозы и галак-тозо-1-фосфата. В тканях глаза (в хрусталике)
Таблица 7-6. Нарушения обмена галактозы
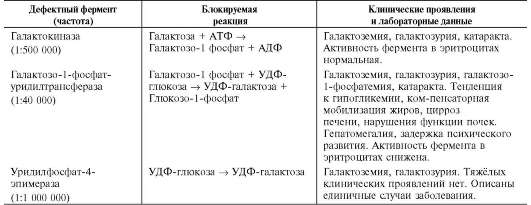
галактоза восстанавливается альдоредуктазой с образованием галактитола (дульцита). В этой реакции в качестве донора водорода используется NADPH. Восстановление галактозы происходит и в ходе нормального метаболизма, но протекает с небольшой скоростью. При галактоземии галактитол накапливается в стекловидном теле и связывает большое количество воды. Вследствие этого нарушается баланс электролитов, а чрезмерная гидратация хрусталика приводит к развитию катаракты, которая наблюдается уже через несколько дней после рождения.
Тяжёлые последствия дефекта ГАЛТ наблюдают в печени. Это связано с накоплением галактозо-1-фосфата и его токсическим действием на гепатоциты. В результате возникают нарушения функции печени: гепатомегалия, жировая дистрофия. В почках таких больных также повышена концентрация галактитола и галактозо-1-фосфата, что влияет на их функции. Отмечают нарушения в клетках полушарий головного мозга и мозжечка, в тяжёлых случаях - отёк мозга, задержку умственного развития, возможен летальный исход.
Для галактоземии, вызванной дефектом га-лактокиназы, тоже характерна катаракта, но при этом заболевании, в отличие от дефекта
ГАЛТ, не отмечают нарушений функций печени, почек, мозга. Наиболее тяжёлые последствия снижения активности ГАЛТ связывают с влиянием галактозо-1-фосфата на активность других ферментов, участвующих в углеводном обмене (фосфоглюкомутазы, глюкозо-6-фосфатдегид-рогеназы).
Известно несколько форм галактоземии, причиной которой является недостаточность
ГАЛТ (табл. 7-7).
Некоторые дефекты в строении ГАЛТ приводят лишь к частичной потере активности фермента. Поскольку в норме ГАЛТ присутствует в организме в избытке, то снижение его активности до 50%, а иногда и ниже может клинически не проявляться.
При диагностике галактоземии исследуют мочу на содержание галактозы, собранную после нескольких кормлений молоком. При обнаружении у ребёнка катаракты его обследуют на недостаточность галактокиназы и ГАЛТ. Наличие галактозы в моче при отсутствии нарушений функции печени указывает на дефект галактокиназы. При обследовании проведение теста с нагрузкой галактозой не рекомендуется, так как этот тест опасен для больных. Лечение заключается в удалении галактозы из рациона.
Таблица 7-7. Некоторые варианты генетического дефекта ГАЛТ
