Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 9 ЭНДОКРИННАЯ СИСТЕМА
Эндокринная система регулирует множество функций разных клеток и органов. Эта регуляторная функция осуществляется при помощи сигнальных молекул - гормонов, вырабатываемых эндокринными клетками, циркулирующих во внутренней среде организма и связывающихся со специфическими рецепторами гормонов на соответствующих клетках-мишенях.
Химия гормонов. По химическому строению различают следующие типы гормонов: олигопептид (например, нейропептиды); полипептид (например, инсулин); гликопротеин (например, тиреотропин); стероид (например, альдостерон и кортизол); производное тирозина (например, йодсодержащие гормоны щитовидной железы: трийодтиронин - Т3 и тироксин - T4); эйкозаноиды (метаболиты арахидоновой кислоты).
Цитология эндокринных клеток. Эндокринные клетки имеют строение, определяемое химической природой синтезируемого гормона.
• Пептиды, белки, гликопротеины, катехоловые амины. Для этих эндокринных клеток характерно наличие гранулярной эндоплазматической сети (здесь происходит сборка пептидной цепи), комплекса Гольджи (присоединение углеводных остатков, формирование секреторных гранул), секреторных гранул.
• Стероидные гормоны. Для клеток, синтезирующих стероидные гормоны, характерно присутствие развитой гладкой эндоплазматической сети и многочисленных митохондрий.
Тропный гормон - гормон, клетками-мишенями которого являются другие эндокринные клетки (например, часть эндокринных клеток передней доли гипофиза синтезирует и секретирует в кровь АКТГ (адренокортикотропный гормон). Мишени АКТГ - эндокринные клетки пучковой зоны коры надпочечников, синтезирующие глюкокортикоиды.
Рилизинг-гормоны (рилизинг-факторы) [от англ. releasing hormone (releasing factor)] - группа синтезируемых в нейронах гипоталамической области мозга гормонов, мишенями которых являются эндокринные клетки передней доли гипофиза (например, рилизинг-гормон для синтезирующих АКТГ клеток передней доли гипофиза - кортиколиберин). Рилизинг-гормоны подразделяют на либерины и статины.
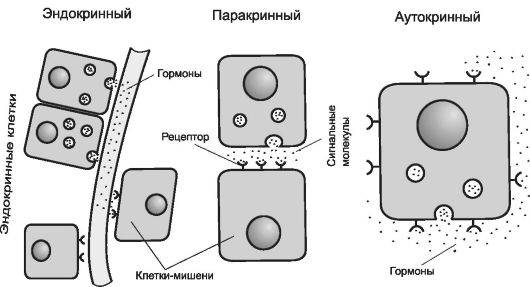 Рис. 9-5. Варианты воздействия гормонов-лигандов на клетки-мишени.
Рис. 9-5. Варианты воздействия гормонов-лигандов на клетки-мишени.
• Либерин - рилизинг-гормон, способствующий усилению синтеза и секреции соответствующего гормона в эндокринных клетках передней доли гипофиза.
• Статин - рилизинг-гормон, в отличие от либеринов ингибирующий синтез и секрецию гормонов в клетках-мишенях.
Варианты эндокринной регуляции. В зависимости от расстояния от продуцента гормона до клетки-мишени различают эндокринный, паракринный и аутокринный варианты регуляции (рис. 9-5).
• Эндокринная, или дистантная регуляция. Секреция гормона происходит во внутреннюю среду, клетки-мишени могут отстоять от эндокринной клетки сколь угодно далеко. Наиболее яркий пример: секреторные клетки эндокринных желёз, гормоны из которых поступают в систему общего кровотока.
• Паракринная регуляция. Продуцент биологически активного вещества и клеткамишень расположены рядом, молекулы гормона достигают мишени путём диффузии в межклеточном веществе. Например, в париетальных клетках желёз желудка секрецию H+ стимулируют гастрин и гистамин, а подавляют соматостатин и простагландины, секретируемые рядом расположенными клетками.
• Аутокринная регуляция. При аутокринной регуляции сама клетка-продуцент гормона имеет рецепторы к этому же гормону (другими словами, клетка-продуцент гормона в то же время является собственной мишенью). В качестве примера приведём эндотелины, вырабатываемые клетками эндотелия и воздействующие на эти же эндотелиальные клетки.
Классификация. Органы эндокринной системы подразделяют на несколько групп:
• гипоталамо-гипофизарная система: нейросекреторные нейроны и аденогипофиз;
• мозговые придатки: гипофиз и эпифиз;
• бранхиогенная группа (имеющая происхождение из эпителия глоточных карманов): щитовидная железа, паращитовидные железы, вилочковая железа;
• надпочечниково-адреналовая система: кора надпочечников, мозговое вещество надпочечников и параганглии;
• островки поджелудочной железы;
• диффузная эндокринная система: эндокринные клетки, рассеянные в различных органах.
ГИПОТАЛАМО-ГИПОФИЗАРНАЯ СИСТЕМА
Эпителиального генеза передняя доля гипофиза (синтез тропных гормонов, экспрессия гена проопиомеланокортина), перикарионы нейросекреторных нейронов гипоталамуса (синтез рилизинг-гормонов, вазопрессина, окситоцина, орексинов), гипоталамо-гипофизарный тракт (транспорт гормонов по аксонам нейросекреторных нейронов), аксо-вазальные синапсы (секреция вазопрессина и окситоцина в капилляры задней доли гипофиза, секреция рилизинг-гормонов в капилляры срединного возвышения), портальная система кровотока между срединным возвышением и передней долей гипофиза в совокупности формируют гипоталамо-гипофизарную систему (рис. 9-6, рис. 9-12).
Гипофиз
Гипофиз анатомически состоит из ножки и тела, а гистологически подразделяется на адено- и нейрогипофиз.
Развитие гипофиза. Гипофиз образуется из двух зачатков эктодермального (карман Ратке) и нейрогенного (processus infundibularis).
• Карман Ратке. На 4-5-й неделе эктодермальный эпителий крыши ротовой бухты образует карман Ратке - вырост, направляющийся к мозгу. Из этого гипофизарного кармана развивается аденогипофиз (передняя, промежуточная и входящая в состав ножки гипофиза туберальная доли).
• Processus infundibularis. Навстречу карману Ратке растёт выпячивание промежуточного мозга, дающее начало нейрогипофизу (задняя доля гипофиза, нейрогипофизарная часть ножки гипофиза и отчасти срединное возвышение).
Кровоснабжение гипофиза. Портальная система кровотока состоит из первичной капиллярной сети срединного возвышения, воротных вен туберальной части аденогипофиза и вторичной капиллярной сети передней доли (рис. 9-9). Приносящие гипофизарные артерии в медиобазальном гипоталамусе (срединное возвышение) образуют первичную капиллярную сеть. Терминали аксонов нейросекреторных клеток
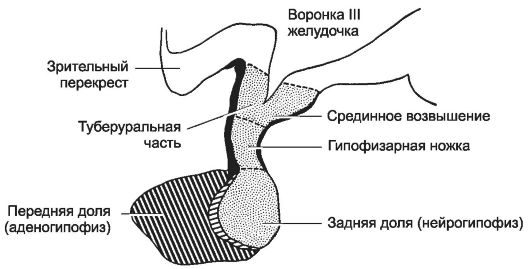 Рис. 9-6. Анатомия гипофиза. [17]
Рис. 9-6. Анатомия гипофиза. [17]
гипоталамуса заканчиваются на этих капиллярах. Кровь из первичной капиллярной сети собирается в портальные вены, идущие по гипофизарной ножке (туберальная часть) в переднюю долю. Здесь портальные вены переходят в капилляры вторичной сети. Обогащённая гормонами передней доли кровь из вторичной капиллярной сети поступает в общую циркуляцию через выносящие вены.
Аденогипофиз (см. рис. 9-6) состоит из передней и промежуточной долей и туберальной части ножки гипофиза. Аденогипофиз покрыт фиброзной капсулой. Передняя доля представлена тяжами эндокринных клеток (аденоцитов), окружённых сетью ретикулиновых волокон. В передней доле ретикулиновые волокна окружают капилляры с фенестрированным эндотелием и широким просветом (синусоиды) вторичной капиллярной сети. Туберальная часть состоит из тяжей эпителиальных клеток, между ними расположены гипофизарные воротные вены (vv. portae hypophysis, см. рис. 9-9), соединяющие первичную капиллярную сеть (срединное возвышение) и вторичную капиллярную сеть (передняя доля гипофиза). Эндокринная функция эпителиальных клеток туберальной части отсутствует, в ней изредка встречаются базофильные аденоциты. Средняя (промежуточная) доля гипофиза у человека выражена слабо.
Аденогипофиз
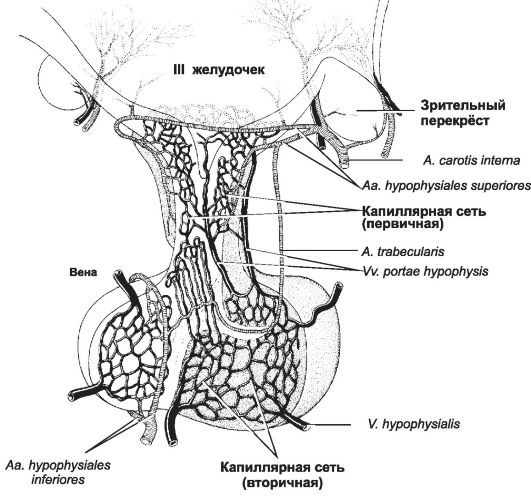 Рис. 9-9. Система кровоснабжения гипофиза. [17] ПЕРЕДНЯЯ ДОЛЯ
Рис. 9-9. Система кровоснабжения гипофиза. [17] ПЕРЕДНЯЯ ДОЛЯ
Передняя доля - эпителиальная эндокринная железа, её клетки синтезируют и секретируют тропные гормоны и продукты экспрессии гена проопиомеланокортина. Разные эндокринные клетки передней доли синтезируют различные пептидные гормоны. Эндокринные клетки передней доли содержат элементы гранулярной эндоплазматической сети, комплекс Гольджи, многочисленные митохондрии и секреторные гранулы различного диаметра. Клетки расположены анастомозирующими тяжами и островками между кровеносными капиллярами с фенестрированным эндотелием. В последние выводятся гормоны, а из капилляров к клеткам поступают либерины и статины.
Классификация эндокринных клеток передней доли (аденоцитов) основана на связывании стандартных красителей, по этому признаку различают хромофильные (базофильные и оксифильные) и хромофобные (плохо окрашивающиеся) клетки. Хромофобные клетки - гетерогенная популяция, включающая дегранулировавшие клетки (оксифилы и базофилы разных типов) и камбиальный резерв. Регенерация аденоцитов происходит из клеток камбиального резерва.
• Базофильные аденоциты подразделяют на кортикотрофы, тиротрофы и гонадотрофы.
♦ Кортикотрофы экспрессируют ген проопиомеланокортина и содержат гранулы диаметром около 200 нм.
♦ Тиротрофы синтезируют тиреотрофный гормон (ТТГ) и содержат мелкие (около 150 нм) гранулы.
♦ Гонадотрофы синтезируют фолликулостимулирующий гормон (фоллитропин) и лютропин, размеры гранул варьируют от 200 до 400 нм. Фоллитропин и лютропин синтезируются в разных подтипах гонадотрофов.
• Ацидофильные аденоциты синтезируют, накапливают в гранулах и секретируют соматотрофин (гормон роста) и пролактин.
♦ Соматотрофы имеют гранулы диаметром до 400 нм.
♦ Лактотрофы содержат мелкие (около 200 нм) гранулы. При беременности и лактации величина гранул может достигать 600 нм.
В передней доле синтезируются СТГ (соматотрофный гормон, соматотро[ф][п]ин, гормон роста), ТТГ (тиреотропный гормон, тиротрофин), АКТГ (адренокортикотропный гормон), гонадотропины (гонадотропные гормоны), а именно лютеинизирующий гормон (лютропин) и фолликулостимулирующий гормон (фоллитропин), а также пролактин. Экспрессия гена проопиомеланокортина приводит к синтезу и секреции ряда пептидов (АКТГ, β- и γ-липотропины, α-, β- и γ-меланотропины, β-эндорфин), из которых гормональная функция установлена для АКТГ и меланотропинов; функции остальных пептидов изучены недостаточно.
Гормоны роста
К этой группе относят гипофизарный гормон роста и хорионический соматомаммотрофин.
♦ Гормон роста гипофизарный (СТГ, соматотрофин, соматотрофный гормон) нормально экспрессируется только в ацидофильных клетках (соматотрофах) передней доли гипофиза.
♦ Хорионическийсоматомаммотрофинсинтезируетсяв клеткахсинцитиотрофобласта. Этот гормон известен также как плацентарный лактоген.
Нативный гормон роста - полипептидная цепь, состоящая из 191 аминокислотных остатков. Синтез и секрецию СТГ стимулирует соматолиберин, а подавляет соматостатин. Эффекты гормона роста
опосредуют соматомедины (инсулиноподобные факторы роста, IGF), синтезируемые преимущественно в гепатоцитах. СТГ - анаболический гормон, стимулирующий рост всех тканей. Наиболее очевидны эффекты СТГ на рост длинных трубчатых костей.
Меланокортины и АКТГ
Адренокортикотропный гормон, α-, β- и γ-меланоцитостимулирующие гормоны (меланотропины), липотропины и β-эндорфин образуются из молекулы-предшественника - проопиомеланокортина (POMC). Продукты гена POMC все вместе называют меланокортинами. Адренокортикотропный гормон. АКТГ состоит из 39 аминокислот. Синтез АКТГ осуществляют кортикотрофы преимущественно передней и в меньшей степени промежуточной доли гипофиза, а также некоторые нейроны ЦНС. Гипоталамический кортиколиберин стимулирует синтез и секрецию АКТГ, а АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (главным образом, глюкокортикоидов).
Гонадотропные гормоны
В эту группу входят гипофизарные фоллитропин и лютропин, а также хорионический гонадотропин (ХГТ) плаценты. Гонадотропные гормоны, а также тиротропин (ТТГ) - гликопротеины, состоящие из двух субъединиц (СЕ). Структура α-СЕ фоллитропина, лютропина, ХГТ и ТТГ идентична, а структура β-СЕ тех же гормонов различна. Гипоталамический гонадолиберин стимулирует синтез и секрецию фоллитропина и лютропина в базофилах (гонадотрофы) передней доли гипофиза. Фоллитропин (фолликулостимулирующий гормон). α-Ингибин - пептидный гормон, вырабатываемый зернистыми клетками фолликулов яичника и сустентоцитами яичка, - подавляет секрецию фоллитропина. Фоллитропин, как и лютропин, регулирует овариальный цикл у женщин. У мужчин мишени фоллитропина - сустентоциты яичка (регуляция сперматогенеза).
Лютропин (лютеинизирующий гормон). У женщин лютропин, как и фоллитропин, регулирует овариальный цикл и эндокринную функцию яичников. У мужчин лютропин стимулирует синтез тестостерона в интерстициальных эндокриноцитах яичек.
Хорионический гонадотропин (ХГТ) - гликопротеин, синтезируемый клетками трофобласта с 10-12 дней развития. При беременности ХГТ взаимодействует с клетками жёлтого тела (синтезирующего и секретирующего прогестерон) яичников.
Тиреотропный гормон
Тиротропин (тиреотропный гормон, ТТГ) синтезируется в базофильных клетках (тиротрофы) передней доли гипофиза. Соматостатин по-
давляет секрецию ТТГ, а гипоталамический тиролиберин стимулирует синтез и секрецию ТТГ. Гормоны щитовидной железы (Т3 и Т4), циркулирующие в крови, регулируют секрецию ТТГ по принципу отрицательной обратной связи. Увеличение содержания свободных T4 и T3 подавляет секрецию ТТГ. Уменьшение содержания свободных T4 и T3 стимулирует секрецию тиротропина. Рецептор ТТГ экспрессируется в фолликулярных клетках щитовидной железы, а также в ретробульбарных тканях. Тиротропин стимулирует дифференцировку эпителиальных клеток щитовидной железы (кроме т.н. светлых клеток, синтезирующих тирокальцитонин) и их функциональное состояние (включая синтез тироглобулина и секрецию Т3 и Т4).
Пролактин
Синтез пролактина происходит в ацидофильных аденоцитах (лактотрофах) передней доли гипофиза. Количество лактотрофов составляет не менее трети всех эндокринных клеток аденогипофиза. При беременности объём передней доли удваивается за счёт увеличения числа лактотрофов и их гипертрофии. Пролактиностатин подавляет секрецию пролактина из лактотрофов. Дофамин ингибирует синтез и секрецию пролактина. Тиролиберин стимулирует секрецию пролактина из лактотрофов. Стимуляция соска и околососкового поля увеличивает секрецию пролактина. Главная функция пролактина - регулирование функции молочной железы.
Нейрогипофиз
Нейрогипофиз (задняя доля гипофиза и нейрогипофизарная часть ножки гипофиза) состоит из клеток нейроглии - питуицитов и кровеносных сосудов. Собственная эндокринная функция питуицитов неизвестна, но нейрогипофиз содержит аксоны гипоталамо-гипофизарного тракта и их окончания на кровеносных капиллярах (аксо-вазальные синапсы). Эти аксоны принадлежат нейронам, расположенным в паравентрикулярном и супраоптическом ядрах гипоталамуса (рис. 9-12). Большие нейроны этих ядер продуцируют вазопрессин и окситоцин, которые по аксонам транспортируются в заднюю долю, где и происходит их высвобождение из нейросекреторных клеток. Следовательно, задняя доля, как и передняя, служит местом выделения пептидных гормонов из гипоталамуса.
Аксо-вазальные синапсы образованы терминальными расширениями аксонов нейросекреторных нейронов гипоталамуса, контактирующими со стенкой кровеносных капилляров срединного возвышения и задней доли гипофиза. Аксоны имеют локальные утолщения (нейросекреторные тельца), заполненные пузырьками и гранулами с гормонами.
Гипоталамус
Нейросекреторные нейроны гипоталамуса - типичные нервные клетки. В перикарионах этих нейронов синтезируются рилизинг-гормоны, орексины, АДГ, окситоцин и другие гормоны. Такие гормон-продуцирующие нервные клетки входят в состав многих ядер гипоталамуса, в т.ч. надзрительного (n. supraopticus) и околожелудочкового (n. paraventricularis).
Гипоталамо-гипофизарный тракт образован аксонами нейросекреторных нейронов гипоталамуса (рис. 9-12). Синтезируемые в нейросекреторных нейронах гормоны при помощи аксонного транспорта достигают аксо-вазальных синапсов нейрогипофиза.
Гипоталамические рилизинг-гормоны
В нейросекреторных нейронах гипоталамуса синтезируются либерины [гонадолиберин (люлиберин), кортиколиберин, соматолиберин, ти- ролиберин] и статины (меланостатин, пролактиностатин, соматостатин).
• Соматостатин синтезируется многими нейронами ЦНС, δ-клетками панкреатических островков, эндокринными клетками пищеварительного тракта и ряда других внутренних органов. Соматостатин - мощный регулятор функций эндокринной и нервной систем, ингибирует синтез и секрецию множества гормонов и секретов.
• Кортистатин продуцируется ГАМКергическими нейронами коры большого мозга и гиппокампа. Этот пептид связывается с рецепторами соматостатина и обладает общими с соматостатином свойствами.
• Соматолиберин стимулирует секрецию гормона роста в передней доле гипофиза.
• Гонадолиберин и пролактиностатин. Ген LHRH кодирует структуру гонадолиберина и пролактиностатина. Мишени гонадолиберина - гонадотрофы, а пролактиностатина - лактотрофы передней доли гипофиза. Гонадолиберин - ключевой нейрорегулятор репродуктивной функции, стимулирует синтез и секрецию фоллитропина и лютропина в продуцирующих гонадотрофы клетках, а пролактиностатин подавляет секрецию пролактина из лактотрофных клеток передней доли гипофиза.
• Тиролиберин синтезируется многими нейронами ЦНС (в т.ч. нейросекреторными нейронами околожелудочкового ядра). Мишени тиролиберина - тиротрофы и лактотрофы передней доли гипофиза. Тиролиберин стимулирует секрецию пролактина из лактотрофов и секрецию тиротропина из тиротрофов.
• Кортиколиберин синтезируется в нейросекреторных нейронах околожелудочкового ядра гипоталамуса, некоторых других нейронах ЦНС, а также в эндометрии, плаценте, матке, яичнике, яичках, желудке, кишечнике, надпочечниках, щитовидной железе и в коже. Кортиколиберин стимулирует синтез АКТГ и других продуктов экспрессии гена проопиомеланокортина (POMC) клетками аденогипофиза. Кортиколиберин, продуцируемый в матке и плаценте, может играть важную роль в нормальном течении беременности.
• Меланостатин подавляет образование меланотропинов.
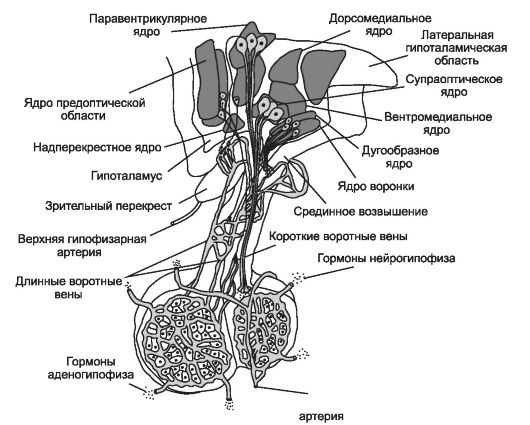
Рис. 9-12. Гипоталамо-гипофизарн^1й тракт. Нейроны с перикарионами больших размеров, локализованные в гипоталамусе, секретируют рилизинг-гормоны в просвет капилляров в области срединного возвышения и воронки, где расположены капилляры первичной сети, собирающие кровь в длинные портальные вены. По ним гипоталамические рилизинг-гормоны поступают в ножку гипофиза и далее в капилляры передней доли (вторичная капиллярная сеть). Аксоны малых нейросекреторных клеток спускаются в ножку гипофиза и выделяют рилизинг-гормоны в капиллярное сплетение, расположенное непосредственно в ножке. Короткие портальные вены переносят рилизинг-гормоны во вторичную капиллярную сеть передней доли. Большие нейроны паравентрикулярного и супраоптического ядер гипоталамуса синтезируют вазопрессин и окситоцин. По аксонам этих нейросекреторных клеток данные гормоны поступают в заднюю долю, где выделяются из нервных терминалей и поступают в просвет многочисленных сосудов, образующих здесь сплетение. [114]
Орексины
В латеральном гипоталамусе расположены нейросекреторные нервные клетки, синтезирующие орексины (гипокретины) A и B. Орексины функционируют как регуляторы сна и бодрствования, участвуют в регуляции пищевого поведения.
Гормоны задней доли
Гормоны задней доли - аргинин вазопрессин (антидиуретический гормон, АДГ), окситоцин, а также нейрофизины - синтезируются в нейросекреторных нейронах надзрительного и околожелудочкового ядер гипоталамуса. Содержащие гормоны мембранные пузырьки транспортируются по аксонам этих нейронов в составе гипоталамогипофизарного тракта в заднюю долю гипофиза и через аксо-вазальные синапсы гормоны секретируются в кровь.
• Окситоцин - циклический нонапептид. Мишени окситоцина - ГМК миометрия и миоэпителиальные клетки молочной железы. Окситоцин стимулирует сокращение ГМК миометрия в родах, при оргазме, в менструальную фазу. Окситоцин стимулирует продукцию и секрецию пролактина, секретируется при раздражении соска и околососкового поля, стимулирует сокращение миоэпителиальных клеток альвеол лактирующей молочной железы (рефлекс молокоотделения). Окситоцин регулирует поведенческую активность, связанную с беременностью и родами.
• Аргинин вазопрессин - нонапетид. Экспрессия АДГ происходит в части нейросекреторных нейронов околожелудочкового и надзрительного ядер гипоталамуса. Секрецию АДГ стимулирует через барорецепторы каротидной области гиповолемия, т.е. уменьшение объёма циркулирующей крови, а ингибируют алкоголь, α-адренергические агонисты, глюкокортикоиды. Аргинин вазопрессин оказывает антидиуретический (регулятор реабсорбции воды в собирательных трубочках почки) и сосудосуживающий (вазоконстриктор) эффекты. Главная функция АДГ - регуляция обмена воды (поддержание постоянного осмотического давления жидких сред организма).
• Нейрофизины I и II кодируются генами окситоцина и АДГ соответственно. Нейрофизины относят к связывающим окситоцин и АДГ белкам.
ЭПИФИЗ
Шишковидная железа - небольшой (5-8 мм) конической формы вырост промежуточного мозга, соединённый ножкой со стенкой третьего желудочка. Капсула органа образована соединительной тканью мягкой мозговой оболочки. От капсулы отходят перегородки, содержащие кровеносные сосуды и сплетения симпатических нервных волокон. Эти перегородки частично подразделяют тело железы на дольки. Паренхима органа состоит из пинеалоцитов и интерстициальных (глиальных) клеток. В интерстиции присутствуют отложения солей кальция, известные как «мозговой песок» (corpora arenacea). Иннервация: орган снабжён многочисленными постганглионарными нервными волокнами от верхнего шейного симпатического узла. Функция органа у человека изучена слабо, хотя железа у ряда позвоночных выполняет различные функции [например, у некоторых амфибий и рептилий эпифиз содержит фоторецепторные элементы (т.н. теменной глаз)], иногда бездоказательно переносимые на человека. Эпифиз у человека,
скорее всего, - звено реализации биологических ритмов, в т.ч. околосуточных.
Пинеалоциты содержат крупное ядро, хорошо развитую гладкую эндоплазматическую сеть, элементы гранулярной эндоплазматической сети, свободные рибосомы, комплекс Гольджи, множество секреторных гранул, микротрубочки и микрофиламенты. Многочисленные длинные отростки пинеалоцитов заканчиваются расширениями на капиллярах и среди клеток эпендимы. Пинеалоциты синтезируют гормон мелатонин и серотонин.
• Мелатонин (N-ацетил-5-метокситриптамин) секретируется в цереброспинальную жидкость и в кровь преимущественно в ночные часы.
• Серотонин (5-гидрокситриптамин) синтезируется преимущественно в дневные часы. Интерстициальные клетки напоминают астроциты, имеют многочисленные ветвящиеся отростки, округлое плотное ядро, элементы гранулярной эндоплазматической сети и структуры цитоскелета: микротрубочки, промежуточные филаменты и множество микрофиламентов. Циркадианный ритм, или околосуточный ритм - один из биологических ритмов (суточная, помесячная, сезонная и годовая ритмика), скоординированный с суточной цикличностью вращения Земли; несколько не соответствует 24 часам. Многие процессы, в т.ч. гипоталамическая нейросекреция, подчиняются околосуточному ритму. Механизмы околосуточного ритма. Изменения освещённости через зрительный тракт оказывают влияние на разряды нейронов надперекрестного ядра (nucleus suprachiasmaticus) ростро-вентральной части гипоталамуса. Надзрительное ядро содержит т.н. эндогенные часы - неизвестной природы генератор биологических ритмов (включая околосуточный), контролирующий продолжительность сна и бодрствования, пищевое поведение, секрецию гормонов и т.д. Сигнал генератора - гуморальный фактор, секретируемый из надзрительного ядра (в т.ч. в цереброспинальную жидкость). Сигналы от надзрительного ядра через нейроны околожелудочкового ядра (n. paraventricularis) активируют преганглионарные симпатические нейроны боковых столбов спинного мозга. Симпатические преганглионары активируют нейроны верхнего шейного узла. Постганглионарные симпатические волокна от верхнего шейного узла секретируют норадреналин, взаимодействующий с α- и β-адренорецепторами плазмолеммы пинеалоцитов. Активация адренорецепторов приводит к увеличению внутриклеточного содержания цАМФ и экспрессии гена CREM, а также к транскрипции арилалкиламин-N-ацетилтрансферазы, фермента синтеза мелатонина.
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа секретирует регуляторы основного обмена - йодсодержащие гормоны - трийодтиронин (Т3) и тироксин (Т4), а также кальцитонин, один из эндокринных регуляторов обмена Ca2+. Йодсодержащие гормоны вырабатывают эпителиальные клетки стенки фолликулов, кальцитонин - светлые клетки.
Развитие. Эпителий бранхиогенной группы желёз (щитовидная, вилочковая, околощитовидные) развивается из энтодермы глоточных карманов. В конце 3-го месяца развития плода начинается синтез йодсодержащих гормонов, появляющихся в амниотической жидкости. Синтезирующие кальцитонин светлые клетки (С-клетки) щитовидной железы развиваются из нервного гребня.
ПАРЕНХИМА
Паренхима щитовидной железы - совокупность секретирующих тиреоидные гормоны клеток и С-клеток, синтезирующих кальцитонин. И те, и другие входят в состав фолликулов и скоплений межфолликулярных клеток.
Тиреоциты и йодсодержащие гормоны
Фолликулы - различной величины и формы (преимущественно округлые) пузырьки, содержащие коллоид. Стенка фолликула образована эпителиальными фолликулярными клетками (продукция йодсодержащих гормонов), прикреплёнными к базальной мембране. Между базальной мембраной и фолликулярными клетками встречаются более крупные светлые клетки (синтез кальцитонина). Фолликулярные клетки, или тироциты образуют стенку фолликула и формируют его содержимое, синтезируя и секретируя в коллоид тироглобулин. Фермент тиропероксидаза и рецептор N-ацетилглюко- замина также синтезируются в фолликулярных клетках. Основная функция фолликулярных клеток - синтез и секреция T4 и T3 - складывается из многих процессов: образование тироглобулина → секреция тироглобулина в полость фолликула → поглощение йода из крови - окисление йода - йодирование тироглобулина в полости фолликула → эндоцитоз и расщепление тироглобулина → секреция T3 и T4. Функцию фолликулярных клеток стимулирует тиротропин (ТТГ). Форма клеток (от низкой кубической до цилиндрической) эпителиальной стенки фолликула зависит от интенсивности их функционирования: высота клеток пропорциональна напряжённости осуществляемых в них процессов.
• Базальная часть клеток содержит ядро, гладкий и шероховатый эндоплазматический ретикулум. В плазмолемму встроены связанные с G-белком рецепторы ТТГ, Na+/I--котранспортёр. Возможна складчатость плазмолеммы (отражает интенсивность обмена между клетками и кровеносными капиллярами - захват йода, поступление метаболитов, секреция гормонов).
• Латеральная часть клеток содержит межклеточные контакты, предупреждающие просачивание коллоида.
• Апикальная часть содержит выраженный комплекс Гольджи (формирование секреторных пузырьков, присоединение углеводов к тироглобулину), разные типы пузырьков [секреторные (содержат тироглобулин), окаймлённые (незрелый тироглобулин из полости фолликула поступает в клетку для рециклизации и выве-
дения в кровоток), эндоцитозные (содержат зрелый тироглобулин для его последующей деградации в фаголизосомах)], микроворсинки (увеличение поверхности обмена между клетками и полостью фолликула). Апикальная плазмолемма содержит рецепторы N-ацетилгалактозамина (связывание незрелого тироглобулина для его интернализации путём опосредованного этими рецепторами эндоцитоза), рецепторы мегалина (интернализация, трансцитоз и секреция в кровь тироглобулина), анионообменники (перемещение йода из цитоплазмы клетки в полость фолликула). В связи с мембранными структурами апикальной части клеток находится тиропероксидаза. Продукция йодсодержащих гормонов. Синтез и секреция йодсодержащих гормонов включает несколько этапов (рис. 9-17). Йодсодержащие гормоны. Тироксин (T4) и трийодтиронин (T3) - водонерастворимые соединения, поэтому сразу после секреции в кровь гормоны образуют комплексы с транспортными белками плазмы, которые не только обеспечивают циркуляцию Т3 и Т4 в крови, но и предотвращают деградацию и экскрецию этих гормонов.
• Тироксин (3,5,3',5'-тетрайодтиронин, C15H11I4NO4, Mr 776,87) - основной йодсодержащий гормон, на долю T4 приходится не менее 90% секретируемых щитовидной железой йодсодержащих гормонов.
♦ L-форма тироксина физиологически примерно вдвое активнее рацемической (DL-тироксин), D-форма гормональной активности не имеет.
♦ Дейодирование наружного кольца тироксина приводит к образованию Т3.
♦ Дейодирование внутреннего кольца тироксина приводит к образованию реверсивного Т3 (rT3), имеющего незначительную физиологическую активность.
• Трийодотиронин (3,5,3'-трийодтиронин, C15H12I3NO4, Mr 650,98). На долю T3 приходится лишь 10% содержащихся в крови йодсодержащих гормонов, но физиологическая активность T3 примерно в четыре раза выше, чем тироксина.
• Функции йодсодержащих гормонов многочисленны. Например, Т3 и Т4 увеличивают обменные процессы, ускоряют катаболизм белков, жиров и углеводов, эти гормоны необходимы для нормального развития ЦНС, они стимулируют рост хряща и поддерживают рост кости, увеличивают частоту сердечных сокращений и сердечный выброс. Крайне разнообразные эффекты йодсодержащих гормонов на клетки-мишени (ими практически являются все клетки организма) объясняют увеличением синтеза белков и потребления кислорода.
С-клетки
С-клетки в составе фолликулов называют также парафолликулярными клетками. В них происходит экспрессия кальцитонинового гена CALC1, кодирующего кальцитонин, катакальцин и относящийся к кальцитониновому гену пептид α. С-клетки крупнее тироцитов, в составе фолликулов расположены, как правило, одиночно. Морфология этих клеток характерна для клеток, синтезирующих белок на экспорт (присутствуют шероховатая эндоплазматическая сеть, комплекс Голь-
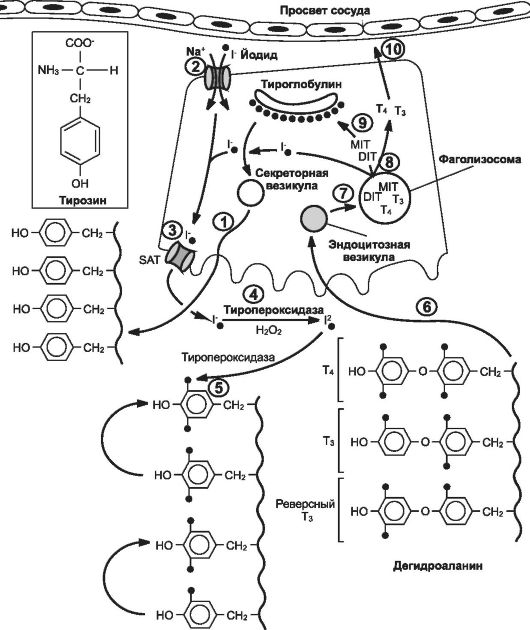 Рис. 9-17. Биосинтез йодсодержащих гормонов. 1. Йод поступает в тироцит через Na+/I--котранспортёр. 2. Из цитоплазмы в полость фолликула йодид транспортируется через анионообменник SAT. 3. На границе апикальной мембраны тироцита и коллоида тиропероксидаза катализирует окисление йодида с образованием молекулы йода. 4. Тиропероксидаза
катализирует йодирование остатков тирозина в молекуле тироглобулина с
образованием монойодтирозина и дийодтирозина. 5. Синтез трийодтиронина и тетрайодтиронина. 6. Интернализация йодированного тироглобулина путём эндоцитоза. 7. Слияние эндоцитозного пузырька с лизосомой и деградация тироглобулина. 8. Высвобождение монойодтирозина, дийодтирозина, Т3 и Т4 в цитоплазму клетки. 9. Дейодирование и реутилизация монойодтирозина и дийодтирозина. 10. Секреция йодсодержащих гормонов в кровь. [114]
Рис. 9-17. Биосинтез йодсодержащих гормонов. 1. Йод поступает в тироцит через Na+/I--котранспортёр. 2. Из цитоплазмы в полость фолликула йодид транспортируется через анионообменник SAT. 3. На границе апикальной мембраны тироцита и коллоида тиропероксидаза катализирует окисление йодида с образованием молекулы йода. 4. Тиропероксидаза
катализирует йодирование остатков тирозина в молекуле тироглобулина с
образованием монойодтирозина и дийодтирозина. 5. Синтез трийодтиронина и тетрайодтиронина. 6. Интернализация йодированного тироглобулина путём эндоцитоза. 7. Слияние эндоцитозного пузырька с лизосомой и деградация тироглобулина. 8. Высвобождение монойодтирозина, дийодтирозина, Т3 и Т4 в цитоплазму клетки. 9. Дейодирование и реутилизация монойодтирозина и дийодтирозина. 10. Секреция йодсодержащих гормонов в кровь. [114]
джи, секреторные гранулы, митохондрии). На гистологических препаратах цитоплазма С-клеток выглядит светлее цитоплазмы тироцитов, отсюда их название - светлые (clear) клетки.
• Кальцит онин - пептид, содержащий 32 аминокислотных остатка.
♦ Регулятор экспрессии - Са2+ плазмы крови, внутривенное его введение существенно увеличивает секрецию кальцитонина.
♦ Функции кальцитонина, как одного из регуляторов кальциевого обмена, определяют как антагонистические функциям гормона паращитовидной железы.
• Катакальцин - пептид, состоящий из 21 аминокислотного остатка, имеет те же функции, что и кальцитонин.
• Относящиеся к кальцитониновому гену пептиды (CGRP) α и β (37 аминокислот) экспрессируются в ряде нейронов ЦНС и периферической нервной системы (особенно в связи с кровеносными сосудами). Их функции - участие в ноцицепции, пищевом поведении, в регуляции тонуса ГМК сосудов (вазодилатация), бронхов (бронхоконстрикция).
Хюртля клетки
Иногда в составе стенки фолликулов или между фолликулами находят крупные клетки с зернистой оксифильной цитоплазмой, содержащие много митохондрий - онкоциты, или клетки Хюртля (Гюртля, также Асканази-Хюртля).
Межфолликулярные клетки
К паренхиме щитовидной железы, помимо образующих фолликулы клеток, относятся также островки клеток, расположенные между фолликулами. Островки образованы способными синтезировать йодсодержащие гормоны клетками (малодифференцированные тироциты, формирующие новые фолликулы), а также С-клетками.
СТРОМА
Строма состоит из вспомогательных структур (капсула, интерстиций, нервные и сосудистые элементы). Капсула сформирована из плотной волокнистой соединительной ткани. От капсулы отходят тяжи (стандартное наименование - септы, или трабекулы) плотной волокнистой соединительной ткани, содержащие кровеносные и лимфатические сосуды, нервы.
Интерстиций. Пространство органа заполняет поддерживающий элементы паренхимы каркас из рыхлой волокнистой соединительной ткани с кровеносными и лимфатическими сосудами, отдельными нервными волокнами и их окончаниями.
Кровоток железы интенсивен и сопоставим с кровоснабжением мозга, перфузией крови через почки и печень. Кровеносные капилляры фенестрированного типа контактируют с эндокринными клетками паренхимы.
Иннервация
• Соматическая чувствительная. В железе найдены чувствительные нервные окончания, образованные ветвлениями периферических отростков чувствительных нейронов.
• Двигательная вегетативная (симпатическая и парасимпатическая). Преобладают сопровождающие кровеносные сосуды и иннервирующие их ГМК варикозные ветвления постганглионарных симпатических нейронов. Эффекты вегетативной иннервации на эндокринную функцию незначительны.
ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Четыре небольшие паращитовидные железы расположены на задней поверхности и под капсулой щитовидной железы. Эпителий нижних двух паращитовидных желёз развивается из энтодермы третьей пары глоточных карманов, верхних двух - из четвёртой пары. Функция желёз - синтез и секреция Са2+-регулирующего пептидного гормона паратиреокрина (паратиреоидного гормона, ПТГ). ПТГ вместе с кальцитонином и катакальцином, а также с витамином D регулирует обмен кальция и фосфатов.
Каждая из четырёх желёз имеет собственную тонкую капсулу, от которой отходят перегородки (септы), содержащие кровеносные сосуды. Паренхима, образованная тяжами и островками эпителиальных клеток, содержит два типа клеток - главные и оксифильные.
• Главные клетки имеют базофильную цитоплазму (развита гранулярная эндоплазматическая сеть), комплекс Гольджи, мелкие митохондрии и секреторные гранулы диаметром 200-400 нм, содержащие
ПТГ.
• Оксифильные клетки равномерно распределены в паренхиме железы или образуют небольшие скопления, содержат крупные митохондрии, слабо выраженный комплекс Гольджи и умеренно развитую гранулярную эндоплазматическую сеть. Функция оксифильных клеток неизвестна, их число с возрастом увеличивается.
• Жировые клетки всегда присутствуют в железе, с возрастом их количество увеличивается.
Паратиреоидный гормон, или паратиреокрин (паратирин, паратгормон, гормон паращитовидной железы, ПТГ, состоит из 84 аминокислотных остатков) поддерживает гомеостаз кальция и фосфатов. Регулятор экспрессии ПТГ - ионы Са2+, взаимодействующие с трансмембранными рецепторами главных клеток паращитовидных желёз. Са2+ сыворотки регулирует секрецию ПТГ по механизму отрицательной обратной связи. Функции. ПТГ поддерживает гомеостаз Ca2+. Паратиреокрин увеличивает содержание Ca2+ в плазме, усиливая его вымывание из костей, реабсорбцию в канальцах почки и всасывание в кишечнике.
НАДПОЧЕЧНИК
Надпочечники (см. рис. 9-24) - парные эндокринные органы, расположенные ретроперитонеально у верхних полюсов почки на уровне Th12 и L1; масса надпочечника - примерно 4 г. Фактически это две железы: кора (на долю коры приходится около 80% массы железы) и мозговая часть. Кора надпочечников синтезирует кортикостероиды (минералокортикоиды, глюкокортикоиды и андрогены), хромаффинная ткань мозговой части - катехоловые амины.
Развитие. На 6-й неделе внутриутробного развития крупные мезодермальные клетки целомического эпителия образуют скопления между основанием дорзальной брыжейки первичной кишки и развивающимися урогенитальными валиками. По направлению к этим скоплениям из ближайших симпатических ганглиев мигрируют клетки нервного гребня - будущие хромаффинные клетки мозгового вещества. В дальнейшем число хромаффинных клеток возрастает вплоть до завершения полового развития. Мезодермальные клетки формируют две зоны коры: наружную - дефинитивную и эмбриональную (фетальную), расположенную на границе с мозговым веществом. Незадолго до рождения начинается дегенерация фетальной коры, и к концу первого года жизни фетальная кора полностью исчезает. В течение первого года жизни в дефинитивной коре различимы клубочковая, пучковая и сетчатая зоны; полностью дифференцировка корковой части надпочечника завершается к третьему году жизни. Регенерация. Клетки коры и мозговой части железы способны поддерживать свою численность как путём их пролиферации, так и за счёт камбиального резерва.
• Кора. Непосредственно под капсулой органа находятся эпителиальные камбиальные клетки, постоянно дифференцирующиеся в эндокринные клетки коры. АКТГ стимулирует пролиферацию камбиального резерва.
• Мозговая часть. Часть мигрировавших сюда клеток нервного гребня сохраняется в виде камбиального резерва. Эти малодифференцированные клетки - источник развития новых хромаффинных клеток.
Кровоснабжение железы осуществляется из трёх источников: верхняя надпочечниковая артерия (ветвь нижней диафрагмальной артерии), средняя надпочечниковая артерия (отходит от аорты), нижняя надпочечниковая артерия (ветвь почечной артерии) (рис. 9-23). Верхняя и средняя надпочечниковые артерии дают начало капиллярам, пронизывающим корковое вещество и заканчивающимися мозговыми венозными синусами в мозговом веществе. Это означает, что гормоны, продуцируемые клетками коркового вещества, покидают кору, проходя через мозговое вещество, при этом глюкокортикоиды коры стимулируют секрецию адреналина из хромаффинных клеток. Это обстоятельство объясняет сочетанное вовлечение органа в развитие стрессовых ситуаций (адаптационный синдром, по Селье). Нижняя надпочечниковая артерия даёт начало мозговой артерии, которая кровоснабжает только мозговое вещество, минуя корковое, и заканчивается на мозговых венозных синусах. Медуллярные венозные синусы открываются в центральную вену.
КОРА НАДПОЧЕЧНИКА
Железа (рис. 9-24) окружена капсулой из плотной волокнистой соединительной ткани, от которой в толщу органа местами отходят соеди-
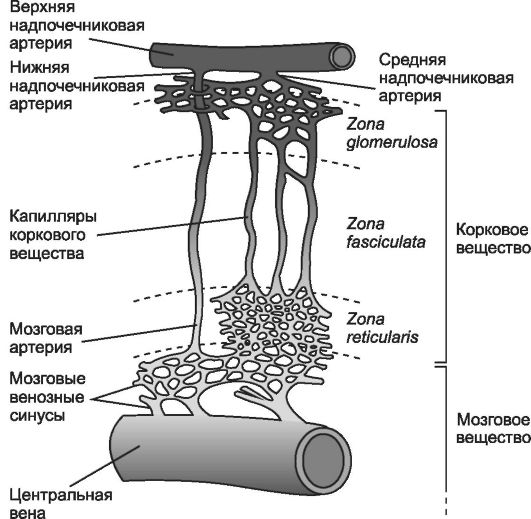 Рис. 9-23. Кровоснабжение надпочечника. [82, 90]
Рис. 9-23. Кровоснабжение надпочечника. [82, 90]
нительнотканные перегородки. Строма железы состоит из поддерживающей эндокринные клетки рыхлой волокнистой соединительной ткани, содержащей огромное количество кровеносных капилляров с фенестированным эндотелием. Паренхима - совокупность эпителиальных тяжей, имеющих различную ориентацию на разном расстоянии от капсулы надпочечника. Это обстоятельство, а также характер гормонального стероидогенеза позволяет выделить в коре клубочковую, пучковую и сетчатую зоны.
Клубочковая зона. Тяжи эндокринных клеток подворачиваются под капсулу и на срезе имеют вид клубочков (15% толщины коры). Здесь синтезируются минералокортикоиды (преимущественно альдостерон). Стимулятор синтеза альдостерона - ангиотензин II и в незначительной степени - АКТГ. Клетки (рис. 9-25Б) имеют плотное округлое ядро с одним или двумя ядрышками, развитую гладкую эндоплазматическую сеть, некрупные митохондрии с пластинчатыми кристами, ри-
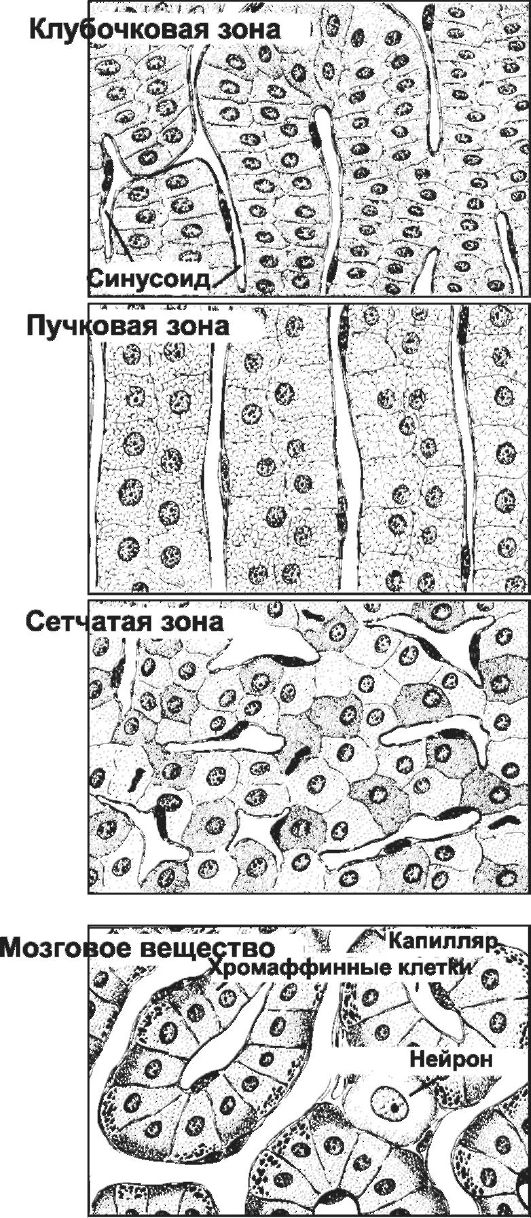 Рис. 9-24. Надпочечник. Непосредственно
под капсулой в составе корковой части находится клубочковая зона. Она
состоит из узких и более мелких по сравнению с другими зонами клеток.
Крупные многоугольные клетки образуют параллельные тяжи пучковой зоны.
Правильный ход тяжей нарушается в сетчатой зоне корковой части
надпочечника. Мозговая часть представлена переплетающимися тяжами
крупных хромаффинных клеток. К тяжам прилегают синусоидные кровеносные
капилляры с широким просветом. [17]
Рис. 9-24. Надпочечник. Непосредственно
под капсулой в составе корковой части находится клубочковая зона. Она
состоит из узких и более мелких по сравнению с другими зонами клеток.
Крупные многоугольные клетки образуют параллельные тяжи пучковой зоны.
Правильный ход тяжей нарушается в сетчатой зоне корковой части
надпочечника. Мозговая часть представлена переплетающимися тяжами
крупных хромаффинных клеток. К тяжам прилегают синусоидные кровеносные
капилляры с широким просветом. [17]
босомы, хорошо развитый комплекс Гольджи и небольшое количество мелких липидных включений.
Пучковая зона занимает около 75% толщины коры. Тяжки эндокринных клеток и находящиеся между ними кровеносные капилляры расположены параллельно друг другу (в виде пучков). Здесь синтезируются глюкокортикоиды (преимущественно кортизол и кортизон), а также андрогены. Синтез глюкокортикоидов регулирует тропный гормон аденогипофиза - АКТГ. Клетки на гистологических препаратах выглядят как вакуолизированные (рис. 9-25А), поэтому их называют спонгиоциты. Вакуолизация клеток на гистологических препаратах отражает присутствие в цитоплазме спонгиоцитов значительного числа липидных капель (содержат преимущественно эфиры холестерина), вымываемых при подготовке препарата. Спонгиоциты содержат округлые митохондрии с кристами в виде трубочек и пузырьков, раз- ветвлённую гладкую эндоплазматическую сеть, элементы гранулярной эндоплазматической сети, лизосомы, многочисленные липидные включения и пигментные гранулы, содержащие липофусцин. Сетчатая зона. В наиболее глубоких частях коры (10% толщины коры) тяжи эндокринных клеток переплетаются, образуя подобие сети. В сетчатой зоне синтезируются глюкокортикоиды и стероидные гормоны типа андрогенов (дегидроэпиандростерон и андростендион). Тропный гормон - АКТГ. Гонадотропные гормоны гипофиза не влияют на секрецию гормонов в сетчатой зоне. В отличие от спонгиоцитов, клетки этой зоны содержат меньше липидных включений, но имеют крупные липофусциновые гранулы. Липофусциновые гранулы содержат лизосомальную кислую фосфатазу и рассматриваются как деградирующие лизосомы.
Стероидогенез гормонов коры надпочечника, а также стероидных гормонов половой сферы - сложный процесс (из железы выделено не менее 50 стероидов), по-разному происходящий в отдельных зонах коры. Стероидные гормоны, их промежуточные продукты, а также фармакологические аналоги гормонов синтезируются на базе холестерина. Процессы стероидогенеза обеспечивают ферменты, локализованные в митохондриях и гладкой эндоплазматической сети.
• Глюкокортикоиды. Основной глюкокортикоид, секретируемый надпочечниками, - кортизол; на его долю приходится 80%. Остальные 20% - кортизон, кортикостерон, 11-дезоксикортизол и 11-дезоксикортикостерон. АКТГ - основной регулятор синтеза глюкокортикоидов. Для синтеза и секреции кортиколиберина, АКТГ и кортизола характерна выраженная суточная периодичность. При нормальном ритме сна увеличение секреции кортизола наступает после засыпания и достигает максимума при пробуждении. Функции глюкокортикоидов разнообразны: от регуляции метаболизма до модификации иммунологического и воспалительного ответов. Наиболее важный метаболический эффект глюкокортикоидов - преобразование жира и мышечных белков в гликоген.
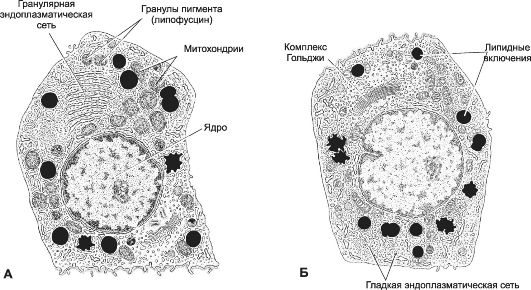 Рис. 9-25. Эндокринные клетки коры надпочечника. А - клетка пучковой зоны, вырабатывающая глюкокортикоиды и андрогены. Клетку называют спонгиоцитом, т.к. она имеет пенистый вид
из-за множества липидных капель в цитоплазме; содержит округлые
митохондрии с кристами в виде трубочек и пузырьков, разветвлённую
гладкую эндоплазматическую сеть. Б - клетка клубочковой зоны,
вырабатывающая альдостерон. Присутствуют развитая гладкая
эндоплазматическая сеть, некрупные митохондрии с пластинчатыми кристами и
небольшое количество мелких липидных включений. [17]
Рис. 9-25. Эндокринные клетки коры надпочечника. А - клетка пучковой зоны, вырабатывающая глюкокортикоиды и андрогены. Клетку называют спонгиоцитом, т.к. она имеет пенистый вид
из-за множества липидных капель в цитоплазме; содержит округлые
митохондрии с кристами в виде трубочек и пузырьков, разветвлённую
гладкую эндоплазматическую сеть. Б - клетка клубочковой зоны,
вырабатывающая альдостерон. Присутствуют развитая гладкая
эндоплазматическая сеть, некрупные митохондрии с пластинчатыми кристами и
небольшое количество мелких липидных включений. [17]
• Минералокортикоиды. Альдостерон - основной минералокортикоид. Другие стероиды надпочечника - кортизол, 11-дезоксикортизол, 11-дезоксикортикостерон, кортикостерон - имеют и минералокортикоидную активность, хотя - сравнительно с альдостероном - их суммарный вклад мал. Ангиотензин II - компонент системы «ренин-ангиотензины» - главный регулятор синтеза и секреции альдостерона. Этот пептид стимулирует выброс альдостерона. Натриуретические факторы ингибируют синтез альдостерона. Функция минералокортикоидов - поддержание баланса электролитов жидкостей организма, осуществляется посредством влияния на реабсорбцию ионов в почечных канальцах.
• Андрогены. В коре надпочечников синтезируются дегидроэпиандростерон и в меньшей степени андростендион.
МОЗГОВАЯ ЧАСТЬ НАДПОЧЕЧНИКА
Эндокринную функцию мозговой части надпочечника выполняют происходящие из нервного гребня хромаффинные клетки. При активации симпатической нервной системы надпочечники выбрасывают в кровь катехоловые амины (адреналин и норадреналин). Катехоламины имеют широкий спектр эффектов (воздействие на гликогенолиз, липолиз, глюконеогенез, существенное влияние на сердечно-сосудистую систему). Вазоконстрикция, параметры сокращения сердечной мышцы и другие эффекты катехоловых аминов реализуются через α- и β-адренергические рецепторы на поверхности клеток-мишеней (ГМК, секреторные клетки, кардиомиоциты). Серьёзные клинические проблемы возникают при опухолях эндокринных клеток и их предшественников (нейробластома, феохромоцитома). Строма. В нежном поддерживающем каркасе, состоящем из рыхлой волокнистой соединительной ткани, расположены многочисленные сосудистые полости - венозные синусы - вариант капилляров типа синусоидов. Их отличительная особенность - значительный диаметр просвета, достигающий десятков и сотен мкм.
Иннервация. Мозговая часть органа содержит множество преганглионарных нервных волокон симпатического отдела нервной системы, хромаффинные клетки расценивают как постганглионарное звено (модифицированные постганглионарные симпатические нейроны) двигательной вегетативной иннервации. Между хромаффинными клетками в мозговом веществе можно также видеть рассеянные небольшие группы ганглионарных клеток с неясной функцией.
Хромаффинные клетки
Хромаффинные клетки (рис. 9-29) содержат гранулы с электронноплотным содержимым, которое с бихроматом калия даёт хромаффинную реакцию. Хромаффинные клетки - основной клеточный элемент мозговой части надпочечников и параганглиев, расположенных
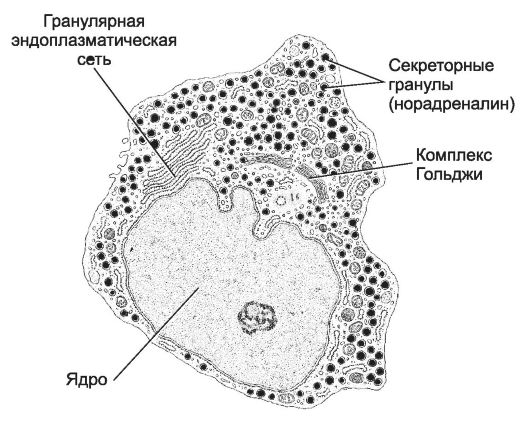 Рис. 9-29. Хромаффинная клетка. Характерны
многочисленные электронно-плотные гранулы с катехоламинами.
Значительный объём клетки занимает крупное ядро. Клетка содержит
митохондрии, выраженный комплекс Гольджи, элементы гранулярной
эндоплазматической сети. [17]
Рис. 9-29. Хромаффинная клетка. Характерны
многочисленные электронно-плотные гранулы с катехоламинами.
Значительный объём клетки занимает крупное ядро. Клетка содержит
митохондрии, выраженный комплекс Гольджи, элементы гранулярной
эндоплазматической сети. [17]
по ходу крупных артериальных стволов (например, каротидное тело). Мелкие скопления и одиночные хромаффинные клетки находят также в сердце, почках, симпатических ганглиях.
Хромаффинные клетки содержат многочисленные митохондрии, выраженный комплекс Гольджи, элементы гранулярной эндоплазматической сети, многочисленные электронно-плотные гранулы, содержащие преимущественно норадреналин и/или адреналин (по этому признаку хромаффинные клетки подразделяют на две субпопуляции), а также АТФ, энкефалины и хромогранины. Адреналин-содержащие гранулы гомогенны. Норадреналин-содержащие гранулы характеризуются повышенной плотностью содержимого в центральной части и наличием светлого ободка по периферии под мембраной гранулы. Секреция гормонов из хромаффинных клеток происходит в результате стимулирующего влияния со стороны преганглионарных симпатических волокон и глюкокортикоидов. Секрет хромаффинных клеток содержит 10% норадреналина и 90% адреналина. Эти катехоламины имеют широкий спектр эффектов (воздействие на гликогенолиз, ли-
полиз, глюконеогенез, существенно влияние на сердечно-сосудистую систему). Вазоконстрикция, параметры сокращения сердечной мышцы и другие эффекты катехоловых аминов реализуются через α- и β-адренергические рецепторы на поверхности клеток-мишеней (ГМК, секреторные клетки, кардиомиоциты).