Биохимия: учебник для вузов/ под ред. Е.С.Северина - 5-е изд., - 2009. - 768 с.
|
|
|
|
РАЗДЕЛ 12 ОБЕЗВРЕЖИВАНИЕ ТОКСИЧЕСКИХ ВЕЩЕСТВ В ОРГАНИЗМЕ
Печень - самая крупная железа пищеварительного тракта. Она выполняет в организме функцию биохимической лаборатории и играет важную роль в белковом, углеводном и липидном обменах (см. ниже). В печени синтезируются важнейшие белки плазмы крови: альбумин, фибриноген, протромбин, церуло-плазмин, трансферрин, ангиотензиноген и др. Через эти белки опосредуется участие печени в таких важных процессах, как поддержание онкотического давления, регуляция АД и объёма циркулирующей крови, свёртывание крови, метаболизм железа и др.
Важнейшая функция печени - детоксикаци-онная (или барьерная). Она имеет существенное значение для сохранения жизни организма. В печени происходит обезвреживание таких веществ, как билирубин и продукты катаболизма аминокислот в кишечнике, а также инакти-вируются лекарственные препараты и токсические вещества экзогенного происхождения, NH3 - продукт азотистого обмена, который в результате ферментативных реакций превращается в нетоксичную мочевину, гормоны и биогенные амины.
Вещества, поступающие в организм из окружающей среды и не используемые им для построения тканей организма или как источники энергии, называют чужеродными веществами, или ксенобиотиками. Эти вещества могут попадать в организм с пищей, через кожу или с вдыхаемым воздухом.
Чужеродные вещества, или ксенобиотики, делят на 2 группы:
• продукты хозяйственной деятельности человека (промышленность, сельское хозяйство, транспорт);
• вещества бытовой химии - моющие средства, вещества для борьбы с насекомыми, парфюмерия.
Гидрофильные ксенобиотики выводятся из организма в неизменённом виде с мочой, гидрофобные могут задерживаться в тканях, связываясь с белками или образуя комплексы
с липидами клеточных мембран. Со временем накопление в клетках тканей чужеродного вещества приведёт к нарушению их функций. Для удаления таких ненужных для организма веществ в процессе эволюции выработались механизмы их детоксикации (обезвреживания) и выведения из организма.
I. МЕХАНИЗМЫ ОБЕЗВРЕЖИВАНИЯ КСЕНОБИОТИКОВ
Обезвреживание большинства ксенобиотиков происходит путём химической модификации и протекает в 2 фазы (рис. 12-1). В результате этой серии реакций ксенобиотики становятся более гидрофильными и выделяются с мочой. Вещества, более гидрофобные или обладающие большой молекулярной массой (>300 кД), чаще выводятся с жёлчью в кишечник и затем удаляются с фекалиями.
Система обезвреживания включает множество разнообразных ферментов, под действием которых практически любой ксенобиотик может быть модифицирован.
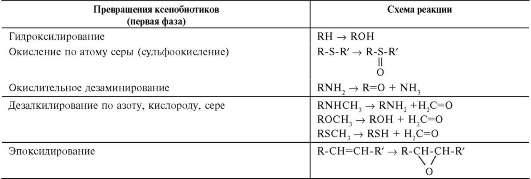
Микросомальные ферменты катализируют реакции С-гидроксилирования, N-гидроксили-рования, О-, N-, S-дезалкилирования, окислительного дезаминирования, сульфоокисления и эпоксидирования (табл. 12-1).
В мембранах ЭР практически всех тканей локализована система микросомального окисления (монооксигеназного окисления). В эксперименте при выделении ЭР из клеток мембрана распадается на части, каждая из которых образует замкнутый пузырёк - микросому, отсюда и название - микросомальное окисление. Эта система обеспечивает первую фазу обезвреживания большинства гидрофобных веществ. В метаболизме ксенобиотиков могут принимать участие ферменты почек, лёгких, кожи и ЖКТ, но наиболее активны они в печени. К группе микросомальных ферментов относят специфические оксидазы, различные гидролазы и ферменты конъюгации.
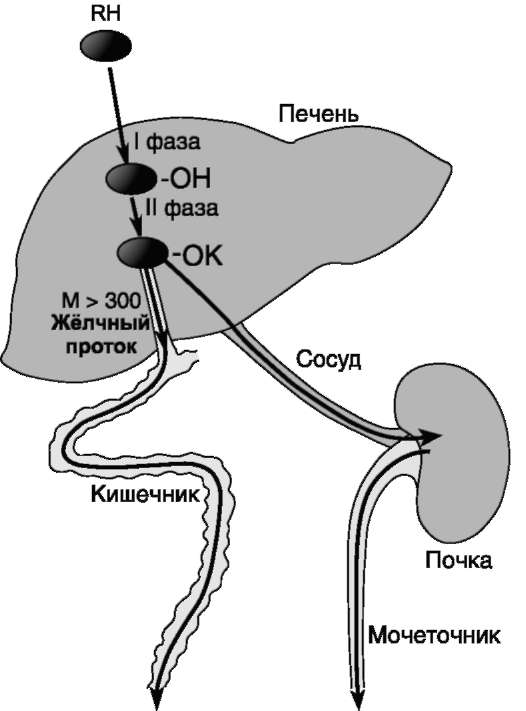
Рис. 12-1. Метаболизм и выведение ксенобиотиков из организма. RH - ксенобиотик; К - группа, используемая при конъюгации (глутатион, глюкуронил и др.); М - молекулярная масса. Из множества цитохром Р450-зависимых реакций на рисунке приведена только одна - схема гидроксилирования ксенобиотика. В ходе первой фазы в структуру вещества RH вводится полярная группа ОН-. Далее происходит реакция конъюгации; конъюгат в зависимости от растворимости и молекулярной массы удаляется либо почками, либо с фекалиями.
Основные функции печени
Обмен углеводов
Глюконеогенез
Синтез и распад гликогена
Обмен липидов и их производных
Синтез жирных кислот и жиров из углеводов Синтез и выведение холестерина Формирование липопротеинов Кетогенез
Синтез жёлчных кислот 25-гидроксилирование витамина D3
Обмен белков
Синтез белков плазмы крови (включая некоторые факторы свёртывания крови) Синтез мочевины (обезвреживание аммиака)
Обмен гормонов Метаболизм и выделение стероидных гормонов Метаболизм полипептидных гормонов
Метаболизм и экскреция билирубина Депонирование
гликогена витамина А витамина В12 железа
Лекарства и чужеродные вещества
Метаболизм и экскреция
Таблица 12-1. Возможные модификации ксенобиотиков в первой фазе обезвреживания
Вторая фаза - реакции конъюгации, в результате которых чужеродное вещество, модифицированное ферментными системами ЭР, связывается с эндогенными субстратами - глюкуроновой кислотой, серной кислотой, глицином, глутатионом. Образовавшийся конъюгат удаляется из организма.
А. МИКРОСОМАЛЬНОЕ ОКИСЛЕНИЕ
Микросомальные оксидазы - ферменты, локализованные в мембранах гладкого ЭР, функционирующие в комплексе с двумя внемитохон-дриальными ЦПЭ. Ферменты, катализирующие восстановление одного атома молекулы О2 с образованием воды и включение другого атома кислорода в окисляемое вещество, получили название микросомальных оксидаз со смешанной функцией или микросомальных монооксигеназ. Окисление с участием монооксигеназ обычно изучают, используя препараты микросом.
1. Основные ферменты микросомальных электронтранспортных цепей
Микросомальная система не содержит растворимых в цитозоле белковых компонентов, все ферменты - мембранные белки, активные центры которых локализованы на цитоплазма-тической поверхности ЭР. Система включает несколько белков, составляющих электронт-ранспортные цепи (ЦПЭ). В ЭР существуют две такие цепи, первая состоит из двух ферментов - NADPH-Р450 редуктазы и цитохрома Р450, вторая включает фермент NADH-цитохром-b5редукта-зу, цитохром b5 и ещё один фермент - стеароил-КоА-десатуразу.
Электронтранспортная цепь - NADPH-Р450 редуктаза - цитохром Р450. В большинстве случаев донором электронов (ē) для этой цепи служит NADPH, окисляемый NADPH-Р450 ре-дуктазой. Фермент в качестве простетической группы содержит 2 кофермента - флавинаде-ниндинуклеотид (FAD) и флавинмононуклеотид (FMN). Протоны и электроны с NADРH переходят последовательно на коферменты NADPH-Р450 редуктазы. Восстановленный FMN (FMNH2) окисляется цитохромом Р450 (см. схему ниже).
Цитохром Р450 - гемопротеин, содержит про-стетическую группу гем и имеет участки связывания для кислорода и субстрата (ксенобиотика). Название цитохром Р450 указывает на то, что максимум поглощения комплекса цитохрома Р450 лежит в области 450 нм.
Окисляемый субстрат (донор электронов) для NADH-цитохром Ь5-редуктазы - NADH (см. схему ниже). Протоны и электроны с NADH переходят на кофермент редуктазы FAD, следующим акцептором электронов служит Fe3+ цитохрома b5. Цитохром b5 в некоторых случаях может быть донором электронов (ē) для ци-тохрома Р450 или для стеароил-КоА-десатуразы, которая катализирует образование двойных связей в жирных кислотах, перенося электроны на кислород с образованием воды (рис. 12-2).
NADH-цитохром b5 редуктаза - двухдоменный белок. Глобулярный цитозольный домен связывает простетическую группу - кофермент FAD, а единственный гидрофобный «хвост» закрепляет белок в мембране.
Цитохром b5 - гемсодержащий белок, который имеет домен, локализованный на поверхности мембраны ЭР, и короткий «заяко-
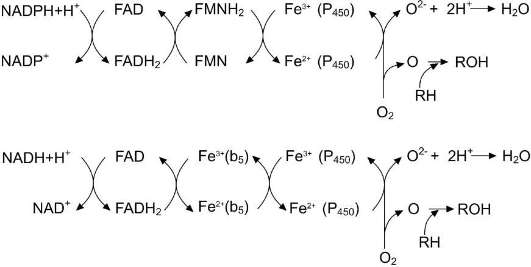
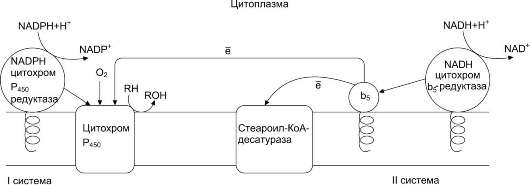
Рис. 12-2. Электронтранспортные цепи ЭР. RH - субстрат цитохрома Р450; стрелками показаны реакции переноса электронов. В одной системе NADPH окисляется NADPH цитохром Р450-редуктазой, которая затем передаёт электроны на целое семейство цитохромов Р450. Вторая система включает в себя окисление NADH цитохром b5-редуктазой, электроны переходят на цитохром b5; восстановленную форму цитохрома b5 окисляет стеароил-КоА-десатураза, которая переносит электроны на О2.
ренный» в липидном бислое спирализованный домен.
NADH-цитохром b5-редуктаза и цитохром b5, являясь «заякоренными» белками, не фиксированы строго на определённых участках мембраны ЭР и поэтому могут менять свою локализацию.
2. Функционирование цитохрома Р450
Известно, что молекулярный кислород в трип-летном состоянии инертен и не способен взаимодействовать с органическими соединениями. Чтобы сделать кислород реакционно-способным, необходимо его превратить в синглетный, используя ферментные системы его восстановления. К числу таковых принадлежит монок-сигеназная система, содержащая цитохром Р450. Связывание в активном центре цитохрома Р450 липофильного вещества RH и молекулы кислорода повышает окислительную активность фермента. Один атом кислорода принимает 2 ē и переходит в форму О2-. Донором электронов служит NADРH, который окисляется NADРH-цитохром Р450 редуктазой. О2- взаимодействует с протонами: О2- + 2Н+ → Н2О, и образуется вода. Второй атом молекулы кислорода включается в субстрат RH, образуя гидроксиль-ную группу вещества R-OH (рис. 12-3).
Суммарное уравнение реакции гидроксилиро-вания вещества RH ферментами микросомаль-ного окисления:
RH + O2 + NADPH + H+ → ROH + H2O + NADP+.
Субстратами Р450 могут быть многие гидрофобные вещества как экзогенного (лекарственные препараты, ксенобиотики), так и эндогенного (стероиды, жирные кислоты и др.) происхождения.
Таким образом, в результате первой фазы обезвреживания с участием цитохрома Р450 происходит модификация веществ с образованием функциональных групп, повышающих растворимость гидрофобного соединения. В результате модификации возможна потеря молекулой её биологической активности или даже формирование более активного соединения, чем вещество, из которого оно образовалось.
3. Свойства системы микросомального окисления
Важнейшие свойства ферментов микросо-мального окисления: широкая субстратная специфичность, которая позволяет обезвреживать самые разнообразные по строению вещества, и регуляция активности по механизму индукции.
Широкая субстратная специфичность. Изоформы Р450
К настоящему времени описано около 150 генов цитохрома Р450, кодирующих различные изоформы фермента. Каждая из изоформ Р450
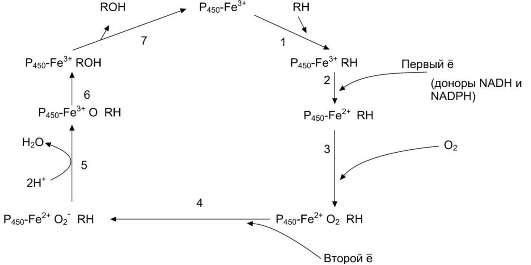
Рис. 12-3. Транспорт электронов при монооксигеназном окислении с участием Р450. Связывание (1) в активном центре цитохрома Р450 вещества RH активирует восстановление железа в геме - присоединяется первый электрон (2). Изменение валентности железа увеличивает сродство комплекса Р450-Fе2+ -RH к молекуле кислорода (3). Появление в центре связывания цитохрома Р450 молекулы О2 ускоряет присоединение второго электрона и образование комплекса Р450-Fе2+О2- -RH (4). H следующем этапе (5) Fе2+ окисляется, второй электрон присоединяется к молекуле кислорода Р450-Fе3+О22-. Восстановленный атом кислорода (О2-) связывает 2 протона, и образуется 1 молекула воды. Второй атом кислорода идёт на построение ОH-группы (6). Модифицированное вещество R-OH отделяется от фермента (7).
имеет много субстратов. Этими субстратами могут быть как эндогенные липофильные вещества, модификация которых входит в путь нормального метаболизма этих соединений, так и гидрофобные ксенобиотики, в том числе лекарства. Определённые изоформы цитохрома Р450 участвуют в метаболизме низкомолекулярных соединений, таких как этанол и ацетон.
Регуляция активности микросомальной системы окисления
Регуляция активности микросомальной системы осуществляется на уровне транскрипции или посттранскрипционных изменений. Индукция синтеза позволяет увеличить количество ферментов в ответ на поступление или образование в организме веществ, выведение которых невозможно без участия системы микросомального окисления.
В настоящее время описано более 250 химических соединений, вызывающих индукцию микросомальных ферментов. К числу этих индукторов относят барбитураты, полицикли-
ческие ароматические углеводороды, спирты, кетоны и некоторые стероиды. Несмотря на разнообразие химического строения, все индукторы имеют ряд общих признаков; их относят к числу липофильных соединений, и они служат субстратами для цитохрома Р450.
Б. КОНЪЮГАЦИЯ - ВТОРАЯ ФАЗА ОБЕЗВРЕЖИВАНИЯ ВЕЩЕСТВ
Вторая фаза обезвреживания веществ - реакции конъюгации, в ходе которых происходит присоединение к функциональным группам, образующимся на первом этапе, других молекул или групп эндогенного происхождения, увеличивающих гидрофильность и уменьшающих токсичность ксенобиотиков (табл. 12-2).
1. Участие трансфераз в реакциях конъюгации
Все ферменты, функционирующие во второй фазе обезвреживания ксенобиотиков, относят к классу трансфераз. Они характеризуются широкой субстратной специфичностью.
Таблица 12-2. Основные ферменты и метаболиты, участвующие в конъюгации

УДФ-глюкуронилтрансферазы
Локализированные в основном в ЭР ури-диндифосфат (УДФ)-глюкуронилтрансферазы присоединяют остаток глюкуроновой кислоты к молекуле вещества, образованного в ходе мик-росомального окисления (рис. 12-4).
В общем виде реакция с участием УДФ-глю-куронилтрансферазы записывается так:
RОH + УДФ-C6H9O6 = RO-C6H9O6 + УДФ. Сульфотрансферазы
Цитоплазматические сульфотрансферазы катализируют реакцию конъюгации, в ходе которой остаток серной кислоты (-SО3Н) от 3'-фосфоаденозин-5'-фосфосульфата (ФАФС) присоединяется к фенолам, спиртам или аминокислотам (рис. 12-5).
Реакция с участием сульфотрансферазы в общем виде записывается так:
RОH + ФАФ-SO3H = RO-SO3H + ФАФ.
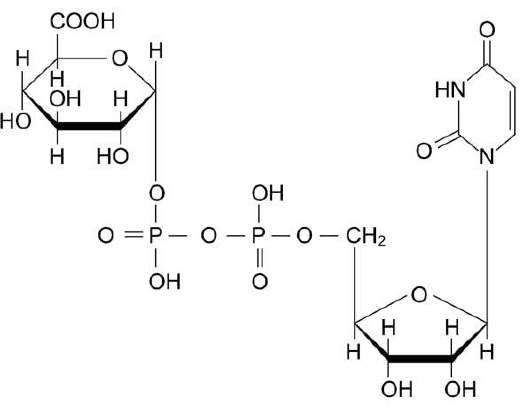
Рис. 12-4. Уридиндифосфоглюкуроновая кислота (УДФ-C6H9O6).
Ферменты сульфотрансферазы и УДФ-глюку-ронилтрансферазы участвуют в обезвреживании ксенобиотиков, инактивации лекарств и эндогенных биологически активных соединений.
Глутатионтрансферазы
Особое место среди ферментов, участвующих в обезвреживании ксенобиотиков, инактивации нормальных метаболитов, лекарств, занимают глутатионтрансферазы (ГТ). Глутатионтранс-феразы функционируют во всех тканях и играют важную роль в инактивации собственных метаболитов: некоторых стероидных гормонов, простагландинов, билирубина, жёлчных кислот, продуктов ПОЛ.
Известно множество изоформ ГТ с различной субстратной специфичностью. В клетке ГТ в основном локализованы в цитозоле, но имеются варианты ферментов в ядре и митохондриях. Для работы ГТ требуется глутатион (GSH) (рис. 12-6).
Глутатион - трипептид Глу-Цис-Гли (остаток глутаминовой кислоты присоединён к цистеину карбоксильной группой радикала).
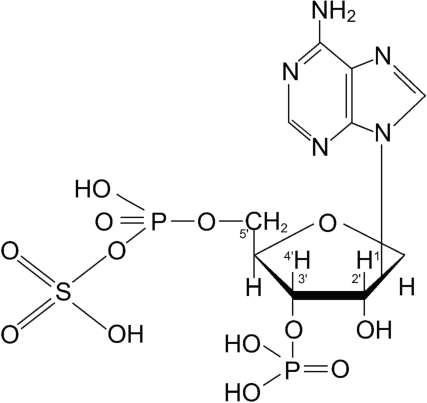
Рис. 12-5. 3'-Фосфоаденозин-5'-фосфосульфат (ФАФ-SО3Н).
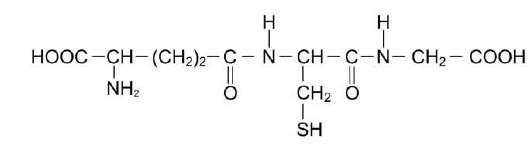
Рис. 12-6. Глутатион (GSH).
ГТ обладают широкой специфичностью к субстратам, общее количество которых превышает 3000. ГТ связывают очень многие гидрофобные вещества и инактивируют их, но химической модификации с участием глутатиона подвергаются только те, которые имеют полярную группу. То есть субстратами служат вещества, которые, с одной стороны, имеют электрофильный центр (например, ОН-группу), а с другой стороны - гидрофобные зоны. Обезвреживание, т.е. химическая модификация ксенобиотиков с участием ГТ, может осуществляться тремя различными способами:
• путём конъюгации субстрата R с глутатио-ном (GSH):
R + GSH → GSRH
• в результате нуклеофильного замещения:
RX + GSH → GSR + НХ,
• восстановления органических пероксидов до спиртов:
R-HC-O-OH + 2 GSH → R-HC-O-OH + GSSG + Н2О.
В реакции: ООН - гидропероксидная группа, GSSG - окисленный глутатион.
Система обезвреживания с участием ГТ и глутатиона играет уникальную роль в формировании резистентности организма к самым различным воздействиям и является наиболее важным защитным механизмом клетки. В ходе биотрансформации некоторых ксенобиотиков под действием ГТ образуются тиоэфиры (конъ-югаты RSG), которые затем превращаются в меркаптаны, среди которых обнаружены токсические продукты. Но конъюгаты GSH с большинством ксенобиотиков менее реакционно-способны и более гидрофильны, чем исходные вещества, а поэтому менее токсичны и легче выводятся из организма (рис. 12-7).
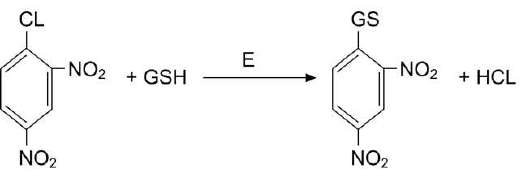
Рис. 12-7. Обезвреживание 1-хлор, 2,4-динитробен-зола с участием глутатиона.
ГТ своими гидрофобными центрами могут нековалентно связывать огромное количество липофильных соединений (физическое обезвреживание), предотвращая их внедрение в липид-ный слой мембран и нарушение функций клетки. Поэтому ГТ иногда называют внутриклеточным альбумином.
ГТ могут ковалентно связывать ксенобиотики, являющиеся сильными электролитами. Присоединение таких веществ - «самоубийство» для ГТ, но дополнительный защитный механизм для клетки.
Ацетилтрансферазы, метилтрансферазы
Ацетилтрансферазы катализируют реакции конъюгации - переноса ацетильного остатка от ацетил-КоА на азот группы -SO2NH2, например в составе сульфаниламидов. Мембранные и цитоплазматические метилтрансферазы с участием SAM метилируют группы -Р=О, -NH2 и SH-группы ксенобиотиков.
2. Роль эпоксидгидролаз в образовании диолов
Во второй фазе обезвреживания (реакции конъюгации) принимают участие и некоторые другие ферменты. Эпоксидгидролаза (эпоксид-гидратаза) присоединяет воду к эпоксидам бензола, бензпирена и другим полициклическим углеводородам, образованным в ходе первой фазы обезвреживания, и превращает их в дио-лы (рис. 12-8). Эпоксиды, образовавшиеся при микросомальном окислении, являются канцерогенами. Они обладают высокой химической активностью и могут участвовать в реакциях неферментативного алкилирования ДНК, РНК, белков (см. раздел 16). Химические модификации этих молекул могут привести к перерождению нормальной клетки в опухолевую.
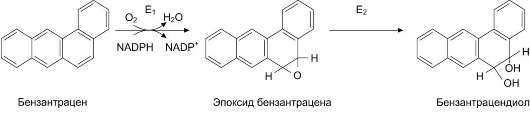
Рис. 12-8. Обезвреживание бензантрацена. Е1 - фермент микросомальной системы; Е2 - эпоксидгидратаза.
В. ГНИЕНИЕ АМИНОКИСЛОТ В КИШЕЧНИКЕ. ОБЕЗВРЕЖИВАНИЕ И ВЫВЕДЕНИЕ ПРОДУКТОВ ГНИЕНИЯ ИЗ ОРГАНИЗМА
Аминокислоты, невсосавшиеся в клетки кишечника, используются микрофлорой толстой кишки в качестве питательных веществ. Ферменты бактерий расщепляют аминокислоты и превращают их в амины, фенолы, индол, скатол, сероводород и другие ядовитые для организма соединения. Этот процесс иногда называют гниением белков в кишечнике. В основе гниения лежат реакции декарбоксилирования и дезаминирования аминокислот.
Образование и обезвреживание n-крезола и фенола
Под действием ферментов бактерий из аминокислоты тирозина могут образовываться фенол и крезол путём разрушения боковых цепей аминокислот микробами (рис. 12-9).
Всосавшиеся продукты по воротной вене поступают в печень, где обезвреживание фенола и крезола может происходить путём конъюгации с сернокислотным остатком (ФАФС) или с глюку-роновой кислотой в составе УДФ-глюкуроната. Реакции конъюгации фенола и крезола с ФАФС
катализирует фермент сульфотрансфераза (рис. 12-10).
Конъюгация глюкуроновых кислот с фенолом и крезолом происходит при участии фермента УДФ-глюкуронилтрансферазы (рис. 12-11). Продукты конъюгации хорошо растворимы в воде и выводятся с мочой через почки. Повышение количества конъюгатов глюкуроновой кислоты с фенолом и крезолом обнаруживают в моче при увеличении продуктов гниения белков в кишечнике.
Образование и обезвреживание индола и скатола
В кишечнике из аминокислоты триптофана микроорганизмы образуют индол и скатол. Бактерии разрушают боковую цепь триптофана, оставляя нетронутой кольцевую структуру.
Индол образуется в результате отщепления бактериями боковой цепи, возможно, в виде серина или аланина (рис. 12-12).
Скатол и индол обезвреживаются в печени в 2 этапа. Сначала в результате микросомального окисления они приобретают гидроксильную группу. Так, индол переходит в индоксил, а затем вступает в реакцию конъюгации с ФАФС, образуя индоксилсерную кислоту, калиевая соль
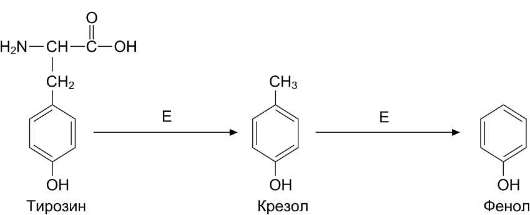
Рис. 12-9. Катаболизм тирозина под действием бактерий. Е - бактериальные ферменты.
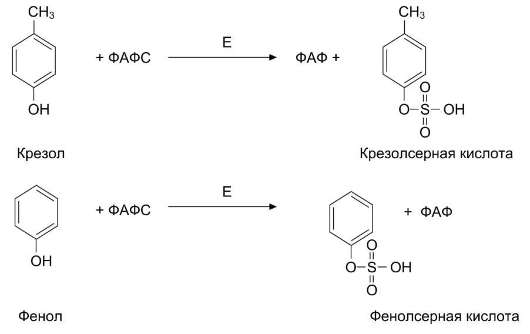
Рис. 12-10. Конъюгация фенола и крезола с ФАФС. Е - сульфотрансфераза.
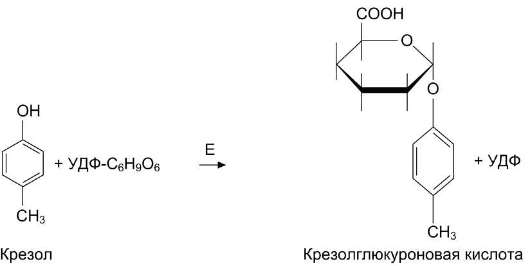
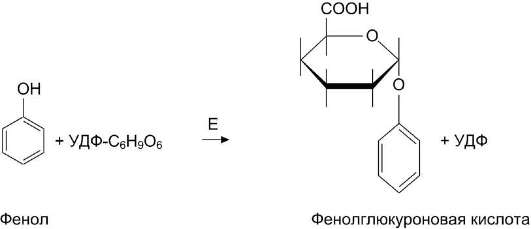
Рис. 12-11. Участие УДФ-глюкуронилтрансферазы в обезвреживании крезола и фенола. Е - УДФ-глюку-ронилтрансфераза.
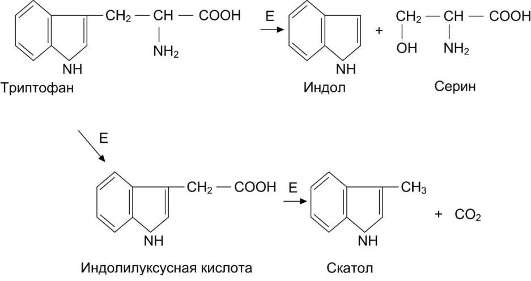
Рис. 12-12. Катаболизм триптофана под действием бактерий. Е - бактериальные ферменты.
которой получила название животного индикана
(рис. 12-13).
Обезвреживание бензойной кислоты
Синтез гиппуровой кислоты из бензойной кислоты и глицина протекает у человека и большинства животных преимущественно в печени (рис. 12-14). Скорость этой реакции отражает функциональное состояние печени.
В клинической практике используют определение скорости образования и выведения гиппуровой кислоты после введения в организм ксенобиотика бензойной кислоты (бензойно-кислого натрия) - проба Квика.
Г. СВЯЗЫВАНИЕ, ТРАНСПОРТ И ВЫВЕДЕНИЕ
КСЕНОБИОТИКОВ
В плазме крови множество как эндогенных, так и экзогенных липофильных веществ транспортируются альбумином и другими белками.
Альбумин - основной белок плазмы крови, связывающий различные гидрофобные вещества. Он может функционировать в качестве белка-переносчика билирубина, ксенобиотиков, лекарственных веществ.
Помимо альбуминов, ксенобиотики могут транспортироваться по крови в составе липопро-теинов, а также в комплексе с кислым α1-глико-протеином. Особенность этого гликопротеина
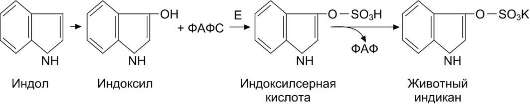
Рис. 12-13. Участие сульфотрансферазы в обезвреживании индола. Е - сульфотрансфераза.
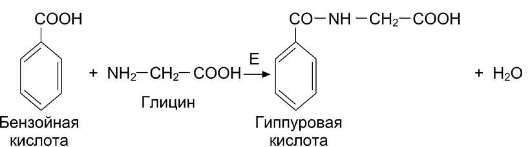
Рис. 12-14. Образование гиппуровой кислоты из бензойной кислоты и глицина. Е - глицинтрансфераза.
состоит в том, что он является индуцируемым белком, участвующим в ответной реакции организма на изменения, происходящие в состоянии стресса, например, при инфаркте миокарда, воспалительных процессах; его количество в плазме увеличивается наряду с другими протеинами. Связывая ксенобиотики, кислый α1-гликопро-теин инактивирует их и переносит в печень, где комплекс с белком распадается, и чужеродные вещества обезвреживаются и выводятся из организма.
Участие Р-гликопротеина в выведении ксенобиотиков
Очень важный механизм выведения из клетки гидрофобных ксенобиотиков - функционирование Р-гликопротеина (транспортная АТФ-аза). Р-гликопротеин - фосфогликопротеин с молекулярной массой 170 кД, присутствующий в плазматической мембране клеток многих тканей, в частности почек и кишечника. Полипептидная цепь этого белка содержит 1280 аминокислотных остатков, образуя 12 трансмембранных доменов и два АТФ-связывающих центра (рис. 12-15).
В норме его функция состоит в экскреции ионов хлора и гидрофобных токсичных соединений из клеток.
Когда гидрофобное вещество (например, противоопухолевое лекарство) проникает в клетку, то оно удаляется из неё Р-гликопротеином с затратой энергии (рис. 12-16). Уменьшение количества лекарства в клетке снижает эффективность его применения при химиотерапии онкологических заболеваний.
Д. ИНДУКЦИЯ ЗАЩИТНЫХ СИСТЕМ
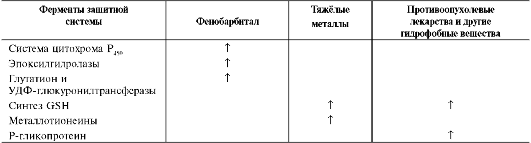
Многие ферменты, участвующие в первой и второй фазе обезвреживания, - индуцируемые белки. Ещё в древности царь Митридат знал, что если систематически принимать небольшие дозы яда, можно избежать острого отравления. «Эффект Митридата» основан на индукции определённых защитных систем (табл. 12-3).
В мембранах ЭР печени цитохрома Р450 содержится больше (20%), чем других мембрано-связанных ферментов. Лекарственное вещество фенобарбитал активирует синтез цитохрома
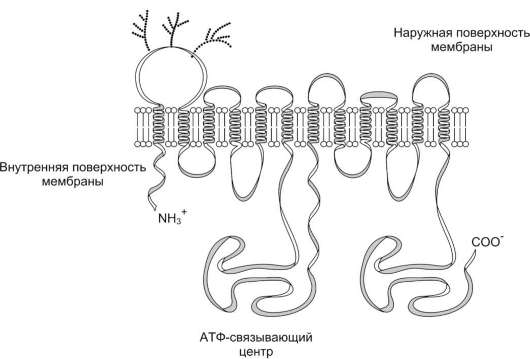
Рис. 12-15. Строение Р-гликопротеина. Р-гликопротеин - интегральный белок, имеющий 12 трансмембранных доменов, пронизывающих бислой цитоплазматической мембраны. N- и С-концы белка обращены в цитозоль. Участки Р-гликопротеина на наружной поверхности мембраны гликозилированы. Область между шестым и седьмым доменами имеет центры для присоединения АТФ и аутофосфорилирования.
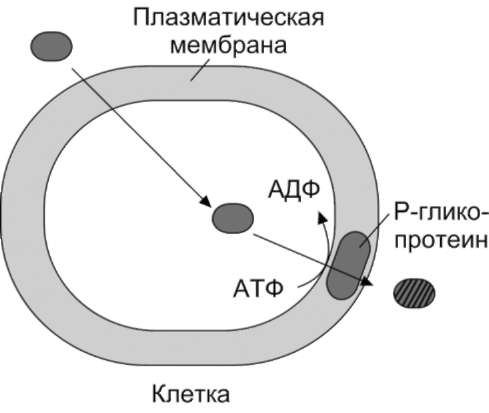
Рис. 12-16. Функционирование Р-гликопротеина.
Заштрихованный овал - противоопухолевое лекарство (гидрофобное вещество).
Р450, УДФ-глюкуронилтрансферазы и эпоксид гидролазы. Например, у животных, которым вводили индуктор фенобарбитал, увеличивается площадь мембран ЭР, которая достигает 90% всех мембранных структур клетки, и, как следствие, - увеличение количества ферментов, участвующих в обезвреживании ксенобиотиков или токсических веществ эндогенного происхождения.
При химиотерапии злокачественных процессов начальная эффективность лекарства часто постепенно падает. Более того, развивается множественная лекарственная устойчивость, т.е. устойчивость не только к этому лечебному препарату, но и целому ряду других лекарств. Это происходит потому, что противоопухолевые лекарства индуцируют синтез Р-глико-протеина, глутатионтрансферазы и глутатиона. Использование веществ, ингибирующих или активирующих синтез Р-гликопротеина, а также
ферменты синтеза глутатиона, повышает эффективность химиотерапии.
Металлы являются индукторами синтеза глутатиона и низкомолекулярного белка метал-лотионеина, имеющих SH-групггы, способные связывать их. В результате возрастает устойчивость клеток организма к ядам и лекарствам.
Повышение количества глутатионтрансфераз увеличивает способность организма приспосабливаться к возрастающему загрязнению внешней среды. Индукцией фермента объясняют отсутствие антиканцерогенного эффекта при применении ряда лекарственных веществ. Кроме того, индукторы синтеза глутатионтрансферазы - нормальные метаболиты - половые гормоны, йодтиронины и кортизол. Катехоламины через аденилатциклазную систему фосфорилируют глу-татионтрансферазу и повышают её активность.
Ряд веществ, в том числе и лекарств (например, тяжёлые металлы, полифенолы, S-алкилы глутатиона, некоторые гербициды), ингибируют глутатионтрансферазу.
ii. биотрансформация лекарственных веществ
Лекарства, поступившие в организм, проходят следующие превращения:
• всасывание;
• связывание с белками и транспорт кровью;
• взаимодействие с рецепторами;
• распределение в тканях;
• метаболизм и выведение из организма.
Механизм первого этапа (всасывание) определяется физико-химическими свойствами лекарства. Гидрофобные соединения легко проникают через мембраны простой диффузией, в то время
Таблица 12-3. Индукция систем, обеспечивающих защиту от ксенобиотиков
как лекарственные вещества, нерастворимые в липидах, проникают через мембраны путём трансмембранного переноса при участии разных типов транслоказ. Некоторые нерастворимые крупные частицы могут проникать в лимфатическую систему путём пиноцитоза.
Следующие этапы метаболизма лекарственного вещества в организме тоже определяются его химическим строением - гидрофобные молекулы перемещаются по крови в комплексе с альбумином, кислым α1-гликопротеином или в составе липопротеинов. В зависимости от структуры лекарственное вещество может поступать из крови в клетку или, являясь аналогами эндогенных веществ, связываться рецепторами клеточной мембраны.
Действие на организм большинства лекарств прекращается через определённое время после их приёма. Прекращение действия может происходить потому, что лекарство выводится из организма либо в неизменённом виде - это характерно для гидрофильных соединений, либо в виде продуктов его химической модификации (биотрансформации).
А. ХАРАКТЕР ИЗМЕНЕНИЙ ПРИ БИОТРАНСФОРМАЦИИ ЛЕКАРСТВЕННЫХ ВЕЩЕСТВ
Биохимические превращения лекарственных веществ в организме человека, обеспечивающие их инактивацию и детоксикацию, являются частным проявлением биотрансформации чужеродных соединений.
В результате биотрансформации лекарственных веществ может произойти:
• инактивация лекарственных веществ, т.е. снижение их фармакологической активности;
• повышение активности лекарственных веществ;
• образование токсических метаболитов.
Инактивация лекарственных веществ
Инактивация лекарственных веществ, как и всех ксенобиотиков, происходит в 2 фазы. Первая фаза - химическая модификация под действием ферментов монооксигеназной системы ЭР. Например, лекарственное вещество барбитурат в ходе биотрансформации превращается в гидроксибарбитурат, который далее участвует в реакции конъюгации с остатком глюкуроновой кислоты. Фермент глюкуронилтрансфераза катализирует образование барбитуратглюкуронида, в качестве источника глюкуроновой кислоты используется УДФ-глюкуронил (рис. 12-17).
В первую фазу обезвреживания под действием монооксигеназ образуются реакционно-способные группы -ОН, -СООН, -NH2, -SH и др. Химические соединения, уже имеющие эти группы, сразу вступают во вторую фазу обезвреживания - реакции конъюгации.
Повышение активности лекарств
В качестве примера повышения активности вещества в процессе его превращений в организме можно привести образование дезметилими-прамина из имипрамина. Дезметилимипрамин обладает выраженной способностью ослаблять депрессивное состояние при психических расстройствах (рис. 12-18).
Химические превращения некоторых лекарств в организме приводят к изменению характера их активности. Например, ипразид - антидепрессант, который в результате дезалкилирования превращается в изониазид, обладающий противотуберкулёзным действием (рис. 12-19).
Образование токсических продуктов в результате реакции биотрансформации. В отдельных случаях химические превращения лекарственных средств в организме могут приводить к появлению у них токсических свойств. Так,
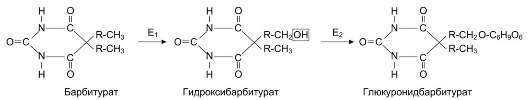
Рис. 12-17. Метаболизм барбитуратов в печени. Е1 - ферменты микросомального окисления; Е2 - глюку-ронилтрансфераза.
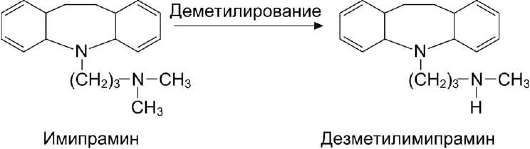
Рис. 12-18. Активация имипрамина в результате реакции деметилирования.

Рис. 12-19. Образование изониазида в ходе дезалкилирования ипраниазида.
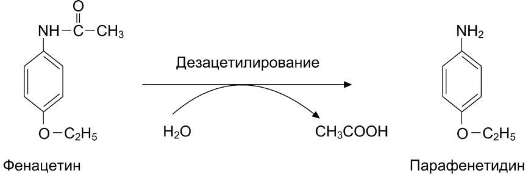
Рис. 12-20. Превращение фенацетина в токсический продукт - парафенетидин.
жаропонижающее, болеутоляющее, противовоспалительное средство фенацетин превращается в парафенетидин, вызывающий гипоксию за счёт образования метгемоглобина - неактивной формы Нb (рис. 12-20).
Реакции конъюгации лекарственных веществ
Вторая фаза инактивации - конъюгация (связывание) лекарственных веществ, как подвергшихся каким-либо превращениям на первом этапе, так и нативных препаратов. К продуктам, образованным ферментами микросомального окисления, может присоединяться глицин по карбоксильной группе, глюкуроновая кислота или остаток серной кислоты - по ОН-группе, ацетильный остаток - к NH2-группе.
В превращениях второй фазы инактивации лекарственных веществ принимают участие эндогенные соединения, образующиеся в организме с затратой энергии SAM: (АТФ), УДФ-
глюкуронат (УТФ), Ацетил-КоА (АТФ) и др. Поэтому можно сказать, что реакции конъюгации сопряжены с использованием энергии этих макроэргических соединений.
Примером реакции конъюгации может служить глюкуронирование гидроксибарбитурата под действием глюкуронилтрансферазы, описанным ранее (см. рис. 12-17). В качестве примера О-метилирования лекарства можно привести один из этапов биотрансформации препарата метилдофа, нарушающего образование адренер-гического медиатора и применяемого в качестве гипотензивного средства (рис. 12-21).
В неизменённом виде выделяются главным образом высокогидрофильные соединения. Из липофильных веществ исключение составляют средства для ингаляционного наркоза, основная часть которых в химические реакции в организме не вступает. Они выводятся лёгкими в том же виде, в каком были введены.
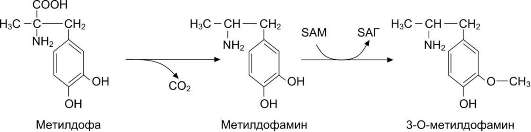
Рис. 12-21. Биотрансформация лекарственного вещества (метилдофа).
Б. ФАКТОРЫ, ВЛИЯЮЩИЕ НА АКТИВНОСТЬ
ФЕРМЕНТОВ БИОТРАНСФОРМАЦИИ ЛЕКАРСТВ
Лекарственные средства в результате химической модификации, как правило, теряют свою биологическую активность. Таким образом, эти реакции лимитируют во времени действие лекарств. При патологии печени, сопровождающейся снижением активности микросомальных ферментов, продолжительность действия ряда лекарственных веществ увеличивается.
Некоторые препараты снижают активность монооксигеназной системы. Например, левоми-цетин и бутадион ингибируют ферменты мик-росомального окисления. Антихолинэстеразные средства, ингибиторы моноаминооксидазы, нарушают функционирование фазы конъюгации, поэтому они пролонгируют эффекты препаратов, которые инактивируются этими ферментами. Кроме того, скорость каждой из реакций биотрансформации лекарственного вещества зависит от генетических, физиологических факторов и экологического состояния окружающей среды.
Возрастные особенности
Чувствительность к лекарственным средствам меняется в зависимости от возраста. Например, у новорождённых активность метаболизма лекарств в первый месяц жизни существенно отличается от взрослых. Это связано с недостаточностью многих ферментов, участвующих в биотрасформации лекарственных веществ, функции почек, повышенной проницаемостью гематоэнцефалического барьера, недоразвитием ЦНС. Так, новорождённые более чувствительны к некоторым веществам, влияющим на ЦНС (в частности, к морфину). Очень токсичен для них левомицетин; это объясняется тем, что в печени
у новорождённых малоактивны ферменты, необходимые для его биотрансформации.
В пожилом возрасте метаболизм лекарственных веществ протекает менее эффективно: снижается функциональная активность печени, нарушается скорость экскреции препаратов почками. В целом чувствительность к большинству лекарственных средств в пожилом возрасте повышена, в связи с чем их доза должна быть снижена.
Генетические факторы
Индивидуальные различия в метаболизме ряда препаратов и в реакциях на препараты объясняют генетическим полиморфизмом, т.е. существованием в популяции изоформ некоторых ферментов биотрансформации.
В ряде случаев повышенная чувствительность к лекарственным средствам может быть обусловлена наследственной недостаточностью некоторых ферментов, участвующих в химической модификации. Например, при генетической недостаточности холинэстеразы плазмы крови длительность действия миорелаксанта дитилина резко возрастает и может достигать 6-8 ч и более (в обычных условиях дитилин действует в течение 5-7 мин). Известно, что скорость ацетилирования противотуберкулёзного средства изониазида варьирует довольно широко. Выделяют лиц с быстрой и медленной метаболизирующей активностью. Считают, что у лиц с медленной инактивацией изониазида нарушена структура белков, регулирующих синтез фермента ацетилтрансферазы, обеспечивающего конъюгацию изониазида с ацетильным остатком.
Факторы окружающей среды
Существенное влияние на метаболизм лекарственных веществ в организме оказывают
также факторы окружающей среды, такие как ионизирующая радиация, температура, состав пищи и особенно различные химические вещества (ксенобиотики), в том числе и сами лекарственные вещества.
III. МЕТАБОЛИЗМ ЭТАНОЛА В ПЕЧЕНИ
Катаболизм этилового спирта осуществляется главным образом в печени. Здесь окисляется от 75% до 98% введённого в организм этанола.
Окисление алкоголя - сложный биохимический процесс, в который вовлекаются основные метаболические процессы клетки. Превращение этанола в печени осуществляется тремя путями с образованием токсического метаболита - ацет-альдегида (рис. 12-22).
А. ОКИСЛЕНИЕ ЭТАНОЛА NAD-ЗАВИСИМОЙ АЛКОГОЛЬДЕГИДРОГЕНАЗОЙ
Основную роль в метаболизме этанола играет цинксодержащий NAD+-зависимый фермент - алкогольдегидрогеназа, локализующаяся в основном в цитозоле и митохондриях печени (95%). В ходе реакции пpoиcxoдит дегидрирование этанола, образуются ацеталь-дегид и восстановленный кофермент NADH. Алкогольдегидрогеназа катализирует обратимую реакцию, направление которой зависит от концентрации ацетальдегида и соотношения NADH/NAD+ в клетке.
С9H5ОН + NAD+ ↔ CH3CHO + NADH + H+.
Фермент алкогольдегидрогеназа - димер, состоящий из идентичных или близких по первичной структуре полипептидных цепей, кодируемых аллелями одного гена. Существуют 3 изоформы алкогольдегидрогеназы (АДГ): АДГ1, АДГ2, АДГ3, различающиеся по строению протомеров, локализации и активности. Для европейцев характерно присутствие изоформ АДГ1 и АДГ3. У некоторых восточных народов преобладает изоформа АДГ2, характеризующаяся высокой активностью, это может быть причиной их повышенной чувствительности к алкоголю. При хроническом алкоголизме количество фермента в печени не увеличивается, т.е. он не является индуцируемым ферментом.
Б. ОКИСЛЕНИЕ ЭТАНОЛА ПРИ УЧАСТИИ ЦИТОХРОМ Р450-ЗАВИСИМОЙ МИКРОСОМАЛЬ-НОЙ ЭТАНОЛОКИСЛЯЮЩЕЙ СИСТЕМЫ
Цитохром Р450-зависимая микросомальная эта-нолокисляющая система (МЭОС) локализована в мембране гладкого ЭР гепатоцитов. МЭОС играет незначительную роль в метаболизме небольших количеств алкоголя, но индуцируется этанолом, другими спиртами, лекарствами типа барбитуратов и приобретает существенное значение при злоупотреблении этими веществами. Этот путь окисления этанола происходит при участии одной из изоформ Р450 - изофермента Р450 II Е1. При хроническом алкоголизме окисление этанола ускоряется на 50-70% за счёт гипертрофии ЭР и индукции ци-тохрома Р450 II Е1.
C9H5OH + NADPH + Н+ + О2 → CH3CHO + NADP+ + 2 Н2О.
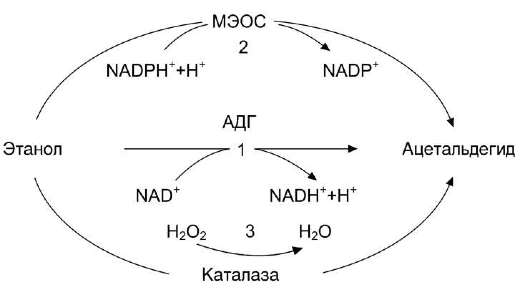
Рис. 12-22. Метаболизм этанола. 1 - окисление этанола NAD+-зависимой алкогольдегидрогеназой (АДГ); 9 - МЭОС - микросомальная этанолокисляющая система; 3 - окисление этанола каталазой.
Кроме основной реакции, цитохром Р450 катализирует образование активных форм кислорода (О2-, Н2О2), которые стимулируют ПОЛ в печени и других органах (см. раздел 8).
в. окисление этанола каталазой
Второстепенную роль в окислении этанола играет каталаза, находящаяся в пероксисомах цитоплазмы и митохондрий клеток печени. Этот фермент расщепляет примерно 2% этанола, но при этом утилизирует пероксид водорода.
СН3СН2ОН + Н2О2 → СН3СНО +2 Н2О.
г. метаболизм и токсичность ацетальдегида
Ацетальдегид, образовавшийся из этанола, окисляется до уксусной кислоты двумя ферментами: FAD-зависимой альдегидоксидазой и NАD+-зависимой ацетальдегиддегидрогеназой (АлДГ).
CH3CHO + О2 + Н2О → CH3COOH +Н2О2.
Повышение концентрации ацетальдегида в клетке вызывает индукцию фермента альдегид-оксидазы. В ходе реакции образуются уксусная кислота, пероксид водорода и другие активные формы кислорода, что приводит к активации
ПОЛ.
Другой фермент ацетальдегиддегидрогеназа (АлДГ) окисляет субстрат при участии кофер-мента NАD+.
CH3CHO + Н2О + NAD+ → CH3COOH + + NADH + H+.
Полученная в ходе реакции уксусная кислота активируется под действием фермента ацетил-КоА-синтетазы. Реакция протекает с использованием кофермента А и молекулы АТФ. Образовавшийся ацетил-КоА, в зависимости от соотношения АТФ/АДФ и концентрации окса-лоацетата в митохондриях гепатоцитов, может «сгорать» в ЦТК, идти на синтез жирных кислот или кетоновых тел.
В разных тканях организма человека встречаются полиморфные варианты АлДГ. Они характеризуются широкой субстратной специфичностью, разным распределением по клеткам тканей (почки, эпителий, слизистая оболочка
желудка и кишечника) и в компартментах клетки. Например, изоформа АлДГ, локализованная в митохондриях гепатоцитов, обладает более высоким сродством к ацетальдегиду, чем цито-зольная форма фермента.
Ферменты, участвующие в окислении этанола, - алкогольдегидрогеназа и АлДГ по разному распределены: в цитозоле - 80%/20% и митохондриях - 20%/80%. При поступлении больших доз алкоголя (более 2 г/кг) из-за разных скоростей окисления этанола и ацетальдегида в цитозоле резко повышается концентрация последнего. Ацетальдегид - очень реакционно-способное соединение; он неферментативно может ацетилировать SH-, NН2-группы белков и других соединений в клетке и нарушать их функции. В модифицированных (ацетилиро-ванных) белках могут возникать «сшивки», нехарактерные для нативной структуры (например, в белках межклеточного матрикса - эластине и коллагене, некоторых белках хроматина и липопротеинов, формирующихся в печени). Ацетилирование ядерных, цитоплаз-матических ферментов и структурных белков приводит к снижению синтеза экспортируемых печенью в кровь белков, например альбумина, который, удерживая поддерживает коллоидно-осмотическое давление, а также участвует в транспорте многих гидрофобных веществ в крови (см. раздел 14). Нарушение функций альбумина в сочетании с повреждающим действием ацетальдегида на мембраны сопровождается поступлением в клетки по градиенту концентрации ионов натрия и воды, происходит осмотическое набухание этих клеток и нарушение их функций.
Активное окисление этанола и ацетальдегида приводит к увеличению отношения NADH/ NAD+, что снижает активность NAD+-зависи-мых ферментов в цитозоле и менее значительно в митохондриях.
Равновесие следующей реакции смещается вправо:
Дигидроксиацетонфосфат + NADH + H+ ↔ Глице-рол-3-фосфат + NAD+,
Пируват + NADH + H+ ↔ Лактат +NAD+.
Восстановление дигидроксиацетонфосфата, промежуточного метаболита гликолиза и глю-конеогенеза, приводит к снижению скорости
глюконеогенеза. Образование глицерол-3-фос-фата повышает вероятность синтеза жира в печени. Увеличение концентрации NADH по сравнению с NAD+ (NADH>NAD+) замедляет реакцию окисления лактата, увеличивается соотношение лактат/пируват и ещё больше снижается скорость глюконеогенеза (см. раздел 7). В крови возрастает концентрация лактата, это приводит к гиперлактацидемии и лактоацидозу
(рис. 12-23).
NADH окисляется ферментом дыхательной цепи NADH-дегидрогеназой. Возникновение трансмембранного электрического потенциала на внутренней митохондриальной мембране не приводит к синтезу АТФ в полном объёме. Этому препятствует нарушение структуры внутренней мембраны митохондрий, вызванное мембранотропным действием этилового спирта
и повреждающим действием ацетальдегида на мембраны.
Можно сказать, что ацетальдегид опосредованно активирует ПОЛ, так как связывая SH-группы глутатиона, он снижает количество активного (восстановленного) глутатиона в клетке, который необходим для функционирования фермента глутатионпероксидазы (см. раздел 8), участвующего в катаболизме H2О2. Hакопление свободных радикалов приводит к активации ПОЛ мембран и нарушению структуры липидного бислоя.
На начальных стадиях алкоголизма окисление ацетил-КоА в ЦТК - основной источник энергии для клетки. Избыток ацетил-КоА в составе цитрата выходит из митохондрий, и в цитоплазме начинается синтез жирных кислот. Этот процесс, помимо АТФ, требует участия NADPH,
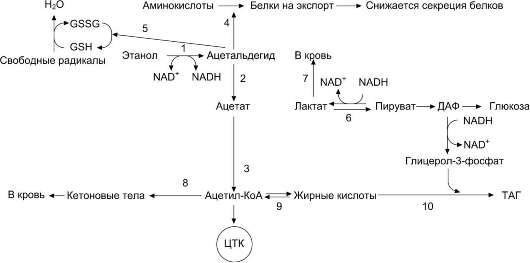
Рисунок 12-23. Эффекты этанола в печени. 1→2→3 - окисление этанола до ацетата и превращение его в ацетил-КоА
(1 - реакция катализируется алкогольдегидрогеназой, 2 - реакция катализируется АлДГ). Скорость образования ацетальдегида (1)часто при приёме большого количества алкоголя выше, чем скорость его окисления (9), поэтому ацетальальдегид накапливается и оказывает влияние на синтез белков (4), ингибируя его, а также понижает концентрацию восстановленного глутатиона (5), в результате чего активируется ПОЛ. Скорость глюконеогенеза (6) снижается, так как высокая концентрация NADH образованного в реакциях окисления этанола (1, 9), ингибирует глюконеогенез (6). Лактат выделяется в кровь (7), и развивается лактоацидоз. Увеличение концентрации NADH замедляет скорость ЦТК; ацетил-КоА накапливается, активируется синтез кетоновых тел (кетоз) (8). Окисление жирных кислот также замедляется (9), увеличивается синтез жира (10), что приводит к ожирению печени и гипертриацилглицеролемии.
который образуется при окислении глюкозы в пентозофосфатном цикле. Из жирных кислот и глицерол-3-фосфата образуются ТАГ, которые в составе ЛПОHП секретируются в кровь. Повышенная продукция ЛПОHП печенью приводит к гипертриацижлицеролемии. При хроническом алкоголизме снижение синтеза фосфолипидов и белков в печени, в том числе и апобелков, участвующих в формировании ЛПОHП, вызывает внутриклеточное накопление ТАГ и ожирение печени.
Однако в период острой алкогольной интоксикации, несмотря на наличие большого количества ацетил-КоА, недостаток оксало-ацетата снижает скорость образования цитрата. В этих условиях избыток ацетил-КоА идёт на синтез кетоновых тел, которые выходят в кровь. Повышение в крови концентрации лактата, ацетоуксусной кислоты и β-гидроксибутирата служит причиной метаболического ацидоза при алкогольной интоксикации.
Как уже было сказано ранее, реакция образования ацетальдегида из этанола протекает под действием алкогольдегидрогеназы. Поэтому при повышении концентрации ацетальдегида и NADH в клетках печени направление реакции меняется - образуется этанол. Этанол - мемб-ранотропное соединение, он растворяется в ли-пидном бислое мембран и нарушает их функции. Это негативно отражается на трансмембранном переносе веществ, межклеточных контактах, взаимодействиях рецепторов клетки с сигнальными молекулами. Этанол может проходить через мембраны в межклеточное пространство и кровь и далее в любую клетку организма.
д. влияние этанола и ацетальдегида на метаболизм ксенобиотиков и лекарств в печени
Характер влияния этанола на метаболизм ксенобиотиков и лекарств зависит от стадии алкогольной болезни: начальная стадия алкоголизма, хронический алкоголизм или острая форма алкогольной интоксикации.
Микросомальная этанолокисляющая система (МЭОС) наряду с метаболизмом этанола участвует в детоксикации ксенобиотиков и лекарств. На начальной стадии алкогольной болезни биотрансформация лекарственных веществ протекает более активно вследствие индукции ферментов системы. Этим объясняют феномен лекарственной «устойчивости». Однако при острой интоксикации этиловым спиртом тормозится биотрансформация лекарственных веществ. Этанол конкурирует с ксенобиотиками за связывание с цитохромом Р450 II Е1, вызывая гиперчувствительность (лекарственную «неустойчивость») к некоторым принятым одновременно с ним лекарственным препаратам.
Кроме того, у людей, страдающих хроническим алкоголизмом, наблюдают избирательную индукцию изоформы Р450 II Е1 и конкурентное ингибирование синтеза других изоформ, принимающих участие в метаболизме ксенобиотиков и лекарств. При злоупотреблении алкоголем индуцируется также синтез глюку-ронил-трансфераз, но снижается образование УДФ-глюкуроната.
Алкогольдегидрогеназа обладает широкой субстратной специфичностью и может окислять разные спирты, в том числе и метаболиты сердечных гликозидов - дигитоксина, дигоксина и гитоксина. Конкуренция этанола с сердечными гликозидами за активный центр алкогольде-гидрогеназы приводит к снижению скорости биотрансформации этой группы лекарств и повышает опасность их побочного эффекта у лиц, принимающих большие дозы алкоголя.
Повышение концентрации ацетальдегида вызывает целый ряд нарушений в структуре белков (ацетилирование), мембран (ПОЛ), модификацию глутатиона, необходимого для одного из самых важных ферментов обезвреживания ксенобиотиков - глутатионтрансферазы и фермента антиоксидазной защиты глутатионперок-сидазы. Таким образом, представленные данные свидетельствуют, что алкогольное поражение печени сопровождается нарушением важнейшей функции этого органа - детоксикационной.