Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. - 480 с.
|
|
|
|
ГЛАВА 7 МЫШЕЧНЫЕ ТКАНИ
Мышечная ткань осуществляет двигательные функции организма. У части гистологических элементов мышечной ткани имеются сократительные единицы - саркомеры (см. рис. 6-3). Это обстоятельство позволяет различать два типа мышечных тканей. Один из них - по- перечно-полоcатая (скелетная и сердечная) и второй - гладкая. Во всех сократительных элементах мышечных тканей (поперечно-полосатое скелетное мышечное волокно, кардиомиоциты, гладкомышечные клетки - ГМК), а также в немышечных сократительных клетках функционирует актомиозиновый хемомеханический преобразователь. Сократительную функцию скелетной мышечной ткани (произвольная мускулатура) контролирует нервная система (соматическая двигательная иннервация). Непроизвольные мышцы (сердечная и гладкая) имеют вегетативную двигательную иннервацию, а также развитую систему гуморального контроля. Для ГМК характерна выраженная физиологическая и репаративная регенерация. В составе скелетных мышечных волокон присутствуют стволовые клетки (клетки-сателлиты), поэтому скелетная мышечная ткань потенциально способна к регенерации. Кардиомиоциты находятся в фазе G0 клеточного цикла, а стволовые клетки в сердечной мышечной ткани отсутствуют. По этой причине погибшие кардиомиоциты замещаются соединительной тканью.
Скелетная мышечная ткань
У человека более 600 скелетных мышц (около 40% массы тела). Скелетная мышечная ткань обеспечивает осознанные и осознаваемые произвольные движения тела и его частей. Основные гистологические элементы: скелетные мышечные волокна (функция сокращения) и клетки-сателлиты (камбиальный резерв).
Источники развития гистологических элементов скелетной мышечной ткани - миотомы и нервный гребень.
Миогенный клеточный тип последовательно складывается из следующих этапов: клетки миотома (миграция) → миобласты митотические (пролиферация) → миобласты постмитотические (слияние) → мы-
шечные трубочки (синтез сократительных белков, формирование саркомеров) → мышечные волокна (функция сокращения).
• Мышечная трубочка. После ряда митотических делений миобласты приобретают вытянутую форму, выстраиваются в параллельные цепи и начинают сливаться, образуя мышечные трубочки (миотубы). В мышечных трубочках происходит синтез контрактильных белков и сборка миофибрилл - сократительных структур с характерной поперечной исчерченностью. Окончательная дифференцировка мышечной трубочки наступает только после её иннервации.
• Мышечное волокно. Перемещение ядер симпласта на периферию завершает формирование поперечно-полосатого мышечного волокна.
• Kлетки-сaтеллиты - обособившиеся в ходе миогенеза G1-миобласты, расположенные между базальной мембраной и плазмолеммой мышечных волокон. Ядра этих клеток составляют 30% у новорождённых, 4% у взрослых и 2% у пожилых от суммарного количества ядер скелетного мышечного волокна. Клетки-сателлиты - камбиальный резерв мышечной ткани скелетного типа. Они сохраняют способность к миогенной дифференцировке, что обеспечивает рост мышечных волокон в длину в постнатальном периоде. Клетки-сателлиты также участвуют в репаративной регенерации скелетной мышечной ткани.
СКЕЛЕТНОЕ МЫШЕЧНОЕ ВОЛОКНО
Структурно-функциональная единица скелетной мышцы - симпласт - скелетное мышечное волокно (рис. 7-1, рис. 7-7), имеет форму протяжённого цилиндра с заострёнными концами. Этот цилиндр достигает в длину 40 мм при диаметре до 0,1 мм. Термином «оболочка волокна» (сярколемма) обозначают две структуры: плазмолемму симпласта и его базальную мембрану. Между плазмолеммой и базальной мембраной расположены клетки-сателлиты с овальными ядрами. Палочковидной формы ядра мышечного волокна лежат в цитоплазме (саркоплазма) под плазмолеммой. В саркоплазме симпласта расположен сократительный аппарат - миофибриллы, депо Ca2+ - саркоплазматическая сеть (гладкий эндоплазматический ретикулум), а также митохондрии и гранулы гликогена. От поверхности мышечного волокна к расширенным участкам саркоплазматического ретикулума направляются трубковидные впячивания сарколеммы - поперечные трубочки (Т-трубочки). Рыхлая волокнистая соединительная ткань между отдельными мышечными волокнами (эндомизий) содержит кровеносные и лимфатические сосуды, нервные волокна. Группы мышечных волокон и окружающая их в виде чехла волокнистая соединительная ткань (перимизий) формируют пучки. Их совокупность образует мышцу, плотный соединительнотканный чехол которой именуют эпимизий (рис. 7-2).
Миофибриллы
Поперечная исчерченность скелетного мышечного волокна определяется регулярным чередованием в миофибриллах различно преломляю-
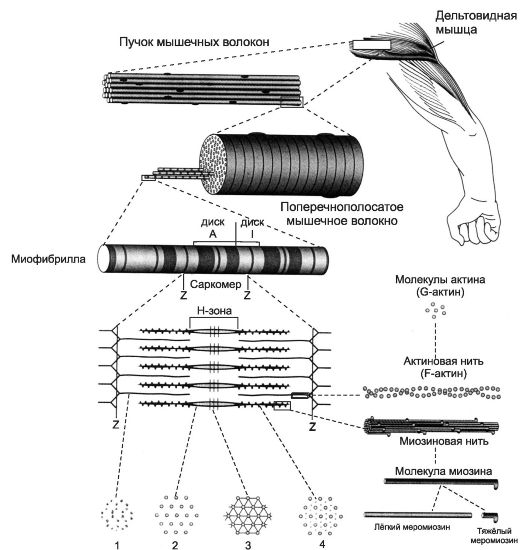 Рис. 7-1. Скелетная мышца состоит из поперечно-полосатых мышечных волокон.
Рис. 7-1. Скелетная мышца состоит из поперечно-полосатых мышечных волокон.
Значительный объём мышечного волокна занимают миофибриллы. Расположение светлых и тёмных дисков в параллельных друг другу миофибриллах совпадает, что приводит к появлению поперечной исчерченности. Структурная единица миофибрилл - саркомер, сформированный из толстых (миозиновых) и тонких (актиновых) нитей. Расположение тонких и толстых нитей в саркомере показано справа и внизу. G-актин - глобулярный, F-актин - фибриллярный актин. [26]
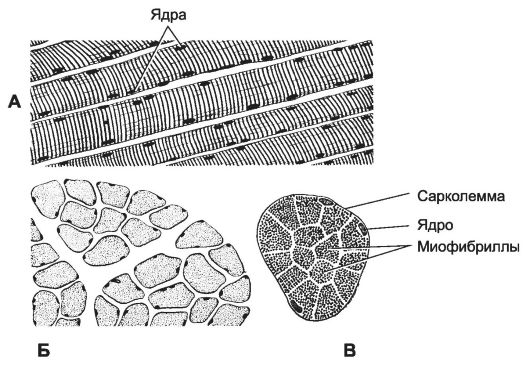 Рис. 7-2. Скелетная мышца в продольном и поперечном разрезе. А - продольный разрез; Б - поперечный разрез; В - поперечный срез отдельного мышечного волокна. [17]
Рис. 7-2. Скелетная мышца в продольном и поперечном разрезе. А - продольный разрез; Б - поперечный разрез; В - поперечный срез отдельного мышечного волокна. [17]
щих поляризованный свет участков (дисков) - изотропных и анизотропных: светлые (Isotropic, I-диски) и тёмные (Anisotropic, А-диски) диски. Разное светопреломление дисков определяется упорядоченным расположением по длине саркомера тонких и толстых нитей; толстые нити находятся только в тёмных дисках, светлые диски не содержат толстых нитей. Каждый светлый диск пересекает Z-линия. Участок миофибриллы между соседними Z-линиями определяют как саркомер. Саркомер. Структурно-функциональная единица миофибриллы, находящаяся между соседними Z-линиями (рис. 7-3). Саркомер образуют расположенные параллельно друг другу тонкие (актиновые) и толстые (миозиновые) нити. I-диск содержит только тонкие нити. В середине I-диска проходит Z-линия. Один конец тонкой нити прикреплён к Z-линии, а другой конец направлен к середине сaркомера. Толстые нити занимают центральную часть сaркомера - А-диск. Тонкие нити частично входят между толстыми. Содержащий только толстые нити участок сaркомера - Н-зона. В середине Н-зоны проходит М-линия. I-диск входит в состав двух сaркомеров. Следовательно, каждый сaр- комер содержит один А-диск (тёмный) и две половины I-диска (светлого), формула саркомера - 1/2 I + А + 1/2 I.
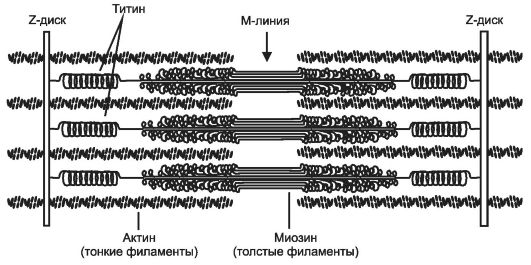 Рис. 7-3. Саркомер содержит
один А-диск (тёмный) и две половины I-диска (светлого). Толстые
миозиновые нити занимают центральную часть саркомера. Титин соединяет
свободные концы миозиновых нитей с Z-линией. Тонкие актиновые нити одним
концом прикреплены к Z-линии, а другим направляются к середине
сяркомера и частично входят между толстыми нитями. [17]
Рис. 7-3. Саркомер содержит
один А-диск (тёмный) и две половины I-диска (светлого). Толстые
миозиновые нити занимают центральную часть саркомера. Титин соединяет
свободные концы миозиновых нитей с Z-линией. Тонкие актиновые нити одним
концом прикреплены к Z-линии, а другим направляются к середине
сяркомера и частично входят между толстыми нитями. [17]
• Толстая нить. Каждая миозиновая нить состоит из 300-400 молекул миозина и С-белка. Половина молекул миозина обращена головками к одному концу нити, а вторая половина - к другому. Гигантский белок титин связывает свободные концы толстых нитей с Z-линией.
• Тонкая нить состоит из актина, тропомиозина и тропонинов (рис. 7-6).
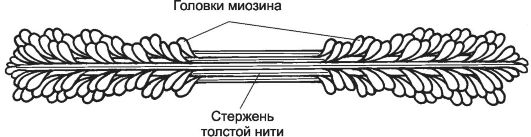 Рис. 7-5. Толстая нить. Молекулы миозина способны к самосборке и формируют веретенообразный агрегат диаметром 15 нм и длиной 1,5 мкм. Фибриллярные хвосты молекул образуют стержень толстой нити, головки миозина расположены спиралями и выступают над поверхностью толстой нити. [17]
Рис. 7-5. Толстая нить. Молекулы миозина способны к самосборке и формируют веретенообразный агрегат диаметром 15 нм и длиной 1,5 мкм. Фибриллярные хвосты молекул образуют стержень толстой нити, головки миозина расположены спиралями и выступают над поверхностью толстой нити. [17]
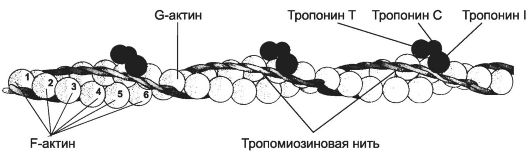 Рис. 7-6. Тонкая нить -
две спирально скрученные нити F-актина. В канавках спиральной цепочки
залегает двойная спираль тропомиозина, вдоль которой располагаются
молекулы тропонина. [17]
Рис. 7-6. Тонкая нить -
две спирально скрученные нити F-актина. В канавках спиральной цепочки
залегает двойная спираль тропомиозина, вдоль которой располагаются
молекулы тропонина. [17]
Саркоплазматическая сеть
Каждая миофибрилла окружена регулярно повторяющимися элементами сaркоплазматического ретикулума - анастомозирующими мембранными трубочками, заканчивающимися терминальными цистернами (рис. 7-7). На границе между тёмным и светлым дисками две смежные терминальные цистерны контактируют с Т-трубочками, образуя так называемые триады. Саркоплазматический ретикулум - модифицированная гладкая эндоплазматическая сеть, выполняющая функцию депо кальция.
Сопряжение возбуждения и сокращения
Сарколемма мышечного волокна образует множество узких впячиваний - поперечных трубочек (Т-трубочки). Они проникают внутрь мышечного волокна и, залегая между двумя терминальными цистернами сaркоплазматического ретикулума, вместе с последними формируют триады. В триадах происходит передача возбуждения в виде потенциала действия плазматической мембраны мышечного волокна на мембрану терминальных цистерн, т.е. процесс сопряжения возбуждения и сокращения.
ИННЕРВАЦИЯ СКЕЛЕТНЫХ МЫШЦ
В скелетных мышцах различают экстрафузальные и интрафузальные мышечные волокна.
• Экстрафузальные мышечные волокна, осуществляющие функцию сокращения мышцы, имеет прямую двигательную иннервацию - нервно-мышечный синапс, образованный терминальным ветвлением аксона α-мотонейрона и специализированным участком плазмолеммы мышечного волокна (концевая пластинка, постсинаптическая мембрана, см. рис. 8-29).
• Интрафузальные мышечные волокна входят в состав чувствительных нервных окончаний скелетной мышцы - мышечных веретён. Интрафузальные мышеч-
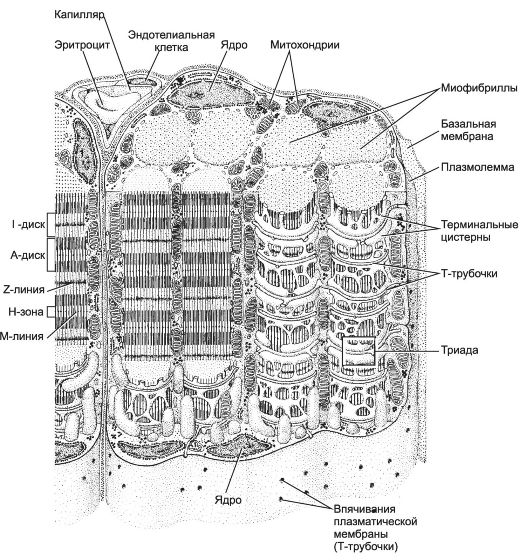 Рис. 7-7. Фрагмент скелетного мышечного волокна. Цистерны
саркоплазматического ретикулума окружают каждую миофибриллу. Т-трубочки
подходят к миофибриллам на уровне границ между тёмными и светлыми
дисками и вместе с терминальными цистернами саркоплазматического
ретикулума образуют триады. Между миофибриллами залегают митохондрии.
[17]
Рис. 7-7. Фрагмент скелетного мышечного волокна. Цистерны
саркоплазматического ретикулума окружают каждую миофибриллу. Т-трубочки
подходят к миофибриллам на уровне границ между тёмными и светлыми
дисками и вместе с терминальными цистернами саркоплазматического
ретикулума образуют триады. Между миофибриллами залегают митохондрии.
[17]
ные волокна образуют нервно-мышечные синапсы с эфферентными волокнами γ-мотонейронов и чувствительные окончания с волокнами псевдоуниполярных нейронов спинномозговых узлов (рис. 7-9, рис. 8-27). Двигательная соматическая иннервация скелетных мышц (мышечных волокон) осуществляется α- и γ-мотонейронами передних рогов спин-
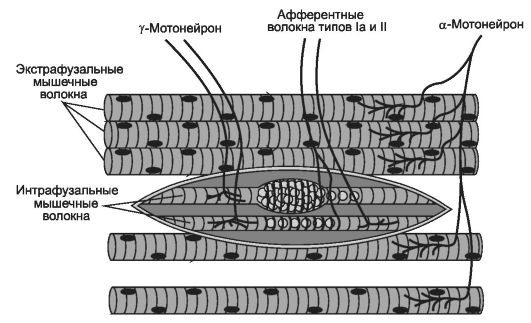 Рис. 7-9. Иннервация экстрафузальных и интрафузальных мышечных волокон. Экстрафузальные
мышечные волокна скелетных мышц туловища и конечностей получают
двигательную иннервацию от α-мотонейронов передних рогов спинного мозга.
Интрафузальные мышечные волокна в составе мышечных веретён имеют как
двигательную иннервацию от γ-мотонейронов, так и чувствительную
(афферентные волокна Iа и II типов чувствительных нейронов
спинномозгового узла). [74]
Рис. 7-9. Иннервация экстрафузальных и интрафузальных мышечных волокон. Экстрафузальные
мышечные волокна скелетных мышц туловища и конечностей получают
двигательную иннервацию от α-мотонейронов передних рогов спинного мозга.
Интрафузальные мышечные волокна в составе мышечных веретён имеют как
двигательную иннервацию от γ-мотонейронов, так и чувствительную
(афферентные волокна Iа и II типов чувствительных нейронов
спинномозгового узла). [74]
ного мозга и двигательных ядер черепных нервов, а чувствительная соматическая иннервация - псевдоуниполярными нейронами чувствительных спинномозговых узлов и нейронами чувствительных ядер черепных нервов. Вегетативная иннервация мышечных волокон не обнаружена, но ГМК стенки кровеносных сосудов скелетных мышц имеют симпатическую адренергическую иннервацию.
СОКРАЩЕНИЕ И РАССЛАБЛЕНИЕ
Сокращение мышечного волокна происходит при поступлении по аксонам двигательных нейронов к нервно-мышечным синапсам (см. рис. 8-29) волны возбуждения в виде нервных импульсов и выброса нейромедиатора ацетилхолина из концевых разветвлений аксона. Дальнейшие события развёртываются следующим образом: деполяризация постсинаптической мембраны → распространение потенциала действия по плазмолемме → передача сигнала через триады на саркоплазматическую сеть → выброс ионов Ca2+ из саркоплазмати-
ческой сети → взаимодействие тонких и толстых нитей, в результате чего происходит укорочение саркомера и сокращение мышечного волокна → расслабление.
ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
Скелетные мышцы и образующие их мышечные волокна различаются по множеству параметров. Традиционно выделяют красные, белые и промежуточные, а также медленные и быстрые мышцы и волокна.
• Красные (окислительные) мышечные волокна небольшого диаметра, окружены массой капилляров, содержат много миоглобина. Их многочисленные митохондрии имеют высокой уровень активности окислительных ферментов (например, сукцинатдегидрогеназы).
• Белые (гликолитические) мышечные волокна имеют больший диаметр, в саркоплазме содержится значительное количество гликогена, митохондрии немногочисленны. Для них характерны низкая активность окислительных ферментов и высокая активность гликолитических ферментов.
• Промежуточные (окислительно-гликолитические) волокна имеют умеренную активность сукцинатдегидрогеназы.
• Быстрые мышечные волокна имеют высокую активность АТФазы миозина.
• Медленные волокна имеют низкую АТФазную активность миозина. Реально мышечные волокна содержат комбинации различных характеристик. Поэтому на практике различают три типа мышечных волокон - быстросокращающиеся красные, быстросокращающиеся белые и медленносокращающиеся промежуточные.
РЕГЕНЕРАЦИЯ И ТРАНСПЛАНТАЦИЯ МЫШЦ
• Физиологическая регенерация. В скелетной мышце постоянно происходит физиологическая регенерация - обновление мышечных волокон. При этом клетки-сателлиты вступают в циклы пролиферации с последующей дифференцировкой в миобласты и их включением в состав предсуществующих мышечных волокон.
• Репаративная регенерация. После гибели мышечного волокна под сохранившейся базальной мембраной активированные клетки-сателлиты дифференцируются в миобласты. Далее постмитотические миобласты сливаются, образуя мышечные трубочки. Синтез сократительных белков начинается в миобластах, а в мышечных трубочках происходят сборка миофибрилл и образование саркомеров. Миграция ядер на периферию и формирование нервно-мышечного синапса завершают образование зрелых мышечных волокон. Таким образом, в ходе репаративной регенерации происходит повторение событий эмбрионального миогенеза.
• Трансплантация. При пересадке мышц используют лоскут из широчайшей мышцы спины. Извлечённый из ложа вместе с собствен-
ными сосудами и нервом лоскут трансплантируют в место дефекта мышечной ткани. Начинают применять и перенос камбиальных клеток. Так, при наследственных мышечных дистрофиях в дефектные по гену дистрофина мышцы вводят нормальные по этому признаку в0-миобласты. При таком подходе рассчитывают на постепенное обновление дефектных мышечных волокон нормальными.
Сердечная мышечная ткань
Поперечно-полосатая мышечная ткань сердечного типа образует мышечную оболочку стенки сердца (миокард). Основной гистологический элемент - кардиомиоцит.
Кардиомиогенез. Миобласты происходят из клеток спланхнической мезодермы, окружающей эндокардиальную трубку. После ряда митотических делений Gj-ми- областы начинают синтез сократительных и вспомогательных белков и через стадию G0-миобластов дифференцируются в кардиомиоциты, приобретая вытянутую форму. В отличие от поперечно-полосатой мышечной ткани скелетного типа, в кардиомиогенезе не происходит обособления камбиального резерва, а все кардиомиоциты необратимо находятся в фазе G0 клеточного цикла.
КАРДИОМИОЦИТЫ
Клетки (рис. 7-21) расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры бассейна венечных сосудов и терминальные ветвления двигательных аксонов нервных клеток вегетативного отдела нервной
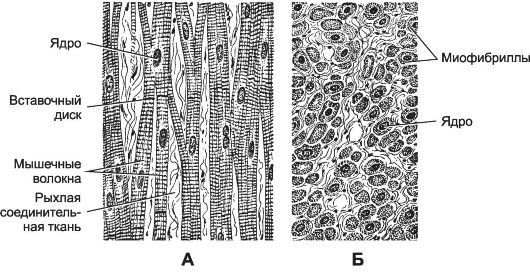 Рис. 7-21. Сердечная мышца в продольном (А) и поперечном (Б) разрезе. [17]
Рис. 7-21. Сердечная мышца в продольном (А) и поперечном (Б) разрезе. [17]
системы. Каждый миоцит имеет сарколемму (базальная мембрана + плазмолемма). Различают рабочие, атипичные и секреторные кардиомиоциты.
Рабочие кардиомиоциты
Рабочие кардиомиоциты - морфо-функциональные единицы сердечной мышечной ткани, имеют цилиндрическую ветвящуюся форму диаметром около 15 мкм (рис. 7-22). При помощи межклеточных контактов (вставочные диски) рабочие кардиомиоциты объединены в так называемые сердечные мышечные волокна - функциональный синцитий - совокупность кардиомиоцитов в пределах каждой камеры сердца. Клетки содержат центрально расположенные, вытянутые вдоль оси одно или два ядра, миофибриллы и ассоциированные с ними цистерны саркоплазматического ретикулума (депо Ca2+). Многочисленные митохондрии залегают параллельными рядами между миофибриллами. Их более плотные скопления наблюдают на уровне I-дисков и ядер. Гранулы гликогена сконцентрированы на обоих полюсах ядра. Т-трубочки в кардиомиоцитах - в отличие от скелетных мышечных волокон - проходят на уровне Z-линий. В связи с этим Т-трубочка контактирует только с одной терминальной цистерной. В результате вместо триад скелетного мышечного волокна формируются диады.
• Сократительный аппарат. Организация миофибрилл и саркомеров в кардиомиоцитах такая же, что и в скелетном мышечном волокне. Одинаков и механизм взаимодействия тонких и толстых нитей при сокращении.
• Вставочные диски. На концах контактирующих кардиомиоцитов имеются интердигитации (пальцевидные выпячивания и углубления). Вырост одной клетки плотно входит в углубление другой. На конце такого выступа (поперечный участок вставочного диска) сконцентрированы контакты двух типов: десмосомы и промежуточные. На боковой поверхности выступа (продольный участок вставочного диска) имеется множество щелевых контактов (nexus, нексус), передающих возбуждение от кардиомиоцита к кардиомиоциту.
Предсердные и желудочковые кардиомиоциты. Предсердные и желудочковые кардиомиоциты относятся к разным популяциям рабочих кардиомиоцитов. Предсердные кардиомиоциты относительно мелкие, 10 мкм в диаметре и длиной 20 мкм. В них слабее развита система Т-трубочек, но в зоне вставочных дисков значительно больше щелевых контактов. Желудочковые кардиомиоциты крупнее (25 мкм в диаметре и до 140 мкм в длину), они имеют хорошо развитую систему Т-трубочек. В состав сократительного аппарата миоцитов предсердий и желудочков входят разные изоформы миозина, актина и других контрактильных белков.
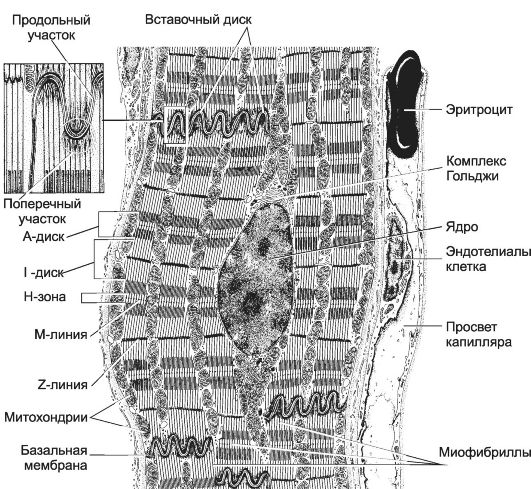 Рис. 7-22. Рабочий кардиомиоцит -
удлинённой формы клетка. Ядро расположено центрально, вблизи ядра
находятся комплекс Гольджи и гранулы гликогена. Между миофибриллами
лежат многочисленные митохондрии. Вставочные диски (на врезке) служат
для скрепления кардиомиоцитов и синхронизации их сокращения. [17]
Рис. 7-22. Рабочий кардиомиоцит -
удлинённой формы клетка. Ядро расположено центрально, вблизи ядра
находятся комплекс Гольджи и гранулы гликогена. Между миофибриллами
лежат многочисленные митохондрии. Вставочные диски (на врезке) служат
для скрепления кардиомиоцитов и синхронизации их сокращения. [17]
Секреторные кардиомиоциты. В части кардиомиоцитов предсердий (особенно правого) у полюсов ядер располагаются хорошо выраженный комплекс Гольджи и секреторные гранулы, содержащие атриопептин - гормон, регулирующий артериальное давление (АД). При повышении АД стенка предсердия сильно растягивается, что стимулирует предсердные кардиомиоциты к синтезу и секреции атриопептина, вызывающего снижение АД.
Атипичные кардиомиоциты
Этот устаревший термин относится к миоцитам, формирующим проводящую систему сердца (см. рис. 10-14). Среди них различают водители ритма и проводящие миоциты.
Водители ритма (пейсмейкерные клетки, пейсмейкеры, рис. 7-24) - совокупность специализированных кардиомиоцитов в виде тонких волокон, окружённых рыхлой соединительной тканью. По сравнению с рабочими кардиомиоцитами они имеют меньшие размеры. В саркоплазме содержится сравнительно мало гликогена и небольшое количество миофибрилл, лежащих в основном по периферии клеток. Эти клетки имеют богатую васкуляризацию и двигательную вегетативную иннервацию. Главное свойство водителей ритма - спонтанная деполяризация плазматической мембраны. При достижении критического значения возникает потенциал действия, распространяющийся через электрические синапсы (щелевые контакты) по волокнам проводящей системы сердца и достигающий рабочих кардиомиоцитов. Проводящие кардиомиоциты - специализированные клетки предсердно-желудочкового пучка Гиса и волокон Пуркинье образуют длинные волокна, выполняющие функцию проведения возбуждения от водителей ритма.
• Предсердно-желудочковый пучок. Кардиомиоциты этого пучка проводят возбуждение от водителей ритма к волокнам Пуркинье, содержат относительно длинные миофибриллы, имеющие спиральный ход; мелкие митохондрии и небольшое количество гликогена.
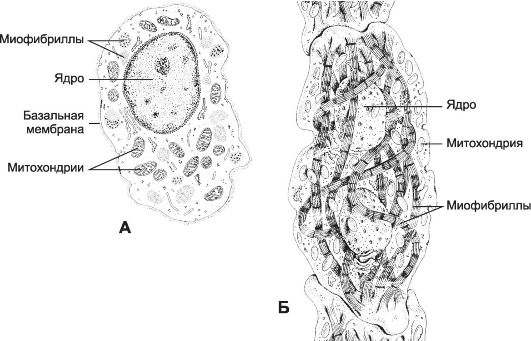 Рис. 7-24. Атипичные кардиомиоциты. А - водитель ритма синусно-предсердного узла; Б - проводящий кардиомиоцит предсердно-желудочкового пучка. [17]
Рис. 7-24. Атипичные кардиомиоциты. А - водитель ритма синусно-предсердного узла; Б - проводящий кардиомиоцит предсердно-желудочкового пучка. [17]
• Волокна Пуркинье. Проводящие кардиомиоциты волокон Пуркинье - самые крупные клетки миокарда. В них содержатся редкая неупорядоченная сеть миофибрилл, многочисленные мелкие митохондрии, большое количество гликогена. Кардиомиоциты волокон Пуркинье не имеют Т-трубочек и не образуют вставочных дисков. Они связаны при помощи десмосом и щелевых контактов. Последние занимают значительную площадь контактирующих клеток, что обеспечивает высокую скорость проведения импульса по волокнам Пуркинье.
ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА
На деятельность сердца - сложной авторегуляторной и регулируемой системы - оказывает модулирующее влияние множество факторов, в т.ч. двигательная вегетативная иннервация - парасимпатическая и симпатическая.
Парасимпатическая иннервация осуществляется блуждающим нервом, а симпатическая - адренергическими нейронами шейного верхнего, шейного среднего и звездчатого (шейно-грудного) ганглиев. Терминальные отделы аксонов вблизи кардиомиоцитов имеют варикозные расширения (см. рис. 7-29), регулярно расположенные по длине аксона на расстоянии 5-15 мкм друг от друга. Вегетативные нейроны не образуют нервно-мышечных синапсов, характерных для скелетной мышцы. Варикозности содержат нейромедиаторы, откуда и происходит их секреция. Расстояние от варикозностей до кардиомиоцитов в среднем составляет около 1 мкм. Молекулы нейромедиаторов высвобождаются в межклеточное пространство и путём диффузии достигают своих рецепторов в плазмолемме кардиомиоцитов. Парасимпатическая иннервация сердца. Преганглионарные волокна, идущие в составе блуждающего нерва, заканчиваются на нейронах сердечного сплетения и в стенке предсердий. Постганглионарные волокна преимущественно иннервируют синусно-предсердный узел, предсердно-желудочковый узел и предсердные кардиомиоциты. Парасимпатическое влияние вызывает уменьшение частоты генерации импульсов пейсмейкерами (отрицательный хронотропный эффект), снижение скорости проведения импульса через предсердно-желудочковый узел (отрицательный дромотропный эффект) в волокнах Пуркинье, уменьшение силы сокращения рабочих предсердных кардиомиоцитов (отрицательный инотропный эффект). Симпатическая иннервация сердца. Преганглионарные волокна нейронов интермедиолатеральных столбов серого вещества спинного мозга образуют синапсы с нейронами паравертебральных ганглиев. Постганглионарные волокна нейронов среднего шейного и звездчатого ганглиев иннервируют синусно-предсердный узел, предсердно-желудочковый узел, предсердные и желудочковые кардиомиоциты. Активация симпатических нервов вызывает увеличение частоты спонтанной деполяризации мембран водителей ритма (положительный хронотропный эффект), облегчение проведения импульса через предсердно-желудочковый узел (положи-
тельный дромотропный эффект) в волокнах Пуркинье, увеличение силы сокращения предсердных и желудочковых кардиомиоцитов (положительный инотропный эффект).
Гладкая мышечная ткань
Основной гистологический элемент гладкомышечной ткани - гладкомышечная клетка (ГМК), способная к гипертрофии и регенерации, а также к синтезу и секреции молекул межклеточного матрикса. ГМК в составе гладких мышц формируют мышечную стенку полых и трубчатых органов, контролируя их моторику и величину просвета. Регуляцию сократительной активности ГМК осуществляют двигательная вегетативная иннервация и множество гуморальных факторов. Развитие. Камбиальные клетки эмбриона и плода (спланхномезодерма, мезенхима, нейроэктодерма) в местах закладки гладкой мускулатуры дифференцируются в миобласты, а затем - в зрелые ГМК, приобретающие вытянутую форму; их сократительные и вспомогательные белки формируют миофиламенты. ГМК в составе гладких мышц находятся в фазе G1 клеточного цикла и способны к пролиферации.
ГЛАДКОМЫШЕЧНАЯ КЛЕТКА
Морфо-функциональная единица гладкой мышечной ткани - ГМК. Заострёнными концами ГМК вклиниваются между соседними клетками и образуют мышечные пучки, в свою очередь формирующие слои гладкой мускулатуры (рис. 7-26). В волокнистой соединительной ткани между миоцитами и мышечными пучками проходят нервы, кровеносные и лимфатические сосуды. Встречаются и единичные ГМК, например, в подэндотелиальном слое сосудов. Форма ГМК - вытя-
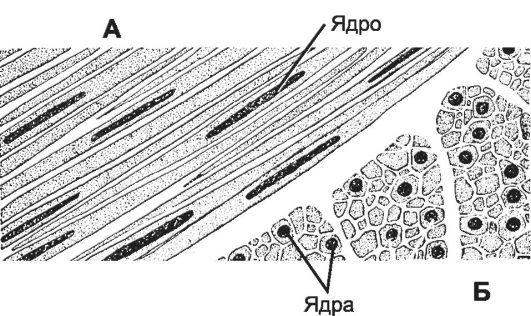 Рис. 7-26. Гладкая мышца в продольном (А) и поперечном (Б) разрезе. На поперечном срезе миофиламенты видны как точки в цитоплазме гладкомышечных клеток. [17]
Рис. 7-26. Гладкая мышца в продольном (А) и поперечном (Б) разрезе. На поперечном срезе миофиламенты видны как точки в цитоплазме гладкомышечных клеток. [17]
нутая веретеновидная, часто отростчатая (рис. 7-27). Длина ГМК от 20 мкм до 1 мм (например, ГМК матки при беременности). Овальное ядро локализовано центрально. В саркоплазме у полюсов ядра расположены хорошо выраженный комплекс Гольджи, многочисленные митохондрии, свободные рибосомы, саркоплазматический ретикулум. Миофиламенты ориентированы вдоль продольной оси клетки. Базальная мембрана, окружающая ГМК, содержит протеогликаны, коллагены типов III и V. Компоненты базальной мембраны и эластин межклеточного вещества гладких мышц синтезируются как самими ГМК, так и фибробластами соединительной ткани.
Сократительный аппарат
В ГМК актиновые и миозиновые нити не формируют миофибрилл, характерных для поперечно-полосатой мышечной ткани. Молекулы
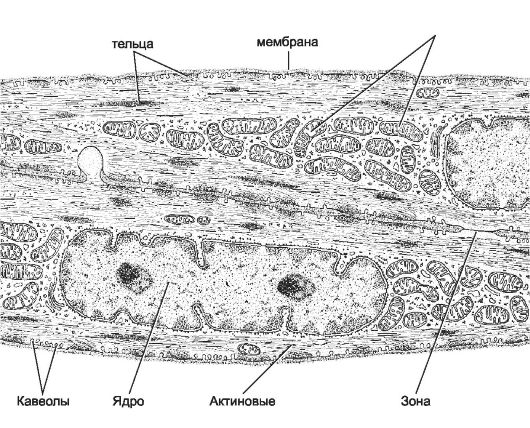
Рис. 7-27. Гладкомышечная клетка. Центральное положение в ГМК занимает крупное ядро. У полюсов ядра находятся митохондрии, эндоплазматический ретикулум и комплекс Гольджи. Актиновые миофиламенты, ориентированные вдоль продольной оси клетки, прикреплены к плотным тельцам. Миоциты формируют между собой щелевые контакты. [17]
гладкомышечного актина образуют стабильные актиновые нити, при- креплённые к плотным тельцам и ориентированные преимущественно вдоль продольной оси ГМК. Миозиновые нити формируются между стабильными актиновыми миофиламентами только при сокращении ГМК. Сборку толстых (миозиновых) нитей и взаимодействие актиновых и миозиновых нитей активируют ионы кальция, поступающие из депо Са2+. Непременные компоненты сократительного аппарата - кальмодулин (Са2+-связывающий белок), киназа и фосфатаза лёгкой цепи гладкомышечного миозина.
Депо Ca2+ - совокупность длинных узких трубочек (саркоплазматический ретикулум) и находящихся под сарколеммой многочисленных мелких пузырьков (кавеолы). Са2+-АТФаза постоянно откачивает Са2+ из цитоплазмы ГМК в цистерны саркоплазматического ретикулума. Через Са2+-каналы кальциевых депо ионы Са2+ поступают в цитоплазму ГМК. Активация Са2+-каналов происходит при изменении мембранного потенциала и при помощи рецепторов рианодина и инозитолтрифосфата. Плотные тельца (рис. 7-28). В саркоплазме и на внутренней стороне плазмолеммы находятся плотные тельца - аналог Z-линий попереч-
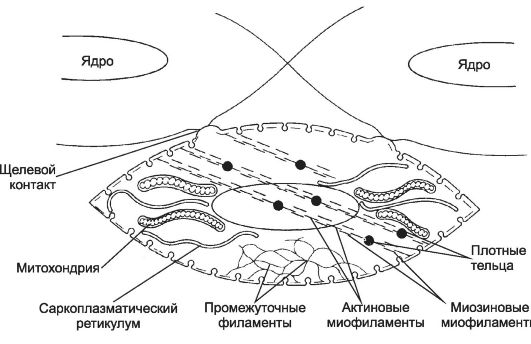 Рис. 7-28. Сократительный аппарат гладкомышечной клетки. Плотные
тельца содержат α-актинин, это аналоги Z-линий поперечно-полосатой
мышцы. В саркоплазме они связаны сетью промежуточных филаментов, в
местах их прикрепления к плазматической мембране присутствует винкулин.
Актиновые нити прикреплены к плотным тельцам, миозиновые миофиламенты
формируются при сокращении. [17]
Рис. 7-28. Сократительный аппарат гладкомышечной клетки. Плотные
тельца содержат α-актинин, это аналоги Z-линий поперечно-полосатой
мышцы. В саркоплазме они связаны сетью промежуточных филаментов, в
местах их прикрепления к плазматической мембране присутствует винкулин.
Актиновые нити прикреплены к плотным тельцам, миозиновые миофиламенты
формируются при сокращении. [17]
но-полосатой мышечной ткани. Плотные тельца содержат α-актинин и служат для прикрепления тонких (актиновых) нитей. Щелевые контакты связывают соседние ГМК и необходимы для проведения возбуждения (ионный ток), запускающего сокращение ГМК.
Сокращение
В ГМК, как и в других мышечных тканях, работает актомиозиновый хемомеханический преобразователь, но АТФазная активность миозина в гладкомышечной ткани приблизительно на порядок величины ниже активности АТФазы миозина поперечно-полосатой мышцы. Медленное образование и разрушение актин-миозиновых мостиков требуют меньшего количества АТФ. Отсюда, а также из факта лабильности миозиновых нитей (их постоянная сборка и разборка при сокращении и расслаблении соответственно) вытекает важное обстоятельство - в ГМК медленно развивается и длительно поддерживается сокращение. При поступлении сигнала к ГМК сокращение клетки запускают ионы кальция, поступающие из кальциевых депо. Рецептор Са2+ - кальмодулин.
Расслабление
Лиганды (атриопептин, брадикинин, гистамин, VIP) связываются с их рецепторами и активируют G-белок (Gs), который в свою очередь активирует аденилатциклазу, катализирующую образование цАМФ. Последний активирует работу кальциевых насосов, откачивающих Са2+ из саркоплазмы в полость саркоплазматического ретикулума. При низкой концентрации Са2+ в саркоплазме фосфатаза лёгких цепей миозина осуществляет дефосфорилирование лёгкой цепи миозина, что приводит к инактивации молекулы миозина. Дефосфорилированный миозин теряет сродство к актину, что предотвращает образование поперечных мостиков. Расслабление ГМК заканчивается разборкой миозиновых нитей.
ИННЕРВАЦИЯ
Симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна иннервируют ГМК. Нейромедиаторы диффундируют из варикозных терминальных расширений нервных волокон в межклеточное пространство. Последующее взаимодействие нейромедиаторов с их рецепторами в плазмолемме вызывает сокращение либо расслабление ГМК. Существенно, что в составе многих гладких мышц, как правило, иннервированы (точнее находятся рядом с варикозными терминалями аксонов) далеко не все ГМК. Возбуждение ГМК, не имеющих иннервации, происходит двояко: в меньшей степени - при медленной диффузии нейромедиаторов, в большей степени - посредством щелевых контактов между ГМК.
ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ
Рецепторы плазмолеммы ГМК многочисленны. В мембрану ГМК встроены рецепторы ацетилхолина, гистамина, атриопептина, ангиотензина, адреналина, норадреналина, вазопрессина и множество других. Агонисты, связываясь со своими ре-
цепторами в мембране ГМК, вызывают сокращение или расслабление ГМК. ГМК разных органов различно реагируют (сокращением либо расслаблением) на одни и те же лиганды. Это обстоятельство объясняется тем, что существуют разные подтипы конкретных рецепторов с характерным распределением в разных органах.
ТИПЫ МИОЦИТОВ
• В основе классификации ГМК находятся различия в их происхождении, локализации, иннервации, функциональных и биохимических свойствах. По характеру иннервации гладкие мышцы делятся на единично и множественно иннервированные (рис. 7-29). Единично иннервированные гладкие мышцы. Гладкие мышцы желудочно-кишечного тракта, матки, мочеточника, мочевого пузыря состоят из ГМК, образующих многочисленные щелевые контакты друг с другом, формируя большие функциональные единицы для синхронизации сокращения. При этом прямую двигательную иннервацию получают лишь отдельные ГМК функционального синцития.
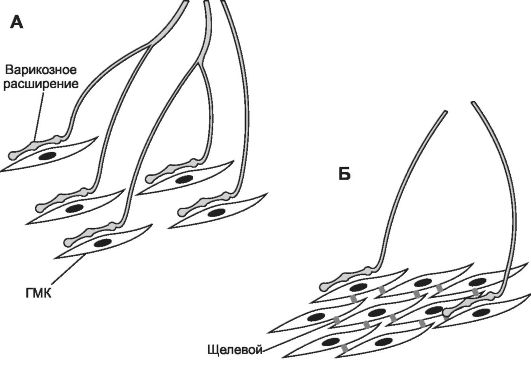
Рис. 7-29. Иннервация гладкомышечной ткани. А. Множественно иннервированная гладкая мышца. Каждая ГМК получает двигательную иннервацию, щелевые контакты между ГМК отсутствуют. Б. Единично иннервированная гладкая мышца. Ин-
нервированы лишь отдельные ГМК. Смежные клетки связаны многочисленными щелевыми контактами, образующими электрические синапсы. [114]
• Множественно иннервированные гладкие мышцы. Каждая ГМК мышцы радужки (расширяющие и суживающие зрачок) и семявыносящего протока получает двигательную иннервацию, что позволяет осуществлять тонкую регуляцию сокращения мышц.
• Висцеральные ГМК происходят из мезенхимных клеток спланхнической мезодермы и присутствуют в стенке полых органов пищеварительной, дыхательной, выделительной и половой систем. Многочисленные щелевые контакты компенсируют сравнительно бедную иннервацию висцеральных ГМК, обеспечивая вовлечение всех ГМК в процесс сокращения. Сокращение ГМК медленное, волнообразное. Промежуточные филаменты образованы десмином.
• ГМК кровеносных сосудов развиваются из мезенхимы кровяных островков. ГМК образуют единично иннервированную гладкую мышцу, но функциональные единицы не такие большие как в висцеральной мускулатуре. Сокращение ГМК сосудистой стенки опосредуют иннервация и гуморальные факторы. Промежуточные филаменты содержат виментин.
РЕГЕНЕРАЦИЯ
Вероятно, среди зрелых ГМК присутствуют недифференцированные предшественники, способные к пролиферации и дифференцировке в дефинитивные ГМК. Более того, дефинитивные ГМК потенциально способны к пролиферации. Новые ГМК возникают при репаративной и физиологической регенерации. Так, при беременности в миометрии происходит не только гипертрофия ГМК, но и значительно увеличивается их общее количество.
Немышечные сокращающиеся клетки Миоэпителиальные клетки
Миоэпителиальные клетки имеют эктодермальный генез и экспрессируют белки, характерные и для эктодермального эпителия (цитокератины 5, 14, 17), и для ГМК (гладкомышечные актин, α-актинин). Миоэпителиальные клетки окружают секреторные отделы и выводные протоки слюнных, слёзных, потовых, молочных желёз, прикрепляясь при помощи полудесмосом к базальной мембране. От тела клетки отходят отростки, охватывающие эпителиальные клетки желёз (рис. 7-30). Стабильные актиновые миофиламенты, прикреплённые к плотным тельцам, и нестабильные миозиновые, формирующиеся в процессе сокращения, - сократительный аппарат миоэпителиальных клеток. Сокращаясь, миоэпителиальные клетки способствуют продвижению секрета из концевых отделов по выводным протокам желёз. Ацетил-
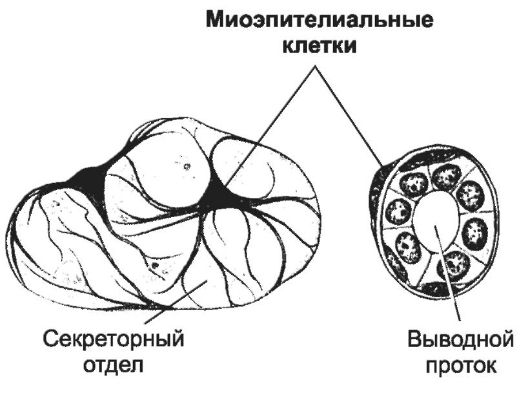 Рис. 7-30. Миоэпителиальная клетка. Корзинчатой
формы клетка окружает секреторные отделы и выводные протоки желёз.
Клетка способна к сокращению, обеспечивает выведение секрета из
концевого отдела. [17]
Рис. 7-30. Миоэпителиальная клетка. Корзинчатой
формы клетка окружает секреторные отделы и выводные протоки желёз.
Клетка способна к сокращению, обеспечивает выведение секрета из
концевого отдела. [17]
холин стимулирует сокращение миоэпителиальных клеток слёзных и потовых желёз, норадреналин - слюнных желёз, окситоцин - лактирующих молочных желёз.
Миофибробласты
Миофибробласты проявляют свойства фибробластов и ГМК. Их находят в разных органах (например, в слизистой оболочке кишечника эти клетки известны как «перикриптальные фибробласты»). При заживлении раны часть фибробластов начинает синтезировать гладкомышечные актины и миозины и тем самым способствуют сближению раневых поверхностей.