Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 8 НЕЙРОГИСТОЛОГИЯ
Нервная система человека содержит не менее триллиона (1012) нервных, около 1013 глиальных клеток и не меньшее количество (>1013) синапсов. Число клеточных типов неизвестно (не менее 100). Это множество образует сложную пространственную структуру - единую сеть с многочисленными связями как на уровне отдельной клетки, так и клеточных ансамблей (ЦНС).
НЕРВНАЯ ТКАНЬ
Гистологические элементы нервной ткани (нейроны и глиоциты) и органов чувств развиваются из производных нейроэктодермы, а именно из нервной трубки, нервного гребня и нейрогенных плакод. Нейруляция, в ходе которой образуется нейроэктодерма, рассмотрена в главе 3 (см. рис. 3-13).
Нервная трубка содержит стволовые нейральные клетки (они же матричные, или вентрикулярные) - источник почти всех клеток ЦНС. Вентрикулярные клетки размножаются и дают начало нейробластам и глиобластам, это клетки-предшественницы нейронов и глиоцитов. Часть вентрикулярных клеток остаётся in situ, это будущая эпендима. Нервный гребень. Если нервная трубка служит источником для развития ЦНС, то нервные элементы периферии и ряд других важных структур (см. табл. 3-3) происходят из нервного гребня. Нейрогенные плакоды. Из них образуются нейроны обонятельной выстилки, вестибулярного и слухового ганглиев, а также чувствительные нейроны коленчатого, каменистого, узловатого и тройничного ганглиев черепных нервов.
• Нейробласты - клетки с большим округлым ядром, плотным ядрышком и бледной цитоплазмой - дают начало нейронам. Нейроны относятся к статической популяции. Ни при каких условиях они in vivo не способны к пролиферации и обновлению. Исключение составляют обонятельные нейроны эпителиальной выстилки носо-
вых ходов, а также некоторые нейроны гиппокампа и обонятельной луковицы.
• Глиобласты - предшественники макроглии (астроциты и олигодендроциты). Все типы макроглии способны к пролиферации.
• Микроглия. Клетки микроглии относят к системе мононуклеарных фагоцитов.
Нейроны
Нейроны - главные клеточные типы нервной ткани. Эти возбудимые клетки проводят электрические сигналы и обеспечивают способность мозга к переработке информации. Перикарион (тело нейрона), отходящие от него отростки (аксон и дендриты) - стандартные части нейронов (рис. 8-13).
• Перикарион содержит ядро, комплекс Гольджи, гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета.
• Ядро нейрона имеет мелкодисперсный хроматин и ядрышко. В силу относительно большого диаметра ядро при светооптической микроскопии выглядит как оптически пустое. Ядрышко крупное и резко базофильное.
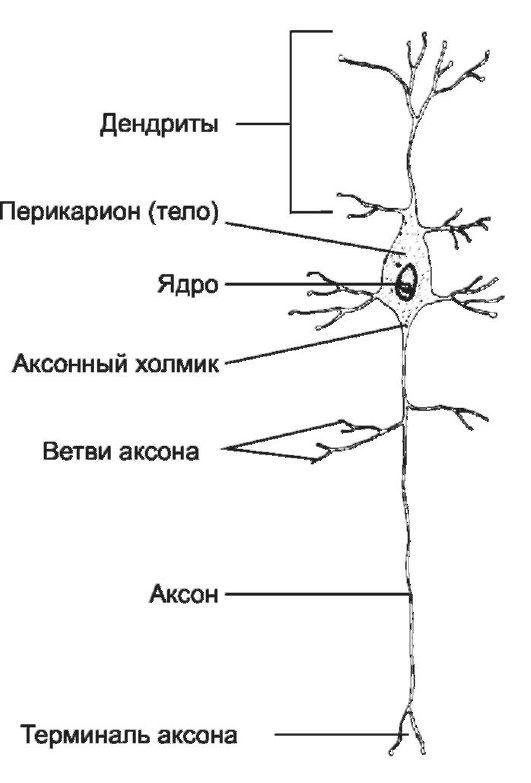 Рис. 8-13. Мультиполярный нейрон. Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них - аксон, все другие - дендриты. [17]
Рис. 8-13. Мультиполярный нейрон. Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них - аксон, все другие - дендриты. [17]
• Комплекс Гольджи хорошо развит. Его особенность - расположение между ядром и местом отхождения аксона, что отражает мощный транспорт белков, синтезированных в гранулярной эндоплазматической сети перикариона, в аксон.
• Аксонный холмик - свободная от гранулярной эндоплазматической сети и рибосом область перикариона, содержащая много микротрубочек и нейрофиламентов, это место, где начинается аксон и генерируется потенциал действия.
• Гранулярная эндоплазматическая сеть. В перикарионе и дендритах развита гранулярная эндоплазматическая сеть. Она соответствует глыбкам хроматофильного вещества (вещество Ниссля).
• Митохондрии многочисленны. Значительные энергетические потребности нервных клеток обеспечивает преимущественно аэробный метаболизм, почему нейроны крайне чувствительны к гипоксии.
• Пигменты. В нейронах (особенно с возрастом) накапливается липофусцин. Нейроны некоторых ядер мозга нормально содержат иные пигменты, почему эти образования и получили своё название (substantia nigra, locus coeruleus).
Отростки, отходящие от перикариона, - аксон и дендриты (рис. 8-13).
• Аксон (нейрит) - длинный отросток, как правило, не ветвящийся на всём его протяжении, но образующий концевые разветвления, содержащие синаптические пузырьки. Объём аксона может достигать 99% суммарного объёма нейрона. Длина аксона может быть весьма значительной, до десятков сантиметров.
• Дендриты - ветвящиеся отростки, заканчивающиеся вблизи от тела нейрона. В плазмолемму встроены постсинаптические рецепторы, дендриты проводят возбуждение к перикариону. Проксимальные области дендритов - продолжение перикариона. Поэтому они содержат рибосомы, компоненты гранулярной и гладкой эндоплазматической сети, элементы комплекса Гольджи. Дендриты интегрально образуют до 95% всей рецепторной поверхности (рецептивного поля) нейрона.
Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофиламенты) и микрофиламентов.
Микротрубочки - наиболее крупные элементы цитоскелета, их диаметр 24 нм. С ними связывают внутриклеточный, в том числе аксонный транспорт. От перикариона по отросткам перемещаются различные вещества (белки, нейромедиаторы и т.д.), органеллы (митохондрии, элементы цитоскелета, везикулы и т.д.). Микротрубочки в перикарионе и дендритах (в отличие от аксона) не имеют направленной ориентации. Большинство микротрубочек аксона (+)-концом направлено к терминали, а (-)-концом - к перикариону (рис. 8-14). Характер ориентации микротрубочек имеет важное значение для распределения по отросткам различных органелл. К (+)-концу перемещаются митохондрии и секреторные пузырьки, а к (-)-концу - рибосомы, мультивезикулярные тельца, элементы комплекса Гольджи.
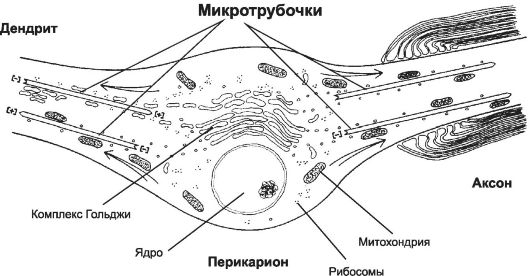 Рис. 8-14. Ориентация микротрубочек в отростках нейрона. Аксон
отличается от дендритов полярной ориентацией микротрубочек. В нём
микротрубочки своими (+)- концами направлены к терминали, а (-)-концами -
к перикариону. [17]
Рис. 8-14. Ориентация микротрубочек в отростках нейрона. Аксон
отличается от дендритов полярной ориентацией микротрубочек. В нём
микротрубочки своими (+)- концами направлены к терминали, а (-)-концами -
к перикариону. [17]
Классификации нейронов. Нейроны отличаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в функциональных сетях и множеству других характеристик. По этой причине классификации нейронов многочисленны. Приводим некоторые классифицирующие критерии и соответствующие примеры.
• Количество отростков. По этому критерию различают нейроны аполярные (отростков нет, ранние нейробласты), униполярные (единственный отросток, формально одноотростчатыми можно считать псевдоуниполярные нейроны спинномозговых узлов), биполярные (два отростка - аксон и дендрит, обонятельные рецепторные клетки), мультиполярные (число отростков >2, один аксон, остальные - дендриты, мотонейроны передних рогов спинного мозга).
• Химия нейромедиатора. Критерий классификации - синтез, накопление в синаптических пузырьках и экскреция в синаптическую щель конкретного нейромедиатора. При этом к имени нейромедиатора добавляют «ергический». Иногда в качестве критерия применяют тип мембранного рецептора, регистрирующего наличие нейромедиатора (в этом случае добавляют «цептивный»). Примеры: холинергические нейроны, адренергические нервные клетки.
• Позиция нейрона в рефлекторной дуге позволяет выделять чувствительные (воспринимающие сигнал из внешней или внутренней среды) нейроны, двигательные (иннервирующие сократительные и секреторные элементы) нейроны и находящиеся между ними вставочные (ассоциативные в нейронных сетях) нервные клетки (интернейроны).
• Направление проведения возбуждения: к центру - афферентные нервные клетки, к периферии - эфферентные нейроны.
• Модальность - характер воспринимаемого и передаваемого сигнала (например, механорецепторные, зрительные, обонятельные нейроны и т.д.).
Синапсы
Синапсы - специализированные межклеточные контакты, передающие сигналы от одного нейрона к другому при помощи нейромедиаторов. В синапсе выделяют пресинаптическую и постсинаптическую части, разделённые синаптической щелью шириной 20-30 нм. Пресинаптические нейроны синтезируют, хранят и секретируют нейромедиаторы. При изменении мембранного потенциала нейромедиатор выделяется в синаптическую щель и связывается со своими рецепторами в постсинаптической мембране, вызывая изменение мембранного потенциала постсинаптического нейрона.
Классификация синапсов. Различают синапсы аксодендритические, аксосоматические, аксо-аксональные и дендродендритические.
• Аксодендритические - синапсы между аксоном одного нейрона и дендритами другого нейрона (рис. 8-15). Вариантом являются аксо-шипиковые синапсы.
• Аксо-шипиковый синапс. Большинство возбуждающих синапсов локализуется в выростах дендритов, содержащих большое количество актина и называемых шипиками.
• Аксо-аксональные - синапсы между аксонами разных нейронов.
• Аксосоматические - синапсы между терминалями аксона одного нейрона и телом другого нейрона.
• Дендродендритические - синапсы между дендритами нейронов. Нейромедиаторы. Большинство нейромедиаторов - аминокислоты и их производные. Одни нейроны модифицируют аминокислоты с образованием аминов (норадреналин, серотонин, ацетилхолин), другие секретируют нейромедиаторы пептидной природы (эндорфины, энкефалины, разные нейропептиды). Часть нейронов используют в качестве нейромедиатора ацетилхолин. Некоторые нейроны могут синтезировать более одного нейромедиатора.
Нейроглия
Клетки нейроглии составляют почти половину объёма мозга. Среди глиальных клеток мозга выделяют эпендимную глию, макроглию и микроглию (рис. 8-16). Макроглия состоит из астроцитов и олигодендроцитов. В периферической нервной системе присутствуют шванновские клетки и группа окружающих нейроны вспомогательных клеток в ганглиях.
Астроциты. Отростки этих звездчатой формы клеток отходят от тела клетки в разных направлениях, оплетают нейроны, сосуды, клетки (эпендимы) желудочков мозга, образуя расширения в виде концевой ножки. Маркёр астроцитов - глиальный фибриллярный кислый бе-
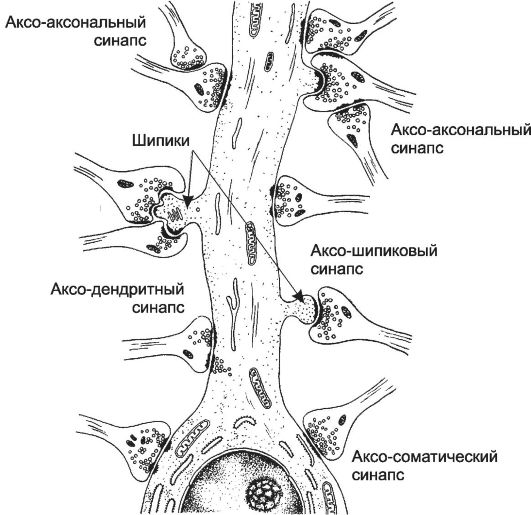 Рис. 8-15. Межнейронные синапсы. [17]
Рис. 8-15. Межнейронные синапсы. [17]
лок промежуточных филаментов. Различают волокнистые и протоплазматические астроциты.
• Волокнистые астроциты имеют длинные, слабо или совсем не ветвящиеся отростки; присутствуют в белом веществе мозга. Концевые расширения их отростков окружают области перехватов в миелиновых волокнах.
• Протоплазматические астроциты находятся в сером веществе мозга. Клетки имеют многочисленные короткие и ветвящиеся отростки. Концевые расширения отростков окружают области синапсов.
Миелинобразующие клетки. Миелин - компактная структура из мембран, спирально закрученных вокруг аксонов. 70% массы миелина составляют липиды. Именно миелин придаёт белому веществу характерный цвет, отличающий его от серого вещества. Миелинобразующие клетки - шванновские и олигодендроциты.
• Олигодендроциты (рис. 8-16Г), как правило, более мелкие клетки, чем астроциты. В сером веществе мозга олигодендроциты находятся
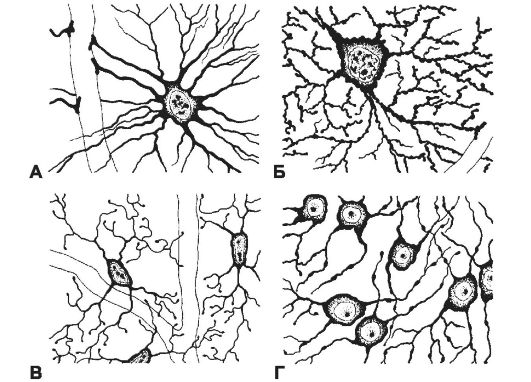 Рис. 8-16. Глиальные клетки: А - волокнистый астроцит; Б - протоплазматический астроцит; В - микроглия; Г - олигодендроглиоциты. [17]
Рис. 8-16. Глиальные клетки: А - волокнистый астроцит; Б - протоплазматический астроцит; В - микроглия; Г - олигодендроглиоциты. [17]
в непосредственном контакте с перикарионами и отростками нейронов. В белом веществе олигодендроциты расположены рядами между нервными волокнами. При помощи тонких неветвящихся отростков олигодендроциты контактируют с аксонами и, продвигаясь относительно аксона уплощёнными концами отростков, окружают его циркулярной пластиной миелина (хорошая аналогия - вращаясь вокруг аксонов, наматывают миелин на аксон). Каждый олигодендроцит при помощи своих отростков миелинизирует несколько аксонов.
• Шванновские клетки входят в состав периферических нервных волокон и образуют миелин. Миелинизация аксонов в периферической нервной системе рассмотрена на рисунке 8-17. Каждая шванновская клетка миелинизирует один аксон. Эпендимная глия. Эпендимные клетки имеют кубическую форму и образуют эпителиоподобный пласт, выстилающий центральный канал и желудочки мозга. Клетки имеют хорошо развитые реснички, а также формируют промежуточные, плотные и щелевые контакты, тем са-
мым образуя барьер проницаемости. Модифицированные эпендимоциты секретируют цереброспинальную жидкость. Микроглия. Клетки микроглии (рис. 8-16В) имеют небольшие размеры, неправильную форму, многочисленные ветвящиеся отростки, ядро с крупными глыбками хроматина, множество лизосом, гранулы липофусцина и плотные пластинчатые тельца. Функция микроглии в интактном мозге неясна.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
НЕРВНЫЕ ВОЛОКНА
В зависимости от того, формируют ли шванновские клетки вокруг осевого цилиндра миелин, выделяют безмиелиновые и миелиновые волокна.
Безмиелиновые волокна состоят из аксонов нервных клеток (осевых цилиндров), окружённых шванновскими клетками. При погружении осевого цилиндра в шванновскую клетку её клеточная мембрана смыкается и образует мезаксон (рис. 8-17) - сдвоенные мембраны шванновской клетки. Каждая шванновская клетка подобным образом окружает несколько осевых цилиндров.
Миелиновое волокно состоит из осевого цилиндра, вокруг которого шванновские клетки образуют миелин за счёт удлинения и концентрического наслаивания мембран мезаксона. Каждая шванновская клетка миелинизирует небольшой сегмент только одного аксона. Миелин прерывается через регулярные промежутки, это узловые перехваты. Фактически это границы между двумя соседними шванновскими клетками. В миелине периферических нервов присутствуют небольшие просветления - насечки миелина. Снаружи от миелина располагаются тонкий слой цитоплазмы шванновской клетки и её ядро.
• Осевой цилиндр содержит митохондрии, элементы гладкой эндоплазматической сети, пузырьки, а также элементы цитоскелета - микротрубочки, нейрофиламенты и микрофиламенты. Диаметр аксона, а следовательно и скорость проведения импульсов по этому аксону, определяются количеством в нём нейрофиламентов.
• Аксонный транспорт различных компонентов обеспечивает тубулин микротрубочек. Различают быстрый (100-1000 мм/сутки) и медленный аксонный транспорт (1-10 мм/сутки), а также антероградный (транспорт от перикариона) и ретроградный (к перикариону). Основной материал антероградного транспорта - белки, синтезированные в перикарионе (например, белки ионных каналов, ферменты синтеза нейромедиаторов).
• Узловые перехваты. На границе между соседними шванновскими клетками участок плазматической мембраны аксона (аксолемма) не прикрыт миелином. Здесь шванновские клетки образуют многочисленные переплетающиеся отростки. Ак-
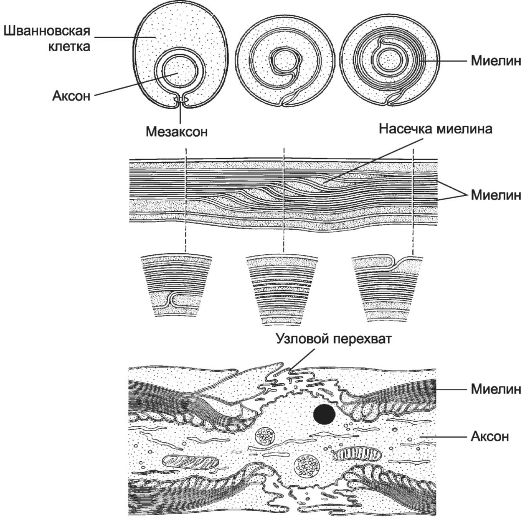 Рис. 8-17. Образование миелинового волокна. В
верхней части рисунка показаны ранние стадии образования миелина. По
мере удлинения мезаксона происходит спиральное наслаивание мембраны
шванновской клетки. При этом её цитоплазма смещается на периферию.
Насечка миелина - узкая полоса, в пределах которой мембраны миелина
расходятся, и между ними расположены небольшие островки цитоплазмы
шванновской клетки. В нижней части рисунка дана схема продольного среза
миелинового волокна в области узлового перехвата - границы между
соседними шванновскими клетками, где они соединяются при помощи
переплетающихся коротких отростков. В перехвате аксолемма осевого
цилиндра не покрыта миелиновой оболочкой. [17]
Рис. 8-17. Образование миелинового волокна. В
верхней части рисунка показаны ранние стадии образования миелина. По
мере удлинения мезаксона происходит спиральное наслаивание мембраны
шванновской клетки. При этом её цитоплазма смещается на периферию.
Насечка миелина - узкая полоса, в пределах которой мембраны миелина
расходятся, и между ними расположены небольшие островки цитоплазмы
шванновской клетки. В нижней части рисунка дана схема продольного среза
миелинового волокна в области узлового перехвата - границы между
соседними шванновскими клетками, где они соединяются при помощи
переплетающихся коротких отростков. В перехвате аксолемма осевого
цилиндра не покрыта миелиновой оболочкой. [17]
солемма перехватов содержит множество потенциалозависимых ?+-каналов, необходимых для поддержания импульсной активности.
• Насечки миелина - участки расслоения миелина, образовавшиеся при миелинизации; в них присутствует цитоплазма шванновских клеток. Функция насечек неясна.
Периферические нервы состоят из миелиновых и безмиелиновых волокон, сгруппированных в пучки, и соединительнотканных оболочек.
• Оболочки нерва. К оболочкам нерва относятся эндоневрий, периневрий и эпиневрий.
♦ Эндоневрий - рыхлая соединительная ткань между отдельными нервными волокнами.
♦ Периневрий содержит наружную часть - плотную соединительную ткань, окружающую каждый пучок нервных волокон, и внутреннюю часть - несколько концентрических слоёв плоских периневральных клеток, снаружи и изнутри покрытых исключительно толстой базальной мембраной, содержащей коллаген типа IV, ламинин и фибронектин.
Периневральный барьер необходим для поддержания гомеостаза в эндоневрии, его образует внутренняя часть периневрия - эпителиоподобный пласт периневральных клеток, соединённых при помощи плотных контактов. Барьер контролирует транспорт молекул через периневрий к нервным волокнам, предотвращает доступ в эндоневрий инфекционных агентов.
♦ Эпиневрий - волокнистая соединительная ткань, объединяющая все пучки в составе нерва.
• Кровоснабжение. Периферический нерв содержит разветвлённую сеть кровеносных сосудов. В эпиневрии и в наружной (соединительнотканной) части периневрия - артериолы и венулы, а также лимфатические сосуды. Эндоневрий содержит кровеносные капилляры.
• Иннервация. Периферический нерв имеет специальные нервные волокна - nervi nervorum - тонкие чувствительные и симпатические нервные волокна. Их источник: сам нерв или сосудистые нервные сплетения.
• Дегенерация и регенерация нерва. При повреждении нерва центральный отрезок (связанный с перикарионами) и периферический отрезок (дистальнее места повреждения) претерпевают разные изменения. Дегенерация нервных волокон (рис. 8-18) происходит на небольшом протяжении центрального и на всём протяжении периферического отрезка. Позднее происходит регенерация периферического отрезка. В этом процессе ведущую роль играют шванновские клетки. Шванновские клетки стимулируют удлинение аксона и контролируют его направленный рост к мишени. При отсутствии шванновских клеток аксоны не могут расти на значительные расстояния.
ЧУВСТВИТЕЛЬНЫЕ НЕРВНЫЕ УЗЛЫ
Спинномозговой узел (ganglion spinale) покрыт соединительнотканной капсулой. Внутри узла находятся группы псевдоуниполярных чувствительных нейронов, между которыми проходят пучки миелиновых
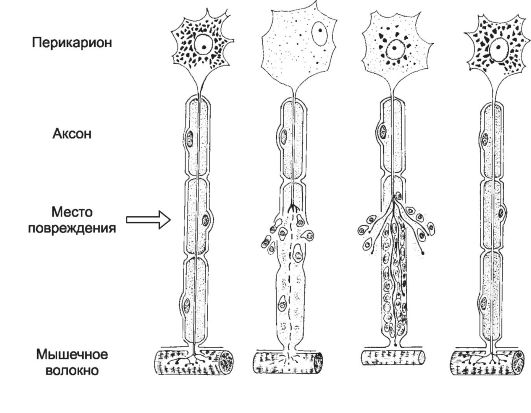
Рис. 8-18. Регенерация нервного волокна. А - волокно до повреждения; Б - в периферическом отрезке аксон дегенерирует, клетки в месте повреждения пролиферируют; В - регенерация аксона в центральном отрезке, прорастание веточек аксона в периферический отрезок; Г - полная регенерация нервного волокна и восстановление связей. [17]
волокон. Перикарионы нейронов имеют округлую форму и окружены клетками-сателлитами. Популяция нейронов ганглия неоднородна. Спинномозговые узлы содержат более 20 различных подтипов чувствительных нейронов.
НЕРВНЫЕ ОКОНЧАНИЯ
Окончания аксонов периферических нервов подразделяют на чувствительные (афферентные) и двигательные (эфферентные).
Чувствительные нервные окончания
Внешние раздражители, а также сигналы о состоянии внутренней среды и мышц организма регистрируют чувствительные нервные окончания - сенсорные рецепторы. Различают свободные и несвободные
воспринимающие приборы, а среди последних инкапсулированные тельца.
• Свободные нервные окончания - терминальные ветвления периферического отростка чувствительного нейрона.
• Несвободные нервные окончания, помимо терминальных ветвлений, содержат специальные клетки. Практически все окончания этого типа - механорецепторы.
• Инкапсулированные механорецепторы (инкапсулированные тельца) - несвободные окончания, имеющие оформленную соединительнотканную капсулу.
Свободные нервные окончания (рис. 8-21) расположены в прослойках соединительной ткани внутренних органов и кожи, а также в базальном и шиповатом слоях эпидермиса, а в областях кожи с высокой тактильной чувствительностью достигают зернистого слоя. Большинство свободных нервных окончаний - механорецепторы. Некоторые окончания в эпидермисе определённо специализированы для регистрации изменений температуры. Имеются также рецепторы, способные определять изменения рН, рО2 и рСО2.
Комплекс тактильного эпителиоцита с нервной терминалью. Тактильные эпителиоциты - округлые или удлинённые клетки, расположенные преимущественно в эпителии кожи пальцев, губ и наружных половых органов (рис. 8-22). Эти клетки формируют контакты с нервными терминалями. В области контакта терминаль резко расширена, содержит множество митохондрий, нейрофиламентов и микротрубочек. Тактильные эпителиоциты крупнее соседних кератиноцитов, в цитоплазме в умеренном количестве содержатся митохондрии, лизосомы, мультивезикулярные тельца, вакуоли, микрофиламенты. Для тактильных эпителиоцитов характерны специфические осмиофильные гранулы размером от 80 до 200 нм. Они сосредоточены преимущественно в обращённых к нервной терминали участках цитоплазмы. В тактильных эпителиоцитах обнаружены пептиды и нейроноспецифические вещества (например, метионин-энкефалин, VIP, вещество Р), что свидетельствует об эндокринной функции клеток и позволяет рассматривать их как компонент диффузной нейроэндокринной системы. Тактильные эпителиоциты участвуют в распознавании формы объекта, его краёв, текстуры поверхности.
Пластинчатые тельца (рис. 8-24) обнаружены в соединительной ткани кожи и различных органов, имеют овальную форму, размер до 0,5- 1,0 мм. Внутренняя колба, наружная капсула и терминальное нервное волокно - основные компоненты тельца.
Адекватная механическая стимуляция пластинчатых телец приводит к появлению рецепторного потенциала в терминальной части нервного окончания. Достигнув критической величины, рецепторный потенциал в первом узловом перехвате вызывает появление потенциалов действия. Наружная капсула рецептора - фильтр, пропускающий только динамическую составляющую механического воздействия.
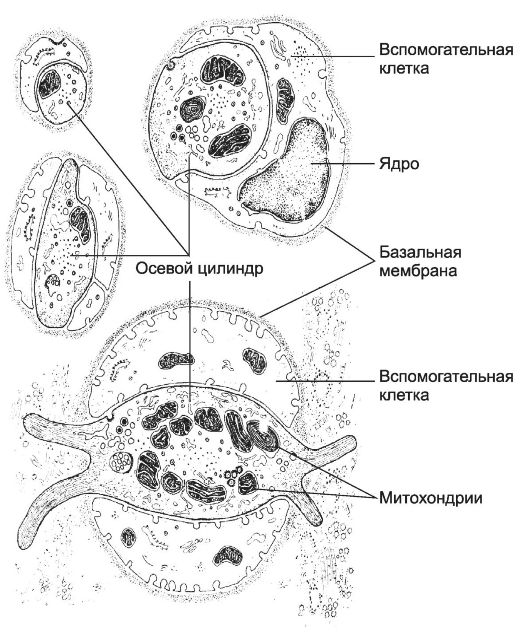 Рис. 8-21. Свободные нервные окончания в соединительной ткани. Осевой
цилиндр контактирует с вспомогательными клетками на значительном
протяжении или полностью окружён ими. Свободное нервное окончание,
изображённое в нижней части рисунка, имеет билатеральную организацию: в
центре расположена эллиптической формы нервная терминаль, покрытая
вспомогательными клетками. [17]
Рис. 8-21. Свободные нервные окончания в соединительной ткани. Осевой
цилиндр контактирует с вспомогательными клетками на значительном
протяжении или полностью окружён ими. Свободное нервное окончание,
изображённое в нижней части рисунка, имеет билатеральную организацию: в
центре расположена эллиптической формы нервная терминаль, покрытая
вспомогательными клетками. [17]
Тактильные тельца (рис. 8-26) присутствуют в сосочковом слое кожи, наиболее компактно располагаясь в кончиках пальцев. Тельце имеет удлинённую форму. Сердцевина тельца образована пластинчатыми вспомогательными клетками, чередующимися с расширенными нервными терминалями.
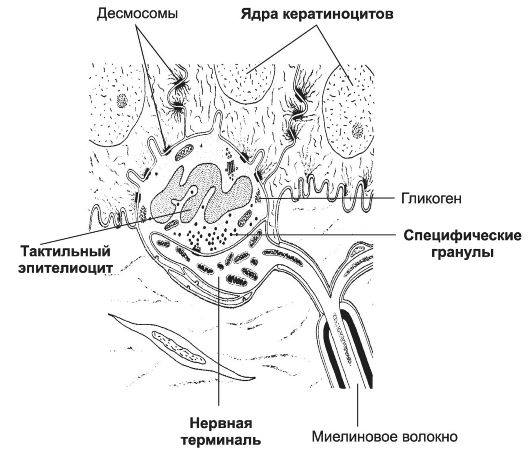 Рис. 8-22. Комплекс тактильного эпителиоцита с нервной терминалью. Тактильный
эпителиоцит расположен в базальном слое эпидермиса, образует отростки и
связывается с кератиноцитами при помощи десмосом. Расширенная нервная
терминаль вступает в контакт с тактильным эпителиоцитом. Специфические
гранулы расположены в обращённой к нервной терминали части клетки. [17]
Рис. 8-22. Комплекс тактильного эпителиоцита с нервной терминалью. Тактильный
эпителиоцит расположен в базальном слое эпидермиса, образует отростки и
связывается с кератиноцитами при помощи десмосом. Расширенная нервная
терминаль вступает в контакт с тактильным эпителиоцитом. Специфические
гранулы расположены в обращённой к нервной терминали части клетки. [17]
Другие тельца (например, веретеновидные и генитальные тельца, колбы Краузе) встречаются реже.
Рецепторы мышц и суставов. К ним относятся мышечные веретёна, сухожильные органы и чувствительные нервные окончания в капсуле суставов.
• Мышечные веретёна (рис. 8-27, см. также рис. 7-9) - чувствительные воспринимающие приборы скелетной мышцы. Основные структурные элементы мышечного веретена - интрафузальные мышечные волокна, нервные волокна и капсула. Мышечное веретено содержит от 1 до 10 коротких интрафузальных мышечных волокон. В экваториальной их части ядра образуют компактное скопление (волокна с
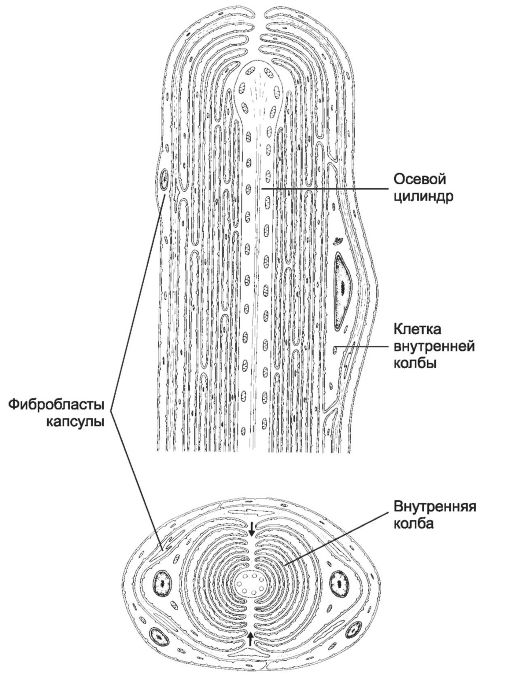 Рис. 8-24. Пластинчатое тельце состоит
из билатерально организованной внутренней колбы и наружной
соединительнотканной капсулы. Клетки внутренней колбы образуют отростки,
концентрическими полукольцами окружающие чувствительную нервную
терминаль. Стрелками указана щель внутренней колбы, проходящая
параллельно короткой оси эллипса нервной терминали. [17]
Рис. 8-24. Пластинчатое тельце состоит
из билатерально организованной внутренней колбы и наружной
соединительнотканной капсулы. Клетки внутренней колбы образуют отростки,
концентрическими полукольцами окружающие чувствительную нервную
терминаль. Стрелками указана щель внутренней колбы, проходящая
параллельно короткой оси эллипса нервной терминали. [17]
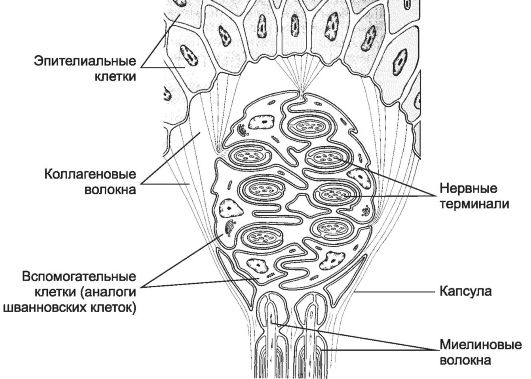 Рис. 8-26. Тактильное тельце окружено
соединительнотканной капсулой с вплетён- ными в неё коллагеновыми
волокнами окружающей соединительной ткани. Подходящие к тельцу нервные
волокна теряют миелин и заходят внутрь тельца, где формируют
многочисленные терминали, окружённые вспомогательными клетками. [17]
Рис. 8-26. Тактильное тельце окружено
соединительнотканной капсулой с вплетён- ными в неё коллагеновыми
волокнами окружающей соединительной ткани. Подходящие к тельцу нервные
волокна теряют миелин и заходят внутрь тельца, где формируют
многочисленные терминали, окружённые вспомогательными клетками. [17]
ядерной сумкой) или располагаются цепочкой (волокна с ядерной цепочкой). Терминали Ια-волокон образуют спираль в пределах экваториальной зоны обоих типов интрафузальных мышечных волокон (первичные, или аннулоспиральные окончания). Терминали более тонких II-волокон заканчиваются на интрафузальных волокнах в области, расположенной рядом с экваториальной.
• Сухожильные органы расположены в концевой части сухожилия на границе с мышцей, а также в связках капсулы суставов. Рецептор имеет веретеновидную форму и окружён капсулой, состоящей из нескольких слоёв плоских клеток. Капсула рецептора - продолжение периневрия и содержит капилляры. В образовании сухожильного органа участвуют терминали афферентных миелиновых волокон (Ib), они ветвятся среди пучков спиралевидных коллагеновых волокон, расположенных в заполненном жидкостью пространстве.
• Чувствительные нервные окончания капсулы суставов - важный элемент проприоцептивной системы организма. Веретеновидные тельца
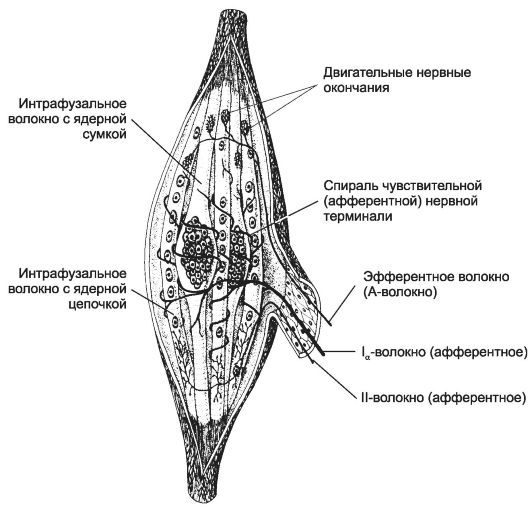 Рис. 8-27. Мышечное веретено. Интрафузальные
мышечные волокна с компактным скоплением ядер - волокна с ядерной
сумкой, в интрафузальных волокнах с ядерной цепочкой ядра распределены
по длине волокна более равномерно. Первичные (аннулоспиральные)
окончания расположены в экваториальной области. Ближе к концам
интрафузальных волокон расположены терминали тонких афферентных
II-волокон - вторичные окончания. Эфферентные волокна образуют
нервно-мышечные синапсы с интрафузальными волокнами в концевой их части.
[17]
Рис. 8-27. Мышечное веретено. Интрафузальные
мышечные волокна с компактным скоплением ядер - волокна с ядерной
сумкой, в интрафузальных волокнах с ядерной цепочкой ядра распределены
по длине волокна более равномерно. Первичные (аннулоспиральные)
окончания расположены в экваториальной области. Ближе к концам
интрафузальных волокон расположены терминали тонких афферентных
II-волокон - вторичные окончания. Эфферентные волокна образуют
нервно-мышечные синапсы с интрафузальными волокнами в концевой их части.
[17]
расположены в периферических участках капсулы. Мелкие пластинчатые тельца, а также свободные нервные окончания - терминали тонких миелиновых волокон и безмиелиновых волокон, среди которых, по-видимому, присутствуют и болевые рецепторы, широко представлены во всех компонентах сустава, но наибольшей плотности достигают в мениске и суставном диске.
Двигательные нервные окончания
Характеристики двигательных нервных окончаний рассмотрены ниже на примере нервно-мышечных синапсов. Как и в других синапсах, здесь различают пресинаптическую и постсинаптическую области, разделённые синаптической щелью.
Пресинаптическая область. Двигательная нервная терминаль снаружи покрыта шванновской клеткой, имеет диаметр 1-1,5 мкм и образует пресинаптическую область нервно-мышечного синапса (рис. 8-29).
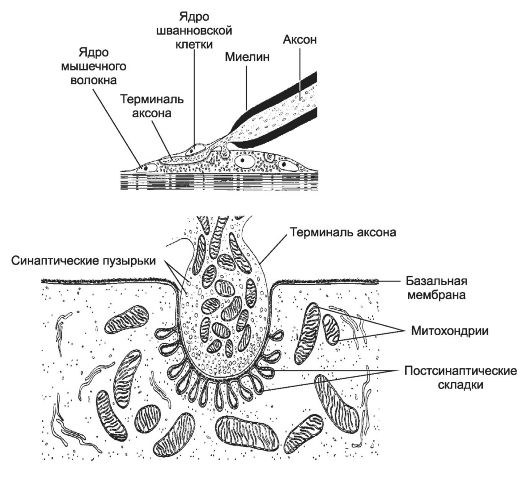 Рис. 8-29. Нервно-мышечный синапс. Пресинаптическая
часть образована терминалью аксона мотонейрона и содержит скопление
синаптических пузырьков вблизи пресинаптической мембраны, а также
митохондрии. Постсинаптические складки увеличивают площадь поверхности
постсинаптической мембраны. В синаптической щели находится синаптическая
базальная мембрана (продолжение базальной мембраны мышечного волокна),
она заходит в постсинаптические складки. В синаптической щели также
находятся молекулы ацетилхолинэстеразы. Этот фермент расщепляет
ацетилхолин и устраняет эффект деполяризующего сигнала на мышечное
волокно. [17]
Рис. 8-29. Нервно-мышечный синапс. Пресинаптическая
часть образована терминалью аксона мотонейрона и содержит скопление
синаптических пузырьков вблизи пресинаптической мембраны, а также
митохондрии. Постсинаптические складки увеличивают площадь поверхности
постсинаптической мембраны. В синаптической щели находится синаптическая
базальная мембрана (продолжение базальной мембраны мышечного волокна),
она заходит в постсинаптические складки. В синаптической щели также
находятся молекулы ацетилхолинэстеразы. Этот фермент расщепляет
ацетилхолин и устраняет эффект деполяризующего сигнала на мышечное
волокно. [17]
В пресинаптической области в большом количестве присутствуют синаптические пузырьки и митохондрии.
• Пресинаптическая мембрана - специализированная часть аксолеммы нервной терминали. В пресинаптической мембране выявлены так называемые активные зоны - участки утолщения мембраны, имеющие прямое отношение к секреции медиатора. Рядом с активными зонами в пресинаптической мембране находятся углубления, количество которых коррелирует с уровнем секреции ацетилхолина. Пресинаптическая мембрана содержит потенциалозависимые Са2+-каналы. При деполяризации мембраны каналы открываются, и ионы Са2+ входят в терминаль, запуская секрецию ацетилхолина.
• Синаптические пузырьки присутствуют практически в любой области нервной терминали, но в непосредственной близости от пресинаптической мембраны они образуют выраженное скопление. Размеры пузырьков варьируют, их средний диаметр равен 50 нм. В холинергических синапсах светлые синаптические пузырьки содержат ацетилхолин.
Постсинаптическая область. Для постсинаптической области характерно наличие крупных митохондрий с хорошо развитыми кристами и большого количества рибосом.
Постсинаптическая мембрана - специализированная часть плазмолеммы мышечного волокна - образует многочисленные инвагинации, от которых на глубину 0,5-1,0 мкм отходят постсинаптические складки, чем существенно увеличивается площадь мембраны. В постсинаптическую мембрану встроены н-холинорецепторы, их концентрация достигает 20-30 тысяч на 1 мкм2.
Синаптическая щель. Через синаптическую щель проходит синаптическая базальная мембрана, она удерживает в области синапса терминаль аксона и контролирует расположение холинорецепторов в постсинаптической мембране.
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Нейроны парасимпатического отдела происходят из нервного гребня на уровне 1-7 сомитов (отдел блуждающего нерва) и каудальнее 28 сомита (пояснично-крестцовый отдел). Нейроны симпатического отдела и хромаффинные клетки мозгового вещества надпочечников развиваются из нервного гребня на уровне сомитов 8-28. Вегетативная иннервация (симпатическая и парасимпатическая) каждого внутреннего органа рассмотрена в соответствующих главах учебника. Здесь мы представляем общую схему анатомических связей симпатического отдела (рис. 8-32).
Строение периферических отделов вегетативной нервной системы рассмотрим на примере вегетативной иннервации пищеварительного тракта (см. также раздел «нервный аппарат пищеварительного тракта» главы 12, рис. 12-19 и рис. 19-20). В стенке пищеварительной трубки имеются нервные сплетения, содержащие ганглии. В последних сосредоточены перикарионы вегетативных нейронов. Количество нейронов в ганглии варьирует от нескольких до сотен. Совокупность нервных элементов пищеварительного тракта составляет энтеральную нервную систему.
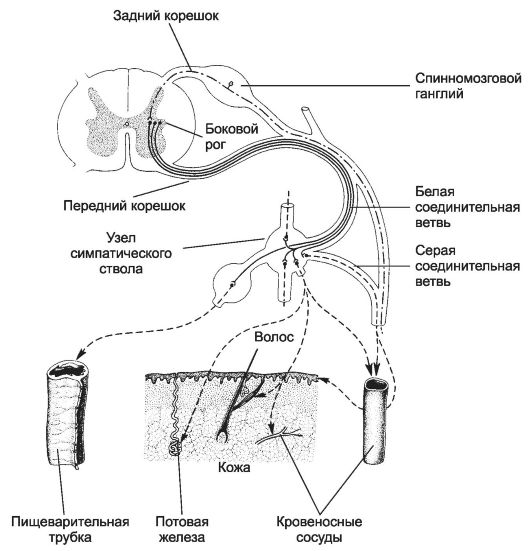 Рис. 8-32. Симпатический отдел вегетативной нервной системы. Перикарионы
первого нейрона расположены в боковых столбах спинного мозга. Их аксоны
проходят в составе передних корешков и белой соединительной ветви и
заканчиваются на перикарионах второго нейрона в паравертебральных
ганглиях симпатической цепочки, превертебральных ганглиях и ганглиях
(терминальных), расположенных вблизи иннервируемых ими органов. Аксоны
второго нейрона паравертебральных, превертебральных и терминальных
ганглиев заканчиваются во внутренних органах, коже, стенке кровеносных
сосудов. Аксоны некоторых нейронов паравертебральных ганглиев проходят
через серую соединительную ветвь. [17]
Рис. 8-32. Симпатический отдел вегетативной нервной системы. Перикарионы
первого нейрона расположены в боковых столбах спинного мозга. Их аксоны
проходят в составе передних корешков и белой соединительной ветви и
заканчиваются на перикарионах второго нейрона в паравертебральных
ганглиях симпатической цепочки, превертебральных ганглиях и ганглиях
(терминальных), расположенных вблизи иннервируемых ими органов. Аксоны
второго нейрона паравертебральных, превертебральных и терминальных
ганглиев заканчиваются во внутренних органах, коже, стенке кровеносных
сосудов. Аксоны некоторых нейронов паравертебральных ганглиев проходят
через серую соединительную ветвь. [17]
Вегетативные нейроны. В энтеральной нервной системе выделено три типа нейронов.
• Энтеральные нейроны типа I: перикарионы имеют уплощённую форму, длинный аксон и большое количество коротких дендритов с расширенным основанием.
• Энтеральные нейроны типа II имеют перикарион овальной формы с гладкой поверхностью и длинные отростки.
• Энтеральные нейроны типа III: перикарионы имеют овальную или неправильную форму, один длинный аксон и большое количество сравнительно коротких дендритов различной длины.
Связи нейронов. Нейроны типа I образуют синапсы с ГМК и с нейронами типа II. Считают, что нейроны типа I - двигательные, а нейроны типа II - чувствительные. Отростки нейронов типа III не только вступают в контакт с нейронами соседних ганглиев, но и проникают в слизистую и подслизистую оболочки.
Синапсы. Аксон постганглионарного вегетативного нейрона образует многочисленные локальные утолщения - варикозные расширения, содержащие синаптические пузырьки (см. рис. 7-29). Эти утолщения - места секреции нейромедиатора.
Вегетативные нейроны синтезируют и секретируют различные биологически активные вещества (нейромедиаторы и модуляторы). Нейромедиаторы (ацетилхолин, норадреналин и серотонин) вызывают сокраще- ние/расслабление ГМК, возбуждение/торможение нейронов энтеральной нервной системы, усиление/подавление секреции экзокринных желёз и энтероэндокринных клеток. В некоторых нейронах ганглиев межмышечного сплетения совместно присутствуют серотонин и вещество P.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Головной отдел нервной трубки образует три расширения, или первичных мозговых пузыря: передний мозг (prosencephalon), средний мозг (mesencephalon) и ромбовидный мозг (rhombencephalon). На 5-й неделе передний мозг разделяется на две части: конечный мозг (telencephalon), образованный первичными полусферами мозга, и промежуточный мозг (diencephalon) с глазными пузырями. Средний мозг и ромбовидный мозг разделены глубоким перешейком. Ромбовидный мозг разделяется на задний мозг (metencephalon) и продолговатый мозг (myelencephalon). Из заднего мозга развиваются мост (pons cerebri) и мозжечок (cerebellum). Остальная часть нервной трубки формирует спинной мозг.
Спинной мозг
Спинной мозг состоит из двух симметричных половин (рис. 8-35), соединённых узкой перемычкой, содержащей центральный канал, остаток полости нервной трубки. На поперечном разрезе легко различимы лежащее снаружи белое вещество и расположенная глубже тёмная масса - серое вещество.
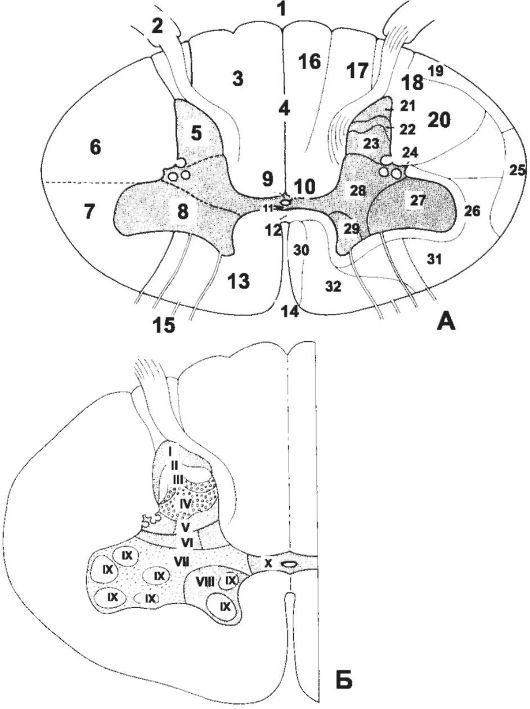 Рис. 8-35. Поперечный срез спинного мозга. А - структуры серого и белого вещества. Б - топография пластинок в сером веществе. 1 - Sulcus medianus posterior; 2 - Radix dorsalis; 3 - Funiculus posterior; 4 - Septum medianum posterius; 5 - Cornu posterius; 6 - Funiculus posterolateralis; 7 - Funiculus anterolateralis; 8 - Cornu anterius; 9 - Commissura grisea posterior; 10 - Canalis centralis; 11 - Commissura grisea anterior; 12 - Commissura alba; 13 - Funiculus anterior; 14 - Fissura mediana anterior; 15 - Radix ventralis; 16 - Fasciculus gracilis; 17 - Fasciculus cuneatus; 18 - Fasciculus dorsolateralis; 19 - Tractus spinocerebellaris posterior; 20 - Tractus pyramidalis lateralis; 21 - Cellulae marginales; 22 - Substantia gelatinosa; 23 - N. proprius; 24 - Processus reticularis; 25 - Tractus spinocerebellaris anterior; 26 - Fasciculi proprii; 27 - Cellulae motoriae laterales; 28 - Substantia intermedia; 29 - Cellulae motoriae mediales; 30 - Tractus pyramidalis anterior; 31 - Fasciculus anterolateralis: tractus spinotectalis, tractus spinothalamicus, tractus spinoannularis, tractus spino-olivaris; 32 - Fasciculus
longitudinalis medialis: tractus vestibulospinalis medialis, tractus
vestibulospinalis lateralis, tractus reticulospinalis, tractus
tectospinalis, tractus interstitiospinalis. [17]
Рис. 8-35. Поперечный срез спинного мозга. А - структуры серого и белого вещества. Б - топография пластинок в сером веществе. 1 - Sulcus medianus posterior; 2 - Radix dorsalis; 3 - Funiculus posterior; 4 - Septum medianum posterius; 5 - Cornu posterius; 6 - Funiculus posterolateralis; 7 - Funiculus anterolateralis; 8 - Cornu anterius; 9 - Commissura grisea posterior; 10 - Canalis centralis; 11 - Commissura grisea anterior; 12 - Commissura alba; 13 - Funiculus anterior; 14 - Fissura mediana anterior; 15 - Radix ventralis; 16 - Fasciculus gracilis; 17 - Fasciculus cuneatus; 18 - Fasciculus dorsolateralis; 19 - Tractus spinocerebellaris posterior; 20 - Tractus pyramidalis lateralis; 21 - Cellulae marginales; 22 - Substantia gelatinosa; 23 - N. proprius; 24 - Processus reticularis; 25 - Tractus spinocerebellaris anterior; 26 - Fasciculi proprii; 27 - Cellulae motoriae laterales; 28 - Substantia intermedia; 29 - Cellulae motoriae mediales; 30 - Tractus pyramidalis anterior; 31 - Fasciculus anterolateralis: tractus spinotectalis, tractus spinothalamicus, tractus spinoannularis, tractus spino-olivaris; 32 - Fasciculus
longitudinalis medialis: tractus vestibulospinalis medialis, tractus
vestibulospinalis lateralis, tractus reticulospinalis, tractus
tectospinalis, tractus interstitiospinalis. [17]
СЕРОЕ ВЕЩЕСТВО
Серое вещество состоит из отростков нервных клеток и их перикарионов, образующих скопления - ядра, объединённые в пластинки. Каждая половина серого вещества формирует на протяжении всего спинного мозга выступы - серые столбы: передний столб - columna anterior, задний столб - columna posterior и боковой столб - columna lateralis. Столб на поперечном разрезе получает название рога, соответственно передний (cornu anterius), задний (cornu posterius) и боковой (cornu laterale). Перикарионы нейронов серого вещества по длине спинного мозга картированы по десяти пластинкам (см. рис. 8-35). Топография ядер соответствует топографии пластинок, хотя они не всегда совпадают.
Нейроны
В сером веществе спинного мозга находятся тела двигательных, вставочных и вегетативных нейронов. Различают α-мотонейроны (крупные) и γ-мотонейроны (мелкие).
• α-Мотонейроны входят в состав медиальных и латеральных ядер. Это наиболее крупные клетки спинного мозга. Их аксоны образуют нервно-мышечные синапсы (рис. 8-29) с поперечно-полосатыми волокнами скелетной мышцы.
• γ-Мотонейроны иннервируют интрафузальные волокна мышечных веретён (рис. 8-27, см. также рис. 7-9).
• Вегетативные нейроны расположены в висцеральных ядрах грудного и поясничного отделов (n. intermediomedialis), а также в крестцовом отделе (крестцовые парасимпатические ядра) промежуточной зоны (substantia intermedia) серого вещества. Нейроны n. intermediomedialis образуют синаптические контакты с афферентными волокнами общей висцеральной чувствительности (общие висцеральные афференты), а их аксоны вступают в контакты с нейронами n. intermediolateralis.
БЕЛОЕ ВЕЩЕСТВО
Белое вещество состоит из нервных волокон и клеток нейроглии. Рога серого вещества разделяют белое вещество на три канатика (задние, боковые и передние). Проводящие пути образованы цепью нейронов, соединённых последовательно своими отростками и обеспечивают проведение возбуждения от нейрона к нейрону (от ядра к ядру). Различают пути восходящие, нисходящие и смешанные.
Восходящие пути
Восходящие пути образованы центральными отростками чувствительных нейронов спинномозговых узлов и аксонами вставочных нейронов. Центральные отростки чувствительных нейронов спинномозговых узлов образуют проводящие пути проприоцептивной и тактильной чувствительности, а именно тонкий пучок (fasciculus gracilis) и клиновидный пучок (fasciculus cuneatus).
Аксоны вставочных нейронов образуют проекционные афферентные пути спиноталамические, спиномозжечковые, спинооливный, спинопокрышечный, спиноретикулярный.
• Спиноталамический путь вентральный (tractus spinothalamicus ventralis) проходит в переднем канатике и проводит импульсы от механорецепторов кожи.
• Спиноталамический путь латеральный (tractus spinothalamicus lateralis) проходит в боковом канатике, это главный путь болевой и температурной чувствительности. Передача болевых импульсов. Вещество P участвует в передаче болевых стимулов в качестве возбуждающего нейромедиатора в синапсах между центральными отростками чувствительных нейронов спинномозгового узла и перикарионами нейронов спиноталамического пути. Блокирование секреции вещества P и снятие болевых ощущений реализуются через рецепторы опиоидных пептидов, встроенных в мембрану терминали центрального отростка чувствительного нейрона. Источник опиоидного пептида энкефалина - вставочный нейрон.
• Спиномозжечковый путь передний (tractus spinocerebellaris anterior) и спиномозжечковый путь задний (tractus spinocerebellaris posterior). По спиномозжечковым путям в мозжечок поступает информация о всех афферентных сигналах глубокой чувствительности и о всех изменениях мышечного тонуса.
• Спинопокрышечный путь (tractus spinotectalis) - проекционный восходящий путь общей чувствительности.
• Спиноретикулярный путь (tractus spinoreticularis) - путь поступления афферентной информации соматических и висцеральных рефлексов.
Нисходящие пути
Нисходящие пути (супраспинальные и спинальные) заканчиваются исключительно на мотонейронах.
Супраспинальные пути проводят сигналы от структур, расположенных выше спинного мозга. К ним относятся пирамидные пути (корковоспинномозговой путь передний (tractus corticospinalis ventralis; корковоспинномозговой путь латеральный, tractus corticospinalis lateralis). тектоспинальный путь (покрышечно-спинномозговой, tractus tectospinalis), красноядерно-спинномозговой путь (tractus rubrospinalis, нисходящий проекционный путь экстрапирамидной системы), ретикулоспинальный путь (tractus reticulospinal, эфферентный путь экстрапирамидной системы, контролирует тонус скелетной мускулатуры и висцеральные двигательные функции), преддверно-спинномозговой путь (tractus vestibulospinalis), оливо-спинномозговой путь (tractus olivospinalis). Спинальные пути расположены в пределах спинного мозга.
• В задних канатиках располагаются нисходящие волокна центральных отростков чувствительных нейронов спинномозговых узлов: в шейных и верхних грудных сегментах они образуют пучок в форме запятой (fasciculus interfascicularis), а в поясничном отделе - септомаргинальный пучок (fasciculus septomarginalis).
• В передних канатиках шейного и грудного отделов спинного мозга волокна медиального преддверно-спинномозгового пути образуют бороз-
дчато-краевой пучок (fasciculus sulcomarginalis), контролирующие тонус мышц шеи в соответствии с различными положениями головы.
Смешанные пути
К смешанным восходящим и нисходящим путям относятся краевой пояс, собственные пучки спинного мозга, а также генератор центрального паттерна.
Генератор центрального паттерна - локальная сеть нейронов в пределах спинного мозга, контролирующая и координирующая сложный цикл стереотипных движений при ходьбе, беге, прыжках, плавании и т.д.
Головной мозг
МОЗЖЕЧОК
Мозжечок (cerebellum) координирует движения и равновесие, расположен над продолговатым мозгом и мостом и связан со стволом мозга тремя парами ножек, по которым проходят афферентные и эфферентные проводящие пути. В глубине белого вещества мозжечка лежат скопления нейронов - ядра мозжечка. Извилины мозжечка, образующие на разрезе фигуру разветвлённого дерева (arbor vitae), разделены глубокими бороздами.
Кора мозжечка
Каждая извилина содержит узкую пластинку белого вещества, полностью покрытую серым веществом (кора мозжечка, рис. 8-37), в котором различают три слоя: наружный - молекулярный, средний - ганглионарный и внутренний - зернистый.
• Ганглионарный слой, или слой грушевидных нейронов (слой клеток Пуркинье) образуют тела крупных нервных клеток. Их перикарионы расположены примерно на одном уровне от поверхности коры. От тела в молекулярный слой отходят 2-3 сильно ветвящихся дендрита. Через зернистый слой в белое вещество от тела грушевидных нейронов отходит аксон. Аксоны грушевидных нейронов - единственные эфферентные волокна, выходящие из коры мозжечка. Они заканчиваются на нейронах ядер мозжечка. Вблизи тела клетки от аксона отходят коллатерали, направляющиеся обратно в ганглионарный слой и глубокие части молекулярного слоя. На грушевидных нейронах так или иначе заканчиваются все афферентные пути мозжечка (рис. 8-37). Молекулярный слой содержит корзинчатые и звездчатые клетки. Корзинчатые клетки образуют многочисленные длинные и сравнительно мало раз- ветвлённые дендриты. Их аксон направлен параллельно поверхности мозжечка в той же плоскости, в которой расположены ветвления дендритов грушевидных нейронов. На всём протяжении аксон образует ветви, заканчивающиеся в виде корзинок на телах грушевидных нейронов. Активность корзинчатых нейронов вызывает торможение грушевидных нейронов.
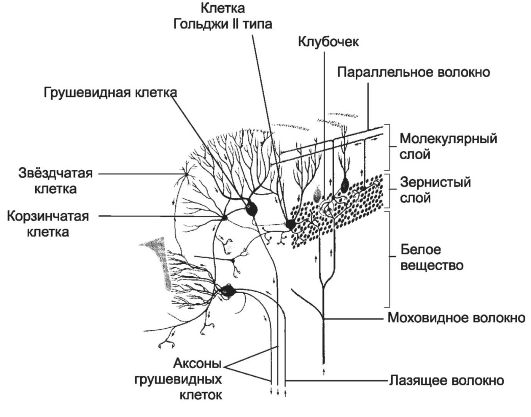 Рис. 8-37. Организация коры мозжечка. Кора
образована тремя слоями: наружный - молекулярный, средний -
ганглионарный и внутренний - зернистый. Из подлежащего белого вещества в
кору проходят афферентные лазящие и моховидные волокна, а выходят
аксоны грушевидных клеток. Стрелками указано направление распространения
возбуждения. [17]
Рис. 8-37. Организация коры мозжечка. Кора
образована тремя слоями: наружный - молекулярный, средний -
ганглионарный и внутренний - зернистый. Из подлежащего белого вещества в
кору проходят афферентные лазящие и моховидные волокна, а выходят
аксоны грушевидных клеток. Стрелками указано направление распространения
возбуждения. [17]
• Звездчатые клетки расположены ближе к поверхности коры. Их аксоны образуют синаптические контакты с дендритами грушевидных нейронов. Эти нейроны вместе с корзинчатыми образуют систему вставочных нейронов, передающую тормозные импульсы на дендриты и тела грушевидных клеток в плоскости, поперечной извилинам.
Зернистый слой содержит клетки-зёрна, звездчатые клетки Гольджи и веретеновидные горизонтальные нейроны.
• Клетки-зёрна. Тело их весьма невелико и практически полностью занято ядром. 3-4 очень коротких дендрита образуют концевые разветвления, напоминающие птичьи лапки. Аксоны клеток-зёрен поднимаются в молекулярный слой, где образуют T-образные разветвления, идущие параллельно поверхности мозжечка в плоскости, совпадающей с направлением извилины. Это параллельные волокна, образующие синапсы с дендритами: грушевидных нейронов, корзинчатых клеток, звездчатых клеток, клеток Гольджи типа II.
• Звездчатые клетки Гольджи. Различают звездчатые нейроны с короткими и длинными аксонами. Крупные перикарионы звездчатых нейронов с короткими аксонами нередко лежат непосредственно под ганглионарным слоем, а большая часть дендритов разветвляется в молекулярном слое и образует синапсы с параллельными волокнами - аксонами клеток-зёрен. Их короткие аксоны, входя в состав клубочков мозжечка (glomeruli cerebellosi), заканчиваются синаптическими контактами на розетках моховидных волокон.
• Веретеновидные горизонтальные нейроны имеют небольшой вытянутый перикарион, от которого отходят длинные горизонтальные дендриты, заканчивающиеся в слое грушевидных нейронов и зернистом слое. Аксоны этих нейронов образуют коллатерали в зернистом слое и уходят в белое вещество.
Афференты мозжечка
В кору мозжечка входят многочисленные волокна из различных отделов мозга. В зернистом слое находятся моховидные волокна. Лазящие волокна заканчиваются в молекулярном слое на дендритах грушевидных нейронов.
• Моховидные волокна, проникнув в зернистый слой, ветвятся и формируют концевые розетки, вступающие в контакт с дендритами клеток-зёрен в составе клубочков мозжечка. Моховидные волокна образуют также синапсы с дендритами клеток Гольджи типа II. Следовательно, моховидные волокна вступают в контакт как с короткими аксонами звездчатых нейронов зернистого слоя, так и с их дендритами.
• Лазящие волокна подходят к телам грушевидных нейронов и здесь распадаются на несколько тонких веточек, оплетающих дендриты. На один грушевидный нейрон приходится одно лазящее волокно.
КОРА БОЛЬШОГО МОЗГА
Кора большого мозга (неокортекс) содержит 6 слоёв нервных клеток
(рис. 8-39).
Слои коры
Снаружи внутрь слои коры располагаются в следующем порядке: молекулярный, наружный зернистый, наружный пирамидный, внутренний зернистый, внутренний пирамидный (ганглионарный) и полиморфный (мультиформный).
I. Молекулярный. Содержит редкие перикарионы, здесь проходят аксоны и дендриты.
II. Наружный зернистый. В наружном зернистом слое присутствуют небольшие пирамидные и звездчатые нейроны.
III. Наружный пирамидный. Представлен многочисленными пирамидными нейронами средней величины; размеры их перикариона возрастают в глубоких частях слоя.
IV. Внутренний зернистый. Содержит мелкие звездчатые клетки.
V. Внутренний пирамидный (ганглионарный). Состоит из крупных пирамидных нейронов и небольшого количества звездчатых клеток.
VI. Полиморфный. Образован множеством нейронов различной величины и формы, а также некоторым количеством пирамидных и зернистых нейронов.
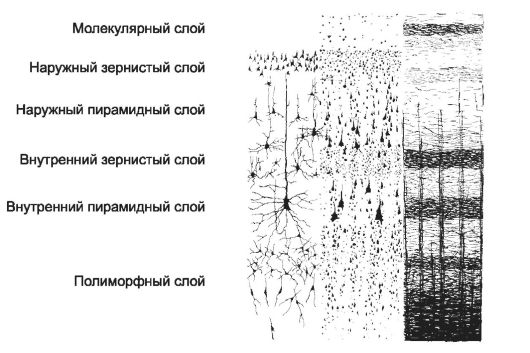 Рис. 8-39. Цитоархитектоника и миелоархитектоника коры большого мозга. Используя
различные методы окрашивания, можно получить картину строения коры с
хорошо различимыми отростками нейронов (левая часть рисунка), только
перикарионами (средняя часть рисунка) или миелиновыми волокнами (правая
часть рисунка). В последнем случае на уровне внутреннего зернистого и
внутреннего пирамидного слоёв видны компактные скопления миелиновых
волокон в виде соответственно полоски наружной и внутренней зернистой
пластинки. [17]
Рис. 8-39. Цитоархитектоника и миелоархитектоника коры большого мозга. Используя
различные методы окрашивания, можно получить картину строения коры с
хорошо различимыми отростками нейронов (левая часть рисунка), только
перикарионами (средняя часть рисунка) или миелиновыми волокнами (правая
часть рисунка). В последнем случае на уровне внутреннего зернистого и
внутреннего пирамидного слоёв видны компактные скопления миелиновых
волокон в виде соответственно полоски наружной и внутренней зернистой
пластинки. [17]
Нейроны коры
Пирамидные нейроны. Размеры перикариона - 10-100 мкм, имеют длинный апикальный дендрит, выходящий из вершины пирамиды, и другие дендриты, отходящие от боковых поверхностей перикариона. От основания пирамиды отходит аксон, уходящий в белое вещество. Возвратные коллатеральные ветви аксона заканчиваются на других пирамидных нейронах или вставочных корковых нейронах. Гигантские пирамидные нейроны расположены в слое V двигательной коры, это самые крупные нейроны коры. Размеры их перикариона более 100 мкм, они дают начало крупным миелинизированным аксонам пирамидного тракта. Крупные пирамидные нейроны расположены также в слое V зрительной коры затылочной доли. Они посылают аксоны в ствол мозга и участвуют в рефлексе движения глаза.
Пирамидная система. В осознанном контроле за произвольными движениями принимает участие кора лобных долей головного мозга (предцентральная извилина), откуда нервные импульсы через мотонейроны спинного мозга поступают к ске-
летным мышцам конечности. Важнейшим нейромедиатором в передаче импульсов между нейронами в данной системе является ацетилхолин. Звездчатые нейроны. Их перикарионы имеют округлую, полигональную или треугольную форму, 4-8 мкм в диаметре. Аксон и дендриты отходят на короткое расстояние от перикариона и участвуют в образовании внутрикорковых связей.
Веретеновидные нейроны чаще встречаются в слое VI. От противоположных концов перикариона отходят дендриты. Аксон уходит глубоко в белое вещество. Инвертированные пирамидные нейроны присутствуют во всех слоях, кроме первого. Имеют перикарион полигональной формы и короткие дендриты. Аксон направляется вертикально к поверхности коры, отдавая коллатерали во всех слоях. Горизонтальные нейроны находятся в слое I. От их веретеновидного перикариона отходит длинный аксон, который вместе с дендритами образует горизонтальные связи в пределах слоя I.
Модули и цилиндры коры
Кора больших полушарий состоит из модулей и цилиндров.
• Модуль имеет в поперечнике около 0,1 мм и пронизывает всю толщу коры. В зрительной коре типичный модуль включает более 100 тысяч синаптически связанных клеток, образующих локальные нейронные сети. Модули контактируют друг с другом коллатералями дендритов и аксонов.
• Цилиндры. Существует представление о вертикальных (радиальных) связях от одной до трёх сотен нейронов коры, в совокупности образующих некую функциональную единицу, называемую цилиндром. Подобная структура может достигать в диаметре нескольких сотен мкм. Цилиндры наиболее наглядно прослеживаются в сенсорных полях коры, где они получают информацию от определённых групп афферентных нейронов.
БАЗАЛЬНЫЕ ГАНГЛИИ
Базальные ганглии образуют комплекс подкорковых скоплений нейронов, расположенных в центральном белом веществе полушарий большого мозга. Эти ганглии обеспечивают регуляцию двигательных и вегетативных функций, участвуют в осуществлении интегративных процессов высшей нервной деятельности. Базальные ганглии объединяют хвостатое ядро (nucleus caudatus), скорлупу (putamen), бледный шар (globus pallidus) и чёрное вещество (substantia nigra). Базальные ганглии получают импульсацию от лобной коры, ответственной за контроль произвольных движений, и опосредуют обратный непроизвольный контроль за движениями через премоторную кору и таламус.
• Двигательные ядра базальных ганглиев подразделяют на несколько функционально различных групп. Одна из них называется полосатым телом. Оно включает хвостатое ядро и скорлупу. Обе эти структуры полосатого тела собирают информацию из других областей мозга. Нейроны практически всех областей неокортекса направляют аксоны в полосатое тело. В скорлупе заканчиваются аксоны
нейронов, расположенных в височной, теменной и лобной коре, а также в компактной части чёрного вещества.
• Экстрапирамидная система. Для целостного процесса движения важным является контроль не только за произвольными, но и за непроизвольными движениями. Эту функцию выполняет экстрапирамидная система, обеспечивающая плавность движений и возможность прервать начатое действие. Эта система объединяет базальные ганглии, располагающиеся за пределами продолговатого мозга вне пирамид (отсюда термин "экстрапирамидная система"). Бессознательный контроль за движениями, осуществляемый этой системой, обеспечивает нейромедиатор дофамин.
ОБОЛОЧКИ МОЗГА
Головной и спинной мозг защищены оболочками (рис. 8-42). Мягкая мозговая оболочка (pia mater) непосредственно прилегает к мозгу. Снаружи проходит твёрдая мозговая оболочка (dura mater). Между ними расположена средняя, паутинная оболочка (t. arachnoidea). Все оболочки образованы волокнистой соединительной тканью. Мягкую и паутинную оболочки можно рассматривать как одно целое под общим названием pia-arachnoidea, или leptomeninx.
• Мягкая мозговая оболочка содержит переплетающиеся пучки коллагеновых волокон и сеть эластических волокон, множество равномерно распределённых кровеносных сосудов. Снаружи оболочка покрыта слоем плоских клеток. Оболочка повторяет ход борозд и извилин мозга.
• Паутинная оболочка - сеть тонких соединительнотканных перегородок (трабекул), состоящих преимущественно из коллагеновых волокон и небольшого коли-
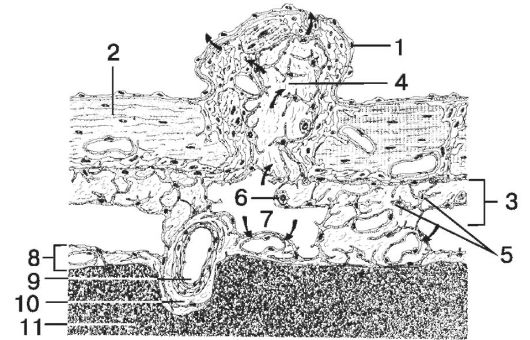 Рис. 8-42. Мозговые оболочки. 1
- эндотелиальная клетка; 2 - твёрдая мозговая оболочка; 3 - паутинная
оболочка; 4 - соединительная ткань; 5 - клетки паутинной оболочки; 6 -
макрофаг; 7 - субарахноидальное пространство; 8 - мягкая мозговая
оболочка; 9 - кровеносный сосуд; 10 - периваскулярная соединительная
ткань; 11 - мозг. [17]
Рис. 8-42. Мозговые оболочки. 1
- эндотелиальная клетка; 2 - твёрдая мозговая оболочка; 3 - паутинная
оболочка; 4 - соединительная ткань; 5 - клетки паутинной оболочки; 6 -
макрофаг; 7 - субарахноидальное пространство; 8 - мягкая мозговая
оболочка; 9 - кровеносный сосуд; 10 - периваскулярная соединительная
ткань; 11 - мозг. [17]
• чества эластических волокон. Паутинная оболочка изнутри и снаружи выстлана непрерывным слоем тонких уплощённых клеток. Пространство между трабекулами заполнено цереброспинальной жидкостью и называется субарахноидальным. Паутинная оболочка в области борозд мозга не прилегает вплотную к мягкой мозговой оболочке. Здесь присутствуют цистерны, содержащие большое количество цереброспинальной жидкости. Твёрдая мозговая оболочка состоит из плотной волокнистой соединительной ткани, в которой преобладают коллагеновые волокна. Пространство между твёрдой и паутинной оболочками называется субдуральным. Оно содержит небольшое количество жидкости, отличной от цереброспинальной. Снаружи твёрдая мозговая оболочка покрыта рыхлой соединительной тканью с большим количеством вен. Твёрдая мозговая оболочка образует складки, которые вместе с надкостницей формируют синусы, выстланные эндотелием и содержащие венозную кровь. Пространство между твёрдой мозговой оболочкой и надкостницей называется эпидуральным. Оно содержит рыхлую соединительную ткань и внутреннее позвоночное венозное сплетение.
ОРГАНЫ ЧУВСТВ
Органы чувств - сенсорные структуры для восприятия зрительных, слуховых, обонятельных и вкусовых раздражений. Соответственно это органы зрения, слуха и равновесия, обоняния и вкуса.
Зрение
Орган зрения состоит из глазного яблока, соединённого через зрительный нерв с мозгом, и включает вспомогательный аппарат в виде век, слёзной железы и поперечно-полосатых глазодвигательных мышц.
Глазное яблоко
Стенка глазного яблока образована тремя оболочками: наружной фиброзной (в задней части непрозрачная склера, переходящая в прозрачную роговицу в передней; лимб - граница между роговицей и склерой), средней - собственно сосудистой (choroidea) и внутренней - сетчатой (рис. 8-45).
Роговица (cornea) - прозрачная оболочка передней стенки глаза, состоит из пяти слоёв. На поверхности роговицы находится плёнка секретов слёзных и слизистых желёз, содержащая лизоцим, лактоферрин, иммуноглобулины. Переднюю поверхность роговицы образует многослойный плоский неороговевающий эпителий. Не содержащий клеток гомогенный слой основного вещества и неупорядоченно ориентированных тонких коллагеновых и ретикулиновых волокон - передняя пограничная мембрана - поддерживает форму роговицы. Собственное вещество представлено правильно расположенными коллагеновыми пластинками и уплощёнными фибробластами, погружёнными в мат-
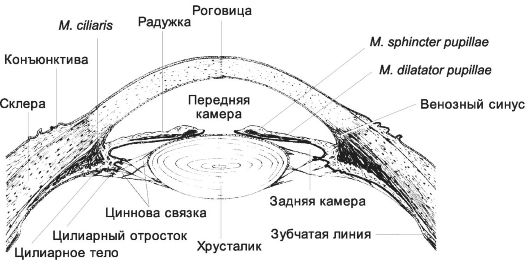 Рис. 8-45. Передний отдел глаза. [17]
Рис. 8-45. Передний отдел глаза. [17]
рикс из гликозаминогликанов. Задняя пограничная мембрана - прозрачный слой роговицы, расположен между собственным веществом и эндотелием задней поверхности роговицы, состоит из коллагеновых волокон типа VIII и аморфного вещества. Эндотелий роговицы ограничивает спереди переднюю камеру глаза, это один слой практически непролиферирующих клеток, выполняющий барьерную и «насосную» функции, необходимые для гидратации стромы роговицы и для поддержания её прозрачности.
Склера - наружная непрозрачная оболочка глазного яблока. Склера построена из плотных тяжей коллагеновых волокон, между которыми находятся уплощённой формы фибробласты. В месте соединения склеры с роговицей расположены небольшие сообщающиеся полости, в совокупности образующие венозный синус склеры (sinus venosus sclerae), обеспечивающий отток жидкости из передней камеры глаза. Сосудистая оболочка осуществляет питание сетчатки. Эта оболочка состоит из нескольких пластинок (слоёв): надсосудистой, сосудистой, хориокапиллярной и базальной.
• Надсосудистая пластинка (lamina suprachoroidea) расположена на границе со склерой, состоит из рыхлой волокнистой соединительной ткани с многочисленными пигментными клетками.
• Сосудистая пластинка (lamina vasculosa) содержит сплетение артерий и вен; в рыхлой соединительной ткани располагаются пигментные клетки и ГМК.
• Сосудисто-капиллярная (lamina choroidocapillaris) пластинка образована сплетением капилляров синусоидного типа.
• На границе с сетчаткой располагается базальная пластинка (lamina basalis) (базальная мембрана и эластические волокна). В передней части глаза сосудистая оболочка образует радужку и цилиарное тело.
Радужка (iris) - продолжение сосудистой оболочки глаза, расположена между роговицей и хрусталиком, разделяет переднюю и заднюю камеры глаза и содержит эндотелий, наружный пограничный, сосудистый, внутренний пограничный и задний пигментный слои. Эндотелий (передний эпителий) - продолжение эндотелия роговицы. Наружный и внутренний пограничные слои имеют сходное строение и содержат фибробласты и меланоциты, погружённые в основное вещество. Сосудистый слой - рыхлая соединительная ткань, содержащая многочисленные сосуды и меланоциты. Задний пигментный слой переходит в двуслойный эпителий сетчатки, покрывающий ресничное тело. В составе радужки ГМК образуют сфинктер зрачка (m. sphincter pupillae) и дилататор зрачка (m. dilatator pupillae). При раздражении парасимпатических (холинергических) нервных волокон зрачок сужается, симпатическая стимуляция приводит к расширению зрачка. Ресничное тело. В области угла глаза сосудистая оболочка утолщается, образуя ресничное тело (corpus ciliare), имеющее на срезе вид треугольника, обращённого основанием в переднюю камеру. Основную массу ресничного тела занимает ресничная мышца (m. ciliaris). В её составе ГМК проходят в трёх взаимно перпендикулярных направлениях. От ресничного тела отходят по направлению к хрусталику ресничные отростки (processus ciliares). Они содержат массу капилляров, покрыты двумя слоями эпителия - пигментным и секреторным, продуцирующим водянистую влагу. К цилиарным отросткам прикрепляется ресничный поясок (zonula ciliaris). Постганглионарные холинергические волокна из ресничного ганглия заканчиваются в ресничной мышце. При сокращении ресничной мышцы ресничный поясок расслабляется, и выпуклость хрусталика увеличивается.
Хрусталик (lens) имеет вид двояковыпуклой линзы. Его передняя поверхность покрыта однослойным кубическим эпителием, который по направлению к экватору становится выше. Эпителиальные клетки хрусталика связаны щелевыми контактами. Прозрачные хрусталиковые волокна (fibrae lentis) составляют основную часть хрусталика и содержат кристаллины. Капсула хрусталика (capsula lentis) - толстая базальная мембрана со значительным содержанием ретикулиновых волокон.
Передняя камера спереди ограничена роговицей, сзади - радужкой, а в пределах зрачка - центральной частью передней поверхности хрусталика. Глубина передней камеры наибольшая в центральной части, где достигает 3-3,5 мм. Угол между задней поверхностью периферической части роговицы и передней поверхностью корня радужки - угол передней камеры, он расположен в области перехода роговицы в склеру и радужки в цилиарное тело.
Задняя камера - пространство за радужкой, ограниченное хрусталиком, ресничным и стекловидным телом.
Водянистая влага (humor aquosus) - жидкость, содержащая растворимые белки плазмы и деполимеризованную гиалуроновую кислоту. Водянистая влага не содержит фибриногена и гипертонична по отношению к крови. Большая часть водянистой влаги поступает из капилляров и эпителия ресничных отростков в заднюю камеру и проходит между радужкой и хрусталиком в переднюю камеру. Трабекулярная сеточка (reticulum trabeculare) образована элементами роговицы, радужки и ресничного тела и имеет важное значение для оттока водянистой влаги из передней камеры глаза.
• Внутриглазное давление (23 мм рт.ст.) зависит от равновесия между количеством образуемой и удаляемой водянистой влаги; скорость её образования и всасывания составляет примерно 2 мм3/мин. Главный путь оттока водянистой влаги: трабекулярная сеть → венозный синус склеры → венозные сосуды глаза.
• Гематотканевые барьеры регулируют обмен между кровью и тканями глаза. Обмен между кровью и водянистой влагой осуществляется в радужке и ресничном теле. Клетки ресничного эпителия связаны плотными контактами, ограничивающими транспорт макромолекул. Капилляры радужки не имеют фенестр.
Стекловидное тело (corpus vitreum) - прозрачная среда глаза, заполняет полость между хрусталиком и сетчатой оболочкой. Стекловидное тело - гель, содержащий воду, коллагены типа II, IX и XI, белок витреин и гиалуроновую кислоту. Через стекловидное тело от сетчатки к хрусталику проходит стекловидный канал (canalis hyaloideus) - остаток эмбриональной сосудистой системы глаза. Стекловидная мембрана (membrana vitrea) - скопление коллагеновых волокон по периферии стекловидного тела, как бы формирующее его капсулу.
Сетчатая оболочка
Сетчатая оболочка (сетчатка, retina) - внутренняя оболочка глаза, имеет зрительный отдел, по зубчатому краю (ora serrata) переходящий в слепой отдел, покрывающий сзади ресничное тело и радужку. У заднего края оптической оси глаза сетчатка имеет округлое жёлтое пятно (macula lutea) диаметром около 2 мм. Центральная ямка (fovea centralis) - углубление в средней части жёлтого пятна, место наилучшего восприятия. Зрительный нерв (nervus opticus) выходит из сетчатки медиальнее жёлтого пятна. Здесь образуется диск зрительного нерва (discus nervi optici). В центре диска имеется углубление, в котором видны питающие сетчатку сосуды.
Слои сетчатки. Зрительная часть сетчатки образована последовательно (от наружной поверхности глазного яблока) расположенными слоями (рис. 8-46): пигментный, наружный ядерный, наружный сетчатый, внутренний ядерный, внутренний сетчатый и ганглионарный.
• Пигментный слой (stratum pigmentosum). Клетки полигональной формы, прилежащие к сосудистой оболочке. Одна клетка пигментного эпителия взаимодействует
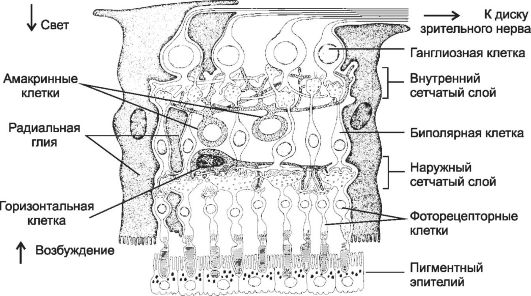 Рис. 8-46. Сетчатка. Стрелками
слева обозначены направления светового потока и возбуждения. Пигментный
эпителий окружает наружные сегменты фоторецепторных клеток, образующих
синаптические контакты с биполярными нейронами. Информация от биполярных
клеток передаётся ганглиозным клеткам и по их аксонам, образующим
зрительный нерв, уходит в мозг. Промежутки между нейронами заполняют
крупные клетки радиальной глии. Их наружные отростки заканчиваются на
границе между наружными и внутренними сегментами фоторецепторных клеток.
[17]
Рис. 8-46. Сетчатка. Стрелками
слева обозначены направления светового потока и возбуждения. Пигментный
эпителий окружает наружные сегменты фоторецепторных клеток, образующих
синаптические контакты с биполярными нейронами. Информация от биполярных
клеток передаётся ганглиозным клеткам и по их аксонам, образующим
зрительный нерв, уходит в мозг. Промежутки между нейронами заполняют
крупные клетки радиальной глии. Их наружные отростки заканчиваются на
границе между наружными и внутренними сегментами фоторецепторных клеток.
[17]
с наружными сегментами десятков фоторецепторных клеток - палочек и колбочек. Клетки пигментного эпителия запасают витамин А, участвуют в его превращениях и передают его производные фоторецепторным клеткам для образования зрительного пигмента.
• Наружный ядерный слой (stratum nucleare externum) включает ядросодержащие части фоторецепторных клеток. Колбочки концентрируются в области жёлтого пятна. Глазное яблоко организовано таким образом, что на колбочки падает центральная часть светового пятна от визуализируемого объекта. По периферии от жёлтого пятна расположены палочки.
• Наружный сетчатый (stratum plexiforme externum). Здесь осуществляются контакты внутренних сегментов палочек и колбочек с дендритами биполярных клеток.
• Внутренний ядерный (stratum nucleare internum). Содержит биполярные клетки, связывающие палочки и колбочки с ганглиозными клетками, а также горизонтальные и амакринные клетки. Перикарионы амакринных клеток расположены во внутренней части внутреннего ядерного слоя.
• Внутренний сетчатый (stratum plexiforme internum). В нём биполярные клетки контактируют с ганглиозными клетками, амакринные клетки выступают в качестве
вставочных нейронов. Популярна концепция о том, что ограниченное число биполярных клеток передает информацию 16 типам ганглиозных клеток при участии не менее 20 типов амакринных клеток.
• Ганглионарный слой (stratum ganglionicum) содержит ганглиозные нейроны. Общая схема передачи информации в сетчатке такова: рецепторная клетка → биполярная клетка → ганглиозная клетка и одновременно амакринная клетка → ганглиозная клетка.
Фоторецепторные клетки - палочки (рис. 8-47) и колбочки. Периферические отростки фоторецепторных клеток состоят из наружного и внутреннего сегментов, соединённых ресничкой. Различают центральное и периферическое зрение, что связано с характером распределения в сетчатке палочек и колбочек. В области центральной ямки расположены преимущественно колбочки. Каждая колбочка центральной ямки образует синапс только с одним биполярным нейроном. Центральное зрение, а также острота зрения реализуются колбочками. Периферическое зрение, а также ночное зрение и восприятие подвижных объектов - функции палочек.
• Наружный сегмент имеет множество уплощённых замкнутых дисков, содержащих зрительные пигменты: родопсин - в палочках; красный, зелёный и синий пигменты - в колбочках.
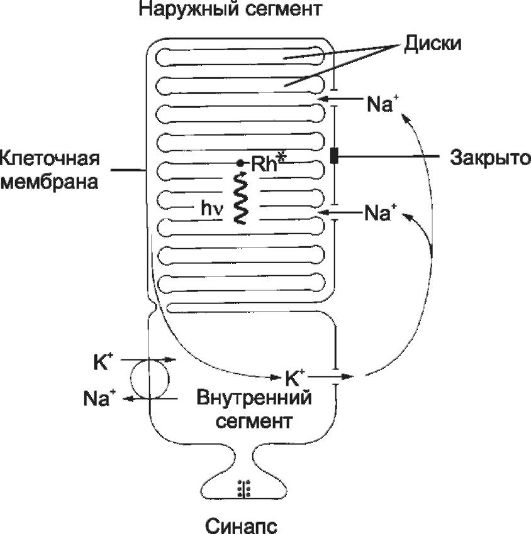 Рис. 8-47. Схема палочки. В
наружном сегменте расположена стопка дисков, содержащих зрительный
пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет
(h) активирует родопсин Rh* в дисках. [17]
Рис. 8-47. Схема палочки. В
наружном сегменте расположена стопка дисков, содержащих зрительный
пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет
(h) активирует родопсин Rh* в дисках. [17]
• Внутренний сегмент заполнен митохондриями и содержит базальное тельце, от которого в наружный сегмент отходит 9 пар микротрубочек.
• Цветовосприятие - функция колбочек. Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов.
• Зрительный пигмент состоит из апопротеина (опсин), ковалентно связанного с хромофором (11-цис-ретиналь или 11-^мс-дегидроретиналь).
Нейроны сетчатки (биполярные, горизонтальные, амакринные и ганглиозные клетки) синтезируют ацетилхолин, дофамин, L-глутамино- вую кислоту, глицин, γ-аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды.
• Горизонтальные клетки. Перикарионы расположены в наружной части внутреннего ядерного слоя, а отростки входят в область синапсов между фоторецепторными и биполярными клетками. Горизонтальные клетки получают информацию от колбочек и передают её также колбочкам.
• Амакринные клетки. Их перикарионы находятся во внутренней части внутреннего ядерного слоя в области синапсов между биполярными и ганглиозными клетками.
• Биполярные клетки реагируют на контрастность изображения. Некоторые биполяры сильнее реагируют на цветной, нежели на чёрно-белый контраст. Одни получают информацию преимущественно от палочек, другие - от колбочек.
• Ганглиозные клетки - крупные мультиполярные нейроны многих разновидностей. Их аксоны образуют зрительный нерв. Ганглиозные клетки реагируют на множество свойств зрительного объекта (например, на светлые и тёмные объекты, однородность освещения, цвет объекта, его ориентацию).
Глия сетчатки. Кроме нейронов, сетчатка содержит крупные клетки радиальной глии (см. рис. 8-46). Их ядра расположены во внутреннем ядерном слое. Наружные отростки заканчиваются микроворсинками, образуя наружный пограничный слой. Внутренние отростки имеют расширение (ножку) во внутреннем пограничном слое на границе со стекловидным телом. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки.
Слёзная железа
Слёзная железа образована группой сложных трубчато-альвеолярных желёзок; их секреторные отделы окружены миоэпителиальными клетками. Секрет железы, слёзная жидкость, по 6-12 протокам поступает в свод конъюнктивы и омывает роговицу и конъюнктиву.
Обоняние
Периферический отдел обонятельного анализатора (рис. 8-49) представлен обонятельной выстилкой (area olfactoria), которая занимает среднюю часть верхней носовой раковины и соответствующий ей участок слизистой оболочки перегородки носа площадью 2,5 см2. Обо-
нятельный эпителий содержит рецепторные клетки, общее количество которых достигает 50 млн. Их центральные отростки (аксоны) передают информацию в обонятельную луковицу. Обонятельные рецепторные клетки окружены опорными клетками. Обонятельный эпителий содержит также пигментные клетки, от которых зависит жёлтый цвет обонятельной выстилки. В подэпителиальной соединительной ткани расположены концевые отделы боуменовых желёз, кровеносные сосуды и пучки безмиелиновых волокон обонятельного нерва. Слизь, секретируемая боуменовыми железами, покрывает поверхность обонятельной выстилки слоем толщиной 60 мкм. В процессе хемовосприятия участвуют обонятельные реснички, погружённые в слизь. Обонятельный нерв - совокупность тонких обонятельных нитей, проходящих через отверстия решётчатой кости к обонятельным луковицам и образующих пучки по 10-100 нитей.
Обонятельная клетка
Морфология. Тело обонятельной клетки содержит многочисленные митохондрии, цистерны эндоплазматической сети с рибосомами, элементы комплекса Гольджи, лизосомы. Обонятельные клетки, кроме центрального (аксона), имеют короткий периферический отросток (дендрит), заканчивающийся на поверхности обонятельного эпителия сферическим утолщением - обонятельной булавой диаметром 1- 2 мкм. В ней присутствуют митохондрии, мелкие вакуоли и базальные тельца для отходящих от вершины булавы 20-80 обонятельных волосков длиной 30-200 мкм, имеющих строение типичных ресничек. В мембране обонятельных ресничек присутствуют рецепторы пахучих веществ.
Функция. Рецепторные клетки обонятельной выстилки регистрируют 25-35 первичных запахов, но их комбинации образуют много миллионов воспринимаемых запахов. Нейроны обонятельной выстилки полости носа человека располагают 350 различными рецепторами пахучих веществ. В каждом нейроне обонятельной выстилки экспрессируется только один ген рецептора пахучего вещества, но каждый рецептор распознает множество пахучих веществ. Таким образом, конкретный запах зависит от активации определённой комбинации рецепторов. Регенерация. Продолжительность жизни обонятельных клеток - 40 дней. Обонятельные рецепторные клетки составляют исключение среди нейронов: они постоянно обновляются за счёт клеток-предшественниц, т.е. относятся к обновляющейся клеточной популяции. Предшественницы рецепторных клеток - базальные клетки эпителия обонятельной выстилки.
Вкус
Периферическая часть вкусового анализатора - вкусовые почки (рис. 8-51). Они обнаружены в эпителии рта, языка, переднего отдела глот-
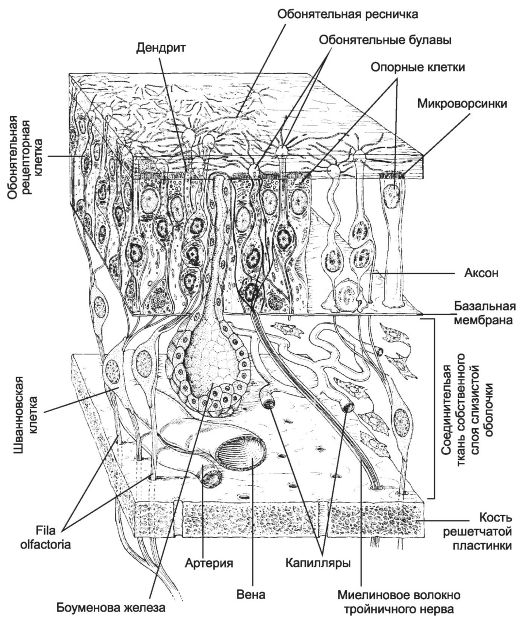 Рис. 8-49. Орган обоняния. В
эпителии обонятельной выстилки расположены обонятельные рецепторные
нейроны. Их наружные отростки заканчиваются обонятельными булавами, от
которых в разные стороны параллельно поверхности эпителия отходят
обонятельные волоски. Аксоны рецепторных нейронов в составе обонятельных
нитей (fila olfactoria) обонятельного нерва проходят через lamina cribrosa решётчатой кости и образуют синапсы с нейронами обонятельной луковицы (bulbus olfactorius). Многочисленные опорные клетки окружают рецепторные клетки. [17]
Рис. 8-49. Орган обоняния. В
эпителии обонятельной выстилки расположены обонятельные рецепторные
нейроны. Их наружные отростки заканчиваются обонятельными булавами, от
которых в разные стороны параллельно поверхности эпителия отходят
обонятельные волоски. Аксоны рецепторных нейронов в составе обонятельных
нитей (fila olfactoria) обонятельного нерва проходят через lamina cribrosa решётчатой кости и образуют синапсы с нейронами обонятельной луковицы (bulbus olfactorius). Многочисленные опорные клетки окружают рецепторные клетки. [17]
ки, пищевода и гортани. Их основная локализация - хемочувствительные сосочки языка: грибовидные, желобоватые и листовидные. У детей и реже у взрослых вкусовые почки встречаются в эпителии губ, надгортанника и даже голосовых связок.
Вкусовая почка
Вкусовая почка (gemma gustatoria) имеет эллипсоидную форму, высоту 27-115 мкм и ширину 16-70 мкм. В её апикальном отделе находится заполненный аморфным веществом вкусовой канал, открывающийся на поверхность эпителия вкусовой порой. Вкусовая почка образована 30-80 удлинёнными клетками, тесно прилегающими одна к другой. Большинство этих клеток вступает в контакт с нервными волокнами, проникающими в почку из подэпителиального нервного сплетения. Клетки вкусовых почек морфологически неоднородны. Выделяют четыре типа клеток.
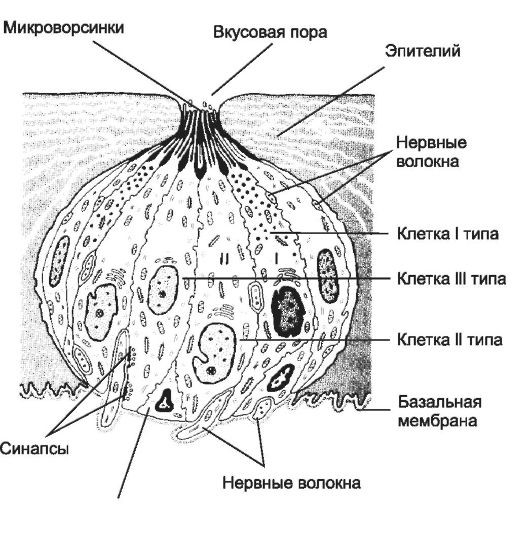
Рис. 8-51. Вкусовая почка состоит из удлинённых светлых клеток различных типов. Их апикальная поверхность открывается во вкусовую пору на поверхности эпителиального пласта. Клетки вкусовой почки образуют синапсы с периферическими отростками чувствительных нейронов. [17]
Хеморецепторные клетки. Хотя контакты с афферентными волокнами образуют все типы клеток, функцию хемовосприятия связывают с клетками типа III. В пресинаптической области вкусовых клеток гранулярные пузырьки содержат серотонин, нейромедиатор афферентного синапса. Хеморецепторные клетки воспринимают сладкое, кислое, горькое, солёное и вкус глутамата (юмами, umami). Регенерация. Во вкусовом рецепторе происходит постоянное обновление клеток. Из периферической области вкусовой почки клетки перемещаются в центральную её часть со скоростью 0,06 мкм/час. Средняя продолжительность жизни клеток вкусового рецепторного органа составляет 250±50 часов.
Слух и равновесие
ОРГАН СЛУХА
Наружное ухо включает ушную раковину, наружный слуховой проход и барабанную перепонку, передающую звуковые колебания на слуховые косточки среднего уха. Ушная раковина образована эластическим хрящом, покрытым тонкой кожей. Наружный слуховой проход выстлан кожей, содержащей волосяные фолликулы, типичные сальные железы и церуминозные железы - видоизменённые сальные железы, вырабатывающие ушную серу. Наружная поверхность барабанной перепонки покрыта кожей. Изнутри, со стороны барабанной полости (среднее ухо), барабанная перепонка (см. рис. 8-62) выстлана однослойным кубическим эпителием, расположенным на тонкой соединительнотканной пластинке.
Среднее ухо содержит слуховые косточки - молоточек, наковальня и стремя, которые передают колебания с барабанной перепонки на мембрану овального окна (см. рис. 8-62). Барабанная полость выстлана многослойным эпителием, который переходит в однослойный цилиндрический мерцательный у отверстия слуховой трубы. Кость медиальной стенки барабанной полости имеет два окна, овальное и круглое, которые отделяют барабанную полость от костного лабиринта внутреннего уха.
Внутреннее ухо образовано костным лабиринтом височной кости, который содержит повторяющий его рельеф перепончатый лабиринт (рис. 8-62, рис. 8-63). Костный лабиринт - система полукружных каналов и сообщающаяся с ними полость - преддверие (vestibulum). Перепончатый лабиринт - система тонкостенных соединительнотканных трубок и мешочков, расположенная внутри костного лабиринта. В костных ампулах перепончатые каналы расширяются. В преддверии перепончатый лабиринт образует два сообщающихся между собой мешочка: эллиптический мешочек (маточка, utriculus), в который открываются перепончатые полукружные каналы, и сферический мешочек (мешочек, sacculus). Перепончатые полукружные каналы и мешочки преддверия заполнены эндолимфой и сообщаются с улиткой, а также с расположенным в полости черепа эндолимфатическим мешком, где эндолимфа резорбируется. Эпителиальная выстилка эндолимфатического мешка содержит цилиндрические клетки с плотной цитоплазмой и ядрами неправильной формы, а также цилиндрические клетки со светлой цитоплазмой, высокими микроворсинками, многочисленными пиноцитозными пузырьками и вакуолями.
Улитка
Улитка (cochlea) - спирально закрученный костный канал. Улитка образует 2,5 завитка длиной около 35 мм. Базилярная (основная) и вестибулярная мембраны, расположенные внутри канала улитки, делят его полость на три части, это барабанная лестница (scala tympani), лестница преддверия (scala vestibuli) и перепончатый канал улитки (scala media, средняя лестница, улитковый ход) (рис. 8-56). Эндолимфа заполняет перепончатый канал улитки, а перилимфа - лестницу преддверия и барабанную лестницу. Лестница преддверия и барабанная лестница сообщаются у вершины улитки с помощью отверстия (helicotrema). В перепончатом канале улитки на базилярной мембране расположен рецепторный аппарат улитки - спиральный орган.
• Эндолимфа - вязкая жидкость, заполняет перепончатый канал улитки и соединяется через специальный канал (ductus reuniens) с эндолимфой вестибулярного аппарата. Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе и перилимфе; концентрация Na+ в эндолимфе в 10 раз меньше, чем в перилимфе.
• Перилимфа по химическому составу близка к плазме крови и спинномозговой жидкости и занимает промежуточное положение между ними по содержанию белка.
Спиральный орган
Спиральный орган (рис. 8-57) содержит несколько рядов волосковых клеток, контактирующих с покровной мембраной. Различают внутренние и наружные волосковые и поддерживающие клетки. Внутренние волосковые клетки - рецепторные, они образуют синаптические контакты с периферическими отростками чувствительных нейронов спирального ганглия.
Внутренние волосковые клетки образуют один ряд. Клетки с латеральной стороны прикрыты внутренними клетками-столбами, а другими своими поверхностями контактируют с фаланговыми клетками. В апикальной части расположены проходящие через кутикулу стереоцилии, состоящие из 20-50 неподвижных микроворсинок. Стереоцилии расположены полукругом (или в виде буквы V), открытым в сторону наружных структур спирального органа. Внутренние волосковые клетки возбуждаются в ответ на звуковой раздражитель и передают возбуждение афферентным волокнам слухового нерва. Смещение покровной мембраны вызывает деформацию стереоцилий, в мембране которых открываются механочувствительные ионные каналы и возникает деполяризация клеточной мембраны.
Наружные волосковые клетки высотой от 20 до 70 мкм расположены в 3-5 рядов, имеют цилиндрическую форму и стереоцилии. Базальная часть размещается в углублении, сформированном наружными фаланговыми клетками. Боковая поверхность клеток омывается жидкостью, заполняющей наружный туннель (cuniculus externus) и опорно-
 Рис. 8-56. Перепончатый канал и спиральный орган. Канал
улитки разделён на лестницу преддверия, барабанную лестницу и
перепончатый канал, в котором расположен спиральный орган. Перепончатый
канал отделён от барабанной лестницы базилярной мембраной. В её составе
проходят периферические отростки нейронов спирального ганглия,
образующие синаптические контакты с наружными и внутренними волосковыми
клетками. [17]
Рис. 8-56. Перепончатый канал и спиральный орган. Канал
улитки разделён на лестницу преддверия, барабанную лестницу и
перепончатый канал, в котором расположен спиральный орган. Перепончатый
канал отделён от барабанной лестницы базилярной мембраной. В её составе
проходят периферические отростки нейронов спирального ганглия,
образующие синаптические контакты с наружными и внутренними волосковыми
клетками. [17]
волосковое клеточное пространство (cuniculus intermedius). Наружные волосковые клетки образуют синапсы с эфферентными волокнами оливоулиткового тракта. Секреция ацетилхолина вызывает гиперполяризацию наружных волосковых клеток, что оказывает модулирующее влияние на высоту клеток и последующие реакции в цепочке передачи звукового раздражения.
 Рис. 8-57. Спиральный орган. Механочувствительные
волосковые клетки образуют несколько рядов: один ряд внутренних и 3-5
рядов наружных. Внутренние и наружные волосковые клетки разделены
туннелем. Его образуют крупные наружные и внутренние клетки-столбы. Со
стереоцилиями волосковых клеток соприкасается покровная мембрана. [17]
Рис. 8-57. Спиральный орган. Механочувствительные
волосковые клетки образуют несколько рядов: один ряд внутренних и 3-5
рядов наружных. Внутренние и наружные волосковые клетки разделены
туннелем. Его образуют крупные наружные и внутренние клетки-столбы. Со
стереоцилиями волосковых клеток соприкасается покровная мембрана. [17]
Поддерживающие клетки. Среди поддерживающих (опорных) клеток различают внутренние фаланговые клетки, наружные фаланговые клетки, наружные пограничные столбчатые клетки, наружные поддерживающие столбчатые клетки, наружные поддерживающие кубические клетки, наружные базальные железистые клетки. Фаланговые клетки вступают в контакт с волосковыми, располагаясь на базилярной мембране. Отростки наружных фаланговых клеток проходят параллельно наружным волосковым клеткам, не соприкасаясь с ними на значительном протяжении, но на уровне апикальной части волосковых клеток вступают с ними в контакт. Поддерживающие клетки связаны щелевыми контактами. Щелевые контакты участвуют в восстановлении уровня К+ в эндолимфе в ходе следовых реакций после возбуждения волосковых клеток.
Покровная мембрана (membrana tectoria) построена из белков (преимущественно коллагены), углеводов и гликозаминогликанов, которые формируют матрикс и волокна.
Базилярная мембрана представлена несколькими частями. Средняя часть - дугообразная зона (pars tecta), латеральная часть - гребенчатая зона (pars pectinata). Дугообразная зона частично встроена в костную спиральную пластинку. Поэтому она менее мобильна при перемещении жидкостей улитки в ответ на звуковое раздражение. Напротив, гребенчатая зона более подвижна и вибрирует в ответ на звуковые сигналы. Путь передачи слухового раздражения. Цепочка передачи звукового давления выглядит следующим образом: барабанная перепонка → молоточек → наковальня → стремя → мембрана окна преддверия → перилимфа → базилярная и покровная мембраны - мембрана окна улитки (рис. 8-62). При смещении стремени частицы перилимфы перемещаются по лестнице преддверия и затем через геликотрему по барабанной лестнице к круглому окну. Жидкость, сдвинутая смещением мембраны окна преддверия, создаёт избыточное давление в вестибулярном канале. Под действием этого давления базальный участок основной мембраны смещается в сторону барабанной лестницы. Колебательная реакция в виде волны распространяется от базальной части основной мембраны к геликотреме. Смещение покровной мембраны относительно волосковых клеток при действии звука вызывает их возбуждение. Смещение мембраны относительно сенсорного эпителия отклоняет стереоцилии волосковых клеток, что открывает механочувствительные каналы в клеточной мембране и приводит к деполяризации клеток. Возникающая электрическая реакция (микрофонный эффект) по своей форме повторяет форму звукового сигнала.
ОРГАН РАВНОВЕСИЯ
В каждом ампулярном расширении полукружного канала находятся кристы, или гребешки (рис. 8-62, 8-63). Чувствительные области в мешочках называются пятнами.
В состав эпителия пятен и крист входят чувствительные волосковые и поддерживающие клетки (рис. 8-64). Волосковые клетки образуют группы из нескольких сот единиц. Внутри каждой группы киноцилии ориентированы одинаково, но их ориентация между группами различна. Эпителий пятен покрыт студенистой отолитовой мембраной. Отолиты - кристаллы карбоната кальция. Эпителий крист окружён желатинообразным прозрачным куполом.
• Волосковые клетки (рис. 8-65) присутствуют в каждой ампуле полукружных каналов и в пятнах мешочков преддверия. Различают два типа волосковых клеток. Клетки обоих типов в апикальной части содержат 40-110 неподвижных волосков (стереоцилии) и одну ресничку (киноцилия), расположенную на периферии пучка стереоцилий. Самые длинные стереоцилии находятся вблизи киноцилии, а длина остальных уменьшается по мере удаления от киноцилии. Клетки типа I расположены в центре гребешков, имеют форму амфоры с закруг- лённым дном и размещены в бокалообразной полости афферентного нервного окончания. Эфферентные волокна образуют синаптические окончания на афферентных волокнах, связанных с клетками I типа.
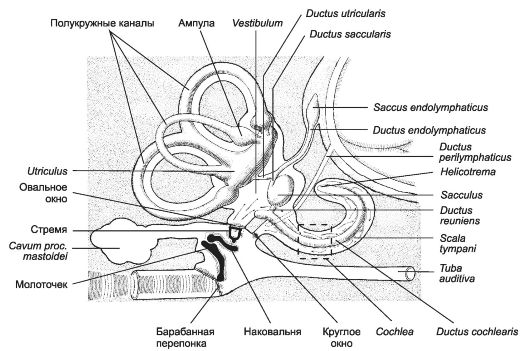 Рис. 8-62. Преддверно-улитковый орган. Звуковое
давление через барабанную перепонку по цепочке косточек среднего уха
(молоточек, наковальня, стремя) передаётся на мембрану окна преддверия и
далее к перилимфе сообщающихся через геликотрему лестниц к окну улитки.
Колебания перилимфы передаются покровной мембране. Её смещение вызывает
раздражение волосковых клеток. [17]
Рис. 8-62. Преддверно-улитковый орган. Звуковое
давление через барабанную перепонку по цепочке косточек среднего уха
(молоточек, наковальня, стремя) передаётся на мембрану окна преддверия и
далее к перилимфе сообщающихся через геликотрему лестниц к окну улитки.
Колебания перилимфы передаются покровной мембране. Её смещение вызывает
раздражение волосковых клеток. [17]
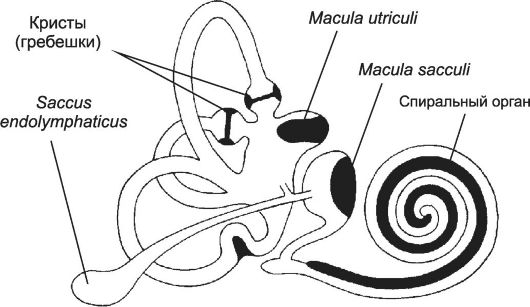 Рис. 8-63. Нейросенсорные области внутреннего уха выделены
чёрным. В нейросенсорных областях расположены механочувствительные
волосковые клетки. Это спиральный орган (орган слуха), кристы и пятна,
или макулы (орган равновесия). [17]
Рис. 8-63. Нейросенсорные области внутреннего уха выделены
чёрным. В нейросенсорных областях расположены механочувствительные
волосковые клетки. Это спиральный орган (орган слуха), кристы и пятна,
или макулы (орган равновесия). [17]
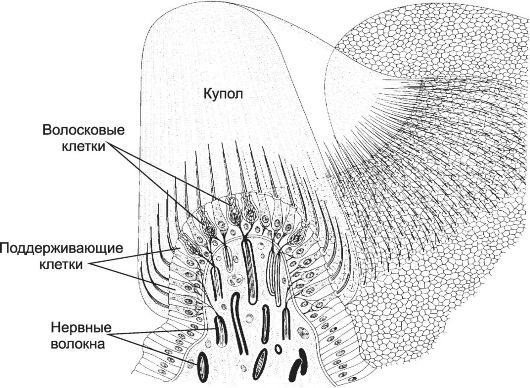 Рис. 8-64. Гребешок. Нейросенсорный
эпителий образован волосковыми и поддерживающими клетками. В центре
гребешка расположены волосковые клетки типа I, а по периферии - типа II.
Волосковые клетки образуют синапсы с нервными окончаниями. В куполе
гребешка отолиты отсутствуют. [17]
Рис. 8-64. Гребешок. Нейросенсорный
эпителий образован волосковыми и поддерживающими клетками. В центре
гребешка расположены волосковые клетки типа I, а по периферии - типа II.
Волосковые клетки образуют синапсы с нервными окончаниями. В куполе
гребешка отолиты отсутствуют. [17]
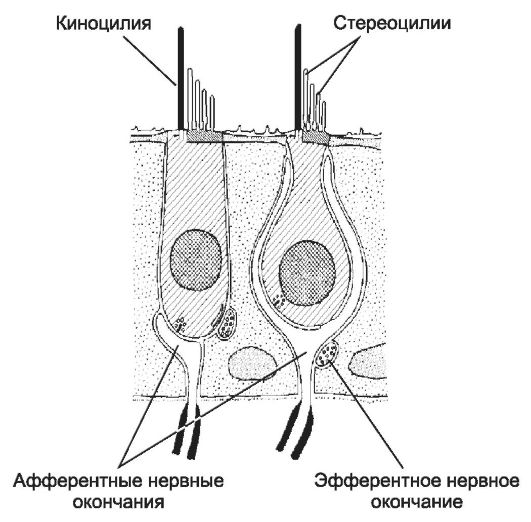 Рис. 8-65. Волосковые клетки. Справа - клетка типа I; слева - клетка типа II. [17]
Рис. 8-65. Волосковые клетки. Справа - клетка типа I; слева - клетка типа II. [17]
• Клетки типа II расположены по периферии гребешков, имеют вид цилиндров с округлым основанием. Нервные окончания на клетках типа II могут быть как афферентными (большинство), так и эфферентными. Дирекционная чувствительность Волосковые клетки чувствительны к направлению действия стимула. При направлении раздражающего воздействия от стереоцилий к киноцилии волосковая клетка возбуждается. При противоположном направлении стимула происходит угнетение ответа.
Вестибулярный нерв образован отростками биполярных нейронов в составе вестибулярного ганглия. Периферические отростки этих нейронов подходят к волосковым клеткам каждого полукружного канала, маточки и мешочка, а центральные направляются в вестибулярные ядра продолговатого мозга.