Общая неврология А.С. Никифоров, Е.И. Гусев - 2007г. - 720 с
|
|
|
|
ГЛАВА 5 ЭКСТРАПИРАМИДНАЯ СИСТЕМА. АКИНЕТИКО-РИГИДНЫЙ СИНДРОМ
5.1. ПОНЯТИЕ ОБ ЭКСТРАПИРАМИДНОЙ СИСТЕМЕ
Движения обеспечиваются поперечнополосатыми мышцами. На их состояние влияют периферические мотонейроны, функция которых определяется суммарным воздействием на них многообразной импульсации. Длительное время при изучении движений сначала признавалось влияние на них, главным образом, больших пирамидных клеток (клетки Беца), входящих в состав V слоя двигательной зоны коры передней центральной извилины (главным образом поля 4, по Бродману). Считалось, что связи между центральными (корковы- ми) и периферическими двигательными нейронами, которые сейчас иногда называют, соответственно, верхними и нижними мотонейронами, могут быть только моносинаптическими, так как они осуществляются лишь через посредство аксонов клеток Беца. Эфферентные проводящие пути, связующие эти нейроны, принято именовать пирамидными, в связи с тем, что они участвуют в формировании находящихся на вентральной поверхности продолговатого мозга пирамид.
Когда
наличие пирамидной системы уже стало общепризнанным, исследователи
обратили внимание на то, что в обеспечении двигательных функций
принимают участие и многие другие клеточные структуры, расположенные на
разных уровнях ЦНС, которые стали именовать экстрапирамидными (термин был введен в
В дальнейшем было установлено, что большинство связей между центральными и периферическими мотонейронами полисинаптические, так как в них включаются и клетки, которые находятся в различных экстрапирамидных структурах, расположенных в подкорковых отделах больших полушарий и в стволе мозга.
По предложению Р. Гранита (Granit R., 1973) структуры так называемых пирамидных путей, от которых главным образом зависят активные движения тела и его частей, были названы фазическими. Экстрапирамидные структуры, влияющие на двигательные акты, положение, поддержание равновесия тела и его позу, названы Р. Гранитом тоническими.
Фазические и тонические структуры находятся между собой в отношении взаимного реципрокного контроля. Они образуют единую систему регуляции движений и позы, состоящую из фазической и тонической подсистем. На всех уровнях этих подсистем, начиная от коры и заканчивая мотонейронами спинного мозга, существуют коллатеральные связи между ними.
Тоническая и фазическая подсистемы являются не только взаимодополняющими, но и в определенном смысле взаимоисключающими. Так, тоническая система, обеспечивая сохранение позы, фиксирует положение тела напряжением «медленных» мышечных волокон, а также предотвращает возможные движения, которые могут привести к перемещению центра тяжести и, следо- вательно, изменению позы. С другой стороны, для осуществления быстрого движения необходимо не только включение фазической системы, ведущее к сокращению определенных мышц, но и снижение тонического напряжения мышц-антагонистов, что обеспечивает возможность выполнения быстрого и точного двигательного акта. В связи с этим статическое состояние, гиподинамия характеризуется гиперактивностью тонической системы и избыточным коллатеральным торможением фазической системы. Вместе с тем патологические синдромы, характеризующиеся быстрыми фазическими, избыточными, непроизвольными движениями (хорея, гемибаллизм и т.п.), обычно сочетаются с атонией.
Принятое
деление нервных структур, обеспечивающих двигательные акты, на
пирамидные и экстрапирамидные - не бесспорно, а названия им даны
случайно. Проводящий путь, именуемый пирамидным, был выявлен на спи-
нальном уровне и обозначен этим термином П. Флексигом в
В
В
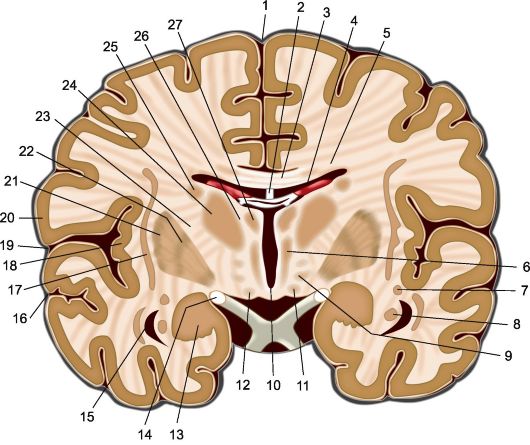
Рис 5.1. Фронтальный срез головного мозга на уровне сосцевидных тел. 1 - межполушарная продольная щель; 2 - свод; 3 - мозолистое тело; 4 - сосудистое сплетение бокового желудочка; 5 - лучистость мозолистого тела; 6 - медиальное ядро таламуса; 7 - хвост хвостатого ядра; 8 - гиппокамп; 9 - субталамическое ядро; 10 - III желудочек; 11 - сосцевидные тела; 12 - основание ножки мозга; 13 - миндалевидное тело; 14 - зрительный тракт; 15 - нижний рог бокового желудочка; 16 - верхняя височная борозда; 17 - ограда; 18 - островок; 19 - латеральная борозда, 20 - покрышка; 21 - скорлупа; 22 - бледный шар; 23 - внутренняя капсула; 24 - латеральные ядра таламуса; 25 - хвостатое ядро; 26 - мозговая пластинка таламуса; 27 - передние ядра таламуса.
этими системами. Не обособлены они и анатомически, за исключением короткой части пути через продолговатый мозг».
Высказанное П. Милнером мнение вполне логично, однако пока по традиции большинство нейрофизиологов и клиницистов признают целесообразность выделения пирамидной и экстрапирамидной системы. К экстрапирамидной системе обычно относят многочисленные клеточные образования, расположенные в больших полушариях мозга, в промежуточном мозге и в стволе мозга, а также афферентные и эфферентные связи между этими образованиями
(рис. 5.1, 5.2).
Основной частью экстрапирамидной системы принято считать подкорковые узлы или базальные ганглии, находящиеся в глубине больших полушарий. Прежде всего это такие парные образования, как чечевицеобразное ядро (nucleus lentiformis) и хвостатое ядро (nucleus caudatus), а также миндалевидное тело (corpus amygdaloideum).
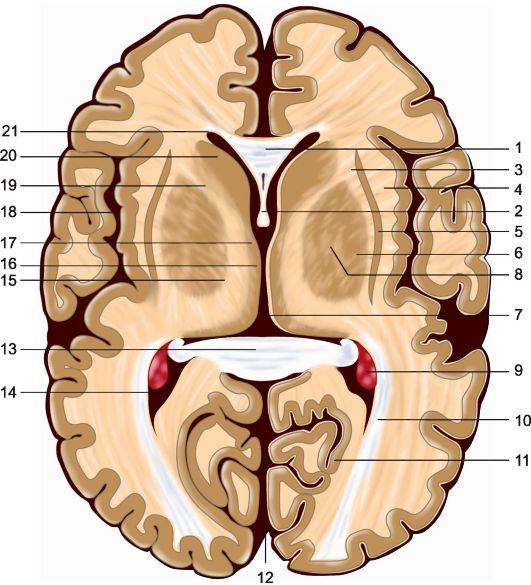
Рис 5.2. Горизонтальный срез головного мозга на уровне мозолистого тела. 1 - колено мозолистого тела; 2 - свод; 3 - наружная капсула; 4 - самая наружная капсула; 5 - ограда; 6 - чечевицеобразное ядро; 7 - III желудочек; 8 - внутренняя капсула; 9 - сосудистое сплетение бокового желудочка; 10 - задняя таламическая лучистость; 11 - шпорная борозда; 12 - продольная межполушарная щель; 13 - валик мозолистого тела; 14 - задний рог бокового желудочка; 15 - латеральные ядра таламуса; 16 - медиальные ядра таламуса; 17 - передние ядра таламуса; 18 - островок; 19 - внутренняя капсула
Кроме того, к экстрапирамидной системе относятся субталамическое ядро Льюиса (nucleus subthalamicus), расположенное в промежуточном мозге; черная субстанция и красное ядро (substantia nigra et nucleus ruber), находящиеся в среднем мозге; вестибулярные ядра и нижняя олива (nucleus vestibularis et oliva inferior) - образования продолговатого мозга; а также ретикулярная формация ствола, мозжечок и участки главным образом медиобазальных отделов коры больших полушарий, имеющие связи с перечисленными образованиями мозга.
5.2. СТРУКТУРЫ И ОСНОВНЫЕ ФУНКЦИИ ЭКСТРАПИРАМИДНОЙ СИСТЕМЫ
Чечевицеобразное ядро - самое крупное из расположенных в глубине большого полушария мозга ядерное образование, состоит из трех члеников, сформированных из серого вещества. Два из них (медиальные), более светлые, составляют так называемый бледный шар (globus pallidus). Бледный шар состоит из крупных клеток, расположенных в петлях, которые образованы миелиновыми волокнами, находящимися здесь в большом количестве и обусловливающими его «бледность». Латерально расположенный членик чечевицеобразного ядра называется скорлупой (putamen). Скорлупа и находящееся поблизости хвостатое ядро состоят из большого количества мелких клеток с короткими ветвящимися отростками и больших мультиполярных нейронов между ними, имеющих длинные аксоны.
Сходство фило- и онтогенеза, гистологического строения и биохимического состава, а также определенная общность функций служат основанием для объединения скорлупы и хвостатого ядра в полосатое тело (corpus striatum seu neostriatum), или стриарную систему. Исчерченность полосатого тела обусловлена наличием чередования в нем участков серого и белого вещества. Стриарной системе противопоставляется паллидарная система, которая известна и как paleostriatum, так как является более древней в филогенетическом отношении и раньше формируется в процессе онтогенеза.
Стриарная и паллидарная системы имеют различное происхождение, разную структуру и в какой-то степени противоположные функции. Скорлупа и хвостатое ядро происходят из паравентрикулярных структур, расположенных вблизи бокового желудочка, тогда как бледный шар, располагаясь вблизи III желудочка, имеет общее происхождение с субталамическим ядром. В паллидарной и стриарной системах предполагается наличие элементов соматотопического представительства.
Хвостатое ядро повторяет очертания бокового желудочка и имеет форму эллипса, при этом хвост его почти достигает миндалевидного ядра. Скорлупа находится снаружи бледного шара и отделена от него слоем миелинизированных волокон - боковой медуллярной пластинкой бледного шара. Латеральная сторона скорлупы отграничена от ограды наружной капсулой (capsula externa). В ее состав входят ассоциативные волокна, связывающие слуховую область коры височной доли с моторной и премоторной корой.
Паллидарная и стриарная структуры объединяются понятием стриопаллидар- ная система. Это объединение обусловлено тем, что при нормальной жизнеде- ятельности организма функции их взаимно уравновешивают друг друга, и благо- даря этому стриопаллидарная система оказывает влияние на двигательные акты как единое целое. Причем в этой единой функциональной системе паллидарные структуры обычно признаются активирующими, а стриарные - тормозящими. Стриопаллидарная система - составная часть экстрапирамидной системы, понятия более широкого, включающего в себя и ряд других структур мозга.
Структуры стриопаллидарной системы имеют связи между собой, а также афферентные и эфферентные связи с другими отделами экстрапирамидной системы, в частности с черной субстанцией, красным ядром, ретикулярной формацией, мозжечком, а также с корой больших полушарий и периферическими мотонейронами ствола и спинного мозга. Через переднюю спайку
мозга (комиссуру Мейнерта) осуществляется взаимодействие подкорковых узлов правого и левого полушарий. Тесная связь стриопаллидарной системы с ядрами гипоталамического отдела мозга определяет ее роль в механизмах эмоциональных реакций.
Полосатое тело получает импульсы от многих отделов мозговой коры, при этом особенно значительны ее ипсилатеральные связи с двигательными зонами (заднелобные отделы, предцентральная извилина, парацентральная долька). Нервные волокна, обеспечивающие эти связи, расположены в определенном порядке. Поступающая по ним импульсация оказывает в основном тормозное действие на клетки полосатого тела. Другая система афферентных волокон обеспечивает передачу импульсов в полосатое тело из центромедианного ядра таламуса. Эти импульсы оказывают на собственные клетки полосатого тела, скорее всего, активирующее действие.
Афферентные пути из хвостатого ядра и из скорлупы, составляющие полосатое тело, направляются к латеральному и медиальному сегментам бледного шара, разделенным тонкой медуллярной пластинкой. Кроме того, полосатое тело имеет прямые и обратные связи с черной субстанцией, которая обеспечивается соответственно аксонами стрионигральных и нигростриарных нейронов. Нигростриарные нейроны являются допаминергическими, тормозящими функцию стриарных холинергических нейронов и уменьшающими таким образом их тормозное влияние на структуры паллидума. ГАМКергические стрионигральные нейроны тормозят активность клеток черной субстанции. Они оказывают тормозное действие как на дофаминергические нигростриарные нейроны, так и на нигроспинальные нейроны, аксоны которых направляются к гамма-мотонейронам спинного мозга, регулируя таким образом тонус поперечнополосатых мышц. Часть нервных волокон, идущих от полосатого тела, обеспечивает его влияние на многие ядерные образования, относящиеся к экстрапирамидной и к лимбико-ретикулярной системам.
Из эфферентных волокон, исходящих из медиального сектора бледного шара, состоит, в частности, так называемая чечевичная петля (ansa lenticularis). Ее волокна идут вентромедиально вокруг задней ножки внутренней капсулы к таламусу, гипоталамусу и к субталамическому ядру. После перекреста эти проводящие пути, несущие импульсы из паллидарной системы, направляются в ретикулярную формацию ствола, откуда начинается цепь нейронов, формирующих ретикулоспинномозговой путь, заканчивающийся у мотонейронов передних рогов спинного мозга.
Основная масса волокон, исходящих из бледного шара, входит в состав таламического пучка (fasciculus thalamicus), состоящего из паллидоталамических и таламопаллидарных волокон, обеспечивающих прямую и обратную связи между паллидумом и таламусом. Обоюдными являются и нервные связи между правым и левым таламусами и корой больших полушарий. Существование таламокортикальных и кортикостриарных связей обеспечивает образование реверберирующих кругов, по которым нервные импульсы могут распространяться в обоих направлениях, обеспечивая согласованность функций таламуса, коры и полосатого тела. Импульсация, направляющаяся к коре со стороны таламуса и стриарной системы, по всей вероятности, влияет на степень активности двигательных зон коры больших полушарий. Регуляцию двигательной активности, адекватность темпа, амплитуды и координации движений обеспечивают также связи подкорковых узлов с вестибулярной, мозжечковой и проприоцептивной системами.
Кора больших полушарий воздействует на функциональное состояние стриопаллидарной системы. Влияние коры на экстрапирамидные структуры осуществляется через посредство эфферентных, нисходящих проводящих путей. Большинство их проходит через внутреннюю капсулу, меньшая часть - через наружную капсулу. Из этого следует, что поражение внутренней капсулы обычно прерывает не только пирамидные пути и корково-ядерные связи, но и ведет к изменению функционального состояния экстрапирамидных образований, в частности вызывает характерное в таких случаях выраженное повышение мышечного тонуса в контралатеральной части тела.
Деятельность сложно организованной экстрапирамидной системы, а также нервных пучков, составляющих корковоспинальный путь, в конце концов, направлена на обеспечение отельных движений и их коррекцию, а также на формирование сложных двигательных актов. Реализация влияния экстрапирамидных структур на мотонейроны спинного мозга осуществляется эфферентными системами. Эфферентные импульсы, идущие из образований стриопаллидарной системы, направляются к клеткам ретикулярной формации, вестибулярных ядер, нижней оливы и других структур экстрапирамидной системы. Переключившись в них с нейрона на нейрон, нервные импульсы направляются в спинной мозг и, проходя по ретикулоспинальному, тектоспинальному (начинающемуся в ядрах четверохолмия), руброспинальному пути Монакова, меди- альному продольному пучку (начинающемуся от ядер Даркшевича и Кахаля), вестибуло-спинномозговому и другим экстрапирамидным проводящим путям, достигают клеток его передних рогов.
Большая часть проводников (по пути следования от подкорковых узлов к клеткам передних рогов спинного мозга) совершает перекрест на разных уровнях мозгового ствола. Таким образом, подкорковые узлы каждого полушария мозга и другие клеточные образования головного мозга, относящиеся к экстрапирамидной системе (кроме мозжечка), оказываются связанными, главным образом, с альфа- и гамма-мотонейронами противоположной половины спинного мозга. Через проводящие пути, относящиеся к экстрапирамидной системе, а также через пирамидные полисинаптические пути они контролируют и регулируют состояние мышечного тонуса и двигательную активность.
От деятельности экстрапирамидных структур зависит способность человека принимать оптимальную для предстоящего действия позу, поддерживать необходимое реципрокное соотношение тонуса мышц-агонистов и антагонистов, двигательную активность, а также плавность и соразмерность двигательных актов во времени и пространстве. Экстрапирамидная система обеспечивает преодоление инерции покоя и инерции движений, координацию произвольных и непроизвольных (автоматизированных) и, в частности, локомоторных движений, спонтанную мимику, влияет на состояние вегетативного баланса.
В случаях нарушения функций той или иной структуры экстрапирамидной системы могут возникать признаки дезорганизации деятельности всей системы, что ведет к развитию различных клинических феноменов: изменения побуждения к движениям, полярные изменения мышечного тонуса, нарушение способности к осуществлению рациональных, экономичных, оптимальных по эффективности как автоматизированных, так и произвольных двигательных актов. Такие изменения, в зависимости от места и характера обусловившего их патологического процесса, могут варьировать в широких пределах, проявляясь в различных случаях подчас диаметрально противоположной симптоматикой:
от двигательной аспонтанности до различных вариантов насильственных, избыточных движений - гиперкинезов.
Много ценной информации о сущности деятельности нервных структур, относящихся к экстрапирамидным структурам, внесло изучение медиаторов, обеспечивающих регуляцию их функций.
5.3. КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
5.3.1. Общие положения
Сложность строения и функций стриопаллидарной системы, наличие в ней определенных элементов соматотопического представительства, обусловливает большое многообразие клинических проявлений ее поражения. Прежде всего выделяют две группы экстрапирамидных синдромов. Основой одной из них является акинетико-ригидный синдром, для другой ведущими оказываются различные варианты гиперкинезов.
Уже к
Расстройства баланса паллидарной и стриарной систем оказываются более выраженными в случае поражения стриопаллидарной системы. Нарушение функции ее стриарного отдела ведет к развитию быстрых гиперкинезов, возникающих на фоне понижения мышечного тонуса (например, хореические гиперкинезы). Если же поражается паллидум и доминирующей оказывается функция стриарной системы, развивается акинетико-ригидный синдром, характерный, в частности, для паркинсонизма. Для паркинсонизма экстрапира- мидного акинетико-ригидного синдрома ведущими клиническими признаками являются снижение двигательной активности и ригидность.
Этой гипотезой врачи руководствовались длительное время.
Третья группа экстрапирамидных нарушений обусловлена поражением мозжечка и его связей, однако ее из дидактических соображений принято рассматривать отдельно, и мы по той же причине уделили ей главу 7.
5.3.2. Акинезия и ригидность
Вариантами снижения двигательной активности являются: акинезия - отсутствие движений, брадикинезия - замедленность движений, олигокинезия -
бедность движений, гипокинезия - недостаточность двигательной активности. При этих изменениях двигательных функций проявляются также инерция покоя и движений, удлинение латентного периода между стимулом и ответной реакцией на него, ухудшением способности к регуляции скорости движения, к смене характера и темпа повторных двигательных актов. Все эти клинические явления «скрадывают выразительность» движений, действий и не имеют прямой зависимости от выраженности обычно сопутствующего им повышения мышечного тонуса по пластическому типу (мышечной ригидности).
Снижение двигательной активности при паркинсонизме сопряжено с недостаточностью побуждения, инициативы к движению, с трудностью для больного начинать движение, преодолевая при этом чрезмерную по степени выраженности инерцию покоя. Сила мышц при этом сохраняется, хотя достижение ее максимума проявляется с запозданием. В итоге у больного развиваются двигательная пассивность, медлительность, иногда он может часами сохранять принятое положение, фиксированную позу, напоминая в таких случаях больного в ступорозном состоянии.
Проявлением снижения двигательной активности и повышенного напряжения мышц могут быть гипомимия - бедность мимики, гипофония - ослабление звучности и монотонность речи, микрография - мелкий почерк. Характерно нарушение физиологических автоматизированных, содружественных движений - синкинезий (например, ахейрокинез - отсутствие содружественных движений рук при ходьбе).
Маскообразность лица в сочетании с общей гипокинезией, при которой теряются типичные для каждого человека индивидуальные особенности походки, жестикуляции, мимики, присущая каждому индивидуальная манера держаться, разговаривать, делают больных с характерным для паркинсонизма акинетико-ригидным синдромом похожими друг на друга. При выраженном акинетико-ригидном синдроме лишь глаза, точнее - взор, сохраняют свою подвижность.
Изучение акинезии
подтверждает, что базальные ядра имеют важное значение в осуществлении
начала (запуска) движения и автоматизированного выполнения действий в
соответствии с ранее приобретенными двигательными навыками.
Нейрохимические исследования позволили установить, что гипокинезия -
следствие возникающего в стриарной системе дефицита допамина,
обусловленное недостаточностью функции нигростриарных нейронов,
расположенных в черной субстанции. Причиной такой неврологической
патологии является развитие дегенеративных процессов в черной субстанции, что было установлено в
Кроме того, на развитие акинезии может влиять и повреждение содержащихся в черной субстанции допаминергических, нигроретикулярных нейронов, аксоны которых направляются в ретикулярную формацию (РФ) ствола. Там происходит переключение импульсов на нервные клетки, аксоны которых участвуют в формировании ретикулоспинномозгового пути. Снижение интенсивности импульсации, проходящей по ретикулоспинальным путям, вызывает торможение клеток гамма-мотонейронов, что способствует повышению тонуса поперечнополосатой мускулатуры и при этом ведет к развитию мышечной

Рис. 5.3. Акинетико-ригидный синдром при паркинсонизме.
ригидности. Нельзя исключить и того, что в патогенезе гипокинезии-акинезии и замедленности мышления (акайрии), опре-
деленную роль играет угнетение функций коры больших полушарий,
возникающее вследствие подавления влияния на нее активирующей
ретикулярной формации, описанной Г. Мэгуном и Р. Моруцци (Magoun H.,
Moruzzi R.,
Ригидность - постоянное пребывание мышц в состоянии тонического напряжения, которое свойственно как мышцамагонистам, так и мышцам-антагонистам, в связи с чем и проявляется пластический характер повышения мышечного тонуса. При пассивных движениях в конечностях больного обследующим ощущается не меняющееся, вязкое, воскоообразное сопротивление. Сам больной предъявляет прежде всего жалобы на скованность.
При акинетико-ригидном синдроме в начальной стадии его развития мышечная ригидность при болезни Паркинсона обычно асимметрична, может проявляться в какой-либо одной части тела, однако в дальнейшем, по мере прогрессирования заболевания, она становится все более распространенной и со временем генерализованной.
Меняется поза больного (рис. 5.3): голова и туловище наклоняются вперед, при этом подбородок нередко почти касается груди, руки прижаты к туловищу, согнуты в локтевых и лучезапястных суставах, пальцы рук согнуты в пястно-фаланговых и разогнуты в межфаланговых суставах, при этом большой палец находится в состоянии оппозиции к остальным. Повышение тонуса в мышцах шеи приводит к тому, что уже на раннем этапе заболевания на оклик больные склонны поворачиваться всем туловищем или максимально поворачивать взор, оставляя голову неподвижной.
Основные различия между ригидностью и спастичностью:
1. Распределение зон повышенного мышечного тонуса: ригидность проявляется как в мышцах-сгибателях, так и в мышцах-разгибателях, но более выражена в сгибателях туловища, значительна и в мелких мышцах лица, языка и глотки. Спастичность сочетается с парезом или параличом и при гемипарезе имеет тенденцию к формированию позы Вернике-Манна (рука согнута, нога разогнута).
2. Качественные показатели гипертонуса: ригидность - сопротивление пассивным движениям постоянное, тонус «пластический», положителен симптом «свинцовой трубки» (при пассивных движениях мышечное сопротивление равномерное, как при сгибании свинцовой трубки). Для спастического состояния мышц характерны симптом отдачи и симптом «складного ножа».
3. Ригидность меньше связана с повышенной активностью дуги сегментарных рефлексов, что характерно для спастичности и больше зависит от частоты разрядов в мотонейронах. В связи с этим сухожильные рефлексы при ри-
гидности не меняются, при спастичности - повышаются, при ригидности не возникает характерных для спастического пареза клонусов и патологических знаков (симптом Бабинского и т.п.).
4. Облигатным проявлением ригидности является феномен «зубчатого колеса» , при спастическом парезе этот феномен не возникает.
При паркинсонизме выраженность гипокинезии и мышечной ригидности могут в определенной степени зависеть от общего состояния больного. В покое гипокинезия и мышечная ригидность выражены резче, при медленных пассивных движениях иногда наблюдается некоторое ослабление ригидности. На гипокинезию и ригидность в значительной степени влияет психическое состояние больного, особенно отрицательные эмоции, которые иногда резко усиливают мышечный тонус. Вместе с тем по утрам, после сна может значи- тельно уменьшаться выраженность обоих компонентов акинетико-ригидного синдрома. Это же иногда проявляется и в некоторых экстремальных ситуациях (кратковременные проявления парадоксальной кинезии). Некоторое снижение выраженности ригидности мышц отмечается и во время пребывания больного в теплой ванне или при лечебном массаже. Все это позволяет судить о том, что функциональный дефект при акинезии и при ригидности в определенных пределах изменчив, в некоторых случаях он может колебаться в степени выраженности: от состояния общей обездвиженности до эпизодов почти полного восстановления функциональных возможностей моторной сферы.
5.4. ДОПАМИНЕРГИЧЕСКАЯ ТЕОРИЯ РАЗВИТИЯ АКИНЕТИКО-РИГИДНОГО СИНДРОМА
С
расширением возможностей нейрохимического и нейрофизиологического
обследования больных было установлено, что при паркинсонизме снижена
концентрация допамина в структурах стриарной системы. Это
обстоятельство привело к проведению серии исследований, обусловивших создание в
В начале этого биохимического ряда, в котором каждый предшествующий элемент трансформируется в последующий с участием определенного фермента, стоит аминокислота фенилаланин (Ф). Катехоламиновый ряд биохимических реакций может быть представлен следующим образом: Ф - тирозин - ДОФА (диоксифенилаланин) - ДА - НА - А. Каждый этап приведенных биохимических превращений осуществляется при участии определенного фермента. Так, превращение тирозина в ДОФА происходит с помощью фермента тирозингидроксилазы; ДОФА превращается в ДА благодаря дофадекарбоксилазе и т.д.
Установлено, что ДА продуцируется клетками черной субстанции. Его дегенерация при паркинсонизме была обнаружена в
в связи с повреждением или гибелью нигростриарных нейронов в стриатум попадает недостаточное количество медиатора допамина, холинергические нейроны стриарного тела оказываются расторможенными и их собственное тормозное влияние на клетки паллидарной системы становится чрезмерным. Снижение же функции структур паллидума провоцирует ригидность мышц и сказывается подавлением двигательной активности, проявляющимся гипокинезией или акинезией.
Кстати, изложение теории Р. Хасслера демонстрирует и примеры феноменов, часто наблюдаемых в ЦНС: 1) феномен гетерогенности нейронов единой нейронной цепи (ее составляют нейроны, различные по продуцируемым ими медиаторам); 2) феномен анатомо-биохимической диссоциации (поражение одной морфологической структуры ведет к биохимическим изменениям в других структурах мозга и нарушению их функций).
Таким образом, в норме ДА-ергические нейроны черной субстанции оказывают тормозное влияние на холинергические нейроны стриатума, сдерживая их тормозящее действие на паллидум. В случае же поражения черной субстанции в подкорковых структурах нарушается баланс между содержанием ДА и АХ (дефицит ДА при относительном избытке АХ), при этом растормаживается стриатум и его тормозящее влияние на паллидум становится чрезмерным, что и ведет к развитию характерного для паркинсонизма акинетико-ригидного синдрома.
Восстановить нарушенный таким образом медиаторный баланс между концентрацией ДА и АХ в экстрапирамидной системе можно путем уменьшения в стриопаллидарной системе уровня АХ или увеличения содержания ДА. Отсюда понятна эффективность лечения паркинсонизма лекарственными препаратами из группы М-холинолитиков (циклодол и пр.). Вместе с тем очевидна и возможность лечения паркинсонизма путем увеличения концентрации в мозговой ткани ДА. С этой целью в клинической практике обычно применяют предшественник допамина в катехоламиновом ряду биохимических реакций - левовращающего изомера диоксифенилаланин (препарат L-ДОФА) и агонисты допамина.
Следует отметить, что допаминергическая теория Р. Хасслера имеет несомненное большое практическое значение, так как помогает в большинстве случаев подобрать оптимальную схему лечения больного, однако и она не отражает всей полноты патогенетических проявлений, определяющих многообразие вариантов клинической картины синдрома паркинсонизма.
Краткая информация о патологических насильственных движениях приведена в следующей главе.