Биология: учебник: в 2 т. / под ред. В. Н. Ярыгина. - 2011. - Т. 1. - 736 с. : ил.
|
|
|
|
Раздел I. ЖИЗНЬ КАК ЯВЛЕНИЕ МАТЕРИАЛЬНОГО МИРА. Глава 1. ВВЕДЕНИЕ В БИОЛОГИЮ
1.1. биология - область естествознания, комплекс научных дисциплин о жизни во всех ее проявлениях
Термин «биология» (греч. bios - жизнь, logos - слово, учение, наука) предложен в начале XIX в. Ж.-Б. Ламарком и Г. Тревиранусом для обозначения науки о жизни как особом природном явлении. За минувшие два столетия биология проделала плодотворный путь развития. В настоящее время она представляет комплекс дисциплин. Предметом изучения одних остается жизнь как явление окружающего мира, других - проявления жизни на том или ином уровне организации или в том или ином ее сегменте, то есть все живое на планете в его конкретном пространственно-временном воплощении.
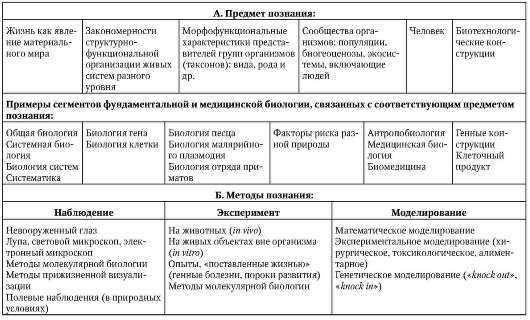
Каждая биологическая дисциплина характеризуется предметом исследования (познания), преимущественно используемыми методами научного анализа, идеями общего порядка, оформленными в виде теорий или гипотез, и методологическими подходами, отражающими отношение исследователя к предмету познания (табл. 1.1).
Таблица 1.1. Процесс научного познания: предмет, методы, идеи общего порядка и методологические принципы
В англоязычной учебной литературе называют еще 2 методологических подхода, характерные для современной биологии, - индуктивный и дедуктивный. Индуктивный подход - это обобщения, вытекающие из результатов изучения «частностей». В европейской науке он стал доминирующим с XVII в., что связано с именами Ф. Бэкона и И. Ньютона, заложившими в основание сформулированных ими законов результаты конкретных опытов (см. закон всемирного тяготения - «яблоко, упавшее с яблони на голову ученого»). Дедуктивный подход исходит из возможности предсказать «частности», имея представления об общих характеристиках объекта познания.
К классическим биологическим дисциплинам относятся общая и системная биология, зоология, ботаника, микология, протистология, микробиология, вирусология, морфология (анатомия, гистология, цитология - в зависимости от структурного уровня), физиология, биохимия и биофизика, этология, биология развития (эмбриология, геронтология), палеонтология, антропология, генетика, экология.
Осознание того, что живое представлено формами, объединенными в группы (таксоны), представители которых различаются по степени исторического родства или же не состоят в таком родстве вовсе, дало систематику. Последняя относит организм к определенному виду, роду, семейству, отряду, классу, типу, порядку. С появлением новых данных положение группы живых существ в системе органического мира пересматривается. Так, использование методов макромолекулярной систематики («молекулярных часов») показало, что генетическое расстояние между орангутан(г)ом и африканскими человекообразными обезьянами (шимпанзе, горилла), относимыми приматологией к одному семейству Pongidae, превосходит названное расстояние между последними и человеком. Поставлен вопрос о выделении орангутан(г)а в отдельное семейство.
Закономерности исторического развития жизни в виде ее отдельных форм или их природных совокупностей изучаются в рамках эволюционного направления (эволюционной теории или учения).
В масштабе реального времени жизнь организована в виде сменяющихся поколений организмов. Механизмы, обеспечивающие указанное явление, изучает репродуктивная биология.
Вторая половина ХХ в. отмечена успехами в познании фундаментальных механизмов жизнедеятельности. Описан в деталях поток биологической информации в живых системах, в основных чертах поняты
молекулярные механизмы энергетического обеспечения процессов жизнедеятельности. Исследования по названным направлениям - задача таких оформившихся во второй половине ХХ столетия биологических дисциплин, как молекулярная биология и молекулярная генетика, биоинформатика, биоэнергетика. Молодой дисциплиной является клеточная биология, возникшая на рубеже третьей и последней четвертей минувшего века как следствие развития цитоморфологии, цитохимии и цитофизиологии первой половины-середины ХХ в.
Объединение молекулярно-генетического, клеточно-биологического, популяционно-клеточного и системного подходов породило современную иммунологию, предметом изучения которой являются механизмы иммунологического надзора с функцией защиты целостности и биологической индивидуальности организма, включая реакцию на выход собственных клеток из-под общеорганизменных регуляторных влияний (онкотрансформация), проникновение в него инфекционных агентов (бактерий, вирусов) и чужеродных белков (факты совместимости по группам крови АВ0, резус и др.)
Разработки в области молекулярной биологии, генетики и клеточной биологии, ориентированные на решение практических проектов в интересах промышленности, медицины и сельского хозяйства, оформились в научно-практическое биотехнологическое (греч. bios - жизнь, techne - ремесло, искусство, мастерство) направление - генную, клеточную, тканевую инженерию. Биотехнологическое направление, по крайней мере, в части генной инженерии базируется на принципах природного явления - горизонтальном (латеральном) переносе генов между представителями разных систематических групп. Это явление распространено в природе, особенно в мире прокариот. В здравоохранении используется ряд лекарственных средств генно-инженерной природы, например инсулин.
Перспективы развития биотехнологического направления в обозримом будущем связывают с нанотехнологиями, в том числе медицинского назначения. Их основу составляют конструкции, не превосходящие по размерам десятки-сотни нанометров (1 нм = 10-9 м) и, следовательно, способные «работать» в качестве диагностических, терапевтических или «надзирающих» (нанороботы) агентов с отдельными клетками и внутриклеточно. Наноподход используется также при создании новых лекарственных средств.
На рубеже ХХ-ХХ1 вв. в биологии произошли события, кульминационным моментом которых стал проект «Геном человека». В результате
его осуществления установлены последовательности нуклеотидов
всех 25 (включая митохондриальную) молекул ДНК клеток человека. Таким образом, практически в полном объеме прочитаны ДНК-тексты и, следовательно, открыт доступ к содержанию генетической информации, управляющей биологической составляющей индивидуального развития и жизнедеятельности людей. Ведутся работы по определению последовательности нуклеотидов (секвенирова-нию; англ. sequence - последовательность) в геномах других существ, включая ближайших эволюционных «родственников» людей (шимпанзе) и возбудителей паразитарных и инфекционных болезней. В итоге в новейшей биологии появилась дисциплина геномика (нем. genom - совокупность генов или, более точно, нуклеотидных последовательностей ДНК гаплоидного набора хромосом).
К носителям генетической информации в клетке, кроме нуклеиновых кислот, относятся белки или протеины (греч. protos - первый; простые белки являются первыми функционально значимыми продуктами активности многих генов; первооснову любой биологической функции составляют белки). Закономерности реализации генетической информации на уровне белков - предмет изучения «сверхновой» биологической дисциплины протеомики (протеом - совокупность белков, образуемых клетками организмов определенного вида).
Количество структурных (смысловых) генов, кодирующих аминокислотные последовательности белков в геноме человека, меньше числа конкретных белков, обнаруживаемых в клетках (см. здесь же, ниже). Это пробудило интерес к превращениям или процессингу (англ. processing - обработка, переработка; лат. procedo - прохожу, продвигаюсь) пре-РНК транскриптов, образующихся в результате считывания информации с ДНК. Результат - «сверхновая» биологическая дисциплина транскриптомика (транскриптом - набор информационных РНК, образуемых клетками организмов конкретного вида на основе соответствующего генома).
Исследования в области транскриптомики и протеомики не могут осуществляться в отрыве от исследований в области геномики. Геном человека содержит 30-35 тыс. (по некоторым последним сообщениям - 20 тыс.) участков ДНК, кодирующих структуру полипептидов и некоторых видов РНК, то есть генов в понимании классической генетики. Количество белков в клетках людей уже сейчас уверенно оценивается цифрой 200-300 тыс. Ожидаемое же количество составляет по предварительным оценкам не менее 1 млн. В связи с этим протеомику следует
рассматривать как элемент функциональной геномики. В таком случае транскриптомика служит «связующим звеном» между собственно геномикой (структурная геномика), поставляющей сведения о ну-клеотидных последовательностях ДНК, и протеомикой, дающей представление о «полном протеомном портрете» или ассортименте белков, образуемых клеткой (организмом). В компетенцию функциональной геномики входит также получение ответов на вопросы: когда, где, при каких условиях и с какой интенсивностью в организме экспрессируются разные гены (образуются разные белки).
Необходимость представлять феномен реализации генетической информации в процессах жизнедеятельности не столько в биохимических терминах (ДНК, РНК, белки, метаболиты), но раскрывая вклад этой информации в структуру и функцию реальных биологических объектов (ресничка, жгутик, механохимическая сократительная система мышцы) привела к зарождению в современной науке о жизни направления биология систем (systems biology), в рамках которого редукционистский методологический принцип (см. здесь же, выше), доминировавший в биологии XX в., сменяется принципами интегративным и системным.
Исследование внутриклеточного обмена веществ (метаболизма) как существенной составляющей потоков информации, энергии и веществ проводится в рамках «сверхновой» биологической дисциплины мета-боломики (греч. metabole - перемена, превращение; метаболизм или обмен веществ - совокупность процессов биохимических превращений веществ и энергии в клетке, организме, экосистеме) или биохимического профилирования. Метаболомика изучает химические взаимодействия, в том числе межбелковые в процессе обмена веществ или, что одно и то же, в процессе жизнедеятельности. При этом метаболом определяется как совокупность всех метаболитов, присутствующих в клетке или ткани в известных условиях.
Поток биологической информации в его структурно-временном оформлении невозможен вне клеточной организации, что дает основание ожидать нарождения в науке о жизни в XXI в. еще одной дисциплины - целлюломики (лат. cellula - клетка) или цитомики (греч. cytos - клетка). В отличие от клеточной биологии, фиксирующей внимание на раскрытии существенных черт клеточной структуры и функций, а также закономерностей организации и динамики клеточных тканевых систем (клеточных популяций), задача цитомики (целлюло-мики) видится в расшифровке механизмов генетического обеспечения и контроля клеточной дифференцировки и гистогенезов, а также геноти-
пических и фенотипических основ разнообразия клеток одного морфо-функционального типа в свете данных геномики, транскриптомики и протеомики.
Выше названы фундаментальные биологические дисциплины. Между тем существуют области исследования биологических объектов, порождаемые практическими соображениями, и таким образом являющиеся в терминах науковедения прикладными. Так, изучается структура паразитоценозов в интересах медицины или животноводства. Прикладной характер имеют биология человека (антропобио-логия), медицинская биология, биомедицина (см. Предисловие). Прикладные исследования опираются на достижения фундаментальной биологии. Вместе с тем есть много указаний на относительность деления научных разработок на фундаментальные и прикладные.
1.2. история представлений о мире жизни.
научный базис биологии
Интерес к познанию мира жизни сопровождает человечество на протяжении всей его истории. На заре этой истории интерес к живому окружению отражал практические нужды людей. Желание узнать, следует ли избегать встреч с теми или иными животными и растениями или, наоборот, использовать их в своих целях, объясняет, почему первоначально внимание к живым формам выливалось в попытки их подразделения (классификации) на полезные и опасные, болезнетворные, представляющие пищевую ценность, пригодные для изготовления одежды, орудий труда, жилищ, предметов обихода, удовлетворения эстетических запросов.
Характерная черта человека - его способность сохранять и передавать потомкам опыт наблюдений за природными явлениями, благодаря чему этот опыт со временем преумножается, приводя периодически к качественно новым решениям ресурсных и иных проблем. Дошедшие до нас памятники еще «донаучного» периода истории человечества свидетельствуют об активном отношении людей к происходящему, их тонкой наблюдательности, стремлении к систематизации опыта в целях извлечения наибольшей пользы. На рис. 1.1 представлена печатка с таблицей, которая воспринимается как инструкция по разведению лошадей по результатам оценки фенотипов в ряду поколений. Символы расположены горизонтальными рядами, а головы лошадей по форме относятся к трем разным типам (генеалогический подход или метод родословных современной генетики).
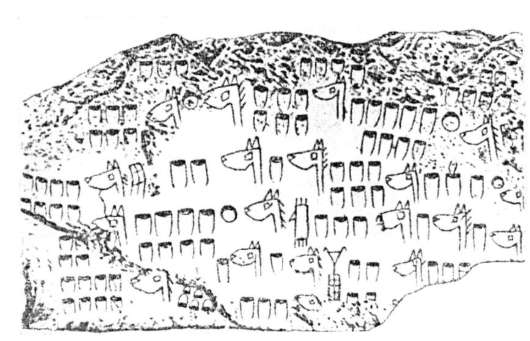
Рис. 1.1. Изображение на печатке: запись о разведении лошадей в Двуречье, 6000 лет назад
На определенной стадии знакомства с живой природой в умах людей, наряду с представлениями о разнообразии организмов, возникает идея единства всего живого, включая людей. Одновременно проясняются роль и истоки разнообразия в живой природе. Возникает понимание
непротиворечивости биологического единообразия и многообразия.
Решающим научным доказательством единства всего живого стала клеточная теория (см. п. 2.1) Т. Шванна и М. Шлейдена (1839). Открытие клеточного принципа строения растительных и животных организмов положило начало плодотворному изучению общих закономерностей, составляющих основу морфологии, физиологии, репродукции и индивидуального развития живых существ.
Открытием фундаментальных законов наследственности биология обязана Г. Менделю, описавшему правила наследования признаков на основе передачи в поколениях дискретных наследственных задатков (1865), Г. де Фризу, К. Корренсу и К. Чермаку, переоткрывшим в 1900 г. и сделавшим достоянием науки правила наследования Г. Менделя, Г. де Фризу, открывшему мутационную изменчивость (1901), основателям популяционной генетики Г. Харди и В. Вайнбергу, сформулировав-
шим закон генетического равновесия в популяциях организмов (1908), Т.Г. Моргану и его научному коллективу, создавшим хромосомную теорию наследственности (1910-1916), Дж. Уотсону, Ф. Крику, М. Вилкин-су и Р. Франклин, открывшим двойную спираль ДНК (1953). Названные законы раскрывают механизм передачи наследственной информации от клетки к клетке, а через клетки - от особи к особи и перераспределения ее в пределах вида в череде поколений, принципы структурно-функциональной организации генетического аппарата. Благодаря этим открытиям становится понятной роль таких биологических явлений, как половое размножение, смена поколений, онтогенез (индивидуальное развитие) и филогенез (историческое развитие).
Заключение о единстве всего живого подтверждают исследования биохимических (обменных, метаболических) и биофизических механизмов жизнедеятельности клеток. Начало этих исследований датируется второй половиной XIX в., однако наиболее весомы достижения молекулярной биологии (вторая половина ХХ в.). Благодаря молекулярно-биологическим исследованиям, уделяющим главное внимание закономерностям хранения, передачи и использования клетками биологической информации, были раскрыты физико-химические основы таких универсальных свойств живого, как наследственность и изменчивость, специфичность биологических макромолекул, структур и функций, закономерное воспроизведение в ряду поколений клеток и организмов определенного типа структурно-функциональной организации.
В контексте идеи единства мира жизни важно то, что живые формы
принципиально одинаковым образом хранят наследственную информацию, передают ее в ряду поколений или используют в своей жизнедеятельности, обеспечивают жизненные процессы энергией и переводят энергию в работу.
Клеточная теория, достижения генетики, биохимии, биофизики и молекулярной биологии обосновывают тезис о единстве органического мира в его современном состоянии. То, что живое на планете едино в историческом плане, обосновывается теорией эволюции (эволюционное учение). Естественно-научные основы теории заложены Ч. Дарвином (1858). Дальнейшее развитие, связанное с достижениями генетики и популяционной биологии, сравнительной эмбриологии и морфологии, палеонтологии, она получила в трудах А.Н. Северцова, Н.И. Вавилова, Р. Фишера, С.С. Четверикова, Ф.Р. Добжанского, Н.В. Тимофеева-Ресовского, С. Райта, И.И. Шмальгаузена, чья научная деятельность
относится к первой половине - середине ХХ в. Эволюционисты рубежа XX-XXI вв. развивают идеи о новых, в том числе «недарвиновских» факторах, механизмах и формах эволюционного процесса.
Эволюционная идея называет пути, способы и механизмы, которые за несколько миллиардов лет привели к наблюдаемому ныне разнообразию живых форм, в одинаковой мере приспособленных к среде обитания и различающихся по уровню структурно-функциональной организации. Другой важный итог эволюционной парадигмы состоит в признании, что живые формы связаны друг с другом общностью происхождения (генетическое родство). Степень родства различается для представителей разных групп, а свое выражение оно находит в преемственности и общности фундаментальных молекулярных, клеточных и системных механизмов развития и жизнедеятельности. Такая преемственность (наследственность) сочетается с изменчивостью, позволяющей осваивать в пространстве и времени новые жизненные условия (экологическая и эволюционная пластичность), достигать высоких уровней структурно-функциональной организации (см. п. 1.4.4).
Восходящие к Ч. Дарвину представления об эволюции, в немалой степени инициированные потребностью объяснить разнообразие живых форм и природу механизмов их смены и нарождения, не касаются вопроса о том, что делает жизнь жизнью во все времена. Их необходимо дополнить с учетом специфической функции живых форм в «экономике» природы как фактора интенсификации и стабилизации земных вещественно-энергетических круговоротов и потоков (см. п. 1.4.3) - планетарная геохимическая роль живого вещества (В.И. Вернадский). В связи со сказанным эволюцию живого (или жизни) следует представлять не только как видообразование, но так же как преобразование биосферы, в ходе которого эволюционируют сообщества (экосистемы, биоценозы), историческая динамика которых обусловлена эволюцией видов. Сближение двух эволюционных парадигм - эволюции видов (таксонов) и эволюции экосистем и биосферы делает вклад эволюционной идеи в обоснование тезиса о единстве мира жизни особенно весомым.
Теория эволюции обращает внимание на условность граней между неживой и живой природой планеты, между живой природой и человеком. В соответствии с геохимической гипотезой происхождения жизни обосновано допущение, что важнейшие атрибуты жизни - самовоспроизведение на базе аутокатализа (матричный синтез), использование высокомолекулярных соединений углерода (нуклеиновые кислоты,
белки), сохранение во времени существующей и наработка новой биологической информации, прогрессивное усложнение структур на основе случайной изменчивости и отбора могли возникнуть на «добиологиче-ском» этапе истории планеты.
Закономерностям эволюции биологических форм не противоречит появление человека - социального и одухотворенного существа, жизнь которого неотделима от принципа клеточной организации структур и функций, молекулярно-биологических, генетических и экологических законов бытия. Эволюционная теория показывает истоки биологических механизмов развития и жизнедеятельности, предпосылки интеллектуальной и трудовой деятельности людей, то есть того, что относится к их биологическому «наследству».
Эволюционная идея в ее современном виде, обосновывая заключение о единстве мира жизни, одновременно обращает внимание на разнообразие групп (рис. 1.2) и вариантов структурно-функциональной организации живых форм. С одной стороны, разнообразие обусловливает высокий эволюционный потенциал жизни, то есть способность, изменяясь в деталях, сохранять себя во времени, несмотря на периодическую радикальную смену абиотических условий (сравни покровные оледенения и межледниковые периоды). С другой стороны, необходимость разнообразия диктуется «инфраструктурой экономики» живой природы, которая в целях реализации феномена биогенной миграции химических элементов должна иметь необходимый набор видов (продуценты, деструкторы, консументы), выполняющих в биогеохимических круговоротах разные функции.
Хотя термин «экология» (греч. oikos - дом, жилище, местообитание) введен в биологический словарь Э. Геккелем во второй половине XIX в., по времени реального рождения экологию следует называть наукой XX в. По перспективам и силе влияния на умы и действия людей в ней также должно видеть одну из лидирующих научно-практических парадигм XXI в., причем настолько, что допустимо говорить о формировании экологического стиля мышления. Центральное положение в экологической науке занимают близкие по сути концепции биогеоценоза (В.Н. Сукачев) и экологической системы (А. Тенсли), датируемые первой половиной ХХ в. Обе концепции характеризуют глобальный принцип существования живых форм - только в составе сообществ в закономерном взаимодействии друг с другом. Сосуществование в таких сообществах представляет собой способ включения организмов в естественные вещественно-энергетические (биогеохимические) кру-
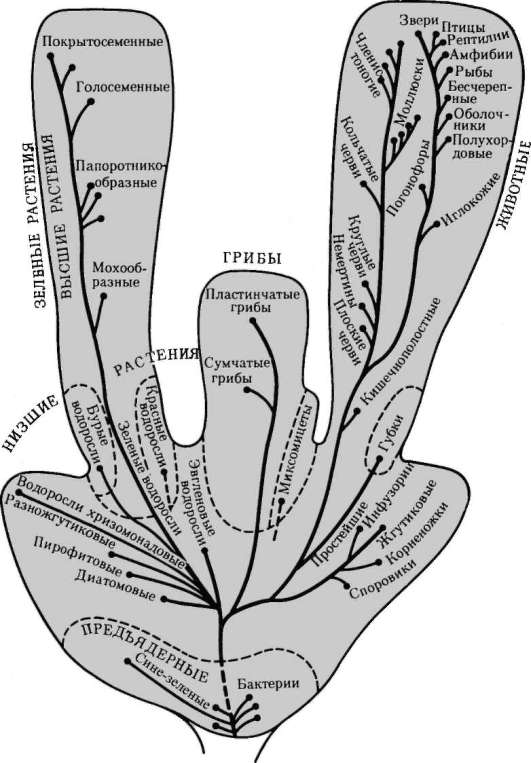
Рис. 1.2. Филогенетические отношения основных групп ядерных эукариотиче-ских организмов - растений, грибов, животных и предъядерных организмов - прокариот. Пунктиром обозначено предполагаемое положение групп
говороты, результатом чего становится интенсификация и стабилизация последних. Для достижения этого результата, однако, необходимо, чтобы сообщество включало продуцентов, возможно, консументов и редуцентов (биоценоз) и находилось во взаимосвязи с элементами атмосферы, гидросферы и литосферы (биогеоценоз).
Крупнейшим обобщением, обозначающим главные тенденции развития биологической науки в условиях присутствия на Земле продвигающегося в интеллектуально-экономическом отношении человеческого общества, является учение о биосфере (термин введен Э. Зюссом в 1875 г. для обозначения живой оболочки Земли), закономерно эволюционирующей в ноосферу (термин введен Э. Леруа в 1927 г. для обозначения оболочки Земли, отмеченной присутствием человеческого общества со всеми атрибутами разумной деятельности). Свой вклад в разработку концепции биосферы на «досапиентной», то есть до появления Я. sapiens, и «сапиентной» стадиях ее существования внес В.И. Вернадский, который обосновал планетарную роль живого вещества (биосферы) через его участие в вещественно-энергетических круговоротах. Охарактеризовав человечество как самостоятельную геологическую силу, он назвал направления и возможные последствия преобразующей деятельности людей.
1.3. определение и фундаментальные свойства жизни
Многообразие форм жизни создает трудности для ее определения как явления. Первые подходы к решению этой задачи основаны на поисках свойств, качественно отличающих живое от неживого. Жизнь определяли как «питание, рост и одряхление» (Аристотель), «стойкое единообразие процессов при различии внешних условий» (Г. Тревира-нус), «совокупность функций, сопротивляющихся смерти» (М. Биша) или «способ борьбы с энтропией» (исследователи начала-середины минувшего столетия), «химическую функцию» (А. Лавуазье), «сложный химический процесс» (И.П. Павлов). Неудовлетворенность этими определениями понятна. Свойства живого, взятые каждое отдельно, лишены исключительности и обнаруживаются в объектах неживой природы.
Определение жизни как «особой, очень сложной формы движения материи» (А.И. Опарин), правильно утверждая ее качественное своеобразие и несводимость биологических законов к законам физическим и химическим, не раскрывает природу своеобразия. Есть мнение, что
жизнь столь же вечна и повсеместна во Вселенной, как материя. Поэтому ее следует считать одним из фундаментальных свойств последней, чем-то сродни гравитации.
Полезны определения, основанные на выделении комплекса свойств, обязательного для живых существ. Одно из них характеризует жизнь как макромолекулярную открытую систему, которой свойственны иерархическая организация, способность к самовоспроизведению, обмен веществ, регулируемый поток энергии. И далее - жизнь представляет собой ядро упорядоченности, распространяющееся в менее упорядоченной Вселенной.
Рассмотрим типичные свойства жизни по отдельности. Живым формам присущ особый способ взаимодействия с окружающей средой - обмен веществ (метаболизм). Его содержание составляют процессы анаболизма (ассимиляция, пластический обмен) и катаболизма (диссимиляция, энергетический обмен). Задачей анаболизма является продукция веществ, необходимых организму в его жизнедеятельности, образовании и обновлении структур. Задача катаболизма - разложение органических соединений на составляющие, которые затем используются для целей энергообеспечения или служат «строительным» материалом. Катаболизм и анаболизм связаны таким образом, что высвобождаемая в ходе катаболических (экзэргонических) процессов энергия используется для приведения в действие энергопотребляющих (эндэргонических) анаболических процессов.
По типу обмена веществ земные живые существа подразделяются на аутотрофные и гетеротрофные организмы. Первые образуют сложные органические соединения из простых неорганических веществ, используя для этого энергию солнца (фотоавтотрофы) или энергию химических связей (хемоавтотрофы). В «экономике» природы ауто-трофам принадлежит важнейшая роль первичных продуцентов (производителей) органики. К ним относятся некоторые бактерии и все зеленые растения. Гетеротрофы не способны синтезировать органику из неорганики. Они получают ее в виде пищи. Окисление пищевых веществ дает этим организмам необходимую для их жизнедеятельности энергию. В «экономике» природы гетеротрофам отведена роль кон-сументов и деструкторов. К ним принадлежат все животные. Люди - также гетеротрофы, правда, своеобразные. Для обеспечения своей жизнедеятельности и, без чего немыслимо бытие человека, общественного обустройства они используют органику не только современного им исторического периода, но и ту, которая образовалась в далеко отстоя-
щие во времени от наших дней минувшие эпохи. Речь идет об ископаемых видах топлива - каменном угле, нефти, природном газе, сланцах.
В многоклеточном организме выделяется внешний и внутренний (внутриклеточный) обмен веществ. Первый происходит между организмом и окружающей средой и заключается в поступлении в него питательных веществ, кислорода, воды, витаминов, минеральных солей и выделении в окружающую среду конечных продуктов обмена, прежде всего углекислого газа и воды. Во втором случае двусторонний обмен устанавливается между клетками и внеклеточной средой, являющейся, по существу, внутренней средой организма.
Для осуществления обмена, с одной стороны, необходим приток веществ извне, а с другой, - неутилизируемые продукты обмена должны выделяться во внешнюю среду. При этом поток веществ через организм неразрывно связан с потоком энергии. Таким образом, организм или клетка в вещественно-энергетическом плане относительно окружающей или внеклеточной среды являются открытыми системами.
Процессы катаболизма и анаболизма представлены химическими реакциями, объединенными в метаболические циклы и каскады (химические превращения), структура которых отличается упорядоченностью во времени и пространстве (прежде всего, речь идет об объеме клетки). Итогом метаболического цикла является определенный биологически значимый результат - на рибосоме из аминокислот образуется полипептид, а поступающий в митохондрию из основного вещества цитоплазмы пируват (анион пировиноградной кислоты) через ряд превращений в матриксе (цикл Кребса) и во внутренней мембране (цепь переноса электронов) органеллы доводится до углекислого газа и воды с образованием высокоэнергизированных (макроэргических) молекул аденозинтрифосфата (АТФ).
Упорядоченность различных составляющих обмена веществ достигается благодаря структурированности (компартментации) объема клетки. Так, предшествующий клеточному делению синтез ДНК происходит в ядре, гидролитическое расщепление поступающих в клетку веществ или разрушение «износившихся» внутриклеточных структур - в лизосомах, упаковка в оболочку выделяемых экзокринной железистой клеткой гранул секрета - в пластинчатом комплексе Гольджи.
Правило компартментации распространяется на оба типа клеточной организации - прокариотический и эукариотический, хотя способы его реализации различны. Различна также эволюционная и экологическая стратегия про- и эукариот. Для первых, не без
оснований называемых доминирующей формой жизни во все времена, выживание и широкое расселение путем освоения разнообразных (в том числе и с «экстремальными» абиотическими характеристиками; бактерия Pyralobus fumaris, например, растет при температуре 113 °С) экологических ниш связаны с исключительной метаболической (биохимической) лабильностью (гибкостью), тогда как для вторых типично преобразование структур при относительной консервативности обменных процессов. Есть мнение, что в биохимии полезны не признаки, а цепи реакций. Оно указывает на возможное своеобразие способов компартментации у прокариот, уходящих корнями в область межмолекулярных взаимодействий. Так, согласно одной из точек зрения, регуляция функциональной (матричной) активности ДНК связана с циклом «компактизация-декомпактизация» двойной спирали. Первый шаг в компактизации ДНК эукариот заключается в образовании нуклеосом. Многими бактериями гистоновые белки не образуются и нуклеосом-ный способ компактизации не используется. Но степень компактиза-ции прокариотической ДНК (укорочение в 1000-10 000 раз) вполне сопоставима со степенью компактизации ДНК эукариотических клеток. Предположительно, свою роль в сверхспирализации ДНК прокариот играют специальные белки, в том числе не гистоны, но основного характера, взаимодействующие с ней.
Значение пространственно-временной упорядоченности процессов в клетке трудно переоценить. К примеру, тело микоплазмы - микроорганизма, занимающего по размерам промежуточное положение между вирусами и типичными бактериями, - по диаметру превосходит атом водорода всего в 1000 раз. Существование микоплазмы, тем не менее, обеспечивается примерно сотней согласованных биохимических реакций. Жизнедеятельность клетки млекопитающего требует для своего обеспечения порядка 10 000 реакций.
В соответствии со вторым законом термодинамики, в энергетически изолированных системах количество энтропии (величина, обратная упорядоченности) с течением времени нарастает. Одним из характерных свойств живых объектов является их способность противостоять росту энтропии, поддерживая присущую им организацию. Образование элементов и сборка из них внутриклеточных структур (анаболизм), отличающихся высокой упорядоченностью, происходят с уменьшением энтропии. Однако параллельно осуществляется окисление пищевых веществ (катаболизм), источником которых является внешняя среда, что сопровождается адекватным увеличением ее энтропии. Поэтому для
полноразмерной биологической системы - «организм и среда его обитания» - изменение энтропии в целом положительно. Ситуация в мире жизни, таким образом, не противоречит упомянутому второму закону термодинамики. Высокая упорядоченность структуры и функций живого существа «оплачивается» энергией, высвобождаемой за счет деструктивных процессов, производимых этим существом в окружающей среде.
Связь живых форм со средой обитания - еще одно непременное свойство жизни. Важное следствие прогрессивной эволюции заключается в том, что параметры внутренней среды организма выводятся из под прямого влияния факторов внешней среды (сравни, температура тела у холодно- и теплокровных животных, онтогенез низших позвоночных, например, рыб и внутриутробное развитие млекопитающих). Свойство живых существ сохранять постоянство внутренней среды, несмотря на колебания показателей окружающей среды, соответствует биологическому понятию гомеостаза.
Жизнь - это процессы самовоспроизведения (размножения) и самообновления. В результате этих процессов, с одной стороны, воспроизводятся организмы определенного типа структурно-функциональной организации, а с другой, - воссоздаются структуры, сменяющие те, которые утрачиваются в ходе жизнедеятельности особи.
Свойства самовоспроизведения и самообновления обеспечиваются использованием живыми формами для сохранения своей организации генетической (биологической) информации. Последняя отбирается по признаку биологической целесообразности в процессе эволюции видов, накапливается в их гено(аллело)фондах и служит основой воспроизведения и жизнедеятельности организмов соответствующего вида в каждом следующем поколении.
Химия биоинформационного обеспечения развития и существования живых форм заключается в использовании уникальных химических соединений, которые в современных условиях не обнаруживаются в неживой природе. Это информационные макромолекулы (биополимеры). Белки, большинство которых в клетке играет роль катализаторов (ферменты). Они обеспечивают реализацию метаболических процессов с достаточной скоростью и требуемым результатом при мягких условиях температуры и давления. Ферменты отличаются специфичностью. Они катализируют превращения групп веществ определенной химической структуры или даже отдельного соединения. Специфичность ферментов, так же как и белков, не несущих каталитической функции (например,
«строительных» - коллагены и эластин соединительной ткани, «транспортных» - гемоглобин), зависит от их первичной структуры, то есть от последовательности аминокислот. Каждое очередное «поколение молекул» определенного белка воспроизводит заданную первичную структуру и, следовательно, несет в себе идентичную информацию. Последнее гарантирует выполнение требуемой функции. Постоянство информации на уровне молекул белка достигается тем, что в основе их воспроизведения лежит матричный синтез. Роль матриц выполняет другой вид информационных биополимеров - нуклеиновые кислоты. Информация, сохраняемая в молекулах ДНК, будучи записанной в виде последовательности троек нуклеотидов, переносится на белок и переводится на язык аминокислотных последовательностей при помощи молекул РНК. Наличие, хранение и реализация специфической биологической (генетической) информации на основе использования уникальных биоинформационных макромолекул - нуклеиновых кислот и белков - составляет еще одно свойство жизни. Информационные биополимеры представляют собой высокомолекулярные соединения углерода.
Оценивая структуру внутриклеточного потока информации (ДНК - РНК - белок - структура и функция), отметим, что его начальное звено (ДНК) соответствует сохраняемой, оберегаемой и закономерно предоставляемой для нужд клетки, но не действующей непосредственно форме информации (назовем ее потенциальной). Информация, перенесенная на молекулы белка, напрямую реализует себя в процессах жизнедеятельности и, следовательно, является действующей формой (назовем ее актуализированной). В приведенной схеме легко узнаются две непременные стороны любого живого существа - его генотип (у прокариот геном) и фенотип.
Жизнь представлена совокупностью пространственно отграниченных друг от друга и от окружающей среды форм - организмов, что соответствует такому ее свойству как дискретность (отграниченность).
Воплощение стартовой наследственной информации (генотип), с которой начинает свое существование организм, в действующую информацию его рабочих молекул и структур (фенотип) происходит в процессе индивидуального развития, или онтогенеза, этого организма, обязательность которого соответствует еще одному универсальному свойству живых форм.
Живые конструкции разного уровня (клетки, клеточные тканевые системы многоклеточных, организм в целом, популяции организмов,
экосистемы) обладают свойством изменять свое состояние в зависимости от колебаний параметров среды, в которой они существуют. При этом совершается работа. Такие изменения в целом имеют приспособительное значение и происходят благодаря механизмам регистрации соответствующих колебаний, анализа поступающих данных, выработки «решения» по содержанию и интенсивности ответа. Названное свойство позволяет рассматривать живые конструкции как кибернетические устройства, в существовании которых задействованы законы информатики - передачи и переработки информации, механизм обратной связи. Термин информация употребляется здесь в широком смысле. Биологическая информация, о которой речь шла выше, качественно и количественно соответствует генетической информации ДНК. Информация в кибернетическом смысле включает и «личный» опыт живого объекта.
В перечне универсальных свойств жизни периода классической биологии значатся раздражимость и возбудимость. В случае раздражимости речь идет о способности живого вещества как такового (протоплазма) реагировать на воздействия, в случае возбудимости - о способности этого вещества переходить в состояние, сопровождающееся выполнением работы. В качестве примера назовем механохимическую систему, обусловливающую мышечное сокращение. Оно происходит в ответ на стимул (нервный импульс) путем сочетанных конформацион-ных (пространственных, объемных) изменений комплекса белков (миозин, актин, тропонин, тропомиозин). По завершении акта сокращения комплекс возвращается в исходное состояние готовности вновь совершить работу.
В процессе жизнедеятельности выполняется не только механическая, но и другие виды работы - химическая (синтезы), осмотическая, электрохимическая, регуляторная. Это требует адекватного энергообеспечения. Решение проблемы энергообеспечения проявлений жизнедеятельности состоит в том, что в эволюции был найден общий энергетический интермедиат (звено, связывающее ката- и анаболизм), который принимает энергию от всех реакций окисления пищи и доставляет ее к месту совершения работы. Наличие универсального переносчика энергии (высокоэнергетический фосфат или АТФ) - одно из свойств жизни.
Среди свойств жизни 2 представляются изначальными, то есть лежащими у ее истоков, ее создавшими и сохраняющими как особое явление материального мира. Это включенность живых форм в процесс
исторического развития (эволюция) и их существование во взаимодействии друг с другом в составе сообществ - биоценозов или экосистем, объединенных в живую оболочку планеты - биосферу.
1.4. происхождение жизни
Гипотезы о происхождении земной жизни нельзя рассматривать даже как рабочие в силу невозможности их экспериментальной проверки. Можно, однако, оценить большую или меньшую вероятность предлагаемых версий, соответствие их общей логике мироздания, законам динамики материи и энергии, в частности термодинамическим. Следуя таким путем, удается составить представление об условиях, факторах и движущих силах, которые, с одной стороны, могли привести к возникновению жизни, а с другой - задают главный вектор ее исторического развития.
Знакомство с проблемой происхождения земной жизни возможно ограничить рассмотрением гипотез панспермии, абиогенеза и биогеохимической.
1.4.1. ГИПОТЕЗА ПАНСПЕРМИИ
Гипотеза панспермии связана с именами выдающихся естествоиспытателей С. Аррениуса, Г. Гельмгольца, В.И. Вернадского, У. Томсо-на (лорда Кельвина). Согласно их видению, жизнь, представляя собой явление космического масштаба, столь же вечна и повсеместна во Вселенной, как и материя. Появление ее на Земле объясняется проникновением «зародышей», постоянно «странствующих» в космическом пространстве. Известны расчеты С. Аррениуса, подтверждающие возможность межпланетного переноса живых конструкций, сходных по размерно-весовым параметрам с бактериальными спорами, под давлением света. В 20-е гг. ХХ в. В.И. Вернадский на основании анализа структуры и геохимии осадочных пород пришел к выводу, что в истории Земли не существовало периода, когда названные породы были бы образованы исключительно абиогенным путем. Углерод заведомо органического происхождения (отличается более высоким содержанием легкого изотопа 12С относительно тяжелого 13С) обнаружен в древнейших на Земле осадочных породах (геологическая формация Исуа в Гренландии, возраст 3,85 млрд лет). Следуя этой линии, допустимо предположить, что жизнь существовала на планете всегда, причем «инфицирование» могло случиться на стадии газопылевого облака.
Прямых свидетельств в пользу космического происхождения земной жизни не существует. Нет также надежных доказательств существования «межзвездной жизни» и ее повсеместного распространения во Вселенной. Хотя метеоритное вещество богато органикой, внеземное происхождение последней требует подтверждения. Во-первых, необходимо исключить возможность загрязнения метеоритов микроорганизмами после достижения ими земной поверхности. Во-вторых, органический материал метеоритов лишен свойства хиральности. Особенностью земных биополимеров является то, что белковые молекулы образованы исключительно левовращающими оптическими изомерами аминокислот, а молекулы нуклеиновых кислот содержат только правовращающие изомеры сахаров - феномен хиральной чистоты. Хотя хиральность биополимеров - непреложный факт, биологический смысл и корни явления далеки от понимания. Одно из объяснений состоит в том, что коль скоро молекула белка представляет собой спираль, то она должна обладать предпосылкой к закручиванию лишь в одну сторону. Спиральная конфигурация позволяет белковым молекулам «ввинчиваться» в жидкокристаллическую структуру воды. Хиральность рибозы и дезок-сирибозы нуклеиновых кислот необходима для образования двойных спиралей. Предполагается также, что хиральность мономеров является условием оформления макромолекулярной системы с белок-поли-нуклеотидным взаимосоответствием. В-третьих, метеоритная органика не воспроизводит еще одной характеристики, которая доказывала бы ее биогенное происхождение. Выше упоминалось, что органический углерод, прошедший фотосинтез, обогащается более легким изотопом 12С. Последнее дает увеличение значений отношения 12С/13С, что не показано для метеоритного материала. Сообщения о наличии на Марсе углерода со смещенным в сторону легкого изотопа отношением требуют подтверждения.
Как бы ни решился вопрос о природе метеоритной органики, гипотеза панспермии не содержит ответа на вопрос, каким образом, в связи с какими обстоятельствами, когда и в каких точках Вселенной жизнь возникла впервые. Открытым остается вопрос о факторах, обусловивших историческое развитие уже земной жизни, давшее современные ее формы. Приведенные вопросы не требуют ответа, если жизнь объявляется одним из фундаментальных свойств материи. В таком случае проблема происхождения жизни ставится в один ряд с проблемой происхождения, например, гравитации.
1.4.2. ГИПОТЕЗА АБИОГЕНЕЗА
У истоков гипотезы абиогенеза стоял Э. Геккель. Суть гипотезы заключается в признании абиогенного (случившегося в отсутствие живых существ) образования органических веществ из неорганических непосредственно на планете.
В середине ХIХ в. Л. Пастер доказал невозможность самозарождения жизни в современных условиях. В 20-х гг. ХХ в. биохимики А.И. Опарин и Дж. Холдейн предположили, что в условиях, имевших место на Земле несколько миллиардов лет назад, образование живого вещества (органики) из неживого было возможным. Среди этих условий - атмосфера восстановительного типа, вода и источники энергии (ультрафиолетовое и космическое излучение, тепло остывающей земной коры, вулканическая деятельность, радиоактивный распад элементов, атмосферные электрические явления), приемлемая температура, отсутствие живых существ. Линия жизни выстраивалась следующим образом:
1) образование атмосферы из газов (метан, оксид и диоксид углерода, аммиак, сероводород, цианиды), служащих «сырьем» для синтеза органических соединений;
2) абиогенное образование простых органических веществ, в том числе мономеров современных биополимеров (аминокислоты, азотистые основания, сахара, АТФ и другие мононуклеотиды);
3) полимеризация мономеров в полимеры - белки (полипептиды) и нуклеиновые кислоты (полинуклеотиды);
4) концентрация абиогенной органики в водной среде с образованием «первичного бульона»;
5) обособление в «бульоне» предбиологических дискретных форм более или менее сложного химического состава - протобионтов, проявляющих некоторые свойства живых форм (уплотненный поверхностный слой, имитирующий мембрану, рост за счет поступления веществ извне, «размножение» путем распада с сохранением особенностей химической организации, предбиологический отбор на стабильность и эффективность «закачки» органики из окружающей среды - прообраз обмена веществ);
6) возникновение простейших форм с полной совокупностью свойств жизни - примитивных клеток-гетеротрофов, питающихся органикой «бульона»;
7) биологическая эволюция возникших существ.
Гипотеза абиогенного происхождения жизни прошла по-своему плодотворный путь. Доказано образование органических соединений, в том числе сложных - полипептидов и полинуклеотидов - из неорганических веществ через стадию молекул-предшественниц в условиях, имитирующих те, которые могли существовать на планете в соответствующий период ее истории. Созданы модели протобионтов - коацерваты (гетерогенный химический состав - полипептиды, полинуклеотиды) и микросферы (однородное белковое содержимое).
Вместе с тем рассматриваемая гипотеза не дает ответа на вопрос о природе хиральной чистоты современных биополимеров. Получаемые в эксперименте абиогенным путем аминокислоты представляют собой смесь лево- и правовращающих изомеров, которые имеют равные химические шансы быть включенными в полипептид. Среди синтезированных в лабораторных условиях сахаров оптическая изомерия вообще не встречается. Согласно проведенным расчетам, вероятность случайного возникновения «осмысленной» полинуклеотидной последовательности из смеси мононуклеотидов «бульона» настолько низка, что потребное на это время на много порядков (!) превосходит время существования даже не Земли, но Вселенной (15-20 млрд лет). Далее, «первичный бульон», являясь исчерпываемым ресурсом, был бы быстро уничтожен, если бы, следуя предлагаемой сторонниками гипотезы схеме, первыми живыми существами оказались гетеротрофы. Данные об исключительно раннем появлении на планете углерода со смещенным в сторону легкого изотопа соотношением 12С/13С (см. выше - формация Исуа в Гренландии) указывают на существование уже тогда фотосинтеза, что говорит об одновременном возникновении авто- и гетеротрофности. Это подтверждает сравнение современных строматолитов цианобактериаль-ных матов и ископаемых строматолитов докембрийского периода (см. п. 1.4.4). В обоих случаях есть основания говорить о высокоинтегриро-ванных сообществах прокариотических организмов, различающихся по типу питания, то есть о присутствии среди первых «обитателей» планеты одновременно продуцентов (производители органики из неорганики), возможно, консументов (потребители органики для создания собственной органики) и редуцентов или деструкторов (разрушители органики), автотрофов и гетеротрофов. В свете сказанного жизнь на Земле сразу же складывалась как экосистема. В рамках гипотезы абиогенеза не нашел удовлетворительной проработки также вопрос о переходе от «осмысленных» биополимеров (полипептиды и полинуклеотиды), даже если признать факт их образования в «бульоне», к клеточной организации.
1.4.3. ГЕОХИМИЧЕСКАЯ ГИПОТЕЗА
Наиболее, видимо, правдоподобная на настоящее время гипотеза о происхождении жизни исходит из того, что живая составляющая формировалась как дополнение к существовавшим на планете геохимическим вещественно-энергетическим круговоротам.
Эта гипотеза согласуется с положениями неравновесной термодинамики (И. Пригожин) и теории самоорганизующихся систем (У. Эшби). В открытых системах в сильно неравновесных условиях (например, земные геохимические круговороты, провоцируемые разницей температур космического пространства и вещества планеты) могут самопроизвольно образовываться упорядоченные структуры. Последние получили название диссипативных в связи с их свойством рассеивать пропускаемую через себя энергию. В таких системах диссипация (рассеивание и, следовательно, безвозвратная потеря системой энергии) выполняет не деструктивную, а созидательную роль, придавая им статус самоорганизующихся. Отличительная черта диссипативных самоорганизующихся систем, к которым принадлежат живые организмы, заключается в их способности эволюционировать и, таким образом, сохранять состояние приспособленности и, следовательно, себя самих при изменении условий. Для этого они используют, с одной стороны, информацию об опыте самосохранения, приобретенную ранее, а с другой - создают новую информацию как предпосылку к самоизменению или самокоррекции под предстоящий опыт. В процессе такого самоизменения происходит усложнение системы.
Для достижения этих результатов (сохранение во времени состояния приспособленности или самосохранение и, параллельно, усложнение в ходе самокоррекции) необходимо выполнение ряда требований. Так, должен действовать автокаталитический механизм самовоспроизведения диссипативной структуры. Им может быть, например, матричный синтез (см. образование ДНК, РНК и полипептидов в клетке). Для сохранения структуры в перспективе необходим постоянно действующий механизм пополнения и обновления информации. Новая информация появляется, если самовоспроизведение системы происходит на фоне «информационного шума» (см. мутации) с участием механизма естественного отбора. Отбор называют «естественным», если материалом для него служит набор случайных по информационному содержанию вариантов (см. феномен неопределенной генотипической изменчивости, резерв наследственной изменчивости). Эффект прогресса воз-
никает, если отбирается и сохраняется информация, способствующая интенсификации функции диссипации (см. адаптации общего значения, обеспечивающие приспособление к широкому кругу условий). Описываемые события вполне соответствуют тому, что происходит в эволюционирующем мире земной жизни.
«Отбираемые», самоинструктирующиеся и самовоспроизводящиеся системы, способные менять структуру в сторону ее усложнения, известны и вне биологии, в частности, в химии. Они получили название (авто) каталитических гиперциклов (М. Эйген).
Сторонники геохимической гипотезы появление жизни связывают с существовавшими на заре истории Земли геохимическими вещественно-энергетическими круговоротами, приводимыми в движение температурным градиентом между более теплой планетой и холодным космосом. Добавлением биологической составляющей, превращающим геохимические процессы в биогеохимические, достигалась интенсификация движения химических элементов и, как следствие, стабилизация структуры круговоротов. Первое - за счет использования постоянного внешнего источника энергии (солнце), второе - благодаря возможности создать в световую фазу резерв энергии (органика фотосинтетического происхождения), расходуемый в темновую фазу, а также повышению степени замкнутости циклов. «Борьба» за энергию составляла основу конкурентных отношений между гиперциклами. Один из выигрышных путей состоял в объединении и согласованных изменениях (коэволюция) различающихся по исходным параметрам гиперциклов. Результат - образование более эффективных (как движущая сила вещественно-энергетических круговоротов) и устойчивых конструкций следующего порядка сложности - иерархия структур, принцип экосистемы.
Усложнение самоорганизующихся систем характеризуется важным следствием. По достижении критического уровня структура становится самоподдерживающейся (Дж. фон Нейман). В происхождении земной жизни этому критическому уровню соответствовал, видимо, «сверхгиперцикл» с типом организации, описываемым сейчас словом «клетка».
Рассматриваемая гипотеза происхождения жизни отличается качественным своеобразием. Она является геохимической, в то время как гипотезы панспермии и абиогенеза - биохимическими. Привлекательность геохимического подхода состоит в том, что он возвращает жизнь как явление в область «обычных», то есть термодинамически по-
нятных событий. Определяя специфическое место жизни в «экономике» планеты как эффективного инструмента оптимизации и стабилизации вещественно-энергетических круговоротов, геохимический подход позволяет отойти от фактора случайности в ее возникновении (гипотеза абиогенеза) или же умозрительной оценки ее как изначального и неотъемлемого свойства материи (гипотеза панспермии).
Геохимическая гипотеза, тем не менее, оставляет без рассмотрения события, которые могли происходить на стадии абиогенного образования ключевых органических молекул и специфических биополимеров. Так, закономерен вопрос, каким образом было преодолено противоречие между необходимостью иметь одновременно фермент (полиме-раза), управляющий образованием информационной макромолекулы (ДНК или РНК), и саму эту молекулу, кодирующую структуру названного фермента (вопрос средневековых схоластов: «Что первично - яйцо или курица?»).
1.4.4. ЖИЗНЬ ВОЗНИКАЕТ КАК СООБЩЕСТВО
Геохимическая гипотеза не противоречит палеонтологическим данным, согласно которым с самого начала жизнь была представлена в виде сообществ (ценозов). Эти «первые» в истории жизни (био) ценозы представляли собой не набор организмов, а систему (био)химических реакций. Природу указанных реакций в наши дни охарактеризовать нельзя. Первый шаг биопоэза (биогенеза), таким образом, состоял в возникновении метаболизма («доорганизменный» этап). Средой формирования первичного метаболизма, включавшегося в геохимические (неорганические) круговороты, мог быть водный гель.
Организмы как обособленные единицы жизни появились лишь на определенной стадии биопоэза (биогенеза). Найденные биохимические следы жизни (3,85 млрд лет, Гренландия) на 400 млн лет старше, чем следы самых древних клеток (3,4-3,5 млрд лет, Австралия и Южная Африка). На пути от первых ценозов к примитивным организмам оформилось такое неотъемлемое свойство жизни, как наследственность. Первичным материальным субстратом этого свойства считают не ДНК,
а РНК.
Примером простейших сообществ, сохраняющихся поныне, служат гидротермы (гидротермали), находящиеся на значительных глубинах в зонах выброса горячих газов, растворов и взвесей из разломов океанического дна - «черные и белые курильщики». В устьях гидро-
терм обнаружены прокариотические биоценозы из литотрофных (в переводе с греческого - «камнееды», хотя пищей им служат газы - водород, углекислый газ и др.; предпочтителен перевод - «питающиеся неорганикой») бактерий из наиболее древней по времени появления на Земле группы архебактерий (Archaea). Они, восстанавливая углекислый газ и серу, окисляя водород, выделяя метан и сероводород, создают запас энергии. Последняя используется в реакциях хемосинтеза, органическим продуктом которого может быть глюкоза. Часть органики потребляется членами гидротермальных биоценозов, другая - выносится из зоны гидротермали в воду с приемлемой для живых форм температурой и растворенным в ней кислородом. Геохимическая функция описанных литотрофных бактерий состоит в том, что они, образуя из неорганики земных недр органику и выполняя, таким образом, роль истинных первичных продуцентов, включают в биологические круговороты основные химические элементы - серу, азот, углерод и водород. Так как не вся производимая в гидротермалях органика утилизируется на месте, круговорот оказывается незамкнутым. Известно, что описанные литотрофы и продукты их жизнедеятельности служат пищей массе организмов, среди которых есть и крупные беспозвоночные, например, головоногие моллюски. Эти организмы завершают утилизацию, возвращая планетарным круговоротам химические элементы.
Сходными событиями был, видимо, отмечен в истории Земли переход от геохимических к биогеохимическим круговоротам. Ценозы литотрофных архебактерий как первичных продуцентов органики формировали базу для более сложной и разнообразной жизни. Следующий шаг мог состоять в появлении аэробных гетеротрофных прокариот, окисляющих выбрасываемую из зоны гидротермалей органику до углекислого газа и воды, тем самым замыкая теперь уже биогеохимические круговороты.
На реликтовую природу гидротермальных сообществ указывают, с одной стороны, принадлежность их членов к группе архебактерий, а с другой - условия существования. Оптимум роста литотрофов - 110 °С (так как в океанических глубинах давление достигает колоссальных величин, точка кипения воды сдвигается на десятки и сотни градусов - принцип скороварки). Во времена зарождения жизни мировой океан был представлен кислым кипятком и под большим давлением дули метановые и аммиачные ветры.
Примером происхождения земной жизни по геохимическому сценарию, причем сразу в виде сообществ, являются цианобактериальные
маты, обнаруживаемые на мелководье в пересоленных лагунах (Сиваш в Крыму, оз. Балхаш в Казахстане). Мат (в переводе с греческого - «каменный ковер») - это сообщество прокариотических организмов, обитающих на небольших по размерам рифовых структурах (строматолиты). Такие сообщества присутствуют в современной биоте Земли, но их следы находят также в докембрийских (более 540 млн лет назад) осадочных породах. В структуре мата выделяют верхний слой или поверхность роста, который заселен, с одной стороны, автотро-фами - производящими кислород фотосинтезирующими цианобак-териями, а с другой - гетеротрофными бактериями - облигатными (обязательными, строгими) аэробами. Одна группа микроорганизмов следующего слоя или подкладки - это автотрофы, которые, используя в качестве источника водорода не воду, а, например, сероводород, осуществляют бескислородный фотосинтез. Вторая группа образована микроорганизмами - факультативными аэробами и гетеротрофами. Ниже поверхности роста и подкладки находится слой, заселенный детритояд-ными анаэробами. Цианобактериальный мат, таким образом, является сообществом прокариот со сложной трофической структурой. Два верхних слоя - это продуценты и консументы первого порядка (аналоги, соответственно, «растений» и «травоядных»), соединенные в цепь пастбищного типа. Третий слой - это редуценты («падальщики»), которые получают органику из верхних слоев по детритной цепи. В сравнении с гидротермальным биоценозом, он характеризуется замкнутыми круговоротами кислорода и органических веществ.
В приведенных примерах один из предполагаемых аналогов раннего биогенеза (гидротермы) размещается в океанических глубинах, тогда как второй (цианобактериальные маты) - это высыхающие лужи. С учетом условий, характеризовавших планету несколько миллиардов лет назад, здесь нет противоречия. Атмосфера Земли того времени, состоявшая, в том числе, из углекислого газа, создавала гораздо более высокое давление (такое, как сейчас на Венере), а гребни океанических хребтов с разломами нередко поднимались над поверхностью тогда неглубокого горячего океана, богатого отмелями.
Причисление живых форм к самоорганизующимся системам, отвечающим правилам, распространяющимся на природу в целом, позволяет увидеть механизм естественного перехода от неживого к живому - через эволюцию и коэволюцию автокаталитических геохимических гиперциклов с включением в них биогеохимических сегментов (за счет активности древнейших прокариотических сообществ,
аналогичных цианобактериальным матам или литотрофным ценозам «курильщиков» современности).
Палеонтологический факт состоит в том, что в раннекембрийских отложениях (возраст 540 млн лет или 1/7 длительности существования земной жизни) обнаруживаются ископаемые представители почти всех подразделений и типов современного животного мира, включая хордовых. Это означает, что в предшествующие эры, объединяемые в докембрий, нашли решение все принципиальные задачи вещественно-энергетического, информационного и структурно-функционального обеспечения жизни. При этом был создан высочайший эволюционный и экологический потенциал. Гиперциклы на основе углеродсодержащих высокомолекулярных соединений (нуклеиновая кислота-полипептид), матричный синтез и естественный отбор среди самовоспроизводящихся с «информационным шумом» (мутации) структур как геохимическая сила возникли с первыми водоемами - 4,0-3,8 млрд лет назад. Переход от геохимических к биогеохимическим круговоротам, оформление первых экосистем и первичной биосферы датируют временем в 3,1 млрд лет назад. Это был прокариотный мир, уже освоивший «традиционные» пути энергообеспечения жизненных процессов (анаэробный и аэробный, брожение, фото- и хемосинтезы) и типы питания (автотрофность, гетеротрофность, детритофагия). В биоинформационной области стабилизировалась величина «информационного шума» (10-5-10-7 мутаций на локус ДНК за поколение). Тогда же сложились, по-видимому, механизмы коррекции ошибок в молекулах ДНК.
1.4.5. ОТ ПРЕДЖИЗНИ К ЖИЗНИ: ОФОРМЛЕНИЕ ПОТОКА БИОЛОГИЧЕСКОЙ ИНФОРМАЦИИ
Живые формы представляют собой дискретные, структурированные, гомеостазированные, самоинструктирующиеся, самовоспроизводящиеся и «отбираемые» системы, что требует наличия у них свойства наследственности. В земной жизни оно обеспечивается информационными макромолекулами - ДНК, РНК и белками, являющимися главными участниками потока биологической (генетической) информации и находящимися в закономерных функциональных отношениях. Вопрос о том, как возник механизм информационного обеспечения жизненных процессов или передачи биоинформации в ряду поколений, до сих пор не имеет однозначного ответа. С ним, однако, связано понимание, каким образом совершался переход от «преджизни» к жизни, от добиоло-
гического к биологическому периоду истории планеты. Три момента здесь представляют интерес. Первый - в силу каких обстоятельств возникли «осмысленные» нуклеотидные последовательности, являющиеся молекулярными «текстами» жизни. Второй - каковы корни механизма редупликации «осмысленных» полинуклеотидов. Третий - как произошло сопряжение двух систем записи биологической (генетической) информации («языков жизни») - использующей нуклеотиды (ДНК, РНК) и аминокислоты (белки). Случайное возникновение из смеси предшественников «осмысленной» нуклеотид-ной последовательности исключается, так как вероятность указанного события ничтожна. Время, необходимое для того, чтобы несколько килограммов типографского шрифта, сброшенного с крыши небоскреба, сложились в текст на известной странице романа «Война и мир», даже не в разы, но в порядки превосходит время существования Вселенной (15-20 млрд лет).
Из предлагаемых версий, содержащих суждения по указанным моментам и иногда откровенно называемых учеными «сказками», приведем одну. На переходном этапе на планете существовал РНК-овый мир, в котором веществом наследственности была РНК. Ее преимущество в сравнении с ДНК состоит в том, что, кроме способности быть носителем информации, РНК обладает ферментативной активностью. Одни рибозимы (РНК-ферменты), катализировавшие на этапе преджиз-ни собственный синтез (самовоспроизведение, ауторепликация), дали элементы аппарата наследственности. Другие катализировали образование полипептидов (протеиноидов). В дальнейшем они дали аппарат синтеза белка (рибосомы, транспортные РНК). Ряд минорных (редких) нуклеотидов, в частности из числа производных 5-оксиметилурацила, являются стереохимическими аналогами аминокислот - цистеина, аргинина, тирозина. Это могло привести к «согласованному» отбору (а в дальнейшем к функциональному сопряжению) полинуклеотидов и про-теиноидов по критерию химического подобия, возможно, вплоть до некоторой взаимозаменяемости в геохимических круговоротах. Информация о каталитической активности конкретных протеиноидов могла фиксироваться полинуклеотидами. В сравнении с настоящим временем, когда в составе РНК обнаруживается, в основном, 4 нуклеотида, в период зарождения жизни число активно используемых «строительных блоков» было, по-видимому, больше - не менее 30 - за счет редких форм. В существовавших тогда условиях (температура порядка 100 °С) такие формы возникали чаще. Это увеличивало разнообразие РНК-
полинуклеотидов. В тот же период количество аминокислот (всего их обнаружено порядка 180), используемых для построения полипептидов (протеиноидов), возможно, также превосходило известное сейчас стандартное число 20, повышая разнообразие в этом классе макромолекул. Все это оптимизировало условия для отбора и подбора.
Переход функции практически исключительного носителя биологической (генетической) информации к ДНК был, возможно, обусловлен ее большей в сравнении с РНК химической устойчивостью. Макромолекуляр-ная организация ДНК в виде двойной спирали, однако, существенно усложняет процесс репликации. Хотя это не имеет эволюционно-биологического объяснения, в пользу изложенных выше представлений косвенно свидетельствует тот факт, что инициация репликации ДНК современных эука-риот требует РНК-затравки (РНК-праймера).
Предполагается, что в развитии жизни на планете существовал этап, представленный группой «околоклеточных» или «предклеточных» форм, отличающихся благодаря несовершенству механизма репликации значительным разнообразием организации и метаболизма и давших начало клеточной форме жизни как таковой. На этом этапе заметную роль в формировании геномов мог играть «горизонтальный» перенос фрагментов ДНК. По мере совершенствования биоинформационных механизмов интенсивность обмена генетическим материалом между живыми формами «одного поколения» снижалась с переходом приоритетов к «вертикальному» (в ряду поколений) переносу биоинформации. В таких условиях сложились предпосылки к дивергентному развитию и, как следствие, возникновению нескольких относительно самостоятельных направлений эволюции.
Оформление структуры потока биологической информации и, таким образом, переход к собственно биологической эволюции означает появление на Земле прокариотической клетки. Согласно сравнительным данным о последовательности нуклеотидов в геномах микроорганизмов среди современных прокариот выделяют две парафилитические, то есть развивающиеся исторически независимо, группы (две подимперии или два царства в разных систематизациях) - Bacteria (Eubacteria) и Archaea (Archaebacteria). Представители этих групп не имеют общего эволюционного предка и, следовательно, не состоят в эволюционном родстве. Последнее хорошо согласуется с выделением в истории становления жизни этапа, представленного «предклеточными» формами с лабильными геномами. Именно на этом этапе могли оформиться два самостоятельных «корня» названных групп прокариот.
Группа Archaea отличается разнообразием и представлена, в основном, экстремофилами, т.е. формами, обитающими в экстремальных условиях (температура, соленость, значения рН). В этой группе, однако, встречаются формы, заселяющие среды с обычными для других прокариот условиями. Недавно описаны представители группы Archaea (продуценты метана), предположительно вовлеченные в развитие периодонтита (periodontitis) у людей.
Эукариотическая клеточная форма организации жизни возникла позже. Геномы и механизмы потока информации у эукариот несут черты, указывающие на эволюционную связь, в одной своей части с группой Bacteria, тогда как в другой - с группой Archaea.
1.4.6. УЗЛОВЫЕ ПУНКТЫ ИСТОРИЧЕСКОГО РАЗВИТИЯ ЖИЗНИ
Переход к более эффективным механизмам энергообеспечения, основанным на использовании кислорода, не мог произойти ранее 1,9-1,7 млрд лет, когда концентрация кислорода в атмосфере Земли достигла и превысила 1% (точка Пастера). Появление в биоте планеты эу-кариот датируется временем 2,0-1,5 млрд лет назад. С их появлением открылась перспектива наращивания в клетках объема генетического материала и, следовательно, биологической информации. Одновременно кольцевая форма молекул ДНК прокариот меняется на линейную, реализуется хромосомный принцип упаковки («расфасовки») наследственного материала, обеспечивший использование биоинформации по частям, более тонкую регуляцию генетических функций, гарантирующий получение дочерними клетками в процессе митоза количественно и качественно полноценной наследственной информации.
Следующий перспективный шаг состоял в появлении многоклеточных форм, что датируется поздним докембрием (600-540 млн лет назад). Многоклеточность стала путем к преодолению физиологических пределов размеров тела (суть проблемы - соотношение объема и площади поверхности). Освоение многоклеточности потребовало решения ряда задач - регуляции численности клеток, дифференцировки и интеграции их деятельности в интересах организма, клеточной рецепции и межклеточного общения, охраны постоянства внутренней среды. С этим типом структурно-функциональной организации связана линия дальнейшего прогресса: хордовые (от 600 млн лет назад), позвоночные (495-445 млн лет назад, появляется иммунный контроль клеточного и белкового состава организма), амниоты (354-290 млн лет назад, опти-
мизируется эмбриогенез, исключительное значение приобретает легочное дыхание, полностью разделяется артериальная и венозная кровь, возникают теплокровность, активная терморегуляция, эффективный гомеостаз как путь к снижению зависимости от среды), млекопитающие (250-200 млн лет назад, оформление экосистем современного типа, включая антропоэкосистемы).
Представлениям, которыми жизни отводится роль способа оптимизации планетарных вещественно-энергетических процессов, не противоречит появление в ходе эволюции человека. «Необычность» последнего в сравнении с другими формами обусловлена оригинальным решением проблем как вещественно-энергетического, так и информационного обеспечения жизнедеятельности и развития. Во-первых, - это доступ к закрытым для остальной жизни источникам энергии (ископаемые виды топлива, энергия пара, ветра или падающей воды, ядерная энергия) и возможность осознаваемого контроля над ее (энергии) использованием. Во-вторых, - это расширение информационной базы за счет программы социального (культурного) наследования. Благодаря этим особенностям человечество, оставаясь в биологическом плане одним видом, заняло в «экономике» Земли место самостоятельной преобразующей геологической силы. Не имеющее аналогов свойство вида Я. sapiens заключается в способности на протяжении всей истории существования увеличивать свою численность. В контексте геохимической гипотезы происхождения жизни появление людей означает трансформацию биогеохимических вещественно-энергетических круговоротов в антропобиогеохимические, а биосферы в ноосферу.
Выделение линии гоминид произошло 5-6 млн лет назад. «Архаичные формы» человека жили на Земле 300-40 тыс. лет назад. Люди современного типа появились 70-30 тыс. лет назад. Приводимые датировки допускают корректировку в зависимости от новых находок науки.
1.5. стратегия жизни. приспособление и прогресс, согласованная эволюция, принцип экосистемы
Знакомство с обитателями (биотой) планеты на любом из этапов развития жизни свидетельствует о многообразии форм сосуществовании организмов, различающихся по общему плану и деталям строения, времени появления в эволюции, месту, занимаемому в структуре биоценозов и экосистем. И в наши дни мир жизни представлен, наряду с эу-кариотами, микроорганизмами и сине-зелеными водорослями, то есть
прокариотами. На фоне разнообразия многоклеточных эукариотиче-ских форм имеется значительное число видов одноклеточных эукариот. Варианты живых форм, возникающие когда-либо, сохраняются в биоте планеты так долго, как долго существуют, пусть на ограниченной территории, климатические, биогеографические, гидроминералогические и иные условия, удовлетворяющие жизненным запросам организмов соответствующего структурно-функционального типа. Такое возможно благодаря закономерно реализуемой в ходе эволюции приспособительной (адаптационной) составляющей стратегии жизни.
Под приспособлением или адаптацией понимают структурное, функциональное, поведенческое или любое другое свойство живых форм, повышающее их шансы на «биологический успех» (выживание + эффективная репродукция) в соответствующем местообитании. Следует различать понятия - приспособление или адаптация (англ. - adaptation) и приспособленность или адапти-рованность (англ. - fitness). Первое используется для обозначения отдельных свойств и признаков, с помощью которых решаются конкретные жизненные задачи - например, ротовой аппарат сосущего или колющего типа насекомых, короткий пищеварительный тракт хищных и длинный со специализированными отделами - травоядных млекопитающих, термо-, хемо-, вибросенсорные системы животных. Второе понятие обозначает соответствие организма всему комплексу условий, которым характеризуется среда обитания. Ранг возникающих в процессе исторического развития адаптаций различается. Одни, гарантируя успех в конкуренции за определенный (иногда «экзотический»), например, пищевой ресурс, способствуют выживанию в узких рамках условий существования. Так, свиные, лошадиные или человеческие аскариды выживают и развиваются, что зафиксировано в названии видов, в организме только «своего» хозяина. Есть приспособления иного ранга - общего значения. Они обеспечивают «биологический успех» в широком диапазоне условий. К таковым относятся, например, амниотическая оболочка сухопутных животных, создающая для зародыша «искусственный водоем», трубчатая нервная система высших животных в сравнении с узловой или ганглионар-ной членистоногих. И среди млекопитающих, и среди насекомых имеются «общественные» формы, но человек - это млекопитающее. Необходимым условием существования живых существ в данный период времени является их соответствие среде (приспособленность) благодаря возникающим адаптациям, слагающимся в определенный комплекс. Ранг адаптаций (приспособлений) проявляется в ходе последующей эволюции.
В процессе исторического развития органического мира закономерно отмечается усложнение структурно-функциональной организации живых форм, что отвечает прогрессивной составляющей стратегии жизни. Названное направление приводит к появлению организмов, удовлетворяющих требованиям морфофизиологического прогресса. Это состояние, приобретаемое группой в ходе эволюции, дает возможность части ее представителей расселиться в адаптивной зоне (среде обитания) с более разнообразными и сложными, оказывающими более сильное давление на организм условиями (суша в сравнении с водной средой, трехмерное пространство в сравнении с плоскостью, социум в сравнении с природой досапиентного периода). Такое достигается благодаря появлению существенных изменений в строении, метаболизме, физиологии и, в немалой степени, в поведении организмов, расширяющих их приспособительные возможности за рамки обычных для пред-ковой группы. Из трех главных сред обитания - водная, воздушная и наземная - последняя является наиболее нагрузочной. Соответственно, выход на сушу в группе позвоночных был отмечен радикальными преобразованиями дыхательной (переход к исключительно легочному типу дыхания), сердечно-сосудистой (см. разделение венозной и артериальной крови путем изоляции малого и большого круга кровообращения, четырехкамерное сердце) систем, конечностей (тетраподизация или четвероногость), кожных покровов (избавление их от функции дыхания), скелета головы (увеличение объема мозгового черепа).
Есть немало примеров, когда численность особей в группе от поколения к поколению растет, территория (ареал) расселения расширяется, в ней увеличивается количество таксонов более низкого ранга. Все перечисленное, однако, происходит в одной адаптивной зоне, то есть комплекса условий, сходных по степени сложности и силе давления на организм. В таких случаях говорят о состоянии биологического прогресса группы, которое в принципе соответствует состоянию процветания. Его критерии: увеличение количества особей в группе; расширение ареала расселения; интенсивное видообразование в роде, увеличение родов в семействе и т.д. Из ныне существующих крупных таксонов к процветающим относятся классы насекомых и млекопитающих. Период процветания класса пресмыкающихся (динозавры) завершился 6070 млн лет тому назад.
Прогрессивная составляющая в эволюции не означает простого движения от «низших» к «высшим» формам. Это сложное явление, происходящее в историческом масштабе времени во всех группах живых существ,
включая и те, которые традиционно относят к низкоорганизованным. В современном понимании бактерия (в общем виде прокариотический организм) - это сложное структурно-функциональное единство. Прогрессивная составляющая эволюционного процесса не обошла «доя-дерные» клеточные формы жизни, но эволюционно обусловленные усложнения здесь относятся исключительно к клеточному уровню. Подавляющее большинство прокариот - это одноклеточные организмы. Выделяемые среди прокариот многоклеточные конструкции (миксо-бактерии, существуя часть жизни как одноклеточные, на период размножения образуют колонии, которые дают плодовые тела) никогда не демонстрируют той сложности, которая наблюдается среди истинно многоклеточных эукариотических форм.
Прогрессивная составляющая в эволюции одноклеточных эукариот или простейших проявилась в усложнении внутриклеточного обустройства. Среди клеток высших многоклеточных животных не найти столь сложную внутреннюю организацию, как, например, у инфузории. С другой стороны, организацию метаболизма прокариот следует оценить, в сравнении с эукариотами, как прогрессивную.
Прогрессивные изменения обнаруживаются у современных представителей крупных таксонов истинно многоклеточных животных, независимо от их ранга согласно критериям уровня морфофизиологической организации. В любом случае они касаются структурных характеристик. При этом отмечается качественное своеобразие указанных изменений у низших и высших форм. В первом случае изменяется, усложняясь, структура клеток (сравни клетки внешних покровов плоских червей и позвоночных), во втором - ткани как материал для построения функциональных систем (см. опорно-двигательная, кровеносная, нервная системы, система иммунитета теплокровных позвоночных).
Для образования высокоорганизованных форм, отвечающих требованиям морфофизиологического прогресса, необходим, как подсказывает интуиция, больший объем наследственной информации. Сравнение количества ДНК в ядрах клеток у представителей разных классов подтипа позвоночных не подтвердило это предположение. Место рекордистов Природой отдано рыбам и амфибиям: содержание ДНК в клетке рыбы - американского чешуйчатника - составляет 3540%, а в клетке амфибии - американского протея - 2780% от количества, типичного для клетки млекопитающего (с-парадокс, где с - количество ДНК в гаплоидном наборе хромосом).
Положительная корреляция с уровнем структурно-функциональной организации наблюдается, однако, для уникальных нуклеотидных последовательностей, представленных в геномах однократно (рис. 1.3). Ситуация осложняется тем, что в этой группе есть как кодирующие (смысловые), выполняющие информационно-генетическую функцию непосредственно, так и некодирующие последовательности. На долю первых в геноме человека приходится порядка 2%, а на долю вторых - 49%. На настоящий момент определено количество кодирующих генов у эукариот, очевидно различающихся по уровню морфофункциональ-ной организации - дрожжи (одноклеточные) Saccharomyces cerevisiae, многоклеточные - круглый червь (нематода) Caenorhabditis elegans и человек Яото sapiens: 6 тыс., 20 тыс. и 30-35 тыс., соответственно. Количество смысловых (структурных, кодирующих) генов у человека превосходит аналогичный показатель для нематоды всего в 2 раза, а число структурных генов у червя всего втрое выше, чем у дрожжевой клетки. Таким образом, у эволюционно далеких, прежде всего в понятиях прогрессивной эволюции, видов - дрожжей и круглого червя, нематоды и человека различия в количестве кодирующих генов незначительны. Вместе с тем при совпадающем порядке числа структурных генов репертуар белков (протеом) человека в сравнении с нематодой отличается гораздо большим разнообразием. Основная причина - альтернативный сплайсинг (см. п. 2.4.5.5). Если у человека альтернативный сплайсинг распространяется на пре-РНК-транскрипты 35-60% генов с образованием в среднем 2-3 дефинитивных транскриптов или и(м) РНК на ген, то у круглого червя это 22% генов со средним выходом дефинитивных транскриптов менее двух на ген. Можно заключить, что кодирующий потенциал генома в клетках организмов, эволюционно более продвинутых, выше и проявляется на уровне транскриптома и протеома. Из сопоставления данных о числе структурных генов у одноклеточного эукариота (дрожжевая клетка), у низкоорганизованного (нематода) и высокоорганизованного (человек) многоклеточных эу-кариот следует вывод, что, начиная с определенного этапа, решение проблемы прогресса в эволюции перешло из плоскости приобретения новых генов в плоскость использования других генетических механизмов. Кроме альтернативного сплайсинга, увеличение разнообразия белков при фиксированном числе структурных генов у млекопитающих достигается путем химической модификации ДНК (избирательное полиаденилирование, метилирование).
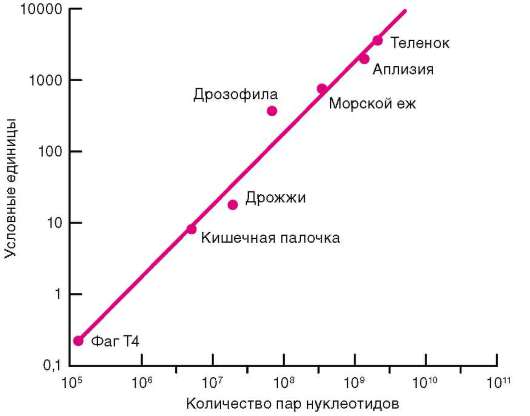
Рис. 1.3. Изменение объема уникальных нуклеотидных последовательностей ДНК в геномах в процессе прогрессивной эволюции
Можно думать, что чем выше уровень морфофизиологической организации, тем больше энергии потребляет живое существо. Показательно в этом плане явление стабилизации уровня удельного потребления энергии в классе млекопитающих, характеризующихся теплокровностью и выделяющихся среди амниот наиболее высоким уровнем организации. Представители названного класса (за исключением человека, для которого рассматриваемый показатель примерно в 4 раза выше), независимо от видовой принадлежности, расходуют за свою жизнь одно и то же количество энергии - 191 600 ккал/кг живого веса. Это наивысший в мире жизни уровень энергопотребления. Указанное количество энергии расходуется с разной скоростью. Животные с высокими показателями основного обмена и активным образом жизни делают это быстрее, отличаясь меньшей продолжительностью жизни.
Можно заключить, что прогрессивное направление стратегии жизни, в целом, характеризуется адекватным биоинформационным и биоэнергетическим обеспечением.
Революционизирующие эволюционные «находки», такие, как эука-риотический тип клеточной организации, многоклеточность, появление хордовых, позвоночных и, наконец, млекопитающих животных, давших в своем развитии приматов, благодаря появлению на планете человека, выделяются в отдельную линию неограниченного прогресса.
Появление в биоте людей означает качественно новое состояние мира жизни. Переход к этому состоянию, хотя он был подготовлен ходом и «наработками» биологической эволюции, означает смену законов, которым следует развитие человечества, с биологических на социальные. Потенциал научных знаний уже сейчас позволяет человеку, правда все еще в ограниченном масштабе, вмешиваться в фундаментальные процессы жизнедеятельности, в том числе генетические (биоинформационные), меняя их ход по своему усмотрению (см. генетически модифицированные организмы). Можно думать, что в связи с активностью человека органический мир Земли вступает в новую фазу своего развития - биотехнологический прогресс.
Согласно главным составляющим эволюционной стратегии (адаптация, прогресс), жизнь представляет собой постоянно приспосабливающийся и самоусложняющийся феномен. Такое восприятие соответствует эволюционной парадигме - методологии, задающей тон в биологической науке со второй половины XIX в. и сложившейся под воздействием идей Ч. Дарвина. Центральное место в эволюционных построениях традиционно отводится процессу видообразования. На второй план отходит очевидное обстоятельство - существование отдельно взятого вида, сколь бы высоким уровнем организации он не отличался, невозможно иначе как во взаимодействии с представителями других видов в составе биоценоза (экосистемы). Эволюционную парадигму следует дополнить экологической или экосистемной, а в перечень главных составляющих, характеризующих стратегию жизни как особого явления материального мира, включить еще одну - биогеохимическую. Согласно этой составляющей, необходимыми атрибутами жизни являются коэволюция и сосуществование организмов разного плана строения и образа жизни, играющих в вещественно-энергетических круговоротах разные, но в одинаковой мере необходимые роли (принцип экосистемы). В таком случае естественное объяснение получают палеонтологические находки, говорящие о том, что, начиная с момента возникновения, мир жизни существует в виде биосферы, глобальное предназначение которой состоит в стабилизации планетарного круговорота веществ и энергии. При этом адаптивная, прогрессивная и биогеохимическая стратегиче-
ские составляющие хорошо соответствуют друг другу. С одной стороны, эффективным фактором стабилизации геохимических круговоротов является интенсификация миграции химических элементов благодаря включению в них живых форм. С другой - возникающие вследствие морфофизиологического прогресса все более высокоорганизованные существа, по определению отличаясь от эволюционных предшественников сниженными значениями энтропии, характеризуются более высоким уровнем вещественно-энергетического обмена.
1.6. иерархическая система жизни. понятие об уровнях организации
Мир жизни представляет собой систему, которой свойственна иерархическая организация. Под системой в науке понимают единство, составленное из значительного числа элементов, которые находятся в закономерных отношениях и связях друг с другом. Главные биологические категории - геном (генотип), клетка, организм, популяция, биогеоценоз, биосфера - представляют собой системы. Иерархической называется система, элементы которой расположены в порядке от высшего к низшему. Так, биосфера слагается из биоценозов (экосистем), представленных популяциями организмов разных видов, совокупность родственных видов образует род, совокупность родов - семейство и т.д. С другой стороны, популяции представлены особями, тела которых имеют клеточное строение, а жизнедеятельность клеток находится под контролем взаимодействующих друг с другом генов. Иерархический принцип организации предусматривает наличие в системе уровней, самостоятельных по происходящим в них процессам, но связанных общностью главной решаемой задачи. Знакомство с уровнями придает конкретность и наглядность представлениям о логике сложных природных явлений (жизнь, организм, клетка, геном, ген) и вносит порядок в изучение этих явлений.
В биологии и медицине вплоть до настоящего времени широко используется классификация уровней организации живого объекта , ориентированная на многоклеточный организм. Такая классификация соответствует состоянию науки о жизни в XIX - первой половине ХХ вв., характеризующемуся стремительным прогрессом в области методов изучения, дающих, с одной стороны, все большее разрешение (микроскоп, электронный микроскоп, индикаторная цито- и гистохимия, рентгеноструктурный анализ), а с другой, - позволяющих визуализа-
цию анатомических и субанатомических структур на живом организме (рентгенография, ультразвуковая диагностика, томография).
В названной классификации выделяемые уровни соответствуют важнейшим, прежде всего с точки зрения функциональной морфологии и патологии, структурам или частям многоклеточного организма, которые, к тому же, являются в разных дисциплинах объектами изучения. Это объясняет, почему выделяемые уровни коррелируют как с размерами структур, так и с разрешающей способностью методов, прежде всего морфофункциональных, биохимических и биофизических, которыми широко пользуются биологи и врачи (табл. 1.2).
Таблица 1.2. Уровни организации (изучения), выделяемые в многоклеточном организме (по Э. Де Робертсу и др., 1967, с изменениями)
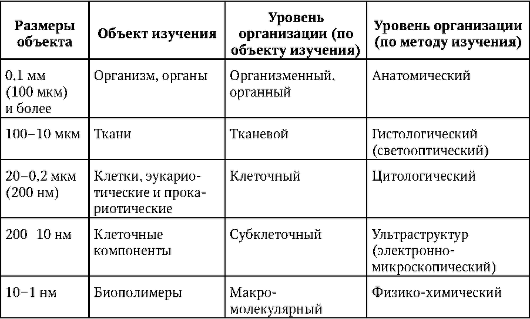
Примечание: 1 м = 103 мм, 1 мм = 103 мкм, 1 мкм = 103 нм, 1 м = 109 нм.
Взаимопроникновение идей и методов различных областей естествознания (физики, химии, биологии), возникновение наук на стыке этих областей (биофизика, биохимия, молекулярная биология) привели к расширению классификации, вплоть до выделения молекулярного и атомарно-электронного уровней. Введение в классификацию молекулярного и атомарного уровней не изменяет главного ее принципа - ориентации на отдельно взятый организм, что не охватывает всего многообразия жизненных явлений.
Возможность исследовать фундаментальные биологические процессы на клеточном, субклеточном и молекулярном уровнях, использование их результатов для создания лечебно-диагностических технологий не является единственной чертой биологии второй половины ХХ в. Типичен плодотворный с точки зрения практических перспектив интерес к процессам в сообществах организмов - популяциях, экосистемах. Это также нашло отражение в современных вариантах приведенной выше классификации, в которую стали включать видовой, биогеоценотиче-ский, биосферный уровни. Такая надстройка носит формальный характер, так как не просматривается объединяющая биологическая задача, решение которой с необходимостью требовало бы интеграции происходящего на заявляемых уровнях (суборганизменных, организмен-ном, надорганизменных) и, при этом, вклад каждого из них был бы специфичен и не дублировался.
Сформулированным выше условиям отвечает классификация уровней организации жизни как особого природного явления, в основу которой положена идея об объединяющей роли эволюционного процесса.
В этой классификации обозначены молекулярно-генетический, клеточный, организменный (онтогенетический), популяционно-видовой, биогеоценотический (экосистемный) уровни (Н.В. Тимофеев-Ресовский, 1958-1961). Ее особенность заключается в том, что отдельные уровни определяются на основе выделения для каждого из них только ему присущих элементарной единицы и элементарного явления. Элементарная единица - это структура (объект), закономерные изменения которой, обозначаемые как элементарное явление, обусловливают специфический для соответствующего уровня вклад в процесс эволюции, т.е. в решение глобальной биологической задачи сохранения и развития жизни. Соответствие выделяемых уровней узловым моментам эволюционного процесса, вне которого не стоит ни одно живое существо, делает их неформальными, распространяющимися на всю область жизни - от до-клеточных форм и прокариот до млекопитающих и человека.
Элементарной единицей на молекулярно-генетическом уровне служит ген - фрагмент молекулы нуклеиновой кислоты (преимущественно, ДНК; генетический материал ряда вирусов представлен РНК), в котором записан определенный в качественном и количественном отношении объем генетической (биологической) информации. Элементарное явление заключается в феномене конвариантной редупликации гена, что означает его самовоспроизведение с ограниченным,
но закономерно случающимся искажением закодированной в гене информации. Путем репликации ДНК происходит копирование (тиражирование) генетической информации, чем обеспечивается сохранность и преемственность, иными словами, консерватизм свойств организмов определенного вида в ряду поколений. Время от времени в силу ограниченной устойчивости к внешним воздействиям (тепловые колебания, ультрафиолетовое и ионизирующие излучения, химически агрессивные свободные радикалы) или вследствие ошибок синтеза, рекомбинации или репарации структура молекул ДНК нарушается, вследствие чего генетическая информация изменяется. В последующих репликациях ДНК нарушения воспроизводятся в молекулах-копиях и наследуются клетками и организмами следующих поколений, чем обусловлена лабильность свойств живых форм. Указанные нарушения, известные в генетике как генные (истинные) мутации, возникают и тиражируются закономерно, что делает процесс репликации ДНК конвариантным, т.е. с изменениями. Благодаря феномену конвариантной редупликации на молекулярно-генетическом уровне обеспечиваются два непременных условия эволюции - наследственность и мутационная (по Ч. Дарвину, неопределенная) изменчивость.
Биологическая информация ДНК (геном, генотип) не участвует в процессах жизнедеятельности непосредственно, но только будучи перенесенной на белок («первичный» фенотип). Учитывая, что естественный отбор происходит по фенотипам особей, такой перенос информации является необходимым условием эволюции. Он осуществляется благодаря механизму матричного синтеза, в котором ДНК выполняет функцию матрицы, но уже для образования молекулы-посредника м(и)РНК, контролирующей образование белка. Отмеченное дает основание рассматривать матричный синтез информационных макромолекул (ДНК, РНК, белки) как принципиальный момент в ряду элементарных явлений молекулярно-генетического уровня.
В реализации генетической информации ДНК в конкретные проявления жизнедеятельности синтез белков представляет собой лишь первый шаг. Он, также как и репликация ДНК, требует для своего осуществления надлежащего морфологического, вещественного (субстратного) и энергетического обеспечения. Структурной, функциональной и генетической основой процессов жизнедеятельности является клетка, представляющая собой элементарную единицу клеточного уровня. Для выполнения своих функций клеткой простейшего микроорганизма микоплазмы требуется порядка 100, а клеткой млекопитающего - при-
мерно 10 тыс. биохимических превращений. Несмотря на различия в объеме выполняемой химической работы (2 порядка), в основе жизнедеятельности клеток обоих типов лежит согласованное по времени и объему сопряженное функционирование потоков информации, энергии и веществ. В таком сопряжении и состоит элементарное явление клеточного уровня. Клетка представляет собой биологическую структуру в одно и то же время необходимую и достаточную, чтобы жизнь существовала и развивалась во всей ее полноте. В силу этого элементарное явление клеточного уровня служит биоинформационной, энергетической и вещественной предпосылкой процессов жизнедеятельности на всех других уровнях организации. Клеточный уровень не назывался Н.В. Тимофеевым-Ресовским и выделяется нами, исходя из дидактических соображений.
Элементарной единицей организменного, более точно, онтогенетического уровня является особь, рассматриваемая во времени, то есть как ее (особи) онтогенез. Закономерные видоспецифичные изменения в продуктивную фазу индивидуального развития составляют элементарное явление рассматриваемого уровня. Эти изменения обеспечивают рост организма, дифференциацию его частей и одновременно интеграцию развития в целостный процесс, морфофунк-циональную специализацию клеток, тканей и органов. Благодаря онтогенезу, происходящему в определенных условиях среды, наследственная информация воплощается в структуры и процессы. На основе генотипа формируется фенотип особи.
Элементарной единицей популяционно-видового уровня служит популяция - совокупность особей одного вида. Объединение особей в популяцию происходит благодаря общности гено(аллело)фонда, используемого в процессе полового размножения для создания генотипов особей следующего поколения. Действие на гено(аллело)фонд популяции так называемых элементарных эволюционных факторов (главные из них - мутационный процесс, колебания численности особей в популяции, миграции, генетико-автоматические процессы, естественный отбор, изоляция) приводит к эволюционно значимым изменениям гено(аллело) фонда в ряду поколений, которые соответствуют элементарному явлению на этом уровне.
Организмы одного вида населяют территорию (ареал) с известными абиотическими параметрами (климат, химизм почв и вод, высота над уровнем моря и т.д.) и взаимодействуют с организмами своего и других видов. В процессе совместного исторического развития (коэволю-
ция) на данной территории представителей разных систематических групп образуются устойчивые во времени, но динамичные сообщества - биогеоценозы (экосистемы), которые служат элементарной единицей биогеоценотического (экосистемного) уровня. Элементарное явление на рассматриваемом уровне представлено вещественно-энергетическими круговоротами и потоками. Ведущая роль в них принадлежит живым организмам. Биогеоценозы, различаясь по видовому составу и характеристикам абиотической части, представляют собой открытые в вещественном и энергетическом плане системы. В своей совокупности они образуют на планете единый комплекс - область распространения жизни - биосферу.
Рассмотренная система включает уровни, отражающие важнейшие биологические явления, вне которых невозможна эволюция и, следовательно, само существование жизни. Хотя элементарные единицы и явления выделяемых уровней различны, все они тесно взаимосвязаны, решая свою специфическую задачу в рамках единого эволюционного процесса.
С конвариантной редупликацией (молекулярно-генетический уровень) связаны элементарные основы этого процесса в виде наследственности, мутационной изменчивости, матричного синтеза. Особая роль клеточного уровня состоит в энергетическом, вещественном и информационном обеспечении происходящего на других уровнях. На онтогенетическом уровне биологическая информация генотипа преобразуется в комплекс фенотипических признаков организма. Возникающий фенотип становится объектом действия естественного отбора. На популяционно-видовом уровне через механизм отбора и избирательной репродукции организмов определяется эволюционная ценность изменений, относящихся по своему происхождению к молекулярно-генетическому, клеточному или онтогенетическому уровню. При этом проявляется иерархический характер межуровневых отношений. К примеру, мутационные изменения ДНК в первую очередь сказываются на функциональных параметрах клеток, далее через клеточные механизмы онтогенеза оказывают влияние на ход индивидуального развития, структурно-функциональный итог которого в виде показателей жизнеспособности и репродуктивного потенциала организма проявляется на уровне популяции. Специфическая роль биогеоценотического уровня состоит в образовании сообществ организмов, приспособленных к совместному проживанию в известной среде обитания и различающихся по своей роли в природных вещественно-энергетических круговоротах.
На этом уровне осуществляется планетарная геохимическая функция жизни.
Уровни, о которых идет речь, отражают структуру эволюционного процесса, одним из результатов которого является человек. Из этого следует, что типичные для выделяемых уровней элементарные единицы и явления характеризуют и людей, правда, с особенностями в силу социальной сущности последних. В пользу сказанного свидетельствует то, что факторы здоровья и патологии человека распределены по всем уровням. Воспроизводство из поколения в поколение людей с достаточным генетическим здоровьем обеспечивается элементарными явлениями молекулярно-генетического уровня - тиражированием и передачей от родителей потомству эволюционно выверенной благоприятной биологической информации, тогда как конвариантный характер редупликации (мутации) несет в себе перспективы наследственной патологии. Благодаря способности клеток к реализации биоинформации в конкретных процессах жизнедеятельности и в организации требуемого взаимодействия организма и среды, устанавливается уровень жизнеспособности, отвечающий критериям здоровья. Нарушение нормального хода внутриклеточных потоков информации, энергии и веществ, приводит к патологии - клеточным дистрофиям, снижению функционального потенциала, малигнизации. Зрелый в структурно-функциональном отношении, жизнеспособный, здоровый организм возникает как результат нормального индивидуального развития, тогда как нарушения последнего приводят к уродствам или снижению общей жизнеспособности. Говоря о вкладе в здоровье онтогенетического уровня, специальное внимание следует уделить старению, которое уже само по себе предусматривает недужность. Здоровье потомства зависит от особенностей гено(аллело)фонда родительской популяции, учитывая наличие в нем неблагоприятных в их фенотипическом проявлении аллелей, составляющих генетический груз. Свою лепту в здоровье на популяционном уровне вносит исторически сложившаяся в популяциях людей система брачных отношений. Первостепенную роль в достижении желаемого уровня здоровья современное здравоохранение отводит условиям и образу жизни, то есть факторам, характеризующим биогеоценотический уровень. К этому уровню относится большинство из идентифицированных на настоящий момент факторов риска утраты здоровья.
1.7. проявление главных свойств жизни по уровням ее организации
В пункте 1.3 рассмотрены свойства жизни как особого природного явления. Полезно вернуться к этой теме с учетом представлений о многоуровневой организации жизни.
Такие свойства, как дискретность, структурированность, про-тивоэнтропийная направленность, вещественно-энергетическая открытость в равной мере присущи клеткам, особям, популяциям и биоценозам (экосистемам), т.е. проявляют себя на всех уровнях.
Наличие генотипа и фенотипа формально характеризует элементарные единицы клеточного и организменного (онтогенетического) уровней. Однако и это свойство относится к жизни в целом, так как генотипы как совокупности генов (нуклеотидных последовательностей ДНК) особей на уровне популяций объединяются в гено(аллело)фонды, которые являются источником генов для особей следующего поколения.
Биоценозы представляют собой не случайные ассоциации популяций особей разных видов, а исторически складывающиеся сообщества взаимоприспособленных организмов разного типа структурно-функциональной организации и разного места в «экономике» природы. Взаимоприспособленность как следствие коэволюции живого населения определенной территории закреплена наследственно в гено(аллело) фондах соответствующих популяций. Совокупность таких гено(аллело) фондов может рассматриваться в качестве общей генетической предпосылки существования биоценоза. Биологическая информация генотипов соответствует потенциальной, а фенотипов - актуализированной, действующей информации. Те же соотношения распространяются на биологическую информацию гено(аллело)фондов популяций и экосистем, с одной стороны, и информацию, актуализированную в фенотипах членов популяции или биоценоза, - с другой.
Непосредственными носителями генетической (биологической) информации являются нуклеиновые кислоты и белки, составляющие элементарную макромолекулярную основу, соответственно, генотипа и фенотипа. С учетом рассуждений, приведенных выше, наличие информационных макромолекул с полным основанием рассматривается в качестве необходимой характеристики не только клетки или организма, но распространяется на всю область жизни.
Способность к росту связывают с индивидуальным развитием организма. Но закономерные циклы развития, включающие изменения
размеров, объемов или распространенности (например, по территории), характеризуют элементарные единицы всех уровней. Репликация ДНК, образование четвертичных структур белков путем объединения полипептидов в функциональный комплекс, рост клетки между делениями, изменение численности особей в популяции, сукцессия биогеоценоза (экосистемы) - вот примеры приложимости рассматриваемого свойства ко всей области жизни.
Результатом временной динамики элементарных единиц разных уровней организации жизни нередко бывает увеличение их количества, т.е. размножение в буквальном смысле. Репликация приводит к увеличению числа биспиралей ДНК, митотические (пролиферативные) циклы - количества клеток, размножение на популяционном уровне - числа особей. Вместе с тем размножение в биологическом понимании - это обязательно самовоспроизведение. Универсальный биологический принцип воспроизведения «себе подобного» лежит в основе сохранения во времени элементарных структур всех уровней и, следовательно, тех элементарных явлений, которые с ними связаны. На молекулярно-генетическом уровне - это двойная спираль ДНК, клеточном - клетка, онтогенетическом - особь, популяционно-видовом - популяция с присущим ей гено(аллело)фондом, возрастной и половой структурой, биогеоценотическом - определенный видовой состав, включающий «своих» продуцентов, консументов и деструкторов. Во всех приведенных примерах самовоспроизведение сопряжено с определенной степенью изменчивости.
Упорядоченность возникает на основе информации, которая, собственно, и воспроизводится в соответствующей структуре или процессе. Первичная («потенциальная») биологическая информация записана в молекулах ДНК. Расчеты показывают, что ее одной недостаточно для кодирования всего многообразия живых структур и комплексов от белковых молекул до биоценозов (экосистем). Дополнительная информация появляется вследствие того, что живые объекты относятся к категории самоорганизующихся, диссипативных систем. Важная черта последних состоит в их целостности, которая проявляется в том, что поведение и судьба элементов, строящих систему, в большей степени определяется ее структурой и в меньшей - свойствами самих элементов. В своем развитии такие системы проходят ряд устойчивых состояний, разделенных периодами неустойчивости (критические периоды), с которыми связано приобретение дополнительной информации. В каждом из таких периодов, отличающихся повышенной чувствительностью
к действию внешних агентов, происходит выбор варианта дальнейшего развития. Эквифинальность, т.е. закономерное достижение системой в итоге развития требуемого результата, определяется внутренне присущими ей «правилами», в соответствии с которыми осуществляются многократные последовательные взаимодействия и изменения элементов разных уровней. Примером поведения самоорганизующейся системы является упорядоченная динамика структур и сменяемость стадий в эмбриональном развитии.
Воссоздание любой структуры сопряжено с выполнением работы. Принцип адекватного энергообеспечения биологических процессов на основе универсального переносчика энергии (высокоэнергетический фосфат, АТФ) проявляется на всех уровнях организации жизни.
1.8. проявление общебиологических закономерностей у людей. биосоциальная природа человека
Уникальное место людей в биоте планеты обусловлено их особым качеством - социальностью. Уже не биологические механизмы, а общественное устройство, производство, труд, интеллект, наука обеспечивают выживание, всесветное и даже космическое проникновение, благополучие человечества. Социальность не противопоставляет людей остальной живой природе. Это качество означает лишь то, что благодаря ему историческое развитие вида Homo sapiens подчиняется не биологическим, а социальным законам.
Человек остается включенным в систему мира жизни. Этот мир складывался и развивался на протяжении большей части истории планеты независимо от человеческого фактора, более того, на определенном этапе своего развития он этот фактор породил. Человечество составляет неотъемлемый компонент биосферы. Благодаря животному происхождению жизнедеятельность человеческого организма основывается на фундаментальных биологических механизмах, которые составляют его биологическое «наследство».
Развитие жизни в одной из ее ветвей привело к появлению примерно 40 тыс. лет назад современного человека, объединяющего в себе биологическое, социальное и духовное. Характер отношений социального и биологического в человеке нельзя представить как их сочетание в некоторой пропорции или подчинение одного другому. Особенностью человеческого биологического является то, что оно проявляется в условиях
действия законов общественного развития (т.е. социальных). Биологические процессы с необходимостью совершаются в организме человека. Им принадлежит фундаментальная роль в определении важнейших сторон жизнеобеспечения и развития. Вместе с тем эти процессы в популяциях людей не дают закономерного результата, обязательного для остального мира живых существ.
Гено(аллело)фонды популяций людей и в настоящее время изменяются в результате мутаций, комбинативной изменчивости, неслучайного подбора брачных пар, дрейфа генов, изоляции, стабилизирующей формы естественного отбора. Вместе с тем в условиях социального бытия естественный отбор утратил свою главную биологическую функцию - видообразование. В таком случае исключается возможность завершенного эволюционного цикла путем достижения закономерного биологического результата - появление новых видов рода Homo. Сохраняющееся же действие элементарных эволюционных факторов оборачивается в отношении человеческих популяций необычными с эволюционно-биологической точки зрения последствиями, например не имеющим по масштабам равных среди других видов организмов генетическим и, следовательно, фенотипическим разнообразием.
1.9. современная система мира живых существ
На любом из этапов своей истории мир живых существ характеризуется многообразием образующих его организмов. Это многообразие обусловлено тем, что в процессе исторического развития появляются разные, в одинаковой степени обеспечивающие приспособление к условиям окружающей среды типы структурно-функциональной организации.
Ч. Дарвин в «Происхождение видов» (1859) указывал на факт, не находящий объяснения с позиций предложенной им же эволюционной теории. В известных на то время науке кембрийских отложениях (возраст 540-520 млн лет) палеонтологи обнаруживали ископаемых представителей почти всех крупных подразделений современного животного царства, тогда как в докембрийских отложениях ископаемых остатков не находили вовсе. Возникал вопрос, как могла появиться кембрийская биота в отсутствие предковых форм, т.е. в отсутствие доказательств предшествующей эволюции. С течением лет палеонтологический «пробел» заполнялся. Выяснилось, что к концу протерозойской эры (2500-540 млн лет) на планете уже существовали представители
всех современных типов животного мира (рис. 1.4), а узловые моменты прогрессивной эволюции животных (эукариотический формат клеточной организации, многоклеточность, двусторонняя симметрия тела, три зародышевых листка, вторичная полость тела или целом, регуляторный тип эмбрионального развития, внутренний скелет) были пройдены.
Согласно распространенной классификации, в современном органическом мире выделяют империю неклеточных или доклеточных форм, к которой относят вирусы и фаги (вирусы, паразитирующие в бактериальных клетках) и империю клеточных форм. Филогенетические отношения между этими формами окончательно не выяснены. Возможно, что вирусы возникли как облигатные (обязательные) паразиты
Рис. 1.4. Основные типы животного царства и их филогенетические отношения
от более организованных живых существ за счет упрощения в процессе адаптации к паразитизму. Нельзя исключить возможность их пребывания на планете как фрагментов нуклеиновых кислот еще на добиоло-гическом (доклеточном) этапе исторического развития и приобретения ими статуса живого организма позже, при «освоении» в качестве среды жизни такой экологической ниши, как бактериальная, растительная или животная клетка.
Империя клеточных форм представлена двумя подимпериями (царства, в некоторых систематиках) - прокариот, или доядерных организмов, в которую включены бактерии и цианобактерии (сине-зеленые водоросли), и эукариот, или ядерных организмов (термины про- и эу-кариоты предложены Э. Шаттоном в 1925 г.). Общепризнанной является гипотеза об эволюционном происхождении вторых от первых. Подимпе-рия эукариот делится на царства растений, животных и грибов.
Распределение организмов по названным царствам согласуется с тремя специализированными способами питания, выявившимися на стадии древних одноклеточных эукариот. Одна группа организмов добывала пищу путем активного ее поиска и захвата (голозойный способ), вторая получала для своей жизнедеятельности вещества простого химического строения за счет их всасывания из окружающей среды (голофитный способ), третья потребляла продукты разложения органических веществ (сапрофитный способ). В ходе эволюции на основе голозойного способа питания возникло царство животных, главными чертами которых являются способность к перемещению и активный захват пищи. Эволюция на основе голофитного способа была связана с приобретением полисахаридной целлюлозной оболочки и возможности всасывать из среды воду, углекислый газ и неорганические ионы, что сочетается с важнейшей с позиций существования земной жизни в целом чертой - фотосинтезом. Указанный путь дал царство растений с присущим им оседлым образом жизни. Питающиеся сапрофитным способом организмы также ведут неподвижный образ жизни. В процессе эволюции они приобрели оболочку, которая включает полисахарид хитин. Благодаря этой оболочке организмы, представляющие царство грибов, способны всасывать высокомолекулярные вещества.
В царствах животных, растений и грибов выделяют подцарства одноклеточных, или простейших, и многоклеточных организмов, а также группы низших и высших.
В представлениях ученых разных поколений и разных направлений биологической науки принципиальная структура системы живой при-
роды воспринимается неоднозначно. В первой половине ХХ в. грибы и бактерии относили к царству растений. В настоящее время встречаются систематики, в которых категории империй и подимперий упразднены, а все живые обитатели Земли распределяются по пяти царствам - вирусы, прокариоты, животные, растения, грибы. При этом некоторые биологи находят настолько существенные различия между грибами и лишайниками, что склонны выделить последние в шестое царство. Причина кроется в объективных трудностях, прежде всего в выборе критериев и подходов, с которыми сталкивается исследователь, пытающийся упорядочить представления о мире жизни.
Этими же трудностями обусловлен тот факт, что высшим таксоном царства животных является тип, тогда как царства микроорганизмов и растений делят на подцарства, в которых затем в качестве высших таксонов выделяют группы и отделы. В царстве грибов таксонам высшего ранга соответствуют отделы.
В практической систематике живых существ (в частности, животных и растений) основной единицей классификации является вид. Исключительность положения вида среди других классификационных единиц определяется тем, что он представляет собой группировку, в которой особи существуют реально, то есть физически - рождаются, развиваются и, достигая половой зрелости, участвуют в репродуктивном процессе, исполняя главную биологическую функцию. Близкородственные виды образуют роды, которые, в свою очередь, объединяются в семейства. Близкие семейства собраны в отряды, совокупность которых образует классы, которые формируют типы или порядки (в царствах растений и грибов - отделы). Под близостью таксонов надвидового ранга понимается степень их эволюционного родства.
В классической биологии для определения степени эволюционного родства организмов, относящихся к разным таксонам, применялся метод тройного параллелизма. Он заключается в сравнении морфо-функциональных показателей взрослых представителей сравниваемых групп, их эмбрионального развития, поиске переходных ископаемых форм. Современная биология подходит к решению этой задачи также путем изучения различий в нуклеотидных последовательностях ДНК и аминокислотных последовательностях белков (молекулярные часы). За исключением деталей, картины эволюции, воссоздаваемые на основе применения классического и молекулярно-биологического подходов, совпадают (рис. 1.5).
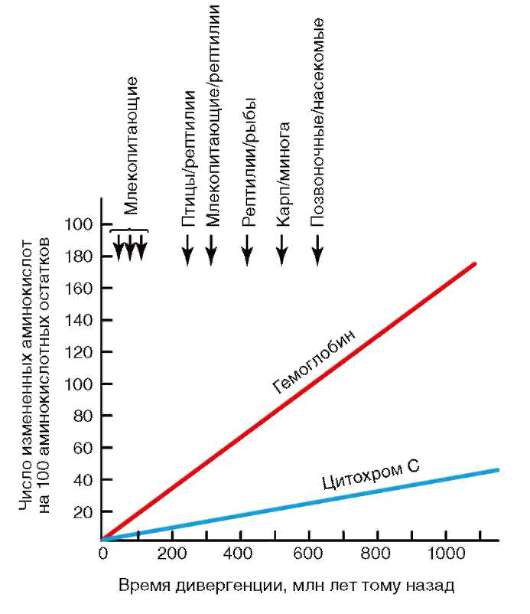
Рис. 1.5. Время дивергенции различных групп животных согласно данным молекулярно-биологических исследований
В современном органическом мире насчитывают порядка 2,5 млн видов организмов, среди которых 350-500 тыс. видов растений, не менее 100 тыс. видов грибов, более 1 млн видов насекомых, примерно 40 тыс. видов простейших (одноклеточных). Есть мнение, что называемая цифра (2,5 млн) занижена. Ожидается ее рост, в частности, за счет обнаружения новых видов, прежде всего, насекомых.
Несмотря на впечатляющие названия (империя, царство), видовой состав группы может быть скромным. Выделяемое в царстве (подим-перии) прокариот подцарство архебактерий (Archaea), например, насчитывает нескольким более 40 видов. Количество видов организмов, населяющих Землю в настоящее время, составляет не более 10% от
общего числа видов, появлявшихся на планете за время существования жизни. Что касается наиболее крупных таксонов, то в царстве животных палеонтологами и зоологами идентифицируются представители порядка 35 типов.
Среди событий, характеризующих развитие мира жизни и его современное состояние, два заслуживают особого внимания, коль скоро именно они соответствовали узловым точкам прогрессивного направления эволюции - возникновение клеток эукариотического типа и многоклеточных живых существ. При этом появление клеток с оформленным ядром (эукариотность) расценивается как более фундаментальное событие, нежели разделение живых существ на растения и животных.
1.10. эукариотическая клетка: шанс прогрессивной эволюции
В настоящее время популярна симбиотическая гипотеза происхождения эукариотической клетки. Согласно этой гипотезе (рис. 1.6, I), базовым элементом или клеткой-хозяином в эволюции клеточной организации эукариотического типа стал факультативно анаэробный прокариот-гетеротроф, способный к амебоидному движению. Переход к аэробному дыханию связан с появлением в клетке митохондрий, которые возникли вследствие изменений симбионтов - аэробных бактерий-гетеротрофов. Сходное происхождение предполагают для жгутиков, предками которых послужили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков могло иметь важное следствие общего значения. Речь идет о центриолях - структурах, играющих важную роль в митозе, и их эволюции от базальных телец жгутиков прокариот. Отметим, однако, что центриоли отсутствуют в клетках растений, которые, тем не менее, делятся митозом. Способность клеток зеленых растений к фотосинтезу связана с присутствием в них хлоропластов. Считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили цианобактерии (сине-зеленые водоросли).
Доводом в пользу симбиотического происхождения митохондрий и хлоропластов является то, что эти органеллы имеют собственную ДНК. Кроме того, ДНК митохондрий и хлоропластов - это не линейные, а кольцевые молекулы. В генах названных органелл не обнаруже-
ii
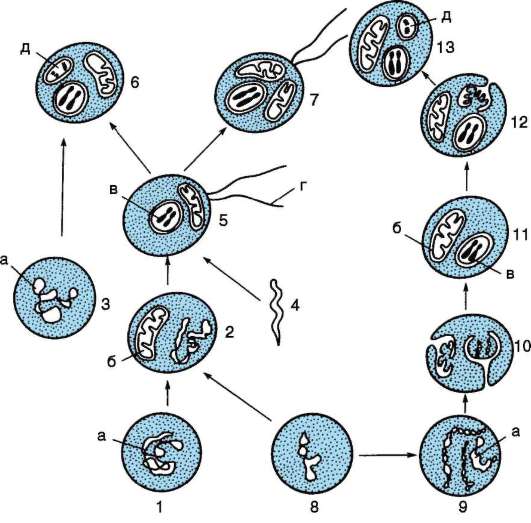
Рис. 1.6. Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам: 1 - анаэробный прокариот (клетка-хозяин); 2 - прокариоты, имеющие митохондрии; 3 - сине-зеленая водоросль (презумптивный хлоропласт); 4 -спирохетообразная бактерия (презумптивный жгутик); 5 - примитивный эукариот со жгутиком; 6 - растительная клетка; 7 - животная клетка со жгутиком; 8 - аэробный прокариот (презумптивная митохондрия); 9 - аэробный прокариот (клетка-родоначальница согласно гипотезе II); 10 -инвагинации клеточной оболочки, давшие ядро и митохондрии; 11 - примитивный эукариот; 12 - впячивание клеточной оболочки, давшее хлоропласт; 13 - растительная клетка; а-ДНК прокариотической клетки; б - митохондрия; в -ядро эукариотической клетки; г -жгутик; д - хлоропласт
но интронов. Вместе с тем бациллин и тубулин, из которых построены жгутики современных, соответственно, прокариот и эукариот, - это различные белки. У бактерий не найдено структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек «9 + 2» или «9 + 0».
Есть свидетельства высокой автономности органелл предположительно симбиотического происхождения, граничащей с их самодостаточностью. Выделенные из клеток хлоропласты, например, способны существовать и размножаться в курином яйце.
Внутриклеточные мембраны гладкой и шероховатой эндоплазма-тической сети, пластинчатого комплекса Гольджи, пузырьков и вакуолей сторонники симбиотической гипотезы рассматривают как производные наружной мембраны ядерной оболочки, которая действительно способна образовывать выпячивания. Центральным, но трудным является вопрос о происхождении ядра. В принципе оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК (у человека - несколько более 3 млрд п.н.), на порядки превышающее ее количество в митохондрии (у человека - 16 569 п.н.) или у бактерии (кишечная палочка, около 4 млн п.н.), происходило постепенно. Первоначально - путем перемещения групп генов из геномов симбионтов, а затем путем полиплоидизации (скачкообразный рост ДНК, пропорциональный гаплоидному количеству) или последовательных дупликаций нуклеотидных последовательностей (относительно плавный рост). Определенная роль в увеличении количества наследственного материала эукариот принадлежит горизонтальному переносу фрагментов ДНК. Сравнение секвенированных геномов представителей разных групп организмов свидетельствуют о том, что сотни генов человека унаследованы от бактерий, а десятки генов ведут свое происхождение от так называемых мигрирующих генетических элементов («прыгающих» генов) - еще одного участника горизонтального переноса генетической информации. Предложена альтернативная рассмотренной инвагина-ционная гипотеза, в соответствии с которой предковой формой эука-риотической клетки был аэробный прокариот (рис. 1.6, II). Внутри такой клетки-хозяина одновременно находилось несколько геномов. Последние, возможно, образовались вследствие репликации исходной ДНК, не сопровождающейся цитотомией, и первоначально прикреплявшейся к клеточной оболочке. Описаны микоплазмы, у которых единственное «кольцо» ДНК прикреплено к особой бляшке (полярный диск). Этот диск способен делиться, причем дочерние диски способны
«расползаться» по клеточной мембране без деления клетки. Среди современных бактерий известны формы с двумя, тремя и большим числом кольцевых молекул ДНК. На следующем этапе возникли органеллы, имеющие собственную ДНК, а также ядро путем впячивания и отшну-ровывания участков клеточной оболочки, представленной у микоплазм только цитоплазматической мембраной (безоболочечные «мягкокожие» формы, или молликуты). Вследствие такого отшнуровывания произошло разделение мембран, с одной стороны, отграничивающих ДНК-содержащие внутриклеточные структуры, а с другой, - выполняющих разграничительную, защитную и другие функции. В процессе дальнейшей эволюции произошло усложнение ядерного генома и упрощение геномов органелл за счет несимметричного перемещения нуклео-тидных последовательностей, возможно, с использованием механизма мобильных генетических элементов. Появилась система внутриклеточных мембран. Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрии и хлоропласта двух мембран. Однако она не дает ответа на вопрос, почему биосинтез белка в хлоропластах и митохондриях в существенных деталях соответствует таковому современных бактериальных клеток (формилметионин в качестве инициирующей аминокислоты, 22 разновидности транспортных РНК (тРНК), меньшие размеры рибосом), отличаясь по названным признакам от биосинтеза белка в цитоплазме клеток эукариотического типа. Последнее формально вполне согласуется с представлениями о симбиотическом происхождении ДНК-содержащих органелл. Вместе с тем показано, что ни один белок митохондрии, включая белки мембран, не образуется без участия ядерного генома, а каждый фермент собирается из собственных и цитоплазматических по происхождению субъединиц. Есть различия в строении митохондриального генома простейших, грибов, растений и высших животных. Геном митохондрий растений, например, длиннее генома грибов и, особенно, генома животных, а ДНК митохондрий многих простейших не замкнута в кольцо. Для геномов митохондрий грибов и жгутиковых найдены различия в генетическом коде.
Когда, сколько раз и как конкретно возникал эукариотический тип клеточной организации, неизвестно. Нельзя исключить, что существующие варианты эукариотических клеток - грибная, растительная, животная - произошли в эволюции независимо, будучи собранными из не всегда совпадающих прокариотических генетических структур или блоков.
История показала, что эволюционные возможности эукариотиче-ских клеток несравнимо выше, чем прокариотических. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по объему геном прокариот. Если сравниваемые геномы уподобить печатному тексту, то геном, например, кишечной палочки соответствовал бы книжке, имеющей 300 страниц, а геном человека - изданию из 200 томов по 1000 страниц каждый. Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов (аллелей), в повторении (амплификации) некоторых генов, а также в обилии нуклеотидных последовательностей ДНК, не выполняющих непосредственно кодирующей функции. С одной стороны, диплоидность расширяет масштабы мутационной изменчивости без угрозы катастрофического снижения жизнеспособности, так как благодаря ей из-под действия естественного отбора выводятся укрываемые в гетерозиготах «неблагоприятные» рецессивные аллели. Эво-люционно значимым следствием этого является образование резерва наследственной изменчивости, чем создается состояние генотипиче-ской преадаптации («приспособление впрок») и что интенсифицирует процесс исторического развития живых форм. Наличие в геноме повторяющихся участков открывает перспективу их дивергенции в разных функционально-генетических направлениях - см. семейства α(альфа)-и β(бета)-глобинов.
Особенностью эукариотического генома в сравнении с прокариоти-ческим является то, что у большинства генов кодирующие участки (эк-зоны) разделены некодирующими (интроны). Этим создаются условия для образования эукариотической клеткой за счет различных комбинаций экзонов (точнее, их транскриптов при осуществлении процессинга и сплайсинга пре-РНК транскрипта) большего числа белков, чем есть реальных структурных (смысловых, кодирующих) генов. Последнее при формально одной и той же генотипической базе повышает уровень фенотипического разнообразия, то есть создает больше возможностей для естественного отбора.
Возросший при переходе к эукариотическому клеточному типу объем генома, к тому же включающий в себя участки ДНК, свободные от кодирования аминокислотных последовательностей белков, открыл перспективу для использования живыми системами принципиально иных генетических механизмов регуляции жизнедеятельности. На уровне генетического аппарата это проявилось в увеличении доли ре-гуляторных сайтов ДНК, усложнении структуры промоторов (области,
с которых запускается транскрипция), смене кольцевых молекул ДНК на линейные, что допускает существенно большие возможности регуляции генетических функций ДНК, в т.ч. за счет комплексирования с различными белками (нуклеопротеидный комплекс ДНК и основных белков гистонов, нуклеосомная организация первичной хромосомной нити, эу- и гетерохроматиновые участки интерфазных хромосом, расширенный спектр ферментов репликации, транскрипции, репарации, белки-транскрипционные факторы, включая, наряду с общими, ткане-специфичные). В итоге стало возможным считывать информацию генома по частям с разных групп генов в разном их сочетании в разных типах клеток и в разное время. Так, если в бактериальной клетке одновременно считывается 80-100% информации генома, то, например, у человека от 8-10% в клетках печени и почек до 44% в клетках головного мозга. Использованию биологической информации клетки по частям принадлежит исключительная роль в возникновении в биоте Земли и дальнейшей эволюции многоклеточных организмов.
В условиях усложнения генетического аппарата эукариот трудно переоценить значение возникновения в ходе исторического развития митоза как механизма воспроизведения в поколениях генетически однотипных клеток.
Среди цитофизиологических особенностей эукариот, увеличивших их эволюционные возможности, необходимо назвать используемое на регулярной основе аэробное дыхание, обеспечивающее более высокий уровень энергообеспечения жизнедеятельности (на каждую молекулу глюкозы 38 молекул АТФ в сравнении с 2 при анаэробном процессе).
Большое значение в определении эволюционного потенциала эука-риотического типа клеточной организации отводят появлению у них цитоскелета и эластичной оболочки. С такой оболочкой связывают образование устойчивых клеточных ассоциаций и, следовательно, появление многоклеточных живых форм. С цитоскелетом - феномен структурно-функциональной дифференцировки клеток, то есть еще одно безусловное свойство многоклеточных существ.
Появление вследствие эволюционных преобразований митоза мей-оза наилучшим образом решило проблему полового размножения многоклеточных организмов с обеспечением эффективного использования в интересах эволюции явления комбинативной генотипической изменчивости.
1.11. многоклеточность: прогрессивная составляющая стратегии жизни получает
импульс
Эффективным эволюционным механизмом совершенствования жизненно важных отправлений организма является дифференциация (специализация) функций, предусматривающая их обособление и разделение. Примеры реализации этого механизма, в частности, путем функциональной специализации внутриклеточных образований, обнаруживаются уже у одноклеточных эукариот (см. рис. 2.3). Ограниченность размеров тел организмов-клеток (не более 2-3 мм в диаметре), возникающий в связи с этим лимит структур, которые могли бы служить целям специализации и наращивания количества функции, делают механизм дифференциации функций на уровне простейших эволюционно бесперспективным. В полной мере для решения задач прогрессивной эволюции возможности указанного механизма были использованы в историческом развитии многоклеточных живых существ.
Палеонтологическая летопись свидетельствует о примерно 17 независимых «попытках» эволюции многоклеточных от одноклеточных эукариот. Для водорослей (красных, зеленых, золотистых), низших грибов-аскомицетов удается воспроизвести преемственные ряды от одноклеточных через их колониальные формы к многоклеточным конструкциям с признаками наличия тканей. Последнее обстоятельство принципиально, так как многоклеточность в многотканевом варианте рассматривается как истинная.
Многоклеточность ведет к позитивным приобретениям в интересах как организмов (эволюционная парадигма), так и экосистемы (биогеохимическая парадигма). Во-первых, возникает возможность, в зависимости от конкретных эволюционных обстоятельств, достигать различных размеров тела. Последнее дает шанс освоить большее число экологических ниш, по-разному строить отношения, например, в системе «хищник-жертва» или «паразит-хозяин». Во-вторых, истинно многоклеточные организмы - это макроскопические существа, что позволяет им создавать в своем теле запас питательных веществ. Резервная биомасса крупных организмов делает их популяции менее зависимыми от ресурсной базы и повышает устойчивость экосистем.
Многоклеточность означает ускорение эволюционных преобразований путем более полного использования резерва наследственной изменчивости. Это обусловлено объединением у многоклеточных по-
лового процесса и размножения в единое целое - половое размножение. Хотя цикл индивидуального развития имеют все живые формы, не исключая вирусы, только у многоклеточных организмов отчетливо выделяется эмбриональный период. Значение этого периода заключается в том, что в нем отражен процесс исторического развития биологического вида, а эволюционно значимые изменения происходят путем преобразования эмбриогенеза.
Эволюционными предшественниками многоклеточных считают колониальные формы простейших. В качестве аналога предковой формы из ныне существующих форм называют Volvox - колониальный организм, для которого установлены дифференциация на клетки переднего и заднего полушария, разделение клеток на соматические (телесные) и гаметы (половые), наличие эмбриогенеза. Вместе с тем у Volvox нет тканей в общепринятом смысле. Предполагается, что губки ведут свою родословную от одного предка, тогда как все другие многоклеточные формы - от другого.
Палеонтологические находки второй половины ХХ в. указывают время появления ранних форм многоклеточных в биоте Земли. Богатая фауна бесскелетных (беспозвоночных) живых существ размером до 1,5 м названа по месту обнаружения эдикарской (Южная Австралия). Сходные формы, например, Kimberella - то ли медуза, то ли моллюск, обнаружены на Белом море в России. Все они включены в группу вен-добионтов или фауны вендского периода (680-570 млн лет). Для них характерен не типичный для современных многоклеточных алло-метрический, а изометрический рост, когда увеличение размеров тела происходит без изменения пропорций его частей. Само тело соответствует по форме длинной ленте, в силу чего величины площадей его внутренней и внешней поверхностей близки (аналог среди современных животных - плоские черви). Этим достигается оптимальное при газообмене и питании путем диффузии соотношение объема тела и площади его поверхности. У вендобионтов не было органов. Последнее допускает предположение, что в их жизнедеятельности немаловажная роль принадлежала эндосимбионтам-прокариотам. В рассматриваемый исторический период из современных типов животных были кишечнополостные, кольчатые черви, примитивные членистоногие и моллюски. Жизнью того времени освоены радиальная и билатеральная симметрия тела. Во времена эдикарской биоты возникали типы, которые затем бесследно сошли с эволюционной арены. Позже, начиная со среднего кембрия, бесследного вымирания типов в биоте Земли не наблюдалось.
В последней трети ХХ в. в отложениях из области Хайнань (Китай), возраст которых 840-740 млн лет, обнаружены останки макроскопических бесскелетных существ червеобразной формы. Основное отличие от вендобионтов - меньшие размеры. Хайнаньская биота, по-видимому, вымерла, не дав предковых форм ни для эдикарской, ни для современной биот. Периодом становления окружающей нас многоклеточной жизни называют поздний докембрий и кембрий (570-500 млн лет).
Существование многоклеточных живых конструкций сопряжено с большими энергозатратами, чем одноклеточных. Так, эдикарская фауна могла возникнуть при содержании О2 в атмосфере не менее 6-10%.
Между представителями хайнаньской, эдикарской и современной многоклеточных биот эволюционно-генетическая связь, по-видимому, отсутствует. Эволюционные и экологические преимущества многокле-точности, о которых речь шла выше, были реализованы в ходе исторического развития третьей из названных биот - современной. Важнейшие черты ее представителей:
• тело состоит из большого количества клеток;
• клетки дифференцированы на половые и соматические (телесные), которые, в свою очередь, также подразделяются по структуре и функциям на определенное число клеточных типов (у человека, 220-250);
• клетки расположены в организме в несколько слоев;
• клетки интегрированы в целостную систему благодаря наличию внутренней жидкостной среды, нервной системы и ряда других механизмов.
Первая из приведенных характеристик была приобретена многоклеточными в эволюции достаточно просто и, по-видимому, одинаковым путем во всех случаях независимого возникновения многоклеточных живых конструкций: среди простейших эукариот имеется множество видов колониальных организмов. Остальные черты многоклеточности связаны с формированием многослойности, благодаря чему клетки, располагающиеся на поверхности и внутри тела, оказываются в разных условиях и, кроме того, появляется объединяющая их внутренняя среда.
Родоначальником многоклеточных современной биоты считают шаровидную колонию жгутиковых, половые клетки которой смещены в глубь колонии, а соматические на первых порах обеспечивают функции перемещения колонии в пространстве и внутриклеточного пищеварения (рис. 1.7). Так как осуществление одной и той же клеткой одновременно функций движения и пищеварения малоэффективно, можно предполагать, что дальнейшие эволюционные преобразования заключались
Рис. 1.7. Происхождение многоклеточных организмов (по И.И. Мечникову). I, II - сферические колонии жгутиковых; III-V - фагоцителлы разной степени сложности: 1 - кинобласт; 2 - рыхлый фагоцитобласт; 3 - скопление чувствительных клеток на переднем конце тела; 4 - ротовое отверстие; 5 - половые клетки; 6 - эпителизованный фагоцитобласт
в специализации разных клеток в направлении преимущественно пищеварительной или двигательной функции. Результатом таких преобразований явилось оформление внутреннего слоя амебоидных клеток, осуществляющих пищеварение - фагоцитобласт, и наружного слоя клеток, обеспечивающих движение, - кинобласт.
Дифференцировка соматических клеток по названным функциям, возникшая первоначально вследствие выделения двух слоев, оказалась ключевым моментом в дальнейшем развитии многоклеточных. С двуслойностью связано появление жидкой внутренней среды, посредством которой клетки могли обмениваться химическими сигналами. Обособление и специализация части наружных клеток в направлении регистрации внешних раздражителей и передачи возбуждения на другие клетки создало предпосылки к формированию нервной системы. Гипотетический предок многоклеточных, характеризующийся наличием гамет и организацией соматических клеток в виде кинобласта и фаго-цитобласта, назван фагоцителлой (И.И. Мечников, 1877-1880). Фаго-цителлу (см. рис. 1.7 или рис. 1.8) допустимо также рассматривать как двухслойный организм с дифференцировкой клеточной массы на экто-
и энтодерму, воспроизводимые в эмбриогенезе всех ныне существующих многоклеточных в виде одноименных зародышевых листков.
Большое значение в эволюции потомков фагоцителлы имели, по-видимому, освоение ими придонного образа жизни или перемещение к поверхности, изменение характера питания (поглощение мелких или более крупных, живых или мертвых пищевых частиц) и движения (прикрепление к субстрату, пассивное или активное движение в определенном направлении). Перечисленными обстоятельствами может быть объяснено присутствие в современной биоте Земли организмов с лучевой и двусторонней (билатеральной) симметрией тела.
Важным шагом стало образование третьего зародышевого листка - мезодермы. Последний становится источником клеточного материала для мышечной и соединительной тканей, структур внутреннего скелета. С появлением мезодермы создаются предпосылки для образования у многоклеточных половых желез, чем обеспечивается надежная защита созревающих половых клеток от прямых воздействий внешней среды. Практически все животные с тремя зародышевыми листками характеризуются подвижным образом жизни и билатеральным типом симметрии. Присущая им двигательная активность с использованием мышц предусматривает напряженный обмен веществ, что, в свою очередь, требует решения проблемы выведения из организма значительного объема продуктов диссимиляции. Поэтому у трехслойных животных впервые появляется и прогрессивно эволюционирует выделительная система.
Принципиальное событие прогрессивной эволюции многоклеточных заключается в возникновении вторичной полости тела или целома, первоначально выполнявшего важную с точки зрения интенсификации двигательной активности функцию гидростатического скелета. Параллельно оптимизируются половая и выделительная функции: прежде чем быть выведенными во внешнюю среду, гаметы и продукты диссимиляции попадают в целом.
Существенное событие в историческом развитии многоклеточных - приобретение регуляторного типа эмбрионального развития. Для него в отличие от мозаичного типа типичен высокий уровень интеграции формообразовательных (морфогенетических) преобразований в развивающемся зародыше. Следствием стала высокая степень автономности индивидуального развития относительно условий, в которых оно протекает, и, таким образом, приоритет генетических факторов над средовыми в достижении требуемого результата эмбриогенеза.
Среди современных многоклеточных животных выделяют группы первичноротых и вторичноротых, различающихся ходом эмбрионального развития. Именно вторая группа соответствует прогрессивному направлению эволюции, хотя конкретные механизмы и факторы этого далеки от понимания. Для вторичноротых, в отличие от первичноротых, характерен регуляторный тип эмбриогенеза (см. здесь же, выше).
Многоклеточные формы современной биоты, кайнозойской по времени оформления (540-495 млн лет), отличаются структурной особенностью, сыгравшей существенную роль в обеспечении прогрессивного направления эволюции. Речь идет о минерализованном скелете. Трудно составить полный список преимуществ, открывающихся у обладателей твердого скелета. Прежде всего, речь идет об исключительных показателях двигательной активности на новой морфофунк-циональной основе. Биоэнергетическая база для этого в виде аэробного дыхания была готова.
В процессе исторического развития живой природы на Земле возникало не менее 35 типов многоклеточных организмов, из которых в настоящее время существует 26.
1.12. экологические кризисы в истории земной жизни
Об экологическом кризисе говорят, когда среда жизни становится непригодной для дальнейшего сохранения ранее сложившихся экосистем, выживания крупных групп организмов или даже биот. В зависимости от протяженности территории, кризис бывает глобальным или региональным. На протяжении большей части истории жизни на планете причины экологических кризисов заключались в радикальном изменении природных условий (смена анаэробной среды на аэробную, покровные оледенения). С появлением людей стали возможны кризисы антропогенной природы.
Один из ранних глобальных экологических кризисов датируется временем, отстоящим от наших дней на 1,8-1,7 млрд лет. Около 1,9 млрд лет тому назад концентрация О2 в атмосфере Земли достигла 1%. «Кислородная революция» заняла порядка 100-200 млн лет. В связи с этой революцией сложившаяся ранее почти исключительно анаэробная био-та вследствие «отравления» кислородом претерпела катастрофические изменения. Многие из экосистем погибли, другие перестроились. Мир жизни в целом из анаэробного превратился в аэробный.
Правилом является то, что после очередного кризиса жизнь не только сохраняет себя, но входит в следующий этап расцвета на основе уже иных форм. Так, на смену прокариотической протерозойской (докем-брийской) биосфере пришла современная фанерозойская (кембрийская) биота с переходом лидирующих позиций к эукариотам и истинно многоклеточным скелетным организмам.
Современный этап истории планеты характеризуется развитием глобального экологического кризиса, но антропогенной природы. В его основе лежит нарастающее давление человечества на среду жизни, обусловленное неуклонным ростом народонаселения и появлением новых технологий, расширяющих доступ людей к природным, нередко невосполнимым ресурсам, прежде всего энергетическим. Суммарная численность древнейшего человека умелого (Homo habilis) достигала 100 тыс., древнего человека прямоходящего, в частности, синантропов (Homo erectus pekinensis) - 1 млн (по другим данным 300 тыс.), современного человека разумного (Homo sapiens) на момент появления вида (кроманьонцы) - 3 млн, а к 1500 г. н.э. - 350 млн особей. В настоящее время население Земли составляет порядка 6 млрд.
Первый антропогенный экологический кризис датируется верхним палеолитом и связан с эффективным использованием людьми техники загонно-облавной охоты на крупных млекопитающих. Позже, уже в мезолите (примерно 15 тыс. лет назад) изобретены лук и стрелы, к участию в охоте приобщаются собаки, что повышает ее результативность (рис. 1.8 или рис. 1.9). Итог деятельности охотников того времени - уничтожение мамонта, носорога Мерка, пещерных медведя, льва, гиены, сокращение поголовья зубров, диких лошадей и многих других видов «промысловых» животных. Активность первых людей внесла существенные изменения в фауну Земли, лишило человечество тех времен главного пищевого ресурса и создала кризисную ситуацию. В мезолите в жизнь людей вошли войны. Выход человечества из палеолитического кризиса был обусловлен «неолитической революцией» (неолит, начало от Х-IX вв. до н.э., датировка различается для разных регионов). Она состояла в переходе к одомашниванию (синантропизация) животных (рис. 1.9 или рис. 1.10) и культурному земледелию.
Неолитическая революция выявила важную закономерность. Бесконтрольная деятельность людей по обеспечению своего «сиюминутного» благополучия, немыслимая без изъятия природных ресурсов во все возрастающих объемах, в сочетании с ростом народонаселения порождает антропогенные кризисы. Так, одним из разрушительных

Рис. 1.8. Загонно-облавная охота. Наскальный рисунок в Сахаре, примерно 7 тыс. лет назад

Рис. 1.9. Дойка коров на стоянке. Наскальный рисунок в Сахаре, примерно 7 тыс. лет назад
результатов неолитического скотоводства, обусловленным перевыпасом, стало появление на карте африканского континента пустыни Сахара, на месте которой 10 тыс. лет назад была саванна. «Ноу-хау» неолита стало поливное земледелие, которое стабильно обеспечивало более высокие урожаи. Ирригация, однако, вела к засолению и смыву почв с прогрессирующим снижением их продуктивности, заиливанию и высыханию рек. Опустынивание обширных территорий в неолите, приведшее к неолитическому антропогенному экологическому кризису, было следствием порочности господствующей тогда системы обработки земли, а рост численности домашних животных близ поселений людей способствовал ускорению процесса.
Антропогенные экологические кризисы, если их оценивать с позиций интересов не природы, а общества, являются экономическими. История человечества дает достаточно примеров, подтверждающих правило, - освоение новых прогрессивных технологий, расширяющих возможности людей в плане интенсификации потребления природных ресурсов, с одной стороны, служит преодолению возникающих кризисных ситуаций и обеспечению дальнейшего роста народонаселения, но с другой - с течением времени неизбежно порождает очередные экологические проблемы.
Вопросы для самоконтроля
1. Что изучает биология?
2. Перечислите основные свойства жизни.
3. Каковы основные гипотезы происхождения жизни и в чем заключается их суть?
4. Перечислите уровни организации жизни. Каковы элементарные единицы и элементарные явления на каждом из них?
5. Изложите основные положения симбиотической и инвагинаци-онной гипотез происхождения эукариотической клетки.
6. Какие прогрессивные возможности существования организмов предоставляет многоклеточность?
7. В чем заключается биосоциальная природа человека?