Общая неврология А.С. Никифоров, Е.И. Гусев - 2007г. - 720 с
|
|
|
|
ГЛАВА 2 ВВЕДЕНИЕ В КЛИНИЧЕСКУЮ НЕВРОЛОГИЮ
2.1. ОБЩИЕ ПОЛОЖЕНИЯ
Нервная система обеспечивает интеграцию структур, составляющих организм, регуляцию происходящих в них метаболических, трофических процессов и поддержание таким образом относительного постоянства внутренней среды организма (гомеостаза). Кроме того, нервная система осуществляет контроль за состоянием и деятельностью отдельных органов и тканей, благодаря наличию обратных связей с ними обеспечивает рациональный уровень их функциональной активности. С помощью нервной системы осуществляется взаимодействие организма с внешней средой, получение информации о воздействующих на него внешних факторах, которые могут непосредственно контактировать с организмом или находиться на некотором расстоянии от него. На основании этой информации под влиянием нервных структур в организме возникают реакции, направленные на его адаптацию к окружающей среде. Нервная система осуществляет контроль за положением частей тела в пространстве и через посредство опорно-двигательного аппарата обеспечивает все виды двигательных актов и их коррекцию в процессе выполнения. Нервная система осуществляет, наконец, все виды психической деятельности: сознание, эмоциональное реагирование, интеллектуально-мнестические процессы, включая высшие психические функции. От состояния нервной системы, а также особенностей воспитания и обучения зависят особенности человеческой личности.
2.2. НЕКОТОРЫЕ ИСТОРИЧЕСКИЕ АСПЕКТЫ ИЗУЧЕНИЯ НЕРВНОЙ СИСТЕМЫ
Всегда ли люди понимали значимость нервной системы, ее роль в развитии и существовании человека, в обеспечении его физического и психического здоровья? Всегда ли осознавали, что особенности личности человека, его душевного состояния, самосознание и понимание происходящего вокруг, возможность чувствовать и действовать зависят от состояния нервной системы, в частности от функций головного мозга? Вовсе нет. К примеру, в древние времена отношение к мозгу у египтян проявлялось при бальзамировании тел умерших. По описаниям древнегреческого историка Геродота известно, что при бальзамировании тел умерших египтян важные для человека органы, в
частности органы брюшной полости, старались сохранить, а с мозгом обращались просто: введенным через ноздри металлическим предметом разрушалась решетчатая кость, после этого мозг, который рассматривался как скопление носового секрета, выделяемого в случаях насморка, из полости черепа вымывали водой. Чувства и разум в Древней Ассирии, Древней Греции, Древнем Риме связывали главным образом с состоянием печени, диафрагмы, сердца.
Признается, что первым о мозге как о вместилище души писал еще в VI в. до н.э. Алкмеон из Кретоны. Гиппократ (460-около
Однако длительно существовало и мнение, которое в свое время разделяли такие великие философы древности, как Аристотель и Платон, о том, что ведущая роль в обеспечении психической жизни человека осуществляется его сердцем. Уже в начале XVII в. великий В. Шекспир устами одного из героев пьесы «Три веронца» задавал вопрос: «Скажи мне, где мечты начало? Мозг, сердце ль жизнь ей даровало?» Да и в настоящее время мы, оценивая душевные качества человека, нередко характеризуем его как сердечного или бессердечного, пользуемся такими выражениями, как «от всего сердца», «легко на сердце» и т.п. В XVII в. Рене Декарт (Descartes R., 1596-1650) высказывался о том, что живые организмы можно сравнивать с механическими автоматическими устройствами, которые управляются мозгом, при этом Р. Декарт признавал, что материально существующий мозг человека в отличие от мозга животных одушевлен. В связи с этим Р. Декарт признается основателем философского направления дуализма. Он ввел представление о дуге (дуга Декарта), которая в последующем стала именоваться рефлекторной дугой. Рефлекс (от лат. reflexus - отражение) - реакция организма на раздражение. Человека Р. Декарт рассматривал как единение телесного механизма с душой, обладающей мышлением и волей, расположенной в эпифизе. Он признавал психическую деятельность, мысль, знание - основой существования («мыслю, следовательно, существую»).
В
лексов. Через три года В.М. Бехтерев (1857-1927) описал на базе клинических наблюдений наличие аналогичных рефлексов у человека и назвал их сочетательными. На основании этого открытия он создал научное направление - объективную психологию, или рефлексологию, основные идеи которой в настоящее время активно разрабатываются наукой, именуемой физиологической психологией.
Во второй половине ХIX в. и в ХХ в. получил развитие комплекс нейронаук (нейроморфология, нейрофизиология, нейрохимия, нейропсихология, нейрогенетика), который сейчас нередко именуется нейробиологией.
В ХХ в. особое внимание обращается на изучение биохимических основ, происходящих в нервной ткани физиологических процессов, в частности на углубленное познание сущности медиаторных систем и их роли в нейрофизиологии в условиях нормы и патологии. Развитие этого научного направления особенно перспективно, так как способствует пониманию деятельности нервной системы, патогенеза ее заболеваний, способствует раскрытию возмож- ностей их лечения.
2.3. ОСНОВЫ ФИЛОГЕНЕЗА НЕРВНОЙ СИСТЕМЫ
Одним из важнейших свойств живой материи является раздражимость. Ее совершенствование привело к специализации некоторых структур простейших живых организмов, отдельных клеточных элементов и обусловило развитие нервной ткани.
Раздражимостью обладают уже простейшие одноклеточные организмы, в частности амеба. Раздражимость у амебы имеет поверхностно-внутреннюю полярность. Луч света раздражает поверхность амебы, что ведет к ее перемещению из освещенной зоны в тень, осуществляемому благодаря повторяющимся изменениям формы организма (метод таксиса). Жгутиковым одноклеточным организмам свойственна уже определенная специализация участков ее поверхности и обусловленная этим переднезадняя (апико-базилярная) полярность.
Многоклеточные простейшие живые существа, например гидра, содержат дифференцированные, имеющие повышенную раздражимость клетки; они находятся в поверхностном слое организма, а их отростки направлены внутрь него и образуют асинаптическую сеть (синцитий). Раздражение поверхности гидры ведет к возникновению у нее слабо дифференцированных двигательных реакций.
Более сложно нервная система организована у червей, в теле которых продольно оси находятся две параллельные цепочки нервных узлов (ганглиев), при этом каждая их пара, расположенная в плоскости одного и того же поперечника, обеспечивает чувствительность и сокращение тканей соответствующего ей фрагмента (сегмента) тела червя. По сути, на данном этапе развития живого проявляется метамерное строение организма. Метамеры (сегменты тела) обладают определенной автономией. В связи с этим части перерезанного лопатой дождевого червя способны к самостоятельному существованию. У членистоногих (например, у пчелы) наряду с парой параллельно расположенных цепочек ганглиев имеется интегрирующий их функцию непарный надглоточный (головной) узел, без которого живое существо оказывается нежизнеспособным.
У позвоночных нервная система развивается из медуллярной трубки, образующейся из наружного (эктодермального) зародышевого листка, при этом у
прародителя позвоночных - ланцетника - наблюдается метамерность строения нервной трубки и всего организма. У более сложно организованных позвоночных в процессе их эволюции происходит утолщение переднего конца нервной (медуллярной) трубки, образование так называемых нервных пузырей, из которых происходит формирование головного мозга. Головной мозг в процессе формирования более высоких уровней развития позвоночных (рыбы, амфибии, рептилии, птицы, млекопитающие) имеет все более сложное строение, при этом прогрессирующая цефализация нервной системы сочетается с метамерией, которая отчетливо выражена у всех млекопитающих, в том числе и у человека.
2.4. ОНТОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ
В процессе онтогенеза человека на дорсальной поверхности наружного (эктодермального) зародышевого листка дифференцируется нервная, или мозговая, борозда (медуллярный желобок). Она постепенно углубляется, края ее сближаются, образуя нервную, или мозговую, трубку. В трубке герминативный (ростковый, зародышевый) клеточный слой, находящийся в наружном (эктодермальном) зародышевом листке снаружи, оказывается внутри.
Нервная трубка развивается неравномерно. Передний отдел ее постепенно утолщается и уже на 4 нед развития зародыша здесь образуется три мозговых пузыря: передний - prosencephalon, средний - mesencephalon, задний - rombencephalon.
В дальнейшем (на 6 нед) передний и задний мозговые пузыри делятся на две части. Таким образом, стадия 3 пузырей сменяется стадией 5 пузырей (рис. 2.1), из которых в последующем и происходит формирование головного мозга (рис. 2.2), при этом в процессе онтогенеза из переднего, точнее - из
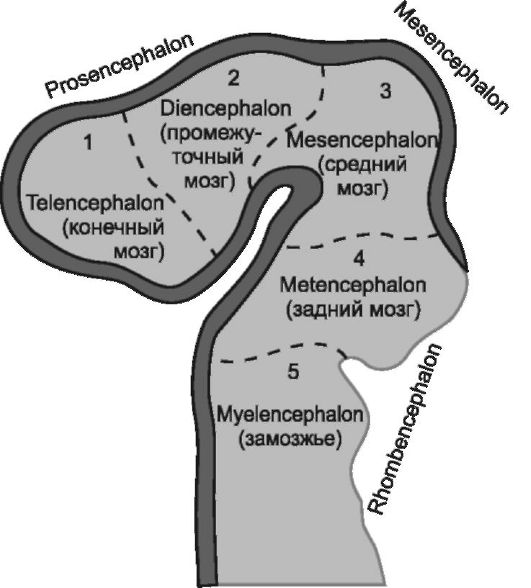
Рис. 2.1. Развитие головного мозга в онтогенезе. Стадия пяти пузырей.
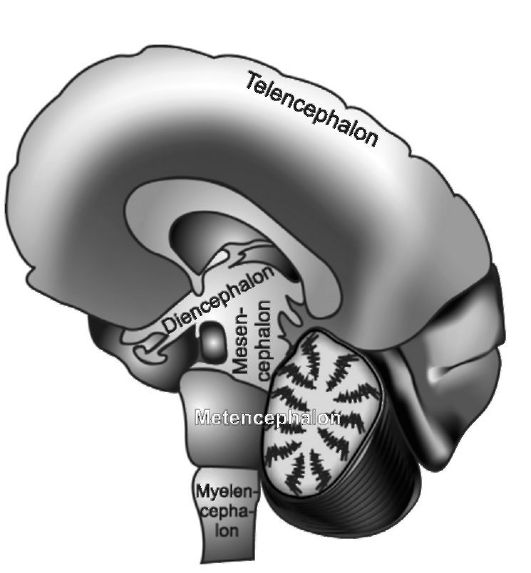
Рис. 2.2. Головной мозг человека. Сагиттальный срез.
первичного переднего мозгового пузыря, выделяется и быстро увеличивается вторичный передний мозговой пузырь, или конечный мозг (telencephalon). Из него формируются главным образом парное образование - большие полушария, а полость его превращается в боковые желудочки мозга (левый признается первым, правый - вторым); каждый из них имеет центральную часть и три рога, при этом центральная часть бокового желудочка оказывается в глубине теменной доли большого полушария, передний рог - в лобной доле, нижний рог - в височной доле, задний рог - в затылочной. Производными вторичного переднего мозгового пузыря оказываются и периферические структуры обонятельного анализатора. Первичный передний мозговой пузырь становится источником формирования главным образом структур промежуточного мозга (diencephalon), а полость его преобразуется в непарный III мозговой желудочек. С каждой стороны промежуточного мозга в дальнейшем вырастает по глазному пузырю, из которого формируются зрительные тракты, зрительные нервы и сетчатка. Из среднего мозгового пузыря образуется средний мозг (mesencephalon); полость его превращается в водопровод мозга. Из заднего (rombencephalon) мозгового пузыря создаются два отдела. Один из них - задний мозг (metencephalon) идет на формирование моста мозга и мозжечка, а из оставшейся части ромбовидного мозга (замозжья) формируется продолговатый мозг (myelencephalon, medulla oblongata, bulbus). Полость ромбовидного мозга при этом превращается в IV мозговой желудочек, дно которого имеет форму ромба (ромбовидная ямка).
В процессе дифференциации зародышевых (герминативных) клеток, составляющих внутренний слой медуллярной трубки, часть их, проходя стадию нейробласта, превращается в нейроциты (нейроны) - зрелые нервные клетки. Другая часть зародышевых клеток, пройдя определенные стадии созревания, трансформируется в зрелые глиальные элементы (астроциты, олигодендроциты, эпендимоциты).
К моменту рождения ребенка нервные клетки достигают зрелости и уже неспособны к делению. В связи с этим в дальнейшем их число не увеличива- ется, и если нервная ткань в процессе онтогенеза развивается нормально, в последующем не возникает опухолей, состоящих из нейронов. Глиальные же клетки во всех стадиях своего развития способны к митотическому делению, и в течение жизни человека в мозге его могут быть обнаружены глиальные элементы разной степени зрелости. Из подвергшихся мутации глиальных клеток возможно формирование опухолей практически в любом возрасте человека. Это истинные мозговые опухоли, известные как глиальные опухоли, или глиомы. В зависимости от степени зрелости составляющих их глиальных клеток глиомы могут дифференцироваться на незрелые (злокачественные) и зрелые (доброкачественные).
В мозговых желудочках главным образом железистыми клетками сосудистых ворсинчатых (хориоидальных) сплетений осуществляется секреция цереброспинальной жидкости (ЦСЖ) или ликвора. Другим источником продукции ЦСЖ признаются эпителиальные клетки и клетки интерстициальной соединительной ткани, также находящиеся в желудочках мозга. Полости мозговых пузырей, трансформируясь в желудочки мозга, сохраняют связи друг с другом, при этом парные боковые желудочки соединяются с непарным III мозговым желудочком через межжелудочковые отверстия (отверстия Монро). III мозговой желудочек через водопровод мозга (aqueductus cerebri, сильвиев водопровод) соединяется с образующимся из полости ромбовидного мозга IV мозговым желудочком.
К 8 нед развития плода возникает переполнение ЦСЖ желудочковой системы формирующегося мозга, т.е. появляется тенденция к развитию внутренней гидроцефалии, при этом в желудочковой системе повышается давление ЦСЖ, что способствует раскрытию апертур (отверстий) IV мозгового желудочка - непарной срединной апертуры (отверстие Мажанди) и парной латеральной апертуры (отверстие Люшки). После раскрытия этих апертур ЦСЖ получает возможность выхода в мозжечково-мозговую (большую) цистерну и боковые цистерны моста, при этом давление ЦСЖ в желудочках головного мозга нор- мализуется, а центральный канал спинного мозга, формирующегося из той части нервной трубки, которая не участвовала в развитии мозговых пузырей, постепенно запустевает и редуцируется.
2.5. НЕРВНАЯ ТКАНЬ
Собственно нервную ткань составляют нервные клетки (нейроны) с их отростками и клетки глии. Сосуды и оболочки мозга имеют мезодермальное происхождение.
2.5.1. Историческая справка
Успехи
оптики, приведшие к созданию светового микроскопа, и достижения химии,
позволившие получить анилиновые и другие красители, привели к
возникновению и развитию гистологии. Это позволило во второй половине
XVIII в. обнаружить в мозге нервные волокна (Монро А., 1787; Фонтан Ф., 1781; Рейли И., 1796); в 30-х годах XIX в. были описаны и нервные клетки (Эренберг Х., в 1833; Пуркинье Я., Шванн Т., в 1838). В
В
В
Уже при макроскопическом осмотре мозга на разрезе выявляется неоднородность составляющей его ткани. В головном и спинном мозге выделяются участки серого и белого вещества. Серое вещество - места скопления тел нервных клеток и протоплазматической астроцитарной глии. Белое вещество состоит в основном из нервных волокон и окружающих их глиальных клеток - главным образом олигодендроцитов и волокнистых астроцитов, при этом белый цвет белого вещества мозга обусловлен цветом миелиновой оболочкой нервных волокон, формирующих проводящие проекционные пути, комиссуральные и ассоциативные связи.
2.5.2. Нервная клетка
Нервная клетка (нейрон), признающаяся основной структурной и функциональной единицей нервной системы (рис. 2.3), принципиально отличается от клеток, составляющих другие органы и ткани.
Но функциональная самостоятельность нейрона условна. Так, например, гибель периферических двигательных нервных клеток, расположенных в передних рогах спинного мозга, может лишить смысла активность сопряженных с ними корковых моторных нейронов, так как прерывается путь между двигательными клетками коры и исполнительным органом - в данном случае с определенными поперечнополосатыми мышцами (ситуация, возникающая, к примеру, при эпидемическом полиомиелите). Особенности функции нейронов сказываются на их форме (рис. 2.4) и составе содержащихся в них цитоплазматических органелл.
Каждая нервная клетка (нейрон) имеет тело (перикарион) и отростки. Один из них - маловетвящийся и обычно самый длинный - аксон (нейрит); другие, короткие, имеющие много ответвлений, - дендриты, в типичных случаях характеризующиеся древовидным строением. Форма и размеры нейронов ва- риабельны. По форме тел их делят на звездчатые, корзинчатые, пирамидные и пр. Размеры тел нейронов варьируют от 4 до 150 мкм в диаметре. Нейроны с большим количеством отростков называют мультиполярными, их большинство. Кроме того, существуют биполярные нейроны с аксоном и одним дендритом, находящиеся главным образом в составе обонятельной, зрительной и слуховой систем, и так называемые псевдоуниполярные клетки, расположенные в спинальных ганглиях и их аналогах, находящихся в составе черепных нервов. Псевдоуниполярные клетки также имеют по два отростка - аксон и дендрит1, но проксимальные части этих отростков прочно прилежат друг к другу, что на препаратах, импрегнированных серебром, создает впечатление униполярности клеток. Принято считать, что только в головном мозге человека насчитывается до 10 млрд нейронов. Возможна классификация нервных клеток и по длине аксонов [клетки с длинными аксонами, выходящими за пределы данного скопления клеток (ядра), называют клетками Гольджи I, клетки с короткими аксонами - клетки Гольджи II]. Классифицируются нейроны также и по их функции: сенсорные, моторные, ассоциативные. Особенно значимой для понимания многих клинических проблем является, пожалуй, классификация
1 У псевдоуниполярных клеток, тела которых расположены в спинномозговых ганглиях или их аналогах на краниальном уровне, дендриты обычно длинные, при этом они имеют строение и функциональные особенности, характерные для аксонов.
нейронов по характеру вырабатываемого в них нейромедиатора (нейротрансмиттера). По этому принципу нейроны дифференцируются на холинергические, серотонинергические, адреналинергические, ГАМКергические, допаминергические и т.п.
Тело клетки и ее отростки покрывает непрерывная сдвоенная мембрана (невролемма), представляющая собой липопротеиновый комплекс и выполняющая разграничительную и транспортные функции. Через нее осуществляется пассивный транспорт воды и некоторых низкомолекулярных веществ, а также перенос ионов и органических молекул против градиента концентрации с затратой энергии, возникающей в основном при расщеплении молекул аденозинтрифосфата (АТФ). Последнее свойство мембраны нервной клетки обеспечивает поддержание в ней постоянного мембранного потенциала покоя, а также возникновение возбуждающего или тормозного постсинаптического потенциалов (ВПСП или ТПСП), определяющих формирование нервного импульса в связи с резким изменением проницаемости клеточных мембран для содержащих биоэлектрический заряд ионов.
Нейрон (рис. 2.3) не только обеспечивает проведение импульсов, но и синтезирует белки, липиды, углеводы, а также нейромедиаторы (нейротрансмиттеры). Некоторые нейроны к тому же продуцируют гормоны (вазопрессин, окситоцин, антидиуретический гормон, рилизинг-факторы). В теле нейрона находится цитоплазма и ядро с расположенным в нем ядрышком, а также базофильные органоиды (органеллы): пластинчатый комплекс (комплекс Голь-
джи), митохондрии, лизосомы, имеющиеся и в соматических клетках, и, кроме того, специфическое для нервных клеток базо- фильное вещество Ниссля, нейрофибриллы и нейротрубочки. Включениями в цито- плазме нервных клеток могут быть гранулы гликогена, каротиноидов, пигмента и пр.
Ядро нервной клетки относительно большое, слабо окрашивается, содержит много дезоксирибонуклеиновой кислоты (ДНК); его окружает двухслойная мембрана с множеством пор, через которые совершается обмен между цитоплазмой и заполняющей ядро нуклеоплазмой. В ядре происходит синтез рибонуклеиновой кислоты (РНК), которая проникает из него в плазму и участвует в формировании органелл клетки. Заключенное в ядре ядрышко представляет собой лишенное мембраны меняющееся по форме, размеру и химическому составу образование, состоящее из РНК, белков, липидов и находящегося внутри слоя ДНК. Изменчивость ядрышка отражает его высокую физиологическую активность.
Пластинчатый комплекс Гольджи (липохондрии), как и ядрышко, в процессе жизнедеятельности клетки подвергается циклическим изменениям. Он состоит из
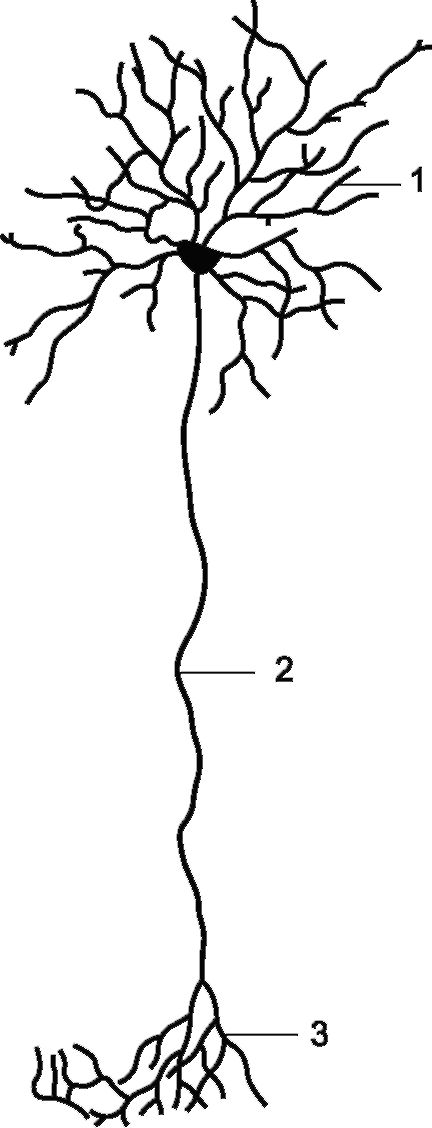
Рис. 2.3. Нейрон, его отростки: 1 - дендриты, 2 - аксон, 3 - разветвления аксона.
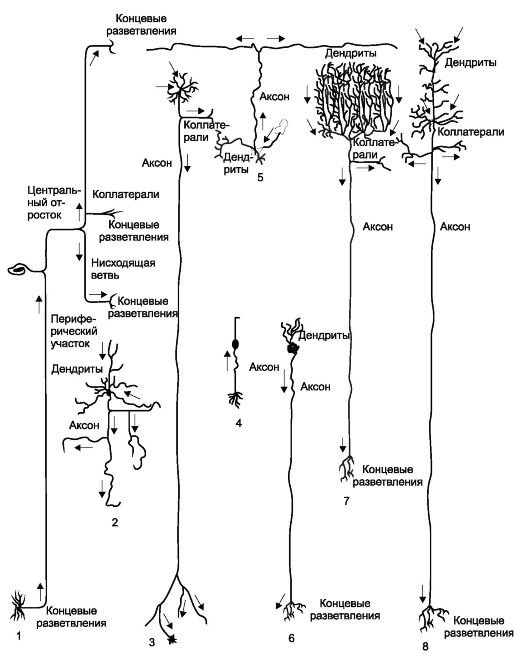
Рис. 2.4. Некоторые виды нейронов (по Бейли).
1 - периферический чувствительный нейрон; 2 - короткоаксонный нейрон типа Гольджи II; 3 - периферический мотонейрон; 4 - обонятельный нейрон; 5 - клетка зернистого слоя мозжечка; 6 - нейрон симпатического узла; 7 - клетка Пуркинье; 8 - пирамидная клетка Беца.
Стрелки показывают направление перемещающихся по клетке нервных импульсов.
плотно упакованных двухслойных мембран и гранул, содержит липиды, фосфатиды, мукополисахариды и участвует в синтезе углеводных полимеров, гормонов.
Митохондрии имеют палочковидную форму, рассеяны по всей цитоплазме нейрона. Особенно много их в наиболее активных частях нейрона: в его теле и окончаниях ветвлений аксона (в пресинаптических пуговках). Митохондрии содержат дыхательные ферменты и играют важную роль в осуществлении дыхания клетки, обеспечивая процесс окислительного фосфорилирования (окисление углеводов и жиров) и участия в гликолизе. Основная функция митохондрий связана с образованием богатой энергией АТФ.
Лизосомы имеют вид вакуолей, содержат гидролитические ферменты (протеиназы, нуклеазы, глюкозидазы, фосфатазы, липазы), расщепляющие различ- ные биополимеры. Основная функция лизосом - расщепление биологических макромолекул внутриклеточного и внеклеточного происхождения на более
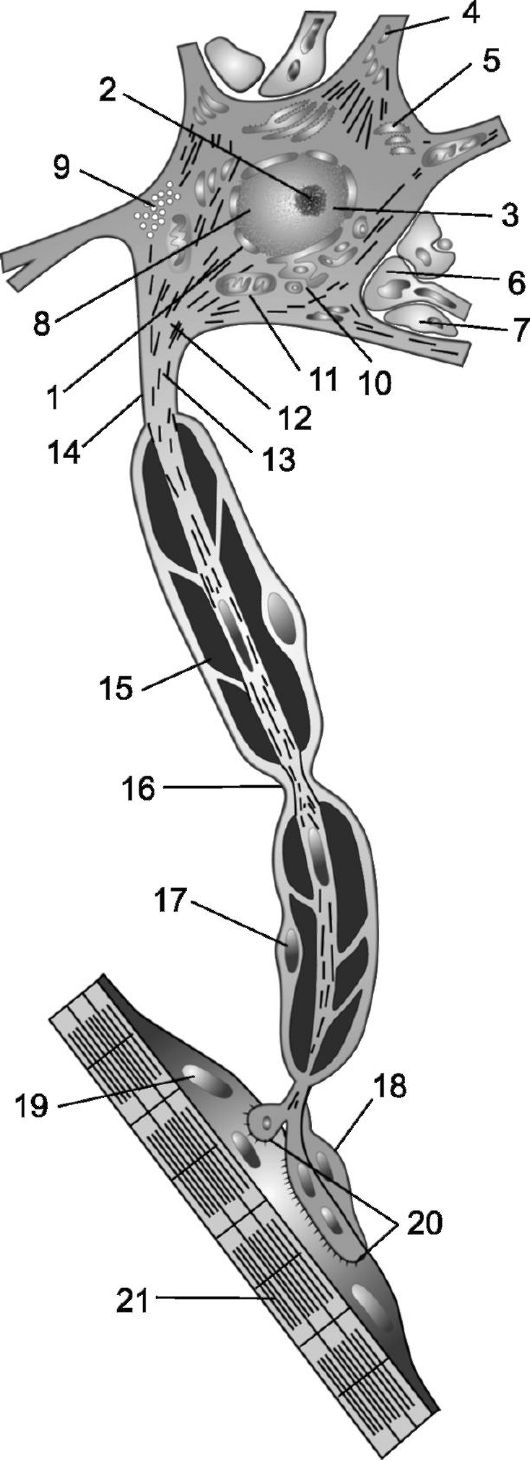
Рис. 2.5. Компоненты периферического мотонейрона [По Дж. Шаде и Д. Форду]. 1 - ядро; 2 - ядрышко; 3 - сателлит ядрышка; 4 - дендрит; 5 - эндоплазматическая сеть с гранулами РНК (вещество Ниссля); 6 - синапс; 7 - ножка астроцита; 8 - гранулы ДНК; 9 - липофусцин; 10 - аппарат Гольджи; 11 - митохондрия; 12 - аксонный холмик; 13 - нейрофибриллы; 14 - аксон; 15 - миелиновая оболочка; 16 - перехват Ранвье; 17 - ядро леммоцита; 18 - леммоцит в области нервно-мышечного синапса; 19 - ядро мышечной клетки; 20 - нервно-мышеч- ное соединение; 21 - мышца.
простые микромолекулы, которые впоследствии могут быть утилизированы в результате происходящего в нейроне биосинтеза более сложных соединений.
Базофильное вещество Ниссля (тигроид) составляют базофильные трубчатые структуры и гранулы из РНК, соединенной с белком (РНК-Б). Глыбки вещества Ниссля рассеяны по всей цитоплазме нейрона и участвуют в формировании ее эндоплазматической сети. Измельчаясь, они проникают в его дендриты, однако в аксоне и в той части тела клетки, от которой начинается аксон, базофильное вещество отсутствует. Признано, что оно осуществляет синтез белков, происходящий под контролем генетического аппарата ядра. Количество базофильного вещества в нервной клетке изменчиво и зависит от ее функционального состояния.
Среди включений в нервной клетке особое значение имеет нейромеланин, скопление которого наблюдается в черном веществе (substantia nigra) среднего мозга. Нейромеланин - необходимое звено в процессе образования катехоламинов.
2.5.3. Нервные волокна
Нервное волокно - в большинстве случаев аксон нервной клетки, состоит из осевого цилиндра, покрытого миелиновой оболочкой. Толщина аксона составляет от 0,3 до 20 мкм и зависит главным образом от толщины миелиновой оболочки, которая образуется за счет многократного обертывания фрагментов осевого цилиндра «избыточной» оболочкой прилежащих к нему глиальных клеток, в центральной нервной системе - олигодендроцитов, в периферической нервной системе - шванновских клеток (леммоциты). Образуемая таким образом миелиновая оболочка состоит из слоев, спиралеобразно окружающих осевой цилиндр аксона, число их может быть 100 и более (рис. 2.6). В состав миелиновой оболочки входят холестерин, фосфолипиды, некоторые цереброзиды и жирные кислоты, а также белковые вещества. Между фрагментами миелиновой оболочки, каждый из которых сформирован за счет оболочки ближайшей глиальной клетки, образуются просветы - перехваты Ранвье (см. рис. 2.5). Скорость проведения по волокну нервного импульса прямо пропорциональна толщине его миелиновой оболочки и варьирует от 0,6 до 20 м/с.
Американские физиологи Г. Гассер (Gasser H.S., 1888-1963) и Дж. Эрлангер (Erlanger G., 1874-1965) в
Зная толщину миелинового волокна, можно определить скорость проведения по нему нервного импульса. Если волокно толще 1 мкм, можно пользоваться формулой: V(скорость проведения нервного импульса) = диаметр волокна (в мкм), умноженный на 6. Полученное произведение обозначается в м/с.
1Г. Гассер и Дж. Эрлангер в
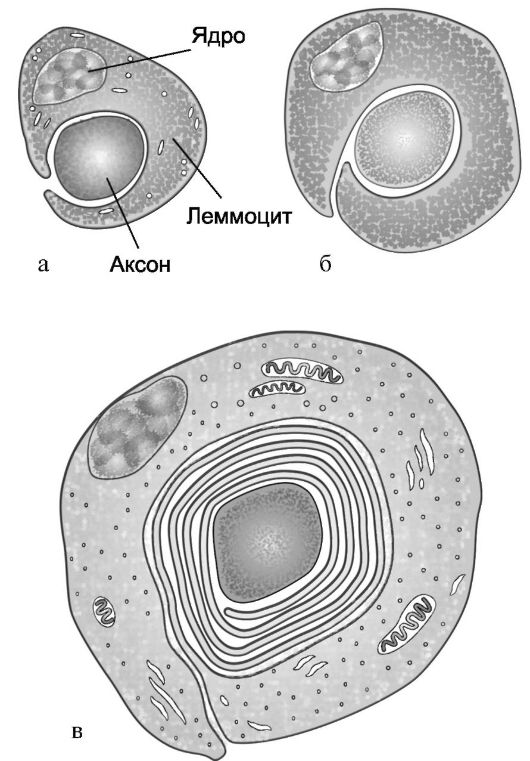
Рис. 2.6. Поперечные срезы нервного волокна на разных этапах процесса его миелинизации (аксон и леммоцит).
а, б, в - стадии формирования миелиновой оболочки.
Для определения скорости проведения импульсов по волокнам С (диаметр меньше 1 мкм) может быть применена другая формула: V = диаметр волокна (в мкм), умноженный на 2. Полученное произведение обозначается в м/с. Д. Ллойд (D. Lloyd) классифицирует аксоны по их диаметру: 1-я группа - 12-20 мкм; 2-я группа - 6-12 мкм; 3-я группа - 1-6 мкм; 4-я группа - меньше 1 мкм.
2.5.4. Аксоток
Для нейрона характерна исключительно высокая активность метаболических процессов. Биосинтез большинства макромолекул клетки осуществляется в ее теле. Отсюда они распространяются по аксону посредством ортоградного аксонального тока. Различают быстрый ортоградный аксональный ток (аксональный транспорт), или быстрый компонент аксотока, благодаря которому по аксону транспортируются молекулы белка, гликопротеиды и фосфолипиды, некоторые ферменты, необходимые для поддержания текущего метаболизма, со скоростью 200-400 мм/сут. Медленный аксональный ток (аксональный транспорт) обеспечивает перенос из тела клетки к периферии аксона материала для восстановления клеточного каркаса (микротубулярно-нейрофиламентарной сети) со скоростью 0,3-1,0 мм/сут и актиносодержащих микрофиламентов и аксоплазматического матрикса со скоростью 2-4 мм/сут.
Большинство переносимых по аксону молекул включается в метаболизм в пределах нейрона; они обеспечивают восстановление его энергетических затрат, рост и регенерацию аксонов. Часть из них может переходить в постсинаптические структуры, а также в прилежащие глиальные клетки, участвуя таким образом, в частности, и в формировании миелиновой оболочки.
Продукты происходящих в аксоне процессов метаболизма посредством ретроградного аксонального тока со скоростью 150-300 мм/сут перемещаются в тело клетки, где подвергаются дальнейшему разрушению ферментами лизосом до составных элементов, используемых при формировании вновь синтезируемых в теле нейрона макромолекул. Таким образом, ретроградный аксональный ток обеспечивает возможность осуществления в теле клетки процессов вторичной утилизации микромолекул.
2.5.5. Синаптическая передача
Место, где окончание аксона сближается с дендритом или телом следующей в нейронной цепи нервной клетки, по предложению Ч. Шеррингтона (Sherrington Ch., 1857-1952), называется синапсом (от греч. sinapto - застежка, соединение). Конечные ветвления аксона - телодендрии заканчиваются утолщением (пресинаптическая пуговка), в котором содержатся митохондрии и пузырьки с квантами медиатора (рис. 2.7). Участок невролеммы пресинаптической пуговки, особенно близко расположенный к структурам следующего нейрона, называется пресинаптической мембраной. Находящийся в непосредственной близости от него участок невролеммы последующего нейрона называется постсинаптической мембраной. Между пресинаптической и постсинаптической мембранами расположена узкая синаптическая щель (ширина ее приблизительно 200 А, или 0,02 мкм).
Когда нервный импульс доходит до пресинаптической пуговки, из расположенных в ней синаптических пузырьков в синаптическую щель выделяется квант нейромедиатора, который достигает постсинаптической мембраны и меняет ее проницаемость для находящихся вокруг положительно и отрицательно заряженных ионов, вызывая, таким образом, в расположенном по другую
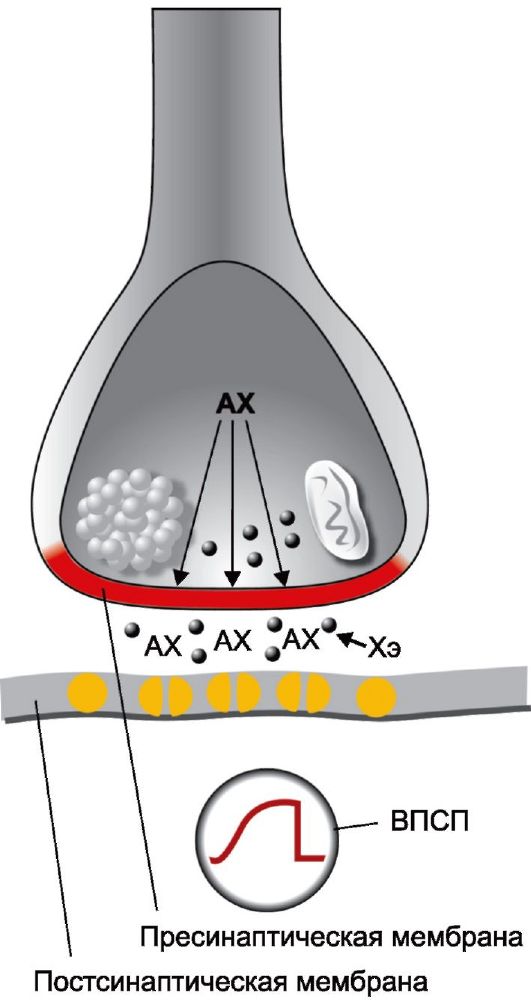
Рис. 2.7. Синаптические процессы в возбужденном синапсе [По Л. Шельцыну, 1980]. А - ацетат; Х - холин; АХ - ацетилхолин; Хэ - холинэстераза; ВПСП - возбудительный постсинаптический потенциал.
сторону синаптической щели нейроне возникновение возбуждающего или тормозного постсинаптического потенциала. В результате нейромедиатор обеспечивает химическую передачу нервного импульса через синаптическую щель и, по сути, служит посредником для передачи нервного импульса от передающего его нейрона к воспринимающему.
Выделившиеся в синаптическую щель кванты медиатора отчасти возвращаются через пресинаптическую мембрану назад (обратный захват) в пресинаптическую пуговку, отчасти медиатор разрушается в синаптической щели под влиянием определенного фермента. Например, в нервно-мышечном синапсе и в синаптическом аппарате других холинергических нейронов таким ферментом является антихолинэстераза. Функции медиаторов (нейротрансмиттеров) могут выполнять многие биологические вещества, чаще аминокислоты. По влиянию на синаптические аппараты нейротрансмиттеры могут быть разделены на возбуждающие и тормозные. К возбуждающим относится глутамат и аспартат, а к тормозным - ГАМК и глицин. Кроме того, выделяется группа нейротрансмиттеров, состоящая главным образом из моноаминов (дофамин, норадреналин, серторонин), при этом одни и те же нейротрансмиттеры могут, воздействуя на одни нейроны, оказывать возбуждающее действие, тогда как влияние их на другие нейроны может быть тормозным. Так, ацетилхолин нервно-мышечного синаптического аппарата возбуждает мышечные волокна, а ацетилхолин как медиатор стриопаллидарных нейронов обеспечивает тормозное влияние на клетки бледного шара.
Помимо трансмиттеров, на синаптическую передачу могут оказывать усиливающее или ослабляющее действие нейромодуляторы (эндорфины, соматостатин, субстанция Р) и нейрогормоны (ангиотензин, вазопрессин и др.), которые, однако, сами по себе не создают деполяризационного эффекта. Нейрогормоны попадают в кровяное русло и разносятся с кровью на большие расстояния. Их действие уступает модуляторам по темпу, но проявляется длительнее.
Синапсы
обеспечивают регуляцию потока нервных импульсов и определяют проведение
их всегда в одном направлении. Цепи нейронов, по которым определенные
нервные импульсы проходят в одном направлении, формируют проводящие пути. Проводящий
путь может состоять из гетерогенных по характеру выделяемого медиатора
нейронов. Химическая передача нервного импульса через синаптический
аппарат была доказана в
2.6. НЕРВНЫЙ ИМПУЛЬС
В
1В
далее в форме потенциала действия. Как это было доказано в
В
Если под влиянием нейромедиатора повышается проницаемость постсинаптической мембраны, происходит прорыв внутрь расположенной после синаптической щели нервной клетки положительно заряженных ионов, главным образом ионов натрия, при этом после проникновения в клетку некоторого количества положительно заряженных ионов возникает прорыв в нее большого их количества, что ведет к моментальному изменению отрицательного внутриклеточного электрического заряда на положительный (приблизительно +40 мВ). Это явление называется деполяризацией клеточной мембраны и с фи- зиологической точки зрения представляет собой процесс возбуждения. В таких случаях возникает потенциал действия, который сразу же распространяется по нервному волокну. Подобная ситуация, порождающая потенциал действия, складывается и на периферии в рецепторных зонах, если интенсивность раз- дражения становится превышающей пороговую величину.
Амплитуда возникшего потенциала действия не зависит от дальнейшей интенсивности и длительности раздражения, так как действует закон «все или ничего». При деполяризации мембраны в зонах перехватов Ранвье происходит «подзарядка» нервного импульса. В результате возникает сальтаторное (скачкообразное) перемещение импульса без декремента, т.е. без потери энергии, без снижения амплитуды биоэлектрического потенциала. Другими словами, нервный импульс перемещается по нервному волокну, не затухая, без потери энергии. После прохождения импульса наступает рефрактерная фаза, во время которой происходит перераспределение ионов натрия и калия по сторонам мембраны нервного волокна. «Утомление» нервных проводников проявляется не изменением биоэлектрического заряда, а снижением частоты нервных импульсов.
Как уже было отмечено, влияние нейромедиатора на постсинаптическую мембрану может обусловить не только ее деполяризацию в связи с прохождением положительно заряженных ионов, но и нарастание отрицательного биопотенциала в тех случаях, когда через нее в клетку проникают в основном ионы с отрицательным биоэлектрическим зарядом (ионы хлора и др.). В таких случаях деполяризации мембраны не происходит, и клетка сохраняет, даже на-
1В
ращивает свой потенциал покоя, при этом она не возбуждается, а тормозится, становится менее возбудимой.
Таким образом, под влиянием воздействий на рецепторный аппарат или на постсинаптическую мембрану нейрона его биоэлектрический заряд может меняться как в сторону превращения отрицательного потенциала покоя в положительный биоэлектрический заряд, так и в сторону сохранения и даже наращивания выраженности отрицательного потенциала покоя. В первом случае речь идет о трансформации потенциала покоя в возбуждающий постсинаптический потенциал (ВПСП), во втором - об усилении потенциала покоя или о создании эффекта тормозного постсинаптического потенциала (ТПСП). В отдельный нейрон, например в периферический двигательный нейрон, расположенный в переднем роге спинного мозга, могут поступать как ВПСП, так и ТПСП. В нейроне происходит сложение подчас большого количества воздействующих на него биоэлектрических потенциалов с разными знаками (+ или -), и от суммы этих слагаемых зависит характер биоэлектрического потенциала, формирующегося внутри клетки и определяющего ее функциональное состояние.
Итак, следует отметить существование двух типов нервной передачи по нервному волокну - импульсного и безымпульсного. Импульсная передача обеспечивается электролитным и нейротрансмиттерным механизмами, безымпульсная - аксотоком, т.е. перемещением аксоплазмы по специальным микротрубочкам аксона, этим вторым путем к иннервируемой ткани доставляются, в частности трофогены - вещества, оказывающие на него трофотропное действие (Карлов В.А., 1999).
2.7. НЕЙРОГЛИЯ
Клетки глии описаны в
Астроглия состоит из астроцитов, которые, участвуя в формировании гематоэнцефалического барьера (ГЭБ), выполняют важную роль в обеспечении метаболических процессов в нервных клетках. Располагаясь между ними и капиллярами, астроциты, подобно своеобразному фильтру, пропускают в нервную клетку лишь то, что ей необходимо, и извлекают из нее продукты обмена.
Олигодендроглия сконцентрирована главным образом в белом веществе моз- га, формирует миелиновую оболочку нервных волокон. На периферии функции олигодендроцитов выполняют леммоциты (шванновские клетки).
Эпендимная глия выстилает расположенные внутри мозга ликворные пути (желудочки и водопровод мозга, а также центральный канал спинного мозга) и выполняет, таким образом, барьерную функцию.
Этим функции глии не ограничиваются. Еще в
Взаимодействуя с нервными клетками, глиальная ткань осуществляет, в частности, своеобразный обменный симбиоз с ними, влияя на состояние происходящих в них метаболических процессов. Она обеспечивает транспортные функции и, кроме того, обладает способностью к накоплению РНК, белка, дыхательных ферментов, которые в дальнейшем (в случае потребности в них нервных клеток) «перекачиваются» из глиальных элементов в нейроны, что обычно совершается в период функциональной активности последних. Глия, таким образом, может рассматриваться и как модулятор нейрональной активности. Некоторые глиальные клетки, в основном астроциты, обеспечивают возможность поглощения и создания запасов медиаторов с последующим их высвобождением. Последнее доказано, в частности, для гамма-аминомасляной кислоты (ГАМК) радиоавтографическим методом.
Глия (главным образом леммоциты) участвует в процессах дегенерации и регенерации нервной ткани. Выявлено химическое сродство аксонов нервных клеток к клеткам глии, обеспечивающим направление регенерации нервных волокон в случае их повреждения. В связи с этим периферические волокна растут по пути, обозначенному сохранившимися леммоцитами. Доказана также секреторная функция леммоцитов, которые высвобождают кванты ацетилхолина (АХ) в условиях регенерации периферических нервов.
Разрастание глиальной ткани при повреждении мозга ведет к формированию мозгового рубца. Глиальная ткань может выполнять и фагоцитарные функции. Есть мнение и о том, что глия является местом хранения закодированной информации, что обеспечивает так называемую долговременную па- мять. В последние десятилетия особое внимание уделяется функциональным возможностям астроцитов и содержащимся в них химическим компонентам. Так, недавно доказано, что астроциты участвуют в метаболизме глутамата (глу- таминовой кислоты) и ГАМК, которые, соответственно, являются возбудительным и тормозным медиаторами. Из синаптической щели часть названных медиаторов попадает в соседние астроциты, где превращается в глутамин, который в последующем используется нейронами для восстановления из него медиаторов глутамата и ГАМК. Установлено также, что астроциты участвуют в поддержании ионного баланса и, следовательно, влияют на генерацию импульса, регулируя уровни Na+ и K+ вокруг нервной клетки. Есть мнение, что формирование глиальных рубцов нарушает забуферивание К+ вокруг нервной клетки. В связи с этим снижается порог возбудимости нервных клеток, что может обусловить эпилептические разряды.
В
мена, находятся в астроцитах. Установлено также, что побочный продукт обмена триптофана - хинолиновая кислота - неблагоприятно влияет на глутамат-рецепторы нейронов, вызывая их гибель. В астроцитах находится фермент хинолинредуктаза, необходимый для синтеза хинолиновой кислоты. При избытке этого фермента в астроцитах возникает возможность избыточного производства хинолиновой кислоты, что ведет к гибели стриарных нейронов и в связи с этим к развитию болезни Гентингтона.
Большая группа демиелинизирующих заболеваний сопряжена с дисфункцией продуцирующих миелин клеток олигодендроглии в ЦНС и леммоцитов в составе периферической нервной системы. Измененный состав миелина, разрушение миелиновых оболочек нервных волокон, гибель олигодендроцитов и леммоцитов являются облигатными признаками таких заболеваний, как рассеянный склероз, лейкоэнцефалиты, полиневропатии, протекающие в форме миелинопатий, которые в большинстве случаев сопряжены с нарушением иммунных процессов в нервной ткани. Астроциты в таких случаях приобретают свойства иммуноэффекторных клеток, экспрессирующих ряд антигенов и иммуномодуляторов, которые вместе с микроглией, имеющей мезодермальное происхождение, формируют в мозговой ткани иммунный ответ.
Микроглия, состоящая из способных к миграции и фагоцитозу клеток мезодермального происхождения, активно участвует в обеспечении клеточного иммунитета. Ограждающий ЦНС гематоэнцефалический барьер не пропускает из крови в ткань мозга антигены и антитела, нет здесь и собственной лимфатической системы, поэтому астроциты и микроглия берут на себя роль иммунной системы при нейроинфекциях, когда полиморфно-ядерные лейкоциты превращаются в зернистые шары и бактериофаги, способствуя очищению мозговой ткани от продуктов распада погибших нервных клеток. Таким образом, изменение состояния и функции глии может влиять на развитие многих физиологических процессов и заболеваний нервной системы.
2.8. НЕРВНАЯ СИСТЕМА
Нервная ткань формирует нервную систему, которая имеет центральный и периферический отделы. Существует, кроме того, деление нервной системы на соматическую (анимальную) и вегетативную (автономную).
2.8.1. Центральная нервная система
Центральную нервную систему составляют расположенный в полости черепа головной мозг и находящийся в позвоночном канале спинной мозг.
2.8.1.1. Головной мозг
Головной мозг располагается в полости черепа, которая наметом мозжечка (tentorium) разделяется на две неравных части: супратенториальную и субтенториальную. Масса головного мозга человека в среднем составляет
физ. Субтенториально размещается мозговой ствол (truncus cerebri), состоящий из среднего мозга, моста мозга и продолговатого мозга, а также мозжечок (cerebellum) (см. главы 7, 9-11).
2.8.1.2. Спинной мозг
Спинной мозг находится в позвоночном кана- ле, его масса от 34 до
Спинной мозг имеет 31-32 пары передних и столько же пар задних корешков. Участок спинного мозга, точнее его серого вещества, анатомически и функционально связанный с парой передних и парой задних корешков, называется сегментом. 8 верхних сегментов образуют шейный отдел спинного мозга, грудной отдел его состоит из 12 сегментов, поясничный и крестцовый отделы имеют по 5 сегментов каждый, последние 1-2 сегмента составляет копчиковый отдел.
Шейные сегменты спинного мозга принято обозначать латинской буквой С (cervicalis) с указанием порядкового номера сегмента (например, V шейный сегмент - CV). Грудные сегменты обозначаются буквами Th (thoracalis), поясничные - L (lumbalis), крестцовые - Со (coccygea) также в сочетании с порядковым номером соответствующего сегмента. Каждый сегмент спинного мозга через посредство своих корешков обеспечивает иннервацию определенного участка тела, именуемого метамером, при этом входящие в метамер мышцы составляют миотом, кости - склеротом, а относящийся к одному метамеру участок кожи называется дерматомом. Название каждого миотома, скле-
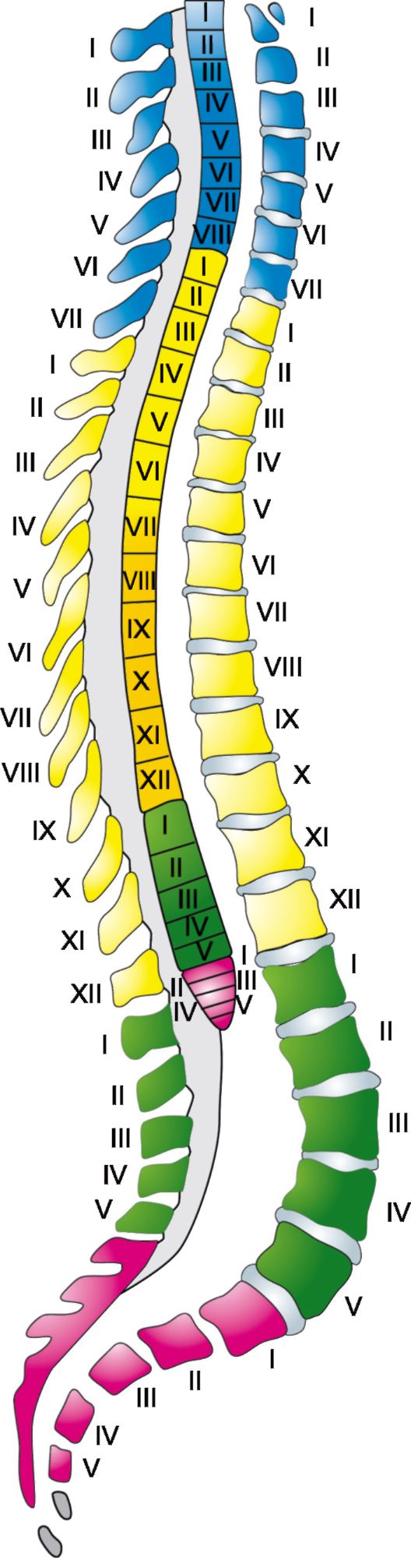
Рис. 2.8. Топографические соотношения между сегментами спинного мозга и позвонками.
ротома и дерматома определяется в соответствии с названием соответствующего ему сегмента спинного мозга.
Толщина спинного мозга не идентична на уровне разных его отделов. На среднем грудном уровне - приблизительно
На поперечном срезе спинного мозга видно, что его центральную часть занимает серое вещество, образующее фигуру, несколько напоминающую бабочку. Ее форма варьирует на разных уровнях спинного мозга. В ней выделяются центральная часть серого вещества, два задних и два передних рога, а на уровне CVIII-LIII - еще и боковые рога спинного мозга. Белое вещество на поперечном срезе спинного мозга занимает не заполненные серым веществом участки. Оно образует передние, задние и латеральные канатики (столбы) спинного мозга, состоящие из нервных волокон, формирующих восходящие (афферентные) и нисходящие (эфферентные) проводящие пути (см. главу 8).
2.8.2. Периферическая нервная система
Нервная ткань, находящаяся вне пределов головного и спинного мозга, относится к периферической нервной системе. Ее образуют главным образом отростки нервных клеток. Имеющиеся в ее составе нейроны в основном сконцентрированы в периферических нервных узлах - ганглиях.
Периферическая нервная система имеет черепной и спинальный уровни. Первый из них составляют 12 пар черепных нервов. Спинальный уровень периферической нервной системы составляют спинальные нервные корешки, нервные ганглии, спинномозговые нервы, их ветви, нервные сплетения, нервные стволы и их ветви, относящиеся к анимальной и вегетативной нервной системе.
Слияние передних и задних спинальных корешков ведет к формированию спинномозгового (спинального) нерва. По составу он смешанный, так как из переднего корешка в него попадают аксоны двигательных клеток, тела которых располагаются в передних рогах, и аксоны симпатических клеток, находящихся в боковых рогах спинного мозга. Кроме того, в его состав входят дендриты чувствительных клеток, тела которых (тела первых чувствительных нейронов) расположены в спинальных ганглиях или их аналогов на краниальном уровне.
Спинномозговой нерв (nervus spinalis) имеет длину около
печивающие иннервацию тканей, расположенных вблизи позвоночника; 4) переднюю ветвь (ramus anterior seu ventralis) - самую крупную, тоже смешанную по функции составляющих ее нервных волокон, осуществляющую иннервацию большей части соответствующего метамера тела человека (миотома, склеротома и дерматома).
Из передних ветвей большей части спинномозговых нервов формируются нервные сплетения. Исключение составляют только передние ветви спинальных нервов ThIII-ThXI, которые непосредственно переходят в соответствующие межреберные нервы. В нервных сплетениях происходит переплетение нервных волокон, проходящих через передние ветви расположенных по соседству спинномозговых нервов. После такого переплетения нервные волокна выходят из сплетений уже в составе периферических нервных стволов (периферические нервы). В результате в состав периферических нервов обычно входят нервные волокна, относящиеся к системе нескольких соседних спинальных сегментов, при этом периферические нервы могут быть смешанными (содержащими двигательные, чувствительные и вегетативные волокна), двигательными или чувствительными.
В связи с тем, что большинство периферических нервов формируется из сплетений, зоны их иннервации не соответствуют сопряженным с определенными сегментами спинного мозга метамерам человеческого тела, в частности его дерматомам. В результате участки тела, в которых возникают признаки нарушения их иннервации при поражении спинномозговых нервов, а также их корешков, не соответствуют участкам тела и, в частности, кожи, в которых возникают неврологические расстройства при поражении периферических нервов, формирующихся в нервных сплетениях.
В периферических нервах нервные волокна группируются в пучки. В пучках они отделяются друг от друга прослойкой соединительной ткани - эндоневрием. Каждый пучок отграничивается от соседних также соединительнотканной перегородкой - периневрием. Соединительнотканная оболочка, окружающая нервный ствол снаружи, называется эпиневрием. Соединительная ткань нервного ствола содержит кровеносные и лимфатические сосуды, обеспечивающие обменные процессы в нерве, а также мелкие нервные веточки (nervi nervorum), содержащие главным образом вазомоторные волокна.
2.9. ЦЕРЕБРОСПИНАЛЬНАЯ ЖИДКОСТЬ И ЕЕ ЦИРКУЛЯЦИЯ
Как уже подчеркивалось, цереброспинальная жидкость (ЦСЖ) продуцируется главным образом сосудистыми сплетениями, расположенными в желудочках мозга (рис. 2.9). Трансформировавшиеся из полости нервной трубки и ее производных (мозговых пузырей) мозговые желудочки сообщаются между собой., при этом из IV желудочка через непарное срединное отверстие (отверстие Мажанди) и парное боковое отверстие (отверстие Люшки) ЦСЖ проникает в ликворные цистерны основания черепа, а затем и в субарахноидальные пространства головного и спинного мозга, расположенные между мягкой и паутинной (арахноидальной) оболочками.
ЦСЖ, попадающая в субарахноидальное пространство, распространяется на основание мозга и его конвекситальные отделы, а также по спинальным
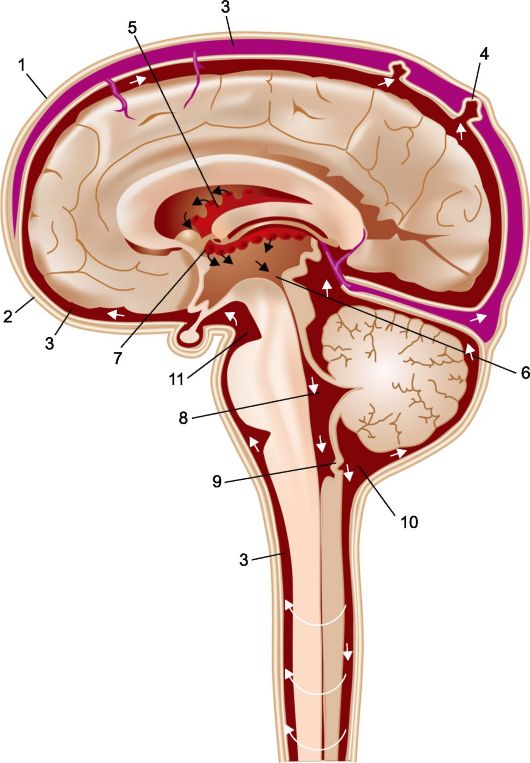
Рис. 2.9. Циркуляция цереброспинальной жидкости [по Netter].
1 - твердая мозговая оболочка; 2 - арахноидальная оболочка; 3 - субарахноидальные пространства; 4 - грануляции арахноидальной оболочки (пахионовы грануляции); 5 - хориоидальные сплетения; 6 - водопровод мозга; 7 - межжелудочковое отверстие; 8 - латеральная апертура IV желудочка мозга; 9 - медиальная апертура IV желудочка мозга; 10 - мозжечково-мозговая цистерна; 11 - межножковая цистерна.
субарахноидальным путям до конечной цистерны, расположенной в каудальной части позвоночного канала, и циркулирует в них. В норме перемещение ЦСЖ от терминальной цистерны к головному мозгу составляет около 1 ч. Приблизительно с той же скоростью осуществляется перемещение ЦСЖ и в противоположном направлении. Циркуляция ЦСЖ обусловлена перепадами гидростатического давления в ликворных путях, пульсацией внутричерепных
артерий, изменениями венозного давления, положения тела в пространстве и пр. Из субарахноидального пространства ЦСЖ проникает в глубину щелевидных мозговых борозд и в периваскулярные, перикапиллярные, а также перицеллюлярные пространства, сопутствующие погружающимся в мозговое вещество сосудам, известные как пространства Вирхова-Робена.
Физиологические функции ЦСЖ многообразны. Она обеспечивает механическую поддержку мозга и создает своеобразную амортизирующую систему, способствующую его механической защите. Кроме того, ЦСЖ участвует в обеспечении относительного постоянства внутричерепного и, в частности, внутрижелудочкового давления, в поддержании оптимального водно-электролитного баланса и других показателей гомеостаза в головном и спинном мозге, в выведении продуктов их метаболизма, в частности лактата и СО2.
В норме секреция ЦСЖ совершается в желудочковой системе мозга со скоростью приблизительно 0,35 мл/мин или 20 мл/ч. В сутки, таким образом, продуцируется около 500-700 мл ЦСЖ. Объем секретируемой ЦСЖ может варьировать в зависимости от вида пищи, водного режима, колебаний активности физиологических процессов. Попадающая в подоболочечные пространства ЦСЖ подвергается резорбции, которая совершается главным образом через арахноидальные ворсины в области пахионовых грануляций, которые в особенно большом количестве находятся в парасагиттальной зоне конвекситального отдела черепа.
Продукция ЦСЖ и ее резорбция в норме достаточно сбалансированы, что способствует поддержанию относительного постоянства объема ЦСЖ в ликворных путях. В норме общий постоянный объем ЦСЖ во всех предназначенных для нее пространствах в полости черепа и позвоночного канала у новорожденного составляет 15-20 мл, у годовалого ребенка - 35 мл, у взрослого человека - приблизительно 140 мл, из которых 30 мл находится в спинальном субарахноидальном пространстве и, в частности, в конечной цистерне. Полная замена ЦСЖ в норме у взрослого человека происходит в течение 5-7 ч, т.е. 4-5 раз в сут.
ЦСЖ обычно получают при поясничном проколе, редко при проколе большой цистерны или бокового желудочка мозга.
При поясничном проколе у лежащего на боку пациента нормальным может быть признано давление ЦСЖ, вытекающей каплями со скоростью 60- 80 капель/мин. Давление ЦСЖ по манометру в норме может быть в пределах от 100 до 180-200 мм вод.ст., при этом у детей оно ниже, чем у взрослых. Подъем системного АД и венозного давления обычно сопровождается повы- шением и ликворного давления, на нем отражаются также колебание пульса и дыхание.
В норме ЦСЖ бесцветная, прозрачная, стерильная, имеет относительную плотность 1,005-1,009, реакция ее нейтральная или слабощелочная, вязкость почти такая же, как у воды. В 1 мм3 ЦСЖ содержится до 5 клеток (в основном лимфоциты). Количество белка в ЦСЖ в норме 0,12-0,45 г/л, при этом величина белкового коэффициента, т.е. отношение альбуминов к глобулинам колеблется в пределах 0,4-0,6. Нормальным считается содержание в ЦСЖ глюкозы в пределах 45,0-79,0 мг% (2,5-4,4 ммоль/л), ионов хлора 115-125 ммоль/л или мэкв/л, хлоридов 7-7,5 г/л, кальция 0,9-1,35 ммоль/л, калия 2,6-2,9 ммоль/л, магния 1,05-1,7 ммоль/л. Состав ЦСЖ во многом оп- ределяется состоянием ГЭБ.
2.10. МОЗГОВЫЕ ОБОЛОЧКИ
Головной и спинной мозг окружают три оболочки: мягкая, плотно прилежащая к ткани головного и спинного мозга и повторяющая все нюансы ее рельефа; паутинная, перебрасывающаяся через мозговые углубления и борозды; и твердая - наиболее плотная, прилежащая к костям черепа, а местами сращенная с ними и выстилающая полость позвоночного канала (см. рис. 2.7). Все мозговые оболочки, как и находящиеся над ними кости черепа и позвоночника, имеют мезодермальное происхождение. Они обеспечивают защиту мозговой ткани и ликворной системы.
Мягкая мозговая оболочка сращена с мозгом и богата сосудами, которые принимают участие в кровоснабжении поверхностных слоев мозговой ткани.
Паутинная оболочка рыхло связана с мягкой посредством соединительнотканных перемычек (трабекул) и перебрасывается через имеющиеся на поверхности мозга углубления с образованием заполненных, как и все субарахноидальные пространства, ЦСЖ так называемых подпаутинных или ликворных цистерн.
Ликворные цистерны представляют собой вместилища ЦСЖ, наиболее важными из них являются базальные, находящиеся на основании черепа: мозжечково-мозговая цистерна (cisterna cerebellomedullaris), цистерна перекреста (cisterna chiasmatis), межножковая цистерна (cisterna interpeduncularis и др.). В позвоночном канале значительная по объему ликворная цистерна имеется в его каудальной части, начиная с уровня окончания спинного мозга, соответствующего проекции LII позвонка, и потому именуемая конечной цистерной (cisterna terminalis). Через конечную цистерну проходит конечная нить и окружающие ее корешки спинного мозга, направляющиеся вниз в «поиске» одноименных межпозвонковых отверстий. Вместе они образуют конский хвост (cauda equina). Именно эта цистерна пунктируется в процессе поясничного прокола.
Твердая мозговая оболочка покрывает арахноидальную и состоит из плотной волокнистой соединительной ткани. Ее составляют два листка. Наружный листок сращен с костями черепа и позвоночника, на основании черепа с костями сращен и внутренний листок твердой мозговой оболочки. В полости черепа она образует складки (дубликатуры), которые принято называть ее отростками. Наиболее крупные отростки твердой мозговой оболочки, находящиеся в сагиттальной плоскости, - серп большого мозга (falx cerebri) между большими полушариями мозга и серп мозжечка (falx cerebelli), расположенный между полушариями мозжечка. В горизонтальной плоскости расположен намет мозжечка (tentorium cerebelli), поддерживающий снизу затылочные доли мозга, отграничивая их от верхней поверхности полушарий мозжечка. Намет мозжечка делит полость черепа на две сообщающиеся между собой неравные части, меньшая располагается под ним и потому называется субтенториальной, а большая - над наметом мозжечка и обозначается как супратенториальное пространство. Дном субтенториального пространства служит задняя черепная ямка, а супратенториального пространства - средняя и передняя черепные ямки.
2.11. КРАТКИЕ СВЕДЕНИЯ О КРОВОСНАБЖЕНИИ ГОЛОВНОГО И СПИННОГО МОЗГА
К головному мозгу кровь притекает по двум парам магистральных сосудов: внутренним сонным и позвоночным артериям (рис. 2.10). Внутренняя сонная ар-
терия (a. carotis interna) отходит от общей сонной артерии (a. carotis communis), проходит через пещеристый синус, образуя в нем два изгиба (сифон внутренней сонной артерии). В полости черепа от внутренней сонной артерии отходят глазная артерия (a. ophtalmica) и передняя ворсинчатая артерия (a. chorioidea anterior). После этого внутренняя сонная артерия делится на конечные ветви - среднюю и переднюю мозговые артерии (aa. cerebri media et anterior), обеспечивающие кровоснабжение лобной, теменной и височной долей большого полушария.
Парная позвоночная артерия (a. vertebralis) представляет собой ветвь под- ключичной артерии. Она проходит через костный канал, образованный отверстиями в поперечных отростках шести верхних шейных позвонков, затем проникает сквозь атлантоокципитальную мембрану и через большое затылочное отверстие входит в полость черепа. В полости черепа от каждой позвоночной артерии отходят задняя спинальная артерия (a. spinalis posterior) и ветвь, участвующая в формировании непарной передней спинальной артерии (a. spinalis anterior). Они покидают полость черепа через большое затылочное отверстие, попадают в позвоночный канал и принимают участие в кровоснабжении спинного мозга. В полости черепа от каждой позвоночной артерии отходит задняя нижняя мозжечко- вая артерия (a. cerebellaris posterior inferior). После этого позвоночные артерии, подойдя к нижнему краю основания моста мозга, сливаются в непарную базилярную артерию (a. basilaris).
Базилярная артерия, пройдя по борозде основания моста, где от нее отходят парные передняя и верхняя мозжечковые артерии (aa. anterior et superior cerebellaris), на уровне среднего мозга делится на две за-
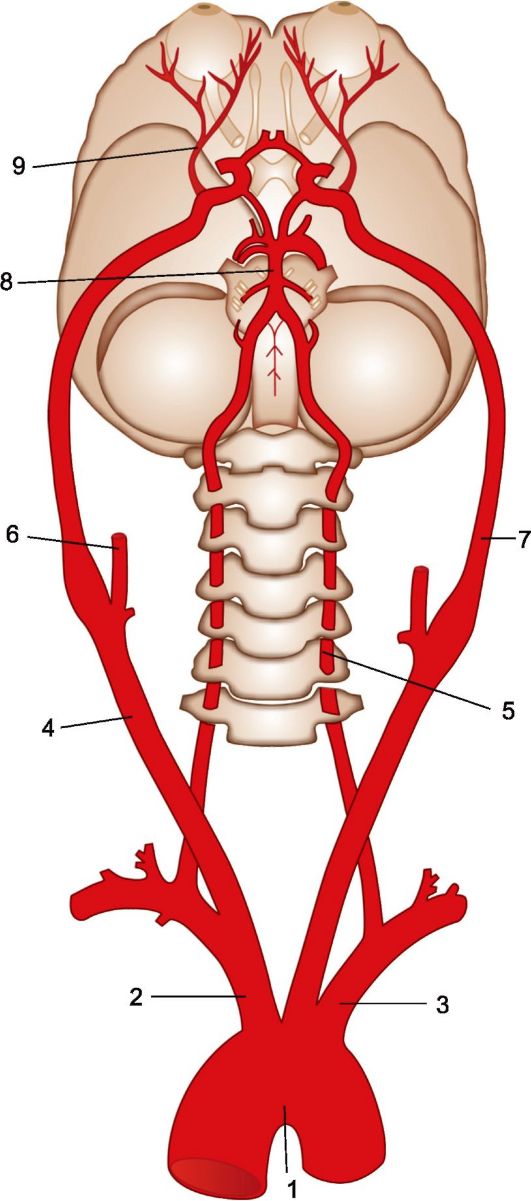
Рис. 2.10. Магистральные артерии головного мозга.
1 - дуга аорты; 2 - плечеголовной ствол; 3 - левая подключичная артерия; 4 - правая общая сонная артерия; 5 - позвоночная артерия; 6 - наружная сонная артерия; 7 - внутренняя сонная артерия; 8 - базилярная артерия; 9 - глазная артерия.
дние мозговые артерии (aa. cerebri posterioris), обеспечивающие кровоснабжение затылочных долей и прилежащих к ним структур больших полушарий. Ветви позвоночных и базилярной артерий снабжают кровью большую часть ствола мозга и мозжечок.
В центре средней черепной ямки находится артериальный круг большого мозга (виллизиев круг), соединяющий между собой каротидные и вертебробазилярную артериальные системы. Его составляют участки внутренних сонных артерий и проксимальные отделы передних и задних мозговых артерий, а также непарная передняя соединительная артерия и парная задняя соединительная артерия.
Кровоснабжение спинного мозга обеспечивается не всеми корешковыми артериями, а лишь несколькими несимметричными, наиболее крупными из них, называемыми в связи с этим корешково-мозговыми (радикуломедуллярными). Их общее число обычно не превышает 5-8. Восходящие и нисходящие ветви этих артерий образуют цепь анастомозов, находящуюся в передней продольной щели спинного мозга и известную под названием передней спинальной артерии. Самый верхний ее участок образуется в результате слияния двух ветвей позвоночных артерий, отходящих от их внутричерепных отделов. Система передней спинальной артерии снабжает кровью 4/5 поперечника спинного мозга, включающих в себя передние, боковые и частично задние его рога, а также передние и боковые канатики.
Заднемедиальные отделы задних рогов и задние канатики спинного мозга снабжаются кровью за счет задних спинальных артерий, в формировании которых принимают участие около 20 задних корешково-мозговых артерий.
Следует отметить, что анастомозы между отдельными сосудистыми бассейнами в головном и спинном мозге в случае нарушения кровообращения в одном из бассейнов нередко не в состоянии компенсировать это нарушение, особенно при остром его развитии. Ветвление артериальных сосудов головного и спинного мозга завершается капиллярной сетью, отсюда кровь попадает в мозговые вены, а затем во внутричерепные венозные синусы и венозные сплетения, расположенные в позвоночном канале. Из полости черепа венозная кровь оттекает главным образом через яремные вены, а из позвоночного канала - через корешковые вены.
2.12. ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР
Эндотелий капилляров и расположенных в мозговых желудочках сосудистых сплетений вместе с элементами глиальной ткани (главным образом с астроцитами) составляют гематоэнцефалический барьер (ГЭБ). Контакты между эндотелиальными клетками достаточно плотные, промежутки между ними, как правило, меньше 1,5 нм, что служит препятствием для большинства молекул, в частности белковых. К тому же между капилляром и нейроном располагается астроцитарная глия. Эти структуры и служат своеобразным фильтром, обеспечивающим относительную защиту нервных клеток и их отростков. Таким образом, ГЭБ регулирует поступление в ЦСЖ и нервную ткань циркулирующих в крови веществ и тем самым обеспечивает относительное постоянство параметров внутренней среды мозга, в частности ионного и осмотического баланса. ГЭБ защищает мозг и цереброспинальную жидкость от всевозможных
случайных эндогенных и экзогенных факторов, которые, попав в кровяное русло, могли бы оказать на нервную систему неблагоприятное воздействие. При патологических состояниях проницаемость ГЭБ может увеличиваться, что обусловливает проникновение в нервную систему веществ, приводящих к нарушению в ней гомеостаза и к развитию таких патологических состояний мозга, как отек, набухание, аутоиммунные процессы, и пр.
Ввиду того, что гематоэнцефалический барьер формируется в основном в постнатальном периоде, у детей в первые годы жизни при различных общих инфекциях развитие неврологических осложнений (менингит, менингоэнцефалит) возникает чаще, чем у взрослых.