Иммунология: структура и функции иммунной системы: учебное пособие / Р.М. Хаитов. - 2013. - 280 с.
|
|
|
|
Глава 6. Т-ЛИМФОЦИТЫ
Основная задача T-лимфоцитов - распознавание чужеродных или изменённых собственных антигенов в составе комплекса с молекулами MHC. Если на поверхности своих клеток будут представлены чужеродные или изменённые свои молекулы, T-лимфоцит запускает их уничтожение.
• В отличие от B-лимфоцитов, T-лимфоциты не продуцируют растворимых форм антигенраспознающих молекул. Более того, большинство T-лимфоцитов не способны распознавать и связывать растворимые антигены.
• Для того чтобы T-лимфоцит «обратил на антиген своё внимание», другие клетки должны каким-то образом «пропустить» антиген через себя и выставить его на своей мембране в комплексе с MHC-I или MHC-II. Это и есть феномен презентации антигена T-лимфоциту. Распознавание такого комплекса T-лимфоцитом - двойное распознавание, или MHC-рестрикция T-лимфоцитов.
АНТИГЕНРАСПОЗНАЮЩИЙ РЕЦЕПТОР T-ЛИМФОЦИТОВ
Антигенраспознающие рецепторы T-клеток - TCR состоят из цепей, принадлежащих к суперсемейству иммуноглобулинов (см. рис. 5-1). Выступающий над поверхностью клетки антигенраспознающий участок TCR - гетеродимер, т.е. состоит из двух разных полипептидных цепей. Известны два варианта TCR, обозначаемые как αβTCR и γδTCR. Эти варианты различаются составом полипептидных цепей антигенраспознающего участка. Каждый T-лимфоцит экспрессирует только 1 вариант рецептора. αβT-клетки были открыты раньше и изучены подробнее, чем γδT-лимфоциты. В связи с этим строение антигенраспознающего рецептора T-лимфоцитов удобнее описывать на примере αβTCR. Трансмембранно расположенный комплекс TCR состоит из 8 полипептидных
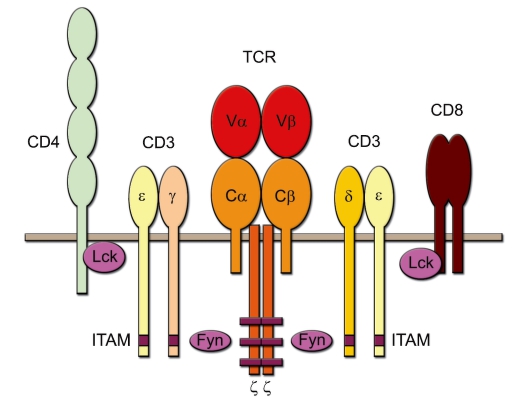
Рис. 6-1. Схема Т-клеточного рецептора и связанных с ним молекул
цепей (гетеродимера α- и β-цепей собственно TCR, двух вспомогательных цепей ζ, а также по одному гетеродимеру ε/δ- и ε/γ-цепей молекулы СD3) (рис. 6-1).
• Трансмембранные цепи α и β TCR. Это 2 примерно одинаковые по размеру полипептидные цепи - α (молекулярная масса 40-60 кДа, кислый гликопротеин) и β (молекулярная масса 40-50 кДа, нейтральный или основный гликопротеин). Каждая из этих цепей содержит по 2 гликозилированных домена во внеклеточной части рецептора, гидрофобную (положительно заряженную за счёт остатков лизина и аргинина) трансмембранную часть и короткий (из 5-12 остатков аминокислот) цитоплазматический участок. Внеклеточные части обеих цепей соединены одной дисульфидной связью.
- V-область. Наружные внеклеточные (дистальные) домены обеих цепей имеют вариабельный аминокислотный состав. Они гомологичны V-области молекул иммуноглобулинов и составляют V-область TCR. Именно V-области α- и β-цепей вступают в связь с комплексом MHC-пептид.
- C-область. Проксимальные домены обеих цепей гомологичны константным областям иммуноглобулинов; это C-области TCR.
- Короткий цитоплазматический участок (как α-, так и β-цепи) не может самостоятельно обеспечить проведение сигнала внутрь клетки. Для этого служат 6 дополнительных полипептидных цепей: γ, δ, 2ε и 2ζ.
• Комплекс CD3. Цепи γ, δ, ε между собой образуют гетеродимеры γε и δε (вместе их называют комплекс CD3). Этот комплекс необходим для экспрессии α- и β-цепей, их стабилизации и проведения сигнала внутрь клетки. Этот комплекс состоит из внеклеточной, трансмембранной (отрицательно заряженной и потому электростатически связанной с трансмембранными участками α- и β-цепей) и цитоплазматической частей. Важно не путать цепи CD3-комплекса с γδ-цепями димера TCR.
• ζ-Цепи соединены между собой дисульфидным мостиком. Большая часть этих цепей расположена в цитоплазме. ζ-Цепи осуществляют проведение сигнала внутрь клетки.
• ITAM-последовательности. Цитоплазматические участки полипептидных цепей γ, δ, ε и ζ содержат 10 последовательностей ITAM (1 последовательность в каждой γ-, ε- и δ-цепях и 3 - в каждой ζ-цепи), взаимодействующих с Fyn - тирозинкиназой цитозоля, активация которой инициирует начало биохимических реакций по проведению сигнала (см. рис. 6-1).
В связывании антигена участвуют ионные, водородные, ван-дерваальсовы и гидрофобные силы; конформация рецептора при этом существенно изменяется. Теоретически каждый TCR способен связывать порядка 105 разных антигенов, причём не только родственных по строению (перекрёстно реагирующих), но и не гомологичных по структуре. Однако в реальности полиспецифичность TCR ограничивается распознаванием всего лишь нескольких структурно схожих антигенных пептидов. Структурной основой этого феномена является особенность одновременного распознавания TCR комплекса «МНС-пептид».
Корецепторные молекулы CD4 и CD8
Помимо самого TCR каждый зрелый T-лимфоцит экспрессирует одну из так называемых корецепторных молекул - CD4 или CD8, которые также взаимодействуют с молекулами MHC на АПК или клеткахмишенях. Каждая из них имеет цитоплазматический участок, связанный
с тирозинкиназой Lck, и, вероятно, вносит свой вклад в проведение сигнала внутрь клетки при распознавании антигена.
• CD4 связывается с инвариантной частью (β2-доменом) молекулы MHC-II (принадлежит к суперсемейству иммуноглобулинов, см. рис. 5-1, б). CD4 имеет молекулярную массу 55 кДа и 4 домена во внеклеточной части. При активации T-лимфоцита одну молекулу TCR «обслуживают» 2 молекулы CD4: вероятно, происходит димеризация молекул CD4.
• CD8 связывается с инвариантной частью (αЗ-доменом) молекулы MHC-I (принадлежит к суперсемейству иммуноглобулинов, см. рис. 5-1, а). CD8 - гетеродимер цепей α и β, соединённых дисульфидной связью. В некоторых случаях обнаруживают гомодимер из двух α-цепей, который также может взаимодействовать с MHC-I. Во внеклеточной части каждая из цепей имеет по одному иммуноглобулиноподобному домену.
Гены T-клеточного рецептора
Гены α-, β-, γ- и δ-цепей (рис. 6-2, также см. рис. 5-4) гомологичны генам иммуноглобулинов и претерпевают при дифференцировке T-лимфоцитов рекомбинацию ДНК, что теоретически обеспечивает генерацию порядка 1016-1018 вариантов антигенсвязывающих рецепторов (реально это разнообразие ограничено числом лимфоцитов в организме до 109).
• Гены α-цепи имеют ~54 V-сегмента, 61 J- и 1 C-сегмент.
• Гены β-цепи содержат ~65 V-сегментов, 2 D-сегмента, 13 J-сегментов и 2 C-сегмента.
• Гены δ-цепи. Между V- и J-сегментами α-цепи расположены гены D-(3), J-(4) и C-(1) сегментов δ-цепи γδTCR. V-сегменты δ-цепи «вкраплены» среди V-сегментов α-цепи.
• Гены γ-цепи γδTCR имеют 2 C-сегмента, 3 J-сегмента перед первым C-сегментом и 2 J-сегмента перед вторым C-сегментом, 15 V-сегментов.
Перестройка генов
• Рекомбинация ДНК происходит при объединении V-, D- и J-сегментов и катализируется тем же комплексом рекомбиназ, что и при дифференцировке B-лимфоцитов.
• После перестройки VJ в генах α-цепи и VDJ в генах β-цепи, а также после присоединения некодируемых N- и P-нуклеотидов с ДНК
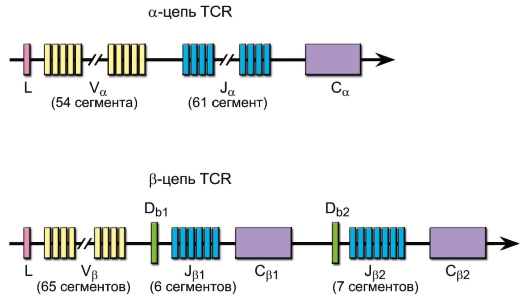
Рис. 6-2. Гены α- и β-цепей антигенраспознающего рецептора T-лимфоцитов человека
транскрибируется РНК. Объединение с C-сегментом и удаление лишних (неиспользуемых) J-сегментов происходит при сплайсинге первичного транскрипта.
• Гены α-цепи могут перестраиваться неоднократно при уже правильно перестроенных и экспрессированных генах β-цепи. Именно поэтому есть некоторая вероятность того, что одна клетка может нести более одного варианта TCR.
• Соматическому гипермутагенезу гены TCR не подвергаются.
ПРОВЕДЕНИЕ СИГНАЛА С АНТИГЕНРАСПОЗНАЮЩИХ РЕЦЕПТОРОВ ЛИМФОЦИТОВ
TCR и BCR имеют ряд общих закономерностей регистрации и проведения в клетку активационных сигналов (см. рис. 5-11).
• Кластеризация рецепторов. Для активации лимфоцита необходима кластеризация антигенраспознающих рецепторов и корецепторов, т.е. «сшивка» нескольких рецепторов одним антигеном.
• Тирозинкиназы. В проведении сигнала играют значительную роль процессы фосфорилирования/дефосфорилирования белков по остатку тирозина под действием тирозинкиназ и тирозинфосфатаз,
ведущие к активации или инактивации этих белков. Эти процессы легко обратимы и «удобны» для быстрых и гибких реакций клетки на внешние сигналы.
• Киназы Src. Богатые тирозином ITAM-последовательности цитоплазматических участков иммунорецепторов подвергаются фосфорилированию под действием нерецепторных (цитоплазматических) тирозинкиназ семейства Src (Fyn, Blk, Lyn в B-лимфоцитах, Lck и Fyn - в T-лимфоцитах).
• Киназы ZAP-70 (в T-лимфоцитах) или Syk (в B-лимфоцитах), связываясь с фосфорилированными ITAM-последовательностями, активируются и начинают фосфорилировать адапторные белки: LAT (Linker for Activation of T cells) (киназой ZAP-70), SLP-76 (киназой ZAP-70) или SLP-65 (киназой Syk).
• Адапторные белки рекрутируют фосфоинозитид-3-киназу (PI3K). Эта киназа в свою очередь активирует серин/треониновую протеинкиназу Akt, вызывая усиление белкового биосинтеза, что способствует ускоренному росту клеток.
• Фосфолипаза Cγ (см. рис. 4-8). Киназы семейства Tec (Btk - в B-лимфоцитах, Itk - в T-лимфоцитах) связывают адапторные белки и активируют фосфолипазу Cγ (PLCγ).
- PLCγ расщепляет фосфатидилинозитдифосфат (PIP2) клеточной мембраны на инозит-1,4,5-трифосфат (IP3) и диацилглицерин
(DAG).
- DAG остаётся в мембране и активирует протеинкиназу С (PKC) - серин/треониновую киназу, которая активирует эволюционно «древний» фактор транскрипции NFκB.
- IP3 связывается со своим рецептором в эндоплазматическом ретикулуме и высвобождает ионы кальция из депо в цитозоль.
- Свободный кальций активирует кальцийсвязывающие белки - кальмодулин, регулирующий активность ряда других белков, и кальциневрин, дефосфорилирующий и тем самым активирующий ядерный фактор активированных T-лимфоцитов NFAT (Nuclear Factor of Activated T cells).
• Ras и другие малые G-белки в неактивном состоянии связаны с ГДФ, но адапторные белки заменяют последний на ГТФ, чем переводят Ras в активное состояние.
- Ras обладает собственной ГТФазной активностью и быстро отщепляет третий фосфат, чем возвращает себя в неактивное состояние (самоинактивируется).
- В состоянии кратковременной активации Ras успевает активировать очередной каскад киназ, называемых MAPK (MitogenActivated Protein Kinase), которые в итоге активируют фактор транскрипции AP-1 в ядре клетки. На рис. 6-3 схематично представлены основные пути передачи сигналов с TCR. Активационный сигнал включается при связывании TCR с лигандом (комплексом молекула МНС-пептид) при участии корецептора (CD4 или CD8) и костимулирующей молекулы CD28. Это приводит к активации киназ Fyn и Lck. Красным цветом отмечены участки ITAM в цитоплазматических частях полипептидных цепей CD3. Отражена роль Src-киназ, связанных с рецептором, в фосфорилировании белков: как рецепторных, так и сигнальных. Обращает на себя внимание чрезвычайно широкий спектр эффектов киназы Lck, связанной с корецепторами; роль киназы Fyn установлена с меньшей определённостью (отражено в прерывистом характере линий).
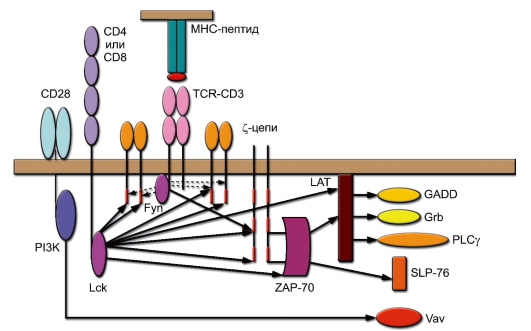
Рис. 6-3. Источники и направление пусковых активационных сигналов при стимуляции Т-лимфоцитов. Обозначения: ZAP-70 (ζ-associated proteinkinase, мол. масса 70 кДа) - протеинкиназа р70, связанная с ζ-цепью; PLCγ (Phospholipase Сγ) - фосфолипаза С, изоформа γ; PI3K (Phosphatidyl Inositol 3-kinase) - фосфатидилинозитол 3-киназа; Lck, Fyn -тирозинкиназы; LAT, Grb, SLP, GADD, Vav - адапторные белки
Ключевую роль в посредничестве между рецепторными киназами и адапторными молекулами и ферментами играет тирозинкиназа ZAP-70. Она активирует (через фосфорилирование) адапторные молекулы SLP-76 и LAT, а последняя передаёт активационный сигнал другим адапторным белкам GADD, GRB и активирует у-изоформу фосфолипазы С (PLCy). До этого этапа в передачу сигнала вовлекаются исключительно факторы, связанные с клеточной мембраной. Важный вклад во включение сигнальных путей вносит костимулирующая молекула CD28, реализующая своё действие через связанную с ней липидную киназу PI3K (Phosphatidyl Inositol 3-kinase). Основной мишенью киназы PI3K служит фактор Vav, связанный с цитоскелетом.
В результате формирования сигнала и передачи его от рецептора Т-клетки к ядру образуются 3 транскрипционных фактора - NFAT, AP-1 и NF-kB, индуцирующие экспрессию генов, контролирующих процесс активации Т-лимфоцитов (рис. 6-4). К образованию NFAT приводит сигнальный путь, не зависящий от костимуляции, который включается благодаря активации фосфолипазы С и реализуется с участием ионов
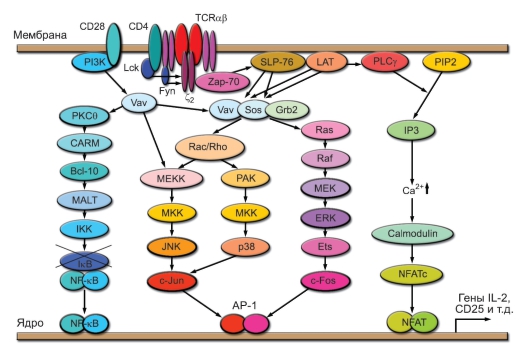
Рис. 6-4. Схема сигнальных путей при активации Т-клеток. NFAT (Nuclear factor of activated T cells), AP-1 (Activation protein-1), NF-κB (Nuclear factor of к-gene of B cells) - факторы транскрипции
Са2+. Этот путь вызывает активацию кальциневрина, который, обладая активностью фосфатазы, дефосфорилирует цитозольный фактор NFAT-Р. Благодаря этому NFAT-Р приобретает способность мигрировать в ядро и связываться с промоторами активационных генов. Фактор АР-1 формируется как гетеродимер из белков с-Fos и с-Jun, образование которых индуцируется благодаря активации соответствующих генов под влиянием факторов, образующихся в результате реализации трёх компонентов МАР-каскада. Эти пути включаются при участии коротких ГТФ-связывающих белков Ras и Rac. Значительный вклад в реализацию МАР-каскада вносят сигналы, зависящие от костимуляции через молекулу CD28. Третий транскрипционный фактор, NF-kB, известен как основной транскрипционный фактор клеток врождённого иммунитета. Он активируется в результате расщепления блокирующей субъединицы IkB киназой IKK, которая в Т-клетках активируется в ходе передачи сигнала, зависимого от изоформы ϴ протеинкиназы С (PKC9). Основной вклад во включение этого сигнального пути вносят костимулирующие сигналы от CD28. Сформировавшиеся транскрипционные факторы, связавшись с промоторными участками генов, индуцируют их экспрессию. Для начальных этапов реакции Т-клеток на стимуляцию особенно важна экспрессия генов IL2 и IL2R, что обусловливает выработку ростового фактора Т-клеток ИЛ-2 и экспрессию его высокоаффинного рецептора на Т-лимфоцитах. В результате ИЛ-2 выступает как аутокринный ростовой фактор, обусловливающий пролиферативную экспансию Т-клеток клонов, вовлечённых в реакцию на антиген.
ДИФФЕРЕНЦИРОВКА T-ЛИМФОЦИТОВ
В основе выделения этапов развития Т-лимфоцитов лежит состояние рецепторных V-генов и экспрессии TCR, а также корецепторов и других мембранных молекул. Схема дифференцировки Т-лимфоцитов (рис. 6-5) аналогична приведённой выше схеме развития В-лимфоцитов (см. рис. 5-13). Приведены ключевые характеристики фенотипа и ростовых факторов развивающихся Т-клеток. Принятые обозначения стадий развития Т-клеток определяются экспрессией корецепторов: DN (от Double-Negative, CD4CD8) - двойные отрицательные, DP (от Double-Positive, CD4+CD8+) - двойные положительные, SP (от Single-Positive, CD4+CD8- и CD4CD8+) - одинарно положительные. Деление DNтимоцитов на стадии DN1, DN2, DN3 и DN4 основывается на характере
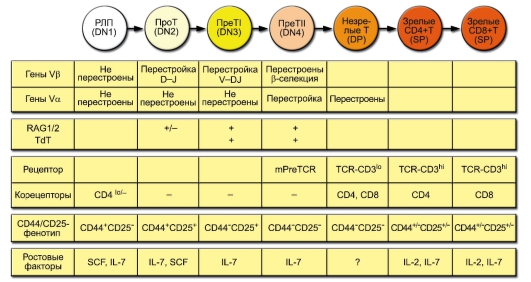
Рис. 6-5. Развитие Т-лимфоцитов
экспрессии молекул CD44 и CD25. Другие условные обозначения: SCF (от Stem Cell Factor) - фактор стволовых клеток, lo (low; метка индекса) - низкий уровень экспрессии. Стадии реаранжировки: D-J - предварительный этап, соединение сегментов D и J (только в генах β- и δ-цепей TCR, см. рис. 6-2), V-DJ - завершающий этап, соединение зародышевого V-гена с объединённым сегментом DJ.
• Тимоциты дифференцируются из общей клетки-предшественника, которая ещё вне тимуса экспрессирует такие мембранные маркёры, как CD7, CD2, CD34 и цитоплазматическую форму CD3.
• Коммитированные к дифференцировке в T-лимфоциты клеткипредшественники мигрируют из костного мозга в субкапсулярную зону коры тимуса, где примерно в течение одной недели медленно пролиферируют. На тимоцитах появляются новые мембранные молекулы CD44 и CD25.
• Затем клетки перемещаются вглубь коры тимуса, молекулы CD44 и CD25 исчезают с их мембраны. В этой стадии начинается перестройка генов β-, γ- и δ-цепей TCR. Если гены γ- и δ-цепей успевают продуктивно, т.е. без сдвига рамки считывания, перестроиться раньше, чем гены β-цепи, то лимфоцит дифференцируется далее как γδT. В противном случае происходит экспрессия β-цепи на мембране в комплексе с pTα (инвариантной суррогатной цепью, заменяющей на этом этапе настоящую α-цепь) и CD3. Это служит
сигналом к прекращению перестройки генов γ- и δ-цепей. Клетки начинают пролиферировать и экспрессировать одновременно CD4 и CD8 - дважды позитивные тимоциты. При этом накапливается масса клеток с уже готовой β-цепью, но с ещё не перестроенными генами α-цепи, что вносит свой вклад в разнообразие αβ-гетеродимеров.
• На следующем этапе клетки перестают делиться и начинают перестраивать Vα-гены, причём несколько раз в течение 3-4 сут. Перестройка генов α-цепи приводит к необратимой делеции δ-локуса, расположенного между сегментами генов α-цепи.
• Происходят экспрессия TCR с каждым новым вариантом α-цепи и отбор (селекция) тимоцитов по силе связывания с комплексом MHC-пептид на мембранах эпителиальных клеток тимуса.
- Позитивная селекция: погибают тимоциты, не связавшие ни одного из доступных комплексов MHC-пептид. В результате позитивной селекции в тимусе погибает около 90% тимоцитов.
- Негативная селекция уничтожает клоны тимоцитов, связывающих комплексы MHC-пептид со слишком высокой аффинностью. Негативная селекция элиминирует от 10 до 70% клеток, прошедших позитивную селекцию.
- Тимоциты, связавшие какой-либо из комплексов MHC-пептид с правильной, т.е. средней по силе, аффинностью, получают сигнал к выживанию и продолжают дифференцировку.
• На короткое время с мембраны тимоцитов исчезают обе корецепторные молекулы, а затем экспрессируется одна из них: тимоциты, распознавшие пептид в комплексе с MHC-I, экспрессируют корецептор CD8, а с MHC-II - корецептор CD4. Соответственно на периферию выходят (в соотношении около 2:1) T-лимфоциты двух типов: CD8+ и CD4+, функции которых в предстоящих иммунных ответах различны.
- CD8+ T-клетки играют роль цитотоксических T-лимфоцитов (ЦТЛ) - они распознают и непосредственно убивают клетки, модифицированные вирусом, опухолевые и другие «изменённые» клетки (рис. 6-6).
- CD4+ T-клетки. Функциональная специализация CD4+ T-лимфоцитов более разнообразна. Значительная часть CD4+ T-лимфоцитов в процессе развития иммунного ответа становится T-хелперами (помощниками), взаимодействующими с В-лимфоцитами, Т-лимфоцитами и другими клетками при
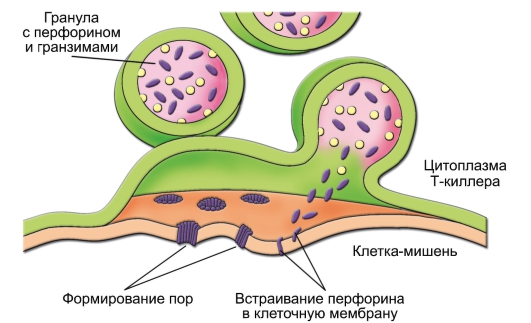
Рис. 6-6. Механизм воздействия цитотоксического T-лимфоцита на клеткумишень. В Т-киллере в ответ на увеличение концентрации Са2+ гранулы с перфорином (фиолетовые овалы) и гранзимами (жёлтые кружочки) сливаются с клеточной мембраной. Освободившийся перфорин встраивается в мембрану клетки-мишени с последующим образованием пор, проницаемых для гранзимов, воды и ионов. В результате клетка-мишень лизируется
прямом контакте или через растворимые факторы (цитокины). В определённых случаях из них могут развиться CD4+ ЦТЛ: в частности, такие T-лимфоциты обнаружены в значительных количествах в коже больных с синдромом Лайелла.
Субпопуляции T-хелперов
С конца 80-х годов XX века было принято выделять 2 субпопуляции T-хелперов (в зависимости от того, какой набор цитокинов они продуцируют) - Th1 и Th2. В последние годы спектр субпопуляций CD4+ Т-клеток продолжает расширяться. Обнаружены такие субпопуляции, как: Th17, T-регуляторы, Tr1, Th3, Tfh и др.
Основные субпопуляции CD4+ Т-клеток:
• Th0 - CD4+ Т-лимфоциты на ранних стадиях развития иммунного ответа, они продуцируют только ИЛ-2 (митоген для всех лимфоцитов).
• Th1 - дифференцированная субпопуляция CD4+ Т-лимфоцитов, специализирующаяся на продукции ИФНγ, ФНОβ и ИЛ-2. Эта субпопуляция осуществляет регуляцию многих реакций клеточного иммунитета, включая гиперчувствительность замедленного типа (ГЗТ) и активацию ЦТЛ. Кроме того, Th1 стимулируют продукцию В-лимфоцитами опсонизирующих антител класса IgG, запускающих каскад активации комплемента. Развитие избыточного воспаления с последующим повреждением тканей напрямую связано с активностью Th1-субпопуляции.
• Th2 - дифференцированная субпопуляция CD4+ Т-лимфоцитов, специализирующаяся на выработке ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13. Эта субпопуляция участвует в активации В-лимфоцитов и способствует секреции ими больших количеств антител разных классов, особенно IgE. Кроме того, Th2-субпопуляция участвует в активации эозинофилов и развитии аллергических реакций.
• Th17 - субпопуляция CD4+ Т-лимфоцитов, специализирующаяся на образовании ИЛ-17. Эти клетки осуществляют противогрибковую и антимикробную защиту эпителиальных и слизистых барьеров, а также играют ключевую роль в патологии аутоиммунных заболеваний.
• Т-регуляторы - CD4+ Т-лимфоциты, подавляющие активность других клеток иммунной системы посредством секреции иммуносупрессорных цитокинов - ИЛ-10 (ингибитора активности макрофагов и Th1-клеток) и ТФРβ - ингибитора пролиферации лимфоцитов. Ингибиторный эффект может также достигаться при непосредственном межклеточном взаимодействии, поскольку на мембране некоторых Т-регуляторов экспрессированы индукторы апоптоза активированных и «отработавших» лимфоцитов - FasL (Fas-лиганд). Существует несколько популяций CD4+ регуляторных Т-лимфоцитов: естественные (Treg), созревающие в тимусе (CD4+CD25+, экспрессируют фактор транскрипции Foxp3), и индуцированные - локализованные преимущественно в слизистых оболочках пищеварительного тракта и переключившиеся на образование ТФРβ (Th3) или ИЛ-10 (Tr1). Нормальное функционирование Т-регуляторов необходимо для поддержания гомеостаза иммунной системы и предотвращения развития аутоиммунных заболеваний.
• Дополнительные хелперные популяции. В последнее время появляется описание всё новых популяций CD4+ Т-лимфоцитов, клас-
сифицированных по типу преимущественно продуцируемого ими цитокина. Так, как оказалось, одной из важнейших популяций являются Tfh (от англ. follicular helper - фолликулярный хелпер). Эта популяция CD4+ Т-лимфоцитов преимущественно расположена в лимфоидных фолликулах и осуществляет хелперную функцию для В-лимфоцитов посредством продукции ИЛ-21, вызывая их созревание и терминальную дифференцировку в плазматические клетки. Кроме ИЛ-21 Tfh могут также продуцировать ИЛ-6 и ИЛ-10, необходимые для дифференцировки В-лимфоцитов. Нарушение функций этой популяции приводит к развитию аутоиммунных заболеваний или иммунодефицитов. Другой «новоявленной» популяцией являются Th9 - продуценты ИЛ-9. По-видимому, это Th2, переключившиеся на секрецию ИЛ-9, способного вызывать пролиферацию Т-хелперных клеток при отсутствии антигенной стимуляции, а также усиливать секрецию В-лимфоцитами IgM, IgG и IgE.
Основные субпопуляции Т-хелперов представлены на рис. 6-7. На рисунке суммированы современные представления об адаптивных субпопуляциях CD4+ Т-клеток, т.е. субпопуляций, формирующих-
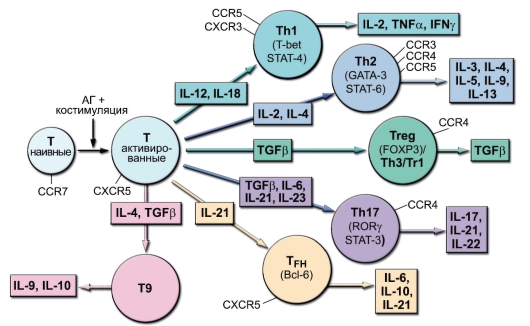
Рис. 6-7. Адаптивные субпопуляции CD4+ Т-клеток (цитокины, дифференцировочные факторы, хемокиновые рецепторы)
ся при иммунном ответе, а не в ходе естественного развития клеток. Для всех разновидностей Т-хелперов указаны цитокины-индукторы (на стрелках, ведущих к кружкам, символизирующим клетки), транскрипционные факторы (внутри кружков), хемокиновые рецепторы, направляющие миграцию (около линий, отходящих от «поверхности клетки»), и продуцируемые цитокины (в прямоугольниках, на которые направлены стрелки, отходящие от кружков).
Расширение семейства адаптивных субпопуляций CD4+ Т-клеток потребовало решения вопроса о природе клеток, с которыми взаимодействуют эти субпопуляции (кому они оказывают «помощь» в соответствии со своей функцией хелперов). Эти представления отражены на рис. 6-8. Здесь же представлен уточнённый взгляд на функции этих субпопуляций (участие в защите от определённых групп патогенов), а также о патологических последствиях несбалансированного усиления активности этих клеток.
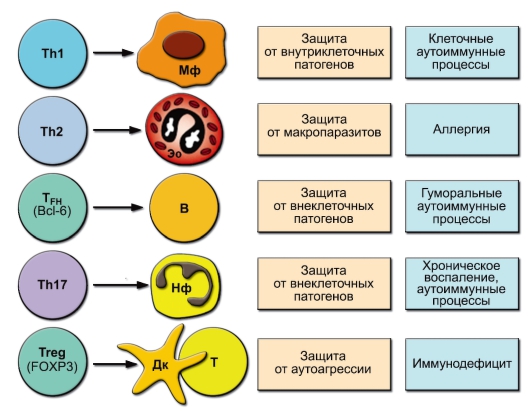
Рис. 6-8. Адаптивные субпопуляции Т-клеток (клетки-партнёры, физиологические и патологические эффекты)
γδT-лимфоциты
Подавляющее большинство (99%) T-лимфоцитов, проходящих лимфопоэз в тимусе, составляют αβT-клетки; менее 1% - γδT-клетки. Последние в большинстве дифференцируются вне тимуса, в первую очередь в слизистых оболочках пищеварительного тракта. В коже, лёгких, пищеварительном и репродуктивном трактах они являются доминирующей субпопуляцией внутриэпителиальных лимфоцитов. Среди всех T-лимфоцитов организма γδT-клетки составляют от 10 до 50%. В эмбриогенезе γδT-клетки появляются раньше αβT-клеток.
• γδT-клетки не экспрессируют CD4. Молекула CD8 экспрессирована на части γδT-клеток, но не в виде ap-гетеродимера, как на CD8+ apT-клетках, а в виде гомодимера из двух a-цепей.
• Антигенраспознающие свойства: γδTCR в большей степени напоминают иммуноглобулины, чем αβTCR, т.е. способны связывать нативные антигены независимо от классических молекул MHC - для γδT-клеток не обязателен или вовсе не нужен предварительный процессинг антигена АПК.
• Разнообразие γδTCR меньше, чем αβTCR или иммуноглобулинов, хотя в целом γδT-клетки способны распознавать широкий спектр антигенов (в основном это фосфолипидные антигены микобактерий, углеводы, белки теплового шока).
• Функции γδT-клеток ещё до конца не изучены, хотя становится преобладающим мнение, что они служат одним из связующих компонентов между врождённым и приобретённым иммунитетом. γδT-клетки - один из первых барьеров на пути патогенов. Кроме того, эти клетки, секретируя цитокины, играют важную иммунорегуляторную роль и способны дифференцироваться в ЦТЛ.
NKT-лимфоциты
Естественные киллерные Т-клетки (NKT-клетки) представляют особую субпопуляцию лимфоцитов, занимающую промежуточное положение между клетками врождённого и адаптивного иммунитета. Эти клетки имеют черты как NK-, так и Т-лимфоцитов. NKT-клетки экспрессируют αβTCR и характерный для NK-клеток рецептор NK1.1, принадлежащий к суперсемье лектиновых гликопротеинов С-типа. Однако TCR-рецептор NKT-клеток имеет существенные отличия от TCR-рецептора обычных клеток. У мышей большинство NKTклеток экспрессирует инвариантный V-домен a-цепи, состоящий из
сегментов Vα14-Jα18, иногда обозначаемый как Jα281. У человека V-домен α-цепи состоит из сегментов Vα24-JαQ. У мышей α-цепь инвариантного TCR преимущественно комплексируется с Vβ8.2, у человека - с Vβ11. Из-за особенностей строения цепей TCR NKTклеток называют инвариантным - iTCR. Развитие NKT-клеток зависит от молекулы CD1d, которая имеет сходство с молекулами МНС-I. В отличие от классических молекул МНС-I, презентирующих Т-клеткам пептиды, CD1d презентирует Т-клеткам только гликолипиды. Хотя считается, что печень является местом развития NKT-клеток, имеются строгие доказательства роли тимуса в их развитии. NKT-клетки играют важную роль в регуляции иммунитета. У мышей и людей с различными аутоиммунными процессами функциональная активность NKT-клеток сильно нарушена. Полной картины значимости таких нарушений в патогенезе аутоиммунных процессов нет. При некоторых аутоиммунных процессах NKT-клетки могут играть супрессорную роль.
Помимо контроля аутоиммунных и аллергических реакций, NKTклетки участвуют в иммунном надзоре, вызывая при повышении функциональной активности отторжение опухолей. Велика их роль в противомикробной защите, особенно на ранних этапах развития инфекционного процесса. NKT-клетки вовлекаются в различные воспалительные инфекционные процессы, особенно при вирусных поражениях печени. В целом NKT-клетки - многофункциональная популяция лимфоцитов, несущая ещё много научных загадок.
На рис. 6-9 обобщены данные о дифференцировке Т-лимфоцитов на функциональные субпопуляции. Представлены несколько уровней бифуркации: γδТ/αβТ, далее для αβТ-клеток - NKT/ остальные Т-лимфоциты, для последних - CD4+/CD8+, для CD4+ Т-клеток - Th/Treg, для CD8+ Т-лимфоцитов - CD8αβ/CD8αα. Показаны также дифференцировочные транскрипционные факторы, ответственные за все линии развития.
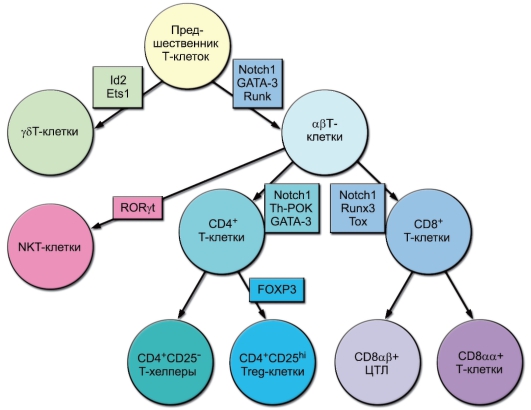
Рис. 6-9. Естественные субпопуляции Т-лимфоцитов и их дифференцировочные факторы