Общая неврология А.С. Никифоров, Е.И. Гусев - 2007г. - 720 с
|
|
|
|
ГЛАВА 4 ДВИЖЕНИЯ. ОСНОВНОЙ ДВИГАТЕЛЬНЫЙ КОРКОВО-МЫШЕЧНЫЙ ПУТЬ
4.1. ОБЩИЕ ПОЛОЖЕНИЯ
Движения - наиболее универсальные проявления жизнедеятельности. Они могут возникать по типу безусловного или условного рефлекса (от лат. reflexus - отражение) или представляют собой различной сложности произвольные и непроизвольные двигательные акты.
Рефлекторная деятельность является отраженной, т.е. осуществляется под влиянием определенных воздействий, исходящей из внешней и внутренней среды. Рефлексы впервые были описаны французским философом и естествоиспытателем Р. Декартом (Descartes R., 1598-1650). И.М. Cеченов (1829- 1905) признавал, что «все акты сознательной и бессознательной жизни по способу своего происхождения суть рефлексы», и считал, что рефлекс - основа нервной деятельности. Эту точку зрения разделяли И.П. Павлов (1849-1936) и В.М. Бехтерев (1857-1927), разработавшие учение об условных (по Павлову), или сочетательных (по Бехтереву) рефлексах.
Каждый рефлекс имеет свою рефлекторную дугу - цепь нейронов. Область локализации рецепторов, раздражение которой ведет к возникновению определенного рефлекса, называется рефлексогенной зоной этого рефлекса. От рефлексогенной зоны, возникшие вследствие ее раздражения, нервные импульсы распространяются вначале по центростремительной (афферентной) части рефлекторной дуги до области ее замыкания - того или иного отдела центральной нервной системы. Здесь импульсы после переработки переходят на центробежную (эфферентную) часть рефлекторной дуги, по которой они и достигают рабочего органа: мышцы или железы.
Рефлекторные дуги врожденных (биологических, безусловных) рефлексов замыкаются в спинном мозге и на различных уровнях ствола и промежуточного отдела головного мозга. Через их посредство на основании врожденных нервных связей осуществляются простые безусловные, а также такие более сложные рефлекторные акты, как пищевой, пассивно-оборонительный, половой и т.п.
Условные рефлексы формируются в процессе жизнедеятельности путем вы- работки временных связей на базе безусловных рефлексов. Рефлекторные дуги их замыкаются главным образом в коре больших полушарий головного мозга. Cовокупность условных рефлексов обеспечивает многообразие сложных реакций организма в его взаимоотношениях с внешней средой.
В процессе формирования двигательного акта кора большого мозга постоянно получает информацию о ходе его выполнения благодаря существующей
обратной связи (обратной афферентации) . Это позволяет мозгу контролировать рефлекторный акт и корригировать его на любом этапе. Признавая наличие обратных связей, следует отметить, что когда мы говорим о рефлекторной дуге, то допускаем упрощенную трактовку понятия, так как логичнее считать, что при обеспечении рефлекторного акта нервные импульсы движутся по «рефлекторным кольцам».
Произвольные двигательные акты могут характеризоваться особой сложностью. В их основе лежат не просто реакции на определенный сигнальный раздражитель, а стремление к удовлетворению потребностей (у человека и социальных потребностей) по определенной программе, составляемой в мозге на основании обширного афферентного синтеза всех тех сигналов из внешнего мира и внутренней среды организма, которые поступают в мозг по различным чувствительным путям (зрительным, слуховым, тактильным, проприоцептивным и пр.), а также на основании использования аппаратов памяти, сохраняющих и воспроизводящих сведения о прошлом опыте.
Известно, что, приступая под влиянием определенных мотиваций к действию, человек обычно прогнозирует его результаты. В связи с этим П. К. Анохин выдвинул гипотезу о существовании физиологического аппарата предвидения и оценки результатов действия, который формируется на основе предшествующего опыта, мотиваций и избранной программы предстоящего действия, а также текущего афферентного синтеза. Этот физиологический аппарат, предназначенный для восприятия информации о намеченном и производимом действии, а также о его результатах и сопоставления их со сложившимся ранее прогнозом, П.К. Анохин (1898-1974) назвал акцептором результатов действия.
Совпадение или несовпадение прогнозируемых и реальных результатов действия оказывает влияние на характер сопутствующих действию эмоциональных реакций. При совпадении реальных результатов с прогнозом они обычно положительны, при несовпадении - отрицательны. Последнее обсто- ятельство является стимулом к коррекции производимых действий или к до- полнительным действиям, направленным на достижение поставленной цели.
4.2. ДВИЖЕНИЯ И ИХ РАССТРОЙСТВА
Виды движений. Движения человека - следствие сокращения и расслаб- ления поперечнополосатых мышц. По характеру обусловленных движением изменений взаимного расположения частей тела в пространстве различают движения сгибательные, разгибательные и вращательные, или ротаторные. Вариантом ротаторных движений можно считать супинацию - вращение ко- нечности вокруг ее оси кнаружи и пронацию - вращение конечности вокруг ее оси внутрь. Движения, сопровождающиеся перемещением всего тела в пространстве, называются локомоторными. Они совершаются путем перемены позиции конечностей с участием почти всей мускулатуры тела. Виды локомоторных движений: ходьба, бег, плавание, ползание, прыжки и т.п.
Движения активные и пассивные . Активными являются движения, которые человек совершает самостоятельно (спонтанно, произвольно или по заданию). Пассивные движения - изменения в пространстве положения частей тела пациента, осуществляемые обследующим, при пассивном отношении больного к этим движениям.
В процессе неврологического обследования пациента проверяется возможность и активных, и пассивных движений в различных суставах. Надо иметь в виду, что для обеспечения активных движений, кроме нормальной иннервации определенных мышц, обеспечивающих данное движение, необходима сохранность этих мышц, а также костно-суставного аппарата, отсутствие значительных боли и болезненности, грубых нарушений проприоцептивной чувствительности. Поэтому нарушение активных движений может быть обусловлено разными причинами, некоторые из которых можно установить исследованием пассивных движений. Они невозможны или затруднены при заболевании или травматическом поражении и обусловленной этим значительной болезненности сустава, а также его анкилозом, при выраженном повышении мышечного тонуса, в частности при рефлекторно-тонических реакциях, характерных для менингита, радикулита и т.п. Пассивные движения могут быть избыточными, и это обычно указывает на значительное снижение мышечного тонуса.
При наблюдении за активными движениями обращают внимание на возможный объем, темп движений, силу, с которой сокращаются мышцы, обеспечивающие движения. При исследовании движений необходимо отчетливо представлять, благодаря сокращению каких мышц они происходят (табл. 4.1). Объем активных движений можно измерить в угловых градусах. Для определения силы кисти пользуются пружинным динамометром. Силу остальных мышц проверяют путем оказания сопротивления движению, производимому больным, или при осуществлении пассивных движений, преодолевая осуществляемое больным сопротивление, при этом обследующий субъективно оценивает силу мышц. Обычно она оценивается по 6-балльной системе (от 0 до 5).
Таблица 4.1. Функции и иннервация мышц
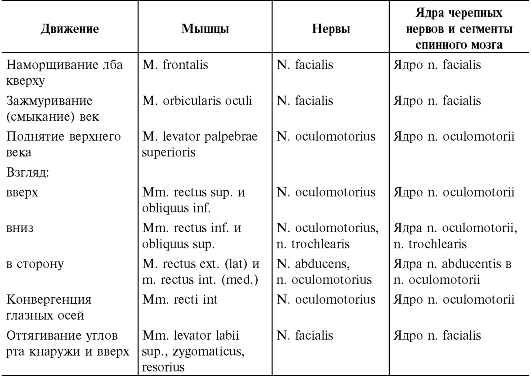
Продолжение табл. 4.1

Продолжение табл. 4.1
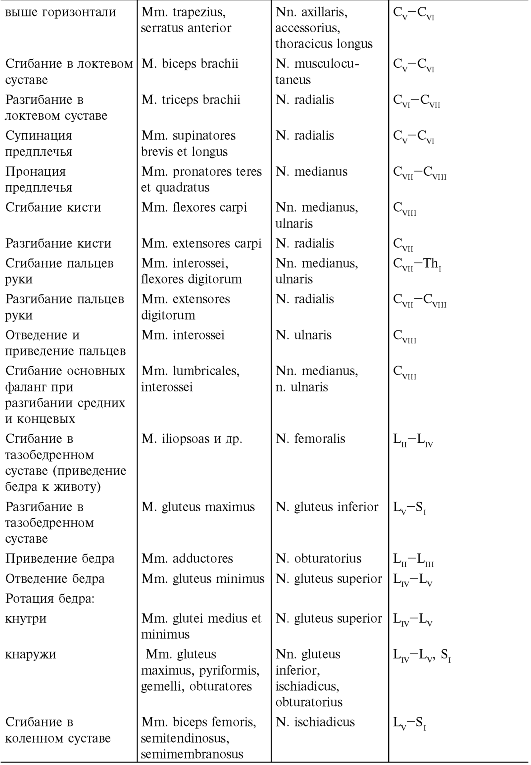
Окончание табл. 4.1

4.2.1. Параличи и парезы
Полное отсутствие активных движений, обусловленное нарушением иннервации мышц, называется параличом (paralisis) или плегией, а ограничение их в связи со снижением мышечной силы - парезом (paresis).
Выявление паралича или пареза обычно не представляет существенных трудностей. Надо лишь исключить ограниченность движений, обусловленную патологией опорно-двигательного аппарата, выраженностью болевого синдрома, нарушением сознания, ступором. Иногда несколько сложнее определить характер пареза или паралича и, в частности, дифференцировать органический парез или паралич от функционального (при истерии или симуляции).
Парез может иметь разную степень выраженности. Для выявления умеренных или слабо выраженных проявлений пареза целесообразно в процессе обследования больного провести определенные пробы. Вот некоторые из них.
1. Если пациент вытягивает вперед руки и удерживает их на горизонтальном уровне в положении супинации, то рука на стороне пареза вскоре переходит в положение пронации и медленно опускается (проба Мингаццини-Барре).
2. Если больной поднимет руки над головой и установит их ладонями кверху так, чтобы при этом средние пальцы рук едва касались друг друга, то на стороне пареза возникает тенденция к ротации предплечья и кисти и находящаяся в состоянии пареза рука постепенно опускается (проба Панченко, или феномен Будды).
3. Для выявления пареза ноги больному предлагается лечь на живот, при этом голени пассивно сгибаются под углом 45? к горизонтальной плоскости; больной должен некоторое время фиксировать голени в таком положении. На
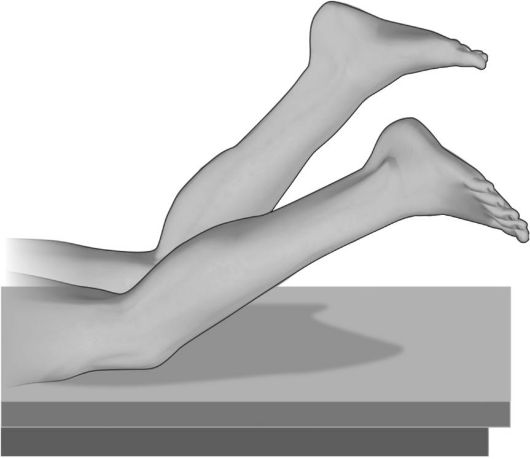
Рис. 4.1. Проба Барре (нижняя). Парез сгибателей левой голени.
стороне пареза удерживать ногу в этом положении труднее, поэтому она постепенно опускается - нижняя проба Барре (рис. 4.1).
По распространенности параличей и парезов различают моноплегию или монопарез - нарушение активных движений в одной конечности; параплегию или парапарез (верхний или нижний) - двигательные расстройства в обеих руках или ногах; гемиплегию или гемипарез - нарушение движений в руке и ноге на одной стороне в сочетании с центральным парезом на той же стороне мышц лица и языка; тетраплегия или тетрапарез - нарушение движений во всех четырех конечностях. Если при тетраплегии или тетрапарезе парализованы и мышцы, иннервируемые черепными нервами, то речь может идти о двойной гемиплегии. Паралич или парез называется альтернирующим (перекрестным) при сочетании паралича (пареза) мышц, иннервируемых каким-либо черепным нервом на стороне патологического процесса, а на противоположной стороне - гемиплегией или гемипарезом. Альтернирующий паралич или парез чаще является следствием одностороннего поражения ствола мозга.
4.2.2. Мышечный тонус
Тонус (от лат. tonus - напряжение) - рефлекторное напряжение мышц, которое зависит от характера достигающих их нервных импульсов (нервномышечный тонус) и от происходящих в них метаболических процессов (собственный тонус мышц).
В норме мышца не бывает совершенно расслабленной. Это обусловлено тем, что в ответ на эфферентную импульсацию, поступающую из головного мозга, и на афферентную, обусловленную раздражением проприорецепторов, поперечнополосатые мышцы находятся в постоянном напряжении (тонусе), которое принято называть контрактильным или физическим мышечным тонусом.
В процессе обследования, чтобы правильно судить о состоянии мышечного тонуса у пациента, производя пассивные движения, следует добиться максимального расслабления мышц. С этой целью необходимо отвлечение его внимания от проводимого обследования: можно вовлечь пациента в разговор, предложить ему решение несложных арифметических задач и т.п. Наличие повышенного контрактильного мышечного тонуса ведет к тому, что каждое
пассивное движение сопряжено с преодолением некоторого сопротивления мышц-антагонистов.
От контрактильного мышечного тонуса и возникающей при его повышении спастики следует отличать пластический тонус мышц, характеризующийся мышечной ригидностью. При повышении тонуса мышц по пластическому типу мышечное сопротивление, выявляемое при пассивных движениях, равномерно, как при сгибании-разгибании свинцовой трубки (симптом свинцовой трубки). Пластический тонус мышц определяется состоянием экстрапирамидных подкорковых структур (см. главу 5).
Мышечный тонус обеспечивает подготовку к движению, сохранность равновесия и позы. При сохранении фиксированной позы и при движениях возникает сокращение одних мышц и расслабление других. Соотношение тонуса мышц, агонистов и антагонистов (их реципрокность) изучал английский физиолог Ч. Шеррингтон (Sherrington Ch., 1857-1952). При нарушении иннервации мышцы тонус ее снижается. Если же периферические двигательные нейроны и сопряженные с ними спинальные рефлекторные дуги сохранны, а нарушен контроль за состоянием периферических двигательных нейронов со стороны церебральных структур, мышечный тонус обычно повышается.
Состояние тонуса мышц оценивается в процессе наблюдения за активными движениями больного, при осмотре и ощупывании его мышц, при пассивных
изменениях положения в пространстве частей тела больного. Снижение или отсутствие мышечного тонуса называется гипотонией или атонией мышц; высокий мышечный тонус - мышечной гипертонией.
При параличах и парезах изменяется контрак- тильный мышечный тонус. В случае гипотонии (и тем более атонии) мышцы вялые, дряблые, рельеф их не контурируется, при пассивных движениях отсутствует мышечное сопротивление, при этом объем движений может быть избыточным (симптом Ольшанского), суставы разболтаны. Со временем в таких случаях развивается гипотрофия мышц.
Для выявления снижения мышечного тонуса можно использовать следующие тесты.
1. Если больному с мышечной гипотонией в положении лежа на животе сгибать ногу в коленном суставе, то пятка ее может коснуться ягодичной области (пяточно-ягодичная проба).
2. Если больному с гипотонией мышц, ле- жащему на спине, одной рукой надавить на коленный сустав, а другой рукой разгибать голень, можно отметить, что пятка легко отрывается от горизонтальной плоскости.
3. При ходьбе у такого больного можно отметить переразгибание ног в коленных суставах (genu recurvatum), что характерно, в частности, для спинной сухотки (рис. 4.2).
Повышение контрактильного мышечного тонуса - мышечная спастика. При мышечной

Рис. 4.2. Genu recurvatum - переразгибание коленных суставов при спинной сухотке.
гипертонии, обусловленной повышением контрактильного тонуса, мышцы плотные, рельеф их отчетливый. Производя пассивные движения, исследующий отмечает определенное напряжение мышц и в связи с этим некоторую тугоподвижность суставов, особенно выраженную в начале движения и к концу пассивного движения - феномен складного ножа и отдачи.
4.2.3. Некоторые врожденные рефлексы и их изменения при параличах и парезах
4.2.3.1. Принципы исследования врожденных рефлексов
При неврологическом обследовании необходимо проверить состояние некоторых врожденных (безусловных) рефлексов, которые обычно выявляются у здорового человека. Если какой-либо из этих рефлексов не вызывается, т.е. об- наруживается арефлексия, или же имеется отчетливое уменьшение двигательной реакции в ответ на раздражение рефлексогенной зоны - гипорефлексия, можно предполагать, что патологический процесс в таких случаях нарушает целостность соответствующей рефлекторной дуги. В связи с этим, если проверка рефлексов проводится с целью уточнения топического диагноза, она имеет смысл лишь при условии, что обследующий знает состав их рефлекторных дуг (табл. 4.2).
Таблица 4.2. Уровни замыкания дуг глубоких и поверхностных рефлексов
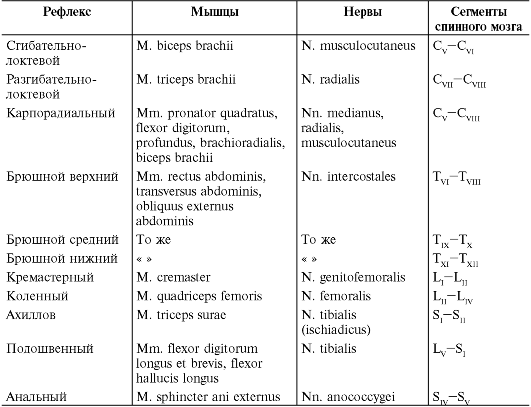
При проверке безусловных рефлексов желательно отвлечь внимание больного от проводимого обследования. Рефлексы проверяют с обеих сторон, постоянно сравнивая между собой рефлекторные реакции, вызванные путем одинакового по интенсивности раздражения симметричных рефлексогенных зон. Надо иметь в виду, что характер ответной реакции при вызывании многих рефлексов подвержен большим индивидуальным колебаниям, поэтому в клинической картине особо важное значение приобретает асимметрия рефлексов, так называемая анизорефлексия, которая практически всегда указывает на органическое поражение нервной системы.
Врожденные рефлексы, состояние которых обычно проверяется при неврологическом осмотре пациента, могут быть разделены на проприоцептивные (сухожильные и надкостничные рефлексы) и экстероцептивные (рефлексы с кожи и слизистых оболочек) в зависимости от того, раздражение каких рецеп- торов ведет к возникновению рефлекторной двигательной реакции.
4.2.3.2. Сухожильные, или миотатические, рефлексы
При проверке состояния сухожильных, или миотатических (от греч. myos - мышца, tatis - напряжение), рефлексов пользуются неврологическим молоточком, которым наносится короткий, отрывистый удар по сухожилию мышцы. Это ведет к ее растяжению, сменяющемуся сокращением, что и проявляется ответной двигательной реакцией. Тонус мышц и сухожильные рефлексы зависят от состояния мышечных веретен и афферентных волокон. Удар по мышечному сухожилию растягивает мышцу, раздражая веретена, активирует афферентные чувствительные нейроны задних рогов, передающие импульс на двигательные альфа-мотонейроны. В результате возникает мышечное сокращение, или миотатический рефлекс.
Обычно проверяются следующие сухожильные рефлексы.
• Рефлекс с двухглавой мышцы плеча (бицепс-рефлекс, сгибательно-локтевой рефлекс) вызывается ударом молоточка по сухожилию мышцы над локтевым сгибом или по ее апоневрозу на надплечье, при этом рука больного должна быть полусогнута и максимально расслаблена. Вызывая рефлекс, обследующий укладывает предплечье пациента на подставленное предплечье своей левой руки или поддерживает руку больного за кисть. Если пациент сидит, то при проверке этого рефлекса его предплечья могут свободно лежать на бедрах. Исследующий может, проверяя рефлекс, нащупать большим пальцем левой руки сухожилие двуглавой мышцы больного, после чего удары молоточком наносятся по ногтевой фаланге своего большого пальца. Ответная реакция при вызывании рефлекса - сгибание предплечья. Афферентная и эфферентная части рефлекторной дуги проходят по мышечно-кожному нерву. Замыкается рефлекторная дуга в CV-CVI сегментах спинного мозга (рис. 4.3а).
• Рефлекс с трехглавой мышцы плеча (трицепс-рефлекс, разгибательно-локтевой рефлекс) вызывается ударом молоточка по сухожилию мышцы над локтевым отростком, при этом рука больного пассивно слегка отводится назад и кнаружи, предплечье в таком случае свободно свисает. Ответная реакция - разгибание предплечья. Афферентная и эфферентная части рефлекторной дуги проходят через локтевой нерв. Замыкается рефлекторная дуга в CvII-CvIII сегментах спинного мозга (рис. 4.3б).
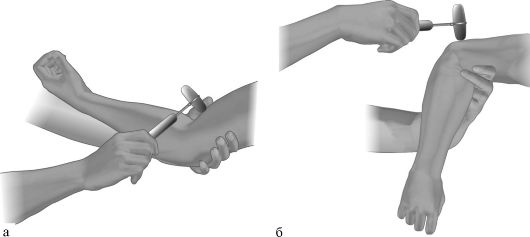
Рис. 4.3. Исследование рефлекса с двуглавой (а) и трехглавой (б) мышц плеча
• Коленный рефлекс (рефлекс с четырехглавой мышцы бедра) вызывается у пациента, находящегося в положении сидя (рис. 4.4) или лежа (рис. 4.5), ударом молоточка по сухожилию четырехглавой мышцы под надколенником, при этом ноги больного полусогнуты в коленных суставах, левая рука обследующего лежит на нижней трети бедер сидящего больного или подведена под его коленные суставы, если больной лежит. Ответная реакция - разгибание голени. Афферентная и эфферентная части рефлекторной дуги проходят по бедренному нерву. Замыкается рефлекторная дуга в LII-LIV сегментах спинного мозга.
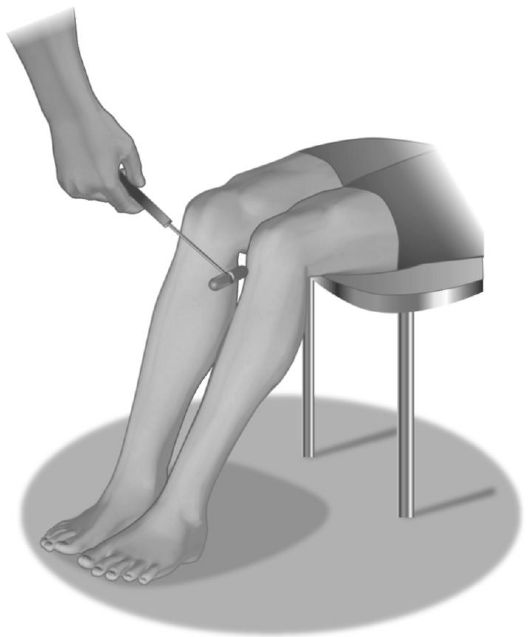
Рис. 4.4. Исследование коленного рефлекса у сидящего пациента.
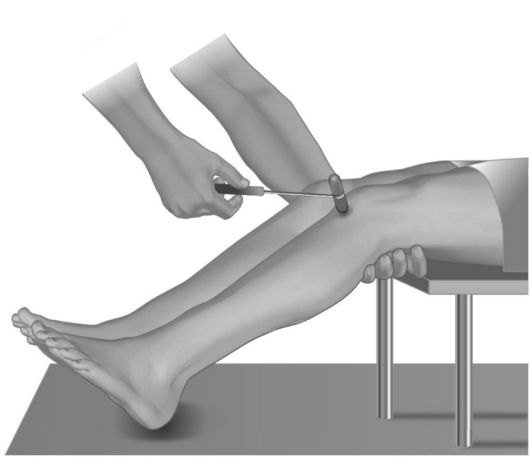
Рис. 4.5. Исследование коленного рефлекса у пациента, лежащего на спине.
• Рефлекс с пяточного сухожилия (ахиллов рефлекс) вызывается ударом молоточка по пяточному (ахиллову) сухожилию (рис. 4.6, а, б). Если больной лежит на спине, то нога его при этом может быть пассивно согнута обследующим в тазобедренном и коленном суставах и фиксирована в этом положении левой рукой. Удобно вызывать рефлекс у больного, стоящего на коленях, например на стуле, со свисающими стопами. Ответная реакция - подошвенное сгибание стопы. Афферентная и эфферентная части рефлекторной дуги проходят по седалищному нерву и его продолжению - большеберцовому нерву. Замыкается рефлекторная дуга в Si и Sii сегментах спинного мозга.
• Нижнечелюстной рефлекс (мандибулярный рефлекс, рефлекс с жевательной мышцы) вызывается ударом молоточка по нижней челюсти или по шпателю, положенному на зубы нижней челюсти пациента, сидящего с приоткрытым ртом. Ответная реакция - смыкание рта. Афферентная часть рефлекторной дуги проходит по третьей ветви тройничного нерва (нижнечелюстной нерв), эфферентная часть рефлекторной дуги - по двигательной порции того же нерва. Замыкается рефлекторная дуга в стволе мозга.
4.2.3.3. Надкостничные рефлексы
• Лучевой рефлекс (запястно-лучевой рефлекс, карпорадиальный рефлекс) вызывается ударом молоточка по шиловидному отростку лучевой кости или над ним, при этом руки больного согнуты под тупым углом в локтевых суставах, полупронированы и лежат на его бедрах, можно предплечье больного уложить на подведенное под него предплечье левой руки обследующего. Ответная реакция - сгибание руки в локтевом суставе в сочетании с ее пронацией. Афферентная и эфферентные части рефлекторной дуги проходят через лучевой и кожно-мышечный нервы. Замыкается рефлекторная дуга в Cv-CvIII сегментах спинного мозга.
• Лопаточно-плечевой рефлекс Бехтерева вызывается
поколачиванием молоточком по внутреннему краю лопатки. Ответная реакция
- приведение плеча и вращение его кнаружи. Афферентная и эфферентная
части рефлекторной дуги проходят по подлопаточному и надлопаточному
нервам. Замыкается рефлекторная дуга в Cv-CvI сегментах спинного мозга. Описал этот рефлекс В.М. Бехтерев в
4.2.3.4. Рефлексы со слизистых оболочек
• Роговичный рефлекс (корнеальный рефлекс) вызывается прикосновением ваткой или мягкой бумажкой к роговице. Ответная реакция при вызывании рефлекса - смыкание век. Афферентная часть рефлекторной дуги проходит по первой ветви тройничного нерва, эфферентная часть - по лицевому нерву. Замыкается рефлекторная дуга в продолговатом мозге.
• Глоточный рефлекс вызывается раздражением слизистой оболочки задней стенки глотки шпателем или свернутой в трубку бумагой. Ответная реакция - глотательное движение. Афферентная и эфферентная части рефлекторной дуги проходят по IX и X черепным нервам. Замыкается рефлекторная дуга в продолговатом мозге.
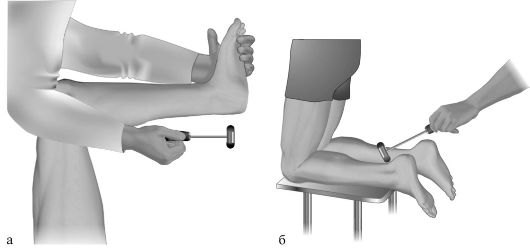
Рис. 4.6. Исследование рефлекса с пяточного сухожилия - ахиллова рефлекса (а, б).
• Нёбный рефлекс (рефлекс с мягкого нёба) вызывается путем прикосновения шпателем к мягкому нёбу. Ответная реакция - поднятие мягкого нёба. Афферентная и эфферентная части рефлекторной дуги проходят по IX и X черепным нервам. Замыкается рефлекторная дуга в продолговатом мозге.
• Анальный рефлекс вызывается покалыванием у края отверстия заднего прохода. Ответная реакция - сокращение круговой мышцы анального отверстия. Афферентная и эфферентная части рефлекторной дуги проходят по анально-копчиковым нервам. Замыкается рефлекторная дуга в SIV и SV сегментах спинного мозга.
4.2.3.5. Кожные рефлексы
Рефлекторные дуги кожных рефлексов замыкаются в коре головного мозга, эфферентная часть их проходит по пирамидным путям. В связи с этим кожные рефлексы появляются не сразу после рождения, а лишь по мере миелинизации пирамидных путей, обычно в возрасте от 6 мес до 2 лет, а исчезновение кожных рефлексов может сигнализировать о поражении не только спинальных рефлекторных дуг, но и проводящих путей, обеспечивающих их связи со структурами головного мозга (обычно при нарушении функции пирамидных путей). Вызываются кожные рефлексы штриховым раздражением кожи определенных частей тела рукояткой неврологического молоточка, спичкой и т.п. Ответная реакция - сокращение мышц, расположенных под раздражаемым участком кожи или вблизи от него. К кожным относятся брюшные, подошвенные и кремастерные рефлексы.
Брюшные рефлексы (рефлексы Розенбаха) вызываются быстрым штриховым раздражением кожи живота по направлению к средней линии, при этом на той же стороне его происходит сокращение соответствующих отделов прямой, поперечной, а также наружной и внутренней косых мышц живота. Для выявления верхнего (надчревного, эпигастрального) брюшного рефлекса штриховое раздражение наносится в верхней части брюшной стенки вдоль реберной дуги,
среднего (чревного, мезогастрального) - по горизонтальной линии на уровне пупка, нижнего (подчревного, гипогастрального) - в нижней части живота параллельно паховой складке. Дуга верхнего брюшного рефлекса проходит через ThVII-ThVIII, среднего - через ThIX-ThX, нижнего - через ThXI-ThXII сегменты спинного мозга и замыкается в головном мозге.
В норме брюшные рефлексы появляются с 5-6 мес жизни и характеризуются большим постоянством, если только брюшная стенка не оказывается чрезмерно дряблой. Они снижаются, быстро истощаются или даже исчезают при поражении указанных сегментов спинного мозга, соответствующих им спинальных корешков и периферических нервов, а также пирамидных путей. В топической диагностике существенное значение может иметь неидентичность степени выраженности (анизорефлексия) брюшных рефлексов справа и слева. Описал брюшные рефлексы немецкий невропатолог О. Розенбах (O. Rosenbach, 1851-1907).
Подошвенный рефлекс вызывается штриховым раздражением подошвенной стороны стопы. Ответная реакция - подошвенное сгибание пальцев стопы. Афферентная и эфферентная части рефлекторной дуги проходят по седалищному и большеберцовому нервам и Lv-Si сегментам спинного мозга.
Кремастерный рефлекс вызывается штриховым раздражением кожи внутренней поверхности бедра. Ответная реакция - подтягивание кверху яичка на стороне раздражения в связи с сокращением m. cremaster. Афферентная и эфферентная части рефлекторной дуги проходят по бедренно-половому нерву и Li-Lii сегментам спинного мозга.
4.3. ОСНОВНОЙ ПРОВОДЯЩИЙ ПУТЬ ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ
4.3.1. Кора двигательной зоны
Основной
проводящий путь, обеспечивающий произвольные движения, это путь, по
которому нервные импульсы от двигательной зоны коры больших полушарий
проходят к поперечнополосатым мышцам. Двигательной зоной коры головного
мозга является, главным образом, кора предцентральной извилины (поле 4
и 6, по Бродману) и прилежащие к этой зоне ее территории. Кора здесь
состоит из 6 клеточных слоев. В v слое расположены большие пирамидные
клетки, описанные в
4.3.2. Корково-ядерные
и корково-спинномозговые связи
Нервные импульсы от центральных мотонейронов (поля 4 и 6), двигательных клеток, расположенных в прилежащих зонах лобной (поле 8) и теменной долей (поля 5 и 7), а также в поясной извилине (поля 23с и 24с), движутся в центробежном направлении по их аксонам, принимающим участие в форми-
ровании лучистого венца, а затем колена и передних двух третей задней ножки внутренней капсулы (рис. 4.7).
Колено внутренней капсулы составляют аксоны клеток центральных мотонейронов, несущие нервные импульсы к двигательным ядрам черепных нервов, расположенным в покрышке ствола мозга. Эта часть аксонов центральных двигательных нейронов формирует корково-ядерный (кортико-нук- леарный) проводящий путь. Волокна его направляются к двигательным ядрам черепных нервов, состоящим из тел периферических двигательных нейронов. Подойдя к этим ядрам, часть волокон корково-ядерного пути переходит на противоположную сторону, т.е. образуется частичный надъядерный перекрест корково-ядерных волокон. Исключение из этого правила составляют лишь волокна, идущие к нижней части ядра лицевого и к ядру подъязычного черепных нервов, так как они над уровнем соответствующих ядер совершают почти полный перекрест. В результате нижняя часть ядра лицевого нерва и ядро подъязычного черепных нервов получают нервные импульсы, как принято считать, только из противоположного полушария мозга, тогда как к остальным периферическим двигательным нейронам, расположенным в ядрах черепных нервов, нервные импульсы поступают как от противоположного большого полушария (по перекрещенным путям), так и из гомонимного полушария мозга (по неперекрещенным путям).
Аксоны центральных двигательных нейронов, составляющие передние 2/3 задней ножки внутренней капсулы, проходят в составе основания ствола мозга, и по пути следования большая их часть (приблизительно 97-98%) заканчивается у нервных клеток ретикулярной формации ствола мозга и в ядерных образованиях, относящихся к экстрапирамидной системе.
Подходя к этим
образованиям, аксоны корковых клеток ветвятся, в результате большинство
идущих от клеток коры эфферентных нервных импульсов возбуждает не
одну, а несколько встречающихся на пути промежуточных клеток.
Происходит процесс дивергенции эфферентных двигательных импульсов и,
в итоге, после вовлечения в состав двигательного пути промежуточных
нейронов, количество волокон, по которым продолжают перемещаться
двигательные импульсы, весьма существенно увеличивается. Эфферентные
двигательные пути, прошедшие и не прошедшие через промежуточные синаптические аппараты (это аксоны клеток Беца, число которых с каждой стороны - 25-30 тыс), в
дальнейшем составляют пирамиды, расположенные на вентральной
поверхности продолговатого мозга (подсчитано, что каждую из пирамид
составляет приблизительно 1 млн нервных волокон). Пройдя пирамиды, эти
волокна, названные в связи с этим в
ку и обеспечивают наибольшую скорость перемещения нервных импульсов по корково-спинальным проводящим путям.
Таким образом, пирамидный путь в настоящее время рассматривается как гетерогенная система, состоящая из нескольких подсистем с различными типами волокон, начинающихся от разных отделов мозговой коры. Подавляющее их большинство прерывается в определенных структурах подкорковых образований и ствола мозга. В связи с этим определение кортикоспинальной системы как моносинаптической, «пирамидной» не соответствует действительности, но оно традиционное, принятое в классической неврологической литературе, поэтому в дальнейшем мы не будем избегать термина «пирамидный путь», но понимание его должно соответствовать современным представлениям.
В спинном мозге пирамидные пути спускаются вниз, при этом на уровне каждого его сегмента часть составляющих их нервных волокон заканчивается у клеток передних рогов, являющихся периферическими (нижними) двигательными нейронами.
Волокна латерального проводящего пути подходят к каждому периферическому мотонейрону, в то время как волокна переднего пирамидного пути заканчиваются только у периферических мотонейронов, которые обеспечивают на той же стороне иннервацию мышц шеи и туловища и, в частности, дыхательной мускулатуры, способствуя сохранению дыхания в случае развития у больного гемиплегии. В связи с этим периферические мононейроны, име- ющие отношение к иннервации мышц шеи и туловища, получают нервные импульсы с обеих сторон, что создает условия для поддержания жизненно важных функций у больных с центральным гамипарезом.
Обобщая изложенное, можно отметить, что в передних рогах сегментов спинного мозга, где располагаются тела периферических двигательных нейронов, заканчиваются аксоны нейронов, идущие от нервных клеток, расположенных в многочисленных супраспинальных двигательных центрах. Здесь же заканчиваются и афферентные структуры, приносящие импульсы от рецепто- ров различных видов чувствительности, расположенных в пределах соответс- твующего сегмента тела.
Эфферентные и афферентные образования непосредственно или опосредованно (через вставочные нейроны) подходят к периферическим двигательным нейронам, в которых происходит суммация приходящих к ним возбуждающих и тормозных постсинаптических потенциалов (ВПСП и ТПСП). В результате синтеза импульсации, поступающей в периферический нейрон, формируется биоэлектрический заряд. Он и определяет характер образующегося в этом нейроне нервного импульса, направляющегося к сопряженным с ним мышечным волокнам.
В осуществлении и регуляции произвольных двигательных актов участвует большое количество нейротрансмиттерных систем. Уточнение характера медиаторов в разных синаптических зонах системы активных движений находится в стадии изучения. Однако уже сейчас известно, что ключевая возбуждающая нейротрансмиссия в центральных мотонейронах осуществляется в основном глутаматом и аспартатом, тогда как ГАМК и таурин способствуют снижению активности двигательных реакций. Нейротрансмиттером периферического двигательного нейрона, обеспечивающим передачу нервных импульсов в нервно-мышечных синапсах, является ацетилхолин.
Так называемый пирамидный путь не является единой, гомогенной системой волокон, он состоит из нескольких подсистем с различными связями
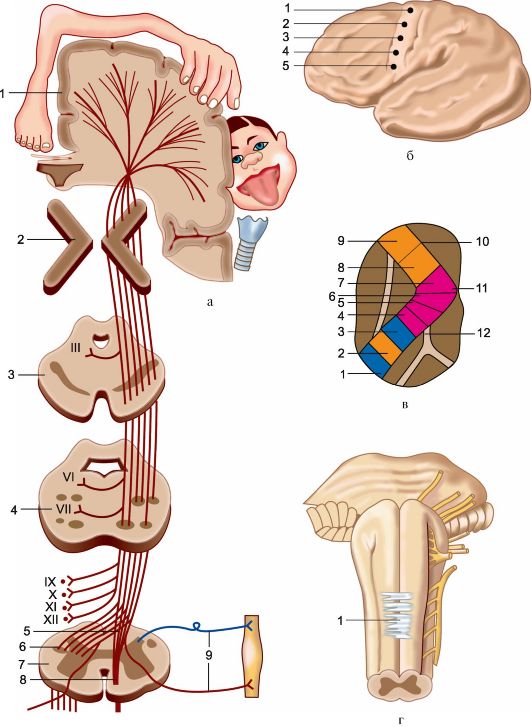
Рис. 4.7. Пирамидная система (схема).
а - пирамидный путь: 1 - кора большого мозга; 2 - внутренняя капсула; 3 - ножка мозга; 4 - мост; 5 - перекрест пирамид; 6 - латеральный корково-спинномозговой (пирамидный) путь, 7 - спинной мозг; 8 - передний корково-спинномозговой путь; 9 - чувствительные двигательные волокна периферического нерва; III, VI, VII, IX, X, XI, XII - соответствующие черепные нервы. б - конвекситальная поверхность большого полушария: двигательная зона коры (поля 4 и 6); топографическая проекция частей
противоположной половины тела: 1 - ноги; 2 - туловища; 3 - руки; 4 - кисти; 5 - лица; в - горизонтальный срез через внутреннюю капсулу, расположение в ней основных проводящих путей. 1 - зрительная и слуховая лучистость; 2 - височно-мостовые и теменно-затылочно-мостовые волокна; 3 - чувствительные таламокортикальные во- локна; 4 - пирамидные пути, проводящие импульсы к мотонейронам пояснично-крес- тцового утолщения спинного мозга; 5 - пирамидные волокна, проводящие импульсы к мотонейронам грудного отдела спинного мозга; 6 - пирамидные пути, проводящие импульсы к мотонейронам шейного утолщения; 7 - корково-ядерные пути; 8 - лоб- но-мостовой путь; 9 - корково-таламический путь; 10 - передняя ножка внутренней капсулы; 11 - колено внутренней капсулы; 12 - задняя ножка внутренней капсулы; г - передняя поверхность ствола мозга: 1 - перекрест пирамидных путей.
и функциями. Прямые корковоядерные и корковоспинномозговые проекции, т.е. волокна, обеспечивающие моносинаптические связи пирамидных клеток коры больших полушарий с периферическими мотонейронами, главным образом с теми из них, которые обеспечивают иннервацию дистальных отделов конечностей, представляют собой наиболее молодую в филогенетическом отношении систему, существующую только у приматов. В настоящее время известно, что в составе проводящего пути, обычно именуемого боковым кортико-спинальным, или пирамидным, аксоны гигантских пирамидных клеток Беца, расположенных в коре передней центральной извилины, и имеющих моносинаптические связи с периферическими мотонейронами составляют всего лишь 2-2,5%.
4.3.3. Периферические (нижние) двигательные нейроны
В
Клетки передних рогов спинного мозга представлены двумя вариантами мотонейронов: 1) мотонейроны, аксоны которых имеют толстую миелиновую оболочку (А-альфа волокна), и 2) мотонейроны, аксоны которых имеют тонкую миелиновую оболочку (А-гамма волокна). Нервные клетки по аналогии с обозначением нервных волокон называются альфа- и гамма-мотонейронами. Аксоны гамма-мотонейронов направляются в мышцы, где проникают в рецепторные аппараты - мышечные веретена, содержащие интрафузальные (находящиеся внутри веретена) волоконца. В случаях растяжения мышцы, например при вызывании сухожильного рефлекса, происходит изменение длины и интрафузальных мышечных волокон: меняется конфигурация мышечных веретен, а вместе с этим происходит раздражение находящихся на них спиралевидных рецепторных структур. Возникающие в этих рецепторных структурах нервные импульсы направляются по афферентным волокнам периферических нервных структур к псевдоуниполярным клеткам спинальных ганглиев. Далее по аксонам этих клеток импульсы попадают в спинной мозг и непосредственно через коллатерали или через вставочные нейроны того же сегмента спинного мозга
обеспечивают возбуждение крупных двигательных клеток (альфа-мотонейронов) передних рогов. Нервные импульсы по имеющим толстую миелиновую оболочку аксонам альфа-мотонейронов направляются к экстрафузальным волокнам, составляющим основную массу ранее подвергшихся растяжению поперечнополосатых мышц. Возникающее при этом возбуждение этих мышечных волокон вызывает напряжение или сокращение мышц, что обеспечивает пребывание в фиксированной позе или обусловливает определенный двигательный акт.
Часть дендритов нервных клеток спинальных ганглиев берет начало в мышечных сухожилиях, точнее, в расположенных в этих сухожилиях рецепторных аппаратах - сухожильных органах Гольджи. Эти рецепторы имеют высокий порог возбуждения, которое наступает лишь при чрезмерном растяжении мышечных сухожилий, угрожающем травматическим повреждением (разрывом, отрывом). В таких случаях из возбужденных сухожильных органов Гольджи в спинной мозг поступают импульсы, обеспечивающие уменьшение активности альфа-мотонейронов и снижение напряжения (тонуса) соответствующих мышц.
Кроме того, снижению мышечного тонуса при перевозбуждении альфа-мотонейронов способствуют импульсы, которые через посредство коллатералей, отходящих от их аксонов, достигают находящихся в составе переднего рога спинного мозга клеток Реншоу. Подвергшись возбуждению, эти клетки посылают тормозные биопотенциалы к альфа-мотонейронам, снижая при этом степень их напряжения.
Таким образом, периферический двигательный нейрон, наряду с влияниями, передающимися на него по эфферентным путям от корковых и экстрапирамидных структур головного мозга, испытывает воздействие импульсов, возникающих в рецепторных аппаратах на уровне того же спинального сегмента или в одноименном с этим сегментом миотоме. Все это в значительной степени способствует регуляции тонуса мышц, его оптимизации в соответствии с меняющимися требованиями, зависящими от характера выполняемых двигательных актов.
Итак, корково-мышечный путь включает центральные и периферические двигательные нейроны. К каждому периферическому нейрону поступает импульсация по нескольким, подчас многим нервным волокнам, идущим из коры большого мозга, из его многочисленных экстрапирамидных структур, а также из тканей одноименного спинальному сегменту метамера тела. Часть из них представляет собой возбуждающие, часть - тормозящие биоэлектрические потенциалы. Все они в периферическом двигательном нейроне подвергаются обработке и суммации. В результате от тела периферического двигательного нейрона к мышце направляются биоэлектрические потенциалы, являющиеся результатом синтеза свойств воздействующей на него импульсации. Перифе- рический двигательный нейрон обеспечивает иннервацию сопряженных с ним поперечнополосатых (от 5 до 1000) мышечных волокон и происходящие в них трофические процессы.
Группа мышечных волокон, иннервируемых одним периферическим мотонейроном и его аксоном, составляет двигательную единицу, при этом нервные волокна (аксон мотонейрона), идущие к мышце, обеспечивающей тонкие движения, иннервируют 5-12 мышечных волокон, тогда как крупные мышцы, осуществляющие грубые движения, получают импульсацию по нервным волокнам, каждое из которых иннервирует от 100 до 1000 их мышечных волокон. По-
нятие о двигательной единице как о функциональной единице нейромоторного аппарата было введено английским невропатологом Ч. Шеррингтоном (Sherrington Ch., 1857-1952). Мышца среднего размера имеет несколько сотен двигательных единиц. Группа периферических мотонейронов, иннервирующих одну мышцу, составляет двигательный пул.
Поражение периферических нейронов ведет к возникновению периферического паралича, который характеризуется полной обездвиженностью соответствующей части тела. Нарушение же функции центральных двигательных нейронов обусловливает центральный паралич, при котором отсутствуют произвольные движения, тогда как рефлекторные движения и синкинезии оказываются избыточными. В случае поражения части центральных двигательных нейронов, пирамидных путей или двигательных единиц, участвующих в иннервации мышц определенной части тела, в ней возникает парез, который может быть центральным или периферическим в зависимости от того, поражением элементов каких нейронов (центральных или периферических) этот парез обусловлен.
4.4. ПЕРИФЕРИЧЕСКИЙ ПАРАЛИЧ
Периферический паралич всегда является следствием поражения периферических двигательных нейронов и возникает как при поражении тел этих нейронов, расположенных в двигательных ядрах черепных нервов или в передних рогах сегментов спинного мозга, так и их аксонов, входящих в состав различных структур периферической нервной системы, а также при блокаде нервно-мышечных синапсов.
Избирательное поражение тел периферических двигательных нейронов характерно, в частности, для эпидемического полиомиелита и бокового амиотрофического склероза. Поражение структур периферической нервной системы может быть следствием травмы, сдавления, инфекционно-аллергического поражения и т.д., ведущих к развитию радикулопатий, плексопатий, невропатий, моноили полиневропатий. Несостоятельность нервно-мышечных синапсов, передающих нервный импульс с нервного окончания на мышцу с помощью медиатора ацетилхолина-Н, возникает при миастении, отравлении токсином ботулизма.
Для периферического, или вялого, паралича характерны следующие признаки.
1. Полная обездвиженность.
2. Атония. Резко выраженное снижение мышечного тонуса. Мышца становится вялой, тестообразной, аморфной, не реагирует на раздражители, лишена силы. При периферическом параличе конечности обычна избыточность пассивных движений в ее суставах.
3. Арефлексия. Исчезает рефлекторная реакция парализованных мышц в ответ на их внезапное раздражение, в частности растяжение, например, при ударе по сухожилию мышцы (сухожильные, или миотатические, рефлексы). Отсутствуют все рефлекторные двигательные реакции, в том числе защитные движения.
4. Атрофия. Если двигательный нейрон или его аксон погибают, то все связанные с ним мышечные волокна претерпевают глубокую денервационную атрофию. Со временем в связи с развитием атрофического процесса умень-
шается масса денервированных мышц. В течение нескольких недель после травмы или начала болезни гипотрофия мышц может быть незаметной, однако в течение первых 4 мес денервированные мышцы теряют до 20-30% исходной массы, а в дальнейшем - до 70-80%.
5. Реакция перерождения, или реакция дегенерации, - извращение реакции на раздражение электрическим током парализованной мышцы и нефункционирующего нерва.
В соответствии со сформулированным в
4.5. ЦЕНТРАЛЬНЫЙ ПАРАЛИЧ
Центральный паралич - следствие поражения как двигательной зоны коры мозга, так и любого отдела нервной системы, связывающего ее с периферическими двигательными нейронами. Наряду с поражением коры моторной зоны больших полушарий причиной центрального паралича может быть нарушение функций корково-ядерных, моно- и полисинаптических корково-спин- номозговых путей на уровне лучистого венца, колена и двух передних третей заднего бедра внутренней капсулы, ножки мозга, ствола мозга, спинного мозга. В большинстве случаев патологический очаг вызывает поражение не всех структур двигательной зоны коры и корково-спинномозговых путей, к тому же часть этих путей не подвергается перекресту. Поэтому на противоположной относительно патологического очага стороне тела никогда не поражаются все мышцы. Чаще в большей степени нарушена функция мышц кисти и плеча, затем - голени.
При центральном параличе сохранены и к тому же расторможены периферические мотонейроны, поэтому при нем, как правило, оказываются чрезмерно высокими сухожильные рефлексы, появляются патологические рефлексы и синкинезии.
Таким образом, центральный паралич - невозможность произвольных движений при сохранности и даже усилении интенсивности элементарных рефлекторных двигательных актов.
Для центрального, или спастического, паралича характерны следующие при- знаки.
1. Повышение мышечного тонуса по контрактильному типу, или спастичность мышц, обусловленная прекращением сдерживающих влияний центральных нейронов на периферические мотонейроны.
2. Повышение сухожильных и надкостничных рефлексов, также сопряженное с нарушением сдерживающих влияний центральных нейронов на периферические мотонейроны.
3. Возникновение клонусов, которые можно рассматривать как результат резко выраженной сухожильной гиперрефлексии, при этом однократное раздражение рефлексогенной зоны ведет к многократному ритмичному повторению стереотипного двигательного ответа.
4. Патологические рефлексы, многие из которых вызываются у детей раннего возраста, а затем исчезают. При расторможенности периферических нейронов в связи с ослаблением сдерживающего влияния церебральных структур эти врожденные рефлексы появляются вновь и уже рассматриваются как патологические.
5. Появление защитных рефлексов и патологических синкинезий (сопутствующих движений).
6. В отличие от периферического паралича при центральном параличе отсутствует реакция перерождения (или дегенерации) мышц и нервов.
Спастичность мышц. Спастическое напряжение мышц, при котором мышечный тонус при движениях неравномерно нарастает и возможны симптомы складного ножа и отдачи, является характерным признаком поражения структур, связывающих центральные двигательные нейроны с периферическими. При остром поражении спастический тонус обычно развивается не сразу, некоторое время парализованные мышцы могут быть вялыми, однако сухожильные рефлексы при этом повышены, возможно раннее появление стопного патологического рефлекса Бабинского. Спастический тонус связан с чрезмерной активностью периферических мотонейронов, освободившихся от тормозящих воздействий структур ретикулярной формации мозгового ствола. Преимущественно в руках повышается тонус в мышцах-сгибателях, в ногах - в разгибателях. У больных с центральным гемипарезом обычно со временем формируются поза и походка Вернике-Манна.
Клонусы. Как уже отмечалось, для центрального пареза или паралича характерно не только повышение сухожильных миотатических рефлексов, вызываемых и в норме, но нередко и появление многократной двигательной реакции на их вызывание (клонус). На практике чаще других вызываются следующие формы клонуса.
Клонус стопы вызывается следующим образом: больной лежит на спине, исследующий сгибает его ногу в коленном и тазобедренном суставах и, при- держивая одной рукой его голень, другой рукой резко производит разгибание (тыльное сгибание) стопы, при этом желательно, чтобы больной слегка упирался подошвенной поверхностью стопы о ладонь обследующего. Ответная реакция - ритмичное чередование сгибания и разгибания стопы в течение всего периода, пока обследующий натягивает пяточное (ахиллово) сухожилие. При центральных парезах, особенно в случаях поражения пирамидных путей спинного мозга, клонус стопы иногда возникает спонтанно, если больной упирается передней частью стопы об пол или о спинку кровати (рис. 4.8а).
Клонус надколенника вызывается у больного, лежащего на спине с выпрям- ленными ногами. Обследующий большим и указательным пальцами захватывает верхний полюс надколенника и после этого резко смещает его в сторону голени и удерживает его в этом положении. Происходит растяжение четырехглавой мышцы бедра и возникает ее ритмичное клоническое сокращение, не останавливающееся до тех пор, пока не будет прекращено растяжение мышечного сухожилия (рис. 4.8б).
Значительно реже описанных форм клонуса удается выявлять клонус нижней челюсти, клонус кисти, клонус ягодичной мышцы.
Патологические рефлексы. В зависимости от места вызывания и характера ответной двигательной реакции патологические рефлексы могут быть разделены на стопные и кистевые, а также на разгибательные (экстензорные) и сгибательные (флексорные).
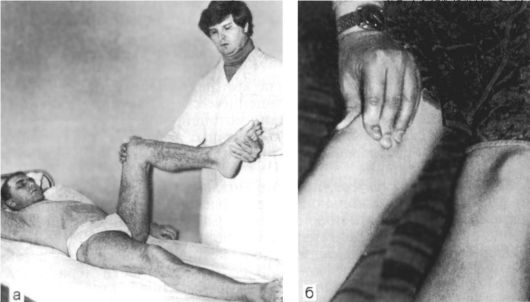
Рис. 4.8. Выявление клонуса стопы (а) и надколенника (б).
При всех патологических стопных разгибательных рефлексах, особенно часто встречающихся при центральных парезах и параличах, ответной реакцией является разгибание большого пальца иногда в сочетании с веерообразным расхождением остальных пальцев стопы. К разгибательным патологическим рефлексам относятся следующие.
Рефлекс Бабинского - вызывается штриховым раздражением кожи наружного края подошвенной поверхности стопы, направленным от пяток к пальцам (рис. 4.9а, б). Является наиболее постоянным при центральных параличах и парезах, но надо иметь в виду, что у детей в возрасте до 2 лет он обычно вызывается и в норме.
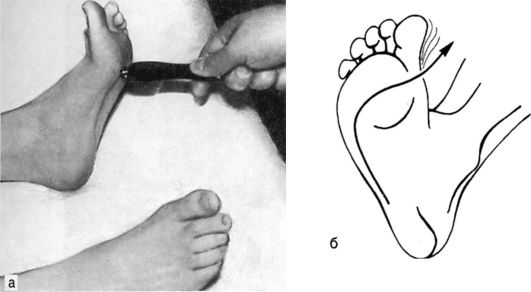
Рис. 4.9. Патологический рефлекс Бабинского (а) и его схема (б).
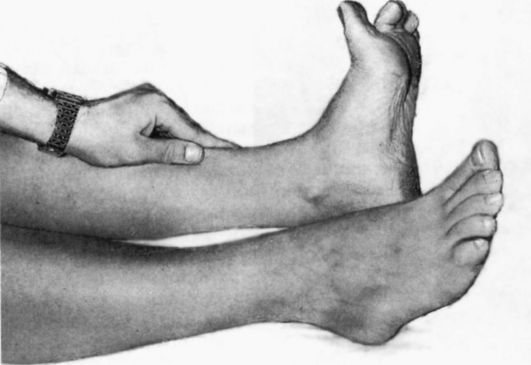
Рис. 4.10. Патологический рефлекс Оппенгейма.
Рефлекс Оппенгейма - вызывается проведением большим и указательным пальцами по гребню большеберцовой кости с некоторым нажимом сверху вниз к голеностопному суставу (рис. 4.10).
Рефлекс Гордона - вызывается сдавлением трехглавой мышцы голени
(рис. 4.11).
Рефлекс Шеффера - вызывается сдавливанием пяточного сухожилия
(рис. 4.12).
При всех сгибательных стопных патологических рефлексах ответная двигательная реакция сводится к быстрому кивательному подошвенному сгибанию пальцев стопы. К сгибательным стопным патологическим рефлексам относятся следующие.
Рефлекс Россолимо - вызывается короткими легкими ударами пальцами руки исследующего по подошвенной поверхности концевых фаланг II-V пальцев стопы больного (рис. 4.13).
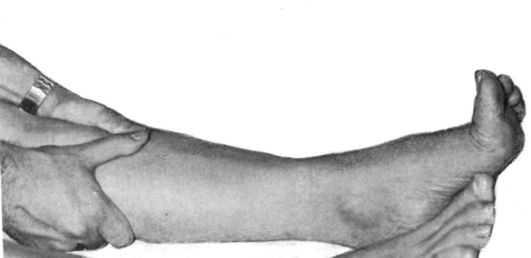
Рис. 4.11. Патологический рефлекс Гордона.
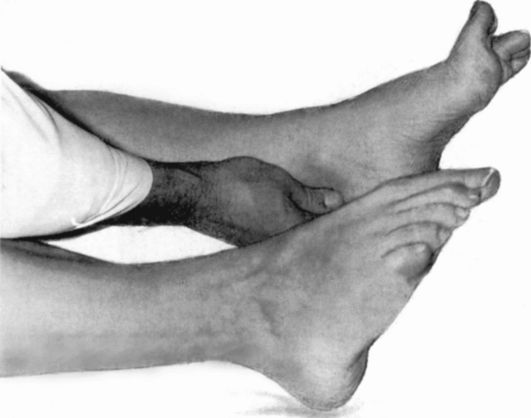
Рис. 4.12. Патологический рефлекс Шефера.
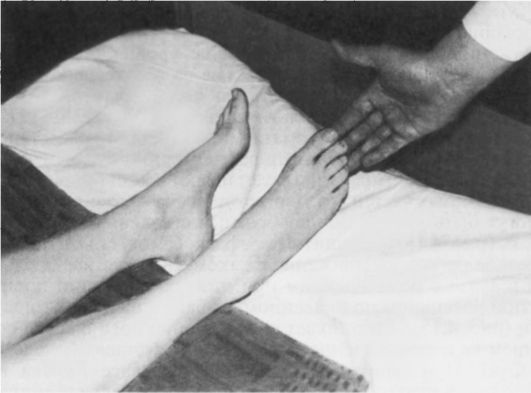
Рис. 4.13. Патологический рефлекс Россолимо.
Рефлекс Бехтерева-Менделя - вызывается постукиванием молоточком по тылу стопы над III-IV плюсневыми костями.
Рефлекс Жуковского-Корнилова - вызывается постукиванием по центральной части подошвы (рис. 4.14).
При поражении пирамидных путей выше уровня шейного утолщения спинного мозга могут вызываться и кистевые патологические рефлексы, которые в основном можно рассматривать как аналоги стопных.
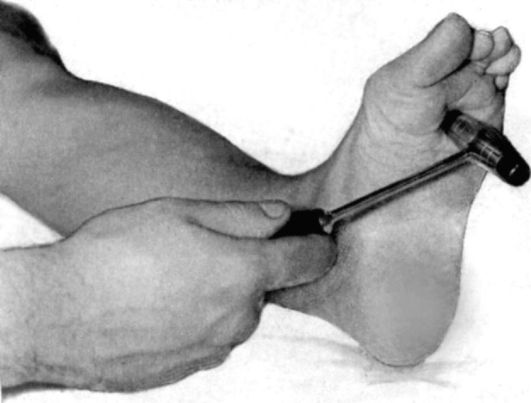
Рис. 4.14. Патологический рефлекс Жуковского-Корнилова.
Защитные рефлексы. Как и патологические рефлексы, защитные рефлексы, или рефлексы спинального автоматизма, возникают вследствие изоляции от расположенных выше отделов мозга рефлекторных дуг, замыкающихся на уровне спинальных сегментов. Они представляют собой непроизвольные движения парализованных частей тела в ответ на их раздражение. Раздражителями при этом могут быть уколы, щипкообразные сдавления кожи, раздражение эфиром и т.п. (рис. 4.15а). Защитные рефлексы могут быть получены также в ответ на резкое пассивное движение в каком-либо суставе парализованной конечнос-
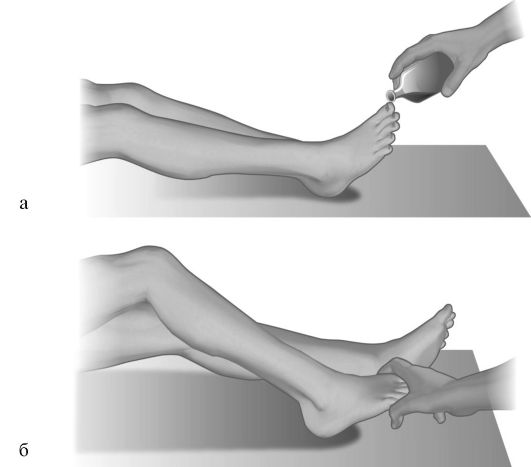
Рис. 4.15. Защитные рефлексы.
а - эфирный метод, б - метод Бехтерева-Мари-Фуа.
ти в связи с возникающим при этом раздражении проприорецепторов. Так, при пассивном подошвенном сгибании пальцев стопы парализованной ноги (рис. 4.15а, б) возникает подтягивание ноги в связи со спонтанным сгибанием ее в тазобедренном и коленном суставах (рефлекс Бехтерева-Мари-Фуа). При защитном бедренном рефлексе Ремака в ответ на штриховое раздражение кожи передней поверхности бедра возникает подошвенное сгибание стопы и пальцев. Защитные рефлексы могут возникать и при случайных раздражениях тела: прикосновении простыни, движении воздуха и т.д. В тех случаях, когда раздражитель остается незамеченным, такие рефлексы нередко называют «спонтанными» защитными рефлексами. Защитные рефлексы, как правило, возникают при сочетанном поражении корково-спинномозговых проводящих путей и экстрапирамидно-спинномозговых полисинаптических связей, бывают особенно отчетливыми при поражении пирамидных и экстрапирамидных путей на спинномозговом уровне. Уровень, ниже которого раздражение тканей ведет к возникновению защитных рефлексов, может способствовать выявлению нижнего полюса патологического очага в спинном мозге.
Патологические синкинезии. Синкинезиями, или содружественными движениями, называются непроизвольные движения, присоединяющиеся к произвольным. Синкинезии могут быть физиологическими и наблюдаются у здоровых людей. Так, сжатию кисти в кулак обычно сопутствует разгибание в лучезапястном суставе, при ходьбе возникают сопутствующие движения рук (хейрокинез) и т.д.
При центральных парезах и параличах вследствие ослабления тормозного влияния на рефлекторные дуги, замыкающиеся на спинномозговом уровне, возникают предпосылки к развитию патологических синкинезий, которые могут быть глобальными, или спастическими, координационными и имитационными.
Глобальные, или спастические, синки- незии - наблюдаются при спастических гемиплегиях. Попытки к активному движению больными конечностями или резкое напряжение мышц здоровой стороны тела вызывает непроизвольное сгибание в парализованной руке и разгибание - в ноге. Глобальные синкинезии в парализованной части тела могут возникать при общих эмоциональных реакциях, а также при зевоте, кашле, чиханье.
Координационные синкинезии - движения, которые больной не может выполнить изолированно, но при сложном двигательном акте они возникают как дополнительные и тогда не могут быть произвольно задержаны. Примером координационной синкинезии может быть симптом Раймиста, заключающийся в

Рис. 4.16. Поза Вернике-Манна при центральном гемипарезе слева.
непроизвольном отведении и приведении парализованной ноги при попытке обследующего привести или отвести здоровую ногу больного, лежащего на спине и оказывающего при этом сопротивление обследующему. Другой пример координационной синкинезии - большеберцовый феномен Штрюмпеля - заключается в том, что у больного, который не мог из-за центрального пареза ноги разгибать стопу, при попытке согнуть в колене больную ногу, преодолевая при этом сопротивление обследующего, возникает непроизвольное разгибание стопы, а иногда и большого пальца.
Имитационные синкинезии - наблюдаются редко, обычно в случаях массивного поражения пирамидных и экстрапирамидных путей. При них в парализованной конечности возникают непроизвольные движения, тождественные тем, которые произвольно совершаются в симметричной здоровой конечности, например сгибание и разгибание пальцев, пронация и супинация предплечья и т.п.
4.6. ПРИЗНАКИ ПОРАЖЕНИЯ РАЗЛИЧНЫХ ОТДЕЛОВ ОСНОВНОГО ДВИГАТЕЛЬНОГО ПУТИ
Поражение различных отделов основного двигательного пути, состоящего из центральных и периферических нейронов и обеспечивающего возможность произвольных движений, имеет свои особенности, выявление которых способствует уточнению топики патологического очага.
• Поражение двигательной зоны коры большого полушария. Двигательная зона коры занимает предцентральную (переднюю центральную) извилину, главным образом поля 4 и 6, по Бродману, ее продолжение на медиальной поверхности полушария - парацентральную дольку, а также прилежащие территории лобной доли - так называемую предцентральную область (поле 8) и участок теменной доли (поля 5 и 7), а также поля 23с и 24с коры поясной извилины. Ввиду больших размеров площади двигательной зоны коры тотальное ее разрушение встречается редко. Обычно имеет место ее частичное поражение, что ведет к развитию двигательных расстройств в той части противоположной половины тела, которая проецируется на пораженный участок коры. Поэтому при корковой локализации патологического очага характерно развитие двигательных расстройств в ограниченной части противоположной половины тела: обычно они проявляются в форме монопареза или моноплегии. Поскольку на двигательную зону коры противоположная половина тела проецируется в перевернутом виде, расстройство функции, например верхних отделов правой предцентральной извилины, ведет к возникновению двигательных расстройств в левой ноге, а поражение нижних отделов левой двигательной зоны коры приводит к центральному парезу мышц правой половины лица и языка. Если патологический очаг расположен на уровне центральных извилин в межполушарной щели, например опухоль, растущая из большого серповидного отростка (фалькс-менингиома), пораженными могут оказаться примыкающие к серповидному отростку парацентральные дольки обоих полушарий, что ведет к развитию центрального нижнего парапареза, обычно в сочетании с нарушением контроля за тазовыми функциями.
В случаях раздражения двигательной зоны коры большого полушария в мышцах соответствующей ей части противоположной половины тела могут
возникать судорожные пароксизмы, что характерно для фокальной эпилепсии джексоновского типа. Судороги эти обычно не сопровождаются расстройством сознания, но они могут распространяться на прилежащие части тела, иногда переходя во вторичный генерализованный судорожный припадок, который, начинаясь как очаговый, трансформируется в большой судорожный припадок с нарушением сознания. Если в патологический процесс вовлекается и прилежащая к пораженному участку передней центральной извилины зона задней центральной извилины, в части противоположной половины тела - мышцы которой находятся в состоянии пареза или паралича, возможны приступы парестезии - сенситивные джексоновские припадки, часто - гипестезия, при этом в большей степени нарушается проприоцептивная чувствительность и сложные виды чувствительности. При джексоновской эпилепсии во время припадка возможно сочетание локальных судорог и парестезий в определенной части тела на стороне, противоположной патологическому очагу.
Поражение дополнительной двигательной зоны в верхней теменной дольке (поля 5 и 7, по Бродману) может обусловить так называемый теменной парез на ограниченной территории противоположной половины тела, который обычно не сопровождается значительным повышением мышечного тонуса.
• Поражение лучистого венца. Лучистый венец представляет собой подкорковое белое вещество мозга, состоящее из аксонов нервных клеток, несущих импульсы в афферентном и эфферентном направлениях. При локализации патологического очага в лучистом венце на противоположной стороне обычно возникает центральный гемипарез, иногда в сочетании с гемигипестезией. Нарушения функции в различных участках противоположной половины тела выражены в разной степени, что зависит от того, какая часть лучистого венца оказалась вовлеченной в патологический процесс.
• Поражение внутренней капсулы. Во внутренней капсуле нервные волокна располагаются компактно, поэтому небольшой патологический очаг в зоне колена и двух передних третей переднего бедра внутренней капсулы может обусловить развитие центральной гемиплегии или центрального гемипареза на противоположной стороне. При более обширном патологическом процессе, распространяющемся на все заднее бедро внутренней капсулы, гемиплегия или гемипарез может сочетаться с возникающими на той же стороне гемианестезией и гемианопсией (выпадение гомонимных половин полей зрения), т.е. развивается так называемый синдром трех «геми». Остро возникающее поражение внутренней капсулы часто развивается при геморрагическом инсульте, проявляющемся медиальной внутримозговой гематомой.
При центральном гемипарезе на руке обычно в большей степени страдают мышцы, отводящие плечо, разгибатели и супинаторы предплечья, разгибатели кисти и пальцев, а на ноге - сгибатели бедра, разгибатели стопы и пальцев, что ведет к развитию у больных в период восстановительной фазы своеобразной позы, известной как поза Вернике-Манна (рис. 4.16). В связи с тем что в руке преобладает тонус мышц-сгибателей, а в ноге - разгибателей, рука, находящаяся в состоянии пареза, оказывается приведенной к туловищу и согнутой в локтевом суставе, кисть ее пронирована, а паретичная нога выпрямлена и кажется несколько длиннее здоровой ноги. Походка больных с центральным гемипарезом при этом оказывается своеобразной. При ходьбе выпрямленная паретичная нога больного совершает движения по дуге, рука на стороне ге-
мипареза остается согнутой и прижатой к туловищу. В таких случаях иногда говорят, что больной «рукой просит, а ногой косит».
• Поражение ствола мозга. При одностороннем поражении различных отделов ствола мозга (средний мозг, мост, продолговатый мозг) характерно развитие альтернирующих (перекрестных) синдромов, при которых на стороне патологического очага возникают признаки поражения отдельных черепных нервов, а на противоположной стороне - гемипарез или гемиплегия по центральному типу, иногда - гемигипестезия. Вариант альтернирующего синдрома в таких случаях определяется уровнем и распространенностью поражения ствола. При двустороннем поражении ствола мозга функции черепных нервов могут быть нарушены с обеих сторон, при этом характерны псевдобульбарный или бульбарный синдромы, тетрапарез, расстройства чувствительности по проводниковому типу.
• Поперечное поражение половины спинного мозга - синдром Броун-Сека- ра. При поражении половины поперечника спинного мозга латеральный пирамидный путь вовлекается в патологический процесс ниже уровня его перекреста. В связи с этим центральный парез или паралич, возникающий ниже уровня поражения спинного мозга, развивается на стороне патологического очага. Двигательные расстройства при этом обычно сочетаются с нарушением чувствительности по проводниковому типу. В таких случаях на стороне патологического процесса оказывается нарушенной проприоцептивная чувствительность, а на противоположной стороне - поверхностная (болевая и температурная).
• Полное поперечное поражение спинного мозга в верхнем шейном отделе (СI- CIV). При двустороннем поражении спинного мозга в верхнем шейном отделе возникает центральная тетраплегия, при этом сочетанное поражение с обеих сторон перекрещенных и неперекрещенных пирамидных путей ведет к тому, что страдают и мышцы туловища, включая дыхательную мускулатуру. Кроме того, в таких случаях ниже уровня расположения патологического очага обычно имеют место нарушения всех видов чувствительности по проводниковому типу, а также тазовые и трофические расстройства.
• Поражение шейного утолщения спинного мозга (CV-ThII). Поражение шейного утолщения спинного мозга также ведет к развитию тетраплегии в сочетании с нарушениями всех видов чувствительности по проводниковому типу ниже уровня патологического очага с тазовыми и трофическими расстройствами. Однако в связи с поражением шейного утолщения спинного мозга паралич или парез рук развивается по периферическому типу, тогда как паралич туловища и ног - по центральному.
• Поражение грудного отдела спинного мозга (ThIII-ThXII). Следствием поперечного поражения грудного отдела спинного мозга является спастическая нижняя параплегия в сочетании с утратой ниже уровня локализации патологического очага всех видов чувствительности, нарушением тазовых функций и расстройством трофики тканей.
• Поражение поясничного утолщения спинного мозга (LII-SII). При поражении поясничного утолщения спинного мозга развивается нижняя параплегия по периферическому типу в сочетании с нарушением чувствительности и трофики тканей на ногах и в аногенитальной зоне, а также с тазовыми расстройствами обычно в виде недержания мочи и кала.
• Избирательное поражение клеток передних рогов спинного мозга и двигательных ядер черепных нервов. В связи с избирательным поражением тел периферических двигательных нейронов возникает периферический паралич мышц, иннервацию которых они обеспечивают, при этом раздражение отдельных еще сохранившихся периферических мотонейронов может вызывать спонтанное сокращение мышечных волокон или их пучков (фибриллярные или фасцикулярные подергивания).
Избирательное поражение периферических двигательных нейронов характерно для эпидемического детского полиомиелита и бокового амиотрофического склероза, а также для спинальных амиотрофий.
• Поражение передних корешков спинного мозга. При поражении передних корешков спинного мозга характерны периферические параличи мышц, входящих в состав миотомов, одноименных пораженным корешкам.
• Поражение спинномозговых нервов. Следствием поражения спинномозговых нервов являются двигательные расстройства по периферическому типу в мышцах, иннервируемых аксонами мотонейронов, входящих в состав этих нервов, а также расстройства чувствительности (боль, гипалгезия, анестезия) в одноименных им дерматомах. Там же возможны вегетативные, в частности трофические, расстройства.
• Поражения нервных сплетений. Поражение нервного сплетения обусловливает развитие двигательных расстройств (паралич или парез) по периферическому типу, обычно в сочетании с нарушениями чувствительности и трофики в зоне иннервации периферических нервов, берущих начало из пораженного сплетения, или его части.
• Поражение периферического нерва. При поражении периферического нерва возникает периферический паралич или парез иннервируемых им мышц, обычно в сочетании с расстройством всех видов чувствительности и трофическими нарушениями в зоне иннервации пораженного нерва (см. главу 8).