Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 15 ПОЛОВАЯ СИСТЕМА
Все клетки организма подразделяют на соматические и половые. Развитие мужских половых клеток - сперматозоидов (сперматогенез) совершается в мужских гонадах (яичках), развитие женских половых клеток - яйцеклеток (овогенез) происходит в женских гонадах (яичниках). Зрелые половые клетки - гаметы (сперматозоид и яйцеклетка) способны объединяться и давать начало новому организму.
Гаметогенез
В гонадах мужские и женские половые клетки проходят следующие стадии развития (гаметогенеза): размножения и роста, созревания, формирования. Часть этих процессов относится к мейозу, общая характеристика мейоза дана в главе 2 (см. также рис. 2-53). Стадии размножения и роста
• Сперматогенез. Первичные половые клетки мигрируют в зачатки гонад, делятся и дифференцируются в сперматогонии. До периода полового созревания сперматогонии остаются в состоянии покоя. Стадия размножения начинается с наступлением половой зрелости. По мере приближения половой зрелости в гипоталамусе усиливается синтез гонадолиберина, активирующего секрецию гонадотропных гормонов аденогипофиза. Под влиянием лютеинизирующего гормона интерстициальные эндокринные клетки яичка увеличивают синтез тестостерона, инициирующего сперматогенез. После ряда митотических делений сперматогонии дифференцируются в сперматоциты первого порядка, вступающие в стадию роста.
• Овогенез. В претерпевающих дифференцировку яичниках овогонии вступают в стадию размножения. К 7-му месяцу внутриутробного развития овогонии прекращают деление и дифференцируются в овоциты первого порядка. Завершив стадию роста, овоциты первого порядка в профазе первого деления мейоза приобретают оболочку из фолликулярных клеток (образуется примордиальный фолликул) и вступают в длительный период покоя, вплоть до наступления по-
ловой зрелости. Количество овоцитов первого порядка в 7 месяцев - до 10 миллионов, при рождении - около 2 миллионов. Стадия созревания
• Сперматогенез. В результате двух делений мейоза образуются четыре сперматиды, имеющие по 22 аутосомы и одной X- или Y-хромосоме.
• Овогенез. С наступлением половой зрелости и установлением овариально-менструального цикла на пике лютеинизирующего гормона завершается первое деление мейоза. Сигнал для завершения второго мейотического деления - оплодотворение, овоцит второго порядка делится с образованием зрелой яйцеклетки и второго полярного (направительного) тельца. Зрелая яйцеклетка имеет 22 аутосомы и одну X-хромосому (рис. 15-8).
Стадия формирования гамет заключается в морфологической дифференцировке сперматид и образовании сперматозоидов. Эта стадия отсутствует в овогенезе.
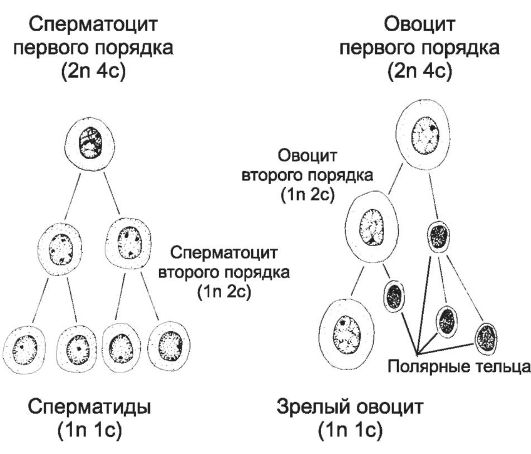 Рис. 15-8. Мейоз включает
два последовательных деления. Перед первым делением генетический
материал удваивается с образованием конъюгированных хромосом (2n2c).
После первого деления мейоза в дочерних клетках уменьшаются количество
хромосом и содержание ДНК; остаётся по 23 конъюгированных (удвоенных)
хромосомы с диплоидным содержанием ДНК (1n2c). После второго деления
мейоза дочерние клетки получают по 23 хромосомы с гаплоидным содержанием
ДНК (1n1c) - 22 аутосомы и одну половую хромосому. n - число хромосом, с
- количество ДНК. [17]
Рис. 15-8. Мейоз включает
два последовательных деления. Перед первым делением генетический
материал удваивается с образованием конъюгированных хромосом (2n2c).
После первого деления мейоза в дочерних клетках уменьшаются количество
хромосом и содержание ДНК; остаётся по 23 конъюгированных (удвоенных)
хромосомы с диплоидным содержанием ДНК (1n2c). После второго деления
мейоза дочерние клетки получают по 23 хромосомы с гаплоидным содержанием
ДНК (1n1c) - 22 аутосомы и одну половую хромосому. n - число хромосом, с
- количество ДНК. [17]
Мужская половая система
Мужская половая система состоит из половых желёз (яички), комплекса половых протоков, добавочных желёз и полового члена.
ЯИЧКО
Яички (testis) располагаются вне полости тела (в мошонке). Это обстоятельство важно для нормального течения сперматогенеза, происходящего при температуре 34 °C и ниже. Снаружи яичко покрыто соединительнотканной белочной оболочкой (рис. 15-10). Внутренний её слой богат кровеносными сосудами - сосудистая оболочка. Утолщение белочной оболочки, вдающееся с одной стороны в паренхиму яичка, называется средостением (mediastinum testis). От средостения внутрь яичка отходят перегородки, разделяющие его на дольки конической формы. Каждая долька содержит от одного до четырёх извитых семенных канальцев, выстланных сперматогенным эпителием. В рыхлой соединительной ткани между семенными канальцами расположены интерстициальные эндокринные клетки. По мере приближения к средостению семенные канальцы продолжаются в прямые канальцы. Их стенка образована кубическим эпителием и базальной мембраной, окружённой тонкой соединительнотканной оболочкой. Прямые канальцы впадают в сеть яичка - систему анастомозирующих тонкостенных трубочек, продолжающихся в выносящие канальцы придатка. Эпителий, выстилающий сеть яичка, представлен кубическими клетками. Некоторые клетки на апикальной поверхности имеют по одной ресничке, способствующей продвижению сперматозоидов. Под
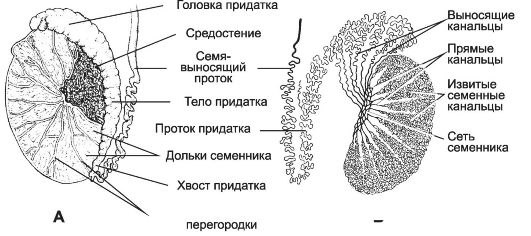
Рис. 15-10. Яичко с придатком. А - соединительнотканная строма яичка и придатка; Б - паренхима яичка и придатка. [17]
базальной мембраной эпителия располагается рыхлая соединительная ткань, окружающая трубочки сети яичка снаружи. Генеративная функция яичка (сперматогенез) осуществляется в извитых семенных канальцах, а эндокринная функция (синтез мужских половых гормонов) - в интерстициальных эндокринных клетках.
Интерстициальные эндокринные клетки
Эндокринные клетки яичка, вырабатывающие андрогены, расположены в рыхлой волокнистой соединительной ткани между семенными канальцами. Это типичные эндокринные клетки, синтезирующие гормоны стероидной природы (см. рис. 9-25). Для этих клеток характерны развитая гладкая эндоплазматическая сеть, многочисленные митохондрии, жировые включения.
Андрогены - мужские половые гормоны. Интерстициальные эндокринные клетки яичка синтезируют 95% циркулирующего в крови тестостерона, дигидротестостерон и предшественник тестостерона андростендион. В эмбриогенезе андрогены контролирует развитие плода по мужскому типу. В период полового созревания они обеспечивают становление физических признаков мужского пола (оволосение по мужскому типу, огрубение голоса, рост мышечной массы), поддерживают рост хряща, стимулируют закрытие эпифизарных пластинок. С наступлением половой зрелости тестостерон необходим для поддержания сперматогенеза. Тестостерон стимулирует рост и секреторную активность предстательной железы, семенных пузырьков и бульбоуретральных желёз. Стимулятор синтеза андрогенов интерстициальными эндокринными клетками яичка - лютеинизирующий гормон (лютропин).
Извитые семенные канальцы
Извитые семенные канальцы выстланы сперматогенным эпителием, содержащим клетки двух типов - гаметы с их предшественниками на различных стадиях дифференцировки и поддерживающие эпителиальные клетки - сустентоциты (рис. 15-12). Снаружи канальцы окружены тонкой соединительнотканной оболочкой. Сустентоциты - поддерживающие эпителиальные клетки. Их широкое основание находится на базальной мембране, а суженная складчатая апикальная часть достигает просвета канальца. При помощи плотных контактов сустентоциты делят сперматогенный эпителий на базальное и адлюминальное пространства. В базальном пространстве находятся только сперматогонии. В адлюминальном пространстве располагаются сперматоциты первого и второго порядков, сперматиды и сперматозоиды. Между сустентоцитами и гаметами устанавливаются адгезионные и щелевые контакты. Сустентоциты оказывают физическую поддержку развивающимся гаметам, обеспечивают их питательными веществами (трофическая функция), поглощают продукты метаболизма, фагоцитируют остатки цитоплазмы формирующихся сперматозоидов и дегенерирующие половые клетки, секретируют жидкость для
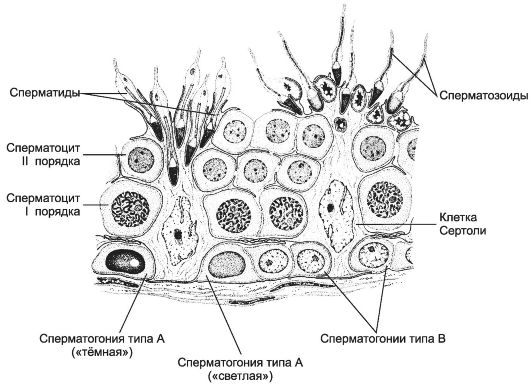 Рис. 15-12. Сперматогенный эпителий. На
базальной мембране располагаются поддерживающие эпителиальные клетки, а
также сперматогонии типов А и В. Ближе к просвету канальца лежат
сперматоциты первого и второго порядков, над которыми находятся
сперматиды на различных этапах развития и сперматозоиды. [17]
Рис. 15-12. Сперматогенный эпителий. На
базальной мембране располагаются поддерживающие эпителиальные клетки, а
также сперматогонии типов А и В. Ближе к просвету канальца лежат
сперматоциты первого и второго порядков, над которыми находятся
сперматиды на различных этапах развития и сперматозоиды. [17]
транспорта сперматозоидов в семенных канальцах. В кровь сустентоциты секретируют эстрогены, ингибин, мюллеров ингибирующий фактор MIF (в плодном периоде). В адлюминальном пространстве поддерживающие эпителиальные клетки накапливают андроген-связывающий белок, фактор стволовых клеток, трансферрин, активаторы плазминогена. Фолликулостимулирующий гормон (фоллитропин) - главный стимулирующий фактор сустентоцитов. Гематотестикулярный барьер. Поддерживающие клетки образуют плотные контакты, формирующие гематотестикулярный барьер. Благодаря такому барьеру, в адлюминальном пространстве создаётся специфическая гормональная среда с высоким уровнем тестостерона. Барьер изолирует созревающие половые клетки от токсических веществ и препятствует развитию аутоиммунного ответа против поверхностных антигенов, экспрессирующихся на мембране созревающих сперматозоидов.
Сперматогенез
Сперматогенез (путь от сперматогонии до сперматозоида) в извитых семенных канальцах длится 65 дней, но окончательная дифференцировка сперматозоидов происходит в протоке придатка яичка в течение следующих двух недель. Только в области хвоста придатка сперматозоиды становятся зрелыми половыми клетками и приобретают способность к самостоятельному передвижению и оплодотворению яйцеклетки. Сперматогенез подразделяют на стадии размножения, роста, созревания и формирования. На стадии размножения выделяют сперматогонии типов А и В (см. рис. 15-12).
Сперматогонии типа А. Среди сперматогоний типа А по степени конденсации хроматина различают тёмные и светлые клетки. Тёмные сперматогонии считаются резервными стволовыми клетками, редко вступающими в митоз. Светлые сперматогонии - полустволовые клетки, находящиеся в непрерывно следующих друг за другом клеточных циклах (интерфаза сменяется митозом). В результате деления светлой клетки типа А образуются либо две клетки типа В (симметричный митоз), либо одна клетка типа В и одна светлая клетка типа А (асимметричный митоз).
Сперматогонии типа В имеют круглое ядро и конденсированный хроматин. Они также вступают в митоз, но при этом остаются связанными друг с другом при помощи цитоплазматических мостиков. После ряда митотических делений сперматогонии типа В дифференцируются в сперматоциты первого порядка, которые из базального пространства перемещаются в адлюминальное и вступают в стадию роста. В стадии роста объём сперматоцитов первого порядка увеличивается в 4 и более раз. Клеточные ассоциации. На стадиях размножения, роста и созревания сперматогенные клетки входят в состав клеточных ассоциаций (рис. 15-14). Светлая сперматогония типа А формирует клон сперматогенных клеток (синцитий), в котором клетки остаются связанными цитоплазматическими мостиками до стадии формирования. Клеточная ассоциация в своем развитии от сперматогоний до сперматозоидов проходит шесть стадий (I-VI), для каждой из которых существует характерный тип сочетания сперматогенных клеток. В ходе сперматогенеза клеточные ассоциации находятся на различных стадиях развития, что обусловливает мозаичное распределение клеточных ассоциаций в составе сперматогенного эпителия.
Созревание. За стадией роста наступает стадия созревания, которая состоит из следующих друг за другом двух делений мейоза. В результате первого деления из одного сперматоцита первого порядка образуется два сперматоцита второго порядка, а после второго деления - четыре сперматиды (см. рис. 15-8 и 15-12). Сперматиды располагаются вбли-
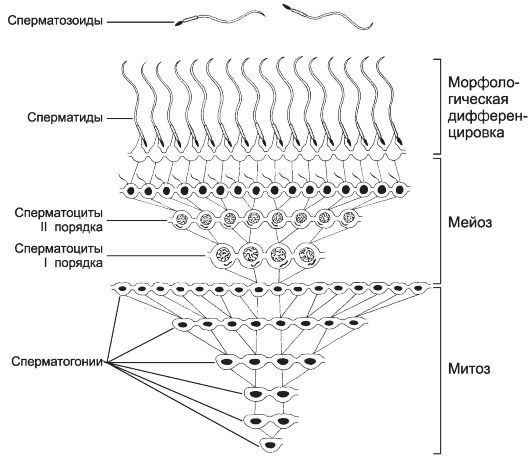 Рис. 15-14. Клон сперматогенных клеток. На
стадиях размножения, роста и созревания сперматогенные клетки связаны
цитоплазматическими мостиками. Только на стадии формирования гамет
цитоплазматические мостики разрываются, и сперматозоиды оказываются
свободными. [17]
Рис. 15-14. Клон сперматогенных клеток. На
стадиях размножения, роста и созревания сперматогенные клетки связаны
цитоплазматическими мостиками. Только на стадии формирования гамет
цитоплазматические мостики разрываются, и сперматозоиды оказываются
свободными. [17]
зи просвета канальца. Каждая сперматида вчетверо мельче сперматоцита первого порядка и имеет ядро с гаплоидным набором хромосом. Стадия формирования. Спермиогенез - постмейотическая стадия морфологической дифференцировки сперматиды в сперматозоид. В составе комплекса Гольджи (на одном из полюсов ядра) появляется акросомный пузырёк с акросомной гранулой, одновременно центриоли мигрируют на противоположную сторону ядра и инициируют сборку аксонемы. Акросомная гранула заполняет акросомный пузырёк и образуется акросомная шапочка. Хроматин конденсируется, ядро вытягивается и приобретает окончательную форму. Цитоплазматические микротрубочки, растущие от центриолей, образуют цилиндрическую
манжетку, способствующую элонгации сперматиды. Митохондрии располагаются спиралеобразно вокруг аксонемы. Сперматида переворачивается хвостом в просвет канальца. Между клетками разрываются цитоплазматические мостики, а сформировавшиеся сперматозоиды оказываются свободными.
Гормональная регуляция сперматогенеза
Гипоталамо-гипофизарная система при помощи гонадолиберина активирует синтез и секрецию гонадотропных гормонов гипофиза, влияющих на активность поддерживающих эпителиальных и интерстициальных эндокринных клеток. В свою очередь, вырабатываемые в яичке гормоны корректируют эндокринную деятельность гипоталамогипофизарной системы. Гонадолиберин поступает в кровь из аксонов нейросекреторных клеток в пульсирующем режиме с пиковыми интервалами около двух часов. Гонадотропные гормоны, как и гонадолиберин, высвобождаются в кровь также в пульсирующем режиме, что особенно характерно для лютропина, пики концентрации которого в крови мужчин наблюдаются с интервалами 90-120 мин. Мишени гонадотропных гормонов - яички. Сустентоциты имеют рецепторы фоллитропина, а интерстициальные эндокринные клетки - лютропина. Тестостерон поступает в сустентоциты, в цитозоле связывается с андроген-связывающим белком и далее секретируется в адлюминальное пространство для поддержания сперматогенеза.
МУЖСКИЕ ПОЛОВЫЕ ПРОТОКИ
К половым протокам мужской половой системы относятся выносящие канальцы придатка яичка, проток придатка, семявыносящий проток, семявыбрасывающий проток.
Придаток яичка
В придатке яичка (epididymis) различают головку, тело и хвост. Головка придатка представлена 10-12 выносящими канальцами (ductuli efferentes). Тело и хвост придатка образованы протоком придатка (ductus epididymis), в который открываются ductuli efferentes. Выносящие канальцы придатка прободают белочную оболочку (tunica albuginea) и соединяют сеть яичка (rete testis) с протоком придатка яичка (см. рис. 15-10). Эти канальцы выстланы эпителием, клетки которого имеют разную высоту (гирляндный эпителий). Высокие цилиндрические клетки снабжены ресничками, способствующими перемещению сперматозоидов по канальцам. Низкие кубические клетки имеют складчатую поверхность с микроворсинками. В апикальной части клеток присутствуют многочисленные пиноцитозные пузырьки и ли-
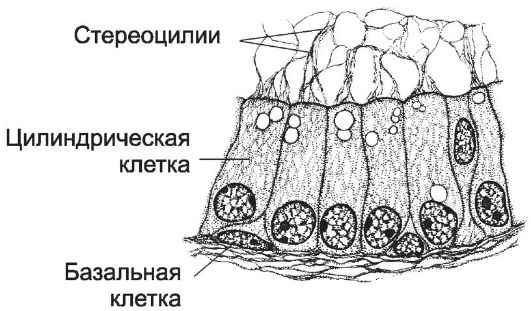 Рис. 15-16. Пламенный эпителий
Рис. 15-16. Пламенный эпителий
ductus epididymis. [17]
зосомы. Функция этих клеток заключается в реабсорбции жидкости, образующейся в извитых канальцах яичка. Снаружи эпителиальной выстилки располагается собственный слой с циркулярно ориентированными ГМК, способствующими продвижению сперматозоидов по выносящим канальцам.
Проток тела придатка - одиночный и сильно извитой каналец длиной до 6 м. Он выстлан многорядным цилиндрическим эпителием (рис. 15-16). В эпителии различают два типа клеток: базальные вставочные и высокие цилиндрические. Цилиндрические клетки снабжены стереоцилиями, склеенными в виде конуса (пламенный эпителий). Между основаниями цилиндрических клеток расположены мелкие вставочные клетки, являющиеся предшественниками цилиндрических клеток. Под эпителием располагается собственный слой, окружённый циркулярно ориентированными ГМК. Мышечный слой протока придатка по мере приближения к семявыносящему протоку становится более выраженным и представлен тремя слоями: внутренним и наружным продольными и средним циркулярным. Сокращения ГМК способствуют продвижению сперматозоидов в семявыносящий проток.
Семявыносящий проток
Из придатка сперматозоиды попадают в семявыносящий проток (ductus deferens). Толстая стенка ductus deferens состоит из трёх оболочек: слизистой, мышечной и адвентициальной. Слизистая оболочка представлена собственным слоем и многорядным эпителием, который в проксимальной части семявыносящего протока по строению сходен с эпителием протока придатка. Мышечная оболочка образована тремя слоями ГМК: внутренним и наружным продольными и средним циркулярным. Мощная мышечная оболочка ductus deferens способствует выбрасыванию сперматозоидов во время эякуляции. Адвентициальная оболочка состоит из волокнистой соединительной ткани с кровеносными сосудами, нервами и группами ГМК. В составе семенного
канатика ductus deferens проходит по паховому каналу в брюшную полость. Вблизи мочевого пузыря семявыносящий проток заканчивается ампулярным расширением. Сразу за ампулой после слияния с семенными пузырьками ductus deferens продолжается в семявыбрасывающий проток.
Семявыбрасывающий проток
Ductus ejaculatorius проникает в предстательную железу и открывается в простатическую часть мочеиспускательного канала (в семенной холмик на гребне мочеиспускательного канала). Эпителий сходен с эпителием семявыносящего протока. Мышечная оболочка отсутствует. Эякуляцию обеспечивают ГМК хвоста придатка яичка и семявыносящего протока.
ДОБАВОЧНЫЕ ЖЕЛЕЗЫ
Семенные пузырьки, предстательную и бульбоуретральные железы принято называть добавочными железами мужской половой системы. Вклад секретов предстательной железы и семенных пузырьков в объ- ём семенной жидкости составляет 95%, на сперматозоиды приходится лишь 5% общего объёма эякулята.
Семенные пузырьки
Семенные пузырьки - две сильно извитые трубки длиной до 15 см, открывающиеся в семявыбрасывающий проток (сразу за ампулой семявыносящего протока). Стенка трубки состоит из трёх оболочек: слизистой, мышечной и соединительнотканной. Слизистая оболочка имеет выраженную складчатость и выстлана однослойным многорядным цилиндрическим эпителием (секреторные и камбиальные базальные клетки).
• Секреторные клетки в апикальной части имеют микроворсинки и секреторные гранулы. Клетки содержат хорошо развитые гранулярную эндоплазматическую сеть и комплекс Гольджи, в цитоплазме присутствуют жировые включения и липохромный пигмент.
• Секрет железы. Семенные пузырьки секретируют вязкий желтоватого цвета (присутствие липохромного пигмента) секрет, поступающий в семявыбрасывающий проток во время эякуляции. Объём семенной жидкости составляет до 70% эякулята.
Мышечная оболочка образована внутренним циркулярным и наружным продольным слоями ГМК. Сокращение ГМК при эякуляции способствует выбрасыванию секрета железы в семявыбрасывающий проток.
Предстательная железа
Дифференцировка и рост предстательной железы находятся под контролем андрогенов. 5а-Редуктаза в эпителиальных клетках простаты катализирует превращение тестостерона в дигидротестостерон, ответственный за пролиферацию и дифференцировку клеток железы. Железа снаружи покрыта соединительнотканной капсулой, содержащей ГМК. Соединительнотканные перегородки с хорошо развитой гладкой мускулатурой разделяют предстательную железу на дольки. Железа состоит из 30-50 разветвлённых трубчато-альвеолярных желёз (рис. 15-18), расположенных в трёх зонах: периферической, центральной и периуретральной. Поскольку каждая отдельная железа простаты - разветвлённая трубчато-альвеолярная, то на срезе концевые отделы имеют самую различную форму: округлую, овальную, вытянутую, разветвлённую. Нередко у мужчин пожилого возраста в просвете секреторных отделов можно обнаружить конкременты - сферической формы тела диаметром до 250 мм, состоящие из белка и солей кальция. Каждая железа имеет собственный выводной проток, открывающийся в просвет уретры. Выводные протоки выстланы многорядным призматическим эпителием, который в дистальных отделах становится переходным.
Секреторные отделы выстланы однослойным кубическим или цилиндрическим эпителием (в зависимости от стадии секреции). В
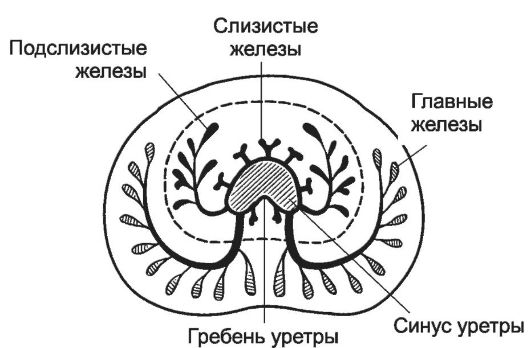 Рис. 15-18. Предстательная железа окружает
проксимальный отдел уретры. В паренхиме вокруг уретры находятся мелкие
слизистые железы (периуретральная зона). Под капсулой расположены
крупные главные железы (периферическая зона). Между слизистыми и
главными находятся средние по размерам подслизистые железы (центральная
зона). Выводной проток каждой железы открывается в просвет уретры. [17]
Рис. 15-18. Предстательная железа окружает
проксимальный отдел уретры. В паренхиме вокруг уретры находятся мелкие
слизистые железы (периуретральная зона). Под капсулой расположены
крупные главные железы (периферическая зона). Между слизистыми и
главными находятся средние по размерам подслизистые железы (центральная
зона). Выводной проток каждой железы открывается в просвет уретры. [17]
составе эпителия присутствуют базальные, железистые и эндокринные клетки.
Железистые клетки имеют выраженный белоксинтезирующий аппарат, в апикальной части клеток присутствуют мелкие светлые секреторные гранулы. Клетки продуцируют секрет, содержащий лимонную кислоту, простато-специфическую кислую фосфатазу, фибринолизин, простато-специфический антиген.
Базальные клетки обеспечивают регенерацию железистого эпителия. Секрет предстательной железы попадает в мочеиспускательный канал за счёт сокращения ГМК, он принимает участие в разжижении семени и способствует его прохождению по мочеиспускательному каналу при эякуляции. Присутствующие в секрете предстательной железы протеолитические ферменты препятствуют склеиванию сперматозоидов.
Бульбоуретральные железы
Бульбоуретральные железы (glandula bulbourethralis) - две трубчатоальвеолярные железы небольшой величины, расположенные между скелетными мышцами мочеполовой диафрагмы и открывающиеся в губчатую часть уретры. Секреторный отдел железы выстлан эпителиальными клетками кубической и цилиндрической формы, которые продуцируют слизистый секрет, содержащий галактозу, галактозамин, галактуроновую и сиаловую кислоты. В соединительнотканной строме, разделяющей железу на многочисленные дольки, встречаются как ГМК, так и поперечно-полосатые мышечные волокна. Вязкий слизистый секрет, выделяемый в период полового возбуждения, служит для смазки уретры перед эякуляцией.
ПОЛОВОЙ ЧЛЕН
Penis формируют три кавернозных тела. Парные, цилиндрической формы corpora cavernosa (пещеристые тела) располагаются на дорсальной стороне полового члена. На вентральной поверхности по средней линии находится corpus cavernosum urethrae (губчатое тело мочеиспускательного канала), имеющее расширенное основание (луковицу) и формирующее на дистальном конце головку полового члена. Кавернозные тела образованы анастомозирующей сетью перегородок (трабекул) из соединительной ткани и ГМК. В свободные пространства между покрытыми эндотелием перегородками открываются капилляры. Головку полового члена формирует плотная волокнистая соединительная ткань, содержащая сеть крупных извитых вен. Снаружи кавернозные тела окружены плотной соединительнотканной белочной оболочкой (tunica albuginea), состоящей из двух слоёв коллагеновых волокон - внутреннего циркулярного и наружного продольного.
Белочная оболочка отсутствует на головке. Головку полового члена покрывает тонкая кожа, в которой присутствуют сальные железы (gl. sebacea preputiales). Все три кавернозных тела объединены фасцией полового члена. Циркулярную складку кожи, покрывающей головку, называют крайней плотью.
Кровоснабжение. В расслабленном состоянии полового члена крупные артерии, проходящие в перегородках кавернозных тел, спирально закручены (aa. helicinae). Эти артерии имеют толстую мышечную оболочку. Продольные утолщения внутренней оболочки, состоящие из пучков ГМК и коллагеновых волокон, выбухают в просвет сосуда и служат клапанами, закрывающими просвет сосуда. Многие из этих артерий прямо открываются в межтрабекулярные пространства. В стенке вен полового члена (кроме циркулярного слоя ГМК в средней оболочке) также присутствуют продольные слои ГМК во внутренней и наружной оболочках. Во время эрекции происходит расслабление ГМК перегородок и спиральных артерий. Артерии несколько распрямляются и кровь с меньшем сопротивлением поступает в свободные пространства кавернозных тел. Одновременное сокращение ГМК глубоких вен вызывает уменьшение просвета сосудов и препятствует оттоку крови из переполненных кровью межтрабекулярных пространств. Детумесценция (расслабление полового члена после эрекции) - следствие меньшего притока крови в результате сокращения ГМК перегородок и сужения спиральных артерий и облегчения оттока крови из кавернозных тел.
Иннервация. Кожа и сосудистое сплетение головки полового члена, фиброзные оболочки кавернозных тел, слизистая и мышечная оболочки перепончатой и простатической частей уретры считаются мощными рефлексогенными зонами, насыщенными разнообразными концевыми чувствительными приборами. Каждая из этих зон играет значительную роль при половом акте, являясь источником нервных импульсов, лежащих в основе безусловнорефлекторных реакций (эрекция, эякуляция, оргазм). Среди чувствительных приборов встречаются как свободные нервные окончания, так и инкапсулированные - пластинчатые, тактильные и генитальные тельца.
МУЖСКАЯ УРЕТРА
Мочеиспускательный канал (трубка длиной около 12 см) проходит через простату (pars prostatica), прободает фасцию мочеполовой диафрагмы (pars membranacea), проникает в губчатое тело мочеиспускательного канала (pars spongiosa) и открывается наружным отверстием уретры на верхушке головки полового члена.
Простатическая часть. Просвет уретры (вследствие выпячивания на задней стенки гребня уретры) имеет V-образную форму (см. рис. 15-18). Вдоль гребня остаются два синуса (sinus urethralis), в которые открываются протоки главных и подслизистых желёз простаты. По обе стороны с боков гребня в семенные холмики открываются семяизвергающие каналы. В области внутреннего отверстия мочеиспускательного канала ГМК наружного циркулярного слоя участвуют в образовании сфинктера мочевого пузыря. Простатическую часть уретры выстилает переходный эпителий, сходный с эпителием мочевого пузыря.
Перепончатая часть. Скелетная мускулатура мочеполовой диафрагмы формирует произвольный сфинктер уретры (наружный сфинктер мочевого пузыря). Переходный эпителий замещается высоким многослойным цилиндрическим. Слизистая и мышечная оболочки простатической и перепончатой частей уретры имеют мощную рецепторную иннервацию.
Губчатая часть. Проходя через луковицу губчатого тела, уретра расширяется, образуя луковицу мочеиспускательного канала. Расширение уретры в головке полового члена называют ладьевидной ямкой. После этого места эпителий становится многослойным плоским, который далее переходит в многослойный плоский ороговевающий, покрывающий головку полового члена. Устья бульбоуретральных желёз открываются в губчатую часть уретры сразу после её прохождения через мочеполовую диафрагму.
Женская половая система
Женская половая система складывается из парных яичников (ovarium) и маточных труб (tuba uterina), матки (uterus), влагалища (vagina), наружных половых органов (organa genitalia feminina externa), а также молочных желёз (mamma).
Все органы половой системы женщины детородного возраста вне беременности подвергаются циклическим изменениям. Комплекс таких изменений - овариально-менструальный цикл. В среднем цикл продолжается 28 дней.
• Овариальный цикл - продолжение фазы созревания овогенеза, овуляция, формирование жёлтого тела. Овариальный цикл регулируют гипофизарные гонадотропины - фолликулостимулирующий и лютеинизирующий гормоны.
• Менструальный цикл - характерные изменения слизистой оболочки матки, имеющие целью возможность имплантации и без наступления последней заканчивающиеся отторжением функционального слоя эндометрия (менструация). Все фазы менструального цикла контролируют гормоны яичника - эстрогены и прогестерон.
В циклических изменениях органов женской половой системы принимают участие женские половые гормоны - эстрогены и прогестерон.
• Эстрогены (эстрадиол, эстрон и эстриол) синтезируются фолликулярными клетками яичника, клетками жёлтого тела, плаценты и печени (эстриол).
• Прогестерон синтезируют клетки жёлтого тела яичника в лютеиновую стадию овариально-менструального цикла, а также клетки хориона при наступлении беременности.
ЯИЧНИК
Поверхность яичника покрыта одним слоем кубических эпителиальных клеток (mesovarium), находящихся на толстой соединительноткан-
ной пластинке - белочной оболочке яичника (t. albuginea). Паренхима яичника (рис. 15-20) состоит из коркового и мозгового вещества. Мозговое вещество органа образовано рыхлой соединительной тканью, богатой эластическими волокнами, кровеносными, лимфатическими и нервными сплетениями. Корковое вещество содержит: примордиальные фолликулы, растущие фолликулы (первичные, вторичные и третичные), зрелые фолликулы, жёлтые тела (corpus luteum), белые тела (corpus albicans) и атретические фолликулы (folliculus atreticus).
Овариальный цикл
Первая половина овариального цикла - фолликулярная (под влиянием фоллитропина происходит развитие части примордиальных фолликулов), вторая половина - лютеиновая (под влиянием гипофизарного лютропина из клеток овулировавшего фолликула формируется эндокринная железа - жёлтое тело). Овуляция приходится примерно на середину цикла. Развитие фолликулов (рис. 15-21) происходит по схеме: примордиальный -> первичный -> вторичный -> третичный -> зрелый (преовуляторный).
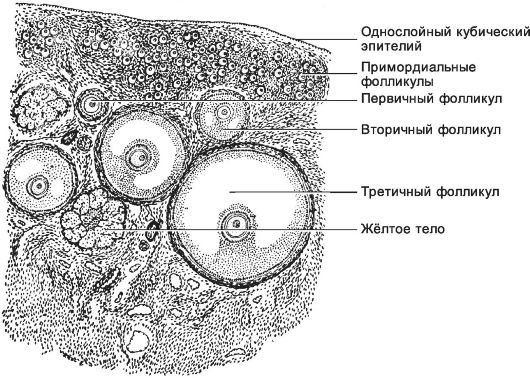 Рис. 15-20. Яичник. В
корковом веществе яичника располагаются фолликулы и их производные
различной степени зрелости. Мозговое вещество представлено рыхлой
соединительной тканью, содержащей сосудистые и нервные сплетения. [17]
Рис. 15-20. Яичник. В
корковом веществе яичника располагаются фолликулы и их производные
различной степени зрелости. Мозговое вещество представлено рыхлой
соединительной тканью, содержащей сосудистые и нервные сплетения. [17]
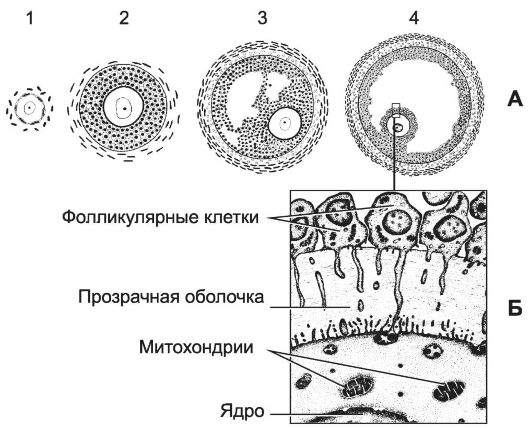 Рис. 15-21. Дифференцировка фолликула. А - стадии развития: 1 -
плоские фолликулярные клетки примордиального фолликула становятся
кубическими, а фолликул дифференцируется в первичный; овоцит покрыт
одним слоем фолликулярных клеток; 2 - вторичный фолликул. Под
влиянием фоллитропина увеличивается количество фолликулярных клеток,
вокруг фолликула формируется оболочка (theca); 3 - третичный фолликул. Между фолликулярными клетками образуются полости, заполненные фолликулярной жидкостью; 4 -
зрелый фолликул (преовуляторный). Сливаются отдельные пространства
между фолликулярными клетками и образуется единая полость, заполненная
фолликулярной жидкостью, яйцеклетка оттесняется к стенке фолликула. Б -
связь фолликулярных клеток и яйцеклетки. Базальная мембрана отделяет
фолликулярные клетки от окружающих тканей. Между фолликулярными клетками
и яйцеклеткой находится прозрачная оболочка. Фолликулярные клетки при
помощи цитоплазматических отростков проникают через прозрачную оболочку и
достигают плазматической мембраны яйцеклетки. [17]
Рис. 15-21. Дифференцировка фолликула. А - стадии развития: 1 -
плоские фолликулярные клетки примордиального фолликула становятся
кубическими, а фолликул дифференцируется в первичный; овоцит покрыт
одним слоем фолликулярных клеток; 2 - вторичный фолликул. Под
влиянием фоллитропина увеличивается количество фолликулярных клеток,
вокруг фолликула формируется оболочка (theca); 3 - третичный фолликул. Между фолликулярными клетками образуются полости, заполненные фолликулярной жидкостью; 4 -
зрелый фолликул (преовуляторный). Сливаются отдельные пространства
между фолликулярными клетками и образуется единая полость, заполненная
фолликулярной жидкостью, яйцеклетка оттесняется к стенке фолликула. Б -
связь фолликулярных клеток и яйцеклетки. Базальная мембрана отделяет
фолликулярные клетки от окружающих тканей. Между фолликулярными клетками
и яйцеклеткой находится прозрачная оболочка. Фолликулярные клетки при
помощи цитоплазматических отростков проникают через прозрачную оболочку и
достигают плазматической мембраны яйцеклетки. [17]
Примордиальные фолликулы располагаются непосредственно под белочной оболочкой яичника в виде компактных групп. Каждый примордиальный фолликул состоит из крупной округлой клетки - овоцита первого порядка, покрытого одним слоем плоских фолликулярных клеток (гранулёза, клетки гранулёзы). Фолликул окружён базальной мембраной. Многочисленные отростки фолликулярных клеток достигают плазмолеммы овоцита и образуют с ним щелевые и адгезионные контакты. При рождении девочки в яичниках содержится около двух миллионов примордиальных фолликулов. С наступлением половой зрелости таких фолликулов остаётся не более 400 тыс. 98% примордиальных фолликулов в течение репродуктивного периода погибает,
около 2% достигает стадии первичного и вторичного фолликулов, не более 400 развивается в преовуляторный фолликул и подвергается овуляции (каждый цикл - 1, много реже - 2).
Каждый овариальный цикл от 3 до 30 примордиальных фолликулов под влиянием фоллитропина вступает в фазу роста и формирует первичные фолликулы. Все начавшие развитие, но не достигшие стадии овуляции фолликулы подвергаются атрезии. Атретические фолликулы содержат погибший овоцит, полость отсутствует, характерна сморщенная прозрачная оболочка, окружённая дегенерирующими фолликулярными клетками.
Первичные фолликулы. В первичных фолликулах фолликулярные клетки из плоских превращаются в кубические. Овоцит увеличивается в размерах.
Вторичные фолликулы. Фолликулярные клетки вторичных фолликулов начинают пролиферировать, в результате чего вокруг овоцита первого порядка образуется несколько слоёв кубических клеток. Между овоцитом и окружающими его фолликулярными клетками появляется толстая прозрачная оболочка (zona pellucida). Растущий фолликул приобретает наружную оболочку из элементов стромы яичника - theca. В составе этой оболочки различают внутренний клеточный слой (theca interna), содержащий синтезирующие андрогены интерстициальные клетки и богатую капиллярную сеть, и наружный фиброзный слой (theca externa), образованный соединительной тканью. Theca interna часто именуют просто тека. Интерстициальные клетки - клетки паренхимы яичника, вероятно, имеют одинаковый генез с клетками theca interna. Они также синтезируют и секретируют андрогены. Рост фолликула стимулируют фоллитропин, эстрогены, норадреналин, инсулиноподобный фактор роста IGF-I.
Третичные фолликулы характеризуются дальнейшим ростом. Между фолликулярными клетками появляются округлые полости, заполненные жидкостью (liquor folliculi). Появляется доминантный фолликул, опережающий в росте остальные, а в его составе формируется выраженная theca.
• Фолликулярные клетки усиливают продукцию эстрогенов. Эстрогены по аутокринному механизму увеличивают плотность рецепторов фоллитропина в мембране фолликулярных клеток.
• Фоллитропин стимулирует появление в мембране фолликулярных клеток рецепторов лютропина.
• Высокое содержание эстрогенов в крови блокирует секрецию фоллитропина, что тормозит развитие других первичных фолликулов и стимулирует секрецию лютеинизирующего гормона (лютропина).
• Уровень лютропина поднимается в конце фолликулярной стадии цикла. Лютеинизирующий гормон стимулирует синтез андрогенов в клетках theca.
• Андрогены из theca через базальную мембрану (стекловидная оболочка на более поздних стадиях развития) диффундируют в клетки гранулёзы, где при помощи ароматазы конвертируются в эстрогены.
Зрелый фолликул
Зрелый фолликул достигает 1,0-2,5 см в диаметре, прежде всего за счёт накопления жидкости в его полости. В полость пузырька вдаётся холмик из фолликулярных клеток (cumulus oophorus), внутри которого находится яйцеклетка. Яйцеклетка (на стадии овоцита первого порядка) окружена прозрачной оболочкой, снаружи от которой располагаются фолликулярные клетки (corona radiata). Стенка зрелого фолликула состоит из прозрачной оболочки, зернистой оболочки, теки.
• Уровень эстрогенов (см. рис. 15-23В) быстро увеличивается, достигая пика приблизительно за 24-36 часов до овуляции.
• Содержание лютеинизирующего гормона увеличивается постепенно до середины цикла. Резкий подъём его уровня происходит через 12 часов после наступления пика эстрогенов. Лютропин вызывает лютеинизацию клеток гранулёзы и theca interna (накопление жёлтого пигмента, липидов) и индуцирует преовуляторный синтез прогестерона.
• Преовуляторное повышение уровня прогестерона (см. рис. 15-23А) облегчает положительное обратное действие эстрогенов (через синтез и секрецию лютропина), а также индуцирует преовуляторный пик фоллитропина за счёт усиления гипофизарного ответа на гонадолиберин.
Овуляция
Овуляция (см. рис. 15-23А, 15-23Б и 15-23В) происходит примерно через 24-36 часов после пика эстрогенов и через 10-12 часов после достижения пика лютеинизирующего гормона, чаще всего на 11-й, 12-й или 13-й дни 28-дневного цикла. Следует, однако, помнить, что при 28-дневном цикле овуляция возможна между 8-м и 20-м днями. Истончение и разрыв стенки фолликула происходят под влиянием простагландинов и протеолитических ферментов гранулёзы.
• Первое мейотическое деление овоцита завершается в зрелом фолликуле перед овуляцией на пике лютеинизирующего гормона. Лютропин стимулирует деление овоцита первого порядка с образованием овоцита второго порядка и первого полярного тельца.
• Второе деление мейоза завершается не ранее оплодотворения.
Жёлтое тело
В лютеиновую стадию овариально-менструального цикла под действием лютеинизирующего гормона на месте лопнувшего зрелого фолликула образуется менструальное жёлтое тело. Жёлтое тело развивается из элементов преовуляторного фолликула и состоит из лютеинизированных фолликулярных клеток (гранулёза-лютеиновые) и клеток theca interna (тека-лютеиновые), между которыми расположены капилляры синусоидного типа. В ходе лютеинизации клетки гипертрофируются, в них увеличивается содержание элементов гладкой эндоплазматической сети, накапливаются жировые включения. Васкуляризация жёл- того тела увеличивает поступление холестерина в клетки гранулёзы.
Лютропин стимулирует в клетках гранулёзы синтез прогестерона, и прогестерон секретируется в кровь.
Менструальное жёлтое тело функционирует в лютеиновую стадию цикла, поддерживая в крови высокий уровень эстрогенов и прогестерона, обеспечивающих подготовку эндометрия к имплантации. Если оплодотворения не произошло, жёлтое тело подвергается инволюции, а в крови резко снижается содержание прогестерона и эстрогенов. Жёлтое тело беременности. Дальнейшее развитие жёлтого тела стимулирует хорионический гонадотропин (ХГТ), вырабатываемый в трофобласте. ХГТ, секретируемый клетками трофобласта, через рецепторы лютропина лютеиновых клеток стимулирует рост жёлтого тела, достигающего в диаметре 5 см, и увеличивает продукцию половых гормонов. Высокий уровень прогестерона и эстрогенов сохраняет развивающуюся беременность. Релаксин (гормон из семейства инсулинов) синтезируется клетками жёлтого тела и цитотрофобластом и в течение беременности оказывает расслабляющий эффект на миометрий, уменьшает плотность лонного сочленения. Жёлтое тело беременности активно в течение первой половины беременности, затем его функция постепенно угасает.
Белое тело - соединительнотканный рубец на месте дегенерировавшего жёлтого тела.
Гормональная регуляция овариального цикла
Овариально-менструальный цикл (рис. 15-23) контролируют гипофизарные гонадотропины - фолликулостимулирующий (фоллитропин) и лютеинизирующий (лютропин). Эндокринную функцию передней доли гипофиза регулирует гипоталамический люлиберин. В свою очередь, гормоны яичника (эстрогены, прогестерон, а также ингибин) вовлечены в регуляцию синтеза и секреции гонадотропинов гипофиза и люлиберина. Таким образом, циклические изменения яичника и эндометрия - иерархическая (гипоталамус → гипофиз → яичники → матка) и саморегулируемая (яичники → гипоталамус и гипофиз) система, функционирующая в течение репродуктивного периода (от менархе до наступления климактерических изменений - менопаузы).
МАТОЧНЫЕ ТРУБЫ
Яйцевод - трубчатый орган длиной 10-12 см и диаметром 0,5-1 см, ответственный за оплодотворение и обеспечивающий перемещение концептуса в матку (см. рис. 3-6). Анатомически яйцевод делят на воронку (infundibulum), от которой отходят многочисленные отростки; ампулу - самый длинный и с наибольшим диаметром просвета отдел, где происходит оплодотворение; перешеек (isthmus) - примыкающий
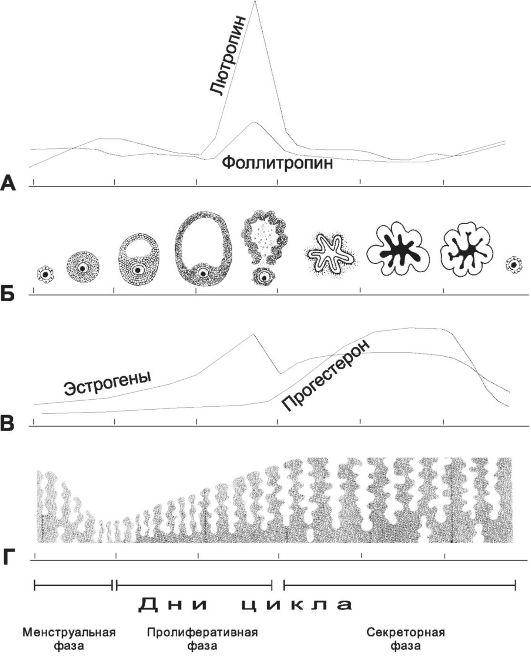
Рис. 15-23. Овариально-менструальный цикл. Циклические изменения содержания в крови гонадотропных гормонов регулируют созревание очередного фолликула и овуляцию (А и Б). По мере развития фолликула в крови повышается уровень эстрогенов, а с момента овуляции и образования жёлтого тела увеличивается концентрация прогестерона (Б и В). Эстрогены и прогестерон вызывают характерные изменения в эндометрии матки в зависимости от стадии цикла (В и Г). [17]
к матке короткий отдел и интрамуральный отдел (pars uterina). В стенке маточной трубы различают три оболочки: слизистую, мышечную и серозную (отсутствует во внутриматочной части трубы). Слизистая оболочка окружает просвет яйцевода, образуя огромное количество ветвящихся складок. Собственный слой слизистой оболоч-
ки построен из рыхлой волокнистой неоформленной соединительной ткани, богатой кровеносными сосудами. Эпителий слизистой оболочки состоит из одного слоя цилиндрических клеток, среди которых различают реснитчатые и секреторные клетки.
• Секреторные клетки имеют выраженную гранулярную эндоплазматическую сеть и комплекс Гольджи. В апикальной части клеток содержится большое количество секреторных гранул. Клетки более активны в секреторную фазу менструального цикла. Они продуцируют жидкость, содержащую питательные вещества для сперматозоидов и развития концептуса.
• Реснитчатые клетки несут на апикальной поверхности реснички, совершающие движения по направлению к матке. Сила и амплитуда биения ресничек зависит от эстрогенов и прогестерона.
Мышечная оболочка состоит из двух слоёв ГМК (внутреннего циркулярного и наружного продольного). Слои отделены друг от друга прослойкой соединительной ткани со значительным количеством кровеносных сосудов.
Перемещение яйцеклетки. Перед овуляцией увеличение тонуса ГМК маточной трубы приближает воронку маточной трубы к поверхности яичника. Биение ресничек эпителиальных клеток фимбрий ампулы маточной трубы создаёт ток жидкости, способствующий перемещению яйцеклетки из брюшной полости в ампулу.
МАТКА
Матка - полый мышечный орган, предназначенный для развития плода. В матке различают тело, дно и шейку. Дно матки располагается над устьем маточных труб. Книзу тело матки продолжается в канал шейки матки, открывающийся во влагалище. Стенка матки образована тремя оболочками: слизистой, мышечной и серозной (рис. 15-26). Слизистая оболочка матки (endometrium) выстлана однослойным цилиндрическим эпителием, лежащим на рыхлой волокнистой неоформленной соединительной ткани собственного слоя. Среди эпителиальных клеток различают секреторные и реснитчатые клетки. В собственном слое имеются маточные железы (крипты) - длинные изогнутые простые трубчатые железы, открывающиеся в просвет матки. Мышечная оболочка матки (myometrium) состоит из трёх слоёв гладкой мускулатуры. Направление удлинённых ГМК в слоях мышечной оболочки различное: продольное в наружном и внутреннем, циркулярное - в среднем. В среднем слое находится множество кровеносных сосудов (stratum vasculare). Величина ГМК, их количество, толщина мышечной оболочки резко увеличиваются при беременности. При родах окситоцин и простагландины (PGE2 и PGF2a) стимулируют сокращение ГМК, облегчая изгнание плода.
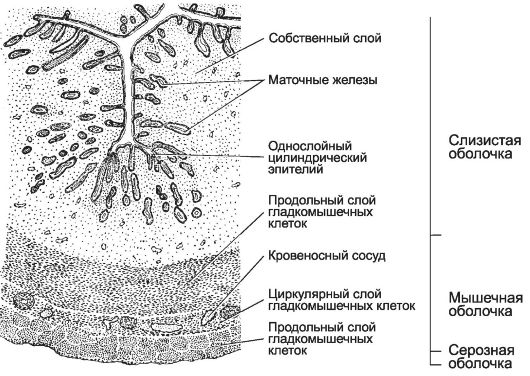 Рис. 15-26. Матка. Слизистая
оболочка состоит из однослойного цилиндрического эпителия и
собственного слоя. В собственном слое слизистой оболочки присутствуют
простые трубчатые железы. Мышечная оболочка представлена внутренним и
наружным продольными слоями ГМК и средним - циркулярным. Снаружи орган
окружает серозная оболочка. [17]
Рис. 15-26. Матка. Слизистая
оболочка состоит из однослойного цилиндрического эпителия и
собственного слоя. В собственном слое слизистой оболочки присутствуют
простые трубчатые железы. Мышечная оболочка представлена внутренним и
наружным продольными слоями ГМК и средним - циркулярным. Снаружи орган
окружает серозная оболочка. [17]
Шейка матки
Это нижний сегмент матки (2,5-3 см), частично выступающий во влагалище (рис. 15-27). Надвлагалищная часть шейки матки, располагающаяся выше места прикрепления стенок влагалища, открывается в полость матки внутренним маточным зевом. Влагалищная порция шейки матки (экзоцервикс) заканчивается наружным маточным зевом. Снаружи влагалищная порция покрыта многослойным плоским эпителием, который полностью обновляется каждые 4-5 дней. Шейка матки имеет узкий и несколько расширенный в средней части канал (эндоцервикс). Стенка шейки образована плотной соединительной тканью. Среди коллагеновых и эластических волокон встречаются продольные пучки ГМК. В родах под действием релаксина плотная соединительная ткань становится мягкой и податливой. Слизистая оболочка канала шейки матки состоит из однослойного цилиндрического эпителия и собственного слоя. В эпителии различают железистые
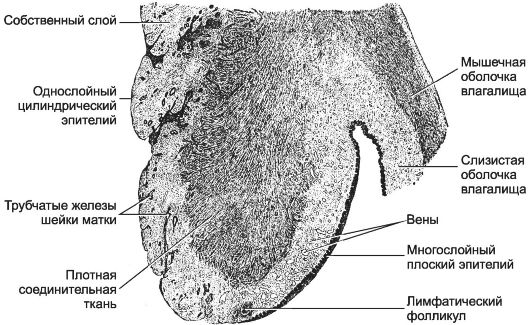 Рис. 15-27. Влагалищная часть шейки матки. Слизистая
оболочка канала шейки матки выстлана однослойным цилиндрическим
эпителием. На поверхность открываются разветвлённые трубчатые железы.
Вблизи наружного маточного зева однослойный цилиндрический эпителий
переходит в многослойный плоский. [17]
Рис. 15-27. Влагалищная часть шейки матки. Слизистая
оболочка канала шейки матки выстлана однослойным цилиндрическим
эпителием. На поверхность открываются разветвлённые трубчатые железы.
Вблизи наружного маточного зева однослойный цилиндрический эпителий
переходит в многослойный плоский. [17]
клетки, продуцирующие слизь, и клетки, имеющие реснички. В просвет канала открываются многочисленные разветвлённые трубчатые железы, располагающиеся в собственном слое слизистой оболочки. Вблизи наружного зева однослойный цилиндрический эпителий слизистой оболочки канала шейки матки переходит в богатый гликогеном многослойный плоский, покрывающий влагалищную порцию шейки матки и продолжающийся далее в составе слизистой оболочки стенки влагалища.
ВЛАГАЛИЩЕ
Влагалище - фиброзно-мышечная трубка, состоящая из слизистой, мышечной и адвентициальной оболочек. Слизистая оболочка представлена многослойным плоским эпителием и собственным слоем, содержащим богатую сеть кровеносных сосудов. Многослойный плоский эпителий изнутри кнаружи состоит из базальных, промежуточных и поверхностных клеток. За счёт базальных (ростковых) клеток происходит постоянная регенерация пласта эпителия. Эпителий подвергается частичному ороговению (поверхностные клетки содержат гранулы
кератогиалина). Рост и созревание эпителия находится под гормональным контролем. Эпителий истончён до менархе и после менопаузы, но становится толще в репродуктивном периоде. Кроме того, толщина эпителия зависит от стадии цикла. В собственном слое присутствуют лимфоциты, зернистые лейкоциты, иногда обнаруживаются лимфатические фолликулы. Мышечная оболочка состоит из двух слоёв ГМК и развитой сети эластических волокон. Внутренний тонкий слой содержит ГМК с циркулярной ориентацией. Наружный толстый слой образован продольными пучками ГМК. Адвентициальная оболочка образована волокнистой соединительной тканью и соединяет влагалище с окружающими структурами.
НАРУЖНЫЕ ПОЛОВЫЕ ОРГАНЫ
Большие половые губы - две кожные складки по боковым сторонам половой щели. С наружной поверхности большие половые губы покрыты волосистой кожей с сальными и потовыми (эккриновыми и апокриновыми) железами. На внутренней поверхности волосяных фолликулов нет. В толще губ расположены венозные сплетения, жировая клетчатка и большие железы преддверия (glandulae vestibulares majores). Парные большие железы преддверия находятся на границе передней и средней трети губ. Это разветвлённые секретирующие слизь трубчато-альвеолярные железы, открывающиеся в преддверие влагалища. Секрет с характерным запахом обычно выделяется при половом возбуждении и увлажняет поверхность преддверия и входа во влагалище.
Малые половые губы расположены кнутри от больших губ и скрыты последними. Малые половые губы лишены жировой ткани, но имеют многочисленные эластические волокна и богатое сосудистое сплетение. Пигментированная кожа содержит сальные железы и небольшие слизистые железы (glandulae vestibulares minores, малые железы преддверия), открывающиеся в преддверие влагалища. Клитор гомологичен дорсальной стороне полового члена, состоит из двух пещеристых тел (corpora cavernosa clitoridis), формирующих на дистальном конце головку (glans clitoridis). Клитор покрыт многослойным плоским эпителием со слабым ороговением; волосы, сальные и потовые железы отсутствуют. Кожа содержит многочисленные свободные и инкапсулированные нервные окончания.
Менструальный цикл
Изменения гормонального фона (содержание в крови эстрогенов и прогестерона в разные дни овариального цикла (см. рис. 15-23) прямо влияют на состояние эндо-
метрия, а также слизистой оболочки маточных труб, цервикального канала и влагалища. Эстрогены контролируют пролиферативную фазу менструального цикла (восстановление функционального слоя эндометрия), а прогестерон - секреторную фазу (подготовку эндометрия к имплантации, уменьшение порога возбудимости ГМК миометрия и поддержание тонуса ГМК шейки матки). Снижение содержания в крови эстрогенов и прогестерона сопровождается отторжением функционального слоя эндометрия и маточным кровотечением (менструальная фаза).
Эндометрий
Слизистая оболочка матки (рис. 15-29) подвергается циклическим изменениям (менструальный цикл). В каждом цикле эндометрий проходит менструальную, пролиферативную и секреторную фазы. В эн-
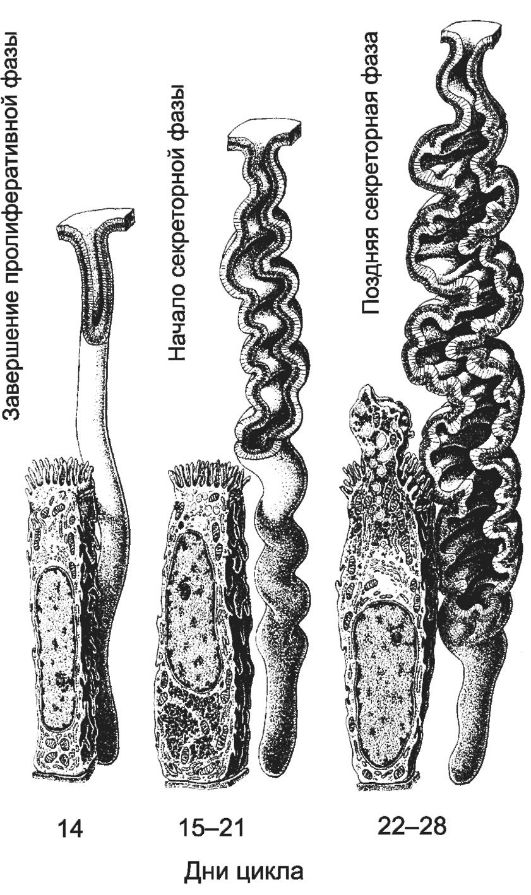 Рис. 15-29. Маточные железы
Рис. 15-29. Маточные железы
на различных фазах менструального цикла. Морфо-функциональное состояние маточных желёз зависит от фазы овариально-менструального цикла. При менструации маточные крипты разрушаются. В пролиферативную фазу за счёт сохранившихся эпителиальных клеток базального слоя происходит восстановление крипт. Железы имеют прямую трубчатую форму. Секреторная фаза характеризуется функциональной активностью маточных желёз. В начале секреторной фазы железы приобретают извилистый ход, в базальной части клеток накапливается гликоген. В поздней секреторной фазе железы расширяются, становятся более извилистыми, клетки приобретают секреторную активность. [17]
дометрии различают функциональный и базальный слои. Базальный слой эндометрия кровоснабжается из прямых артерий и сохраняется в менструальную фазу. Функциональный слой эндометрия, отпадающий при менструации, кровоснабжается из спиральных артерий, склерозирующихся в менструальную фазу, вызывая ишемию функционального слоя.
Менструальная фаза - отторжение функционального слоя эндометрия. Если не произошло имплантации, менструальное жёлтое тело подвергается инволюции, а в крови резко снижается содержание гормонов яичника (прогестерона и эстрогенов). Это приводит к скручиванию, склерозированию и уменьшению просвета спиральных артерий, снабжающих кровью верхние две трети функционального слоя эндометрия. В результате происходит ухудшение кровотока в функциональном слое эндометрия. Развивающаяся ишемия приводит к отторжению функционального слоя и генитальному кровотечению. При менструации функциональный слой отторгается полностью, базальный слой эндометрия сохраняется. При длительности цикла 28 дней менструация в среднем продолжается 5±2 дня.
Пролиферативная фаза следует за менструальной и длится до момента овуляции, т.е. до конца фолликулярной стадии цикла. В это время фоллитропин стимулирует рост и созревание фолликула и усиливает продукцию фолликулярными клетками эстрогенов. Под влиянием эстрогенов (в основном эстрадиола) происходят пролиферация клеток базального слоя и восстановление (регенерация) функционального слоя эндометрия. Эпителиальные клетки желёз базального слоя мигрируют на поверхность, пролиферируют и образуют новую эпителиальную выстилку слизистой оболочки. В эндометрии формируются новые маточные железы, из базального слоя врастают спиральные артерии. Длительность фазы может варьировать. Секреторная фаза продолжается от овуляции до начала менструации (12-16 дней). В эту фазу в яичнике функционирует жёлтое тело, вырабатывающее эстрогены и прогестерон. Высокий уровень прогестерона создаёт благоприятные условия для имплантации бластоцисты.
• Маточные железы расширяются, становятся извилистыми. Железистые клетки прекращают деление, гипертрофируются и начинают секретировать гликоген, гликопротеины, липиды, муцин. Секрет поднимается к устью маточных желёз и выделяется в просвет матки.
• Спиральные артерии приобретают более извитой характер и приближаются к поверхности слизистой оболочки.
• Компактная и губчатая зоны функционального слоя. В поверхностных частях функционального слоя увеличивается количество соединительнотканных клеток, в цитоплазме которых накапливаются гликоген и липиды. Вокруг клеток формируются коллагеновые и ретикулиновые волокна, образованные коллагеном типов I и III. Клетки стромы приобретают черты децидуальных клеток плаценты. Благодаря таким изменениям эндометрия, в функциональном слое образуется две зоны: компактная (zona compacta), обращённая к просвету, и более глубокая - губчатая (zona spongiosa).
Шейка матки
В собственном слое слизистой оболочки шейки матки отсутствуют спиральные артерии, поэтому в менструальную фазу слизистая оболочка шейки матки не отторгается подобно эндометрию тела матки. Вместе с тем, в фолликулярную ста-
дию цикла железы эндоцервикса продуцируют водянистый секрет, а в лютеиновую стадию цикла - вязкий слизистый секрет, заполняющий канал шейки матки наподобие пробки и препятствующий проникновению в матку сперматозоидов и микроорганизмов. В лютеиновую стадию прогестерон поддерживает тонус ГМК шейки матки.
Влагалище
Фолликулярная стадия. В начале стадии влагалищный эпителий тонкий и бледный. Под влиянием эстрогенов происходит пролиферация эпителия, который достигает своей максимальной толщины. В клетках накапливается значительное количество гликогена, используемого влагалищной микрофлорой. Образующаяся при этом молочная кислота препятствует развитию патогенных микроорганизмов. В эпителии появляются признаки частичной кератинизации.
Лютеиновая стадия. Рост и созревание эпителиальных клеток блокируется, на поверхности эпителия появляются лейкоциты и роговые чешуйки.
Молочная железа
Молочная железа - производное эпидермиса и относится к железам кожи. Однако, развитие железы и её функциональная активность зависят от гормонов половой сферы.
Гистологическая картина молочной железы зависит от степени её зрелости и активности. Морфологические различия обнаруживаются в строении ювенильной железы, зрелой неактивной и зрелой функционирующей (лактирующей) желёз (рис. 15-30).
Ювенильная железа представлена выводными междольковыми и внутридольковыми протоками, разделёнными соединительнотканными перегородками; секреторные отделы отсутствуют. Зрелая неактивная железа. При половом созревании под влиянием эстрогенов железа увеличивается в объёме. Выводные протоки становятся более разветвлёнными, а среди соединительнотканных перегородок накапливается жировая ткань. Секреторные отделы, как и в ювенильной железе, отсутствуют.
Лактирующая железа (см. рис. 15-30). Под влиянием прогестерона в сочетании с эстрогенами, пролактином и хорионическим соматомаммотропным гормоном индуцируется дифференцировка секреторных отделов молочной железы. Уже на третьем месяце беременности из растущих концевых отделов внутридольковых протоков формируются почки, дифференцирующиеся в секреторные отделы - альвеолы. Альвеолы выстланы кубическими секреторными клетками (альвеолярные клетки). Снаружи стенку альвеол и выводных протоков окружают миоэпителиальные клетки. Внутридольковые протоки выстланы однослойным кубическим эпителием, который в молочных протоках переходит в многослойный плоский. Соединительнотканные перего-
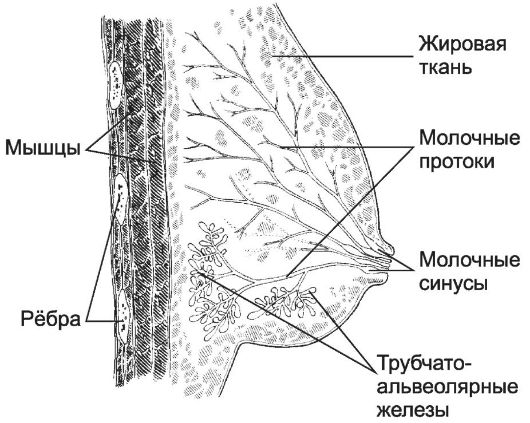 Рис. 15-30. Лактирующая молочная железа состоит
из 15-20 отдельных сложных альвеолярно-трубчатых желёз, каждая из
которых открывается собственным выводным протоком (молочный проток) на
вершине соска. Сосок состоит из плотной соединительной ткани с большим
содержанием эластических волокон и ГМК, располагающихся циркулярно в
основании соска и параллельно молочным протокам. Кожа вокруг соска
содержит крупные сальные железы, образующие возвышения (бугорки
околососкового кружка). Молочные протоки под ареолой образуют молочные
синусы, резервуары для молока. Пигментация эпидермиса соска происходит в
пубертатном периоде, кожа ареолы пигментируется при беременности. [17]
Рис. 15-30. Лактирующая молочная железа состоит
из 15-20 отдельных сложных альвеолярно-трубчатых желёз, каждая из
которых открывается собственным выводным протоком (молочный проток) на
вершине соска. Сосок состоит из плотной соединительной ткани с большим
содержанием эластических волокон и ГМК, располагающихся циркулярно в
основании соска и параллельно молочным протокам. Кожа вокруг соска
содержит крупные сальные железы, образующие возвышения (бугорки
околососкового кружка). Молочные протоки под ареолой образуют молочные
синусы, резервуары для молока. Пигментация эпидермиса соска происходит в
пубертатном периоде, кожа ареолы пигментируется при беременности. [17]
родки, разделяющие дольки молочной железы, становятся менее выраженными.
Секреция. Молоко секретируется по апокриновому типу. Жиры высвобождаются вместе с фрагментами клеточной мембраны, остальные компоненты молока выделяются путём экзоцитоза.