Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 11 ИММУННАЯ ЗАЩИТА
В общем виде главная функция иммунной системы состоит в защите организма в условиях чужеродного окружения. Решение этой стратегической задачи осуществляется несколькими путями, в том числе защитой от «несвоего» (инфекции, трансплантат) и элиминации модифицированного «своего» (опухоли, повреждённые и стареющие клетки).
Основные понятия
Антиген - вещество, несущее признаки генетически чужеродной информации. Антиген можно определить как молекулу, распознаваемую иммунокомпетентными клетками как чужеродную (не свою). Молекула антигена взаимодействует с антителами (АТ), Т- или В-клеточными рецепторами. Если B-лимфоциты распознают свободную молекулу антигена, то T-лимфоциты - фрагмент антигена на поверхности других клеток. Количество разных антигенов (антигенных детерминант) достигает 1018.
Антитело (АТ) - гликопротеин, относящийся к классу иммуноглобулинов (Ig). АТ специфически взаимодействует с комплементарным антигеном. АТ существуют в миллионах разновидностей, и каждая молекула имеет уникальный участок связывания антигенной детерминанты. АТ синтезируются плазматическими клетками в ходе гуморального иммунного ответа. Выделено пять разных классов (изотипов) АТ - IgA, IgD, IgE, IgG и IgM.
Главный комплекс гиcтоcовмеcтимоcти. Синтез молекул главного комплекса гистосовместимости (молекул MHC) контролирует комплекс генов MHC. Гены MHC характеризуются выраженным полиморфизмом и имеют большое количество аллелей. Спектр молекул MHC уникален для каждого организма и определяет его биологическую индивидуальность. Молекулы, кодируемые генами MHC, подразделяют на два класса: молекулы MHC класса I и молекулы MHC класса II. Молекулы MHC - гликопротеины плазматической мембраны - экспрессированы на всех антиген-представляющих клетках и являются мишенями иммунного ответа. Молекулы MHC контролируют иммунный ответ.
Молекулы MHC класса II участвуют в представлении антигенов T-клет- кам и во взаимодействии Т- и В-лимфоцитов. Молекулы MHC классов I и II распознаются поверхностноклеточными дифференцировочными молекулами CD и участвуют в реакциях клеточной цитотоксичности, осуществляемой цитотоксическими Т-лимфоцитами (TC).
• CD8. Молекулы MHC класса I взаимодействуют с молекулой CD8, экспрессируемой на мембране предшественника TC.
• CD4. Молекулы MHC класса II взаимодействуют с молекулой CD4, экспрессируемой на мембране Т-хелпера (TH), что вызывает выделение лимфокинов, стимулирующих пролиферацию и созревание предшественников TC.
ИММУНОКОМПЕТЕНТНЫЕ КЛЕТКИ
К иммунокомпетентным клеткам относят Т- и В-лимфоциты (Т- и В- клетки), NK-клетки и антиген-представляющие клетки. T-лимфоци- ты развиваются в тимусе из костномозговых клеток-предшественниц. В-лимфоциты дифференцируются в печени плода и костном мозге взрослого организма. NK-клетки образуются из предшественников лимфоидных клеток в костном мозге. Все антиген-представляющие клетки имеют костномозговое происхождение.
T-лимфоциты
T-лимфоциты (тимус-зависимые) ответственны за клеточный иммунный ответ, а также помогают реагировать на антиген В-лимфоцитам при гуморальном иммунном ответе. T-лимфоциты узнают антиген, предварительно процессированный и представленный на поверхности антиген-представляющих клеток (рис. 11-5). T-клетки состоят из функциональных подтипов CD4+ и CD8+.
• T-хелперы (TH) - CD4+ T-клетки. При активации синтезируют и секретируют цитокины (ИЛ2, ИЛ4, ИЛ5, ИЛ6, γ-интерферон). В ходе иммунного ответа узнают молекулы MHC класса II.
• Цитотоксические T-лимфоциты (TC) - CD8+ T-клетки, уничтожают инфицированные вирусом, опухолевые и чужеродные клетки при помощи перфорина, взаимодействуя с молекулой MHC класса I, встроенной в плазматическую мембрану клетки-мишени.
• T-супрессоры (TS) - представители CD8+ T-клеток - регулируют интенсивность иммунного ответа, подавляя активность TH клеток; предотвращают развитие аутоиммунных реакций; защищают организм от нежелательных последствий иммунной реакции.
Дифференцировка T-лимфоцитов. Стволовая кроветворная клетка поступает из костного мозга в тимус, где протекает антиген-независимое созревание T-клеток. Здесь предшественники T-лимфоцитов начинают экспрессировать специфические маркёры: T-клеточный рецептор,
CD2, CD3, CD4 или CD8 (см. рис. 11-5).
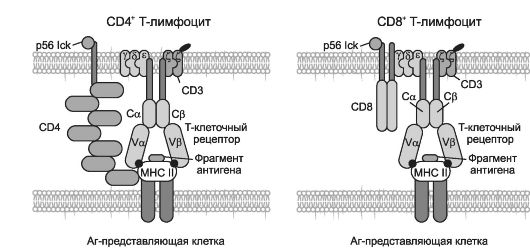 Рис. 11-5. Т-клеточный рецептор и другие специфические маркёры Т-клеток. [100]
Рис. 11-5. Т-клеточный рецептор и другие специфические маркёры Т-клеток. [100]
B-лимфоциты
В-лимфоциты ответственны за гуморальный иммунный ответ. Из красного костного мозга B-лимфоциты мигрируют в тимус-независимые зоны лимфоидных органов. Продолжительность жизни большинства В-лимфоцитов не превышает десяти дней, если они не активируются антигеном. В этом случае зрелые В-лимфоциты (плазматические клетки) вырабатывают АТ - Ig всех известных классов. В-клетки составляют от 5 до 15% лимфоцитов периферической крови и морфологически не отличаются от T-клеток. CD19, CD20, CD22, CD79a и В-клеточный рецептор - основные маркёры, используемые для идентификации В-клеток.
Дифференцировка B-лимфоцитов начинается в печени на 9-й неделе развития, продолжается в красном костном мозге, где и поддерживается на протяжении всей жизни. Дифферон для В-лимфоцита выглядит следующим образом: стволовая кроветворная клетка a общий лимфоидный предшественник В- и T-клеток a ранний про-В-лимфоцит (про от англ. progenitor, предшественник) - поздний про-Влимфоцит - большой пре-В-лимфоцит - малый пре-В-лимфоцит - незрелый В-лимфоцит - зрелый В-лимфоцит.
NK-клетки
NK-клетки составляют до 10% всех лимфоцитов крови, не имеют поверхностных детерминант, характерных для T- и В-лимфоцитов (MHC-нерестригированные киллеры). NK-клетки убивают ауто-, алло- и ксеногенные опухолевые клетки, инфицированные некоторыми вирусами и бактериями клетки. В типичных NK-клетках экспрессируются дифференцировочные молекулы CD2, CD7, CD56 и CD16 (рецептор Fс-фрагмента IgG).
Антиген-представляющие клетки
Антиген-представляющие клетки присутствуют преимущественно в коже, лимфатических узлах, селезёнке и тимусе. Это макрофаги, дендритные клетки, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерханса кожи (внутриэпидермальные дендроциты), эпителиальные клетки вилочковой железы. Эти клетки захватывают, процессируют и представляют антиген (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают интерлейкины и другие цитокины.
Дендритные клетки происходят из костного мозга и образуют популяцию долгоживущих клеток, которые запускают и модулируют иммунный ответ. В костном мозге их предшественники образуют субпопуляцию CD34+ клеток, которые способны дифференцироваться во внутриэпидермальные дендроциты для эпителия и дендритные клетки
для внутренней среды. Дифференцировку дендритных клеток поддерживают колониестимулирующий фактор гранулоцитов и макрофагов GM-CSF и ИЛ3. Дендритные клетки имеют звездчатую форму и в состоянии покоя несут на поверхности относительно небольшое количество молекул MHC.
ВЗАИМОДЕЙСТВИЕ КЛЕТОК ПРИ ИММУННОМ ОТВЕТЕ
Иммунный ответ возможен в результате активации клонов лимфоцитов и состоит из двух фаз. В первой фазе антиген активирует те лимфоциты, которые его распознают. Во второй (эффекторной) фазе эти лимфоциты координируют иммунный ответ, направленный на устранение антигена.
Гуморальный иммунный ответ
В гуморальном иммунном ответе участвуют макрофаги (антиген-представляющие клетки), T-хелперы и В-лимфоциты. Макрофаг поглощает вторгшийся в организм антиген и подвергает его процессингу - расщеплению на фрагменты. Фрагменты антигена выставляются на поверхности клетки вместе с молекулой MHC. Комплекс «антиген-молекула MHC класса II» предъявляется T-хелперу (рис. 11-9).
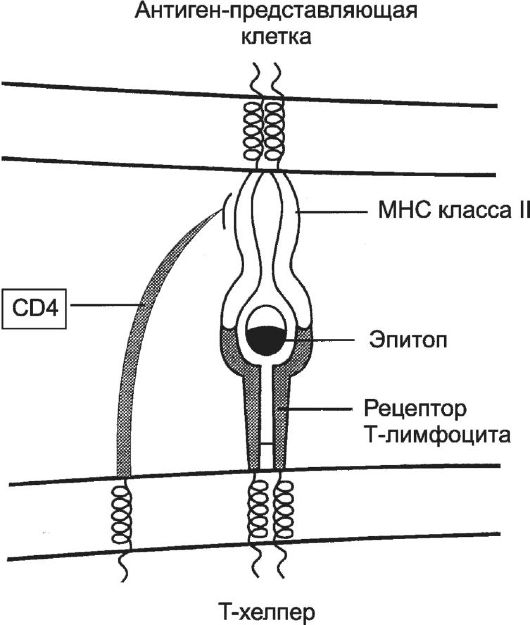 Рис. 11-9. Распознавание антигена рецептором T-лимфоцита. При
Рис. 11-9. Распознавание антигена рецептором T-лимфоцита. При
помощи рецептора T-лимфоцита T-клетка распознаёт антиген, но только находящийся в комплексе с молекулой MHC. В случае ТH-клетки в процессе участвует её молекула - CD4, которая свободным концом связывается с молекулой MHC. Распознаваемый T-клеткой антиген имеет два участка: один взаимодействует с молекулой MHC, другой (эпитоп) связывается с рецептором T-лимфоцита. Подобный тип взаимодействия, но с участием молекулы CD8, характерен для процесса распознавания TC-лимфоцитом антигена, связанного с молекулой MHC класса I. [17]
T-хелпер распознаёт комплекс «антиген-молекула MHC класса II» на поверхности антиген-представляющей клетки. Узнавание T-хелпе- ром нужных молекул на поверхности антиген-представляющей клетки стимулирует секрецию ИЛ1. Активированный ИЛ1 T-хелпер синтезирует ИЛ2 и рецепторы ИЛ2, через которые агонист стимулирует пролиферацию T-хелперов и цитотоксических T-лимфоцитов. В случае T-хелпера речь идёт об аутокринной стимуляции, когда клетка реагирует на тот агент, который сама же синтезирует и секретирует. Таким образом, после взаимодействия с антиген-представляющей клеткой T-хелпер приобретает способность отвечать на действие ИЛ2 всплеском пролиферации. Биологический смысл этого процесса состоит в накоплении такого количества T-хелперов, которое обеспечит образование в лимфоидных органах необходимого количества плазматических клеток, способных вырабатывать АТ против данного антигена. B-лимфоцит. Активация В-лимфоцита предполагает прямое взаимодействие антигена с Ig на поверхности В-клетки. В этом случае сам В-лимфоцит процессирует антиген и представляет его фрагмент в связи с молекулой MHC II на своей поверхности. Этот комплекс распоз- наёт T-хелпер, отобранный при помощи того же антигена, который участвовал в отборе данного В-лимфоцита.
Активированный B-лимфоцит дифференцируется в плазматическую клетку: увеличивается количество рибосом, гранулярная эндоплазматическая сеть и комплекс Гольджи становятся более выраженными. Плазматическая клетка (см. рис. 6-35) синтезирует Ig. ИЛ6, выделяемый активированными T-хелперами, стимулирует секрецию Ig. Часть зрелых В-лимфоцитов после антиген-зависимой дифференцировки циркулирует в организме как клетки памяти.
Клеточный иммунный ответ
Клеточный иммунный ответ характеризуется пролиферацией коммитированных иммунокомпетентных клеток, реагирующих с антигеном в комплексе с молекулой MHC класса I на поверхности чужеродных клеток или эндогенными антигенами в комплексе с молекулой MHC класса I на поверхности собственных вирус-инфицированных и трансформированных (опухолевых) клеток. В клеточном иммунном ответе участвует цитотоксический T-лимфоцит.
Цитотоксический T-лимфоцит (TC). Предъявленный на поверхности клетки-мишени антиген в комплексе с молекулой MHC класса I связывается с рецептором цитотоксического T-лимфоцита. В этом процессе участвует молекула CD8 клеточной мембраны TC. Секретируемый T-хелперами ИЛ2 стимулирует пролиферацию цитотоксических T-лимфоцитов. Цитотоксический T-лимфоцит распознаёт клетку-мишень и прикрепляется к ней. В цитоплазме активированного
цитотоксического T-лимфоцита присутствуют мелкие гранулы с цитолитическим белком перфорином. Выделяемые T-киллером молекулы перфорина полимеризуются в мембране клетки-мишени в присутствии Ca2+. Сформированные в плазматической мембране клетки-мишени перфориновые поры пропускают воду и соли, но не молекулы белка. В итоге клетка гибнет, теряя воду и ионы.
Органы иммунной защиты
Различают центральные (первичные) и периферические (вторичные) органы иммунной защиты. Центральные лимфоидные органы (костный мозг, тимус) - главное место лимфопоэза. Здесь лимфоциты дифференцируются из клеток-предшественниц, размножаются и созревают. T-клетки созревают в тимусе, а В-лимфоциты - в печени плода и костном мозге взрослого организма. В ходе дифференцировки в центральных органах лимфоциты начинают экспрессировать рецепторы, которые в дальнейшем могут связываться с антигеном. В центральных органах отбираются и выживают те лимфоциты, которые толерантны (невосприимчивы) к собственным антигенам. К периферическим (вторичным) лимфоидным органам относят селезёнку, лимфатические узлы, лимфоидную ткань, связанную со слизистой оболочкой (лимфатические фолликулы, миндалины). В периферических лимфоидных органах лимфоциты взаимодействуют между собой, со вспомогательными клетками и с антигенами. Здесь макрофаги, антиген-представляющие клетки и зрелые Т- и В-лимфоциты участвуют в иммунном ответе, образуются эффекторные клетки и клетки памяти. Иммунные реакции с участием циркулирующих в крови антигенов протекают в селезёнке. Клетки лимфатических узлов реагируют с антигеном, циркулирующим в лимфе. Лимфоидная ткань, связанная со слизистой оболочкой, реагирует на антиген, проникающий в неё из внешней среды.
ТИМУС
Тимус (вилочковая железа) относится к железам бранхиогенной группы и развивается из материала 3-го и 4-го глоточных карманов. На 8-й неделе в зачатке тимуса появляются лимфоидные клетки, мигрирующие из костного мозга.
Тимус - двудольчатый орган, расположенный в грудной полости. Каждая доля состоит из долек, разделённых соединительнотканными трабекулами (септами). Капсула и отходящие от неё септы построены из плотной волокнистой соединительной ткани. Объём органа заполнен эпителиальным каркасом, в котором располагаются тимоциты
(так называют лимфоциты тимуса). Эпителиальные (эпителиоретикулярные) клетки в дольке тимуса образуют сеть и участвуют в дифференцировке тимоцитов. В дольке зрелого тимуса различают корковый и мозговой слои.
В тимусе происходит антиген-независимая дифференцировка T-лим- фоцитов. Тимус подвергается возрастной инволюции. Атрофия начинается в пубертатном периоде и продолжается в течение всей жизни. Атрофия тимуса зависит от чувствительности тимоцитов к глюкокортикоидам. Все процессы, приводящие к резкому увеличению уровня глюкокортикоидов (беременность, стресс), ускоряют инволюцию тимуса. Образование T-клеток в тимусе продолжается и во взрослом организме, но с меньшей интенсивностью.
Дифференцировка тимоцитов
Клетка-предшественница T-лимфоцитов поступает в тимус из костного мозга в плодном периоде (рис. 11-11). Она экспрессирует на своей поверхности дифференцировочную молекулу CD7, а позже - CD1 и CD2. Протимоциты имеют фенотип CD7+CD1+CD2+cCD3+CD4-CD8-. Молекула CD1 исчезает, когда клетка становится зрелой, а цитоплазматическая форма CD3 (сCD3) переходит в мембранную. Тимоциты. По мере сборки α- и β-цепей T-клеточного рецептора протимоциты начинают экспрессировать маркёры CD4 и CD8, давая начало большинству тимоцитов с фенотипом CD1+CD2+CD4+CD8+. Эти клетки способны дифференцироваться в двух направлениях: в клетки CD1-CD2+CD4+CD8- и клетки CD1-CD2+CD4-CD8+ при наличии у обоих подтипов мембранного маркёра CD3 и αβ-рецептора T-лим- фоцита. Этим клеткам разрешено покидать тимус, они появляются в периферической крови и лимфоидных органах. Выселившиеся из тимуса T-лимфоциты экспрессируют либо CD4, либо CD8, но клетки фенотипа CD4+CD8+ отсутствуют.
Корковый слой
Субкапсулярная часть коркового слоя содержит делящиеся клетки - лимфобласты (клетки-предшественницы T-лимфоцитов, ранние протимоциты). Они взаимодействуют с эпителиальными клетками (рис. 11-13), при этом происходит селекция лимфоцитов и большинство T- лимфоцитов погибает в корковом слое. В корковом слое присутствует гематотимический барьер.
Внутренняя часть содержит потомки лимфобластов, в большинстве своём неделящиеся малые тимоциты и эпителиальные клетки. По мере созревания протимоциты утрачивают дифференцировочную молекулу CD1, но приобретают CD3, CD4 и CD8. Дальнейшая их дифференцировка протекает в мозговой части дольки тимуса.
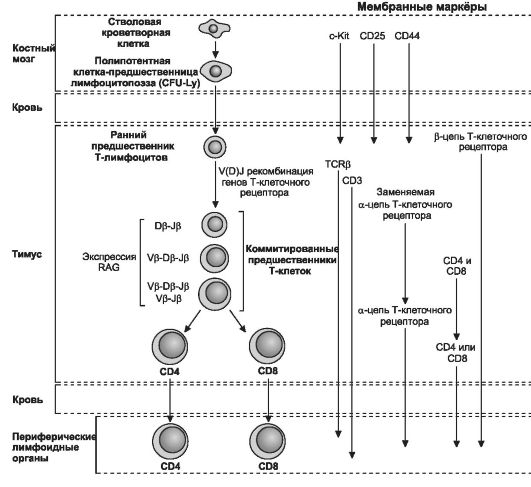 Рис. 11-11. Дифференцировка T-лимфоцитов от стволовой кроветворной клетки до состояния терминальной дифференцировки. [98]
Рис. 11-11. Дифференцировка T-лимфоцитов от стволовой кроветворной клетки до состояния терминальной дифференцировки. [98]
Эпителиальные (эпителиоретикулярные) клетки имеют крупное округлое ядро. Их характерная особенность - наличие длинных отростков, содержащих пучки тонофиламентов. Отростки соединяются между собой при помощи десмосом. В цитоплазме присутствуют гранулы, содержащие тимозины и тимопоэтин. Эпителиальные клетки экспрессируют молекулы MHC класса II.
Гематотимический барьер делает корковую часть недоступной для антигенов из внутренней среды организма и защищает от их действия созревающие здесь T-лимфоциты. Барьер образуют эндотелиальные клетки и эпителиальные клетки со своей базальной мембраной.
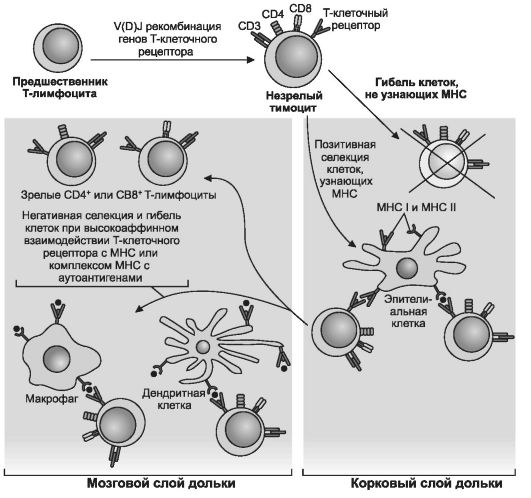 Рис. 11-13. Селекция тимоцитов. Выживающие
в ходе отбора тимоциты превращаются в зрелые T-клетки. В процесс отбора
вовлечены клетки тимуса - эпителиальные, дендритные и макрофаги.
Отобранные в тимусе зрелые T-лимфоциты не реагируют на собственные
молекулы MHC и толерантны к собственным антигенам. Важной сигнальной
молекулой, участвующей в процессах созревания и дифференцировки клеток,
является T-клеточный рецептор. [17]
Рис. 11-13. Селекция тимоцитов. Выживающие
в ходе отбора тимоциты превращаются в зрелые T-клетки. В процесс отбора
вовлечены клетки тимуса - эпителиальные, дендритные и макрофаги.
Отобранные в тимусе зрелые T-лимфоциты не реагируют на собственные
молекулы MHC и толерантны к собственным антигенам. Важной сигнальной
молекулой, участвующей в процессах созревания и дифференцировки клеток,
является T-клеточный рецептор. [17]
Мозговой слой
Тимоциты из коркового слоя поступают в мозговой слой и дифференцируются в CD4+ и CD8+ лимфоциты. Зрелые T-клетки выходят из мозгового слоя по венулам и выносящим лимфатическим сосудам. Только 3-5% клеток, продуцируемых в тимусе, покидает этот орган. Остальные клетки погибают. В мозговом слое присутствуют макрофа-
ги, уничтожающие погибшие или обречённые на гибель тимоциты. Дегенерирующие эпителиальные клетки мозгового слоя образуют слоистые эпителиальные тельца.
Дендритные клетки. Присутствующие в мозговом слое дендритные клетки фенотипически и функционально гетерогенны. Среди них различают типичные дендритные клетки и так называемые плазмацитоидные. Типичные дендритные клетки тимуса представляют собственные антигены тимоцитам и играют важную роль в процессе негативной селекции.
Функции тимуса
В вилочковой железе элиминируются лимфоциты, способные узнавать антигены собственного организма, т.е. происходит селекция лимфоцитов. В тимусе также вырабатываются гормоны иммунной системы.
• Селекция лимфоцитов. Молекулы рецепторов в клеточной мембране тимоцита взаимодействуют с комплексом «MHC-аутоантиген» в мембране эпителиальной клетки (см. рис. 11-13). Клоны тех тимоцитов, рецепторы которых узнают комплекс «MHC-аутоантиген», уничтожаются. Таким образом, нормально функционирующая иммунная система удаляет в вилочковой железе лимфоциты, запрограммированные активироваться антигенами собственного организма.
• Гормоны иммунной системы. В эпителиальных клетках тимуса синтезируются пептидные гормоны: тимозины, тимопоэтин, тимотаксин, тимический сывороточный фактор (тимулин) и др. Кроме того клетки тимуса синтезируют нейрофизины, вазопрессин, окситоцин, соматостатин и хромогранин А. Тимозины (α1, α3, α7, β4 и др.) способствуют дифференцировке T-лимфоцитов и появлению специфических рецепторов в их клеточной мембране, стимулируют выработку многих лимфокинов, в т.ч. ИЛ2. Тимопоэтин - стимулятор дифференцировки предшественников T-лимфоцитов, влияет на дифференцировку T-лимфоцитов, но не на их иммунологический репертуар.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
Сеть лимфатических узлов фильтрует антиген из интерстициальной жидкости и лимфы. Лимфатические узлы появляются в конце 2-го- начале 3-го месяцев внутриутробного развития. Лимфатический узел (рис. 11-16) имеет округлую или почковидную форму размером 2- 10 мм, снаружи покрыт соединительнотканной капсулой, от которой отходят трабекулы. Лимфа поступает в узел через несколько приносящих лимфатических сосудов. Через ворота (hilus) узла проходят кровеносные и выносящий лимфатический сосуд. В лимфатическом узле
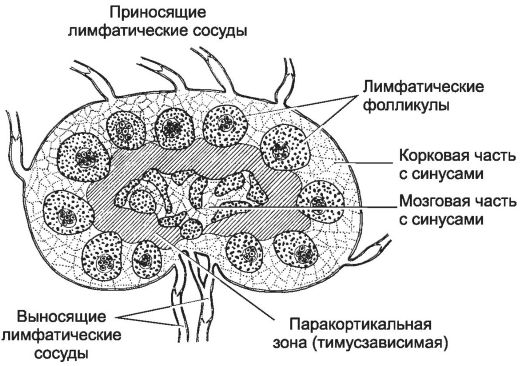 Рис. 11-16. Лимфатический узел разделён
на корковую и мозговую части. В корковой части расположены
лимфатические фолликулы, от которых в мозговую часть отходят мозговые
тяжи. Тимус-зависимая паракортикальная зона заштрихована. [17]
Рис. 11-16. Лимфатический узел разделён
на корковую и мозговую части. В корковой части расположены
лимфатические фолликулы, от которых в мозговую часть отходят мозговые
тяжи. Тимус-зависимая паракортикальная зона заштрихована. [17]
различают корковую и мозговую части, а также синусы. На границе между корковой и мозговой частями расположена тимус-зависимая паракортикальная зона. В лимфатическом узле T-лимфоциты взаимодействуют с B-лимфоцитами и фолликулярными отростчатыми клетками. Из паренхимы лимфатического узла лимфоциты поступают в выносящий лимфатический сосуд. В лимфатическом узле происходит созревание В- и T-клеток, в результате которого B-лимфоциты приобретают способность продуцировать АТ, T-хелперы - поддерживать цитокинами взаимодействие иммунокомпетентных клеток, а цитотоксические T-лимфоциты - уничтожать инфицированные и опухолевые клетки.
Корковая часть
По периферии лимфатического узла, в корковой его части, расположены многочисленные лимфатические фолликулы и корковые синусоиды (синусы). Корковая часть - B-клеточная зона. Лимфатические фолликулы. В ретикулярной строме лимфатического фолликула располагаются B-лимфоциты, дендритные клетки и макрофаги. При активации лимфатического фолликула формируется зародышевый центр (рис. 11-17).
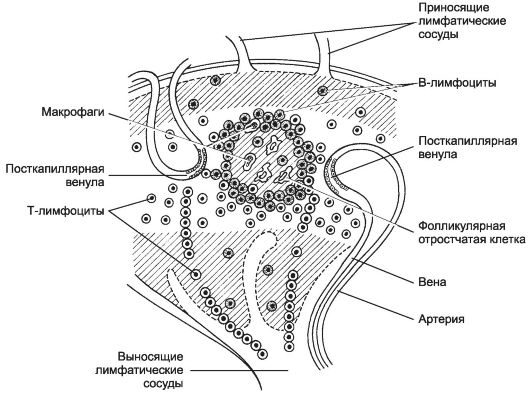 Рис. 11-17. Распределение Т- и B-лимфоцитов в лимфатическом узле. Тимус-зависимая
зона - светлая. Тимус-независимая зона заштрихована. T-лимфоциты
поступают в паренхиму узла из посткапиллярных венул и вступают в контакт
с фолликулярными отростчатыми клетками и B-лимфоцитами. [17]
Рис. 11-17. Распределение Т- и B-лимфоцитов в лимфатическом узле. Тимус-зависимая
зона - светлая. Тимус-независимая зона заштрихована. T-лимфоциты
поступают в паренхиму узла из посткапиллярных венул и вступают в контакт
с фолликулярными отростчатыми клетками и B-лимфоцитами. [17]
• Зародышевый центр - специализированная структура лимфатических фолликулов, появление которого сопровождает иммунный ответ. В центральной части такого активированного фолликула присутствует светлая зона - центр размножения (зародышевый центр), светлую зону окружает мантия из лимфоцитов (тёмная зона). В тём- ной зоне центра в B-клетках осуществляется процесс соматической рекомбинации генов иммуноглобулинов. Этому процессу сопутствует негативная селекция B-клеток в светлой зоне.
• Дендритные клетки . Активированные фолликулы имеют два типа дендритных клеток: фолликулярные отростчатые клетки, которые представляют нативный антиген B-лимфоцитам, и CD11c+-клетки, процессирующие антиген и представляющие его фрагмент T-хелперам.
• Плазматические клетки. В зародышевом центре B-лимфоциты постоянно вступают в апоптоз. Но - если B-лимфоцит встречается со своим антигеном и в пределах светлой зоны зародышевого центра
взаимодействует с активированным T-лимфоцитом - то апоптоз этого B-лимфоцита откладывается, и B-клетка начинает активно пролиферировать со скоростью, удваивающей здесь количество B- лимфоцитов каждые 6 часов. Пролиферирующие B-клетки смещают неактивированные B-лимфоциты на периферию и в совокупности образуют тёмную зону. Часть пролиферирующих B-клеток дифференцируется в плазматические клетки.
Мозговая часть
Центральная часть узла содержит мозговые тяжи и синусоиды. Мозговые тяжи образованы скоплением Т- и B-лимфоцитов, плазматических клеток, многочисленных макрофагов. Большинство клеток в тяжах - мигранты из корковой части.
Тимус-зависимая зона
Тимус-зависимая зона - паракортикальная зона лимфатического узла (см. рис. 11-17). В тимус-зависимой зоне большинство клеток готово к реакции с антигеном. T-лимфоциты располагаются преимущественно в этой зоне, откуда они мигрируют в мозговые синусы и поступают в выносящий лимфатический сосуд. Эта зона содержит многочисленные антиген-представляющие (дендритные) клетки, экспрессирующие на своей поверхности молекулы MHC класса II. В этой же зоне присутствуют посткапиллярные венулы с кубическими (высокими) эндотелиальными клетками, где происходит хоминг лимфоцитов - именно здесь из циркуляции в лимфатический узел поступают В- и T-лимфоциты. Через стенку этих венул в лимфатический узел за одну секунду проходит до 10 000 лимфоцитов. Венулы с кубическим эндотелием - характерная морфологическая особенность всех вторичных лимфоидных органов, кроме селезёнки.
В паракортикальной зоне активируются цитотоксические T-лимфоци- ты. Здесь они распознают антиген на поверхности инфицированных вирусом макрофагов или дендритных клеток и служат мишенями воздействия цитокинов из активированных TH-клеток. Будучи активированными, цитотоксические T-лимфоциты пролиферируют и рециркулируют, часть из них вновь поступает во вторичные лимфоидные органы.
Синусы
Под капсулой расположен краевой синус, куда поступает лимфа из приносящих лимфатических сосудов. Краевой синус через промежуточные синусы переходит в синусы мозгового вещества, а из них лимфа по выносящему лимфатическому сосуду в области ворот выходит из органа.
СЕЛЕЗЁНКА
Селезёнка закладывается на 5-й неделе внутриутробного развития, снаружи покрыта соединительнотканной капсулой, содержащей ГМК и большое количество эластина. От капсулы отходят трабекулы. В паренхиме органа различима белая и красная пульпа. В отличие от всех других вторичных лимфоидных органов, селезёнка не содержит венул с высоким эндотелием и не имеет приносящих лимфатических сосудов, поэтому антигены попадают в орган только через кровь. Следовательно, селезёнка является фильтром для патогенов, циркулирующих в крови. Красная пульпа представлена синусами и тяжами, содержащими ретикулярные клетки, макрофаги, эритроциты, тромбоциты, гранулоциты, лимфоциты и многочисленные плазматические клетки. В ретикулярной строме красной пульпы преобладают эритроциты и присутствуют многочисленные макрофаги, уничтожающие отжившие эритроциты. Белая пульпа - совокупность лимфоидной ткани селезёнки, представленная скоплениями T-лимфоцитов вокруг артерий, выходящих из трабекул, и артериол (тимус-зависимая зона). Лимфатические фолликулы - тимус-независимая зона (рис. 11-19). После взаимодействия в тимус-зависимой зоне с антиген-представляющими клетками T-лимфоциты перемещаются в лимфатические фолликулы, где активируют B-лимфоциты.
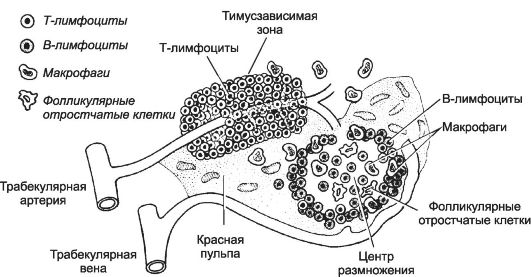 Рис. 11-19. Тимус-зависимая и тимус-независимая зоны селезёнки. Скопление
T- лимфоцитов (светлые клетки) вокруг артерий, вышедших из трабекул,
образует тимус-зависимую зону. Лимфатический фолликул и окружающая его
лимфоидная ткань белой пульпы - тимус-независимая зона. Здесь
присутствуют B-лимфоциты (тёмные клетки), макрофаги и фолликулярные
отростчатые клетки. [17]
Рис. 11-19. Тимус-зависимая и тимус-независимая зоны селезёнки. Скопление
T- лимфоцитов (светлые клетки) вокруг артерий, вышедших из трабекул,
образует тимус-зависимую зону. Лимфатический фолликул и окружающая его
лимфоидная ткань белой пульпы - тимус-независимая зона. Здесь
присутствуют B-лимфоциты (тёмные клетки), макрофаги и фолликулярные
отростчатые клетки. [17]
Лимфатический фолликул. В нём различают центр размножения и краевую зону.
• Центр размножения. В центральной части фолликулов (центр размножения, или реактивный центр) присутствуют макрофаги, фолликулярные отростчатые клетки и B-лимфоциты. Скопления B-лимфоцитов расположены и по периферии фолликула.
• Краевая зона (рис. 11-19) - граница между фолликулом и красной пульпой. Здесь присутствуют многочисленные активно фагоцитирующие макрофаги, антиген-представляющие клетки, медленно рециркулирующие B-лимфоциты и NK-клетки. Во внутренней части краевой зоны расположены синусы, куда поступает кровь из артериальных сосудов фолликула. В краевой зоне кровь вступает в контакт с паренхимой органа. Здесь из кровеносного русла в ткань выходят Т- и B-лимфоциты, распределяющиеся по специфическим для каждого клеточного типа зонам селезёнки.
Кровоток в селезёнке
Артерии, входящие в ворота органа, разветвляются на более мелкие трабекулярные артерии (рис. 11-19, 11-20, 11-21). Они покидают трабекулы и входят в пульпу (пульпарные артерии). Центральные артерии. От пульпарных артерий в фолликулы отходят артериолы (традиционно называемые центральными артериями), разветвляющиеся на капилляры в составе фолликулов белой пульпы. Тимус-зависимая зона - муфты из T-лимфоцитов, окружающие эти ветви пульпарных артерий.
Кисточковые артериолы. Центральные артерии выходят из фолликула в красную пульпу и делятся на расходящиеся ветви - кисточковые артериолы, входящие в состав эллипсоидов. Эллипсоиды содержат скопления макрофагов, окружающих сосуды. В пределах эллипсоидов артериолы переходят в капилляры.
• По теории незамкнутой циркуляции, кровь из капилляров поступает в ретикулярную ткань красной пульпы, а затем - в синусоиды.
• По теории замкнутой циркуляции, капилляры открываются прямо в синусоиды. Данные последних лет подтверждают теорию незамкнутой циркуляции селезёнки.
Синусоиды. Ретикулярная строма красной пульпы пронизана синусоидами диаметром до 40 мкм. Эндотелиальные клетки синусоидов расположены продольно и выпячиваются в просвет сосудов. Между эндотелиальными клетками имеются продольные щели, через которые проходят форменные элементы крови.
Венозный отток. Кровь из синусоидов поступает в пульпарные вены, далее в трабекулярные вены к воротам органа.
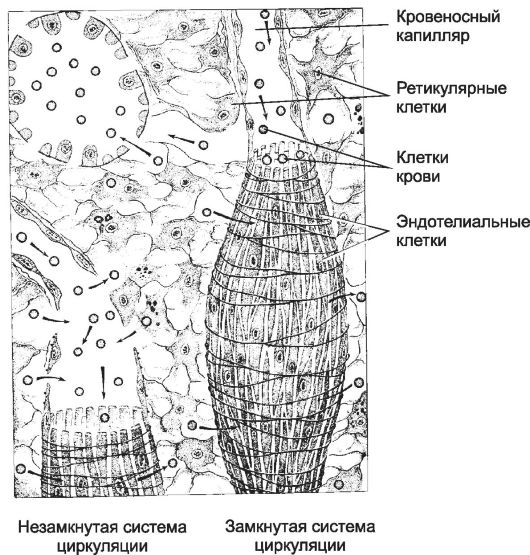 Рис. 11-20. Синусоиды в красной пульпе селезёнки. По
теории незамкнутой циркуляции (слева), кровь из капилляров поступает в
красную пульпу, а затем - в синусоиды. По теории замкнутой циркуляции
(справа), капилляры открываются прямо в синусоиды. [17]
Рис. 11-20. Синусоиды в красной пульпе селезёнки. По
теории незамкнутой циркуляции (слева), кровь из капилляров поступает в
красную пульпу, а затем - в синусоиды. По теории замкнутой циркуляции
(справа), капилляры открываются прямо в синусоиды. [17]
Функции селезёнки
• Селезёнка продуцирует иммуноглобулины. После спленэктомии наблюдают снижение уровня сывороточных АТ.
• Cелезёнка - место образования гуморальных факторов, влияющих на систему мононуклеарных фагоцитов. Так, тафтсин стимулирует активность фагоцитов, а спленин - функциональный аналог тимопоэтина.
• Фагоцитоз повреждённых и старых эритроцитов и тромбоцитов. Закончившие жизненный цикл эритроциты и тромбоциты распоз-
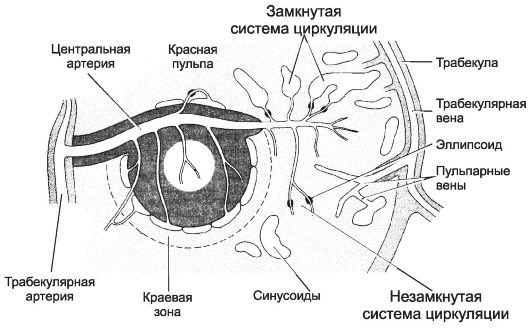 Рис. 11-21. Кровоток в селезёнке. Трабекулярные
артерии → пульпарные артерии → артериолы и капилляры фолликула → синусы
краевой зоны → выход Т- и B-лим- фоцитов из сосудистого русла.
Артериолы фолликула → кисточковые артериолы: красной пульпы →
капилляры-синусоиды. [17]
Рис. 11-21. Кровоток в селезёнке. Трабекулярные
артерии → пульпарные артерии → артериолы и капилляры фолликула → синусы
краевой зоны → выход Т- и B-лим- фоцитов из сосудистого русла.
Артериолы фолликула → кисточковые артериолы: красной пульпы →
капилляры-синусоиды. [17]
• наются и фагоцитируются макрофагами. Этот процесс протекает в красной пульпе. Селезёнка - депо эритроцитов, гранулоцитов и тромбоцитов.
ЛИМФОИДНАЯ ТКАНЬ СЛИЗИСТЫХ ОБОЛОЧЕК
Слизистые оболочки ЖКТ, воздухоносных и мочевыделительных путей содержат значительное количество ретикулиновых волокон, одного из элементов поддерживающего каркаса органов кроветворения. Здесь, как правило, скапливаются лимфоциты, зачастую формирующие лимфатические фолликулы. Типичный пример лимфоидной ткани в слизистой оболочке - агрегаты лимфатических фолликулов в подвздошной кишке (пейеровы бляшки). В лимфоидной ткани слизистых оболочек присутствуют венулы с высоким (кубическим) эндотелием, через стенку которых в ткань поступают лимфоциты. Бляшка имеет выносящие, но (в отличие от лимфатического узла) не имеет приносящих лимфатических сосудов. Антигены из просвета кишки «подаются» в бляшку при помощи М-клеток. Собственный слой слизистой оболочки содержит дендритные клетки, отростки которых про-
никают в эпителий. Они, как и М-клетки, взаимодействуют с бактериальной микрофлорой.
Поверхность слизистой оболочки в дыхательной и пищеварительной системах защищена слизью, дефензинами и секреторным IgA. Транспорт, процессинг и представление антигена осуществляются при тесном взаимодействии организованной лимфоидной ткани и ассоциированного с фолликулами эпителия. Периферия лимфатических фолликулов в слизистой оболочке содержит большое количество T-лимфоцитов. В фолликулах присутствуют также B-клетки, дендритные клетки и венулы с высоким эндотелием. В пейеровых бляшках каждый фолликул отделён от эпителия субэпителиальной куполообразной зоной, богатой Т-, B-лимфоцитами и дендритными клетками. Ассоциированный с фолликулами эпителий содержит М-клетки, которые транспортируют чужеродные макромолекулы и микроорганизмы к антиген-представляющим клеткам, расположенным по другую сторону эпителиального барьера.
М-клетка - типичная эпителиальная клетка с полярной дифференцировкой, формирующая плотные контакты. Отличительной её особенностью является наличие расположенного в базолатеральной части внутриэпителиального кармана. С плазмолеммой этого кармана взаимодействуют интраэпителиальные лимфоциты при участии молекул адгезии. Мембрана апикальной части клетки не участвует в формировании всасывательной каёмки, имеет нерегулярные микроворсинки и микроскладки и приспособлена для прикрепления чужеродных молекул, частиц и микроорганизмов.