Биохимия: учебник для вузов/ под ред. Е.С.Северина - 5-е изд., - 2009. - 768 с.
|
|
|
|
РАЗДЕЛ 10 ОБМЕН НУКЛЕОТИДОВ
Рибонуклеозид- и дезоксирибонуклеозид-фосфаты - существеннейшие компоненты клеток.
• Нуклеозидтрифосфаты (НТФ) используются в качестве субстратов синтеза ДНК и РНК, без которых невозможны образование белков и клеточная пролиферация.
• Природа выбрала цикл АДФ-АТФ в качестве универсального механизма трансформации энергии окисления в энергию биосинтетических процессов. В некоторых биологических процессах и другие НТФ используются в качестве источника энергии.
• Производные нуклеотидов служат донорами активных субстратов в синтезе гомо- и гетерополисахаридов, липидов и белков. Например: УДФ-глюкоза, УДФ-галактоза, ГДФ-манноза, УДФ-N-ацетилглюкозамин или ЦМФ-ацетилнейраминовая кислота принимают участие в синтезе гликогена и гликозаминогликанов; ЦДФ-холин - в синтезе фосфолипидов.
• УДФ-глюкуроновая кислота, ФАФС, S-аде-нозилметионин - наиболее частые участники универсальной системы детоксикации, обеспечивающей последующее выведение ксенобиотиков (чужеродных веществ) и некоторых собственных метаболитов из организма.
• АМФ входит в состав коферментов дегидро-геназ (NAD+, NADP+, FAD) и ацилирования (KoA).
• С помощью циклических форм нуклеотидов (цAMФ, цГМФ) осуществляется передача в клетку сигналов гормонов, факторов роста, нейромедиаторов и некоторых других регу-ляторных молекул.
Практически все клетки организма способны к синтезу нуклеотидов (исключение составляют некоторые клетки крови). Другим источником этих молекул могут быть нуклеиновые кислоты собственных тканей и пищи, однако эти источники имеют лишь второстепенное, вспомогательное значение.
Со строением нуклеотидов и номенклатурой мы знакомились в разделе 4, в данном разделе предстоит рассмотреть метаболизм этих молекул в организме.
I. ПЕРЕВАРИВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ ПИЩИ В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ
Пищевые нуклеопротеины, попадая в организм человека, в желудке отщепляют белковый компонент и денатурируют под действием HCl желудочного сока (рис. 10-1). Далее полинук-леотидная часть этих молекул гидролизуется в кишечнике до мононуклеотидов.
В расщеплении нуклеиновых кислот принимают участие ДНК-азы и РНК-азы панкреатического сока, которые, будучи эндонуклеазами, гидролизуют макромолекулы до олигонуклеоти-дов. Последние под действием фосфодиэстераз панкреатической железы расщепляются до смеси 3'- и 5'-мононуклеотидов. Нуклеотидазы и неспецифические фосфатазы гидролитически отщепляют фосфатный остаток нуклеотидов и превращают их в нуклеозиды, которые либо всасываются клетками тонкого кишечника, либо расщепляются нуклеозидфосфорилазами кишечника с образованием рибозоили дезок-сирибозо-1-фосфата, пуриновых и пиримиди-новых оснований.
Пищевые пурины и пиримидины не являются незаменимыми пищевыми факторами и очень мало используются для синтеза нуклеиновых кислот тканей. В энтероцитах обнаружена высокая активность ксантиноксидазы - фермента, который большую часть пуринов, поступающих в клетки, превращает в мочевую кислоту, удаляющуюся с мочой. Пиримидиновые основания, не успевшие поступить в энтероциты, под действием микрофлоры кишечника расщепляются до NH3, CO2, β-аланина и β-аминоизобутирата.
В различных клетках организма синтезируется до 90% пуриновых и пиримидиновых нуклеотидов из простых предшественников de
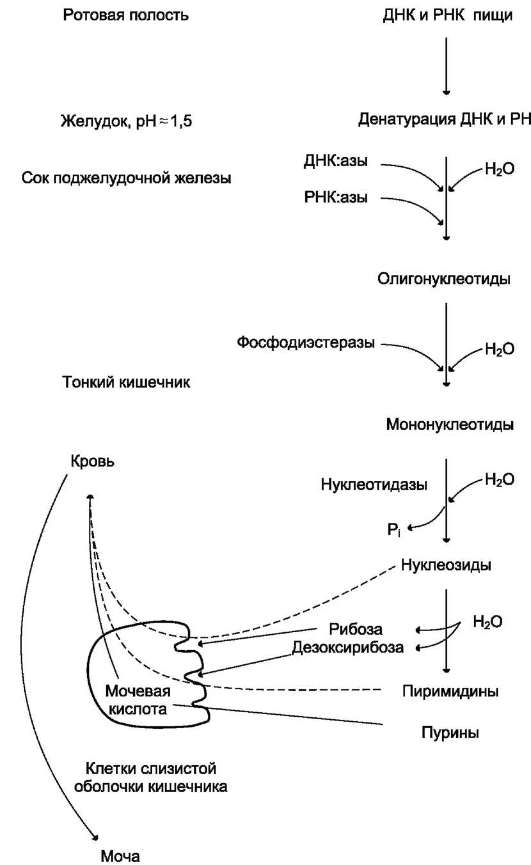
Рис. 10-1. Переваривание нуклеиновых кислот пищи.
novo. Введённые в кровь азотистые основания и нуклеозиды, а также основания и нуклеозиды, образующиеся в результате внутриклеточного разрушения нуклеиновых кислот, в небольшом количестве могут использоваться для повторного синтеза нуклеотидов по так называемым «запасным» путям.
ii. синтез пуриновых
нуклеотидов
В 40-50-х годах ХХ столетия опытами с мечеными изотопами удалось выяснить происхождение атомов пуринового ядра при синтезе пуринов de novo. Было установлено, что в формировании кольца принимают участие аминокислоты
Асп, Гли, Глн, СО2 и два одноуглеродных производных тетрагидрофолата: метенил-Н4-фолат и формил-Н4-фолат. Этим способом образуется основное количество пуриновых нуклеотидов, тогда как нуклеотиды, синтезирующиеся за счёт повторного использования азотистых оснований или нуклеозидов, составляют не более 10-20% общего фонда этих соединений.
А. ОБРАЗОВАНИЕ 5-ФОСФОРИБОЗИЛ-1-ДИФОСФАТА
Фосфорибозилдифосфат (ФРДФ), или фос-форибозилпирофосфат (ФРПФ) занимает центральное место в синтезе как пуриновых, так и пиримидиновых нуклеотидов (рис. 10-2).
Он образуется за счёт переноса β,γ-пирофос-фатного остатка ATФ на рибозо-5-фосфат в реакции, катализируемой ФРДФ-синтетазой.
Источниками рибозо-5-фосфата могут быть: пентозофосфатный путь превращения глюкозы или катаболизм нуклеозидов, в ходе которого под действием нуклеозидфосфорилазы первоначально образуется рибозо-1-фосфат, а затем с помощью соответствующей мутазы фосфатный остаток переносится в 5-положение.
ФРДФ участвует не только в синтезе пуриновых и пиримидиновьгх нуклеотидов из простых предшественников (т.е. de novo), но используется на образование пуриновых нуклеотидов по «запасному» пути и в синтезе нуклеотидных коферментов.
Б. БИОСИНТЕЗ ПУРИНОВЫХ НУКЛЕОТИДОВ DE NOVO
Сборка пуринового гетероцикла осуществляется на остатке рибозо-5-фосфата при участии различных доноров углерода и азота (рис. 10-3).
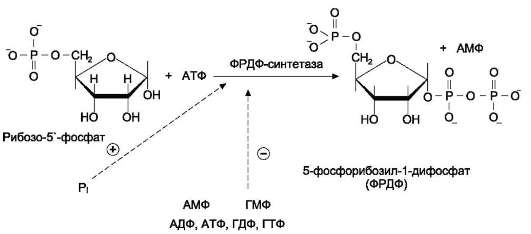
Рис. 10-2. Образование 5-фосфорибозил-1-дифосфата.
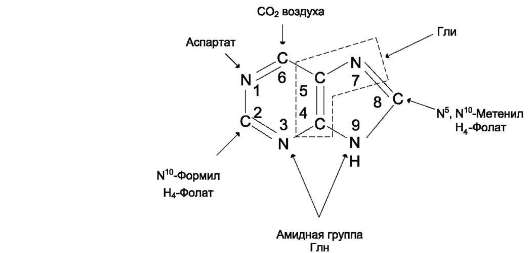
Рис. 10-3. Происхождение атомов С и N в пуриновом кольце.
Включение простых предшественников в пуриновое кольцо с образованием ИМФ
Первая специфическая реакция образования пу-риновых нуклеотидов - перенос амидной группы Глн на ФРДФ с образованием 5-фосфорибозил-1-амина (рис. 10-4). Эту реакцию катализирует фермент амидофосфорибозилтрансфераза. При этом формируется β-Ν-гликозидная связь.
Затем к аминогруппе 5-фосфорибозил-1-амина присоединяются остаток глицина, N5, N10-мете-нил-Н4-фолата ещё одна амидная группа глута-мина, диоксид углерода, аминогруппа аспартата и формильный остаток N10-формил Н4-фолата.
Результатом этой десятистадийной серии реакций является образование первого пуринового нуклеотида - инозин-5'-монофосфата (ИМФ), на синтез которого затрачивается не менее шести молекул АТФ. В отличие от прокариотов, у которых каждую стадию этого процесса катализирует отдельный фермент, у эукариотов за счёт слияния генов возникли полифункциональные ферменты, каждый из которых катализирует несколько реакций. В синтезе пуриновых нук-леотидов de novo это реакции 3, 4 и 6, 7-8 и 10-11 соответственно.
ИМФ в основном используется на синтез АМФ или ГМФ. Небольшое количество этого продукта обнаруживается также в тРНК в качестве одного из минорных нуклеотидов.
Превращение ИМФ в АМФ и ГМФ в обоих случаях включает 2 стадии и идёт с затратой энергии (рис. 10-5).
Аденилосукцинатсинтетаза, используя энергию ГТФ, присоединяет аспартат к ИМФ с образованием аденилосукцината, который в реакции, катализируемой аденилосукциназой, отщепляет фумарат и превращается в АМФ.
Второй пуриновый нуклеотид (ГМФ) образуется также в 2 стадии. Сначала ИМФ окисляется NАD+-зависимой ИМФ-дегидрогеназой с образованием ксантозин-5 -монофосфата (КМФ). Последующее трансамидирование гидроксиль-ной группы при С2-пуринового кольца КМФ катализирует ГМФ-синтетаза с использованием амидной группы Глн и энергии АТФ.
При образовании пуриновых нуклеотидов ГТФ расходуется на синтез АМФ, а АТФ - на синтез ГМФ. Перекрёстное использование пу-риновых нуклеозидтрифосфатов на образование конечных продуктов синтеза помогает
поддерживать в клетках баланс адениловых и гуаниловых нуклеотидов.
Печень - основное место образования пури-новых нуклеотидов, откуда они могут поступать в ткани, не способные к их синтезу: эритроциты, ПЯЛ и частично мозг.
Образование нуклеозидди- и трифосфатов
В образовании нуклеиновых кислот, некоторых коферментов и во многих синтетических процессах нуклеотиды используются в виде ди-и трифосфатов, синтез которых катализируют ферменты класса трансфераз. АМФ и ГМФ превращаются в нуклеозиддифосфаты (НДФ) с помощью специфичных к азотистому основанию нуклеозидмонофосфаткиназ (НМФ-киназ) и АТФ. Так, аденилаткиназа катализирует реакцию:
амф + атф → 2 адф, а гуанилаткиназа:
гмф + атф → гдф + адф.
Аденилаткиназа особенно активна в печени и мышцах, где высок уровень энергоёмких процессов. Функция этого фермента заключается в том, чтобы поддерживать в тканях равновесие фонда адениловых нуклеотидов: AMФ, AДФ и ATФ.
Взаимопревращения нуклеозиддифосфатов и нуклеозидтрифосфатов осуществляет нуклео-зиддифосфаткиназа. Этот фермент в отличие от НМФ-киназ обладает широкой субстратной специфичностью и, в частности, может катализировать реакцию:
гдф + атф → гтф + адф.
Превращение AДФ в ATФ происходит, в основном, за счёт окислительного фосфорили-рования или в реакциях субстратного фосфо-рилирования гликолиза или цитратного цикла.
В. «ЗАПАСНЫЕ» ПУТИ СИНТЕЗА ПУРИНОВЫХ НУКЛЕОТИДОВ (РЕУТИЛИЗАЦИЯ АЗОТИСТЫХ ОСНОВАНИЙ И НУКЛЕОЗИДОВ)
Огромные затраты энергии для синтеза пуриновых нуклеотидов de novo не способны полностью обеспечить субстратами синтез нуклеиновых кислот в период гаструляции и
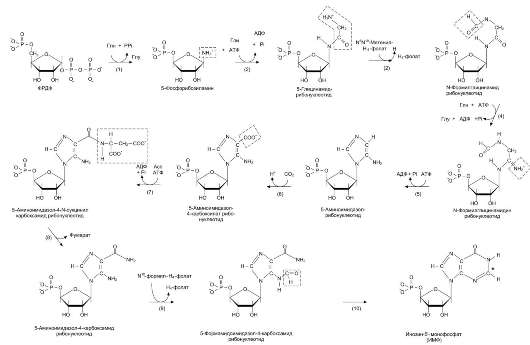
Рис. 10-4. Синтез пуриновых нуклеотидов de novo.
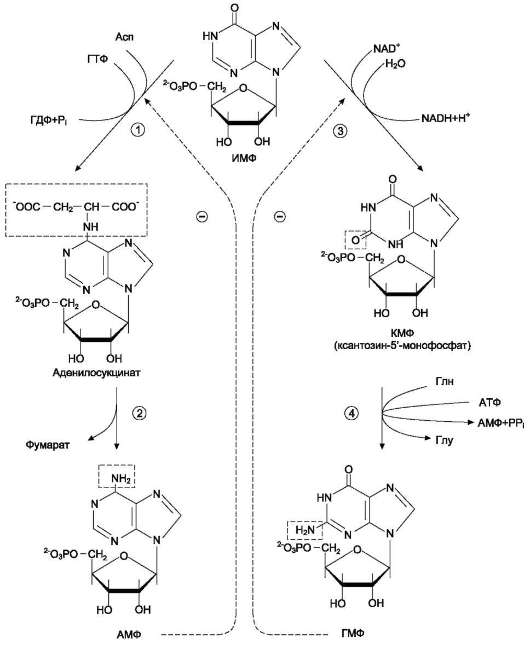
Рис. 10-5. Синтез АМФ и ГМФ из ИМФ. 1 - аденилосукцинатсинтетаза; 2 - аденилосукциназа; 3 - ИМФ-дегидрогеназа; 4 - ГМФ-синтетаза.
раннего роста ребёнка. Потребность в большом количестве нуклеотидов привела к развитию «запасных» путей синтеза этих «дорогих» молекул. Наибольшее значение в этом процессе имеют ферменты, осуществляющие превращение пуринов в мононуклеотиды с использованием ФРДФ как донора остатка фосфорибозы.
Синтез АМФ и ГМФ из аденина и гуанина
ФРДФ-зависимое фосфорибозилирование пуринов катализируют 2 фермента.
Аденинфосфорибозилтрансфераза, ответственная
за образование АМФ (рис. 10-6). Гипоксантин-гуанинфосфорибозилтрансфераза,
катализирующая образование ИМФ и ГМФ из гипоксантина и гуанина соответственно
(рис. 10-7).
Однако в организме при любых ситуациях этот путь синтеза пуриновых нуклеотидов, получивший название «путь спасения», имеет вспомогательное значение.
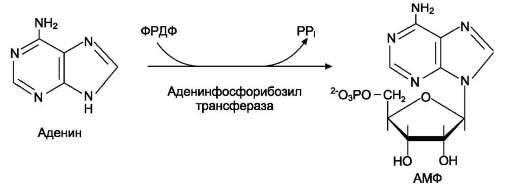
Рис. 10-6. Фосфорибозилирование аденина в АМФ.
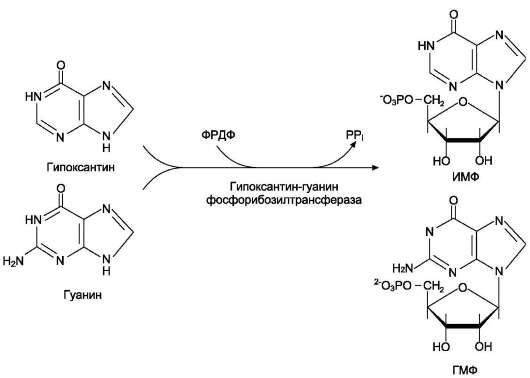
Рис. 10-7. Фосфорибозилирование гипоксантина и гуанина с образованием ИМФ и ГМФ.
Нуклеозидкиназы
Нуклеозиды, получающиеся при катаболизме нуклеиновых кислот из нуклеотидов под действием нуклеотидаз, могут повторно фосфори-лироваться, образуя нуклеозид-5 -монофосфаты за счёт переноса γ-фосфатного остатка АТФ на соответствующий субстрат. У млекопитающих такой путь пополнения запасов пуриновых нук-леотидов в клетке не имеет существенного значения. Основным ферментом этой группы является аденозинкиназа, которая ускоряет реакцию:
Аденозин + АТФ → АМФ + АДФ.
Из всех способов реутилизации пуринов наиболее активна гипоксантин-гуанинфосфори-бозилтрансферазная реакция, поскольку ИМФ, образующийся в этой реакции, вовлекается в синтез АМФ и ГМФ. Использование гипоксан-тина и гуанина по запасному пути становится жизненно важным событием в клетках, не способных к синтезу пуриновых нуклеотидов de novo. Значение аденинфосфорибозилтранс-феразы в повторном использовании аденина
менее существенно. По сравнению с аденозином количество аденина в клетках мало, а первый возвращается в фонд нуклеотидов с помощью аденозинкиназы.
Г. РЕГУЛЯЦИЯ СИНТЕЗА ПУРИНОВЫХ НУК-ЛЕОТИДОВ
Основным показателем, от которого зависит синтез пуриновых нуклеотидов, служит концентрация ФРДФ, которая, в свою очередь, зависит от скорости его синтеза, утилизации и разрушения. Количество ФРДФ определяется доступностью рибозо-5-фосфата и активностью ФРДФ син-тетазы - фермента, чувствительного к концентрации фосфата и пуриновых нуклеотидов.
Внутриклеточная концентрация ФРДФ строго регулируется и обычно низкая. ФРДФ синтета-за - аллостерический фермент. Он активируется неорганическим фосфатом (Pi) и ингибируется пуриновыми нуклеозидмоно-, ди- и трифос-
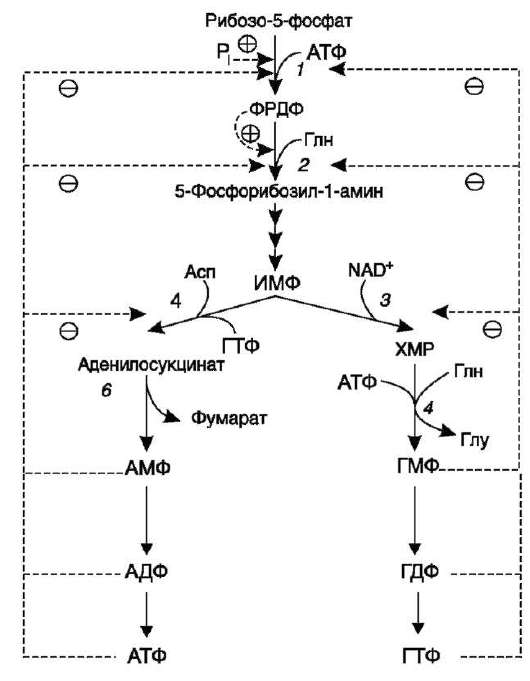
Рис. 10-8. Регуляция синтеза пуриновых нуклеотидов. 1 - ФРДФ синтетаза; 2 - амидофосфорибозил-трансфераза; 3 - ИМФ дегидрогеназа; 4 - аденило-сукцинатсинтетаза.
фатами, которые по эффективности ингибиро-вания распределяются в следующем порядке: НМФ > НДФ > НТФ (рис. 10-8). ФРДФ служит не только субстратом, но и аллостерическим активатором второй реакции синтеза пури-нонуклеотидов de novo, которую катализирует амидофосфорибозилтрансфераза.
Пуриновые нуклеотиды, особенно АМФ и ГМФ по механизму отрицательной обратной связи ингибируют амидофосфорибозилтрансфе-разу, которая катализирует первую специфическую реакцию синтеза пуриновых нуклеотидов de novo.
Метаболическая цепь образования АМФ и ГМФ de novo регулируется также в месте её разветвления: АМФ ингибирует аденилосукци-натсинтетазу, а ГМФ - реакцию образования ксантиловой кислоты, которую катализирует ИМФ дегидрогеназа. Перекрёстная регуляция путей использования ИМФ служит для того, чтобы снизить синтез одного пуринового нук-леотида при дефиците другого.
Помимо ферментов основного пути синтеза пуриновых нуклеотидов de novo, регулируется также активность ферментов «запасных» путей: аденинфосфорибозилтрансфераза ингибируется АМФ, а гипоксантин-гуанинфосфорибозил-трансфераза - ИМФ и ГМФ.
iii. катаболизм пуриновых
нуклеотидов
У человека основной продукт катаболизма пуриновых нуклеотидов - мочевая кислота. Её образование идёт путём гидролитического отщепления фосфатного остатка от нуклеотидов с помощью нуклеотидаз или фосфатаз, фос-форолиза N-гликозидной связи нуклеозидов пуриннуклеозидфосфорилазой, последующего дезаминирования и окисления азотистых оснований (рис. 10-9).
От АМФ и аденозина аминогруппа удаляется гидролитически аденозиндезаминазой с образованием ИМФ или инозина. ИМФ и ГМФ превращаются в соответствующие нуклеозиды: инозин и гуанозин под действием 5 -нуклеотида-зы. Пуриннуклеозидфосфорилаза катализирует расщепление N-гликозидной связи в инозине и гуанозине с образованием рибозо-1-фосфата и азотистых оснований: гуанина и гипоксанти-
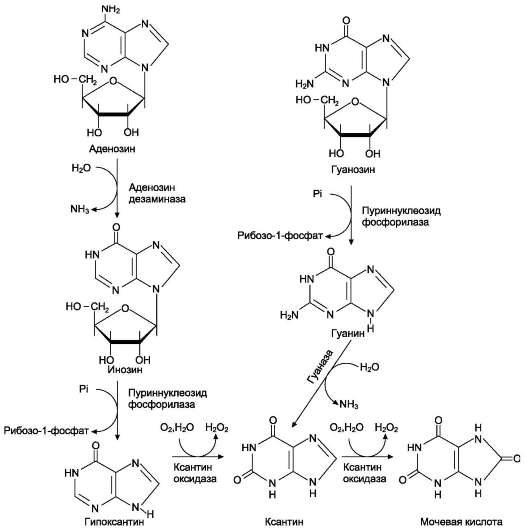
Рис. 10-9. Катаболизм пуриновых нуклеотидов до мочевой кислоты.
на. Гуанин дезаминируется и превращается в ксантин, а гипоксантин окисляется в ксантин с помощью ксантиноксидазы, которая катализирует и дальнейшее окисление ксантина в мочевую кислоту.
Ксантиноксидаза - аэробная оксидоредук-таза, простетическая группа которой включает ион молибдена, железа (Fe3+) и FAD. Подобно другим оксидазам, она окисляет пурины моле-
кулярным кислородом с образованием перок-сида водорода. В значительных количествах фермент обнаруживается только в печени и кишечнике.
Мочевая кислота удаляется из организма главным образом с мочой и немного через кишечник с фекалиями. У всех млекопитающих, кроме приматов и человека, имеется фермент уриказа, расщепляющий мочевую кислоту с
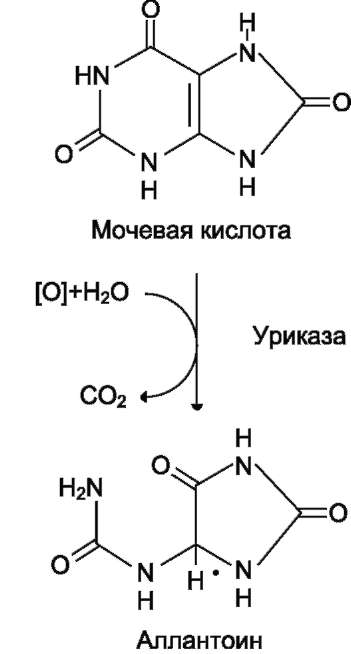
Рис. 10-10. Превращение мочевой кислоты в ал-лантоин.
образованием аллантоина, хорошо растворимого в воде (рис. 10-10).
Амфибии, птицы и рептилии, подобно человеку, лишены уриказы и экскретируют мочевую кислоту и гуанин в качестве конечных продуктов обмена.
Мочевая кислота является слабой кислотой. Содержание недиссоциированной формы и солей (уратов) зависит от рН раствора. При физиологических значениях рН у мочевой кислоты может диссоциировать только один протон из трёх (рК = 5,8), поэтому в биологических жидкостях присутствует как недиссоциированная кислота в комплексе с белками, так и её натриевая соль.
В сыворотке крови в норме содержание мочевой
кислоты составляет 0,15-0,47 ммоль/л или 3-7 мг/дл. Ежесуточно из
организма выводится от 0,4 до
iv. нарушения обмена пуриновых нуклеотидов
Ураты значительно более растворимы, чем мочевая кислота: так, в моче с рН 5,0, когда мочевая кислота не диссоциирована, её растворимость в 10 раз меньше, чем в моче с рН 7,0,
при котором основная часть мочевой кислоты представлена солями. Реакция мочи зависит от состава пищи, но, как правило, она слабокислая, поэтому большинство камней в мочевыводящей системе - кристаллы мочевой кислоты.
А. ГИПЕРУРИКЕМИЯ И ПОДАГРА
Когда в плазме крови концентрация мочевой кислоты превышает норму, то возникает гиперурикемия. Вследствие гиперурикемии может развиться подагра - заболевание, при котором кристаллы мочевой кислоты и уратов откладываются в суставных хрящах, синовиальной оболочке, подкожной клетчатке с образованием подагрических узлов, или тофусов. К характерным признакам подагры относят повторяющиеся приступы острого воспаления суставов (чаще всего мелких) - так называемого острого подагрического артрита. Заболевание может прогрессировать в хронический подагрический артрит.
Поскольку лейкоциты фагоцитируют кристаллы уратов, то причиной воспаления является разрушение лизосомальных мембран лейкоцитов кристаллами мочевой кислоты. Освободившиеся лизосомальные ферменты выходят в цитозоль и разрушают клетки, а продукты клеточного катаболизма вызывают воспаление.
Общий фонд сывороточных уратов в норме составляет ~
Подагра - распространённое заболевание, в разных странах ею страдают от 0,3 до 1,7% населения. А поскольку сывороточный фонд уратов у мужчин в 2 раза больше, чем у женщин, то они и болеют в 20 раз чаще, чем женщины.
Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе ФРДФ синтетазы или ферментов «запасного» пути: гипоксантин-гуа-нинили аденинфосфорибозилтрансфераз.
К другим характерным проявлениям подагры относят нефропатию, при которой наблюдают образование уратных камней в мочевыводящих путях.
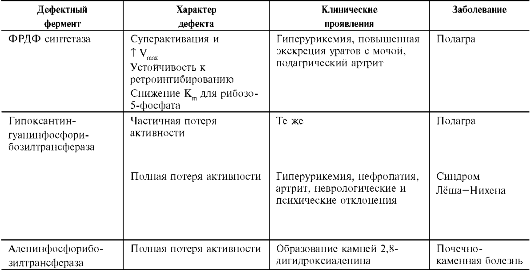
Полиморфные варианты ФРДФ синтетазы
Активность ФРДФ синтетазы, катализирующей образование ФРДФ, строго контролируется пу-риновыми нуклеотидами. Мутации в гене ФРДФ синтетазы привели к появлению полиморфных вариантов фермента, которые характеризуются аномальным ответом на обычные регуляторные факторы: концентрацию рибозо-5-фосфата и пуриннуклеотидов. Как правило, наблюдается суперактивация фермента. Пуриновые нуклеотиды синтезируются со скоростью, почти независимой от нужд клетки. Это вызывает ингибиро-вание запасных «путей спасения», усиление катаболизма избыточного количества нуклеоти-дов, повышение продукции мочевой кислоты, гиперурикемию и подагру (табл. 10-1).
Примерно у 40% больных одной из форм гликогеноза - болезнью Гирке (недостаточностью глюкозо-6-фосфатазы) сопутствующей патологией является подагра. Снижение способности печени секретировать глюкозу в кровь увеличивает использование глюкозо-6-фосфата в пентозофосфатном пути. Образуются большие количества рибозо-5-фосфата, которые могут стимулировать избыточный синтез, а следовательно, и катаболизм пуриновых нуклеотидов.
Б. НЕДОСТАТОЧНОСТЬ ФЕРМЕНТОВ «ЗАПАСНЫХ ПУТЕЙ» СИНТЕЗА ПУРИНОВЫХ НУКЛЕОТИДОВ. СИНДРОМ ЛЁША-НИХЕНА
В ряде случаев причиной гиперурикемии, избыточной экскреции пуринов с мочой и подагры являются нарушения в работе ферментов «пути спасения» пуриновых оснований (табл. 10-1). Гипоксантин-гуанин фосфорибозилтранс-фераза катализирует реакцию превращения гуанина и гипоксантина в соответствующие нук-леотиды (рис. 10-7). Обнаружены полиморфные варианты гипоксантин-гуанинфосфорибозил-трансферазы со сниженной ферментативной активностью, что:
• уменьшает повторное использование пури-новых оснований, и они превращаются в мочевую кислоту;
• увеличивает синтез пуриновых нуклеотидов de novo из-за слабого использования ФРДФ в реакциях реутилизации и увеличения его концентрации в клетке. Адениловые и гуаниловые нуклеотиды образуются в количествах, превышающих потребности клеток, а это способствует усилению их катаболизма.
Таблица 10-1. Гиперурикемия, вызванная дефектами в работе ферментов обмена пуриннуклеотидов
Синдром Лёша-Нихена - тяжёлая форма ги-перурикемии, которая наследуется как рецессивный признак, сцепленный с Х-хромосомой, и проявляется только у мальчиков.
Болезнь вызвана полным отсутствием активности гипоксантин-гуанинфосфорибозилтранс-феразы и сопровождается гиперурикемией с содержанием мочевой кислоты от 9 до 12 мг/дл, что превышает растворимость уратов при нормальном рН плазмы. Экскреция мочевой кислоты у больных с синдромом Лёша-Нихена превышает 600 мг/сут и требует для выведения этого количества продукта не менее 2700 мл мочи.
У детей с данной патологией в раннем возрасте появляются тофусы, уратные камни в моче-выводящих путях и серьёзные неврологические отклонения, сопровождающиеся нарушением речи, церебральными параличами, снижением интеллекта, склонностью к нанесению себе увечий (укусы губ, языка, пальцев).
В первые месяцы жизни неврологические расстройства не обнаруживаются, но на пелёнках отмечают розовые и оранжевые пятна, вызванные присутствием в моче кристаллов мочевой кислоты. При отсутствии лечения больные погибают в возрасте до 10 лет из-за нарушения функции почек.
Полная потеря активности аденинфосфо-рибозилтрансферазы не столь драматична, как отсутствие гипоксантин-гуанинфосфорибозил-трансферазы, однако и в этом случае нарушение повторного использования аденина вызывает гиперурикемию и почечнокаменную болезнь, при которой наблюдается образование кристаллов 2,8-дигидроксиаденина.
В. ЛЕЧЕНИЕ ГИПЕРУРИКЕМИИ
Основным препаратом, используемым для лечения гиперурикемии, является аллопуринол - структурный аналог гипоксантина (рис. 1011).
Аллопуринол оказывает двоякое действие на обмен пуриновых нуклеотидов:
• ингибирует ксантиноксидазу и останавливает катаболизм пуринов на стадии образования гипоксантина, растворимость которого почти в 10 раз выше, чем мочевой кислоты. Действие препарата на фермент объясняется тем, что сначала он, подобно гипоксантину, окисляется в гидроксипуринол, но при этом
• остаётся прочно связанным с активным центром фермента, вызывая его инактивацию; с другой стороны, будучи псевдосубстратом, аллопуринол может превращаться в нуклео-тид по «запасному» пути и ингибировать ФРДФ синтетазу и амидофосфорибозил-трансферазу, вызывая торможение синтеза пуринов de novo. При лечении аллопуринолом детей с синдромом Лёша-Нихена удаётся предотвратить развитие патологических изменений в суставах и почках, вызванных гиперпродукцией мочевой кислоты, но препарат не излечивает аномалии в поведении, неврологические и психические расстройства.
Г. ГИПОУРИКЕМИЯ
Гипоурикемия и возросшая экскреция ги-поксантина и ксантина может быть следствием недостаточности ксантиноксидазы, вызванной нарушениями в структуре гена этого фермента, либо результатом повреждения печени.
v. биосинтез пиримидиновых нуклеотидов
Фонд пиримидиновых нуклеотидов, подобно пуриновым нуклеотидам, в основном синтезируется из простых предшественников de novo, и только 10-20% от общего количества образуется по «запасным» путям из азотистых оснований или нуклеозидов.
А. ОБРАЗОВАНИЕ ПИРИМИДИНОВЫХ
НУКЛЕОТИДОВ DE NOVO
В отличие от синтеза пуринов, где формирование гетероциклического основания осуществляется на остатке рибозо-5-фосфа-
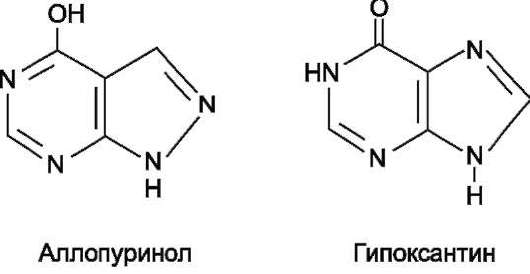
Рис. 10-11. Строение аллопуринола и гипоксантина.
та, пиримидиновое кольцо синтезируется из простых предшественников: глутамина, СО2 и аспарагиновой кислоты и затем связывается с рибозо-5-фосфатом, полученным от ФРДФ.
Процесс протекает в цитозоле клеток. Синтез ключевого пиримидинового нуклеотида - УМФ идёт с участием 3 ферментов, 2 из которых полифункциональны.
Образование дигидрооротата
У млекопитающих ключевой, регуляторной реакцией в синтезе пиримидиновых нуклеотидов является синтез карбамоилфосфата из глута-мина, СО2 и АТФ, в реакции катализируемой карбамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле клеток (рис. 10-12). В реакции NH2-группа карбамоилфосфата образуется за счёт амидной группы глутамина, что отличает эту реакцию от реакции синтеза карбамоилфосфата в митохондриях в процессе синтеза мочевины из CO2, NH3 и АТФ с участием КФС I.
Карбамоилфосфат, использующийся на образование пиримидиновых нуклеотидов, является продуктом полифункционального фермента, который наряду с активностью КФС II содержит каталитические центры аспартаттранскарбамои-лазы и дигидрооротазы. Этот фермент назвали «КАД-фермент» - по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка. Объединение первых трёх ферментов метаболического пути в единый полифункциональный комплекс позволяет использовать почти весь синтезированный в первой реакции карбамо-илфосфат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт - дигидрооротат (рис. 10-13).
Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зависимой
дигидрооротатдегидрогеназой и превращается в свободное пиримидиновое основание - орото-вую кислоту, или оротат.
Образование УМФ
В цитозоле оротат становится субстратом бифункционального фермента - УМФ-синта-зы, которая обнаруживает оротатфосфорибо-зилтранс-феразную и ОМФ-декарбоксилазную активности. Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нуклеотид - оротидин-5 -монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5 -монофосфат (УМФ).
Таким образом, шесть последовательных реакций синтеза пиримидиновых нуклеотидов осуществляются тремя ферментами, которые кодируются в геноме человека тремя различными структурными генами.
Биосинтез УДФ, УТФ и цитидиловых нуклеотидов
УМФ под действием специфических нуклео-зидмонофосфат (НМФ) и нуклеозиддифосфат (НДФ) киназ превращается в УДФ и УТФ в результате переноса γ-фосфатного остатка АТФ на соответствующий субстрат.
НМФ-киназа катализирует следующую реакцию:
умф + атф → удф + адф, а НДФ-киназа:
удф + атф → утф + адф.
ЦТФ синтетаза катализирует амидирование УТФ (рис. 10-14), осуществляя АТФ-зависимое замещение кетогруппы урацила на амидную группу глутамина с образованием цитидин-5 -трифосфата (ЦТФ).
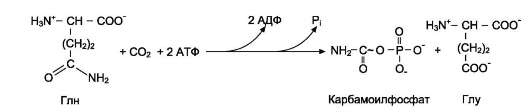
Рис. 10-12. Синтез карбамоилфосфата.
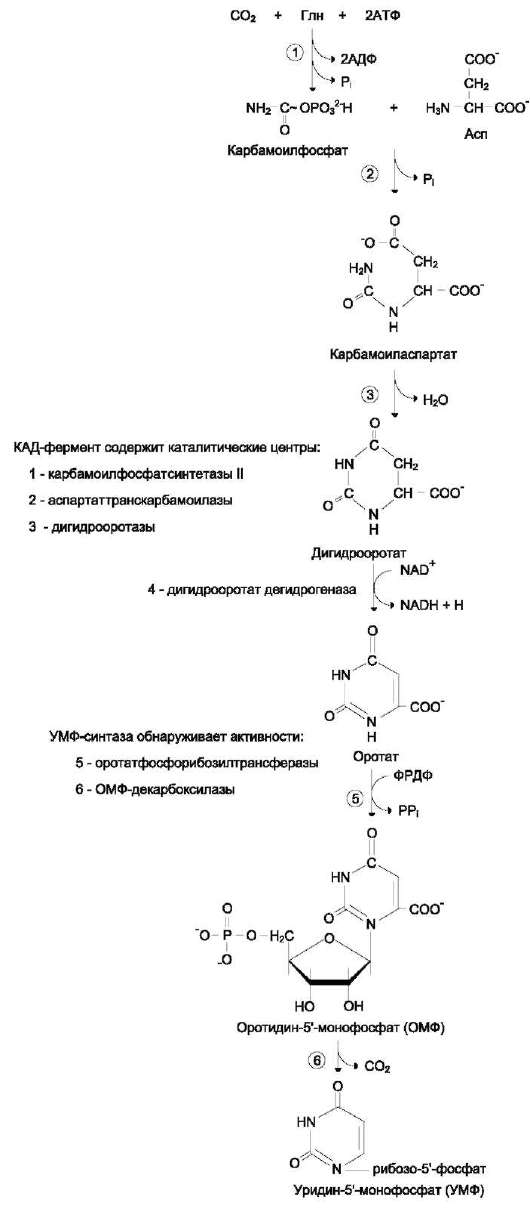
Рис. 10-13. Биосинтез УМФ de novo.
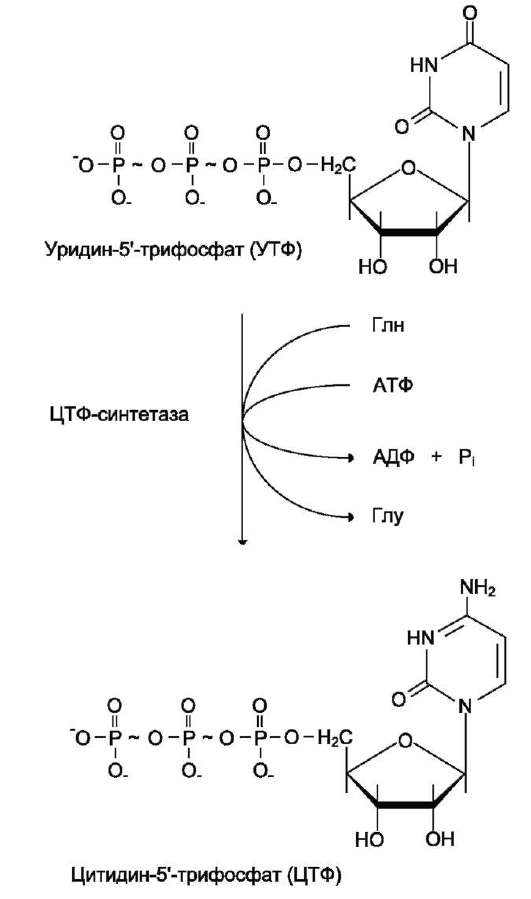
Рис. 10-14. Синтез ЦТФ из УТФ.
Б. «ЗАПАСНЫЕ» ПУТИ СИНТЕЗА ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Использование пиримидиновьгх оснований и нуклеозидов в реакциях реутилизации препятствует катаболизму этих соединений до конечных продуктов с расщеплением пиримидинового кольца. В ресинтезе пиримидинов участвуют некоторые ферменты катаболизма нуклеотидов. Так, уридинфосфорилаза в обратимой реакции может рибозилировать урацил с образованием уридина.
Урацил + Рибозо-1-фосфат → Уридин + Н3РО4.
Превращение нуклеозидов в нуклеотиды катализирует уридин-цитидинкиназа.
Часть ЦМФ может превращаться в УМФ под действием цитидиндезаминазы и пополнять запасы уридиловых нуклеотидов.
ЦМФ + H2O → УМФ + NH3.
в. регуляция синтеза пиримидиновых нуклеотидов
Регуляторным ферментом в синтезе пири-мидиновых нуклеотидов является полифункциональный КАД-фермент. УМФ и УТФ ал-лостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ
(рис. 10-15).
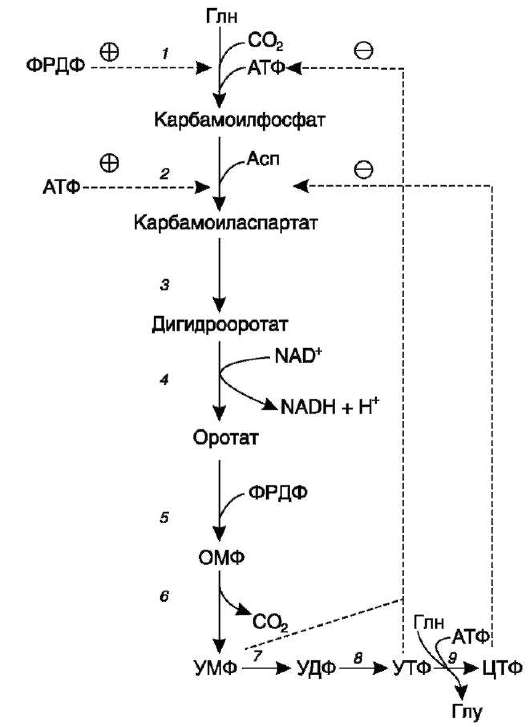
Рис. 10-15. Регуляция синтеза пиримидиновых нуклеотидов. КАД-фермент катализирует реакции 1, 2, 3; дигидрооротатдегидрогеназа - реакцию 4; УМФ син-тетаза - реакции 5 и 6; НМФ киназа - реакцию 7; НДФ киназа - реакцию 8; ЦТФ синтетаза - реацию 9.
Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидино-вых нуклеотидов, необходимых для синтеза
РНК.
VI. КАТАБОЛИЗМ ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Уже говорилось о том, что цитидиловые нук-леотиды могут гидролитически терять аминогруппу и превращаться в УМФ. Когда от УМФ при участии нуклеотидазы (или фосфатазы) и уридинфосфорилазы отщепляются неорганический фосфат и рибоза, то остаётся азотистое основание - урацил. Аналогично расщепляются дезоксирибонуклеотиды, и из dЦМФ образуется урацил, а из dТМФ - тимин (рис. 10-16).
Пиримидиновые основания при участии ди-гидропиримидиндегидрогеназы присоединяют 2 атома водорода по двойной связи кольца с образованием дигидроурацила или дигидроти-мина. Оба гетероцикла могут взаимодействовать с водой в реакции, катализируемой дигидро-пиримидинциклогидролазой, и дигидроурацил превращается в β-уреидопропионовую кислоту, а дигидротимин - в β-уреидоизомасляную кислоту. Оба β-уреидопроизводных под действием общего для них фермента уреидопропионазы расщепляются с образованием СО2, NH4+ и β-аланина или β-аминоизомасляной кислоты соответственно.
β-Аланин обнаруживают в плазме крови и многих тканях. Он используется в мышцах на образование дипептидов: карнозина и анзерина. Под действием бактериальной микрофлоры кишечника β-аланин включается в пантотеновую кислоту, которая всасывается и используется на образование KoA.
Часть β-аланина и β-аминоизобутирата тран-саминируется с α-кетоглутаратом и даёт малонил полуальдегид или метилмалонил полуальдегид соответственно, которые превращаются в ма-лонил-КоА и сукцинил-КоА и используются в соответствующих метаболических путях, либо окисляются до СО2 и Н2О. Частично β-амино-изобутират экскретируется с мочой.
VII. НАРУШЕНИЯ ОБМЕНА ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Описано несколько нарушений, связанных со снижением активности ферментов обмена пиримидиновых нуклеотидов. Одно из них - оро-тацидурия - вызвано дефектом в работе второго бифункционального фермента синтеза нуклеоти-дов de novo - УМФ-синтазы, два других обнаружены в процессе катаболизма пиримидинов.
А. ОРОТАЦИДУРИЯ
Это единственное нарушение синтеза пирими-динов de novo. Оно вызвано снижением активности УМФ-синтазы, которая катализирует образование и декарбоксилирование ОМФ. Поскольку в эмбриогенезе от образования пиримидинов de novo зависит обеспечение синтеза ДНК субстратами, то жизнь плода невозможна при полном отсутствии активности этого фермента. Действительно, у всех пациентов с оротацидурией отмечают заметную, хотя и очень низкую активность УМФ-синтазы. Установлено, что содержание оротовой кислоты в моче пациентов (1 г/сут и более) значительно превосходит количество оро-тата, которое ежедневно синтезируется в норме (около 600 мг/сут). Снижение синтеза пирими-диновых нуклеотидов, наблюдающееся при этой патологии, нарушает регуляцию КАД-фермента по механизму ретроингибирования, из-за чего возникает гиперпродукция оротата.
Клинически наиболее характерное следствие оротацидурии - мегалобластная анемия, вызванная неспособностью организма обеспечить нормальную скорость деления клеток эритро-цитарного ряда. Её диагностируют у детей на том основании, что она не поддаётся лечению препаратами фолиевой кислоты.
Недостаточность синтеза пиримидиновых нуклеотидов сказывается на интеллектуальном развитии, двигательной способности и сопровождается нарушениями работы сердца и ЖКТ. Нарушается формирование иммунной системы, и наблюдается повышенная чувствительность к различным инфекциям.
Гиперэкскреция оротовой кислоты сопровождается нарушениями со стороны мочевы-водящей системы и образованием камней. При отсутствии лечения больные обычно погибают в первые годы жизни. При этом оротовая
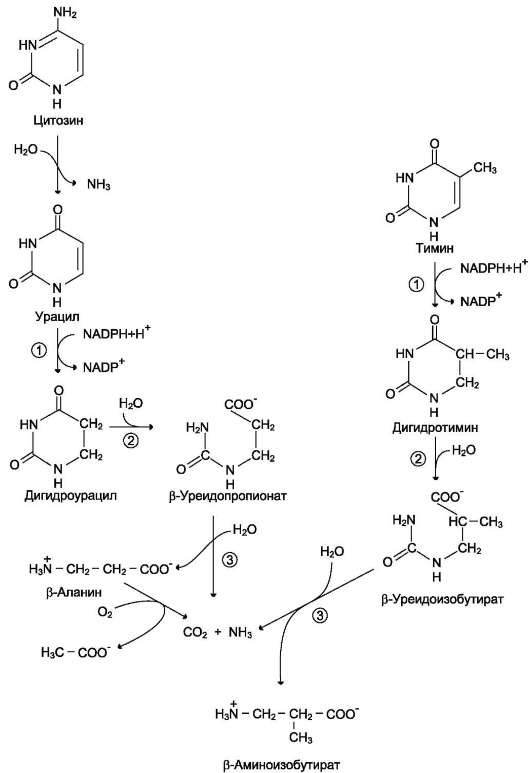
Рис. 10-16. Катаболизм пиримидиновых оснований. 1 - дигидропиримидиндегидрогеназа; 2 - дигидропи-римидинциклогидролаза; 3 - уреидопропионаза.
кислота не оказывает токсического эффекта. Многочисленные нарушения в работе разных систем организма вызваны «пиримидиновым голодом».
Для лечения этой болезни применяют уридин (от 0,5 до 1 г/сут), который по «запасному» пути превращается в УМФ.
Уридин + АТФ → УМФ + АДФ.
Нагрузка уридином устраняет «пиримидино-вый голод», а поскольку из УМФ могут синтезироваться все остальные нуклеотиды пиримиди-нового ряда, то снижается выделение оротовой кислоты из-за восстановления механизма рет-роингибирования КАД-фермента. Для больных оротацидурией лечение уридином продолжается в течение всей жизни, и этот нуклеозид становится для них незаменимым пищевым фактором.
Кроме генетически обусловленных причин, оротацидурия может наблюдаться:
• при гипераммониемии, вызванной дефектом любого из ферментов орнитинового цикла, за исключением карбамоилфосфатсинтетазы I. В этом случае карбамоилфосфат, синтезированный в митохондриях, выходит в цитозоль клеток и начинает использоваться на образование пиримидиновых нуклеоти-дов. Концентрация всех метаболитов, в том числе и оротовой кислоты, повышается. Наиболее значительная экскреция оротата отмечается при недостаточности орнитин-карбамоилтрансферазы (второго фермента орнитинового цикла);
• в процессе лечения подагры аллопурино-лом, который превращается в оксипури-нолмононуклеотид и становится сильным ингибитором УМФ-синтазы. Это приводит к накоплению оротовой кислоты в тканях и крови.
Б. НАРУШЕНИЯ КАТАБОЛИЗМА
ПИРИМИДИНОВ
Известны нарушения в работе 2 ферментов этого метаболического пути.
При недостаточности пиримидин-5'-нуклеоти-дазы нарушаются отщепление неорганического фосфата от пиримидиновых мононуклеотидов и образование нуклеозидов.
Неактивная изоформа пиримидин-5'-нуклео-тидазы обнаружена в эритроцитах. В результате
наблюдается накопление пиримидиновых НТФ, которые ингибируют пентозофосфатный путь превращения глюкозы и тем самым создают предпосылки к гемолизу эритроцитов (см. раздел 14).
Дигидропиримидиндегидрогеназа - ско-рость-лимитирующий фермент катаболизма пи-римидинов. Нарушение работы этого фермента сопровождается отклонениями в функционировании нервной системы и диагностируется на основании повышения уровня свободных пири-мидинов: урацила и тимина в плазме крови.
VIII. БИОСИНТЕЗ ДЕЗОКСИРИБОНУКЛЕОТИДОВ
Синтез дезоксирибонуклеотидов идёт с заметной скоростью только в тех клетках, которые вступают в S-фазу клеточного цикла и готовятся к синтезу ДНК и делению. В покоящихся клетках дезоксинуклеотиды практически отсутствуют. Все дезоксинуклеотиды, кроме тимидиловых, образуются из рибонуклеотидов путём прямого восстановления ОН-группы у второго углеродного атома рибозы в составе рибонуклеозидди-фосфатов до дезоксирибозы. Тимидиловые нук-леотиды синтезируются из сУМФ особым путём с участием N5, N10-метилен-Н4-фолата.
А. РИБОНУКЛЕОТИДРЕДУКТАЗНЫЙ
КОМПЛЕКС
Реакцию восстановления НДФ в дезокси-про-изводные катализирует рибонуклеотидредуктаз-ный комплекс, в состав которого входят: собственно рибонуклеотидредуктаза (РНР), белок тиоредоксин и фермент тиоредоксинредуктаза, обеспечивающий регенерацию восстановленной формы тиоредоксина (рис. 10-17).
Рибонуклеотидредуктаза - олигомерный белок, состоящий из двух В1- и двух В2-субъеди-ниц, и содержит негеминовое железо в качестве кофактора.
Непосредственным донором водорода в реакции восстановления рибозы служит низкомолекулярный белок тиоредоксин. В рабочую часть этого белка входят 2 SH-группы, которые, отдавая водород, окисляются с образованием дисульфидного мостика. Второй фермент комплекса - тиоредоксинредуктаза - катализирует гидрирование окисленного тиоредоксина с использованием NADPH.
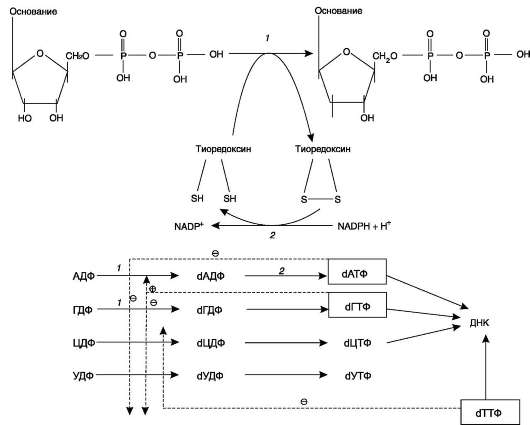
Рис. 10-17. Восстановление рибонуклеозиддифосфатов в 2'-дезоксирибонуклеозиддифосфаты. 1 - рибо-нуклеотидредуктаза (РНР); 2 - тиоредоксинредуктаза.
При участии комплекса РНР образуются: dАДФ, dГДФ, dУДФ и dЦДФ, которые с помощью НДФ-киназ превращаются в дНТФ, 3 из которых (кроме дУДФ) непосредственно используются в синтезе ДНК.
дНДФ + АТФ → дНТФ + АДФ.
Б. БИОСИНТЕЗ ТИМИДИЛОВЫХ НУКЛЕОТИДОВ
Тимидин-5'-монофосфат (дТМФ) образуется из дУМФ в реакции, катализируемой тимиди-латсинтазой (рис. 10-18). Донором метильной группы, появляющейся в 5-положении пири-мидинового кольца в молекуле дТМФ, служит кофермент тимидилатсинтазы - N5, N10-мети-
лен-Н4-фолат. С помощью этого кофермента в молекулу дУМФ включается метиленовая группа и восстанавливается в метильную, используя 2 атома водорода от Н4-фолата.
Образование субстрата тимидилатсинтазной реакции - дУМФ осуществляется двумя путями (рис. 10-19):
• дефосфорилированием дУДФ;
• гидролитическим дезаминированием дЦМФ с помощью дЦМФ дезаминазы. дЦМФ получается при дефосфорилировании дЦДФ - одного из продуктов рибонуклеотидредуктазной реакции. В организме человека это основной путь образования дУМФ.
Скорость синтеза дТМФ зависит также от количества второго субстрата тимидилатсинта-
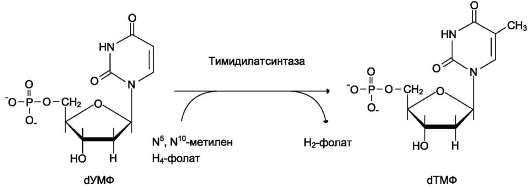
Рис. 10-18. Синтез дТМФ из дУМФ.
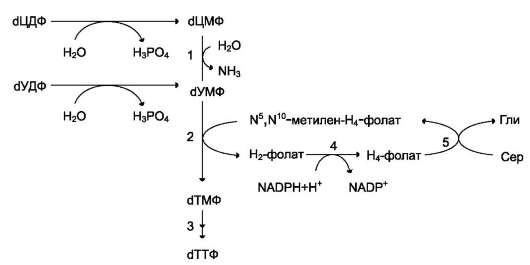
Рис. 10-19. Образование ТТФ из дЦДФ и дУДФ. 1 - дЦМФ дезаминаза; 2 -тимидилатсинтаза; 3 - дНМФ-и дНДФ киназы; 4 - дигидрофолатредуктаза; 5 - серингидроксиметилтрансфераза.
зной реакции - N5, N10-метилен-Н4-фолата, пополнение запасов которого осуществляется при участии 2 ферментов: дигидрофолатредуктазы, которая с участием NADPH восстанавливает Н2-фолат в Н4-фолат, и серин гидроксиметил-трансферазы, осуществляющей перенос β-гид-роксиметиленовой группы серина на Н4-фолат (см. раздел 9). У человека дТМФ образуется, главным образом, из дЦДФ.
В. «ЗАПАСНЫЕ» ПУТИ СИНТЕЗА
ДЕЗОКСИРИБОНУКЛЕОТИДОВ
В быстроделящихся клетках наряду с синтезом дезоксинуклеотидов с помощью рибонуклео-тидредуктазного комплекса и тимидилатсинтазы
активируются реакции, обеспечивающие повторное использование тимина и дезоксицитидина в реакциях, катализируемых ферментами «запасных» путей и обратимых реакций катаболизма. Под влиянием тимидинфосфорилазы протекает следующая реакция:
Тимин + Дезоксирибоза-1-фосфат → Тимидин + H3PO4
Тимидинкиназа катализирует реакцию:
Тимидин + АТФ → дТМФ + АДФ.
Дезоксицитидинкиназа катализирует реакцию образования дЦМФ:
Дезоксицитидин + АТФ → дЦМФ + АДФ.
Г. РЕГУЛЯЦИЯ СИНТЕЗА
ДЕЗОКСИРИБОНУКЛЕОТИДОВ
Рибонуклеотидредуктаза, тимидилатсинтаза и тимидинкиназа - индуцируемые ферменты, их количество в клетке регулируется на генетическом уровне по механизму индукции и репрессии. Синтез этих белков начинает нарастать в G1-пе-риоде, достигает максимума во время активного синтеза ДНК, снижаясь практически до нуля в G2- и М-периоды клеточного цикла.
В то же время активность РНР подвержена сложной аллостерической регуляции, с помощью которой достигается сбалансированное образование всех дНДФ.
РНР осуществляет последовательное восстановление всех рибонуклеозиддифосфатов. Первыми восстанавливаются пиримидиновые нуклеотиды, а последним - дАДФ. дАДФ фосфорилируется в дАТФ, накопление которого полностью прекращает восстановление всех остальных рибонуклео-зиддифосфатов.
Д. НАРУШЕНИЯ В РАБОТЕ РНР, ВЫЗВАННЫЕ НЕДОСТАТОЧНОСТЬЮ ФЕРМЕНТОВ КАТАБОЛИЗМА ПУРИННУКЛЕОЗИДОВ
Аденозиндезаминаза (АДА) и пуриннуклеозид-фосфорилаза (ПНФ) участвуют в превращении пуриновых нуклеозидов в азотистые основания. Их недостаточность сопровождается развитием тяжёлых форм иммунодефицита.
Недостаточность аденозиндезаминазы. АДА катализирует гидролитическое дезаминирование аденозина и дезоксиаденозина:
Аденозин + Н2O → Инозин + NH3,
Дезоксиаденозин + Н2O → Дезоксиинозин + NH3.
Фермент АДА обнаружен во многих органах и тканях, однако его недостаточность имеет наиболее тяжёлые последствия для клеток лим-фоцитарного ряда. Низкая активность этого фермента нарушает пролиферацию и созревание Т- и В-лимфоцитов и сопровождается тяжёлыми формами клеточного и гуморального иммунодефицита. Дети, страдающие этой патологией, как правило, погибают в раннем возрасте от бактериальных, вирусных или грибковых инфекций.
Столь тяжёлые последствия недостаточности АДА для клеток лимфоцитарного ряда объясняют тем, что при снижении скорости дезаминирова-ния адениловых и дезоксиадениловых нуклео-тидов в клетках увеличивается концентрация дАТФ, который ингибирует РНР. Это нарушает синтез всех дНТФ и лишает клетки субстратов для синтеза ДНК. Для нелимфоцитарных клеток недостаточность АДА не сопровождается нарушениями метаболизма в связи с тем, что в них активно работает фосфатаза дАТФ, которая предотвращает накопление основного ингибитора РНР - дАТФ.
Фермент обладает групповой субстратной специфичностью и использует в качестве субстратов некоторые производные аденозина, которые применяются в терапии онкологических и противовирусных заболеваний (аденозинара-бинозид, формицин).
Недостаточность пуриннуклеозидфосфорилазы (ПНФ). ПНФ катализирует фосфоролиз пури-новых рибо- и дезоксирибонуклеозидов с освобождением азотистых оснований и рибозоили дезоксирибозо-1-фосфата. Субстратами служат гуанозин, дезоксигуанозин и инозин.
Нуклеозид + Н3РС4 → Азотистое основание + Рибозо-1-фосфат.
Фермент обнаружен во многих органах и тканях, но особенно активен в клетках-предшественниках Т-лимфоцитов в процессе их созревания в тимусе. При наследственной недостаточности пуриннуклеозидфосфорила-зы, вызванной генными мутациями, в крови снижается образование и количество зрелых Т-лимфоцитов. Нарушение созревания Т-лим-фоцитов вызвано тем, что в этих клетках высокой активностью обладает дезоксигуанозинкиназа, а это приводит к накоплению дГТФ в концентрациях, которые, подобно дАТФ, ингибируют
РНР.
У детей снижен клеточный иммунитет, хотя гуморальный иммунитет не страдает, так как в В-лимфоцитах дезоксигуанозинкиназа малоактивна и накопления дГТФ в токсических концентрациях не отмечают.
Болезнь, вызванная недостаточностью ПНФ, характеризуется более лёгким течением, чем болезнь, обусловленная дефицитом АДА.
IX. ферменты синтеза рибо-и дезоксирибонуклеотидов как мишени для действия
противовирусных
и противоопухолевых
препаратов
В терапии инфекционных и онкологических болезней, научных исследованиях в области медицины и биологии часто используют синтетические аналоги пуринов и пиримидинов. Введение в организм животного или человека аналога, имеющего изменения в структуре гетероциклического кольца или углеводной компоненты, угнетает активность ферментов, участвующих в метаболизме нуклеотидов, скорость синтеза РНК или ДНК из-за нарушения комплементарных взаимодействий азотистых оснований и роста полинуклеотидных цепей. Аналоги пуринов, пиримидинов и их нуклео-зиды нашли применение в качестве антибактериальных, противовирусных и химиотерапевти-ческих средств.
А. ПРОТИВООПУХОЛЕВЫЕ ПРЕПАРАТЫ
Синтезировано очень много аналогов дНТФ, которые включаются ДНК полимеразами в ДНК и ингибируют репликацию. К числу мощных противоопухолевых препаратов принадлежит 5-фторурацил (5-FU) - аналог урацила.
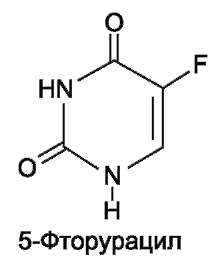
В организме по «запасным» путям 5-FU превращается в 5-F-УМФ либо в реакции, катализируемой оротатфосфорибозилтрансфе-разой, либо через промежуточное образование нуклеозида и последующее фосфорилирование. Превращаясь в нуклеозиддифосфат, 5-FU может участвовать в реакции, катализируемой РНР, и восстанавливаться в соответствующее дезокси-производное. Под действием фосфатазы 5-F-
дУДФ снова теряет фосфат, и образующийся 5-F-дУМФ связывается с тимидилатсинтазой и N5, N10-метилен-Н4-фолатом, образуя комплекс, напоминающий промежуточное соединение в реакции превращения дУМФ в дТМФ. Тими-дилатсинтаза оказывается полностью блокированной, и синтез дТМФ прекращается:
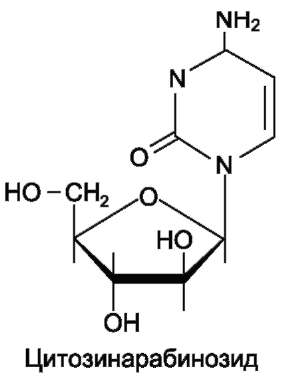
Цитозинарабинозид (или цитарабин) представляет собой соединение, в котором остаток рибозы замещён на стериоизомер - арабинозу. Оно используется в химиотерапии рака, в частности, при острой миелоцитарной лейкемии.
В организме препарат может превращаться в дНТФ, ингибировать ДНК полимеразы и снижать скорость репликации.
Аналоги фолиевой кислоты. В обмене нуклеотидов производные Н4-фолата как доноры одно-углеродных групп участвуют в формировании пуринового гетероциклического кольца и в ключевой реакции синтеза дТМФ из дУМФ, катализируемой тимидилатсинтазой.
В последнем случае N5, N10-метилен-Н4-фолат служит донором метильной группы и в ходе реакции превращается в Н2-фолат. Для активного синтеза тимидиловых нуклеотидов Н2-фолат должен повторно использоваться, проходя стадию восстановления в Н4-фолат (см. формулу ниже).
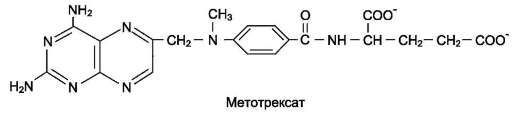
Метотрексат и аминоптерин - структурные аналоги фолиевой кислоты - ингибируют ди-гидрофолатредуктазу и таким образом нарушают синтез пуриновых нуклеотидов и превращение дУМФ в дТМФ, снижая внутриклеточную концентрацию субстратов синтеза ДНК и РНК. Препараты широко используют в химиотерапии опухолей.
Б. АНТИВИРУСНЫЕ И АНТИБАКТЕРИАЛЬНЫЕ
ПРЕПАРАТЫ
Азидотимидин (АZТ, или зидовидин) представляет собой мощный противовирусный препарат, применяющийся в лечении инфекций, которые сопровождают приобретённые формы иммунодефицита. Будучи структурным аналогом тимидина, препарат имеет в 3'-положении дезоксирибозы азидогруппу.
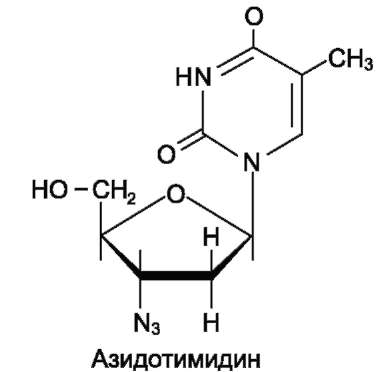
AZT может фосфорилироваться и с помощью ДНК-полимераз включаться в растущую молекулу ДНК. Однако присутствие в З'-положении дезоксирибозы азидогруппы делает синтезирующиеся молекулы ДНК не способными к последующему удлинению. В результате образование новых молекул ДНК прекращается.
Важно, что фосфорилированные производные АZТ утилизируются более эффективно вирусной ДНК-полимеразой или так называемой обратной транскриптазой, чем ДНК-полимеразами эука-риотов, поэтому препарат наиболее эффективно влияет на размножение вирусов и, в частности, ретровируса, вызывающего ВИЧ-инфекцию.
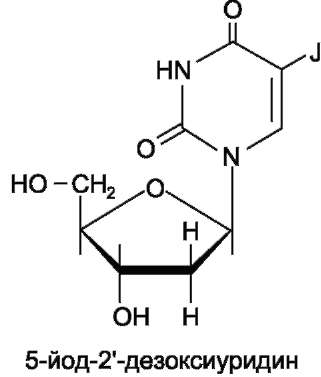
5-Йоддезоксиуридин используют в терапии кератитов и поражений роговицы глаза вирусом герпеса.
Азатиоприн в организме превращается в 6-меркаптопурин, который оказывает мощное им-муносупрессорное действие. Препарат широко используют в трансплантологии для предотвращения развития иммунологических реакций, вызывающих отторжение трансплантата.