Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 3 ВНУТРИУТРОБНОЕ РАЗВИТИЕ
Многоклеточный организм развивается из оплодотворённой яйцеклетки (зигота) не только путём увеличения количества клеток (пролиферация) и увеличения массы зародыша (рост). Одновременно определяется судьба отдельных клеток. Этот процесс известен как детерминация. Детерминированные клетки специализируются (дифференцировка), т.е. приобретают конкретную морфологию и оказываются способными выполнять конкретную функцию. Одновременно осуществляется морфогенез -формирование органов и архитектуры тела. Таким образом, детерминация, дифференцировка, морфогенез, пролиферация, рост, миграция клеток, гибель клеток, т.е разные морфогенетические процессы и составляют суть развития многоклеточного организма.
Периоды пренатального развития
В пренатальном развитии человека выделяют начальный, зародышевый и плодный периоды (табл. 3-2).
Таблица 3-2. Периоды пренатального развития
Период | Продолжительность | Критическое событие |
Начальный (концептус) Зародышевый (эмбрион) Плодный (плод) | Первые 2 недели От 3-й по 8-ю недели включительно До конца беременности | Оплодотворение Образование первичной полоски Плацентация |
* Эмбрион - общность клеток или существо, формирующееся на стадии первичной полоски, но не ранее. По мнению A. McLaren [69], нельзя называть эмбрионом продукт дифференцировки зиготы до стадии первичной полоски. Автор предлагает называть этот продукт концептусом, проэмбрионом, предэмбрионом.
Для начального периода и раннего эмбриона (см. рис. 3-6) выделены такие фазы развития и связанные с ними события: зигота - начало синтеза ДНК и белка; дробление - начало синтеза основных типов РНК; морула - клетки ещё тотипотентны (эмбриональная стволовая клетка); бластоциста - утрата тотипотентности; клетки детермини-
рованы к образованию зародышевых или внезародышевых структур; гаструла - наличие зародышевых листков и стволовых клеток; органогенез - формирование зачатков органов из клеточных клонов.
Половые клетки
Сперматозоид. В эякуляте человека содержится 3х108 сперматозоидов (рис. 3-2). В женских половых путях они сохраняют способность к оплодотворению до 2 суток, но до оплодотворения сперматозоиды должны пройти процесс капацитации. Сперматозоиды направленно
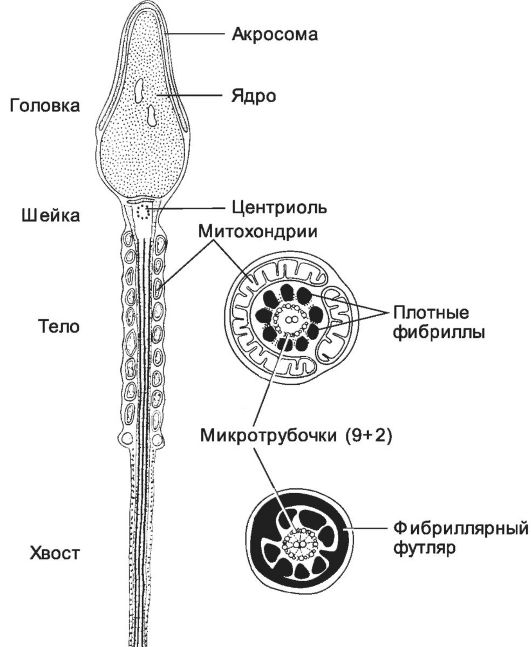 Рис. 3-2. Сперматозоид. Головка
содержит ядро и акросому. В шейке присутствует центриоль. Промежуточный
отдел заполнен митохондриями, формирующими митохондриальную спираль. В
осевой части промежуточного отдела и практически вдоль всего хвоста
располагается аксонема. В хвосте (снаружи от аксонемы) локализуются
наружные плотные фибриллы и фибриллярный футляр. [17]
Рис. 3-2. Сперматозоид. Головка
содержит ядро и акросому. В шейке присутствует центриоль. Промежуточный
отдел заполнен митохондриями, формирующими митохондриальную спираль. В
осевой части промежуточного отдела и практически вдоль всего хвоста
располагается аксонема. В хвосте (снаружи от аксонемы) локализуются
наружные плотные фибриллы и фибриллярный футляр. [17]
мигрируют к воронке маточной трубы, где и происходит их встреча с яйцеклеткой (см. позицию 2 на рис. 3-6).
• Направленная миграция сперматозоида. В феномене встречи сперматозоида и яйцеклетки важную роль играет хемотаксис - направленная миграция сперматозоидов по градиенту химических веществ, выделяемых овулировавшей яйцеклеткой. Хемоаттрактанты попадают в маточную трубу в составе фолликулярной жидкости лопнувшего фолликула, а также секретируются яйцеклеткой и фолликулярными клетками corona radiata.
• Капацитация. До того как сперматозоид встретится с яйцеклеткой, он в течение нескольких часов находится в женских половых путях. При этом на сперматозоид воздействуют факторы женского организма (pH, слизь, прогестерон, хемоаттрактанты и др.). Жизнеспособные и подвижные сперматозоиды не способны оплодотворить яйцеклетку, пока не пройдут окончательное созревание в женских половых путях. Процесс, при котором сперматозоид приобретает способность к оплодотворению яйцеклетки, называется капацитацией. Для успешной капацитации сперматозоиды должны примерно 7 часов находиться в женских половых путях.
• Акросома (см. рис. 3-2) образуется в ходе сперматогенеза как продукт комплекса Гольджи и может рассматриваться как аналог лизосомы. Акросома расположена в головке сперматозоида, кпереди от ядра и тотчас под плазматической мембраной. Спереди мембрана акросомы (наружная) соприкасается с клеточной мембраной сперматозоида, а сзади (внутренняя мембрана) - с ядерной мембраной. При оплодотворении акросома участвует в акросомной реакции.
Яйцеклетка
При разрыве зрелого фолликула (овуляция) из яичника в брюшную полость выбрасывается овоцит второго порядка (рис. 3-3). Его окружают прозрачная оболочка (zona pellucida) и лучистый венец (corona radiata).
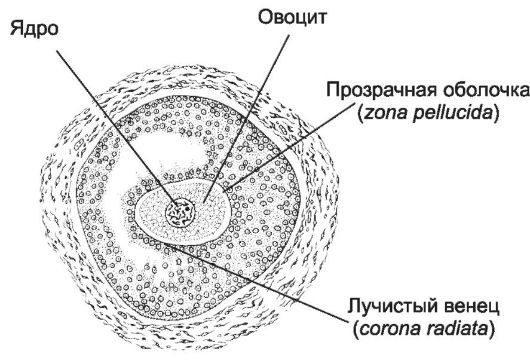 Рис. 3-3. Овоцит. Лучистый
венец, прозрачная оболочка и плазматическая мембрана овоцита - барьеры,
которые должен последовательно преодолеть сперматозоид, чтобы произошло
оплодотворение. [17]
Рис. 3-3. Овоцит. Лучистый
венец, прозрачная оболочка и плазматическая мембрана овоцита - барьеры,
которые должен последовательно преодолеть сперматозоид, чтобы произошло
оплодотворение. [17]
• Перемещение яйцеклетки. Перед овуляцией увеличение тонуса ГМК маточной трубы приближает воронку маточной трубы к поверхности яичника. Биение ресничек эпителиальных клеток фимбрий ампулы маточной трубы создаёт ток жидкости, способствующий перемещению яйцеклетки из брюшной полости в ампулу.
• Способность к оплодотворению. Для успешного оплодотворения яйцеклетка должна встретиться со сперматозоидом в течение одних суток после овуляции. Из практических соображений время, в течение которого овулировавшая яйцевая клетка может быть оплодотворена, оценивают в пять суток.
• Прозрачная оболочка. По мере развития яйцеклетки в ней происходят синтез и последующая секреция гликопротеинов, постепенно формирующих zona pellucida. Зрелая прозрачная оболочка содержит густую сеть тонких нитей, состоящую из гликопротеинов (главным образом, белки ZP); один из них (ZP3) - главный рецептор сперматозоида, а ZP2 - вторичный рецептор, обеспечивающий дополнительное связывание гамет. Связывание сперматозоида с гликопротеинами ZP является сигналом к акросомной реакции.
Оплодотворение
Оплодотворение - слияние мужской и женской гамет - приводит к образованию одноклеточного зародыша - зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы; при этом сливаются их ядра (пронуклеусы), объединяются хромосомы, и возникает диплоидная клетка нового организма - зигота. Начало оплодотворения - момент слияния мембран сперматозоида и яйцевой клетки, окончание оплодотворения - момент объединения материала мужского и женского пронуклеусов.
Для того, чтобы произошло оплодотворение, сперматозоид должен последовательно преодолеть три барьера (рис. 3-4) лучистый венец, состоящий из нескольких слоёв фолликулярных клеток, прозрачную оболочку и, наконец, плазматическую мембрану яйцеклетки. Сперматозоид легко проникает через лучистый венец между рыхло расположенными фолликулярными клетками и достигает прозрачной оболочки. При взаимодействии сперматозоида с прозрачной оболочкой происходят следующие события: связывание сперматозоида с его рецептором → акросомная реакция → расщепление компонентов zona pellucida ферментами акросомы → проникновение сперматозоида через образовавшийся в оболочке канал к плазматической мембране яйцеклетки.
• Акросомная реакция - экзоцитоз содержимого акросомы, предназначенного для разрушения прозрачной оболочки. Начало акросомной реакции - следствие взаимодействия олигосахаридов гликопротеи-
Рис. 3-4. Преодоление сперматозоидом барьеров яйцевой клетки. А - проникновение сперматозоида через лучистый венец; Б - акросомная реакция; В - прохождение сперматозоида через прозрачную оболочку; Г - слияние клеточных мембран сперматозоида и яйцеклетки. [17]
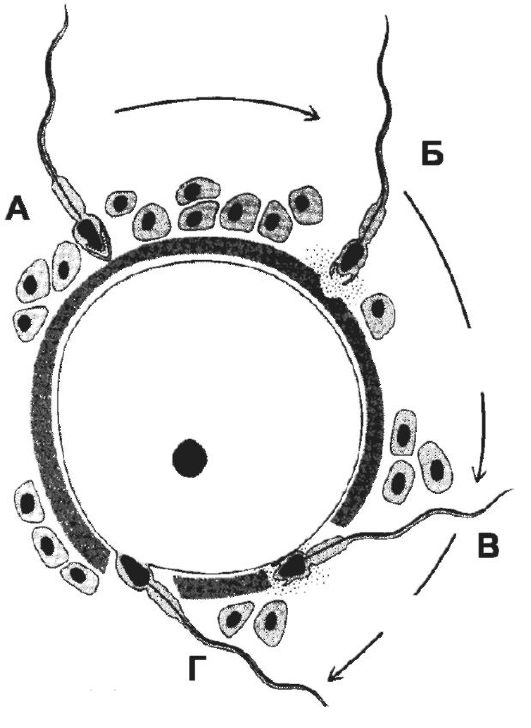 на
ZP3 прозрачной оболочки с его лектин-подобным рецептором в мембране
головки сперматозоида. При акросомной реакции наружная мембрана акросомы
и клеточная мембрана сперматозоида сливаются и формируют мелкие
пузырьки, отделяющиеся от головки. При этом из акросомы освобождаются
гиалуронидаза, протеазы и другие ферменты, расщепляющие молекулы
прозрачной оболочки. Слияние гамет. В результате акросомной
реакции в прозрачной оболочке образуется узкий канал, через который
проходит сперматозоид; плазматические мембраны сперматозоида и
яйцеклетки вступают в соприкосновение и сливаются, а ядро сперматозоида
оказывается погружённым в цитоплазму яйцеклетки.
на
ZP3 прозрачной оболочки с его лектин-подобным рецептором в мембране
головки сперматозоида. При акросомной реакции наружная мембрана акросомы
и клеточная мембрана сперматозоида сливаются и формируют мелкие
пузырьки, отделяющиеся от головки. При этом из акросомы освобождаются
гиалуронидаза, протеазы и другие ферменты, расщепляющие молекулы
прозрачной оболочки. Слияние гамет. В результате акросомной
реакции в прозрачной оболочке образуется узкий канал, через который
проходит сперматозоид; плазматические мембраны сперматозоида и
яйцеклетки вступают в соприкосновение и сливаются, а ядро сперматозоида
оказывается погружённым в цитоплазму яйцеклетки.
Активация яйцеклетки. Оплодотворение активизирует яйцеклетку, в цитоплазме повышается концентрация Ca2+, что служит сигналом для второго деления мейоза. Оплодотворённый овоцит второго порядка после метафазы второго деления завершает мейоз с образованием гаплоидной зрелой яйцеклетки и второго полярного тельца. Объединение пронуклеусов. В течение первых 12 часов после проникновения сперматозоида в яйцеклетку происходит перестройка ядер (пронуклеусов) слившихся гамет. Ядра набухают, появляются ядрышки. Пронуклеусы мигрируют в центр яйцеклетки и сближаются. Их ядерные оболочки исчезают, а материнские и отцовские хромосомы перемешиваются - образуется синкарион. Этот процесс (сингамия) и есть собственно оплодотворение.
• Кортикальная реакция. По периферии яйцеклетки расположены кортикальные гранулы. Их диаметр 200-600 нм; они содержат ферменты, в том числе различные гидролазы. Сразу после проникновения сперматозоида в яйцеклетку начинается кортикальная реакция - выделение содержимого кортикальных гранул в перивителлиновое пространство. Сигнал для кортикальной реакции - повышение концентрации Ca2+ в цитозоле. Под действием ферментов кортикальных гранул происходят протеолиз ZP2 и модификация рецептора сперматозоида ZP3. В модифицированном состоянии молекулы ZP теряют способность к запуску акросомной реакции в других сперматозоидах. Тем самым блокируется полиспермия.
• Оболочка оплодотворения. В результате кортикальной реакции прозрачная оболочка подвергается существенным изменениям - происходит её стабилизация (оболочка оплодотворения). В стабилизированном состоянии оболочка оплодотворения защищает проходящий по маточной трубе концептус. Без оболочки оплодотворения дробление зиготы невозможно.
От зиготы до бластоцисты
Оплодотворённая яйцеклетка в результате дроблений формирует зародыш (концептус), состоящий из бластомеров. В дальнейшем (до начала гаструляции) зародыш проходит стадии морулы и бластоцисты.
Дробление
Дробление - митотическое деление диплоидных клеток без увеличения их суммарного объёма. При дроблении происходит относительно быстрое увеличение количества клеток (бластомеров), от 2 до примерно 12-16 к третьим суткам после оплодотворения, когда концептус достигает стадии морулы и выходит в полость матки из маточной трубы (рис. 3-6). Дробление отличается от обычного клеточного цикла отсутствием фаз G1 и G2 и очень короткой S-фазой, во время которой происходит синтез ДНК. Интервал между делениями дробления составляет примерно 12-24 часа. В ходе дроблений клетки уменьшаются в размерах, так что диаметр концептуса не превышает 100 мкм. На стадии бластоцисты дробления прекращаются и клетки возобновляют обычный клеточный цикл.
Характер дробления определяется количеством и распределением в цитоплазме желточных включений. У человека - плацентарного млекопитающего - в связи с внутриутробным питанием плода через плаценту - нет необходимости в создании больших запасов желтка. Поэтому желток распределён в цитоплазме равномерно (изолецитальная яйцеклетка). Зигота, образованная такой яйцеклеткой, совершает голо-
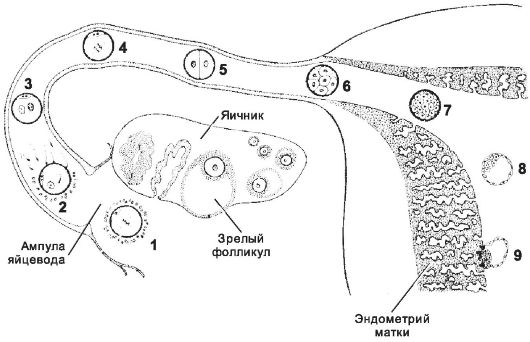 Рис. 3-6. Концептус от оплодотворения до имплантации. Овуляция
наступает на 14-й день менструального цикла, в течение 1 сут происходит
оплодотворение. Трое суток с момента оплодотворения концептус
продвигается по маточной трубе, на 4-е сутки попадает в полость матки и
через 5,5-6 сут имплантируется в эндометрий. В ходе перемещения по
маточной трубе происходит дробление, и в полости матки оказывается
бластоциста, готовая к имплантации. 1 - овоцит тотчас после овуляции; 2 - оплодотворение через 12-24 ч после овуляции; 3 - стадия появления мужского и женского пронуклеусов; 4 - начало дробления; 5 - двухклеточная стадия (30 часов); 6 - морула, состоящая из 12-16 бластомеров (3 сут); 7 - продвинутая морула в полости матки (4 сут); 8 - ранняя бластоциста (4,5-5 сут); 9 - имплантация бластоцисты (5,5-6 сут). [17]
Рис. 3-6. Концептус от оплодотворения до имплантации. Овуляция
наступает на 14-й день менструального цикла, в течение 1 сут происходит
оплодотворение. Трое суток с момента оплодотворения концептус
продвигается по маточной трубе, на 4-е сутки попадает в полость матки и
через 5,5-6 сут имплантируется в эндометрий. В ходе перемещения по
маточной трубе происходит дробление, и в полости матки оказывается
бластоциста, готовая к имплантации. 1 - овоцит тотчас после овуляции; 2 - оплодотворение через 12-24 ч после овуляции; 3 - стадия появления мужского и женского пронуклеусов; 4 - начало дробления; 5 - двухклеточная стадия (30 часов); 6 - морула, состоящая из 12-16 бластомеров (3 сут); 7 - продвинутая морула в полости матки (4 сут); 8 - ранняя бластоциста (4,5-5 сут); 9 - имплантация бластоцисты (5,5-6 сут). [17]
бластическое дробление, т.е. полностью разделяется на два бластомера. Дальнейшие дробления - асинхронные и несколько неравномерные. При
дроблении зигота окружена прозрачной оболочкой, сохраняющейся до стадии бластоцисты.
• Первое дробление (рис. 3-6) начинается примерно через 30 часов после оплодотворения. Плоскость первого деления проходит через область направительных телец. Область отделения направительных телец обычно рассматривают как анимальный полюс. Образующиеся два бластомера несколько различны по размерам. В результате первого деления дробления нарушается симметрия зародыша и образуется два бластомера с различными потенциями развития: один бластомер дает начало «эмбриональной части» бластоцисты (полярный трофобласт и глубокая часть внутренней клеточной массы), а другой бластомер - «неэмбриональ-
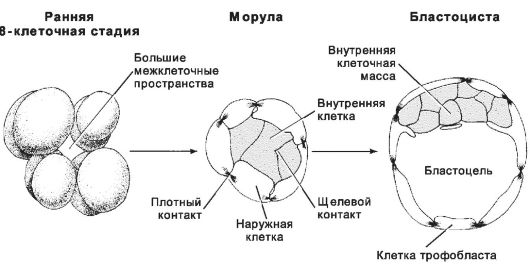 Рис. 3-7. Компакт изация и морула на ранних этапах развития. В
результате компактизации уменьшается размер межклеточных пространств
между бластомерами, они сближаются, и формируется морула. В моруле
различают внутреннюю часть (клетки связаны щелевыми контактами) и
наружную часть (клетки соединены при помощи плотных контактов). Из
клеток внутренней части морулы развивается эмбриобласт, а из клеток
наружной части формируется трофобласт. Внутренняя клеточная масса и
трофобласт - главные структурные элементы бластоцисты. [17]
Рис. 3-7. Компакт изация и морула на ранних этапах развития. В
результате компактизации уменьшается размер межклеточных пространств
между бластомерами, они сближаются, и формируется морула. В моруле
различают внутреннюю часть (клетки связаны щелевыми контактами) и
наружную часть (клетки соединены при помощи плотных контактов). Из
клеток внутренней части морулы развивается эмбриобласт, а из клеток
наружной части формируется трофобласт. Внутренняя клеточная масса и
трофобласт - главные структурные элементы бластоцисты. [17]
ной части» бластоцисты (муральный трофобласт и поверхностный слой внутренней клеточной массы).
• Второе дробление (рис. 3-6). Плоскость второго деления проходит перпендикулярно плоскости первого деления дробления, концептус формально переходит в стадию 4-х бластомеров, но дробление у человека асинхронно, поэтому при переходе от стадии 2-х бластомеров к стадии 4-х бластомеров в течение некоторого времени можно наблюдать трёхклеточный концептус.
• Третье дробление. На этой стадии образуется концептус с различным количеством бластомеров; условно его можно отнести к стадии 8 бластомеров (рис. 3-6, рис. 3-7). До этого бластомеры расположены рыхло, но перед следующим (4-м) делением дробления эмбрион подвергается компактизации. Концептус уплотняется (компактизация), поверхность соприкосновения бластомеров увеличивается, а объём межклеточного пространства уменьшается.
Морула
По мере увеличения числа бластомеров концептус перемещается по маточной трубе и к третьим суткам достигает стадии морулы (рис. 3-6). Морула - группа клеток, возникших в ходе нескольких делений дробления и заключённых внутри прозрачной оболочки (рис. 3-7). Центрально расположенные клетки морулы образуют щелевые контакты.
Именно из этих клеток и возникает собственно эмбрион. Периферические клетки морулы соединяют плотные контакты; они формируют барьер, обособляющий внутреннюю среду морулы.
Бластоциста
Бластоциста возникает с появлением бластоцеля (заполненной жидкостью полости) к четвёртым суткам после оплодотворения (рис. 3-7, рис. 3-9, рис. 3-21А). Объём бластоцеля увеличивается, и концептус приобретает форму пузырька. Прозрачная оболочка истончается и исчезает. Бластоцисту составляют трофобласт и внутренняя клеточная масса (эмбриобласт).
Трофобласт - покров зародышевого комплекса (рис. 3-7, рис. 3-9); образован периферическими клетками морулы. Крупные удлинённые клетки трофобласта соединены при помощи плотных контактов, что стабилизирует состав жидкости в бластоцеле. Клетки трофобласта «накачивают» жидкость в бластоцель.
Внутренняя клеточная масса (эмбриобласт) - компактная масса мелких клеток, выступающих в бластоцель (рис. 3-7, рис. 3-9). Клетки внутренней клеточной массы происходят из центральной части морулы и связаны при помощи щелевых контактов. В дальнейшем из внутренней клеточной массы образуются собственно зародыш и некоторые связанные с ним оболочки. Частичное или полное разделение внутренней клеточной массы приводит к развитию близнецов.
Гаструляция и первичные зародышевые листки
• Гаструляция начинается в конце второй недели развития и характеризуется появлением у клеток способности к перемещениям. С началом гаструляции активируются первые тканеспецифические гены. Эмбриобласт расслаивается на эпибласт (слой цилиндрических клеток) и гипобласт (слой кубических клеток, обращённый к бластоцелю). Эпибласт и гипобласт вместе образуют двухслойный зародышевый диск (бластодиск). В дальнейшем на месте двухслойного зародышевого диска путём миграции и пролиферации клеток развиваются первичные зародышевые листки: эктодерма, мезодерма и энтодерма. Гипобласт. Формирование гипобласта (первичной энтодермы) происходит по каудально-краниальному градиенту. Обращённые к бластоцелю клетки вентральной части внутренней клеточной массы обособляются в тонкий слой - гипобласт (рис. 3-9). Клетки гипобласта выселяются из внутренней клеточной массы вследствие слабого адгезионного взаимодействия между ними. Интенсивно пролиферирующие клетки гипобласта перемещаются по внутренней
поверхности трофобласта и формируют внезародышевую энтодерму прилегающей к трофобласту стенки желточного мешка.
• Эпибласт. Оставшиеся после образования гипобласта клетки внутренней клеточной массы формируют эпибласт (первичная эктодерма). Из него развивается зародыш, а также внезародышевые мезодерма и эктодерма (амниотическая).
• Зародышевый диск (рис. 3-9, рис. 3-10) - основная структура, участвующая в гаструляции. Зародышевый диск (более ранняя стадия - бластодиск) состоит из эпибласта, находящегося в контакте с гипобластом (фактически уже слоем клеток внезародышевой энтодермы желточного мешка). Краниальный отдел зародышевого диска расширен. В дальнейшем (на 15-е сутки) в суженной каудальной части
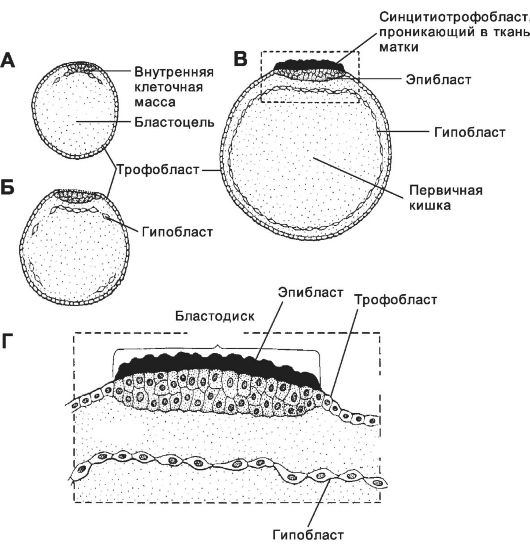 Рис. 3-9. Начало гаструляции. Стадии формирования первичной кишки (А-В), бластодиск - В и Г. Выселяющиеся из гипобласта клетки обрастают изнутри трофобласт, формируя внезародышевую энтодерму. [17]
Рис. 3-9. Начало гаструляции. Стадии формирования первичной кишки (А-В), бластодиск - В и Г. Выселяющиеся из гипобласта клетки обрастают изнутри трофобласт, формируя внезародышевую энтодерму. [17]
зародышевого диска возникает вытянутое в кранио-каудальном направлении утолщение - первичная полоска. Через первичную полоску мигрируют клетки, участвующие в образовании первичных зародышевых листков и внезародышевых структур (рис. 3-10, рис. 3-12). ♦ Зародышевая энтодерма происходит из части эпибласта, расположенной кпереди от первичной полоски.
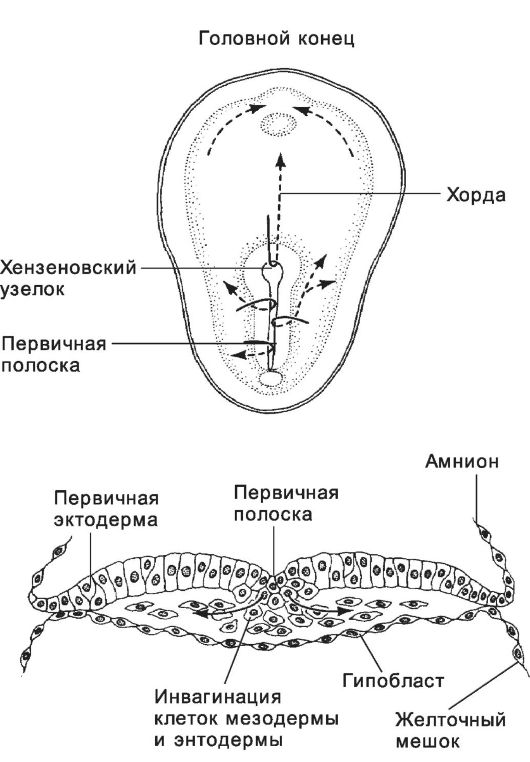 Рис. 3-10. Перемещения клеток при гаструляции. Клетки
первичной эктодермы, проходящие через первичный (хензеновский) узелок,
образуют хорду. Остальные клетки первичной эктодермы, проходящие через
первичную полоску, мигрируют в латеральном направлении и формируют
мезодерму и энтодерму. [17]
Рис. 3-10. Перемещения клеток при гаструляции. Клетки
первичной эктодермы, проходящие через первичный (хензеновский) узелок,
образуют хорду. Остальные клетки первичной эктодермы, проходящие через
первичную полоску, мигрируют в латеральном направлении и формируют
мезодерму и энтодерму. [17]
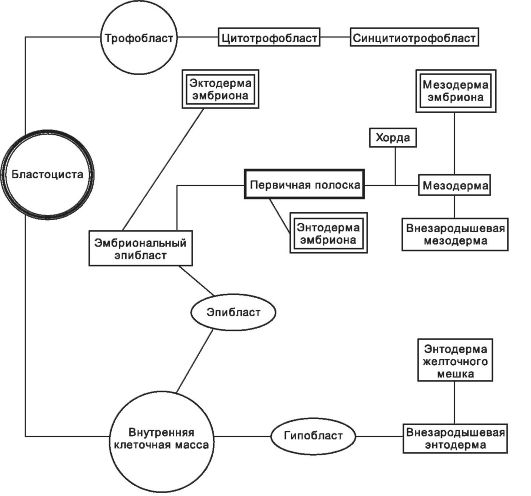 Рис. 3-12. Происхождение зародышевых листков. [17]
Рис. 3-12. Происхождение зародышевых листков. [17]
♦ Зародышевая и внезародышевая мезодерма. Через заднюю часть первичной полоски мигрируют клетки эпибласта, образующие мезодерму. Сначала перемещаются клетки будущей внезародышевой мезодермы, а затем клетки для мезодермы зародыша. Хорду формируют мезодермальные клетки, расположенные вдоль срединной линии зародыша. Они мигрируют через головной конец первичной полоски в области первичного узелка по направлению к будущей головной части зародыша.
♦ Внезародышевая эктодерма. Из периферических участков эпибласта выселяются клетки, выстилающие изнутри полярный трофобласт.
♦ Эктодерма зародыша образуется за счёт не выселяющихся из эпибласта клеток.
Нейруляция и сомиты
В результате гаструляции формируются зародышевые листки, части которых, влияя друг на друга, индуцируют образование новых структур. Пример подобного влияния - первичная эмбриональная индукция; её результат - развитие из дорсальной эктодермы нервной системы (ткани). Нейруляция - процесс закладки нервной системы и осевых структур. Нейруляция начинается с 16-х суток развития (первые признаки формирования нервной пластинки) и в основном завершается к 23-м суткам. Почти одновременно из мезодермы формируются сомиты и нефротом.
Нейруляция
Стадии нейруляции (рис. 3-13) - индукция (первичная эмбриональная индукция) нервной пластинки → приподнимание краёв нервной пластинки и образование нервного желобка → появление нервных валиков - формирование нервного гребня и начало выселения из него клеток - смыкание нервных валиков с образованием нервной трубки - срастание эктодермы над нервной трубкой. Некоторые структуры нервной ткани развиваются из нейрогенных плакод.
• Первичная эмбриональная индукция. Нейральная, или первичная эмбриональная индукция - образование нервной пластинки из дорсальной эктодермы. Этот процесс определяет организатор - хордомезодерма. В ходе первичной эмбриональной индукции детерминируется судьба клеток, дающих начало нервной системе.
• Нервная пластинка - утолщённая часть дорсальной эктодермы, формирующаяся по кранио-каудальному градиенту. Призматические клетки только что сформированной нервной пластинки расположены на базальной мембране, содержащей фибронектин, сульфатированные гликозаминогликаны и ламинин. Клетки нервной пластинки в апикальной части соединены при помощи плотных контактов, а в базальной части - щелевых.
• Нервная трубка. Вскоре после образования края нервной пластинки приподнимаются, и формируются нервные валики (рис. 3-13). Между валиками расположен нервный желобок. Позднее края нервных валиков смыкаются по срединной линии, и образуется замкнутая нервная трубка. Краниальный и каудальный участки нервной трубки долго остаются незамкнутыми, их называют соответственно передним и задним нейропором. Передний нейропор закрывается на 23-26-й день развития, а задний - на 26-30-й день.
• Нервный гребень. После смыкания валиков и образования нервной трубки часть эктодермы, расположенная между нейральной и не-
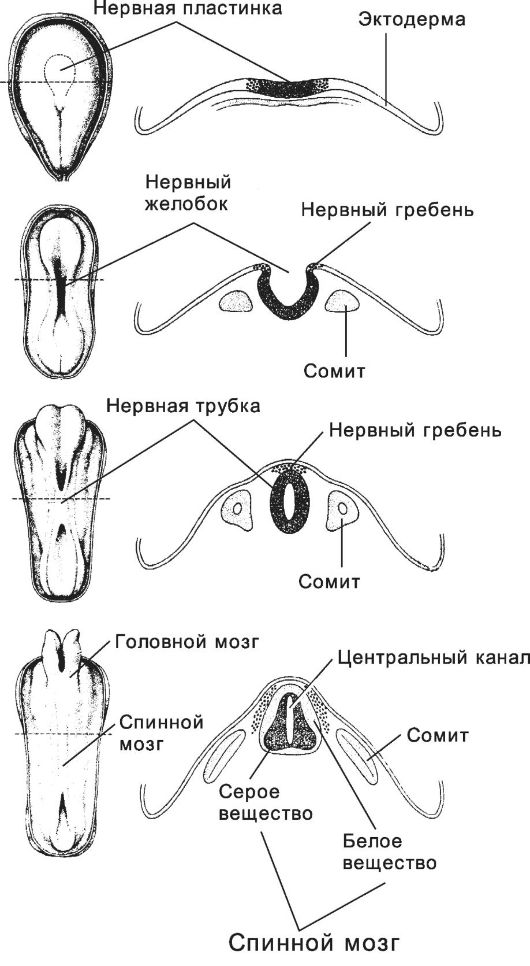 Рис. 3-13. Развитие нервной системы. Последовательные
стадии (сверху вниз): нервная пластинка (19 сут), нервный желобок (20
сут), нервная трубка (22 сут) и зачаток ЦНС (24 сут). Слева - дорсальная
поверхность зародыша, справа - дорсальная часть зародыша в поперечном
разрезе на уровне, обозначенном пунктиром слева. [17]
Рис. 3-13. Развитие нервной системы. Последовательные
стадии (сверху вниз): нервная пластинка (19 сут), нервный желобок (20
сут), нервная трубка (22 сут) и зачаток ЦНС (24 сут). Слева - дорсальная
поверхность зародыша, справа - дорсальная часть зародыша в поперечном
разрезе на уровне, обозначенном пунктиром слева. [17]
нейральной (кожной) эктодермой (рис. 3-13), формирует новую структуру - нервный гребень, его производные см. в таблице 3-3.
Таблица 3-3. Производные нервного гребня
Чувствительные нейроны спинномозговых узлов и ганглиев черепных нервов: верхнего ганглия языкоглоточного нерва, яремного ганглия, часть нейронов ганглия тройничного нерва, часть нейронов ганглия коленца промежуточного нерва
Симпатические нейроны Парасимпатические нейроны
Шванновские клетки и клетки-сателлиты спинномозговых узлов и ганглиев черепных нервов
Меланоциты
Клетки каротидного тельца Продуцирующие кальцитонин клетки Хромаффинные клетки
Хрящ, кости, мышцы и соединительная ткань лица
Окологлоточная мезенхима
Верхне- и нижнечелюстной отростки
Подъязычные дуги и 3-я глоточная дуга
Одонтобласты
Эндотелий роговицы
• Нейрогенные плакоды - утолщения эктодермы, расположенные латерально по обе стороны от формирующейся нервной трубки в краниальном отделе зародыша.
Мезодерма и сомиты
Клетки зародышевой мезодермы выселяются из эпибласта и формируют пресомитную мезодерму, из которой возникают сомиты - симметричные парные структуры по бокам от хорды и нервной трубки (рис. 3-16). Из мезодермы образуется ещё два крупных зачатка: нефротом (промежуточная мезодерма) и латеральная мезодерма. Производные этих структур, а также всех зародышевых листков см. на рисунке 3-12.
• Пресомитная мезодерма. Клетки, прошедшие через первичную полоску, мигрируют в латеральном направлении и образуют непрерывный пласт толщиной в несколько клеток. В непосредственной близости от нервной трубки и хорды мезодермальные клетки образуют скопление - концентрические слои клеток метамерной организации в виде потенциальных сомитов, или сомитомеров. Сомитомеры определяют сегментацию хорды, нервной трубки, промежуточной и латеральной мезодермы (мезодермы латеральной пластинки).
• Сомиты. В результате пролиферации клеток, их миграции и последующей агрегации из сомитомеров формируется дорсальная мезодерма - сомиты (рис. 3-16). Образование сомитов происходит от головного к хвостовому концу зародыша. Новая пара сомитов образуется примерно каждые 6,6 часов. В сомите существует полость, ограниченная клетками, связанными между собой при помощи плотных контактов. В каждом сомите различают склеротом, дерматом и миотом; их клетки имеют свои пути миграции и служат источником для различных структур.
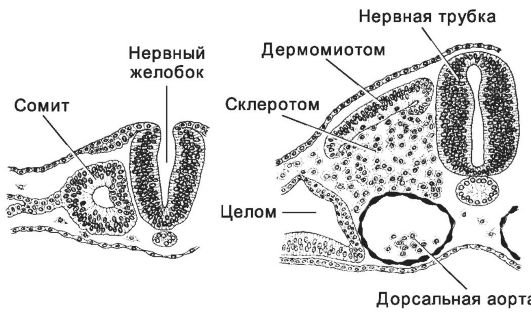 Рис. 3-16. Образование сомита и последующее выселение из него клеток. Слева - мезодермальные клетки сосредоточены латеральнее нервного желобка вокруг небольшой полости; справа -
клетки вентральной и медиальной части сомита, расположенного
латеральнее нервной трубки, начинают мигрировать в направлении хорды;
совокупность этих клеток - склеротом. [17]
Рис. 3-16. Образование сомита и последующее выселение из него клеток. Слева - мезодермальные клетки сосредоточены латеральнее нервного желобка вокруг небольшой полости; справа -
клетки вентральной и медиальной части сомита, расположенного
латеральнее нервной трубки, начинают мигрировать в направлении хорды;
совокупность этих клеток - склеротом. [17]
♦ Склеротом. Под влиянием хорды и нервной трубки клетки вентро-медиальной области сомитов (склеротом) интенсивно размножаются и выселяются из сомита, окружая хорду и вентральную часть нервной трубки. Выселившиеся клетки дифференцируются в хрящевые и образуют позвонки, рёбра и лопатки.
♦ Миотом и дерматом. В оставшейся дорсо-латеральной части сомита выделяют миотом (внутренний слой клеток, образующий впоследствии скелетную мускулатуру) и дерматом (наружный слой, зачаток соединительнотканной части кожи).
• Нефротом. Латеральнее сомита находится скопление клеток промежуточной мезодермы (нефротом) - зачаток мочевыделительной и половой систем.
• Латеральная мезодерма. Расположенная латеральнее нефротома мезодерма (мезодерма латеральной пластинки) расщеплена на два
листка: дорсальный и вентральный. Дорсальный (париетальный) листок - соматическая мезодерма (из неё образуются серозные оболочки). Вентральный (висцеральный) листок - спланхническая мезодерма (из неё образуются сердце, кора надпочечников, строма гонад, соединительная и гладкомышечная ткани внутренних органов и кровеносных сосудов).
Провизорные органы
• На ранних стадиях развития закладываются провизорные органы: хорион, амнион, желточный мешок и аллантоис. Они образуют оболочки зародыша, связывают его с организмом матери и выполняют некоторые специальные функции. Источниками провизорных органов являются структуры бластоцисты, в том числе гипобласт и трофобласт. Гипобласт. Бластоциста состоит из внутренней клеточной массы (эмбриобласт) и трофобласта (рис. 3-9, рис. 3-21). На 8-9-е сутки внутренняя клеточная масса расслаивается на эпибласт (первичная эктодерма) и гипобласт (первичная энтодерма). Клетки гипобласта не принимают участия в образовании структур плода, их потомки присутствуют исключительно в составе провизорных органов (см. рис. 3-22, 3-23 и 3-24). Внезародышевая энтодерма формирует внутренний слой желточного мешка и аллантоиса. Внезародышевая экто-
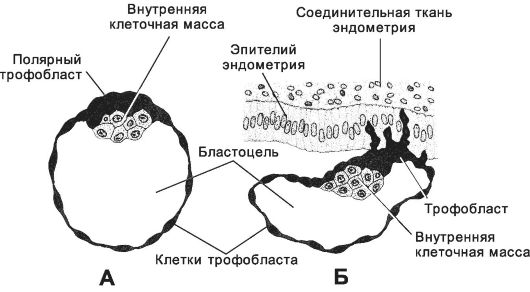 Рис. 3-21. Имплантация. А - бластоциста перед имплантацией (4,5 сут); Б -
начало имплантации. Клетки трофобласта, расположенные на эмбриональном
полюсе бластоцисты (полярная область), на 5-6-е сутки проникают в
слизистую оболочку матки. [17]
Рис. 3-21. Имплантация. А - бластоциста перед имплантацией (4,5 сут); Б -
начало имплантации. Клетки трофобласта, расположенные на эмбриональном
полюсе бластоцисты (полярная область), на 5-6-е сутки проникают в
слизистую оболочку матки. [17]
• дерма участвует в образовании внутреннего слоя амниона. Внезародышевая мезодерма разделяется на внутренний и наружный листки. Внутренний листок вместе с трофобластом формируют хорион, при этом клетки внезародышевой мезодермы обрастают трофобласт, образуя эндоцеломическую полость, или полость хориона. Наружный листок внезародышевой мезодермы участвует в формировании наружных слоёв амниона, желточного мешка и аллантоиса. Трофобласт (рис. 3-22). В трофобласте различают полярную область, покрывающую внутреннюю клеточную массу, и пристеночную (муральную) часть, образующую бластоцель. Клетки мурального трофобласта устанавливают контакт с материнской тканью в имплантационной крипте эндометрия матки. В трофобласте развиваются два слоя: внутренний (цитотрофобласт) и наружный (синцитиотрофобласт). ♦ Цитотрофобласт (слой Лангханса) состоит из интенсивно размножающихся клеток. Их ядра содержат хорошо различимые ядрыш-
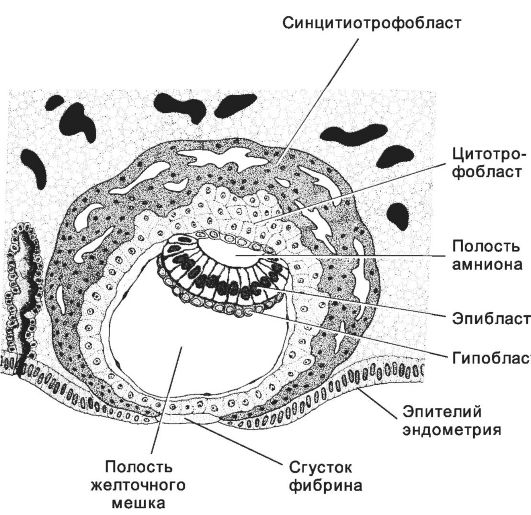 Рис. 3-22. Погружённая в эндометрий бластоциста. Внутренняя
часть трофобласта представлена цитотрофобластом. В наружной части
трофобласта (синцитиотрофобласт) имеется большое количество лакун.
Внутренняя клеточная масса расслоилась на эпи- и гипобласт. [17]
Рис. 3-22. Погружённая в эндометрий бластоциста. Внутренняя
часть трофобласта представлена цитотрофобластом. В наружной части
трофобласта (синцитиотрофобласт) имеется большое количество лакун.
Внутренняя клеточная масса расслоилась на эпи- и гипобласт. [17]
ки, а клетки - многочисленные митохондрии, хорошо развитые гранулярную эндоплазматическую сеть и комплекс Гольджи. В цитоплазме содержится масса свободных рибосом и гранул гликогена.
♦ Синцитиотрофобласт - высокоплоидная многоядерная структура, образуется из клеток цитотрофобласта и служит источником плацентарного соматомаммотропина (плацентарный лактоген), хорионического гонадотропина (ХГТ) и эстрогенов.
Амнион
Амнион (рис. 3-22, 3-23) - плодный пузырь - объёмистый мешок, заполненный амниотической жидкостью (околоплодными водами). В области пупочного кольца амнион переходит на пуповину и далее на плодную часть плаценты, образуя их эпителиальный покров. Зародышевый (эмбриональный) и плодный периоды развития человека происходят внутри плодного пузыря.
• Образование амниона. Одновременно с расслоением внутренней клеточной массы на эпибласт и гипобласт образуется амниотическая полость, ограниченная эпибластом и внезародышевой (амниотической) эктодермой. В ходе гаструляции клетки внезародышевой мезодермы обрастают амниотическую эктодерму, формируя наружный слой амниона.
• Амниотические складки. На краниальном конце амнион образует головную амниотическую складку. С увеличением размеров зародыша его голова растёт вперёд, в амниотическую складку. Боковые амниотические складки формируются по обе стороны зародыша за счёт краёв головной складки. Хвостовая амниотическая складка образуется на каудальном конце зародыша и растёт в краниальном направлении. Головная, боковые и хвостовая амниотические складки сходятся над зародышем и замыкают амниотическую полость. Место соединения амниотических складок - амниотический шов; здесь образуется исчезающий впоследствии тканевый тяж.
• Амниотическая жидкость. Сформированный амниотический мешок наполняется жидкостью, защищающей зародыш при сотрясении, позволяющей плоду совершать движения и предотвращающей слипание растущих частей тела друг с другом и с окружающими тканями. На 99% амниотическая жидкость состоит из воды, 1% приходится на белки, жиры, углеводы, ферменты, гормоны, неорганические соли, а также эпителиальные клетки амниона, кожи, кишечника, дыхательных и мочевыделительных путей. К концу беременности объём жидкости составляет 700-1000 мл.
Желточный мешок
Желточный мешок - вынесенная за пределы зародыша часть первичной кишки. Стенка желточного мешка состоит из двух слоев: внутренний слой образован внезародышевой энтодермой, а наружный - внезародышевой мезодермой. В период наибольшего развития желточного мешка его кровеносные сосуды отделены от стенки матки тонким слоем ткани, что делает возможным поглощение из матки питательных веществ и кислорода. Внезародышевая мезодерма служит местом эмбрионального кроветворения (гемопоэза). Здесь формируются кровяные островки. Во внезародышевой энтодерме желточного мешка временно (на пути их миграции в зачатки гонад) располагаются примордиальные половые клетки. Позднее складки амниона сдавливают желточный мешок; образуется узкая перемычка, соединяющая его с полостью первичной кишки, - желточный стебелёк. Эта структура удлиняется и вступает в контакт с ножкой тела, содержащей аллантоис (рис. 3-23). Желточный стебелёк и дистальная часть аллантоиса вместе со своими сосудами образуют пупочный канатик, отходящий от зародыша в области пупочного кольца. Желточный стебелёк обычно полностью зарастает к концу 3-го месяца развития плода.
Рис. 3-23. Внезародышевые оболочки. Зародыш находится в полости амниона. Желточный мешок связан с зачатком пищеварительной системы, а с каудальной её частью сообщается аллантоис. Все эти структуры находятся в эндоцеломической полости. Хорион топографически и структурно разделяется на гладкий и ворсинчатый. [17]
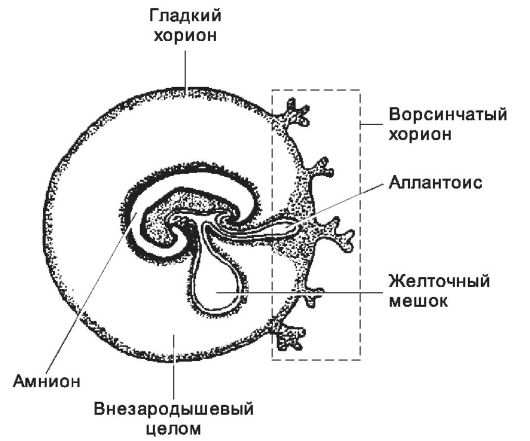 Аллантоис
Аллантоис
Задняя стенка желточного мешка к 16-му дню развития формирует небольшой вырост - аллантоис (греч. alias, колбасовидный), образованный внезародышевыми энтодермой и мезодермой. Устье аллантоиса открывается в клоаку, а его дистальная часть проникает в ножку тела (рис. 3-23). У человека аллантоис рудиментарен, он не функционирует как орган дыхания или резервуар для окончательных продуктов обмена, но имеет важное значение в эмбриональном кроветворении и ангиогенезе.
На 3-5-й неделе развития в стенке аллантоиса происходит гемопоэз и формируются кровеносные сосуды пупочного канатика (две пупочные артерии и одна пупочная вена). На 7-й неделе эмбриогенеза уроректальная перегородка разделяет клоаку на прямую кишку и мочеполовой синус, соединённый с аллантоисом. Поэтому проксимальный отдел аллантоиса имеет отношение к образованию мочевого пузыря. На 2-м месяце эмбриогенеза аллантоис дегенерирует, а на его месте появляется урахус - плотный фиброзный тяж, тянущийся от верхушки мочевого пузыря к пупочному кольцу. В постнатальном периоде урахус организуется в срединную пупочную связку.
Плацента
Плацента связывает плод с организмом матери. Плацента состоит из материнской (базальная часть децидуальной оболочки) и плодной (ворсинчатый хорион) частей.
Имплантация
В полости матки бластоциста находится в течение 1,5-2 суток. Имплантация происходит на 20-21-й день нормального менструального цикла, или через 5,5-6 суток после оплодотворения, когда функциональный слой эндометрия имеет наибольшую толщину (рис. 3-24). В ходе имплантации бластоциста вступает в тесный контакт с эпителием слизистой оболочки матки. Под действием маточного секрета прозрачная оболочка растворяется. После этого бластоциста прикрепляется к эндометрию, как правило, тем полюсом, на котором расположена внутренняя клеточная масса (полярный трофобласт). Через двое суток бластоциста оказывается полностью погружённой в слизистую оболочку матки.
ХОРИОН
Соединение трофобласта и внезародышевой мезодермы приводит к образованию хориона. В формировании хориона различают три периода: предворсинчатый, период образования ворсинок и период котиледонов.
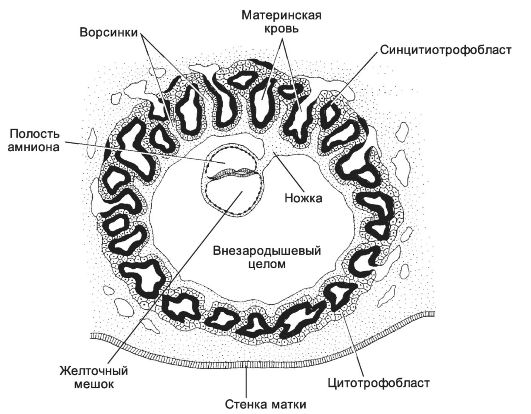 Рис. 3-24. Трёхнедельный эмбрион на стадии гаструлы. Сформированы
полость амниона и желточный мешок. Клетки трофобласта, образующие
плаценту, вступают в контакт с кровеносными сосудами матки. Зародыш
связан с трофобластом происходящей из внезародышевой мезодермы ножкой
тела. В ножку тела прорастает аллантоис, здесь протекает ангиогенез, и в
дальнейшем формируется пупочный канатик с проходящими в его составе
пупочными (аллантоисными) сосудами: двумя пупочными артериями и одной
пупочной веной. [17]
Рис. 3-24. Трёхнедельный эмбрион на стадии гаструлы. Сформированы
полость амниона и желточный мешок. Клетки трофобласта, образующие
плаценту, вступают в контакт с кровеносными сосудами матки. Зародыш
связан с трофобластом происходящей из внезародышевой мезодермы ножкой
тела. В ножку тела прорастает аллантоис, здесь протекает ангиогенез, и в
дальнейшем формируется пупочный канатик с проходящими в его составе
пупочными (аллантоисными) сосудами: двумя пупочными артериями и одной
пупочной веной. [17]
• Предворсинчатый период. В ходе имплантации клетки трофобласта пролиферируют и образуют цитотрофобласт (рис. 3-24, 3-25). По мере взаимодействия с эндометрием трофобласт начинает цитолитически разрушать ткани эндометрия, в результате появляются полости (лакуны), заполненные кровью матери. Лакуны разделены перегородками из клеток трофобласта, это первичные ворсинки. После появления лакун бластоциста может быть названа плодным пузырём.
• Ворсинчатый период. В этот период последовательно образуются первичные, вторичные и третичные ворсинки (рис. 3-25).
♦ Первичные ворсинки - скопления клеток цитотрофобласта, окружённые синцитиотрофобластом.
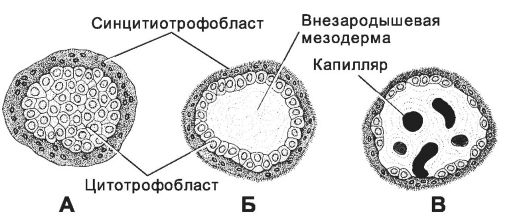 Рис. 3-25. Развитие ворсинок хориона. А - первичная; Б - вторичная; В - третичная ворсинка. [17]
Рис. 3-25. Развитие ворсинок хориона. А - первичная; Б - вторичная; В - третичная ворсинка. [17]
♦ Вторичные ворсинки. На 12-13-й день в первичные ворсинки врастает внезародышевая мезодерма, что приводит к формированию вторичных ворсинок, равномерно распределённых по всей поверхности плодного яйца. Эпителий вторичных ворсинок представлен светлыми клетками округлой формы с крупными ядрами. Над эпителием расположен синцитий с нечёткими границами, тёмной зернистой цитоплазмой, щёточной каёмкой и полиморфными ядрами.
♦ Третичные ворсинки. С 3-й недели развития появляются третичные ворсинки, содержащие кровеносные сосуды. Этот период называют плацентацией. Ворсинки, обращенные к базальной части децидуальной оболочки, кровоснабжаются не только из сосудов, происходящих из хориальной мезодермы, но и из сосудов аллантоиса. Период соединения ветвей пупочных сосудов с местной сетью кровообращения совпадает с началом сердечных сокращений (21-й день развития), и в третичных ворсинках начинается циркуляция эмбриональной крови. Васкуляризация ворсинок хориона заканчивается на 10-й неделе беременности. К этому сроку формируется плацентарный барьер. Не все ворсинки хориона развиты одинаково хорошо. Ворсинки, обращённые к капсулярной части отпадающей оболочки, развиты слабо и постепенно исчезают. Поэтому хорион в этой части называют гладким.
• Период котиледонов. Котиледон - структурно-функциональная единица сформированной плаценты - образован стволовой ворсинкой и её разветвлениями, содержащими сосуды плода. К 140-му дню беременности в плаценте сформировано 10-12 больших, 40-50 мелких и до 150 рудиментарных котиледонов. К 4-му месяцу беременности формирование основных структур плаценты заканчивается (рис. 3-26). Лакуны полностью сформированной плаценты содержат около 150 мл материнской крови, полностью замещаемой 3-4 раза в минуту. Общая поверхность ворсинок достигает 14 м2, что обеспечивает высокий уровень обмена между беременной и плодом.
Децидуальная оболочка
Децидуальная (отпадающая) оболочка образуется на всём протяжении эндометрия. К концу второй недели эндометрий полностью замещает-
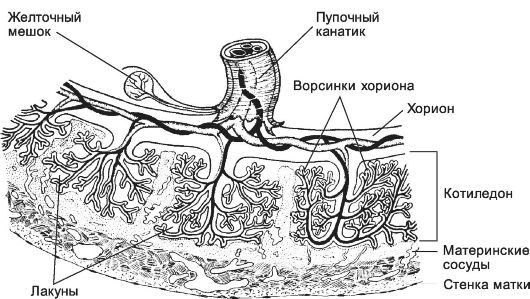 Рис. 3-26. Плацента. С
увеличением срока беременности ворсинки становятся раз- ветвлённее. В
сформированной плаценте присутствуют скопления ворсинок - котиледоны.
Часть ворсинок (закрепляющие, или якорные) более тесно связана с
материнской тканью. [17]
Рис. 3-26. Плацента. С
увеличением срока беременности ворсинки становятся раз- ветвлённее. В
сформированной плаценте присутствуют скопления ворсинок - котиледоны.
Часть ворсинок (закрепляющие, или якорные) более тесно связана с
материнской тканью. [17]
ся децидуальной оболочкой, в которой различают базальную (decidua basalis), капсулярную (decidua capsularis) и пристеночную (decidua parietalis) части. Децидуальная оболочка, окружающая хорион, образует базальную и капсулярную части (рис. 3-27). В остальных областях полость матки выстлана пристеночной децидуальной оболочкой. В децидуальной оболочке выделяют губчатую (спонгиозную) и компактную зоны.
♦ Базальная часть децидуальной оболочки входит в состав плаценты (рис. 3-27). Она отделяет плодное яйцо от миометрия. В губчатом слое много желёз, сохраняющихся до 6-го месяца беременности.
♦ Капсулярная часть. К 18-му дню беременности капсулярная часть полностью смыкается над имплантированным плодным яйцом и отделяет его от полости
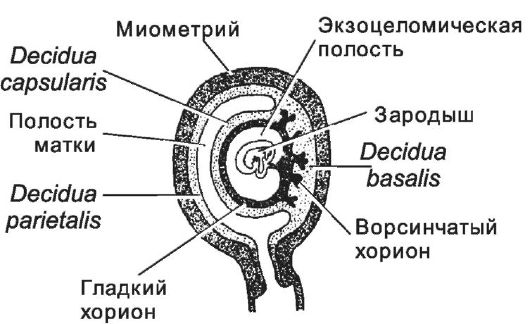 Рис. 3-27. Децидуальная оболочка матки и плацента. Полость
матки выстилает пристеночная часть децидуальной оболочки. Капсулярная и
базальная её части покрывают хорион. Базальная часть децидуальной
оболочки, обращённая к ворсинчатому хориону, входит в состав плаценты.
[17]
Рис. 3-27. Децидуальная оболочка матки и плацента. Полость
матки выстилает пристеночная часть децидуальной оболочки. Капсулярная и
базальная её части покрывают хорион. Базальная часть децидуальной
оболочки, обращённая к ворсинчатому хориону, входит в состав плаценты.
[17]
матки. По мере развития плода капсулярная часть выпячивается в полость матки и к 16-й неделе срастается с пристеночной частью.
♦ Пристеночная часть. До 15-й недели беременности пристеночная часть децидуальной оболочки утолщается за счёт компактной и губчатой зон. K моменту слияния пристеночной и капсулярной частей железы становятся неразличимыми. В конце доношенной беременности пристеночная часть децидуальной оболочки представлена несколькими слоями децидуальных клеток.
♦ Децидуальные клетки. Клетки рыхлой соединительной ткани вокруг сосудов компактной зоны резко увеличены. Это молодые децидуальные клетки, сходные с фибробластами. По мере дифференцировки децидуальные клетки приобретают округлую форму, их ядра становятся светлыми, клетки теснее прилегают друг к другу. Децидуальные клетки продуцируют пролактин и простагландины.
Ворсинчатый хорион
Третичные ворсинки хорошо развиты на стороне, обращённой к миометрию. В ворсинчатом хорионе капилляры приближены к базальной мембране трофобласта и образуют поверхностную капиллярную сеть. Среди клеточных элементов стромы третичных ворсинок присутствуют макрофаги - клетки Хофбауэра. От вершины ворсинок в сторону децидуальной ткани отходят клеточные столбики (колонны), состоящие из клеток цитотрофобласта, контактирующих с поверхностной компактной зоной децидуальной оболочки. В области контакта формируется зона коагуляционного некроза (слой Нитабух). Далее клетки цитотрофобласта проникают в губчатую зону эндометрия, миометрий и стенку сосудов матки. Врастание цитотрофобласта на 6-й неделе беременности в стенку спиральных артерий приводит к вскрытию их просвета и установлению циркуляции материнской крови между ворсинками хориона. Ворсинки, тесно связанные с материнской тканью, называют якорными, или закрепляющими.
Функция плаценты
Функции плаценты многообразны. К ним относятся метаболический обмен между матерью и плодом, транспорт материнских антител, эндокринная функция (синтез ХГТ, прогестерона, хорионического соматомаммотропина, трансферрина, пролактина), детоксикация метаболитов.
Плацентарный барьер
В состав плацентарного барьера (материнская кровь-кровь плода) входят: синцитиотрофобласт → цитотрофобласт → базальная мембрана трофобласта → соединительная ткань ворсинки → базальная мембрана в стенке капилляра плода - эндотелий капилляра плода. Хорион защищает плод от влияния иммунной системы матери.