Иммунология: структура и функции иммунной системы: учебное пособие / Р.М. Хаитов. - 2013. - 280 с.
|
|
|
|
Глава 7. АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
Лейкоцитарные антигены человека (HLA - Human Leukocyte Antigens) - гликопротеины, кодируемые генами главного комплекса гистосовместимости - MHC (Major Histocompatibility Complex). Они определяют биологическую индивидуальность каждого человека. В иммунной системе эти гликопротеины выполняют важнейшую функцию: они участвуют в презентации АПК пептидных антигенов Т-лимфоцитам.
ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ
HLA-молекулы, кодируемые генами MHC, подразделяют на гликопротеины MHC класса I (HLA-A, HLA-B и HLA-C), представленные на поверхности всех соматических клеток за исключением клеток ворсинчатого трофобласта и эритроцитов, и MHC класса II (HLA-DP, HLA-DQ и HLA-DR), экспрессированные преимущественно на мембране АПК (ДК, активированных макрофагов и В-лимфоцитов), а также на некоторых T-хелперах.
Соматические клетки в нормальных условиях молекулы MHC-II не экспрессируют. T-лимфоциты с помощью антигенраспознающих рецепторов способны распознавать антигены только в комплексе с молекулами MHC-I или MHC-II на поверхности клеток своего организма - феномен двойного распознавания. Таким образом, природная функция белков MHC - презентация пептидных антигенов T-лимфоцитам.
Первоначально главный комплекс гистосовместимости (MHC) был открыт в опытах по пересадке органов как некий значительный по размеру комплекс генов, от которых больше, чем от других генов, зависит скорость отторжения трансплантата.
Законы трансплантации, описанные в 1912 г. Георгом Шёне в книге «Гетеропластическая и гомопластическая трансплантация»:
• аллогенные и ксеногенные трансплантаты всегда отторгаются;
• аутологичные (донор и реципиент - один и тот же организм) и сингенные (антигены MHC донора и реципиента одинаковы) трансплантаты приживаются;
• гибриды 1-го поколения - F1 (P1×P2) - не отторгают трансплантаты обеих родительских линий (P1, P2), однако каждая из родительских линий отторгает трансплантат от гибрида (это свидетельствует о кодоминантности в гибридах экспрессии генов, кодирующих продукты, служащие мишенью при отторжении). Кодоминантность означает, что экспрессируются аллели на двух гомологичных хромосомах, т.е. от каждого из родителей:
AA × BB → AB,
где А и В - аллели, перешедшие к гибриду F1 от родителей (Р1 и Р2 соответственно).
• При обратном скрещивании (F1 × P1 или F1 × P2) 50% потомства быстро отторгают трансплантат от второй родительской линии (при трансплантации кожного лоскута быстрое отторжение наступает на 8-е сутки). Из этого по законам менделевской генетики следует, что быстрое отторжение контролирует один локус. Его-то и назвали главным комплексом гистосовместимости. А генетическая формула выглядит так:
AB × AA → AA, AA, AB, AB. Гены главного комплекса гистосовместимости
Комплекс MHC весьма значителен по размеру. Он включает около 2000 аллельных вариантов генов. Расположение в геноме локусов комплекса MHC человека схематично изображено на рис. 7-1.
• MHC-I. Гены групп HLA-A, HLA-B, HLA-C и другие кодируют молекулы MHC класса I.
• MHC-II. Гены групп HLA-DP, HLA-DQ и HLA-DR кодируют молекулы MHC класса II.
• MHC-III. Термин «MHC-III» обозначает область между MHC-I и MHC-II, где картированы гены, кодирующие некоторые компоненты системы комплемента (C4a и C4b, C2, фактор В), цитокины (ФНОα и лимфотоксин), 21-гидроксилазу (фермент, участвующий в биосинтезе стероидных гормонов) и др.
• Неклассические гены не принадлежат ни к одному из классов MHC. Описано 6 таких генов в области расположения генов MHC-I (Е, F, G, Н, J, X) и 6 - в области MHC-II (DM, DO, CLIP, TAP, LMP,
LNA).
Трансмембранные молекулы главного комплекса гистосовместимости
• Молекула MHC-I (см. рис. 5-1, а) состоит из двух полипептидных цепей - α-цепи и β2-микроглобулина.
- α-Цепь состоит примерно из 325 аминокислотных остатков, имеет молекулярную массу около 45 кДа и содержит 3 внеклеточных домена (α1, α2, α3), трансмембранный участок и цитоплазматический участок из 55 аминокислотных остатков. Домены α1 и α2 формируют углубление в виде желоба или «корзины» размером 0,25×0,1×0,11 нм для связывания антигенных пептидов длиной 9-11 аминокислот, презентируемых для распознавания T-лимфоцитам.
- С внеклеточными α-доменами нековалентно ассоциирована лёгкая полипептидная цепь с молекулярной массой 11,5 кДа, называемая β2-микроглобулином, которая кодируется геном, расположенным вне комплекса MHC, на хромосоме 15.
• Молекулы MHC-II (см. рис. 5-1, б) - гетеродимеры из двух трансмембранных гликопротеинов: α-цепи (молекулярная масса 34 кДа) и β-цепи (молекулярная масса 29 кДа). Внеклеточная часть каждой
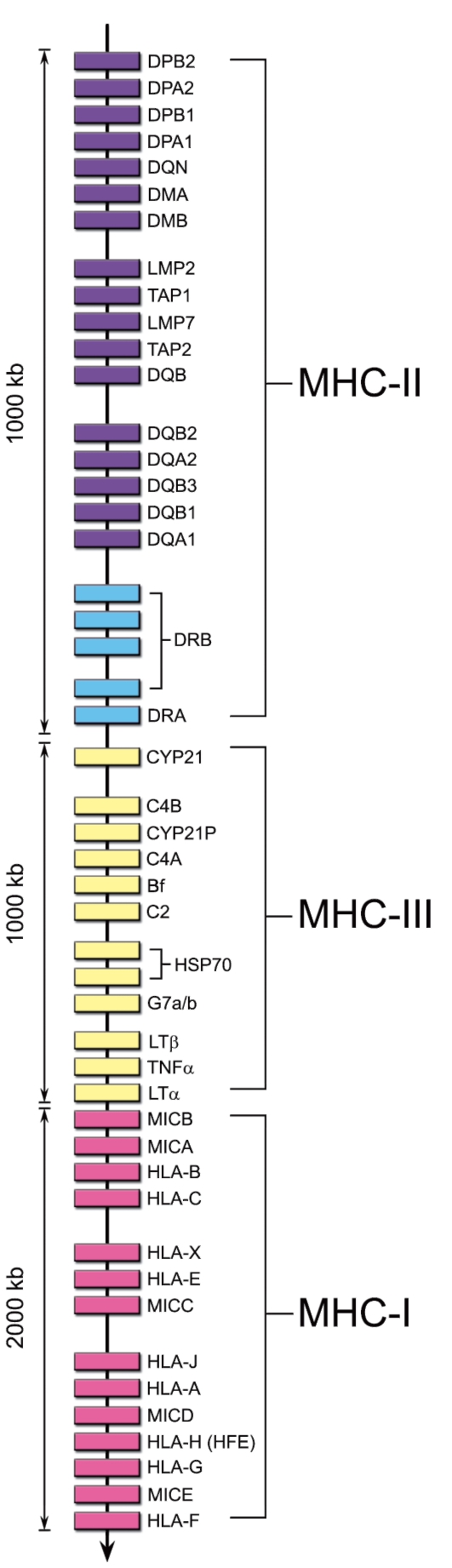
Рис. 7-1. Расположение генов MHC в хромосоме 6 человека
цепи имеет 2 домена. Пептидсвязывающую бороздку формируют совместно α1- и β1-домены. В отличие от аналогичного углубления в составе гликопротеинов MHC-I, пептидсвязывающая щель MHC-II открыта с обеих сторон, что позволяет связывать более длинные пептиды (15-30 аминокислот).
Генетический полиморфизм
Гены комплекса MHC, в отличие от генов TCR и иммуноглобулинов, не подвергаются рекомбинации. Механизм их приспособления к неограниченному множеству потенциальных антигенов заключается в их генетическом полиморфизме (кодоминантность, множественность аллелей, естественный отбор).
• Кодоминантность. Гены MHC кодоминантны, т.е. одновременно экспрессируются гены материнской и отцовской хромосом. Генов MHC-I по 3 (А, В, С) в каждой из гомологичных хромосом, генов MHC-II - также по 3 (DP, DQ, DR). Следовательно, если у матери и отца нет одинаковых аллелей, каждый человек имеет как минимум 12 различных основных аллелей генов MHC.
• Множественность аллелей. Полиморфизм классических генов MHC (классов I и II) означает наличие в популяции множества аллелей - вариантов одноимённого гена у разных особей. Для отдельных локусов MHC известно более сотни аллельных вариантов. Каждый аллельный вариант молекулы MHC преимущественно связывает пептиды с определёнными «якорными» аминокислотными остатками.
• Эволюционный отбор. Каждая особь потенциально способна развить иммунитет только на те пептиды, которые способны связать гликопротеины MHC этой особи. Конкретные варианты MHC закрепляются в эволюции естественным отбором, и каждая отдельная особь оказывается приспособленной к региональным видам и штаммам инфекционных микроорганизмов, на защиту от которых шёл отбор MHC у предков.
• Неклассические гены MHC, видимо, не полиморфны или не столь полиморфны, как гены MHC классов I и II.
Комплексы антигенный пептид-MHC
Формирование комплексов антигенных пептидов с молекулами MHC разных классов происходит в различных клеточных компартментах.
• MHC-I «обслуживают» зону цитозоля, сообщающегося через ядерные поры с содержимым ядра. Здесь происходит фолдинг (принятие правильной конформации) синтезированных белковых молекул. При возникновении ошибок, в том числе и при синтезе вирусных белков, белковые продукты расщепляются в мультипротеазных комплексах (протеосомы). Образующиеся при этом пептиды связываются с молекулами MHC-I. В результате молекулы MHC-I презентируют T-лимфоцитам пептидные антигены, образующиеся внутри клетки. Именно поэтому CD8+ Т-лимфоциты, распознающие комплексы антигенов с MHC-I, участвуют в первую очередь в защите от вирусных, а также от внутриклеточных бактериальных инфекционных заболеваний.
• MHC-II. Зона «обслуживания» MHC-II связана с внеклеточной средой, а также с клеточными органеллами (аппарат Гольджи, эндоплазматический ретикулум, лизосомы, эндосомы и фагосомы). Пептиды, образующиеся в данной зоне, имеют внеклеточное происхождение - это продукты протеолиза белков, захваченных клеткой посредством эндоцитоза. Вновь синтезированные молекулы α- и β-цепей MHC-II с помощью инвариантной полипептидной цепи Ii и кальнексина экспонируются внутрь везикул (эндосом или фаголизосом) и только здесь, связавшись с пептидным антигеном, принимают необходимую конформацию для дальнейшего представления на мембране клетки. Таким образом, молекулы MHC-II презентируют антиген при развитии иммунного ответа на внеклеточные патогены. Главную роль в этом ответе играют CD4+ Т-лимфоциты, распознающие антиген в комплексе с
MHC-II. Суперантигены
В ходе эволюции патогенные микроорганизмы «научились» синтезировать так называемые суперантигены, способные присоединяться к молекулам MHC-II без процессинга и связываться с V-областью Р-цепи TCR. Такое связывание вызывает поликлональную активацию 2-20% периферических CD4+ T-лимфоцитов, что приводит, с одной стороны, к общей интоксикации организма, а с другой - к быстрой гибели активированных T-лимфоцитов путём апоптоза и, следовательно, к иммунодефициту. Например, клиническая картина пищевых отравлений обусловлена главным образом действием энтеротоксинов стафилококков на T-лимфоциты. Иммунодефицит при ВИЧ-инфекции также в
значительной мере объясняется лимфотоксическим действием суперантигенов ВИЧ.
Антигенпрезентирующие молекулы CD1
За пределами MHC имеются гены, кодирующие антигенпрезентирующие молекулы (например, гены кластера CD1, кодирующие CD1a, CD1b, CD1c, CD1d, CD1e). Продукты этих генов - гетеродимеры (состоят из α-цепи и β2-микроглобулина). Они способны связывать и презентировать T-лимфоцитам небелковые антигены - например, миколевую кислоту и липоарабиноманнан оболочки микобактерий. Эти молекулы-гетеродимеры конститутивно экспрессированы на ДК и B-лимфоцитах, а также на тимоцитах и энтероцитах. Антиген в комплексе с CD1 распознают γδT-лимфоциты и NKT-клетки.
АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
Молекулы MHC контролируют иммунный ответ. Так, MHC-II участвуют в презентации антигенов T-клеткам (рис. 7-3, 7-4) и во взаимодействии T- и B-лимфоцитов. Антигены MHC-I и MHC-II связываются с поверхностными маркёрами Т-клеток: MHC-I с CD8, а MHC-II - с CD4.
• Профессиональные АПК. Молекулы MHC-II экспрессированы только на определённых клетках, которые называют профессиональными АПК. Таких клеток у человека 3 типа: ДК костномозгового происхождения (ДК), B-лимфоциты и макрофаги. На их мембранах, помимо молекул MHC-I и MHC-II, присутствуют все корецепторные молекулы, необходимые для презентации антигена Т-клеткам. Они продуцируют цитокины, необходимые для активации T-лимфоцитов и запуска иммунного ответа.
• Эндотелий тоже может выполнять функции АПК. Вероятно, экспрессия комплексов пептид-MHC на клетках эндотелия служит специфическим сигналом, привлекающим эффекторные лимфоциты из циркуляции в очаг поражения, обеспечивая антигенспецифичный хоминг.
Характеристика разных АПК представлена на рис. 7-2.
Дендритные клетки
Из всех клеток, имеющих определение «дендритные» (что означает «клетки с отростками»), к профессиональным АПК относят только
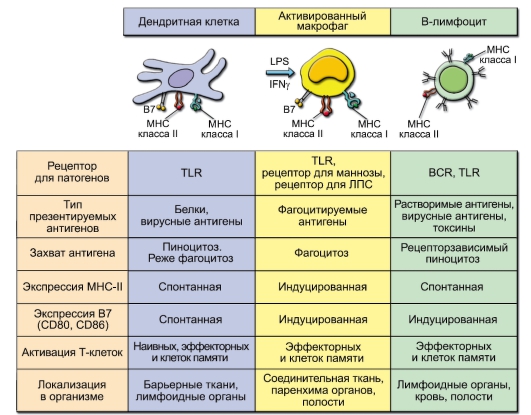
Рис. 7-2. Антигенпрезентирующие клетки. Обозначения: В7 - костимулирующие молекулы (CD80 и CD86)
клетки костномозгового происхождения. Такие клетки широко представлены в организме. Их много в покровных тканях (например, клетки Лангерганса в коже), носоглотке, лёгких, кишечнике и желудке, в крови (незрелые формы) и в лимфоидных органах (зрелые, активированные). Если у B-лимфоцитов и макрофагов есть и иные, причём главные для них, функции - продукция иммуноглобулинов у B-лимфоцитов, фагоцитоз и «переваривание» у макрофагов, - то у ДК нет других функций, кроме презентации антигенов и передачи костимулирующих сигналов лимфоцитам.
• Разновидности ДК: - Миелоидные ДК происходят из моноцитов и экпрессируют маркёр CD11c Их, вероятно, можно рассматривать как разновидность макрофагов, специализирующихся на презентации антигенов T-лимфоцитам.
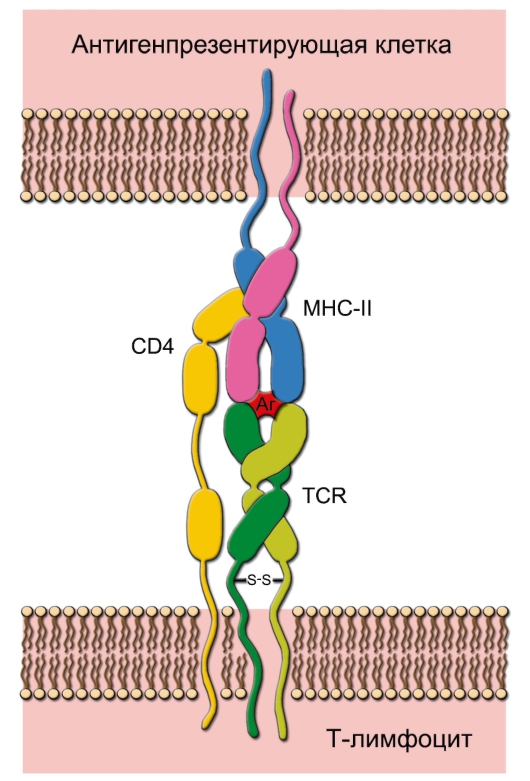
Рис. 7-3. Распознавание антигена рецептором T-лимфоцита. Каждая молекула MHC-II состоит двух цепей - α и β. При помощи TCR T-клетка распознаёт антиген, но только находящийся в комплексе с молекулой MHC. В случае T-хелпера в процессе также участвует CD4, который свободным концом связывается с молекулой MHC. Распознаваемый T-клеткой антиген имеет 2 участка: один взаимодействует с молекулой MHC, другой (эпитоп) связывается с рецептором T-лимфоцита. Подобный же тип взаимодействия, но с участием CD8 характерен для процесса распознавания цитотоксическим T-лимфоцитом антигена, связанного с молекулой MHC-I. Аг - антиген
- Плазмоцитоидные ДК происходят от общей лимфоидной клетки-предшественника, из которой развиваются также T- и B-лимфоциты, NK-клетки. Маркёры предшественников плазмоцитоидных ДК: ИЛ-3Rα (вариант рецептора для ИЛ-3, или CD123), иммуноглобулиноподобные рецепторы - ILT3+ (CD85k) и ILT7+ (CD85g), а также BDCA-2 (CD303), BDCA-3 (CD141) и BDCA-4 (CD304).
• Эндоцитоз. ДК активно и непрерывно поглощают вещества из окружающей среды. При отсутствии патогена ДК поглощают вещества собственных тканей и презентируют этот материал T-лимфоцитам без корецепторного стимула. В результате иммунный ответ в отношении собственных тканей не развивается и поддерживается состояние толерантности к «своему». ДК до активации представлены незрелыми формами, с низкой способностью презентировать антигены Т-лимфоцитам.
• Активация. ДК активируются (созревают) при проникновении патогена в организм и превращаются в зрелые ДК. В распознавании патогена при этом участвует TLR.
- На миелоидных ДК, как и на макрофагах, присутствуют рецепторы для маннозы, ЛПС, а также TLR2 и TLR4, распознающие продукты грамотрицательных и грамположительных бактерий соответственно.
- На плазмоцитоидных ДК присутствуют TLR7 и TLR9 (распознают вирусную и бактериальную ДНК), а также особый лектиновый рецептор, который связывает, например, вирус гриппа.
• Иммунный синапс. Активированные патогеном ДК (зрелые) мигрируют из покровных тканей в региональные лимфоидные органы (в частности, в паракортикальные зоны лимфатических узлов), где презентируют T-лимфоцитам антигены и активируют их через корецепторные молекулы B7.1, B7.2 и CD40. Формирование такого межклеточного - иммунного - синапса (рис. 7-4) - обязательное условие начала развития иммунного ответа.
Взаимодействие T- и В-лимфоцитов
При первичном иммунном ответе единственные эффективные АПК для T-лимфоцитов - ДК. Однако при активации T-лимфоцита антигеном, презентированным ДК, в иммунный ответ будут вовлекаться и рядом расположенные B-лимфоциты, для которых в сложившемся микроокружении тоже найдутся объекты для распознавания. При этом возможны 2 варианта взаимодействия T- и В-лимфоцитов.
• В-лимфоциты через BCR связывают растворимый антиген, поглощают его эндоцитозом, подвергают процессингу и экспонируют на поверхности фрагменты антигена в составе комплексов с молекулами MHC-II. Т-клетка через TCR распознаёт антиген на поверхности B-лимфоцита в комплексе с молекулой МНС-II. Кроме того,
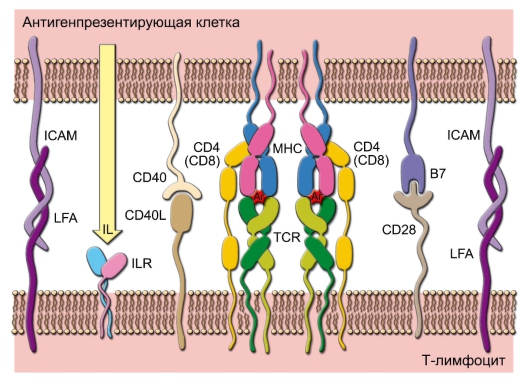
Рис. 7-4. Иммунный синапс между T-лимфоцитом и антигенпрезентирующей клеткой. Минимальный набор взаимодействий, необходимых для начала развития иммунного ответа: связывание агрегированных TCR с комплексами пептид-MHC; связывание корецептора CD4 (или CD8) с MHC; связывание CD40 с CD40L; связывание B7 с CD28; связывание молекул адгезии (ICAM, LFA); взаимодействие цитокинов (IL) с рецепторами для цитокинов (ILR)
устанавливаются все необходимые и достаточные корецепторные взаимосвязи между T- и B-лимфоцитами (табл. 7-1). Такое взаимодействие происходит в T-зависимых зонах периферической лимфоидной ткани в начале развития иммунного ответа. • B-лимфоцит распознаёт специфический антиген, однако недалеко оказывается активированный T-лимфоцит, распознавший антиген на другой АПК. В таком случае взаимодействие Т- и В-лимфоцитов может ограничиться восприятием В-клеткой цитокиновых сигналов от Т-лимфоцита. Взаимодействие же с участием мембранных молекул может наступать или не наступать, по крайней мере, при первичном иммунном ответе. При вторичном иммунном ответе в процесс вовлекаются Т-клетки памяти и обязательно происходит взаимодействие мембранной молекулы B-лимфоцита CD40 с мембранной молекулой T-клетки CD40L. Кроме T-лимфоцитов,
CD40L экспрессируется на тучных клетках, тромбоцитах, макрофагах, естественных киллерах, а также на клетках гладких мышц, эндотелиальных и эпителиальных клетках. Без этого взаимодействия, как показывает опыт, не происходит переключение с IgM на другие изотипы. Вторичный же ответ B2-лимфоцитов характеризуется обязательным переключением изотипа иммуноглобулинов с IgM на IgG, IgA или IgE. Эти взаимодействия Т- и В-клеток происходят уже в B-клеточных зонах - в фолликулах лимфоидных органов.
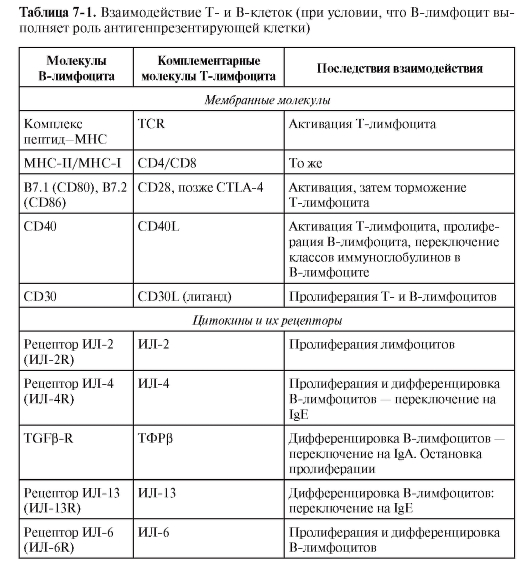
ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ РАЗВИТИЕ РАЗЛИЧНЫХ ТИПОВ ИММУННОГО ОТВЕТА
ДК в качестве АПК не только обеспечивают саму возможность развития иммунного ответа или возникновение иммунной толерантности, но и определяют направление дифференцировки хелперных субпопуляций T-лимфоцитов, а значит, тип иммунного ответа (табл. 7-2).
Характер активации АПК зависит от биохимических характеристик и дозы патогена, его биологических свойств и путей попадания в организм. В результате развивается последовательность событий, представленная на рис. 7-5.
Th1-опосредованный или клеточный иммунный ответ
При этом типе иммунного ответа доминирует секреция ИФНу, приводящая к активации макрофагов и стимуляции секреции В-лимфоцитами антител со свойствами опсонинов. Важную роль в Th1-ответе играют ЦТЛ и NK-клетки, что важно не только для противоинфекционного, но и для противоопухолевого и трансплантационного иммунитета.
Биологические эффекты ИФНу направлены на уничтожение клеток, инфицированных вирусами или бактериями:
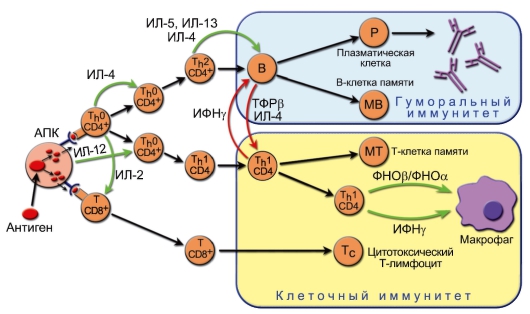
Рис. 7-5. Роль антигенпрезентирующих клеток и Т-хелперов в формировании оптимального варианта иммунного ответа. Этапы: 1 - разные патогены по-разному связываются с разными антигенпрезентирующими клетками, поразному процессируются и по-разному их активируют; 2 - активированные разными способами антигенпрезентирующие клетки продуцируют разные цитокины, воздействующие на T-лимфоциты. Кроме того, концентрации антигена на антигенпрезентирующих клетках могут значительно различаться; 3 - таким образом, направление дифференцировки эффекторного T-лимфоцита определяют и антиген, и цитокиновый сигнал от антигенпрезентирующих клеток; 4 - дифференцированные эффекторные T-лимфоциты начинают продуцировать определённый набор цитокинов, определяющий различные типы иммунного ответа. Перекрёстная регуляция: ИФНγ, образуемый Th1-клетками, подавляет пролиферацию Th2-лимфоцитов. В свою очередь, ИЛ-10, продуцируемый Th2клетками, подавляет образование Th1-цитокинов. Зелёные стрелки - активационные сигналы, красные - ингибиторные
• индукция противовирусного действия на уровне ферментов, расщепляющих нуклеиновые кислоты (2',5'-олигоаденилатсинтетазы и др.);
• сильная стимуляция макрофагов, соответственно повышенный синтез этими клетками токсичных продуктов;
• стимуляция NK-клеток;
• поддержание переключения синтеза иммуноглобулинов в B-лимфоцитах на IgG, что активирует фагоциты (нейтрофилы и макрофаги), т.е. T-лимфоциты как продуценты интерферона обе-
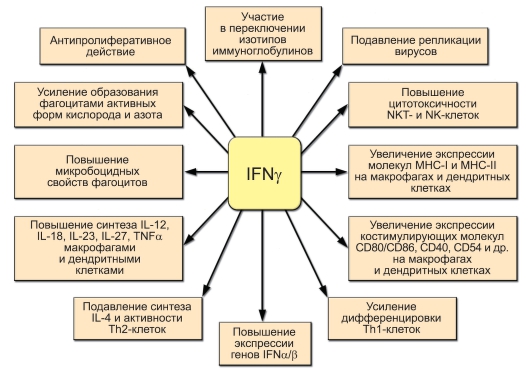
Рис. 7-6. Основные функции IFNy
спечивают макрофагальный и цитотоксический характер иммунного ответа.
Основные функции IFNy представлены на рис. 7-6.
Условия развития иммунного ответа Th1-типа
• Зрелые ДК осуществляют контроль за Т-клеточной дифференцировкой в зависимости от типа патогена, пути заражения, наличия сигналов от клеток врождённого иммунитета и тканевых факторов. В случае контакта с внутриклеточными бактериями, вирусами или паразитами миелоидные ДК, вступив в контакт с Т-лимфоцитами лимфатических узлов, начинают продуцировать ИЛ-12 и ФНОα, стимулируя T-лимфоциты к выработке ИФНγ. ИЛ-12 продуцируют также макрофаги и нейтрофилы.
• На T-лимфоците, TCR которого связал антиген с высокой авидностью, устойчиво экспрессируется рецептор для ИЛ-12, содержащий β2-субъединицу (экспрессия этой субъединицы специфична именно для Th1).
• Плазмоцитоидные ДК сразу после распознавания патогена (ещё до попадания в лимфатический узел) начинают активно продуцировать интерфероны типа I (ИФНα и ИФНβ), которые, с одной стороны, аутокринно стимулируют созревание ДК, с другой - индуцируют образование T-лимфоцитами ИФНγ и ИЛ-10, а не одного только ИФНγ, как ИЛ-12 миелоидных ДК. ИЛ-10 подавляет дифференцировку и активность Th1-лимфоцитов, возможно, не напрямую, а через угнетение активности макрофагов и возникающий при этом недостаток ИЛ-12.
• ИФНγ и ИФНα - кофакторы для дифференцировки Th1лимфоцитов, индуцируемой ИЛ-12. Их источником, помимо Th1-клеток, служат активированные CD8+ T-клетки, т.е. CD8+ Т-лимфоциты вносят свой вклад в дифференцировку CD4+ T-клеток по пути Th1.
• В свою очередь, Th1-хелперы стимулированные ДК, активируют цитотоксические CD8+ Т-лимфоциты и макрофаги, которые «расправляются» с клетками организма, заражёнными внутриклеточными патогенами.
Таким образом, большая нагрузка антигеном и достаточные количества ИЛ-12 - необходимые и, возможно, достаточные условия для индукции дифференцировки T-лимфоцитов в продуценты ИФНγ, т.е. CD4+ Th1-лимфоциты.
Th2-опосредованный или гуморальный иммунный ответ
• Иммунный ответ Th2-типа контролируется другими цитокинами, чем Th1-ответ, - в основном ИЛ-4. Продуценты ИЛ-4 - CD4+ Тh2-клетки, дважды негативные CD4-CD8- T-лимфоциты, тучные клетки. Этот тип ответа направлен на стимуляцию В-лимфоцитов к образованию антител, нейтрализующих бактерии, токсины, паразитов и реализующих другие эффекты гуморального иммунитета.
• Th2-лимфоциты поддерживают переключение синтеза изотипов иммуноглобулинов в B-лимфоцитах на IgE, IgG4 и IgА. Клеткипартнёры для этих изотипов - тучные клетки, базофилы и эозинофилы. При их активации развиваются воспалительные процессы с выраженным вазоактивным компонентом и экссудацией или характерное эозинофильное воспаление.
• За исключением патологических случаев IgE-зависимых аллергических реакций иммунный ответ Th2-типа принято (достаточно условно) рассматривать как противовоспалительный.
Условия развития иммунного ответа Th2-типа
Этот вариант иммунного ответа направлен на «перехват» растворимых компонентов патогенов (токсинов, аллергенов, химиопрепаратов), опсонизацию и фагоцитоз бактерий и вирусов, а также на элиминацию крупных инвазивных патогенов - гельминтов, которые не могут быть фагоцитированы макрофагами из-за своих размеров. При этом активация цитотоксических лимфоцитов не сообразна, и приоритет получает стимуляция B-лимфоцитов к продукции антител. Лидирующая роль в дифференцировке Т-хелперов в направлении Th2 отводится плазмоцитоидным ДК.
Интерлейкин-4 (ИЛ-4)
• СD4+ Т-лимфоциты, не получившие стимула от ИЛ-12, экспрессируют молекулу OX40. Лигандом для неё служит OX40L на мембране активированных ДК и B-лимфоцитов.
• Взаимодействие OX40-OX40L индуцирует образование T-лимфоцитами ИЛ-4 и экспрессию на В-клетках CXCR5 (рецептор для хемокина CXCL13), обеспечивающего миграцию этих В-лимфоцитов в лимфоидные фолликулы лимфатического узла, т.е. в B-клеточную зону, где происходят дальнейшие Т/B-взаимодействия.
• ИЛ-4 и ИЛ-13 аутокринно стимулируют дифференцировку наивных CD4+ T-лимфоцитов в Th2-клетки.
Интерлейкин-5 (ИЛ-5)
• При физиологическом иммунном ответе на антигены, в частности, связанные с гельминтами, Th2-лимфоциты и тучные клетки продуцируют ИЛ-5.
• ИЛ-5 поддерживает дифференцировку эозинофилов и активирует зрелые эозинофилы. Это единственный известный эффекторный механизм, направленный на уничтожение гельминтов, проникших в ткани внутренней среды организма.
Кишечных гельминтов организм «изгоняет» также с участием IgE, индуцирующего дегрануляцию тучных клеток в слизистой оболочке кишки. Выбрасываемый в ткани гистамин инициирует сокращение гладких мышц стенки кишки, т.е. активную перистальтику и выброс гельминтов. Кроме того, цитокины, ассоциированные с Th2-ответом
(ИЛ-4, ИЛ-9, ИЛ-13, ИЛ-25 и ИЛ-33), вызывают активацию эпителиальных клеток кишечника и усиление выработки слизи.
Апоптоз. Для Th2-лимфоцитов характерна экспрессия ещё одной мембранной молекулы - CD30. Взаимодействие её с лигандом CD30L, экспрессированным на B-лимфоцитах, эозинофилах, эпителии мозговой зоны тимуса и клетках-предшественницах миелопоэза в костном мозге, сначала повышает активность T-лимфоцита, а затем индуцирует его апоптоз.
Иммунное отклонение
В условиях нормы дифференцировка Th1- и Th2-клеток определяется функциональными запросами, так как АГ, индуцирующие иммунный ответ, как правило, обеспечивают его развитие в направлении формирования тех типов эффекторных клеток, которые (сами по себе или через гуморальные продукты) участвуют в реализации протективного эффекта.
Направление терминальной дифференцировки наивных CD4+ T-лимфоцитов в сторону преобладания той или иной субпопуляции (Th1 или Th2) в процессе развития иммунного ответа называют иммунным отклонением.
Нарушение баланса активности Th1- и Th2-субпопуляций играет определяющую роль в развитии многих заболеваний. Несбалансированное преобладание эффектов Th1-клеток означает гиперпродукцию ИФНу и других цитокинов, способствующих развитию иммунного воспаления, которое является основой клеточной аутоиммунной патологии - органоспецифических и некоторых системных аутоиммунных процессов. Преобладание влияния Th2-клеток приводит к гиперпродукции ИЛ-4, ИЛ-5, ИЛ-13 и других цитокинов, способствующих развитию аллергической реакции немедленного типа через влияние на выработку IgE-антител, дифференцировку тучных клеток и эозинофилов. Подходы к коррекции Th1/Th2 дисбалансов основаны на воздействиях, оппозитных преобладающему типу цитокинов: при избыточном количестве Th1-факторов необходимо воздействовать цитокинами, продуцируемыми Th2-клетками, или антителами к Th1цитокинам или их рецепторам. Преобладание Th2-цитокинов требует противоположных воздействий.
Патологические процессы с превалированием иммунного ответа типа Th1 или Th2 перечислены ниже.
• Th1 (макрофагальное воспаление - ГЗТ, гранулемы): тиреоидит Хасимото, офтальмопатия, сахарный диабет 1-го типа, рассеянный склероз, ревматоидный артрит, гастрит (возбудитель Helicobacter pylori), боррелиоз Лайма, хронический гепатит С, острое отторжение аллотрансплантата, острая болезнь «трансплантат против хозяина», саркоидоз, апластическая анемия, привычные аборты.
• Th2 (Тh2-зависимое воспаление - экссудативное, эозинофильное и др.): корь, синдром Оменна, атопические заболевания; хроническая болезнь «трансплантат против хозяина»; аллергический кератоконъюнктивит.