Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 2 КЛЕТКА
Клетка - главный гистологический элемент. Эукариотическая клетка состоит из трёх основных компартментов: плазматическая мембрана, ядро и цитоплазма со структурированными клеточными единицами (органеллы, включения). Важное значение для организации клеток имеют биологические мембраны, входящие в состав каждого клеточного компартмента и многих органелл. Мембраны клеток имеют принципиально сходную организацию. Любую клетку снаружи ограничивает плазматическая мембрана.
Плазматическая мембрана
Согласно жидкостно-мозаичной модели, плазматическая мембрана - жидкая динамическая система с мозаичным расположением белков и липидов (рис. 2-1). В плоскости мембраны белки обладают латеральной подвижностью. Интегральные белки перераспределяются в мембранах в результате взаимодействия с периферическими белками, элементами цитоскелета, молекулами в мембране соседней клетки и компонентами внеклеточного вещества. Основные функции плазматической мембраны: избирательная проницаемость, межклеточные взаимодействия, эндоцитоз, экзоцитоз.
Химический состав
В состав плазматической мембраны входят липиды, холестерин, белки и углеводы.
Липиды (фосфолипиды, сфинголипиды, гликолипиды) составляют до 45% массы мембран.
• Фосфолипиды. Молекула фосфолипида состоит из полярной (гидрофильной) части (головка) и аполярного (гидрофобного) двойного углеводородного хвоста. В водной фазе молекулы фосфолипидов автоматически агрегируют хвост к хвосту, формируя каркас биологической мембраны в виде двойного слоя (бислой). Таким образом, в мембране хвосты фосфолипидов направлены внутрь бислоя, а головки обращены кнаружи.
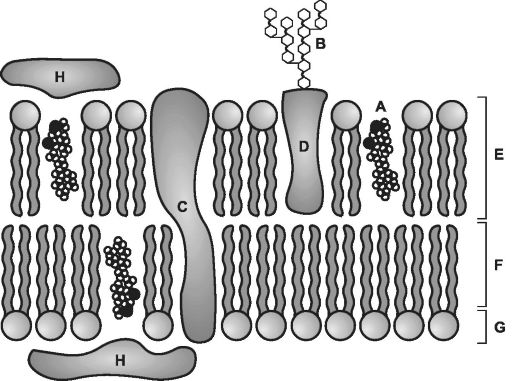 Рис. 2-1. Компоненты плазматической мембраны. A - холестерин; B - олигосахарид в составе гликопротеина на наружной поверхности; C и D - интегральные белки; E - молекулы фосфолипидов; F - хвосты жирных кислот в составе фосфолипидов; G - полярные головки фосфолипидов; H - периферический белок. [17]
Рис. 2-1. Компоненты плазматической мембраны. A - холестерин; B - олигосахарид в составе гликопротеина на наружной поверхности; C и D - интегральные белки; E - молекулы фосфолипидов; F - хвосты жирных кислот в составе фосфолипидов; G - полярные головки фосфолипидов; H - периферический белок. [17]
• Сфинголипиды - липиды, содержащие основание с длинной цепью (сфингозин или сходную с ним группу); сфинголипиды в значительном количестве находятся в миелиновых оболочках нервных волокон, слоёв модифицированной плазмолеммы шванновских клеток и олигодендроглиоцитов ЦНС.
• Гликолипиды - молекулы содержащих олигосахариды липидов, присутствующие в наружной части бислоя, а их остатки сахаров ориентированы к поверхности клетки. Гликолипиды составляют 5% липидных молекул наружного монослоя.
Холестерин имеет чрезвычайно важное значение не только как компонент биологических мембран, на основе холестерина происходит синтез стероидных гормонов - половых, глюкокортикоидов, минералокортикоидов.
Белки составляют более 50% массы мембран. Белки плазмолеммы подразделяют на интегральные и периферические.
• Интегральные мембранные белки прочно встроены в липидный бислой. Примеры интегральных мембранных белков - белки ионных каналов и рецепторные белки
(мембранные рецепторы). Молекула белка, проходящая через всю толщу мембраны и выступающая из неё как на наружной, так и на внутренней поверхности, - трансмембранный белок.
• Периферические мембранные белки (фибриллярные и глобулярные) находятся на одной из поверхностей клеточной мембраны (наружной или внутренней) и нековалентно связаны с интегральными мембранными белками. Примерами периферических мембранных белков, связанных с наружной поверхностью мембраны, могут служить рецепторные и адгезионные белки. Примеры периферических мембранных белков, связанных с внутренней поверхностью мембраны, - белки, ассоциированные с цитоскелетом (например, дистрогликаны, белок полосы 4.1, протеинкиназа С), белки системы вторых посредников.
Углеводы (преимущественно олигосахариды) входят в состав гликопротеинов и гликолипидов мембраны, составляя 2-10% её массы. С углеводами клеточной поверхности взаимодействуют лектины. Цепи олигосахаридов, ковалентно связанных с гликопротеинами и гликолипидами плазмолеммы, выступают на наружной поверхности мембран клетки и формируют поверхностную оболочку толщиной 50 нм - гликокаликс. Гликокаликс участвует в процессах межклеточного узнавания, межклеточного взаимодействия, пристеночного пищеварения.
Избирательная проницаемость
Трансмембранная избирательная проницаемость поддерживает клеточный гомеостаз, оптимальное содержание в клетке ионов, воды, ферментов и субстратов. Пути реализации избирательной проницаемости мембран: пассивный транспорт, облегчённая диффузия, активный транспорт. Гидрофобный характер сердцевины бислоя определяет возможность (или невозможность) непосредственного проникновения через мембрану различных с физико-химической точки зрения веществ (в первую очередь, полярных и неполярных).
• Неполярные вещества (например, холестерин и его производные) свободно проникают через биологические мембраны. По этой причине эндоцитоз и экзоцитоз полярных соединений (например, пептидных гормонов) происходят при помощи мембранных пузырьков, а секреция стероидных гормонов - без участия таких пузырьков. По этой же причине рецепторы неполярных молекул (например, стероидных гормонов) расположены внутри клетки.
• Полярные вещества (например, белки и ионы) не могут проникать через биологические мембраны. Именно поэтому рецепторы полярных молекул (например, пептидных гормонов) встроены в плазматическую мембрану, а передачу сигнала к другим клеточным компартментам осуществляют вторые посредники. По этой же причине трансмембранный перенос полярных соединений осуществляют специальные системы, встроенные в биологические мембраны.
Межклеточные информационные взаимодействия
Клетка, воспринимая и трансформируя различные сигналы, реагирует на изменения окружающей её среды. Плазматическая мембрана - место приложения физических (например, кванты света в фоторецепторах), химических (например, вкусовые и обонятельные молекулы, рН), механических (например, давление или растяжение в механорецепторах) раздражителей внешней среды и сигналов информацион-
ного характера (например, гормоны, нейромедиаторы) из внутренней среды организма. При участии плазмолеммы происходят узнавание и агрегация (например, межклеточные контакты) как соседних клеток, так и клеток с компонентами внеклеточного вещества (например, адгезионные контакты, адресная миграция клеток и направленный рост аксонов в нейроонтогенезе). Информационные межклеточные взаимодействия укладываются в схему, предусматривающую следующую последовательность событий:
сигнал → рецептор → (второй посредник) → ответ.
Сигналы. Передачу сигналов от клетки к клетке осуществляют сигнальные молекулы (первый посредник), вырабатываемые в одних клетках и специфически воздействующие на другие клетки - клетки-мишени. Специфичность воздействия сигнальных молекул определяют присутствующие в клетках-мишенях рецепторы, связывающие только собственные лиганды. Все сигнальные молекулы (лиганды) - в зависимости от их физико-химической природы - подразделяют на полярные (точнее - гидрофильные) и аполярные (точнее - жирорастворимые). Рецепторы регистрируют поступающий к клетке сигнал и передают его вторым посредникам. Различают мембранные и ядерные рецепторы.
• Мембранные рецепторы - гликопротеины. Они контролируют проницаемость плазмолеммы путём изменения конформации белков ионных каналов (например, н-холинорецептор), регулируют поступление молекул в клетку (например, холестерина), связывают молекулы внеклеточного вещества с элементами цитоскелета (например, интегрины), регистрируют присутствие информационных сигналов (например, нейромедиаторов, квантов света, обонятельных молекул, антигенов, цитокинов, гормонов пептидной природы). Мембранные рецепторы регистрируют поступающий к клетке сигнал и передают его внутриклеточным химическим соединениям, опосредующим конечный эффект (вторые посредники). Функционально мембранные рецепторы подразделяют на каталитические, связанные с ионными каналами и оперирующие через G-белок.
• Ядерные рецепторы - белки-рецепторы стероидных гормонов (минерало- и глюкокортикоиды, эстрогены, прогестерон, тестостерон), ретиноидов, тиреоидных гормонов, жёлчных кислот, витамина D3. Каждый рецептор имеет область связывания лиганда и участок, взаимодействующий со специфическими последовательностями ДНК. Другими словами, ядерные рецепторы - активируемые лигандом транскрипционные факторы. В геноме человека имеется более 30 ядерных рецепторов, лиганды которых находятся на стадии идентификации (сиротские рецепторы).
• Внерецепторные низкомолекулярные сигналы. Некоторые низкомолекулярные сигналы (например, оксид азота и монооксид углерода) воздействуют на клетку-мишень, минуя рецепторы.
♦ Оксид азота (NO) - газообразный медиатор межклеточных взаимодействий, образуется из L-аргинина при участии фермента NO-синтазы. В клеткахмишенях активирует гуанилатциклазу, что приводит к увеличению уровня второго посредника - цГМФ.
♦ Монооксид углерода (угарный газ, CO). Как сигнальная молекула CO играет важную роль в иммунной, сердечно-сосудистой системах и периферической нервной системе.
Вторые посредники. Внутриклеточные сигнальные молекулы (вторые посредники) передают информацию с мембранных рецепторов на эффекторы (исполнительные молекулы), опосредующие ответ клетки на сигнал. Стимулы, такие как свет, запах, гормоны и другие химические сигналы (лиганды), инициируют ответ клетки-мишени, изменяя в ней уровень внутриклеточных вторых посредников. Вторые (внутриклеточные) посредники представлены многочисленным классом соединений. К ним относятся циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, диацилглицерол, Ca2+.
Ответы клеток-мишеней. Функции клеток выполняются на разных уровнях реализации генетической информации (например, транскрипция, посттрансляционная модификация) и крайне разнообразны (например, изменения режима функционирования, стимуляция или подавление активности, перепрограммирование синтезов и так далее).
Эндоцитоз
Эндоцитоз - поглощение (интернализация) клеткой воды, веществ, частиц и микроорганизмов (рис. 2-8А). К вариантам эндоцитоза относят пиноцитоз, фагоцитоз, опосредованный рецепторами эндоцитоз с образованием окаймлённых клатрином пузырьков и клатрин-независимый эндоцитоз с участием кавеол.
Пиноцитоз - процесс поглощения жидкости и растворённых веществ с образованием небольших пузырьков. Пиноцитоз рассматривают как неспецифический способ поглощения внеклеточных жидкостей и содержащихся в ней веществ, когда некоторая область клеточной мембраны впячивается, образует ямку и далее пузырёк, содержащий межклеточную жидкость.
Опосредуемый рецепторами эндоцитоз (рис. 2-9) характеризуется поглощением из внеклеточной жидкости конкретных макромолекул, связываемых специфическими рецепторами, расположенными в плазмолемме. Последовательность событий опосредованного рецепторами эндоцитоза такова: взаимодействие лиганда с мембранным рецептором → концентрирование комплекса «лиганд-рецептор» на поверхности окайм- лённой ямки → формирование окаймлённого клатрином пузырька → погружение в клетку окаймлённого пузырька. Обладающий ГТФазной активностью хемомеханический белок динамин на стыке плазмолеммы и окаймлённого пузырька формирует т.н. молекулярную пружину, которая при расщеплении ГТФ распрямляется и отталкивает пузырёк от плазмолеммы (рис. 2-9Б). Подобным образом клетка поглощает трансферрин, холестерин вместе с ЛНП и многие другие молекулы.
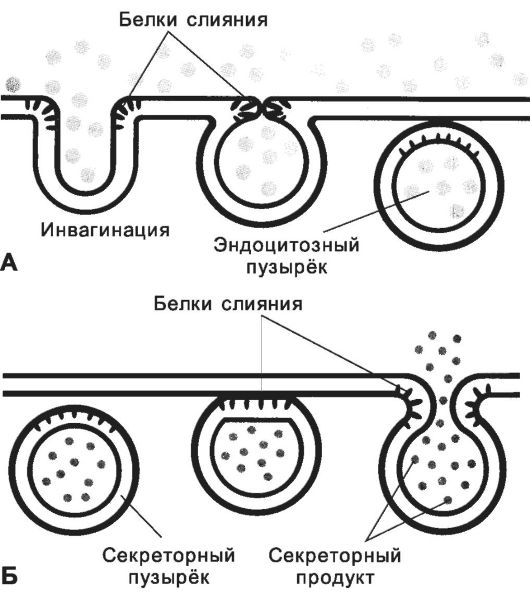 Рис. 2-8. Эндоцитоз (А) и экзоцитоз (Б). При
эндоцитозе участок плазматической мембраны впячивается и замыкается.
Инвагинация плазмолеммы и сближение краёв формирующейся ямки происходит
главным образом за счёт перестройки примембранного F-актина. Образуется
эндоцитозный пузырёк, содержащий поглощённые частицы. При экзоцитозе
мембрана транспортных или секреторных пузырьков сливается с
плазматической мембраной, и содержимое пузырьков высвобождается во
внеклеточное пространство. В слиянии мембран участвуют специальные
белки. [17]
Рис. 2-8. Эндоцитоз (А) и экзоцитоз (Б). При
эндоцитозе участок плазматической мембраны впячивается и замыкается.
Инвагинация плазмолеммы и сближение краёв формирующейся ямки происходит
главным образом за счёт перестройки примембранного F-актина. Образуется
эндоцитозный пузырёк, содержащий поглощённые частицы. При экзоцитозе
мембрана транспортных или секреторных пузырьков сливается с
плазматической мембраной, и содержимое пузырьков высвобождается во
внеклеточное пространство. В слиянии мембран участвуют специальные
белки. [17]
Клатрин-независимый эндоцитоз. Путём клатрин-независимого эндоцитоза происходит поглощение многих объектов и молекул, например, рецептора трансформирующего фактора роста TGFb, токсинов, вирусов и др. Один из путей клатрин-независимого эндоцитоза - поглощение молекул с помощью небольших инвагинаций плазмолеммы диаметром 50-80 нм - кавеол. Кавеолы характерны для большинства клеточных типов; особенно многочисленны в эндотелиальных клетках, где они участвуют в транспорте крупных макромолекул.
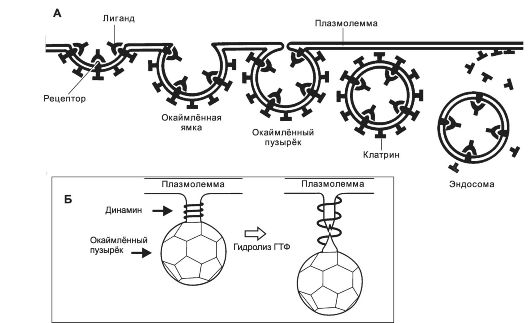 Рис. 2-9. Опосредуемый рецепторами эндоцитоз (А).
Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы и
др.) связываются со своими рецепторами в плазмолемме. Образуются
окаймлённые клатрином ямки, а затем - окаймлённые пузырьки, содержащие
комплекс «лиганд-рецептор». Окаймлённые пузырьки после освобождения от
клатрина - эндосома. Внутри эндосом лиганд отщепляется от рецептора. Погружение окаймлённого пузырька в цитоплазму (Б). Присоединение
к динамину ГТФ инициирует формирование механохимической молекулярной
пружины из плотно упакованных колец динамина с шагом в 11 нм вокруг
шейки эндоцитозного пузырька. При гидролизе ГТФ расстояние между
смежными кольцами увеличивается до 22 нм. Таким образом, ГТФаза динамина
катализирует расщепление ГТФ и тем самым растяжение пружины с
последующим отрывом окружённого клатрином эндоцитозного пузырька от
плазмолеммы. [17]
Рис. 2-9. Опосредуемый рецепторами эндоцитоз (А).
Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы и
др.) связываются со своими рецепторами в плазмолемме. Образуются
окаймлённые клатрином ямки, а затем - окаймлённые пузырьки, содержащие
комплекс «лиганд-рецептор». Окаймлённые пузырьки после освобождения от
клатрина - эндосома. Внутри эндосом лиганд отщепляется от рецептора. Погружение окаймлённого пузырька в цитоплазму (Б). Присоединение
к динамину ГТФ инициирует формирование механохимической молекулярной
пружины из плотно упакованных колец динамина с шагом в 11 нм вокруг
шейки эндоцитозного пузырька. При гидролизе ГТФ расстояние между
смежными кольцами увеличивается до 22 нм. Таким образом, ГТФаза динамина
катализирует расщепление ГТФ и тем самым растяжение пружины с
последующим отрывом окружённого клатрином эндоцитозного пузырька от
плазмолеммы. [17]
Фагоцитоз - поглощение крупных частиц (например, микроорганизмов или остатков клеток). Фагоцитоз осуществляют специальные клетки - фагоциты (макрофаги, нейтрофилы). В ходе фагоцитоза образуются большие эндоцитозные пузырьки - фагосомы. Фагосомы сливаются с лизосомами и формируют фаголизосомы. Фагоцитоз, в отличие от пиноцитоза, индуцирует сигналы, воздействующие на рецепторы в плазмолемме фагоцитов. Подобными сигналами служат АТ, опсонизирующие фагоцитируемую частицу (подробнее см. в разделе «Нейтрофилы» главы 6).
Экзоцитоз
Экзоцитоз (секреция) - процесс, когда внутриклеточные секреторные пузырьки (например, синаптические) и секреторные гранулы сливаются с плазмолеммой, а их содержимое освобождается из клетки (рис. 2-8Б). В ходе экзоцитоза можно выделить следующие последовательные стадии: перемещение везикулы в субплазмолеммальное пространство, установление связи и докинг (от англ. dock - стыковка) к участку плазмолеммы, слияние мембран, высвобождение содержимого гранулы (пузырька) и восстановление (обособление) мембраны гранулы.
• Мембранные пузырьки содержат вещества, подлежащие выведению из клетки (секреции, экзоцитозу). Такие пузырьки образуются в комплексе Гольджи.
• Гранулы - секреторные пузырьки с электронно-плотным содержимым, они присутствуют в хромаффинных клетках (катехоламины), тучных (гистамин) и некоторых эндокринных клетках (гормоны).
Конститутивная и регулируемая секреция. Процесс секреции может быть спонтанным и регулируемым. Одна часть пузырьков постоянно сливается с клеточной мембраной (конститутивная секреция), в то время как другая часть пузырьков накапливается под плазмолеммой, но процесс слияния пузырька и мембраны происходит только под действием сигнала, чаще всего вследствие увеличения концентрации Са2+ в цитозоле (регулируемый экзоцитоз).
Типы секреции
Типы секреции (мерокриновый, или эккриновый, апокриновый и голокриновый) рассмотрены в главе 5.
Трансцитоз
Трансцитоз - транспорт макромолекул через клетку, в ходе которого происходит быстрое и эффективное переключение эндоцитоза на экзоцитоз. Трансцитоз обычно осуществляется с участием кавеол. Кавеолы формируют дискретные пузырьки-переносчики, курсирующие между апикальной и базальной частями клетки, подвергаясь в каждом обороте (круге транспорта) процессу отрыва-слияния. Трансци-
тоз характерен, например, для эндотелиальных клеток, где происходит транспорт макромолекул через клетки из просвета сосуда в ткань.
Ядро
Ядро (см. рис. 2-22) состоит из хроматина, ядрышка и нуклеоплазмы, окружённых ядерной оболочкой. Хранение и реализация генетической информации (транскрипция → процессинг → трансляция → посттрансляционная модификация), а также ряд других функций ядра происходят при участии ДНК и разных видов РНК (рис. 2-15). Хроматин (рис. 2-17). Термином «хроматин» обозначают комплекс ядерной двуцепочечной ДНК с белками (гистоны, негистоновые белки). Хроматин представлен хроматиновыми волокнами диаметром 11 нм, состоящими из сферических структурных единиц - нуклеосом. Различают гетерохроматин и эухроматин.
• Гетерохроматин - транскрипционно неактивный и конденсированный хроматин интерфазного ядра. В световом микроскопе - базофильные глыбки, в электронном микроскопе - скопления плотных гранул. Гетерохроматин располагается преимущественно по периферии ядра и вокруг ядрышек, составляет 10% от общего хроматина. Типичный пример гетерохроматина - тельце Барра.
Тельце Барра. Во всех соматических клетках генетически женского организма одна из X-хромосом инактивирована и известна как половой хроматин (тельце Барра).
• Эухроматин - диспергированная часть хроматина, находится в более светлых участках ядра между глыбками гетерохроматина. Эухроматин составляет 90% от общего хроматина, из них 10% - транскрипционно активная часть.
Хромосомы (рис. 2-17) видны при митозе или мейозе, когда полностью конденсированный хроматин образует многочисленные плотно упакованные петли. Каждая хромосома содержит одну длинную двуцепочечную молекулу ДНК и ДНК-связывающие белки. Результатом взаимодействия ДНК с ДНК-связывающими белками является компактизация хроматина. Длина молекулы ДНК в составе одной хромосомы приблизительно составляет 4 см, тогда как длина метафазных хромосом равняется 4 мкм.
ДНК. Молекула ДНК построена из двух (смысловой и антисмысловой) полинуклеотидных цепей, кодирующих ядерный геном клетки. ДНК служит матрицей для синтеза РНК. При помощи фосфодиэфирных связей нуклеотиды образуют полинуклеотидную цепь. Последовательность нуклеотидов в цепи кодирует наследственную информацию. Геном. Различают ядерный и митохондриальный геномы. Ядерный геном - полный комплект генов в 46 хромосомах диплоидной клетки. Примерно 3 миллиарда пар оснований ДНК кодируют две копии примерно 24 000 генов, причём кодирующая часть ДНК занимает менее 5%.
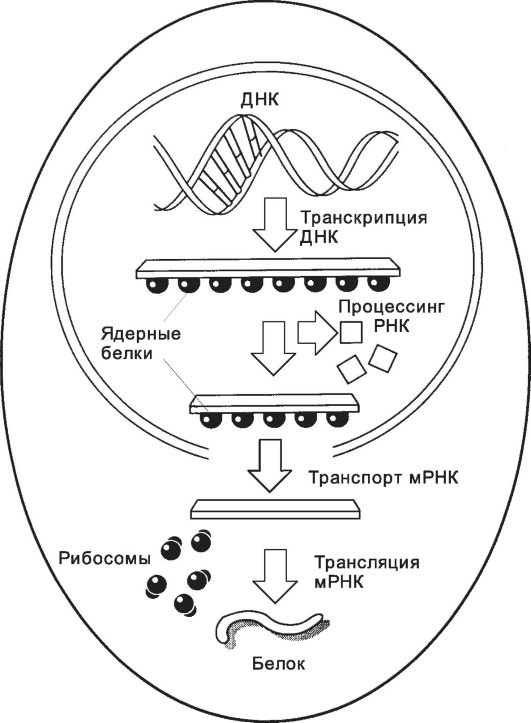 Рис. 2-15. Этапы считывания генетической информации. В
ходе транскрипции на ДНК-матрице синтезируется длинная молекула РНК
(первичный транскрипт), содержащая последовательности экзонов и
интронов. По завершении синтеза РНКтранскрипта последовательности
интронов удаляются, что делает молекулу РНК значительно короче. Эта мРНК
выходит из ядра в цитоплазму и соединяется с рибосомами. Молекула мРНК
продвигается сквозь рибосому, и её нуклеотидная последовательность
транслируется в соответствующую последовательность аминокислот
создаваемой белковой цепи. [17]
Рис. 2-15. Этапы считывания генетической информации. В
ходе транскрипции на ДНК-матрице синтезируется длинная молекула РНК
(первичный транскрипт), содержащая последовательности экзонов и
интронов. По завершении синтеза РНКтранскрипта последовательности
интронов удаляются, что делает молекулу РНК значительно короче. Эта мРНК
выходит из ядра в цитоплазму и соединяется с рибосомами. Молекула мРНК
продвигается сквозь рибосому, и её нуклеотидная последовательность
транслируется в соответствующую последовательность аминокислот
создаваемой белковой цепи. [17]
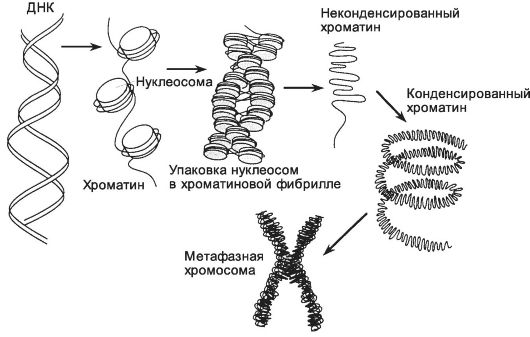 Рис. 2-17. Организация хромосомы. В неконденсированном хроматине двойная
спираль ДНК лежит на поверхности октамера гистонов (H2A, H2B, H3 и H4),
образуя хроматиновые волокна диаметром 11 нм. Смежные нуклеосомы
разделены интервалами в 200 пар оснований. В конденсированном хроматине дополнительно
присутствует гистон H1, соединяющий нуклеосомы с образованием
хроматиновых волокон диаметром 30 нм. Во время митоза хроматин полностью
конденсируется, формируя видимые хромосомы. [17]
Рис. 2-17. Организация хромосомы. В неконденсированном хроматине двойная
спираль ДНК лежит на поверхности октамера гистонов (H2A, H2B, H3 и H4),
образуя хроматиновые волокна диаметром 11 нм. Смежные нуклеосомы
разделены интервалами в 200 пар оснований. В конденсированном хроматине дополнительно
присутствует гистон H1, соединяющий нуклеосомы с образованием
хроматиновых волокон диаметром 30 нм. Во время митоза хроматин полностью
конденсируется, формируя видимые хромосомы. [17]
Ген - участок ДНК, ответственный за образование одной функциональной молекулы РНК. Экспрессия гена, кодирующего последовательность аминокислот в полипептидной цепи, протекает по схеме (см. рис. 2-15): транскрипция (синтез первичного транскрипта на матрице ДНК) → процессинг (образование мРНК) → трансляция (считывание информации с мРНК) → сборка полипептидной цепи (включение аминокислот в полипептидную цепь на рибосомах) → посттрансляционная модификация (добавление к полипептиду разных химических группировок, например, фосфатных [фосфорилирование], карбоксильных [карбоксилирование] и т.д.).
Репликация ДНК. Клетки перед каждым делением воспроизводят (реплицируют) ДНК: дочерние молекулы ДНК воспроизводятся при помощи ДНК-полимеразы одновременно в нескольких, т.н. точках начала репликации.
РНК - полинуклеотид, сходный по химическому составу с ДНК, но содержащий в нуклеотидах рибозу вместо дезоксирибозы и азотистое
основание урацил (U) вместо тимина (T). Различают мРНК, тРНК, рРНК. Синтез полимеров рРНК, мРНК и тРНК на матрице ДНК катализируют соответственно РНК-полимераза I, II и III. Матричная РНК (мРНК, информационная РНК) содержит сотни и тысячи нуклеотидов и переносит генетическую информацию из ядра в цитоплазму и непосредственно участвует в сборке полипептида на рибосомах (трансляция).
Транспортная РНК (тРНК) содержит около 80 нуклеотидов и доставляет аминокислоты к рибосоме, где они присоединяются к растущей полипептидной цепи. Существует минимально одна тРНК для каждой из 20 аминокислот. Таким образом, тРНК переводит последовательность нуклеотидов в последовательность аминокислот. Рибосомная РНК (рРНК) взаимодействует с мРНК и тРНК в ходе сборки полипептида, в комплексе с белками (в т.ч. ферментами) образует рибосому.
Транскрипция (синтез молекул мРНК на матричной ДНК) - первый этап реализации генетической информации в клетке. В результате образуется РНК-копия в виде первичного транскрипта. В дальнейшем он процессируется, в результате образуется зрелая мРНК, выходящая из ядра в цитоплазму через ядерные поры.
Ядрышко
Ядрышко - неокружённая мембраной компактная структура (см. рис. 2-22) в ядре интерфазных клеток, содержащая петли ДНК хромосом 13, 14, 15, 21 и 22. В ядрышке различают фибриллярный центр - слабоокрашенный компонент, содержащий транскрипционно неактивную ДНК; плотный фибриллярный компонент (pars fibrosa) состоит из транскрипционно активных участков ДНК, от которых перпендикулярно отходят различной длины 45S-предшественники рРНК; гранулярный компонент (pars granulosa), содержит зрелые предшественники рибосомных субъединиц. Основные функции ядрышка - синтез рРНК (транскрипция и процессинг рРНК) и образование субъединиц рибосом.
Ядерная оболочка
В состав ядерной оболочки (рис. 2-21) входят наружная и внутренняя ядерные мембраны, перинуклеарные цистерны, ядерная пластинка, ядерные поры.
• Наружная ядерная мембрана. На её поверхности расположены рибосомы, где синтезируются белки, поступающие в перинуклеарную цистерну, рассматриваемую как часть гранулярной эндоплазматической сети.
• Внутренняя ядерная мембрана снаружи граничит с перинуклеарной цистерной, изнутри отделена от содержимого ядра ядерной пластинкой.
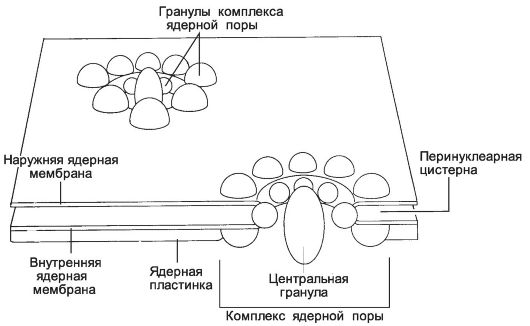 Рис. 2-21. Поры в оболочке ядра. Комплекс
ядерной поры образован 8 большими белковыми гранулами, расположенными
по окружности вблизи края поры и соединяющими обе ядерные мембраны
(внутреннюю и наружную). Часто в центре поры присутствует большая
центральная гранула. Она состоит из вновь синтезированной субъединицы
рибосомы, переносимой в цитоплазму. [17]
Рис. 2-21. Поры в оболочке ядра. Комплекс
ядерной поры образован 8 большими белковыми гранулами, расположенными
по окружности вблизи края поры и соединяющими обе ядерные мембраны
(внутреннюю и наружную). Часто в центре поры присутствует большая
центральная гранула. Она состоит из вновь синтезированной субъединицы
рибосомы, переносимой в цитоплазму. [17]
• Ядерная пластинка толщиной 80-300 нм содержит белки промежуточных филаментов - ламины A, B и C, участвует в организации ядерной оболочки и перинуклеарного хроматина, может разделять комплексы ядерных пор и дезинтегрировать или интегрировать структуру ядерной оболочки в ходе митоза.
• Ядерная пора. Содержимое ядра сообщается с цитозолем через 3-4 тысячи специализированных коммуникаций - ядерных пор, осуществляющих диффузию воды, ионов и транспорт множества макромолекул между ядром и цитоплазмой. Ядерная пора (см. рис. 2-21) имеет диаметр 80-150 нм, содержит канал поры и комплекс ядерной поры. Перенос макромолекул через ядерные поры осуществляют специальные транспортные белки - кариоферины, которые специфически распознают и связывают конкретные молекулы и курсируют между ядром и цитоплазмой, перенося связанную молекулу в одном направлении: из цитоплазмы в ядро (импортины) или из ядра в цитоплазму (экспортины).
♦ Канал поры диаметром 9 нм беспрепятственно пропускает небольшие водорастворимые молекулы.
♦ Комплекс ядерной поры образован 8 большими белковыми гранулами, сформированных примерно из 100 разных белков. Эти белки реагируют на сигналы
ядерного импорта (своего рода входной билет в ядро). Комплекс ядерной поры включает цитоплазматическое кольцо, нуклеоплазматическое кольцо и среднее кольцо.
Нуклеоплазма
Нуклеоплазма заключена в ядерную оболочку, состоит из ядерного матрикса и разных ядерных частиц.
• Ядерный матрикс. Фибриллярные элементы ядра и рибонуклеопротеиновая сеть формируют матрикс, в который погружены ядерные рецепторы, ферменты (АТФаза, ГТФаза, ДНК- и РНК-полимеразы) и множество других молекул, часто образующих ассоциации - ядерные частицы. На транскрипцию и процессинг РНК в матриксе влияют ядерные рецепторы, канцерогены, транскрипционные факторы, белки теплового шока, вирусы.
• Ядерные частицы содержат интерхроматиновые и полихроматиновые гранулы, ядерные рибонуклеопротеиновые частицы и ядерные рецепторы.
Цитоплазма
Жидкая часть цитоплазмы (цитозоль), составляет около половины объ- ёма клетки. Помимо воды, в цитозоле присутствуют ионы, множество химических соединений разной природы, макромолекулы. Цитозоль содержит цитоскелет, органеллы и включения. Органелла (органоид) - специализированный для выполнения конкретной функции и метаболически активный элемент цитоплазмы (рис. 2-22). К органеллам относят свободные рибосомы, гранулярную эндоплазматическую сеть (шероховатый эндоплазматический ретикулум), гладкую эндоплазматическую сеть (гладкий эндоплазматический ретикулум), митохондрии, комплекс Гольджи, центриоли, окаймлённые пузырьки, лизосомы, пероксисомы, а к макромолекулярным комплексам цитозоля относят апоптосомы, формируемые при программированной гибели клеток, а также протеосомы - комплексы нелизосомных протеаз, осуществляющие деградацию белков. Трёхмерная цитоплазматическая сеть волокнистых и трубчатых структур различного типа формирует цитоскелет.
ЦИТОСКЕЛЕТ
К элементам цитоскелета относят микротрубочки, микрофиламенты и промежуточные филаменты. Цитоскелет придаёт клетке определённую форму и выполняет множество других функций (например, обеспечивает подвижность клетки и внутриклеточный транспорт).
Микротрубочки
Микротрубочки состоят из 13 параллельных тубулиновых протофиламентов (нитей), образующих полые цилиндры диаметром 25 нм и длиной в несколько микрометров (рис. 2-24). Каждая нить собрана
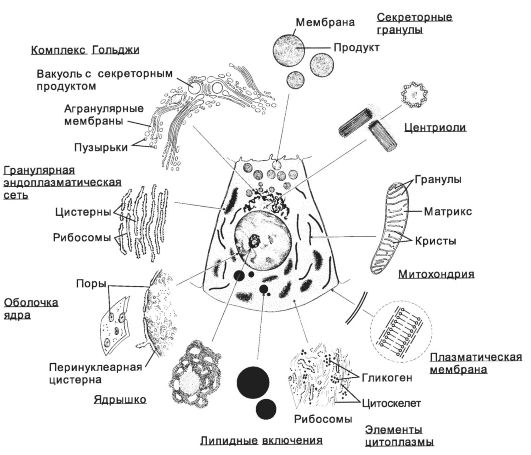 Рис. 2-22. Органеллы и включения. [17]
Рис. 2-22. Органеллы и включения. [17]
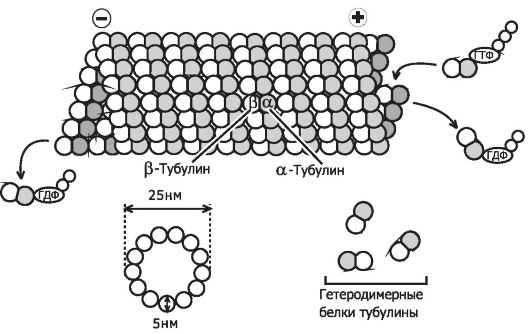 Рис. 2-24. Микротрубочка. 13
параллельно расположенных протофиламентов состоят из отдельных
субъединиц - димеров α- и β-тубулина. 13 параллельно расположенных
тубулиновых протофиламентов формируют полый цилиндр диаметром 25 нм.
Протофиламенты образуются путём полимеризации гетеродимерного белка
тубулина, состоящего из глобулярных субъединиц - α- и β-тубулина. [114]
Рис. 2-24. Микротрубочка. 13
параллельно расположенных протофиламентов состоят из отдельных
субъединиц - димеров α- и β-тубулина. 13 параллельно расположенных
тубулиновых протофиламентов формируют полый цилиндр диаметром 25 нм.
Протофиламенты образуются путём полимеризации гетеродимерного белка
тубулина, состоящего из глобулярных субъединиц - α- и β-тубулина. [114]
из гетеродимерного белка тубулина, состоящего из двух глобулярных субъединиц - α- и β-тубулина. Сборка микротрубочек осуществляется в т.н. центре организации микротрубочек в центросоме. Микротрубочки - динамичные структуры, постоянно подвергающиеся полимеризации и деполимеризации.
• Полимеризация и деполимеризация микротрубочек. Удлинение (рост) микротрубочек происходит за счёт полимеризации молекул тубулина. В каждой микротрубочке различают (+)-конец и (-)-конец. Микротрубочки постоянно подвергаются полимеризации и деполимеризации с (+)-конца, тогда как с противоположного (-)-конца (если он не занят стабилизирующим белком) тубулиновые гетеродимеры отделяются от микротрубочек. Как только прекращается добавление новых димеров к растущему концу, в этом месте сразу начинается разборка полимера. Повторяющиеся раунды полимеризации и деполимеризации характеризуют динамическую нестабильность микротрубочек. Цитозольные белки, способные связываться с концами микротрубочек и стабилизировать их, относят к семейству ассоциированных с микротрубочками белков.
• Функции микротрубочек. Микротрубочки участвуют в поддержании формы клетки, антероградном и ретроградном аксоном транспорте макромолекул, органелл и секреторных везикул, фагоцитозе и функции лизосом. Микротрубочки образуют аксонемы и базальные тельца, обеспечивая подвижность жгутиков и ресничек, в составе центриолей они обеспечивают расхождения хромосом при делении клеток.
• Молекулярные моторы. Применительно к микротрубочкам под этим термином понимают АТФазы (динеины и кинезины), одним доменом связывающиеся с тубулином микротрубочек, а другим - с различными мембранными органеллами (митохондриями, секреторными везикулами из комплекса Гольджи, элементами эндоплазматической сети, эндоцитозными пузырьками, аутофагосомами) или макромолекулами. За счёт расщепления АТФ моторные белки перемещаются вдоль микротрубочек и таким образом транспортируют ассоциированные с ними органеллы и макромолекулы. При этом кинезиновый мотор направлен к (+)-концу, а динеиновый - к (-)-концу микротрубочки (см. рис. 2-28).
♦ Тубулин-кинезиновый хемомеханический преобразователь обеспечивает внутриклеточный транспорт органелл и перемещение хромосом вдоль микротрубочек в ходе клеточного деления. Перемещение органелл вдоль микротрубочек с участием кинезинов осуществляется в направлении (+)-конца микротрубочек.
♦ Тубулин-динеиновый хемомеханический преобразователь отвечает за направленный транспорт макромолекул и органелл к (-)-концу микротрубочек. В составе аксонемы тубулиновый молекулярный мотор приводит в движение жгутик сперматозоида и реснички мерцательных клеток.
• Аксонема (рис. 2-25, рис. 2-26) состоит из 9 периферических пар микротрубочек и двух расположенных центрально одиночных микротрубочек. В каждой периферической паре различают субфибриллу А, содержащую 10-11 тубулиновых протофиламентов, и субфибриллу В, содержащую 13 протофиламентов. Смежные пары микротрубочек соединены между собой эластичным белком нексином. С субфибриллой А связаны наружные и внутренние ручки. В их состав входит белок динеин, содержащий 2-3 глобулярные головки, соединённые с гибкой фибриллярной частью молекулы. Основание фибриллярной части вплетено в микротрубочку (субфибрилла A). Глобулярная головка обладает АТФазной активностью. При расщеплении АТФ она скользит по поверхности микротрубочки (субфибрилла B) соседней пары по направлению к её (-)-концу. Этот механизм аналогичен скольжению элементов актомиозинового хемомеханического преобразователя в мышце. Аксонема - основной структурный элемент реснички и жгутика.
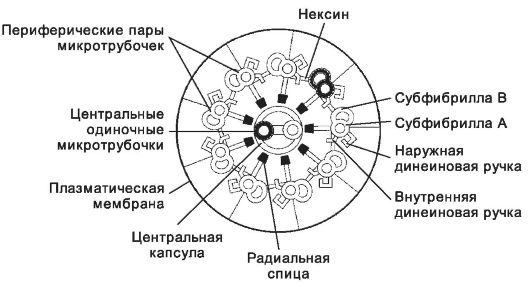 Рис. 2-25. Аксонема состоит из комплекса микротрубочек и связанных с ними белков.
Рис. 2-25. Аксонема состоит из комплекса микротрубочек и связанных с ними белков.
9 пар микротрубочек расположено по окружности, одна пара находится в центре. Каждая периферическая пара образована субфибриллой A и субфибриллой B. Субфибриллы состоят из протофиламентов. Обладающий АТФазной активностью белок динеин - компонент тубулин-динеинового хемомеханического преобразователя - входит в состав ручек, связанных с субфибриллой A. [17]
• Базальное тельце (рис. 2-26) состоит из 9 триплетов микротрубочек, расположенных в основании реснички или жгутика; служит матрицей при организации аксонемы.
• Ресничка - вырост клетки длиной 5-10 мкм и шириной 0,2 мкм, содержащий аксонему (рис. 2-26). Реснички присутствуют в эпителиальных клетках воздухопроводящих и половых путей, перемещают слизь с инородными частицами и остатками отмерших клеток и создают ток жидкости около клеточной поверхности.
• Жгутик, как правило, не встречается в количестве более двух на клетку. В сперматозоиде человека жгутик имеет длину 50-55 мкм, толщину 0,2-0,5 мкм и содержит аксонему.
• Киноцилия - (греч. kinesis, движение; cilium, ресничка) специальная органелла подвижности на поверхности волосковых клеток органа равновесия (см. рис. 8-65).
Микрофиламенты
Две переплетённые нити F-актина (фибриллярного актина), составленные из G-актина (глобулярного актина), формируют микрофиламенты диаметром 6-8 нм. В комплексе с актин-связывающими белками актиновые филаменты образуют различные внутриклеточные структуры (тонкие нити миофибрилл, кортикальный примембранный скелет, опоясывающую десмосому, микроворсинки, стереоцилии).
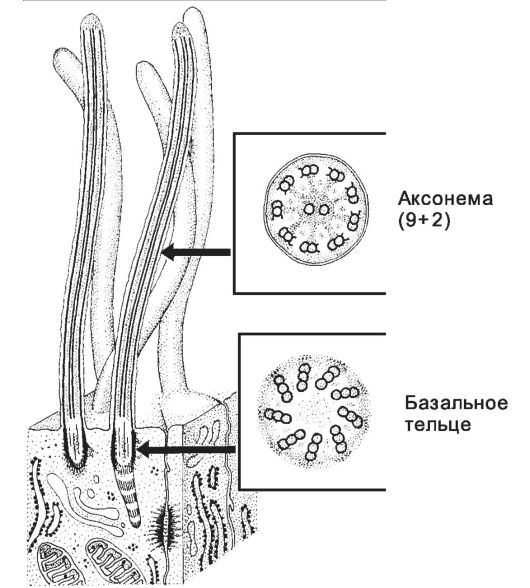 Рис. 2-26. Ресничка - тонкий вырост на поверхности клетки. Стержень
реснички образован аксонемой - системой микротрубочек 9+2. В основании
реснички расположено базальное тельце, служащее матрицей для
формирования аксонемы. [17]
Рис. 2-26. Ресничка - тонкий вырост на поверхности клетки. Стержень
реснички образован аксонемой - системой микротрубочек 9+2. В основании
реснички расположено базальное тельце, служащее матрицей для
формирования аксонемы. [17]
Как и микротрубочки, микрофиламенты полярны; присоединение (полимеризация) новых молекул G-актина происходит на (+)-конце, а деполимеризация (разборка полимера) на (-)-конце микрофиламента. Известны токсины (например, цитохалазины, фаллоидин), связывающиеся с актином и блокирующие его полимеризацию, нарушая тем самым подвижность клеток, фагоцитоз и цитокинез. Микрофиламенты принимают непосредственное участие в образовании микроворсинок клетки.
• Микроворсинки (рис. 2-31). Пучок параллельных микрофиламентов образует сердцевину микроворсинок высотой 1 мкм. Каждый энтероцит (осуществляющая всасывание эпителиальная клетка тонкой кишки) содержит более 1000 микроворсинок, которые увеличивают площадь апикальной поверхности клетки в 20 раз.
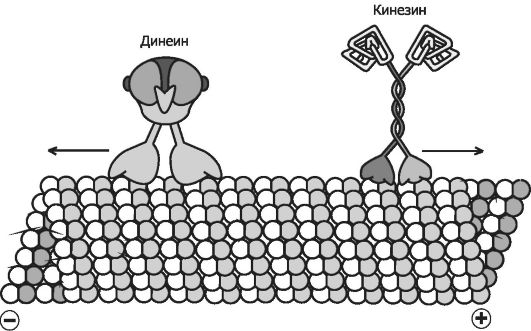 Рис. 2-28. Молекулярные моторы. Тубулин-кинезиновый
хемомеханический преобразователь. Двигательные белки молекулярных
моторов (динеин и кинезин) - ферменты, преобразующие энергию АТФ в
механическую работу. Кинезиновый мотор обеспечивает транспорт органелл к
(+)-концу, а динеиновый - к (-)-концу микротрубочек. [114]
Рис. 2-28. Молекулярные моторы. Тубулин-кинезиновый
хемомеханический преобразователь. Двигательные белки молекулярных
моторов (динеин и кинезин) - ферменты, преобразующие энергию АТФ в
механическую работу. Кинезиновый мотор обеспечивает транспорт органелл к
(+)-концу, а динеиновый - к (-)-концу микротрубочек. [114]
• Стереоцилии (см. рис. 8-57, рис. 15-16). На поверхности эпителиальных клеток придатка яичка и семявыносящего протока, волосковых клеток органа слуха и равновесия имеются высокие микроворсинки, известные как стереоцилии.
• Функции микрофиламентов крайне разнообразны: участие в процессах мышечного сокращения, подвижности немышечных клеток, эндоцитоза, экзоцитоза, изменения консистенции цитозоля, стабилизации локальных выпячиваний плазматической мембраны, цитокинезе.
Промежуточные нити
Нити цитоскелета диаметром 8-12 нм названы промежуточными, поскольку по величине диаметра они находятся между микротрубочками и микрофиламентами. Промежуточные нити (филаменты) состоят из белков, специфичных для определённых клеточных типов. Различают цитокератины эпителия, десмины мышц, виментины мезенхимы, ламины ядерной оболочки, а также нейрофиламенты, глиофиламенты и чётковидные волокна хрусталика.
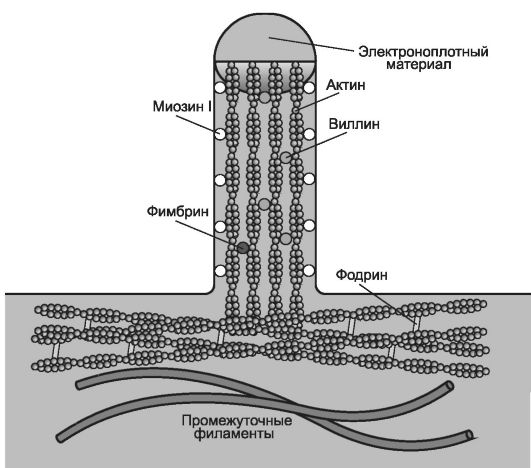 Рис. 2-31. Организация микроворсинки в апикальной части каёмчатой клетки. Около
30 параллельно идущих микрофиламентов образуют стержень микроворсинки.
(+)-Концы двух переплетённых нитей F-актина микрофиламентов направлены к
вершине микроворсинки. Микрофиламенты сшивают актин-связывающие белки
фимбрин и виллин. Микрофиламенты присоединены к внутренней поверхности
плазматической мембраны при помощи миозина I. В основании микроворсинок
актиновые нити (-)-концами заякорены в терминальную сеть - примембранное сплетение микрофиламентов сшитых между собой фодрином. [114]
Рис. 2-31. Организация микроворсинки в апикальной части каёмчатой клетки. Около
30 параллельно идущих микрофиламентов образуют стержень микроворсинки.
(+)-Концы двух переплетённых нитей F-актина микрофиламентов направлены к
вершине микроворсинки. Микрофиламенты сшивают актин-связывающие белки
фимбрин и виллин. Микрофиламенты присоединены к внутренней поверхности
плазматической мембраны при помощи миозина I. В основании микроворсинок
актиновые нити (-)-концами заякорены в терминальную сеть - примембранное сплетение микрофиламентов сшитых между собой фодрином. [114]
Поскольку экспрессия белков промежуточных нитей специфична для определён- ных клеточных типов, для цитодиагностики генеза опухолей в клинико-лабораторной практике используют иммуноцитохимические реакции с АТ против белков конкретных типов промежуточных нитей.
Функции. Промежуточные нити создают внутриклеточный каркас, обеспечивают упругость клетки, поддерживают упорядоченность расположения компонентов цитоплазмы, координируют связи между внеклеточным веществом, цитоплазмой и ядром.
РИБОСОМЫ
Рибосома состоит из большой и малой субъединиц (СЕ), содержащих различные типы рРНК и белки. Малая СЕ связывается с мРНК и активированными тРНК. Функция рибосом - трансляция (считывание кода мРНК и сборка полипептидов). Рибосомы подразделяют на митохондриальные и более крупные цитоплазматические.
• Митохондриальные рибосомы (60S) состоят из 45S и 35S СЕ, содержащих соответственно 16S и 12S рРНК, кодируемые митохондриальной ДНК. Эта ДНК содержит последовательности для митохондриальных мРНК и тРНК. Большинство ферментов, участвующих в трансляции митохондриальной мРНК, кодируется ядерной ДНК.
• Цитоплазматические рибосомы - небольшие электронно-плотные частицы размером 12x25 нм (80S), состоят из синтезируемой в ядрышке рРНК и белков. Цитоплазматические рибосомы подразделяют на свободные и связанные с мембранами эндоплазматической сети (поэтому такая эндоплазматическая сеть и называется гранулярной) и наружной ядерной мембраной. Ассоциаты рибосом формируют полирибосомы.
• Полирибосома (полисома) - комплекс нескольких рибосом, расположенных на одной молекуле мРНК. Полирибосомы, как и отдельные рибосомы, находятся в цитоплазме в свободном состоянии или прикреплены к мембранам эндоплазматической сети. Свободные полирибосомы синтезируют белки и ферменты для самой клетки (конститутивный синтез), а полирибосомы гранулярной эндоплазматической сети - предназначенные для хранения или выведения из клетки (синтез на экспорт).
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ
В эндоплазматической сети вырабатывается, процессируется и транспортируется множество веществ, которые используются клеткой или выделяются из неё. Различают гранулярную (зернистую, шероховатую) и гладкую эндоплазматическую сеть (ретикулум). Цистерны гранулярной и гладкой эндоплазматической сети не сообщаются. Клетки, специализированные на выработку белка, имеют более развитую гранулярную эндоплазматическую сеть. Клетки, продуцирующие липиды и стероидные гормоны, содержат выраженную гладкую эндоплазматическую сеть.
Функции эндоплазматической сети: ❖ поставка липидов другим органеллам (гладкая); ❖ гомеостаз Ca2+ (гладкая); ❖ биогенез органелл (гранулярная); ❖ формирование пространственной (трёхмерной) структуры (укладки) белков (гранулярная); ❖ посттрансляционный контроль качества белка (гранулярная).
Гранулярная эндоплазматическая сеть
Гранулярная эндоплазматическая сеть - система плоских мембранных цистерн с находящимися на их наружной поверхности рибосомами (см. рис. 2-22). В шероховатой эндоплазматической сети происходит синтез белков для плазматической мембраны, лизосом, пероксисом, а также синтез белков на экспорт, т.е. предназначенных для секреции. Мембраны гранулярной эндоплазматической сети связаны с наружной мембраной оболочки ядра и перинуклеарной цистерной. Гранулярная эндоплазматическая сеть располагается в непосредственной близости от ядра и комплекса Гольджи. Она участвует в синтезе и процессинге белков, преимущественно предназначенных для выделения из клетки. Рибосомы при помощи рибофоринов связаны с наружной (обращённой в цитозоль) поверхностью сети. Их количество (например, в гепатоците) достигает 13 млн. Собранные на рибосомах белки поступают внутрь цистерны для последующего процессинга. Концентрация белка здесь может превышать 100 мг/мл. Здесь же происходит укладка белков и формирование правильной трёхмерной структуры. В цистернах сети к белкам присоединяются углеводы с образованием гликопротеинов, а также формируются белковые комплексы с металлами. Из эндоплазматической сети многие белки поступают во все компартменты клетки для выполнения своих функций или направляются в комплекс Гольджи для последующей модификации. Резидентные белки и шапероны. Наряду с покидающими сеть белками, имеются резидентные белки, которые постоянно присутствуют в просвете цистерн и нужны для поддержания функции сети, а именно для узнавания образованных здесь белков, их процессирования и удержания в течение необходимого времени до отправления их по нужному адресу. Примером резидентных белков может служить белок BiP - шаперон иммуноглобулин-связывающего белка, принадлежащий семейству белков теплового шока Hsp70. В контроле качества белка участвуют шапероны. В белковом матриксе эндоплазматической сети шапероны предотвращают агрегацию белков и делают возможной эффективную их укладку.
Гладкая эндоплазматическая сеть
Гладкий ретикулум (гладкий ЭР) - система анастомозирующих мембранных каналов, пузырьков и трубочек - не содержит рибофоринов и по этой причине не связан с рибосомами.
Функции гладкой эндоплазматической сети многообразны: синтез липидов и стероидных гормонов, детоксикация и депонирование ионов кальция.
• Детоксикация. Одной из наиболее важной функцией гладкого ЭР является детоксикация (при помощи оксидаз гепатоцитов) как продуктов клеточного метаболизма, так и поступающих извне веществ, в том числе этанола и барбитуратов. С участием гладкого ЭР вещества конвертируются в водорастворимые соединения, что способствует их выведению из организма. Для эффективной детоксикации гладкий ЭР в течение нескольких дней может удвоить общую площадь своей поверхности.
• Синтез стероидных гормонов. В стероидпродуцирующих клетках (кора надпочечников, половые железы) гладкий ЭР служит для метаболизма стероидов и образования (при участии митохондрий) конечных форм стероидных гормонов.
• Депо кальция. Цистерны гладкой эндоплазматической сети многих клеток специализированы для накопления в них Ca2+ путём постоянного откачивания Ca2+ из цитоплазмы, где нормально содержание Ca2+ не превышает 10-7 М. Подобные депо существуют в скелетной и сердечной мышцах, нейронах, хромаффинных клетках, яйцеклетке, эндокринных клетках и т.д. Различные сигналы (например, гормоны, нейромедиаторы, факторы роста) влияют на функции клеток путём изменения концентрации в цитозоле внутриклеточного посредника - Ca2+. Например, условие сокращения мышечных элементов - резкое повышение концентрации Ca2+ в цитозоле. Для этого необходимо постоянно откачивать ионы кальция из цитозоля и накапливать их в специальных депо, образованных Ca2+-запасающими цистернами гладкой эндоплазматической сети. Внутри цистерн находятся Ca2+-связывающие белки. В мембрану цистерн - депо Ca2+ встроены Ca2+-насосы (Ca2+-АТФаза), постоянно закачивающие Ca2+ внутрь цистерн, и Ca2+-каналы, через которые происходит выброс Ca2+ из депо при поступлении сигнала.
МИТОХОНДРИИ
Митохондрии - преобразователи энергии и её поставщики для обеспечения клеточных функций - занимают значительную часть цитоплазмы клеток и сосредоточены в местах высокого потребления АТФ (например, в эпителии канальцев почки они располагаются вблизи плазматической мембраны [обеспечение реабсорбции], а в нейронах - в синапсах [обеспечение электрогенеза и секреции]). Количество митохондрий в клетке измеряется сотнями. Митохондрии имеют собственный геном. Органелла функционирует в среднем 10 суток, обновление митохондрий происходит путём их деления.
Морфология митохондрий
Митохондрии (рис. 2-39) чаще имеют форму цилиндра диаметром 0,2-1 мкм и длиной до 7 мкм (в среднем около 2 мкм). У митохондрий две мембраны - наружная и внутренняя; последняя образует кристы. Между наружной и внутренней мембранами находится межмембранное пространство. Внемембранный объём митохондрии - матрикс.
• Наружная мембрана проницаема для многих мелких молекул.
• Межмембранное пространство. Здесь накапливаются ионы H+, выкачиваемые из матрикса, что создаёт протонный градиент концентрации по обе стороны внутренней мембраны.
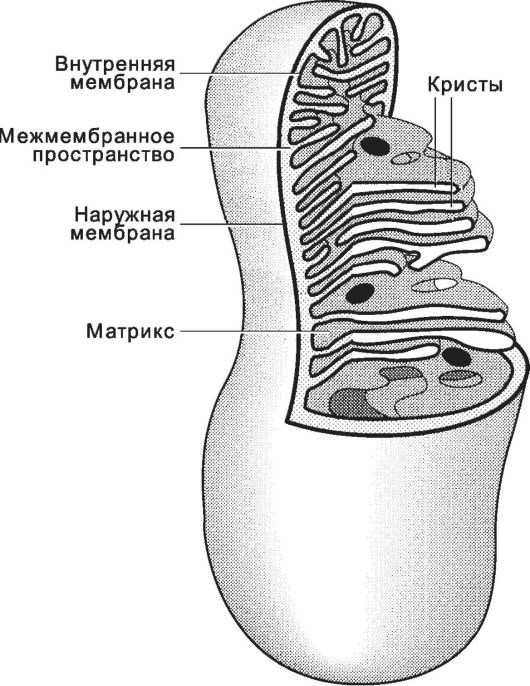 Рис. 2-39. Митохондрия. Органелла
содержит наружную и внутреннюю мембраны с узким межмембранным
пространством. Внутренняя мембрана образует многочисленные выросты -
кристы, окружённые матриксом. В матриксе находятся ДНК, ферменты и
гранулы. |17|
Рис. 2-39. Митохондрия. Органелла
содержит наружную и внутреннюю мембраны с узким межмембранным
пространством. Внутренняя мембрана образует многочисленные выросты -
кристы, окружённые матриксом. В матриксе находятся ДНК, ферменты и
гранулы. |17|
• Внутренняя мембрана избирательно проницаема; содержит транспортные системы для переноса веществ (АТФ, АДФ, Pi, пирувата, сукцината, α-кетоглутарата, малата, цитрата, цитидинтрифосфата, ГТФ, дифосфатов) в обоих направлениях и комплексы цепи переноса электронов, связанные с ферментами окислительного фосфорилирования, а также с сукцинатдегидрогеназой (СДГ).
• Матрикс. В матриксе присутствуют все ферменты цикла Кребса (кроме СДГ), ферменты β-окисления жирных кислот и некоторые ферменты других систем. В матриксе находятся гранулы с Mg2+ и Ca2+.
• Цитохимические маркёры митохондрий - цитохромоксидаза и СДГ.
Функции митохондрий
Митохондрии выполняют в клетке множество функций: окисление в цикле Кребса, транспорт электронов, хемиосмотическое сопряжение, фосфорилирование АДФ, сопряжение окисления и фосфорилирования, функцию контроля внутриклеточной концентрации кальция,
синтез белков, образование тепла. Велика роль митохондрий в программированной (регулируемой) гибели клеток.
• Теплопродукция. Естественный механизм разобщения окислительного фосфорилирования функционирует в клетках бурого жира. В этих клетках митохондрии имеют атипичную структуру (уменьшен их объём, увеличена плотность матрикса, расширены межмембранные пространства) - конденсированные митохондрии. Такие митохондрии могут усиленно захватывать воду и набухать в ответ на тироксин, увеличение концентрации Ca2+ в цитозоле, при этом усиливается разобщение окислительного фосфорилирования, и происходит выделение тепла. Эти процессы обеспечивает специальный разобщающий белок термогенин. Норадреналин из симпатического отдела вегетативной нервной системы усиливает экспрессию разобщающего белка и стимулирует теплопродукцию.
• Апоптоз. Митохондрии играют важную роль в регулируемой (программированной) гибели клеток - апоптозе, выделяя в цитозоль факторы, повышающие вероятность гибели клетки. Одним из них является цитохром C - белок, переносящий электроны между белковыми комплексами во внутренней мембране митохондрий. Выделяясь из митохондрий, цитохром C включается в состав апоптосомы, активирующей каспазы (представители семейства киллерных протеаз).
КОМПЛЕКС ГОЛЬДЖИ
Комплекс Гольджи расположен около ядра и образован стопкой из 3-10 уплощённых и слегка изогнутых цистерн с расширенными концами (рис. 2-43). Эта органелла процессирует белки, синтезированные в гранулярной эндоплазматической сети. Выходящие из цистерн комплекса Гольджи белки находятся внутри транспортных пузырьков. Продукты комплекса Гольджи разделяются на три потока, которые направляются в плазматическую мембрану (интегральные мембранные белки), лизосомы или выделяются из клетки (например, пищеварительные ферменты).
Цистерны комплекса Гольджи образуют три основных компартмента: цис-компартмент, транс-компартмент и промежуточный компартмент. С комплексом Гольджи тесно связан и всегда рассматривается вместе ещё один компартмент - транс-сеть Гольджи.
• Цис-компартмент (формирующийся) включает цистерны, обращён- ные к расширенным элементам гранулярной эндоплазматической сети, а также небольшие транспортные пузырьки.
• Транс-компартмент (зрелый) образован цистернами, обращёнными к вакуолям и секреторным гранулам. На небольшом расстоянии от краевой цистерны транс-компартмента лежит транс-сеть.
• Промежуточный компартмент - небольшое количество цистерн между цис- и транс-компартментами.
• Транс-сеть Гольджи лежит на небольшом расстоянии от краевой цистерны транс-компартмента и участвует в образовании лизосом и сортировке белков для различных транспортных пузырьков.
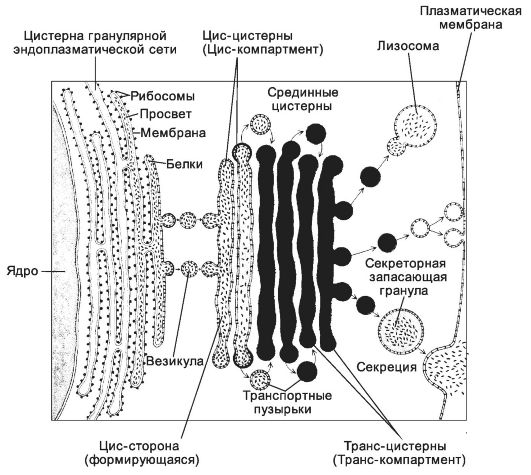 Рис. 2-43. Комплекс Гольджи. Белки
и липиды поступают в комплекс Гольджи через цис-компартмент.
Транспортные пузырьки переносят эти молекулы последовательно из одной
цистерны в другую. Готовый продукт выходит из комплекса через
транс-компартмент, находясь в различных пузырьках. Часть из них содержит
молекулы, обеспечивающие внутриклеточное пищеварение, и сливается с
лизосомами. Другая часть пузырьков участвует в экзоцитозе. Третья группа
пузырьков содержит белки для плазмолеммы. [17]
Рис. 2-43. Комплекс Гольджи. Белки
и липиды поступают в комплекс Гольджи через цис-компартмент.
Транспортные пузырьки переносят эти молекулы последовательно из одной
цистерны в другую. Готовый продукт выходит из комплекса через
транс-компартмент, находясь в различных пузырьках. Часть из них содержит
молекулы, обеспечивающие внутриклеточное пищеварение, и сливается с
лизосомами. Другая часть пузырьков участвует в экзоцитозе. Третья группа
пузырьков содержит белки для плазмолеммы. [17]
Компартменты и направление транспорта. Белки, синтезированные в гранулярной эндоплазматической сети, поступают в специальный компартмент, который располагается между эндоплазматической сетью и комплексом Гольджи. Этот компартмент представлен т.н. везикулярно-трубчатыми кластерами. Здесь происходит умеренное накопление белков и определяется направление их дальнейшего транспорта. Из этого компартмента материал для дальнейшей модификации проходит по следующей цепочке: цис-компартмент → промежуточный компартмент → транскомпартмент → транс-сеть Гольджи. Это направление транспорта продукта обозначается как антероградное. Перемещение продукта по данной цепочке в обратном направлении называют ретроградным транспортом. Некоторые белки после мо-
дификации в комплексе Гольджи транспортируются ретроградно в гранулярную эндоплазматическую сеть для выполнения там своих функций. Функции комплекса Гольджи. В комплексе Гольджи происходят процессы модификации белка (например, гликозилирование, т.е. присоединение к молекуле белка молекул сахаров), концентрирования, сортировки и упаковки секреторных продуктов (образование секреторных гранул).
ПРОТЕОСОМЫ
Содержание белков в клетке определяется не только скоростью их синтеза, но также и скоростью их деградации. Время полураспада клеточных белков варьирует от нескольких минут до нескольких дней. Существуют и дефектно синтезированные или повреждённые белки, подлежащие идентификации и быстрому удалению. Деградация белков (цитозольный протеолиз) происходит по двум основным механизмам - убиквитин-протеосомный путь и опосредованная лизосомным протеолизом деградация.
• Убиквитин. Этот полипептид присутствует во всех клетках всех организмов, кроме бактерий, что и определило его название (лат. ubique - везде). Клетки используют убиквитин в качестве маркёра нежелательных белков, которые после взаимодействия с убиквитином легко распознаются и быстро расщепляются в специальных органеллах - протеосомах.
• Протеосома - комплекс нелизосомных мультикаталитических протеиназ. Протеосомы осуществляют значительную часть цитозольного протеолиза, поэтому протеосомы регулируют множество важных внутриклеточных процессов: контроль клеточного цикла, выживаемость клеток, опухолевый рост, ангиогенез, экспрессия молекул адгезии, продукция цитокинов, удаление повреждённых, окисленных и мутантных белков.
ЛИЗОСОМЫ
Лизосомы - окружённые мембраной округлые пузырьки, содержащие кислые гидролазы. В широком смысле функция лизосом заключается в утилизации «внутриклеточного мусора». Лизосомы образуются путём слияния так называемых поздних эндосом, содержащих лизосомные гидролазы, с везикулами, подлежащими деградации (например, фагосомой).
Поздние эндосомы (или перинуклеарные эндосомы) образуются при слиянии содержащих лизосомные гидролазы везикул и везикул, в мембрану которых встроены специфические лизосомные белки.
Различают следующие типы лизосом: мультивезикулярные тельца, фаголизосомы, аутофаголизосомы, остаточные тельца.
• Мультивезикулярные тельца образуются при слиянии перинуклеарной и периферической эндосом.
• Фаголизосома образуется при слиянии перинуклеарной эндосомы и фагосомы, содержащей экзогенный материал.
• Аутофаголизосома образуется при слиянии перинуклеарной эндосомы и аутофагоцитозной вакуоли, содержащей подлежащие деградации эндогенные молекулы и органеллы.
• Остаточные тельца - лизосомы любого типа, содержащие непереваренный материал (липофусцин, гемосидерин).
Известно более 50 лизосомных ферментов: рибонуклеазы, дезоксирибонуклеазы, катепсины B и L (протеазы), сульфатазы, β-глюкуронидазы, β-галактозидазы, гликозидазы, липазы, эстеразы, фосфатазы и другие. Лизосомные ферменты наиболее активны в кислой среде (pH 5,0), для поддержания которой в мембрану лизосомы встроен протонный насос (Н+,К+-АТФаза). Кислая фосфатаза служит надежным маркёром лизосом.
Лизосомы участвуют во внутриклеточном пищеварении. Лизосомы «работают» внутриклеточно, но некоторые клетки (например, нейтрофилы, остеокласты) для выполнения своих функций секретируют лизосомы во внеклеточное пространство.
Родственные лизосомам органеллы. Некоторые свойства лизосом обнаруживаются в специфических органеллах, характерных для опре- делённого типа клеток. К таким органеллам относят, например, меланосомы пигментных клеток, азурофильные гранулы нейтрофилов, плотные тельца тромбоцитов (δ-гранулы).
ПЕРОКСИСОМЫ
Пероксисомы - мембранные пузырьки размером 0,1-1,5 мкм с электронно-плотной сердцевиной. Они особенно многочисленны в клетках печени и почки; количество пероксисом в одной клетке может составлять до нескольких сотен. В мембране органеллы находятся специфичные для пероксисом белки - пероксины, а в матриксе - матриксные белки, в т.ч. более 40 ферментов (например, каталаза и пероксидаза), катализирующие анаболические (биосинтез жёлчных кислот) и катаболические (β-окисление длинных цепей жирных кислот, Н2О2-зависимое дыхание, деградация ксенобиотиков) процессы. Все компоненты пероксисом поступают из цитозоля. Продолжительность жизни пероксисом - 5-6 суток. Новые органеллы возникают из предсуществующих путём их простого деления.
ВКЛЮЧЕНИЯ
Клеточные включения образованы материалом, который постоянно присутствует в клетке. Это могут быть пигменты, запасы питательных веществ и энергии (жиры и гликоген), продукты расщепления компонентов клетки. К разнородной по морфологии и генезу группе включений относят находящиеся в цитозоле единичные или группы
гранул и капель: гликогена, липидов, пигментов, липофусцина (см. рис. 2-22).
• Пигментные включения. Одним из наиболее распространённых пигментов является меланин. Его особенно много в клетках-продуцентах - меланоцитах. Он присутствует также в клетках эпителия, соединительной ткани и даже в нейронах. Липофусцин образуется в результате деградации компонентов клетки, это продукт лизосомного переваривания, не подвергающийся дальнейшему расщеплению, а именно мембранные пузырьки различных размеров с электронноплотным содержимым. Внутриклеточные включения липофусцина имеют жёлто-коричневый цвет. Пигмент называют также пигментом изнашивания, или старения. Его легко найти в нервной системе, в тканях с высокой активностью макрофагов, в неделящихся клетках (нейроны, кардиомиоциты) и мышечных волокнах. Особенно много липофусцина в волокнах скелетной мышцы, где он располагается преимущественно под сарколеммой и около ядра. Количество пигмента с возрастом нарастает. Накопление липофусцина в жировых клетках придаёт жиру жёлтый цвет.
• Запасы питательных веществ и энергии. Подобные включения образованы гликогеном и липидами. Включения из гликогена чаще встречаются в печени и скелетной мышце. Гранулы гликогена размером 20-30 нм не связаны с мембраной, но часто расположены вблизи гладкой эндоплазматической сети. В отличие от других включений (например, липофусцина), гликоген может быть равномерно распределён по всему объёму цитоплазмы. Липидные (жировые) включения рассматриваются как источник энергии в клетке. Капли различных размеров и плотности, не связанные с мембранами; содержат триглицериды и холестерин. Выраженность липидных включений может быть преходящей. Длительность их пребывания в клетке варьирует от минут и часов (энтероциты кишки) до недель и месяцев (адипоциты).
Клеточный цикл
Клеточный цикл (рис. 2-51) состоит из митоза (М-фаза) и интерфазы. В интерфазе последовательно различают фазы G1, S и G2.
СТАДИИ КЛЕТОЧНОГО ЦИКЛА
Интерфаза
G1 следует за телофазой митоза. В эту фазу клетка синтезирует РНК и белки. Продолжительность фазы - от нескольких часов до нескольких дней.
G0. Клетки могут выйти из цикла и находиться в фазе G0. В фазе G0 клетки начинают дифференцироваться.
S. В фазу S в клетке продолжается синтез белка, происходит репликация ДНК, разделяются центриоли. В большинстве клеток фаза S длится 8-12 часов.
G2. В фазу G2 продолжается синтез РНК и белка (например, синтез тубулина для микротрубочек митотического веретена). Дочерние центриоли достигают размеров дефинитивных органелл. Эта фаза длится 2-4 часа.
Митоз
В ходе митоза делятся ядро (кариокинез) и цитоплазма (цитокинез). Фазы митоза: профаза, прометафаза, метафаза, анафаза, телофаза (рис. 2-52).
Профаза. Каждая хромосома состоит из двух сестринских хроматид, соединённых центромерой, исчезает ядрышко. Центриоли организуют митотическое веретено. Пара центриолей входит в состав ми-
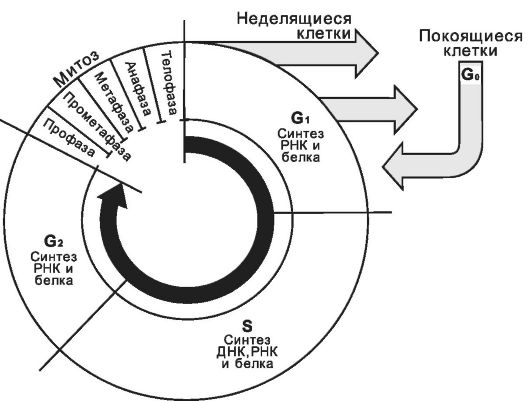 Рис. 2-51. Стадии клеточного цикла. В
клеточном цикле различают митоз - сравнительно короткую фазу M и более
длительный период - интерфазу. Фаза M состоит из профазы, прометафазы,
метафазы, анафазы и телофазы; интерфаза складывается из фаз Gj, S и G2. Клетки, выходящие из цикла, более не делятся и вступают в дифференцировку. Клетки в фазе G0 обычно не возвращаются в цикл. [17]
Рис. 2-51. Стадии клеточного цикла. В
клеточном цикле различают митоз - сравнительно короткую фазу M и более
длительный период - интерфазу. Фаза M состоит из профазы, прометафазы,
метафазы, анафазы и телофазы; интерфаза складывается из фаз Gj, S и G2. Клетки, выходящие из цикла, более не делятся и вступают в дифференцировку. Клетки в фазе G0 обычно не возвращаются в цикл. [17]
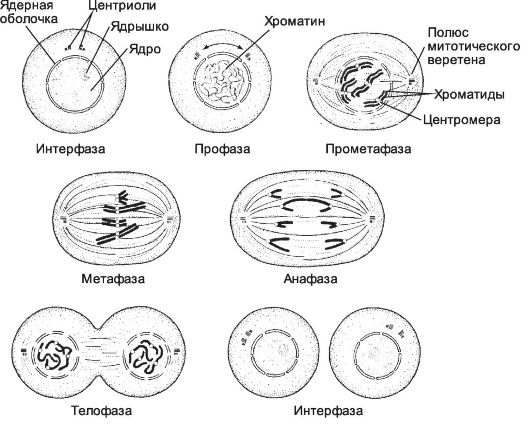 Рис. 2-52. M-фаза клеточного цикла. После фазы G2
начинается M-фаза клеточного цикла. Она состоит из пяти стадий деления
ядра (кариокинез) и деления цитоплазмы (цитокинез). M-фаза заканчивается
к началу фазы G1 следующего цикла. [65]
Рис. 2-52. M-фаза клеточного цикла. После фазы G2
начинается M-фаза клеточного цикла. Она состоит из пяти стадий деления
ядра (кариокинез) и деления цитоплазмы (цитокинез). M-фаза заканчивается
к началу фазы G1 следующего цикла. [65]
тотического центра, от которого радиально отходят микротрубочки. Сначала митотические центры располагаются вблизи ядерной мембраны, а затем расходятся и образуется биполярное митотическое веретено. В этом процессе участвуют полюсные микротрубочки, взаимодействующие между собой по мере удлинения. Центриоль входит в состав центросомы (центросома содержит две центриоли и перицентриольный матрикс) и имеет форму цилиндра диаметром 150 нм и длиной 500 нм; стенка цилиндра состоит из 9 триплетов микротрубочек. В центросоме центриоли расположены под прямым углом друг к другу. В ходе фазы S клеточного цикла центриоли дуплицируются. В митозе пары центриолей, каждая из которых состоит из первоначальной и вновь образованной, расходятся к полюсам клетки и участвуют в образовании митотического веретена. Прометафаза. Ядерная оболочка распадается на мелкие фрагменты. В области центромер появляются кинетохоры, функционирующие как центры организации кинетохорных микротрубочек. Отхождение кинетохор от каждой хромосомы в обе стороны и их взаимодействие
с полюсными микротрубочками митотического веретена - причина перемещения хромосом.
Метафаза. Хромосомы располагаются в области экватора веретена. Образуется метафазная пластинка, в которой каждая хромосома удерживается парой кинетохоров и связанными с ними кинетохорными микротрубочками, направленными к противоположным полюсам митотического веретена.
Анафаза - расхождение дочерних хромосом к полюсам митотического веретена со скоростью 1 мкм/мин.
Телофаза. Хроматиды подходят к полюсам, кинетохорные микротрубочки исчезают, а полюсные продолжают удлиняться. Образуется ядерная оболочка, появляется ядрышко.
Цитокинез - разделение цитоплазмы на две обособляющиеся части. Процесс начинается в поздней анафазе или в телофазе. Плазмолемма втягивается между двумя дочерними ядрами в плоскости, перпендикулярной длинной оси веретена. Борозда деления углубляется, и между дочерними клетками остаётся мостик - остаточное тельце. Дальнейшее разрушение этой структуры приводит к полному разделению дочерних клеток.
Регуляторы клеточного деления
Пролиферация клеток, происходящая путём митоза, жёстко регулируется множеством молекулярных сигналов. Скоординированная деятельность этих многочисленных регуляторов клеточного цикла обеспечивает как переход клеток от фазы к фазе клеточного цикла, так и точное выполнение событий каждой фазы. Главная причина появления пролиферативно неконтролируемых клеток - мутации генов, кодирующих структуру регуляторов клеточного цикла. Регуляторы клеточного цикла и митоза подразделяют на внутриклеточные и межклеточные. Внутриклеточные молекулярные сигналы многочисленны, среди них в первую очередь следует назвать собственно регуляторы клеточного цикла (циклины, циклин-зависимые протеинкиназы, их активаторы и ингибиторы) и онкосупрессоры.
Мейоз
В ходе мейоза образуются гаплоидные гаметы (рис. 2-53, см. также
рис. 15-8).
Первое деление мейоза
Первое деление мейоза (профаза I, метафаза I, анафаза I и телофаза I) - редукционное.
Профаза I последовательно проходит несколько стадий (лептотена, зиготена, пахитена, диплотена, диакинез).
Лептотена - хроматин конденсируется, каждая хромосома состоит из двух хроматид, соединённых центромерой.
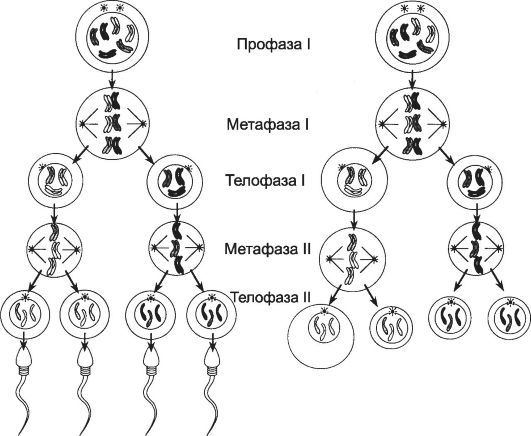 Рис. 2-53. Мейоз обеспечивает переход половых клеток из диплоидного состояния в гаплоидное. [17]
Рис. 2-53. Мейоз обеспечивает переход половых клеток из диплоидного состояния в гаплоидное. [17]
Зиготена - гомологичные парные хромосомы сближаются и вступают в физический контакт (синапсис) в виде синаптонемального комплекса, обеспечивающего конъюгацию хромосом. На этой стадии две лежащие рядом пары хромосом образуют бивалент.
Пахитена - хромосомы утолщаются вследствие спирализации. Отдельные участки конъюгировавших хромосом перекрещиваются друг с другом и образуют хиазмы. Здесь происходит кроссинговер - обмен участками между отцовскими и материнскими гомологичными хромосомами.
Диплотена - разделение конъюгировавших хромосом в каждой паре в результате продольного расщепления синаптонемального комплекса. Хромосомы расщепляются по всей длине комплекса, за исключением хиазм. В составе бивалента чётко различимы 4 хроматиды. Такой бивалент называют тетрадой. В хроматидах появляются участки раскручивания, где синтезируется РНК.
Диакинез. Продолжаются процессы укорочения хромосом и расщепления хромосомных пар. Хиазмы перемещаются к концам хромосом (терминализация). Разрушается ядерная мембрана, исчезает ядрышко. Появляется митотическое веретено. Метафаза I. В метафазе I тетрады образуют метафазную пластинку. В целом отцовские и материнские хромосомы распределяются случайным образом по ту или другую сторону экватора митотического веретена. Подобный характер распределения хромосом лежит в основе второго закона Менделя, что (наряду с кроссинговером) обеспечивает генетические различия между индивидуумами.
Анафаза I отличается от анафазы митоза тем, что при митозе к полюсам расходятся сестринские хроматиды. В эту фазу мейоза к полюсам отходят целостные хромосомы.
Телофаза I не отличается от телофазы митоза. Формируются ядра, имеющие 23 конъюгированные (удвоенные) хромосомы, происходит цитокинез, образуются дочерние клетки.
Второе деление мейоза
Второе деление мейоза - эквационное - протекает так же, как митоз (профаза II, метафаза II, анафаза II и телофаза II), но значительно быстрее. Дочерние клетки получают гаплоидный набор хромосом (22 аутосомы и одну половую хромосому).